
Abstract
The oxygen transport system in the human body is described from atmospheric air to working skeletal muscles. It is illustrated that the pulmonary diffusion capacity becomes critical during whole body exercise as the arterial oxygen tension decreases markedly. Furthermore, the Bohr effect on the oxyhaemoglobin dissociation curve means that, with a lowering of pH to below 7.0, arterial oxygen saturation may decrease to below 90%. In addition, the ability to increase cardiac output limits the oxygen transport capacity when working with several muscle groups at the same time, and muscle blood flow is reduced by approximately 30% compared with times when only a single muscle group is activated. Oxygen diffusion to the myocytes thereby becomes affected, while endurance training enhances blood volume and the number of capillaries surrounding the muscle cells as well as the size of the heart, and thus increases maximal oxygen uptake by approximately 50%.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Exercise physiology describes the adaptive changes taking place in the organism in response to work or exercise. A related area is the biomechanics of human movement, e.g. during walking and running. Similarly, problems associated with labour and mechanisms involved in the development of muscle fatigue and eventual chronic pain are addressed. Exercise physiology evaluates the responses to training vs detraining, the adaptations to special environments such as high altitude, or eventually zero gravity during space flight vs the increased pressure during diving. Finally, it is increasingly realised that regular exercise is beneficial towards weight control and sustained health while competitive sport may be associated with problems related to doping.
2 Skeletal Muscles
Physical work is carried out by the skeletal muscles and two main muscle fibre types have been identified. While fast twitch muscle fibres (FT) are able to generate force quickly they are, in general, unable to maintain a contraction. On the other hand, slow twitch fibres (ST) possess low myosin ATPase (Fig. 1), and can maintain the developed force for longer periods since they are designed for aerobic metabolism with many mitochondria and a large amount of myoglobin. Among all the muscles in the body, distribution of different fibre types vary but it is, on average, around 50%/50%, while endurance trained athletes appear to be selected from among people with a percentage of ST fibres that is about 70% or even larger. The endurance capacity of the ST fibres is reflected not only in the high number of capillaries and mitochondria but also in the enzymes that they encompass, succinate dehydrogenase, etc., that enable them for high aerobic metabolism.

Cross-sectional view of skeletal muscle with identification of the two main fibre types (I slow twitch; IIa, fast twitch trained; IIx, fast twitch untrained) by ATPase staining after incubation at pH as indicated (Jesper Løvig Andersen, Copenhagen Muscle Research Center)
Although it is the skeletal muscles that perform the work, it is the brain that decides to initiate and terminate work. In this context, it may be that the end of exercise is defined by an inability of the brain to access the motor neurons. Such “central fatigue” manifests when exercise becomes strenuous or difficult, such as the first time on a bicycle, and appears, paradoxically, to be associated with difficulty in recruiting ST fibres. Thus, a fundamental aim of training is to automate the motor pattern required for a given movement.
3 Metabolic Demand
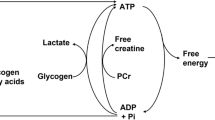
Metabolism is divided into an aerobic and anaerobic pathway for ATP generation. At the onset of exercise it takes some time before aerobic metabolism is fully activated to cover the energy requirements of the work performed (Fig. 2). During these initial seconds of exercise, metabolism is covered, in addition to the creatine phosphate and the oxygen bound to haemoglobin and myoglobin, by the production of lactate. The required energy that is not accounted for by the pulmonary uptake of oxygen (VO2) is termed the oxygen deficit and can be divided into a lactic and an alactic component. Conversely, it takes some time before VO2 returns to the resting value when exercise is terminated. This additional metabolic component, termed “the oxygen debt”, is related not only to the work carried out per se but also to the increase in metabolism associated with an elevated body temperature and is likely to be twice as large as the oxygen deficit.

Oxygen consumption (VO2) and heart rate during a 2,000-m simulated rowing race in two rowers. The energy not accounted for by the VO2 at the beginning of the exercise is termed oxygen deficit
The relative contribution of the aerobic and anaerobic components to the work carried out varies with intensity and distance covered. The aerobic and anaerobic contributions to maximal exercise are approximately equal when the event lasts ∼2 min (Fig. 3). VO2max represents the metabolic rate at the pace of an event that lasts approximately 10 min, while during longer events only a fraction of the VO2max can be utilised. Thus, a successful marathon runner is described not only by his VO2max but also by his ability to maintain metabolism at a high level around VO2max, e.g. 90% rather than 70% for a less qualified runner, and a high percentage of ST fibres is likely to contribute to that ability. In general, the work intensity and the time it can be maintained are related by a reverse hyperbolic function.

The aerobic (circle) and anaerobic (shaded circle) contributions to maximal exercise. Courtesy of K. Jensen, University of Southern Denmark
However, the aerobic and anaerobic capacities are not a full description of an athlete's ability to excel. Much work is performed, for example during running, to support the body in each step and to swing the legs (and the arms). Thus, the characteristics of a successful runner are long light legs and a low body weight. Conversely, in rowing, where the crew is carried by the boat, it is an advantage to be tall and heavy with a relatively long upper body and arms. For comparison of small and large individuals, the VO2max can be expressed relative to body weight to ¾ power (Fig. 4), reflecting the dependence of running performance to VO2/kg, while for rowing the absolute value (L min−1) is most relevant.

Relationship between VO2max and body mass. Regression lines with 95% confidence intervals and correlation coefficients (r) are shown. (A) The VO2max increases with body mass, but when it is expressed relative to body mass (B), those subjects with the largest body mass show the smallest values, when trained for the same sport. When scaling VO2max with body mass to the 0.73 power (i.e. ml kg−0.73 min−1; C), VO2max becomes independent of body mass
3.1 Metabolic Rate
At rest and during exercise, aerobic metabolism, or the sum of mitochondrial respiration rates in various organs, is represented by the rate of VO2. The relative contribution of these organs changes with physiological state, such that internal organs such as the liver, kidneys and brain account for most of the whole-body VO2 at rest, while locomotor muscles account for >90% of VO2max during maximal aerobic exercise. VO2max, which reflects the maximal metabolic rate, is elicited by the energy needs of locomotor activity, and it increases with the volume of muscle mass involved in exercise. In contrast, the basal metabolic rate (BMR) reflects the lowest need of energy at rest that is used for all sorts of housekeeping functions, such as maintaining cell potentials, driving the heart, and maintenance of body temperature, an indirect measure of plasma thyroxin. BMR and VO2max thus define the span over which the aerobic metabolic rate of the organism can vary. Whereas BMR appears to depend on body mass to the ¾ power, VO2max shows large inter-individual and inter-species variability, related to the degree of work or exercise capacity. It is typically about tenfold higher than BMR even though well-trained athletes can achieve a VO2max up to 20 times higher than their BMR, and even greater variation is found in animals.
4 Static and Dynamic Work
During exercise, work is performed by muscle fibres shortening and developing tension against gravity, against the mass that is moved, and through the generation of heat, depending on the intensity of the exercise. The relationship of movement to tension varies depending on the type of work. With static work, which is quasi-isometric and can be of explosive intensity, the movement is small in relation to the tension and thus it differs from dynamic work (quasi-isotonic) in which the movement is relatively large. Dynamic work is mostly produced by repetitive contractions involving primarily ST muscle fibres and the energy cost is mainly met by aerobic metabolism. Static work entails relatively sustained muscular contraction that involves FT muscle fibres which produce lactate, and the energy cost is mainly met by anaerobic metabolism. Most exercise activities or sports entail a mixture of static and dynamic work components and thus, the physiological responses to a given exercise activity are (relatively) unique. For example, power weightlifting has a large static component that limits its duration to relatively short periods.
5 Energy Requirements and Cardiorespiratory Limitations
There are many different physiological and anthropometric parameters that can be used alone or in combination with other parameters to predict the competitive success of endurance athletes. It appears, however, that one of the most important physiological determinants of endurance performance is VO2max. Therefore, obtaining high aerobic power values is of importance for the successful endurance athlete, and training is characterized by large amounts of low intensity training. Anaerobic energy systems are used mainly during the start phase of an endurance race (for approximately 60–80 s), as it takes a minute or so for VO2 to reach its maximal level (Fig. 2), and to a lesser extent also during the finish of an endurance race.
VO2max integrates the ability of the lungs to provide O2 to the blood, the O2-binding capacity of the blood, maximum cardiac output, the muscle type, and the number of capillaries surrounding the muscle fibres. The schematic view of the main organ systems that are involved in the development of VO2max and their trainability is presented in Fig. 5. During exercise no single factor limits the O2 transport cascade expressed as the sum of the resistances presented by each step. Yet, in order to preserve arterial pressure during whole-body exercise, blood flow to working muscles and internal organs, including the brain, is restrained, with adverse outcomes for metabolism and fatigue.

Organ systems and their trainability to impact maximal oxygen consumption (VO2 max) and exercise performance
5.1 Aerobic Metabolism
Over the course of an event lasting 5–7 min, elite athletes exercise with the majority of work performed between 95 and 98% of VO2 max. For example, male elite level heavyweight rowers have a VO2 of up to 6.51 min−1, while female heavyweights have about 4.41 min−1 (Table 1). There is a strong linear relationship (r = 0.99) established between placement in a championship and the crew's average VO2 max (Fig. 6). The highest VO2 max reported is 7.41 min−1 for an elite Finnish cross-country skier.

Regression line between average VO2 max of a crew and placement in a rowing championship. The 95% confidence limits of the regression are also shown
Seasonal changes in VO2 max have been described with increases of 10 mL min−1 kg−1 or ∼10% during the competitive season, even though these increases in relative values of VO2 max be highly dependent on decreases in body mass. VO2 max increases with age, training distance per year, and with sporting experience. Usually, VO2 max levels off at about the age of 18 but increases can be seen as a consequence of endurance training until around the age of 24 (Fig. 7). In parallel, increases can be seen in other endurance parameters.

