
Abstract
A number of protective immune functions decline with age along with physiological and anatomical changes, contributing to the increased susceptibility of older adults to infectious diseases and suboptimal protective immune responses to vaccination. Influenza vaccination is the most cost-effective strategy to prevent complications from influenza viral infections; however, the immunogenicity and effectiveness of currently licensed vaccines in the United States is about 30–50% in preventing complications arising from influenza and preventing death from all causes during winter months in older adults. Hence, it is crucial to understand the molecular mechanisms that lead to immune dysfunction as a function of age so that appropriate strategies can be developed to enhance the disease resistance and immunogenicity of preventive vaccines, including influenza vaccines, for the older adult population.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- West Nile Virus
- Severe Acute Respiratory Syndrome
- Influenza Vaccine
- Influenza Vaccination
- Influenza Viral Infection
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
A dramatic increase in the older adult population is occurring globally due to improved sanitation, preventive vaccination, development of effective antimicrobial drugs, and advances in medical sciences. This growth of the older adult population is having a major impact on healthcare, social services, and public health. In the year 2004, older adults accounted for 12.4% of the total population and required $531.5 billion in primary healthcare costs (Hartman et al. 2008). This represented almost 34% of all healthcare spending, as the cost of providing healthcare for an older adult aged 65 or above is 3–5 times greater than the cost for a younger adult. The older adult population in the USA is projected to almost double by 2030 2007(AoA). A decline in immune function leading to increased susceptibility to infectious diseases and poor adaptive immune response to vaccination is a key characteristic of aging (Miller 1996, 1997). For example, increased colonization of bacteria and yeast on the skin and mucosal surfaces, respiratory, and urogenital tracts, increased susceptibility to viral infections, and reactivation of latent viral and bacterial infections are all well documented (Gardner et al. 2006; Worley 2006; Ely et al. 2007; Htwe et al. 2007; Kovaiou et al. 2007; Simmons et al. 2007; van Duin and Shaw 2007). In general, infectious diseases such as severe acute respiratory syndrome (SARS), West Nile virus, respiratory syncytial virus (RSV), influenza, and pneumococcal infections tend to be more severe (with complications), often resulting in unfavorable outcomes among older adults when compared to those in healthy adults. In addition, the efficacy of preventive vaccines against bacterial and viral targets declines dramatically with the progression of age among older adults, clearly indicating that the dysregulated immune status referred to as “immunosenscence” is the consequence of altered physiological and anatomical functions (Ginaldi et al. 2001; Aw et al. 2007). Hence, in this review, we will address the status of innate and adaptive immune functions in aging, the current state of influenza vaccines and their efficacy in older adults, and strategies that need to be considered to protect them against influenza.
2 Immune Status in Aging
2.1 Innate Immunity
Innate immunity was considered “nonspecific” and received secondary importance when compared to antigen-specific adaptive immune functions until the late 1990s. However, with the discovery of Toll-like receptors (TLRs), innate immunity is now recognized to be crucial to the survival of species. Therefore, understanding innate immunity can offer newer insights into the development of novel immunomodulators and antimicrobials (Hoffmann et al. 1999; Medzhitov and Janeway 2000a,b; Imler and Hoffmann 2001). Since the discovery of TLRs, several other pathogen-sensing receptor families have been identified over the last decade (Bingle and Craven 2002; Kang et al. 2002; Lu et al. 2002; Holmskov et al. 2003; Yoneyama et al. 2004; Martinon and Tschopp 2005; Ting and Davis 2005; Brown 2006; Takaoka et al. 2007). These families evolved to overcome microbial strategies and their metabolic needs in order to eliminate them. TLRs are expressed either as soluble molecules on the cell membrane or in vesicular compartments, or in the cytosol, as shown in Fig. 1. These pathogen sensors recognize structural components of pathogens and activate signal transduction cascades, leading to gene transcription with several outcomes, such as activation of antibacterial and antiviral defenses, secretion of proinflammatory cytokines and chemokines, tissue repair in the event of damage, and activation of adaptive immune responses. In most cases, the precise structure or sequence of the pathogen signature that stimulate the innate immune receptors is not well defined.