The development of maximal oxygen consumption (VO2 max), ventilation and maximal aerobic power (P max) of elite rowers from junior to senior level
A plateau in VO2 max has been observed in highly trained endurance athletes despite increased training volumes. Even with the attainment of a plateau in VO2 max, endurance performance can still be improved. This can be achieved by increasing the endurance capacity — the ability to maintain higher VO2 over the distance — and increased economy, defined as a larger work performed for a given VO2. Since endurance capacity appears to be important in performance prediction when athletes of similar VO2 max values are compared, alternative parameters for the measurement of endurance capacity have been proposed. For example, it is suggested that the power which elicits a blood lactate level of 4.0 mM is the most predictive parameter for competition performance.
5.2 Anaerobic Metabolism
Anaerobic metabolism is indicated by a high peak blood lactate concentration, which increases exponentially with work rate, the muscle mass involved in exercise and the motivation of the athlete. Values of 11 mM have been reported after treadmill running, while values of 15–17 mM are commonly observed after rowing competitions. Accordingly, the buffering system of the blood (bicarbonate) may be eliminated from the blood and the pH of the blood decreases from its normal value of 7.4–7.1. The record low measured pH value is 6.74 and corresponds to a blood lactate level of 32 mM. However, these values give little indication of the quantity of anaerobic metabolism. For that purpose the “oxygen deficit” can be calculated (Fig. 2). For example, in rowers the oxygen deficit has been reported to be 90 ml kg−1, or substantially larger than reported in runners.
6 Endocrine and Metabolic Responses
In response to mental and physical stress, circulating eosinophils decrease from 110 to 20 mm−3 before an endurance race, and even to 3 mm−3 after exhaustive exercise, while plasma ACTH, adrenal cortical hormone and adrenaline increase. After maximal ergometer exercise, involving a large muscle mass, plasma adrenaline and noradrenaline concentrations increase from the resting values of 0.9 and 2.3 nM, respectively, to extremely high values of 19 and 74 nM. These values are about twice as large as those noted during running, suggesting a role for muscle mass in the catecholamine response to maximal exercise. Also, pancreatic polypeptide, a hormone under vagal control, increases 2.5-fold after maximal exercise, suggesting that vagal activity may contribute to the feeling of fatigue including gastrointestinal symptoms experienced at exhaustion.
Protein turnover is larger, while the protein synthesis is not increased, in trained athletes compared with controls, supporting the theory that intense exercise does not increase basal protein turnover rate. Yet, anabolism, and hence muscle build up, may be aided when a protein rich meal is provided soon after exercise. Serum androgen and growth hormone increase from 700ng ml−1 and 5 μU ml−1 respectively at rest, to 850 ng ml−1 and 50 μU ml−1 respectively after maximal running, but do not increase after submaximal exercise. These changes occur despite a constant serum-luteinizing hormone (60 ng ml−1). It may be speculated that the raised level of androgen during exercise acts in association with growth hormone to increase the pubertal growth velocity and contributes to muscular development associated with exercise. Administration of testosterone, however, does not increase muscle glycogen build-up after maximal dynamic exercise. Serum concentration of myoglobin and creatinine kinase (a cellular enzyme) is elevated after exercise lasting 30–40 min and remains elevated 1.5 h after the training bout, indicating skeletal muscle cellular damage which could help explain the subsequent muscle soreness. Consequently, creatine supplementation enhances performance.
6.1 Blood Glucose
Blood glucose is essential for brain metabolism and supplements muscle metabolism during exercise. With the levels of plasma adrenaline observed during maximal exercise, glucolysis in the liver is stimulated and contributes significantly to the maintenance of blood glucose. Thus, intense exercise is associated with optimal blood glucose concentrations, but the liver glucose may become depleted during prolonged exercise lasting ∼12 h. Thus, during cross-country skiing and mountaineering a readily available carbohydrate source, in the form of e.g. chocolate, is essential. In these circumstances, when the blood glucose level decreases brain metabolism is compromised, resulting in an inability to maintain movement, and thus body temperature decreases, with often grave consequences.
7 Ventilation
The lung is the organ for exchange of gases between air and blood. Carbon dioxide produced from metabolism is unloaded to the alveoli while O2 diffuses into the blood where it binds to haemoglobin. The O2 transport is passive, as the driving force is based on the gas pressure gradients between air and blood, and is limited by the diffusion capacity of the lungs.
At rest, ventilation (V E) involves activation of the diaphragm muscle, which operates as a motor piston to generate a negative pressure within the thorax whereby external air passively enters the lung alveoli. During exercise the diaphragm operates with increased force, while the internal intercostal muscles help to expand the ribcage and further decrease the intrathoracic pressure. Thus, increased V E is generated by increased breathing frequency and increased depth of each breath. Very large V E levels, typically greater than 2001 min−1 and sometimes as high as 2601 min−1, are developed in large athletes during competition. During a 6 min all-out effort, V E rises exponentially until the third minute, but nevertheless it continues to increase until the end of the effort. Since breathing frequency does not differ between trained and untrained, performance is more favourable for athletes with large total lung and vital capacities (VC). Hence, the lungs of athletes reflect their large bodies, and a VC of 9 L has been recorded in an elite rower. This observation reflects selection as VC is the only link of the transport O2 chain that does not improve with training once adult stature has been attained. Thus, a correlation between endurance performance and VC is reported regularly, and successful athletes, e.g. rowers, typically demonstrate VC values of 71, compared to 5.51 as expected for their body size.
A large V E could be considered to be a disadvantage as it requires a higher activation of ventilatory muscles which compete with the locomotor muscles for their share of cardiac output. In fact, unloading of breathing by mechanical assistance increases blood flow to skeletal muscles and specific training of the inspiratory (but not the expiratory) muscles may enhance endurance performance.
7.1 Breathing Pattern
During exercise breathing is entrained to the locomotor rhythm, meaning that expiration and inspiration are performed in phase with the limb movements. The V E and the pattern of breathing during exercise are the result of respiratory muscle contraction acting on the mechanical properties of the respiratory system. The tidal volume is determined by the size of the lungs, and both inspiratory and expiratory times are influenced by maximum airflow. Inspiratory flow rate is limited primarily by the ability to generate inspiratory muscle pressure, while expiratory flow rate is limited by airway mechanics, rather than the ability to generate expiratory pressure. Peak expiratory flow rates can reach 151 s−1, but in some of these individuals a plateau in expiratory flow rates suggestive of airway collapse is also observed.
Entrainment, or synchronisation between limb movements and breathing pattern, is a common occurrence during exercise and several factors are associated with it. Since contraction of the thoracic musculature is required both for respiration and stabilisation of the trunk during exercise, it is important that breathing and limb movements are synchronised so that one does not interfere with the other. The physiological benefits from such coupling are an improvement in the efficiency of the respiratory muscles and gas exchange, prevention of diaphragmatic fatigue, and reduction of breathing effort sensation. Hence, limb movements influence respiratory efficiency and, at the same time, respiratory muscles modulate the locomotor rhythm.
During incremental exercise, ventilatory tidal volume progressively increases up to the point where each breath encroaches upon the flat portion of the pulmonary compliance curve. Hence, a greater portion of the negative intrapleural pressure is used to overcome pulmonary elastic work. To circumvent this mechanical limitation of lung distension during inspiration, there is a shift in the breath/stroke ratio, for example from a 1:1 to a 2:1 during rowing, where the respiratory system is confronted with a different mechanical constraint, that of expiratory flow limitation, due to the limited time of each expiratory phase.
7.2 Diffusing Capacity
Oxygen enters the body through the lungs that have an enormous gas diffusion capacity with an alveolar area of 50–100 m2. Increased V E is coupled intimately to exercise but the limiting role of the lungs to O2 transport remains unknown for two reasons. First, lung function is described as a capacity rather than as the actual contribution to transport of O2 during exercise. Furthermore, evaluation of the lungs is combined with the capacity of haemoglobin in pulmonary capillaries to take up O2, expressed as the pulmonary diffusion capacity (DL CO) using carbon monoxide (CO) as test gas. The more relevant value of diffusion capacity for O2 is 23% larger than DLCO and the value for CO2 is 25 times DLCO. DLCO is a non-invasive measure of lung function using CO as test gas because its affinity to haemoglobin is 200 times that of O2. When ∼1% CO is added to inspired air, CO is binding with haemoglobin over the entire length of the pulmonary capillary (Fig. 8) and under these conditions DLCO represents the gas transport capacity.

Diagram of oxygen tension of blood (PcO2) during its course through a pulmonary capillary at two flow rates
Also shown is a cartoon of red cells passing through a pulmonary capillary with carbon monoxide (CO) binding to haemoglobin (bright red). The apparent pulmonary diffusion for oxygen (DmO2) expresses the effective area for diffusion rather than the diffusion capacity established when the whole capillary is engaged in oxygen transfer. Because there is little or no transport of O2 in the venous end of the capillary, the effective capillary diffusion area (s1 or s2) is approximated by the insertion of a line integrating the considered increase in PcO2. When it takes longer for the alveolar-to-capillary O2 equilibrium to be established because of doubled flow during exercise or because of low inspired O2 tension, the venous end of the capillary becomes increasingly important and an enhanced effective diffusion area is expressed as an elevated DmO2. Discrepancy between a doubling of the diffusion capacity for CO and an eightfold increase in DmO2 during exercise indicates that at rest in normoxia, an equilibrium is established between the alveolar O2 tension (PaO2) and PcO2 within one fourth of the length of the pulmonary capillary. During hypoxemia, however, equilibrium is not reached and the entire capillary contributes to diffusion expressed as a maximal DmO2. Any venous admixture to arterial blood is reported as a change in DmO2, representing an integrated measure of pulmonary function rather than of diffusion per se. To validate the absolute value, pulmonary venous O2 tension needs to replace the arterial O2 tension (PaO2) in the calculation of DmO2 since blood serving the pulmonary tissue is drained in the left atrium of the heart.
Conversely, DLCO is dominated by acute changes of the pulmonary capillary blood volume. For example, DLCO decreases by 15% from the supine to the seated position because pulmonary capillary blood volume is reduced while the pulmonary membrane diffusion is enhanced by 30%. During exercise, there is a doubling of DLCO (25–50 ml mmHg−1 min−1), reflecting a support of the muscle pump to the pulmonary capillaries blood volume with recruitment of pulmonary capillaries in the apical parts of the lungs (Fig. 9).

Pulmonary diffusion capacity following 6 min all-out maximal exercise
7.3 Transport of O 2
In normoxia equilibrium between alveolar and capillary O2 tensions is established within a fraction of the capillary blood transit time (Fig. 8). During exercise, as the velocity of red cells in pulmonary capillaries increases, equilibrium takes longer to be established and, ultimately, as the length of the capillary is involved in the diffusion of O2, the diffusion capacity is utilised.
As the length of the capillary involved in diffusion increases during exercise, it becomes difficult to maintain alveolar O2 tension (PAO2), and an increase in V E is required to compensate for O2 taken up by capillary blood. During exercise, V E follows two distinct patterns. Low-intensity exercise is associated with little drive to V E, representing the influence from the central nervous system (central command). Relative hypoventilation is expressed as a ∼4 mmHg elevation of the arterial CO2 tension (PaCO2), which is the dominant stimulus for V E at rest. As workload progresses, V E increases exponentially, influenced by the decreased blood pH. The important effect of blood pH on driving V E is demonstrated by the reduced V E (∼151 min−1) following normalisation of pH with bicarbonate administration despite the 7.5 mmHg increase of PaCO2. During intense exercise the marked hyperventilation decreases PaCO2 below the resting value and PAO2 increases to high levels (130 mmHg).
Even though during light to moderate exercise the lungs facilitate O2 transport by recruiting alveoli and enhancing the within-capillary diffusion area, a diffusion capacity of ∼80 ml mmHg−1 min−1 (an eightfold increase from rest) is insufficient to account for the often more than 15-fold increase in VO2 during maximal exercise in athletes. Further, during intense exercise VO2 depends on a widening of the alveolar-capillary O2 tension difference because gas diffusion is impeded by high pulmonary artery pressure that provokes accumulation of fluid in the alveolar-capillary space. Thus PaO2, expressed as a physiologic shunt or exercise-induced hypoxaemia, decreases (e.g. from 90 to 75 mmHg) although PAO2 increases.
During intense exercise the elevated V E, as demonstrated by the extraordinary large rates observed, increases PAO2 and, consequently, PaO2 is maintained at the highest possible level. Such mechanism may explain the enhanced exercise performance following specific respiratory muscle training that supports a large V E and PaO2, as indicated by the elevated end-tidal O2 tension during exercise. In contrast, pulmonary diffusion capacity does not respond to training and the large reported values for DLCO represent selection of gifted individuals. DLCO varies from 15 to 45 ml mmHg−1 min−1 and this range reflects differences in body size and central blood volume in addition to variation in pulmonary membrane diffusion capacity. Pulmonary membrane diffusion capacity is larger than DLCO because it does not include the resistance to diffusion of O2 in plasma, or the binding of CO to haemoglobin.
7.4 Haemoglobin
Oxygen is dissolved in blood and it also binds to haemoglobin, whereby the O2-carrying capacity is markedly increased. Dissolved O2, described by the PaO2 (normal value 100 mmHg), determines, together with the acidity of the blood (normal blood pH is 7.43), the level of haemoglobin saturation with O2 (normal value 97–99%). The transport of O2 in blood by haemoglobin is described by the oxyhaemoglobin dissociation curve (Fig. 10) reserving ∼2% of the transport to O2 dissolved in plasma during maximal exercise.

The oxyhaemoglobin dissociation curve. With a decrease in pH and increase in temperature, the curve is shifted to the right
At rest and during moderate exercise, pH has little influence on the amount of O2 transported. During maximal exercise where, despite the increased drive to breath, PaO2 is reduced to a level similar to that seen at rest in high altitude, a right-shift (Bohr effect) of the oxyhaemoglobin dissociation curve demonstrates that any deviation in pH affects the ability of haemoglobin to transport O2 and, consequently, VO2max is affected in proportion to the reduction in haemoglobin O2 saturation. During maximal exercise the Bohr effect can reduce arterial haemoglobin O2 saturation to ∼90% and impose a 5–10% restraint on VO2max. By breathing O2-enriched air, where the inspired O2 fraction is increased from 0.21 to 0.30, PaO2 is increased to above normal values and the influence of a low pH on haemoglobin O2 binding is attenuated, resulting in the restoration of arterial haemoglobin O2 saturation. The V E is not affected, but, with a higher O2 pressure gradient and high haemoglobin O2 saturation, blood O2-carrying capacity increases, which induces a higher O2 uptake whereby exercise performance is enhanced by 2%. Arterial haemoglobin O2 saturation is also restored in response to pH normalisation, following bicarbonate administration, and VO2 max also increases. On the other hand, O2 delivery is enhanced both by the effect of exercise-induced body temperature elevation and the Bohr effect, augmented by the lower pH of venous compared to arterial blood (e.g. 7.0 vs 7.2) as CO2 carbon dioxide is exhaled (PCO2 reduced from 90 to 40 mmHg; venous vs arterial).
The impact of the lung in limiting exercise performance is demonstrated as even a V E of more than 2001 is not enough to maintain the PaO2 during maximal rowing, and even higher V E may offer some protection against the reduction in PaO2. Therefore, V E may contribute to limitations of O2 transport and it is also a factor that may discriminate between winners and losers.
8 Blood Lactate
The Bohr effect on the oxyhaemoglobin dissociation curve illustrates the delicate balance between aerobic and anaerobic metabolism during maximal exercise. Appreciating the various tactics endurance athletes apply during a race, it is likely that the degree of O2 transport is affected by acidity. This varies between individuals depending on PaO2, which is influenced by the balance between hyperventilation and pulmonary membrane diffusion capacity. Middle-distance races start at high speed to accelerate the increase in VO2 because the total aerobic metabolism is represented by the accumulated VO2 during the race rather than the highest level VO2 reaches.
More is known about exercise at high altitude. When the inspired O2 tension is low, it is disadvantageous to work at an intensity that provokes lactate acidosis, because any anaerobic contribution to metabolism attenuates the more important O2-carrying capacity of blood. While it is possible to work at an intensity that provokes lactate acidosis, exercise in hypoxia is usually associated with a small deviation in pH, a condition that has been defined as the high-altitude lactate paradox. Following high-altitude acclimatization, where both ventilation and PaO2 increase, plasma lactate during maximal exercise is comparable with the sea-level values. The unproven, but probably beneficial, effect of moderate altitude training (∼2,000 m) for sea-level performance may be explained by the adaptation of the respiratory muscles to the large ventilation in hypoxia. Upon return to sea level, it feels easier to maintain high ventilation and elevate PaO2 during competition, and thus enhance the anaerobic contribution to exercise without affecting the oxygen transport capacity of haemoglobin.
8.1 Lactate Threshold
Interest has also focused on the lactate threshold or the work rate that elicits a blood lactate concentration of 4 mM. This work rate increases with training, and it seems to depend on the muscle fibre composition. Athletes with many slow-twitch fibres are able to exercise at a high intensity with a blood lactate value of no more than 4 mM.
The Bohr effect on the oxyhaemoglobin dissociation curve explains why the work rate, at a given blood lactate level, is a sensitive predictor of endurance performance. Blood lactate increases exponentially with workload but their relationship is right-shifted following training, i.e. blood lactate increases with relative workload. The workload that elicits a given lactate level (often set at 4 mM) is, thereby, an indirect measure of VO2 max that, in itself, is a predictor of performance. Blood lactate is a more precise performance predictor because blood lactate reflects not only VO2 max but also the ability to work without affecting the oxyhaemoglobin dissociation curve.
For a given workload less lactate is produced, with the recruitment of ST rather than FT muscle fibres, as illustrated when ST fibres are prevented from contracting with curare-induced (South American arrow poison) partial neuromuscular blockade. The workload that elicits a given blood lactate level reflects the work capacity of ST muscle fibres. The composition of muscles depends not only on the percent of ST vs FT fibres but also on their relative size. For example, weightlifters develop large FT fibres in adaptation to rapid lifts, while rowers are characterised by large ST fibres reflecting the relatively slow movements involved in rowing (Fig. 3). Furthermore, considering that central fatigue inhibits ST muscle fibre recruitment, in high exercise intensity which requires increased central command a smaller contribution to work from ST muscle fibres necessitates that work has to be carried out with a larger contribution from FT muscle fibres, which results in elevated lactate production. In other words, evaluation of blood lactate during submaximal exercise reflects the mental preparation and automatisation of the movement that determines the central command requirements.
Alongside these considerations it should also be noted that although it is lactate that is measured in blood, it is the deviation in pH that influences oxygen transport. Lactate is a substrate for tissues including muscle, liver, kidney and brain but its exponential accumulation in the blood, as the work rate increases, is a manifestation of attenuated elimination by liver and kidneys. Blood lactate is, thereby, also an indicator of how well organ blood flow is preserved during various exercise intensities.
9 The Heart Rate Response
The heart rate response to exercise is of particular interest because the almost linear relationship between heart rate and work load (or VO2) is applied widely for evaluation of VO2 max in population studies. Furthermore, heart rate is also a determinant of cardiac output that supports blood pressure, which in turn affects perfusion of the working muscles and the brain. Hence, a brief presentation follows of the factors that affect heart rate, cardiac output, blood pressure and blood flow to vascular beds critical for endurance exercise performance.
9.1 Blood Volume and Cardiac Preload
The heart of quadrupedal animals is on the same level as the main portion of blood within the body, but upright humans face a circulatory challenge as the indifference point for volume is at the level of the pelvis and about 80% of the blood volume is positioned below the heart. Thus, in response to reduced central blood volume, cardiovascular reflexes including sympathetic activation and the veno-arterial reflex are important for maintaining the upright position. Yet, it is not possible to remain upright without the muscle pump preventing the accumulation of blood in dependent parts of the body, as is the case in soldiers standing still in line who faint, with a concomitant decrease in heart rate and blood pressure. This reflex, defined as vasovagal syncope, is elicited when central blood volume is reduced by 30% due to gravitational pooling in the upright posture, and the associated reduction in blood pressure is attributed to a Bezold–Jarish-like reflex that induces vasodilatation in skeletal muscles at the expense of flow to the brain.
9.2 Starling's Law of the Heart
The influence of central blood volume, or cardiac preload, on the function of the heart is described by Starling's law of the heart (Figs. 11 and 12). In this context, normovolaemia, defined by the absence of further increase in stroke volume or cardiac output when central blood volume increases, is achieved in the supine posture. During head-down tilt that increases diastolic filling of the heart, there is no further stroke volume increase, indicating that the upper flat part of the Starling curve has been reached. Conversely, when upright, central blood volume is reduced and the heart operates on the ascending part of the Starling curve where cardiac output depends on preload.

Starling curve for the human heart as evaluated during head-up and head-down tilt. During supine rest, the upper flat part of the curve is reached

Starling curve for the human heart as evaluated during head-up and head-down tilt. During supine rest, the upper flat part of the curve is reached
During exercise, central blood volume and the ∼10% increase in working muscles blood volume are supported by the muscle pump, promoting venous return and the redistribution of blood volume by vasoconstriction in the splanchnic area. However, with increasing blood flow requirements in working muscles and the skin, as body temperature increases, splanchnic vasoconstriction does not recruit enough blood to maintain the central blood volume level established at seated or supine rest. Central blood volume contributes to the level of sympathoexcitation established, as illustrated by the heart rate response. For example, at supine rest, heart rate may be 60 bpm and increase to 80 bpm when standing but decrease to 70 bpm during contraction of the legs. Muscle contractions increase sympathetic activity but the concomitant enhancement of central blood volume and central venous pressure by the muscle pump elicits a “paradoxical” reduction in sympathetic activity. Similarly, during running a reduced central blood volume attenuates central venous pressure and distension of the atria, as reflected by the plasma level of atrial natriuretic peptide (ANP).
Consequently, heart rate is higher during running than during rowing at a given exercise VO2. Furthermore, the lowest heart rate response to exercise is observed in the supine posture. This gravitational influence on heart rate is sustained during maximal exercise, as indicated by the lower value during rowing than during running, despite the larger VO2 max established by the larger active muscle mass during rowing. With the enhanced central blood volume following rowing training, heart rate at a given work load, including maximal effort, decreases.
9.3 Cardiac Output
The cardiac output depends on the volume of blood that the heart receives, or its preload. As the total capacitance of the vasculature is larger than the total blood volume, the distribution of blood volume is critical for maintenance of blood pressure and regional flow. Blood volume encompasses both volumes of red cells and plasma that change rapidly in response to exercise. For example, plasma volume is elevated by 20% following short-term training and it decreases during bed rest or during space flight. The enlargement of plasma volume following training, and the reduction in plasma volume when central blood volume remains elevated, demonstrate that central blood volume rather than total blood volume is the regulated variable. During exercise body weight is lost by sweating, but even after weight is restored by drinking, central blood volume remains reduced for many hours following exercise and plasma volume is expanded by further drinking as thirst is maintained. Central blood volume is reduced following exercise due to muscle oedema provoked by the combined effects of elevated perfusion pressure and muscle vasodilatation associated with exercise. Also, cutaneous vasodilatation induced by the elevated body temperature contributes to the attenuation of central blood volume during and after exercise. Even though body temperature normalises and muscle oedema is cleared rapidly post-exercise, muscle blood volume is maintained elevated for almost a day and that volume is recruited from the central circulation, as detected by DLCO (Fig. 8) and thoracic electrical impedance.
The reduced central blood volume following exercise is reflected in the levels of hormones that regulate fluid balance, including plasma vasopressin (alternatively named anti-diuretic hormone, ADH) and plasma ANP. Plasma vasopressin remains elevated while plasma ANP is low following exercise, and both these hormonal changes reduce urine production, resulting in positive fluid balance. It is less clear why the red cell volume increases in response to training. Bone marrow is stimulated by erythropoietin (EPO) released mainly from kidneys to produce haemoglobin. Exposure to high-altitude hypoxia increases haemoglobin production, but the acute increase in haematocrit reflects the loss of plasma volume. Intense exercise, in addition to the exercise-induced hypoxaemia stimulus for EPO production, may stimulate haemoglobin production via the sympathetically induced reduction in kidney blood flow, a mechanism similar to the low EPO production and associated anaemia observed in kidney diseases.
The increase in total haemoglobin is an important adaptation to training because VO2 max is related to red cell volume rather than to the haemoglobin concentration. In a seeming paradox, despite the increased red cell volume, elite endurance athletes often present low haemoglobin concentrations (or haematocrit), due to the training-induced enlarged plasma volume. Elite endurance athletes, e.g. rowers, may have a plasma and red cell volume of 4.8 and 3.11, respectively, compared to control reference values of 3.2 and 2.41, respectively, while, on an average, their haemoglobin concentration is larger than the normal healthy population (Fig. 3)
9.4 The Heart
As known from cardiac diseases, the heart adapts to the load it is exposed to, and that adaptation applies also to training. With endurance training the internal diameters of the heart enlarge with the highest values observed in professional bicyclists at 55 ml, vs 50 ml for untrained subjects. The heart of weightlifters is different because they develop high blood pressure during each maximal effort by a concomitantly performed Valsalva-like manoeuvre that stabilises the spine. To overcome the high blood pressure, the wall thickness of the heart's septum separating the left and right ventricle increases to 10 mm, while the internal diameters of the heart remain unchanged.
In some activities, such as rowing and kayaking, there is a combined demand for a large VO2 max, cardiac output and stroke volume, in addition to the need to overcome the high blood pressure, at the beginning of each rowing stroke. As a result, both the internal dimensions and wall thickness of the heart increase, and athletes of these sports possess the largest sports heart, with values for left ventricular mass of 330 g compared to 142 g for divers (Fig. 13).

Echocardiographic presentation of the heart in diastole of a world champion rower (a) and a control subject (b) of similar size. For the rower, the internal diameter of the left ventricle is 6.4 cm compared to 4.9 cm for the control subject. Similarly, the septum has a width of 1.3 and 0.8 cm, respectively and the posterior wall of the heart a width of 1.2 and 0.9 cm, respectively
About 7% of elite rowers have both a left ventricular wall thickness over the normal limit of 13 mm and an enlarged left ventricular cavity. In elite rowers the heart is so enlarged that myocardial perfusion becomes inhomogeneous, which, together with high vagal tone and low intrinsic heart rate (after combined vagal and sympathetic blockade), creates a complicated electrocardiographic presentation. As in skeletal muscles, training increases capillarization of the heart and, following detraining, the size of the heart returns to its control values.
While it is a problem to maintain an adequate preload to the heart during seated and especially upright exercise, there is little problem for the heart to pump the blood it is provided with. In contrast to the pain experienced in skeletal muscles during sustained exercise, healthy people do not complain of chest pain during exercise, indicating that myocardial oxygen demand does not limit cardiac output. Furthermore, it is energy-efficient for the heart to provide a large cardiac output.
Energy requirements of the heart depend on its rate and (systolic) pressure expressed as the rate–pressure product. Following rowing training the enlarged blood volume ensures filling of the heart and reduces sympathetic activity that attenuates heart rate by the same mechanism observed during supine and seated exercise. Furthermore, enhancement of central blood volume associated with semi- or supine posture, e.g. rowing, attenuates the pressure that arterial baroreceptors control during exercise, as exemplified by the blood pressure reduction when leg exercise is added to arm cranking. Cardiac output is elevated following endurance training with no additional strain on the heart (i.e. same rate–pressure product during maximal exercise), indicating that the enhanced blood flow to active muscles is provided by means of enhanced vascular dilatation (Fig. 14). Yet, the muscle pump cannot provide enough blood to the heart and sometimes, at exhaustion, a restraint on cardiac preload is illustrated by decreasing central venous pressure.

Red cells passing through a capillary, with substances released to dilate the vessels as oxygen is liberated
10 Stroke Volume
The stroke volume of elite endurance athletes is impressive (e.g. in rowers 195 vs 110 m for control subjects), but it is not limited by the capacity of the heart to encompass a large volume of blood. The problem upright humans face in increasing cardiac output during exercise is different from that experienced by quadrupedal animals. In puppies, work capacity increases following pericardiectomy that allows the heart to expand. In upright humans there is not a similar restraint on stroke volume as illustrated by the filling of the heart during supine exercise and by the ability of the heart to increase stroke volume in response to a volume overload. With administration of the plasma expander Macrodex® heart rate during maximal exercise decreases (183 vs 188 bpm) while stroke volume is enhanced by 10% (155–144 ml).
Following volume expansion the enhanced stroke volume confirms that the heart operates at the ascending part of the Starling curve when humans are upright. The size of the heart remains unchanged and it may decrease somewhat when heart rate exceeds 150 bpm, indicating that the heart propels the blood it is provided with. During exercise stroke volume increases by enhanced contractility and the systolic duration shortens to about one half, while the diastolic duration is further limited to one third of the resting value. Exercise tachycardia develops although plasma potassium increases, e.g. to 6–7 mM, signifying that sympathetic activation is needed not only to maintain function of the heart but also to clear plasma potassium during and after exercise.
The short diastolic interval during exercise may present a problem for the filling of the heart, as illustrated in patients with atrial fibrillation where cardiac output is compromised when heart rate exceeds 120 bpm. However, during exercise venous return is enhanced, and combined with the increased contractility of the heart, stroke volume is enhanced. Sympathetic activation is supported by a small increase in free plasma calcium released from albumin as pH decreases. Relaxation of the left ventricle may draw blood into the heart and this action is enhanced when the end-systolic volume is attenuated during vigorous exercise. Yet, the attenuated heart rate response to exercise following training is an advantageous adaptation for the filling of the heart.
10.1 Extreme Exercise
The ability of the heart to cope is less impressive during extreme exercise. Reports on horses running to death are not uncommon. It is also well-known that Pheidippides died after running the original marathon course to report to Athens the Greek victory over the Persians (battle of Marathon, 490 BC) (it may be noted that previously he had been fighting in the battle after running 2 × 250 km (!) in a round trip from Athens to Sparta to request support). While running to exhaustion, the heart of rats is dilated and the blood volume encompassed within the cavities is elevated by 50% and normalises only over days. That degree of exhaustion is further characterised by a state simulating thyroid insufficiency, as illustrated by rats forced to swim to exhaustion that reduce their spontaneous activity, eat more and increase body weight.
The extent that the post-exercise stress syndrome in rats relates to overtraining in humans is only a speculation, but long-distance events such as long-distance running or ultra-marathons provoke cardiac fatigue. Cardiac fatigue is presented primarily as reduced diastolic function of the heart. Both chronotropic and inotropic functions are affected by competitions lasting many hours, as illustrated by attenuated responses to sympathomimetic drugs, e.g. by beta-receptor downregulation or desensitization.
10.2 Cardiac Output
In 1920 cardiac output was measured during rowing in fixed-seat rowing boats by Liljestrand and Lindhard, showing values of 13–171 min−1 reaching up to ∼301 min− 1 in lightweight rowers during ergometer rowing. Cardiac output provides blood flow to tissues including working skeletal muscles and there is, on average, a 7.3:1 coupling between cardiac output and VO2 in untrained humans or 6.5:1 in trained endurance athletes. The largest reported VO2 max of 7.41 min−1 would thereby be expected to require a cardiac output of 491 min−1, and values of 401 min−1 are reported regularly. There are, however, large inter-individual variations in cardiac output both at rest and during exercise. At rest some variation relates to body size and cardiac output is expressed as cardiac index (3.51 min−1 m−2) in cardiology with body surface area based on height and weight (∼1.7 m2). During exercise cardiac output values usually range from 29 to 401 min−1, with the highest values achieved by those athletes who present also large body size: height exceeding 199 cm, body mass around 90 kg, body fat percent below 9% and VO2 max higher than 6.71 min (Table 1). Additionally, cardiac output varies according to the variation in genes, as demonstrated by the arg16gly polymorphism of the ß2-adrenergic receptor, and with haematocrit.
During exercise pulmonary VO2 increases in relation to metabolism in exercising muscles with oxygen uptake of non-exercising tissues being 0.41 min−1. Such observations underscore a tight coupling between cardiac output, regional blood flow and metabolism with some attenuation of blood flow at the highest workloads. Yet, it is not blood flow per se that is regulated but rather oxygen-carrying capacity defined as venous oxygen saturation because red cells liberate vasodilatating substances when they release oxygen (Fig. 14).
The postulate of an oxygen-dependent regulation of cardiac output and regional blood flow assumes that the heart provides the needed cardiac output, but that is not always the case. When the circulating blood volume is reduced, as exemplified by sweating during prolonged exercise or dehydration, cardiac output is smaller than expected according to haematocrit.
11 Blood Pressure
Arterial pressure has two roles in the regulation of blood flow to tissues. Arterial pressure is the key regulated circulatory variable, controlled beat by beat from the arterial baroreceptors that modulate peripheral resistance, as it provides perfusion pressure to the tissues and notably to the brain. Thus, cerebral blood flow is dependent on mean arterial pressure. At rest, variation in blood pressure is related to the cardiac cycle, resulting in a pulse pressure of approximately 45 mmHg. During exercise, however, blood pressure also varies with locomotor rhythm, e.g. the rowing cycle, because of the Valsalva-like manoeuvre performed at the catch of each stroke, giving rise to a “pulse pressure” of more than 100 mmHg. This means that the systolic pressure may approach 200 mmHg during maximal rowing. Regulation of arterial pressure by the arterial baroreceptors during rowing implies that their operating range is right-shifted and elevated by neural influence from central command and the muscle pressor reflex (Fig. 15).

The carotid baroreflex during arm (A), leg (L) and combined arm and leg exercise (A + L). At rest the actual pressure (arrow) corresponds to the maximum gain of the reflex (circle), while during exercise it may be positioned at a slightly lower estimated carotid sinus pressure (ECSP) suggesting that the baroreflex detects hypotension although blood pressure is elevated. HR, heart rate; MAP, mean arterial pressure
Two strategies may be applied to establish the elevated pressure that the baroreceptors are reset to control during exercise. Ideally, the set pressure can be established by an increase in cardiac output to compensate for the marked decrease in total peripheral resistance induced by exercise. However, if that is not possible because of strain on cardiac output either by a restricted preload or by an inability of the heart to produce the required cardiac output, mean arterial pressure is maintained by vasoconstriction not only to internal organs but also to working muscles and to the brain.
12 Regional Blood Flow
Skeletal muscle blood flow is modulated by deoxygenation of haemoglobin adjusting flow to metabolism, and that takes place despite the enhanced sympathetic activity during exercise (Fig. 14). Such sympatholysis depends also on other factors such as the elevated muscle temperature, potassium, nitric oxide and the arterial pyruvate/lactate ratio. However, muscle blood flow is not allowed to increase at the expense of blood pressure.
Priority for blood pressure regulation over regulation of flow is demonstrated when comparison is made between flow to a muscle working in isolation and together with other muscles. As an example, flow to working legs (∼101 min−1) is reduced when the arms are working intensely at the same time. Equally, arm blood flow (4.61 min−1 in untrained vs 6.41 min−1 in rowers) and oxygenation are larger during arm cranking than when arm cranking is performed together with high-intensity cycling exercise (Fig. 16).

Effect of adding arm exercise to leg exercise on leg oxygen uptake (VO2), leg blood flow (LBF), the leg arterial to venous oxygen difference [(a – v)O2 diff], and mean arterial pressure (MAP)
Skeletal muscles seldom receive the blood flow that their vasculature can handle. During exercise involving large muscle mass, especially, flow to the working muscles is reduced by ∼20–40% compared to the flow they receive during exercise involving small muscle mass, and this reduction is manifested primarily via sympathetically mediated enhanced vascular resistance. However, when leg exercise is added to arm cranking, blood pressure decreases and perfusion pressure to the arm accounts for approximately 50% of the reduction in arm blood flow.
13 Muscles
The final step in the oxygen transport chain to muscle is by diffusion. Capillaries are recruited when muscles are activated, suggesting that the capillary network is designed to provide the muscle with oxygen during exercise rather than at rest. In fact, arteries are not gas-impermeable and a considerable amount of gas exchange (oxygen uptake and elimination of CO2) takes place in vessels larger than the capillaries. Yet, during exercise muscle cells are provided with oxygen through diffusion from the capillaries. There is a coupling between VO2 max and capillary density, and more capillaries surround ST and FT muscle fibres. Typical values for the vastus lateralis muscle are 1.8 capillaries per fibre for untrained and 2.6 capillaries per fibre for trained endurance athletes, while the highest capillarization is observed in the internal intercostal muscles with six capillaries per fibre.
As with the lungs, the oxygen diffusion capacity of the muscles can be calculated. As in the lungs, the available oxygen diffusion capacity of the muscles is not always used. During moderate exercise capillary recruitment enhances the gas diffusion capacity, but during maximal exercise blood flow is limited, and thus gas diffusion is restrained. During arm-only exercise, gas diffusion values of the arm may be 20 ml min− 1 mmHg−1 and 50 ml min−1 mmHg− 1 in untrained and trained rowers, respectively, while the value decreases to 32 ml min−1 mmHg−1 when legwork is added to arm-only exercise.
14 Brain
Ultimately, it is the brain that limits performance. Central fatigue was described by the Italian physiologist A. Mosso (1904). Using a finger ergograph he demonstrated that fatigue becomes pronounced after a demanding mental task such as a lecture. Since the work of Mosso central fatigue has been described in a wide range of situations and, conversely, the enhanced performance associated with so-called diverting activities may be seen as alleviating that type of fatigue. The influence of diverting activities on muscle fatigue (Setchenov's phenomenon) refers to the observation that performance is enhanced when exercise with one muscle group (e.g. with one hand) is supplemented by activity with another muscle group. Similarly, if exercise is continued until exhaustion with the eyes closed, opening of the eyes enhances strength and work can be continued for some time.
Another delineation of a limitation to recruitment of muscles is the varying strength established when contractions are performed with either one or both legs (Fig. 17). The force developed during simultaneous contraction of the legs is less than the sum of strength developed during contractions of one leg at a time, and this “leg strength paradox” is modulated by training. Walking and running are characterised by alternate use of the legs, i.e. one leg is extending while the other leg is flexing, whereas when both legs are used concomitantly the developed strength is equal to, or exceeds, the sum of the strength that can be developed with one leg, e.g. during rowing.

Experimental setup and evaluation of leg strength
The ability to resist fatigue is enhanced by training and this effect is, moreover, so specific that to a large extent it must be ascribed to the enhanced ability to recruit motoneurons. Training of repeated one-legged contractions postpones, as expected, onset of fatigue during one-legged contractions whilst performance with both legs remains unchanged (Fig. 18). Conversely, training the extension of both legs benefits contractions involving both legs, whereas the performance of one leg is, surprisingly, unchanged and, under both circumstances, the electromyographic activity over the muscles decreases in parallel with force. Also, after maximal dynamic exercise, central fatigue is important, as demonstrated by electrical stimulation of the motor nerve that yields a greater power than that evoked by voluntary contractions. Finally, the consistent finding that a ∼25% increase in strength takes place without hypertrophy of the muscle fibres confirms that full recruitment of muscle fibres requires a central adaptation or a learning process.

Fatigure during repeated one- and two-legged contractions following one- or two-legged training
With the varying ability of the central nervous system to recruit the motoneurones, the pattern of muscle contractions developed during central fatigue is of interest. During partial neuromuscular blockade, two types of contractions can be delineated. With the use of an acetylcholine agonist drug (decamethonium or suxamethonium), muscle contractions become slow but enduring. Conversely, a non-depolarising neuromuscular blocking agent (e.g. tubocurarine) provokes contractions that maintain a high rate of rise of tension, but the developed force fades off rapidly. Because the two types of neuromuscular blocking agents affect selectively fast- and slow-twitch fibres respectively, the contraction pattern manifested during partial neuromuscular blockade provides some insight into the characteristics of the two main fibre types in humans. From that perspective, it seems that central fatigue affects the recruitment of slow- rather than fast-twitch muscle fibres as the contraction maintains its rate of rise of tension and, at the same time loses its endurance (Fig. 19).

Registration of force during repeated maximal voluntary handgrip contractions. As force decreases, the ability to maintain the contraction also becomes affected, while there is little reduction in the rate of rise of tension. Thus, at exhaustion, the contraction has duration of ∼0.3 s, while the normal maximal contraction lasts for some 2–3 s. During partial neuromuscular blockade by tubocuarine, rate of tension is similarly maintained, while the developed force fades off after 0.15 s to continue at a much lower level after ∼0.3 s. Conversely, after the use of decamethonium or suxamethonium, the rate of rise of tension is affected severely, but the developed force can be maintained for as long as desired
The central nervous system mechanisms responsible for central fatigue remain elusive. One consideration relevant to central fatigue is that stimulation of cortical areas provokes facilitation of reflexes over fast muscles and, conversely, inhibits reflexes involving slow muscles. In the following section, the cerebral metabolic response to exercise will be addressed from the perspective that central fatigue may be provoked by a limited provision of oxygen and substrate to relevant areas of the brain.
14.1 Cerebral Blood Flow During Exercise
While demonstration of central fatigue during different human activities may seem trivial, it has been a challenge to identify changes within the brain that correlate to hampered recruitment of the muscles. With physical activity regional cerebral blood flow (CBF) increases in activated areas of the brain but, for the brain as a whole, there is not much, if any, increase in CBF. A stable global CBF during exercise may imply that increased activity in one cerebral region is compensated by downregulation in other regions, and may explain why it is difficult to do more than one concentrated effort at a time. Furthermore, with the marked hyperventilation associated with maximal whole-body exercise such as rowing, the arterial carbon dioxide tension decreases and this reduces CBF. Together with the arterial hypoxaemia that can develop during intense exercise (arterial oxygen saturation decreasing from 97 to 90%), the reduction in CBF means that cerebral oxygenation decreases by ∼10%. Such a reduction in cerebral oxygenation is significant because it is of the same magnitude as seen during fainting and therefore a likely contributing factor to central fatigue.
14.2 Cerebral Energy Metabolism
Changes in brain metabolism are expressed by the ratio of oxygen to carbohydrate uptake of the brain, known as the cerebral metabolic ratio (MR), which at rest is close to 6. Cerebral metabolism depends primarily on oxidation of carbohydrate as the brain uptake of amino acids and free fatty acids is of little quantitative importance. During brain activation, however, MR decreases and in that respect physical exercise represents an extreme. The largest reduction in MR is established during exhaustive whole-body exercise while MR increases to ∼ 6.5 during anaesthesia (Fig. 20). With intense ergometer rowing-induced activation of the brain, MR decreases to values lower than 2, suggesting that less than 30% of the carbohydrate taken up by the brain is oxidised, or that up to ∼10 mmol glucose-equivalents need to be accounted for.

The cerebral metabolic ratio (brain uptake of oxygen relative to that of carbohydrate; glucose +1/2 lactate) during general anaesthesia, at rest and during various types of brain activation, including several types of exercise with ergometer rowing demonstrating the largest deviation from the resting value of 6
For evaluation of MR during exercise, lactate is important. During resting wakefulness the brain releases small amounts of lactate. In contrast, when exercise provokes a substantial increase in blood lactate, there is an uptake by the brain that is proportional to the arterial concentration. Seemingly, the lactate taken up by the brain is metabolised both by neurones and astrocytes as it is not accumulating within the cerebrospinal fluid or within the brain tissue. Breakdown of MR during exercise, however, does not depend on the lactate taken up by the brain. During prolonged exercise, there is only a modest or no increase in blood lactate, and accordingly, little lactate uptake by the brain. Yet MR decreases at exhaustion during prolonged exercise as during intense exercise of shorter duration.
14.3 Brain Glycogen Metabolism
During muscle contractions part of the energy is derived from glycogen breakdown, and during sustained muscle work fatigue presents when glycogen deposits are depleted. Similarly, the brain glycogen level decreases during neuronal activity. The human brain glycogen concentration is ∼ 6 mM (glucosyl units) in grey and white matter and as high as 13 mM in the hippocampus. Therefore the total glycogen content in the brain is estimated to ∼10 mmol. In the brain glycogen is confined primarily to the astrocytes, thus, the intracellular concentration of glycogen in the astrocytes may approach that of skeletal muscle, underpinning its potential importance as a fast accessible energy reservoir.
In analogy with skeletal muscle, central fatigue could be provoked when the brain glycogen level reaches a critically low level, and the “extra” and unaccounted for carbohydrate uptake by the brain could play a role during such energetic imbalance. By provoking such a crisis during exercise in hypoxia, isometric contractions become more affected than fast contractions, suggesting that recruitment of slow-twitch fibres is more vulnerable to central fatigue than that of fast-twitch muscle fibres. If so, central fatigue represents an exception to the normal “orderly recruitment” of muscle fibres where slow-twitch fibres are recruited before fast-twitch muscle fibres. Inability to maintain slow-twitch muscle fibre recruitment explains why fatigue manifests as a difficulty in preserving a smooth movement.
Brain temperature is also important, and work is stopped when its temperature reaches 40–41°C. Conversely, proper hydration allows for evaporation of water and control of temperature while training, which may be interpreted as preparation of the cardiorespiratory system to provide oxygen and substrates to working muscles so that work can be carried out without the brain being aware of the effort. Taken together, the cardiorespiratory system has the capacity to increase VO2 max by modulating each of the different steps in the transport system, with the exception of the pulmonary system, which is the only organ for which adaptation to endurance training has not been demonstrated.
15 Diving Response
A series of cardiovascular and respiratory adaptations permit mammalian air-breathers to perform extended aquatic dives. Among them is the “diving response”, consisting of selective vasoconstriction, which induces preferential shunting/blood shifts/thoracic filling, and bradycardia, which limits overall oxygen consumption and protects the heart and the brain from asphyxia. In humans, the response is induced by breath-holding while immersing the face in cold water. The magnitude of heart rate reduction is a measure of the diving response and is greater in diving than in terrestrial species. In seals, the response may reduce the heart rate by up to 80%. Humans, untrained in apneic diving, react with a heart rate reduction of between 20 and 30% from the resting level, during diving or apnea with face immersion in cold water. Trained human divers exhibit a bradycardia of 40–50%, which is in the range of the responses found in some semi-aquatic mammals. As the response in man reaches its maximum in water of 10°C, and considering that most apneic diving occurs in relatively warm waters, the practical application of the diving response in humans is questioned.
Even though some studies show an inverse relationship between heart rate and lung volume, it seems of importance for the magnitude of response whether the dive is performed during exhalation or inhalation. It is known that diving mammals dive in the expiratory position: seals, for example, exhale before diving in an apparent attempt to accentuate the bradycardic response. In humans however, this has not been demonstrated, and the issue remains elusive as breath-holding at residual volume tends to increase rather than decrease heart rate.
Even during the extreme physical exertion observed in elite freedivers, bradycardia persists, leading in some cases to a reduction in heart rate of more than 50%. Nevertheless, this mechanism delays the fall in alveolar and arterial PO2 and, thereby, the development of hypoxia in vital organs. In conclusion, it seems that the human diving response has an oxygen-conserving effect during exercise.
16 Altitude
Ascent to altitude entails exposure to a reduced barometric pressure and hence a reduced tension of oxygen in the inspired air that causes arterial hypoxemia, which provokes hyperventilation and leads to respiratory alkalosis. Breathlessness is conspicuous even during mild exercise. During incremental exercise the increase in exercise ventilation leads to the ventilatory capacity being reached at a relatively low rate of work. Exercise is then limited mainly by the function of the lungs. Lactacidemia also occurs, but the rise in blood lactate concentration is less than during maximal exercise at sea level.
During exercise a number of factors contribute to the accentuation of the hypoxemia, which causes tachycardia and an increase in cardiac output relative to oxygen uptake. The pulmonary pressure rises, thus contributing to ventilation–perfusion inequality, which in turn aggravates the hypoxemia further. Also, the conditions for oxygen transfer from the alveolar gas into the blood in the pulmonary capillaries are compromised as there is a diffusion limitation to gas transfer. Under these conditions the maximal work rate can also be compromised.
The acute effects of exposure to altitude are ameliorated by acclimation. The respiratory alkalosis is corrected by the secretion of alcalic urine while the desaturation is corrected by increased erythropoiesis that results in polycythemia. Also, a rise in the plasma concentration of 2–3 diphosphoglycerate partly reverses a displacement of the oxygen dissociation curve, which has been provoked by the prevailing blood gas changes. Following these adaptations submaximal exercise ventilation is further increased and the hypoxemia is slightly reduced as a consequence. The cardiac output returns to its normal level.
Under normal circumstances at an altitude of 1,500 m mild exercise can be performed without acclimation but acclimation increases the exercise capacity. At 3,000 m mild exercise initially provokes an exaggerated physiological response and the ability to perform killed tasks is impaired. In addition if the ascent is rapid acute mountain sickness and pulmonary oedema can develop.
17 Heat and Cold
During sustained heavy exercise blood flow is diverted to the skin at the expense of the active muscles and fluid is lost as sweat. These processes are rendered less effective by a hot, humid environment. Heart rate relative to oxygen uptake is then increased and maximal oxygen uptake is reduced. There is an associated rise in the deep body temperature, which causes reflex tachypnea and increased ventilation during submaximal exercise. The maximal exercise ventilation is little changed and the oxygen cost of different activities is relatively normal.
A cool environment has the converse effect of increasing the capacity for prolonged exercise. However, this beneficial effect does not extend to a very cold environment where shivering increases the oxygen cost of activities. Immersion in water has the additional consequence of heat loss by conduction. In these circumstances the temperature at which thermal equilibrium can be maintained is usually relatively high, around 20°C. Exercise, instead of offsetting the heat loss, may actually augment it by increasing the water flow across the skin. The tolerance of immersion depends critically on the quantity of subcutaneous fat, with fat individuals being much better protected than those who are lean. Endurance training increases tolerance to both heat and cold, but acclimation to heat and cold appear not to affect exercise performance.
18 Genetic Influence
Human form and function are dictated by the interaction of genes with environmental stimuli. Thus variation in environmental exposures will strongly influence phenotype. However, although a core genetic inheritance is common to all humans, small functional variations exist in these genes which dictate that individual (or racial) responses to environmental challenge will differ. Such genetic differences influence exercise performance through associated differences in anatomy and physiology and their response to training stimuli. The human gene map is growing in complexity as there are now in excess of 100 gene variants related to human performance (Rankinen et al. 2006).
For example, even though there is recent evidence suggesting that to some extent VO2 max is heritable, identification of specific genes is less convincing. Despite the initial focus on the angiotensin converting enzyme (ACE) gene and its possible association with endurance performance, it is now clear that many successful endurance athletes do not have the “endurance genotype”. Unfortunately, progress is slow in the field of genetics of fitness and performance, primarily because the number of laboratories and scientists focused on the role of genes and sequence variations in exercise-related traits continues to be quite limited.
19 Health
Several studies show that exercise continued throughout life attenuates the age-related deterioration of physiological functions and that it may favour life expectancy. In 1928 Pierre de Coubertin published a brochure entitled “La cure d'aviron” (health through rowing), and at 72 was still an active oarsman. Harvard University oarsmen have been reported to live three to six years longer than the American life expectancy in the early nineteenth century. The only contradictory finding of a somewhat lower average age at death of Cambridge University crew oarsmen than in a random group of people (67.1 vs 67.4 years) from the same time period can be attributed to their engagement in the First World War. Nevertheless, having considered the interference of world affairs, the death rate from cardiovascular disease is lower in athletes than in a random control group of people, and such observations are now available from participants in a large range of activities. Thus, in perspective studies there is a direct relationship between lifespan and physical activity, including a reduced prevalence of certain types of cancer, such as colon cancer. An early observation by Morris was that the conductors of London's double-decker buses lived longer than the drivers, and similar observations made in Finland show a progressive increase in the lifespan of international championship participants depending on the type of sport, ranging from power sports to ball games and endurance events. It may be, however, that such comparisons reflect evaluations of participants with different genetic make-up. In that light a specific polymorphism in the gene encoding the angiontensin-converting enzyme resulting in low-range values of angiotensin in plasma is over-represented in successful competitive Australian rowers. Also, this “insertion” allele is associated with elite endurance performance among high-altitude mountaineers. On the other hand, the “deletion” allele, producing relatively high levels of angiotensin, has been identified as a risk factor for the development of cardiovascular diseases.
19.1 Height
With ageing, body height decreases by approximately 2 cm due to the progressive curvature of the spine and osteoporotic collapse of the vertebrae. This decrease in body height develops from the age of ∼40 to ∼60 years even in people who continue to be active, and thus, there is no reported difference in height between old athletes and sedentary men. Probably reflecting the increasing height of the population, older active individuals are shorter than their young counterparts, but there is a lack of longitudinal studies evaluating the effect of regular exercise on osteoporosis and the age-related decline of the length of the spinal column.
19.2 Osteoporosis
Peak bone mass is established in both men and women around the age of 30 years and subsequently declines by ∼0.5% per year in men and by ∼1% per year in women. The decline in bone mass observed throughout life contributes to the elevated incidence of osteoporotic fractures. Physical activity, especially weight-bearing activity, is an essential requirement for maintained bone mass.
19.3 Body Mass
As body mass increases with age, probably due to reduced physical activity, the risks of falling, fracture and limb disability, as well as morbidity from major chronic diseases and even mortality, are increased. Regular exercise of sufficient volume and intensity provides for the maintenance of low body mass in the elderly. There is greater prevalence of obesity with age, which is linked with numerous chronic diseases including type II diabetes, hypertension and cardiac disease, and thereby to mortality. Even though body fat is higher in old compared with that in young athletes (e.g. rowers, 18% vs 12%, Table 1), it is similar to that of young sedentary men and lower than in sedentary men matched for age and body size (18% vs 23%, Table 2).
Ageing is also characterised by a decrease in fat-free mass. In spite of lower fat-free mass, elderly individuals who engage in regular physical activity have a larger fat-free mass than elderly sedentary men. In particular, older oarsmen have a larger fat-free mass than habitual runners, and maintain a fat-free mass similar to young sedentary men. This finding reflects the specific adaptive response of each exercise modality. Thus, regular physical activity contributes to attenuate the age-related loss of fat-free mass and reduces obesity and, therefore, morbidity associated with chronic diseases.
19.4 Aerobic Capacity
Elderly individuals who engage in regular physical activity have a slower decline in VO2 max compared with sedentary elderly. For example, elderly oarsmen have a lower VO2 max compared with young oarsmen (3.0 vs 4.11 min−1, Table 1) but their VO2 max is larger than in sedentary men matched for age and body size (3.0 vs 2.21 min−1, Fig. 1) and similar to young sedentary men. Thus, regular physical activity continued in advanced age slows the ageing-related decline in aerobic fitness.
The decay of cardiorespiratory function with age is also represented by a greater increase in blood lactate at a given submaximal exercise intensity. However, elderly individuals who engage in regular physical activity achieve a given lactate level at a higher exercise intensity, expressed by VO2, compared with sedentary men matched for age and body size (e.g. for older oarsman, blood lactate of 4 mmol l−1 at 2.6 vs 1.8l min−1). Also, elderly competitive athletes possess a VO2 similar to young athletes matched for performance time (Table 2, older rowers).
In addition, V Emax decreases with age, which might be explained by loss of elasticity of the lung tissue and the chest wall, both of which demand more work associated with breathing. While regular physical activity does not prevent the age-associated decrease in V Emax (117 vs 162l min−1, in elderly compared with young individuals) physical activity on a daily basis is associated with an attenuated age-related decrease in respiratory function when matched for age and body size (117 vs 74l min−1, Table 1). Interestingly, elderly oarsmen exhibit a V Emax similar to young oarsmen matched for rowing performance and body size (121 vs 124l min−1, Table 2).
19.5 Heart Rate
HR is a simple but informative parameter both in clinical and physiological exercise settings. As HR increases nearly linearly with work rate to maximal exercise, this relationship is used widely to predict work capacity, or VO2max, from values derived at a submaximal work intensity. HRmax decreases progressively with ageing and is lower in older than in young individuals who engage in regular physical activity and sedentary men (176 vs 201 and 198 beats min−1 respectively, Table 1, Fig. 1). However, regular physical activity plays a role in blunting the age-associated decline of HRmax, as demonstrated by higher HRmax in older oarsmen than older sedentary men (166 bpm).
As with the reduction in VO2 max, HR at a submaximal workrate increases with age, as indicated by the achievement of a given blood lactate level at a lower HR in older oarsmen compared with young oarsmen with similar rowing performance (149 vs 165 bpm at 4 mmol 1−1). On the other hand, an age-associated decrease in HR at submaximal work intensity is modified by habitual exercise, as demonstrated by the achievement of a blood lactate of 4 mmol 1−1 at a higher HR in older oarsmen than in older sedentary men (163 vs 157 beats min−1, Fig. 2).
The seated exercise position elicits a lower HR response than running at comparable exercise intensity both in older and young men and women. This posture-induced reduction in the HR response to seated exercise is associated with an expanded central blood volume, as shown by a higher plasma atrial natriuretic peptide (ANP; 131 vs 100pg ml−1, released by atrial stretch) and central venous pressure (CVP; 3.3 vs −5.2 mmHg) compared with running. Consequently, when HRmax predicted by the traditional equation that 220 minus age is applied for estimating exercise intensity relative to VO2 max, the flat relationship between HR and VO2 during seated exercise should be taken into consideration in order not to prescribe too large a workload.
19.6 Structure and Function of the Heart
The weight of the heart increases by an average of 1 g year−1 in men and 1.5 g year−1 in women, reflecting some degree of left ventricular hypertrophy, even in subjects apparently free of hypertension. Regular physical activity induces favourable alterations of the heart, as shown by a cardiac hypertrophy in older oarsmen compared with sedentary men (septum thickness, 11 vs 9 mm; posterior wall thickness, 9 vs 8 mm; left ventricular mass, 227 vs 205 g;left ventricular mass index, 127 vs 103 g m−2). In addition, regular physical activity continued into advanced age offers protection against deterioration of cardiovascular function with age, as demonstrated by enhanced systolic function (left ventricular shortening fraction, 0.45 vs 0.36).
With regard to cardiac function and systemic haemodynamics, the maximal cardiac output declines with advancing age because of a decrease in stroke volume. The oxygen pulse (VO2/HR) is used as an index of stroke volume. The age-related decline in stroke volume is not completely averted by regular physical activity demonstrated by a lower value in older than in young oarsmen, but this reduction is attenuated, as demonstrated by a higher oxygen pulse in older oarsmen than in sedentary men matched for age and body size (17 vs. 12 mL beat−1 at a blood lactate level of 4 mmol L−1; 17 vs. 14 mL beat−1 at maximal effort, Table 1, Figs. 1 and 2). Additionally, there is similar oxygen pulse between older and young oarsmen with similar 2,000 m rowing performance time.
19.7 Blood Lactate Concentration
Ageing reduces muscle oxidative capacity, capillarisation, and oxidative enzymes activity: changes that are reflected in the reduced blood lactate concentration immediately after maximal exercise in sedentary men and even in vigorously trained endurance runners and cyclists. Thus, the age-associated reduction in maximal blood lactate is not prevented by lifelong physical activity, as shown by lower values in older compared with young oarsmen and sedentary men (8.3 vs 10.9 and 10.0 mmol l−1, Table 1, Fig. 2). Nevertheless, the higher maximal blood lactate values in elderly active individuals compared with sedentary men matched for age and body size (7.9 mmol l−1) suggest that physical activity carried out throughout life attenuates the age-associated decline in skeletal muscle adaptation to physical training.
19.8 Risk Factors of Coronary Artery Disease
Dyslipidoproteinaemia, i.e. elevated concentrations of total cholesterol (TC), triglycerides (TG), low-density lipoprotein cholesterol (LDL-C), and a reduced concentration of high-density lipoprotein cholesterol (HDL-C) in plasma, are risk factors for coronary artery disease. The LDL-C is implicated in plaque formation in blood vessels, while the HDL-C is involved in plaque removal. Thus, the ratio of LDL-C to HDL-C, or that of TC to HDL-C, i.e. the atherosclerosis index, is applied to evaluate individual risks for coronary artery disease. Also, ageing is considered as one of the secondary risk factors for development of coronary artery disease. Lifelong physical activity fosters attenuation in risk factors for coronary artery disease in aged men, which is grounded on a lower atherosclerosis index in elderly active individuals (LDL-C/HDL-C, 1.7; TC/HDL-C, 3.1) compared with both older and young sedentary men (LDL-C/HDL-C, 2.1 and 2.1; TC/HDL-C, 3.6 and 3.5, respectively).
Ageing is associated with a progressive increase in body fat, which results from a declining level of physical activity. Leptin is a protein produced by “the obesity (ob) gene” and its secretion from adipose tissue to the circulation is considered to signal the brain with regard to the size and the nutritional status of adipose tissue, a signal that seems to be lost or inhibited in people who become very fat. Physical activity continued into advanced age has a protective effect on the age-associated increase in fat mass and serum concentration of leptin, as reflected by the lower leptin values in older oarsmen compared with older sedentary men matched for body size (fat mass, 13 vs 16 kg, serum leptin, 2.5 vs 4.1 ng ml−1. Thus, the serum concentration of leptin is correlated to percent body fat (r = 0.60) and cross-sectional area of leg fat.
19.9 Risk Factors of Diabetes Mellitus
Diabetes is a disorder of carbohydrate metabolism characterised by hyperglycaemia, and the incidence of type II diabetes increases with advancing age. Insulin is an important hormone that reduces glucose circulating in the blood by facilitating its transport into the cells. There is an inverse relationship between the level of habitual physical activity and the incidence of type II diabetes. Regular physical activity benefits the insulin-stimulated whole-body and skeletal muscle glucose uptake as shown by a lower fasting plasma glucose and insulin in older oarsmen compared with older sedentary men (glucose, 5.3 vs 5.8 mmol l−1, insulin, 6.9 vs 8.7 pmol l−1 respectively).
19.10 Skeletal Muscle Structure and Function
As the rate of protein synthesis declines with ageing, skeletal muscle size decreases, as shown by the reductions in arm, leg, and limb mass even in endurance-trained elderly. Regular physical activity has a profound effect in preventing muscle wasting in older active men, demonstrated by a larger area of leg extensor muscle than in older sedentary men (78 vs 68 cm2). This favourable influence of regular exercise on muscular morphology may be attributed to an increase of type I fibre area in particular, while aging has a detrimental effect on type II muscle fibres.
The skeletal muscle structural changes associated with ageing are also reflected in the arm, leg, and back strength declining at an overall rate of 8% per decade, starting in the third decade of life. The ability of the leg extensor muscles to develop power is of special relevance as it enables the elderly to accomplish daily tasks. The loss of leg extension power increases the risks of falls and limb disability. The age-related decline in muscle power is not prevented by regular physical activity, as shown by the lower bilateral dynamic leg extension power in older than in young oarsmen (1,675 vs 2,358 W) but older oarsmen possess a higher power than older and young sedentary men (1,261 and 1,485 W, respectively). The parallel deterioration of skeletal muscular size and function is supported by a relationship between leg extension power and extensor muscle area (59–89 cm2, r = 0.74). Therefore, regular physical activity has a profound effect in preventing muscle wasting and weakness.
19.11 Benefit of Regular Physical Activity
Regular physical activity contributes to proper body composition, aerobic fitness and healthy cardiovascular and skeletal muscular conditions, and has preventive effects on indices for the development of coronary artery disease and type II diabetes.
References
American College of Sports Medicine (1998b) Exercise and physical activity for older adults. Med Sci Sports Exercise 30:992–1008
Blair SN, Brodney S (1999) Effects of physical inactivity and obesity on morbidity and mortality: current evidence and research issues. Med Sci Sports Exercise 31:S646–S662
Dalsgaard MK (2006) Fuelling cerebral activity in exercising man. J Cereb Blood Flow Metab 26:731–750
Drinkwater BL (1994) Physical activity, fitness and osteoporosis. In: Bouchard C, Shephard RJ, Stephens T (eds) Physical Activity, Fitness, and Health, International Proceedings and Consensus Statement. Human Kinetics Publishers, Champaign IL, pp. 724–736
Fritsch W (2000) Rowing in daily life. In: Fritsch W (ed) Rowing. Meyer & Meyer Sport Ltd, Oxford, pp. 148–183
Ide K, Secher NH (2000) Cerebral blood flow and metabolism during exercise. Prog Neurobiol 61:397–414
Mosso A (1904) Fatigue. Swan Sonnenschein, London
Nybo L, Secher NH (2004) Cerebral perturbations provoked by prolonged exercise. Prog Neurobiol 72:223–261
Secher NH (1983) The physiology of rowing. J Sports Sci 1:23–53
Secher NH, Volianitis S (2006) Are the arms and legs in competition for cardiac output? Med Sci Sports Exercise 38:1797–1803
Volianitis S, McConnell AK, Koutedakis Y, McNaughton L, Backx K, Jones DA (2001) Inspiratory muscle training improves rowing performance. Med Sci Sports Exercise 33:803–809
Wagner PD, Hoppler H, Saltin B (1977) Determinants of maximal oxygen uptake. In: Crystal RG, West JB (eds) The Lung. Lippincott-Raven, Philadelphia, pp 2033–2041
Yoshiga CC, Higuchi M (2003b) Rowing performance of female and male rowers. Scand J Med Sci Sports 13:317–321
Further reading
Pedersen BK, Saltin B (2006) Evidence for prescribing exercise as therapy in chronic disease. Scand J Med Sci Sports 16(Suppl 1):3–63
Rankinen T, Bray MS, Hagberg JM, Pérusse L, Roth SM, Wolfarth B, Bouchard C (2006) The human gene map for performance and health-related fitness phenotypes: the 2005 update. Med Sci Sports Exercise 38:863–1888
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2009 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Volianitis, S., Secher, N.H. (2009). Human Exercise Physiology. In: Glass, M., Wood, S. (eds) Cardio-Respiratory Control in Vertebrates. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-93985-6_20
Download citation
DOI: https://doi.org/10.1007/978-3-540-93985-6_20
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-540-93984-9
Online ISBN: 978-3-540-93985-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)