Pathogen sensors of the innate immune system. Several families of pathogen-sensing receptors that recognize conserved molecular signatures of pathogens are localized in various compartments within the cell as well as in body fluids. Engagement of these receptors leads to the activation of the innate immune system and the elimination of the pathogens
The dynamic barrier against infectious diseases, the epithelial lining of skin, gastrointestinal, respiratory, and urogenital systems, prevents the colonization and entry of potential pathogens into the body’s interior, which is sterile (Ganz 2002). These epithelial cells express several pattern recognition receptors and, upon recognition of the molecular signatures of pathogens, secrete antimicrobial substances that aid in the destruction of pathogenic microbes (Ganz et al. 1992; Schittek et al. 2001; Zanetti 2004). In addition to epithelial cell turnover that reduces the microbial load, mucosal secretions of respiratory, urogenital, and gastrointestinal tracts have antibacterial substances that also facilitate the elimination of colonization. Similarly, the antibacterial components of sweat and skin secretions reduce colonization of microbial load (Schittek et al. 2001; Zanetti 2004). Limited information is available on the status and functionality of skin, lung, and other mucosal epithelial layers among older adults. The epithelial cell turnover rate in skin slows down among the older adults, with a reduced secretion of sweat and sebum resulting in dryness and increased microbial colonization, especially with Pseudomonas and Proteus species (Laube 2004). These altered physiological states may contribute to delayed skin wound healing (Thomas 2001; Reed et al. 2003; Sorensen et al. 2003; Gosain and DiPietro 2004; Laube 2004). Altered physiological and anatomical changes in the lungs also contribute to poor innate immunity, thereby increasing microbial colonization and the incidence of pneumonia (Meyer 2001). These changes include reduced elasticity and function of lung muscles, reduced mucociliary clearance rates, decreased oropharyngeal clearance of bacteria, decreased phagocytic activity of alveolar macrophages, and decreased mucosal secretions (Meyer et al. 1996; Meyer 2004, 2005). Similarly, there is increased localization of Candida species on the oral and urogenital mucosal surfaces with age among older adults (Shay and Ship 1995; Sobel 1997). In the case of influenza infection, no data are available on the status of innate immune responses at the epithelial barriers in aging. Hence, detailed studies addressing the statuses of pathogen-sensing mechanisms with age are required and will enable us to come up with the strategies to reduce microbial load at epithelial surfaces and to enhance disease resistance.
2.2 Pathogen Sensing and Antigen-Presenting Cells
The primary role of innate immunity is to prevent the entry of pathogens into the tissues; however, a number of factors such as dose of infecting pathogen and the immune and nutritional status of the individual determine if innate immunity is able to prevent colonization and infection. Once pathogens overcome the epithelial defenses and gain access into tissues, myeloid lineages of hemopoietic stem cells from bone marrow, namely tissue-resident macrophages and dendritic cells, recognize the pathogens. Innate immune receptors, either directly or through scavenger receptors or pathogens bound to soluble innate immune receptors, initiate phagocytosis and an inflammatory response. These interactions lead to the secretion of proinflammatory cytokines such as IL-6, TNF-α, and IL-8, which attract neutrophils and natural killer cells to the site of infection, thus creating an optimal priming environment to initiate an adaptive immune response. Dendritic cells (DCs) capture antigens from pathogens, mature, differentiate, and migrate to regional draining lymph nodes to stimulate antigen-specific T and B cells, the lymphoid lineages that originate from hemopoietic stem cells. Following antigen-specific clonal expansion of B and T cells, the invading pathogen is or the pathogen-infected cells are removed by specific antibody and T cells. Tissue-resident macrophages play a major role in pathogen sensing, elimination, and tissue repair. We have demonstrated previously that the expression and function of TLRs on peritoneal as well as splenic macrophages decline with age using a murine model or peripheral blood mononuclear cells (PBMCs) from humans (Renshaw et al. 2002; van Duin et al. 2007b; van Duin and Shaw 2007). These findings are consistent with the previous observations that macrophage function declines with age, although the molecular mechanisms were not clear (Plowden et al. 2004a,b; Sebastian et al. 2005). Not only does macrophage function decline with age, but so does their ability to process and present antigens, secrete proinflammatory cytokines and chemokines, provide costimulatory signals, and migrate to the site of infection, as documented in aged animal models (Plowden et al. 2004a,b). Although an age-related decline in the acute proinflammatory response of monocytes has been identified, other studies have demonstrated increased levels of proinflammatory cytokines in serum and in culture supernatants of in vitro stimulated monocyte cultures from healthy older adults compared to younger adults. These observations led Franceschi and colleagues to coin the term “inflammaging” indicating a low-grade chronic inflammatory state as a hallmark of aging (Franceschi et al. 2000; Franceschi 2007) and increased risk for adverse changes in health in older adults. This would predict high levels of proinflammatory cytokines in frail older adults, but just the opposite has been found; low levels of the cytokines have been associated with frailty. Differences in the observations may be accounted for based on the type (polyclonal vs. antigen- or ligand-specific) and duration (acute vs. chronic) of stimulus (van den Biggelaar et al. 2004). Although additional studies need to clarify the observed differences in the secretion of proinflammatory cytokines between aged animal models, healthy older adults and frail older adults, it is clear that there are alterations in pro- and anti-inflammatory cytokine secretion and their balance with aging (Alberti et al. 2006). These alterations will affect both innate and adaptive immune functions (Fig. 2). In addition, the migration of antigen-bearing DCs is severely affected in aged animals, indicating that the priming environment for adaptive immune responses is suboptimal (Linton et al. 2005). Although careful studies are yet to be performed, Langerhans cells in skin appear to decline in numbers with age, and their function also declines with age (Meyerson 1966; Laube 2004). In contrast, bone marrow-derived DCs generated with a cocktail of cytokines from aged animals or humans are found to be as effective as those generated from their younger counterparts in recalling memory T cell responses. This would suggest that in vitro generation bypasses age-related defects that are seen with ex vivo DCs (Lung et al. 2000; Tesar et al. 2006). However, primary CD4 T-lymphocyte responses remain impaired in spite of normal DC function, suggesting increased antigen and costimulation thresholds of aged, naïve T-lymphocytes, consistent with our earlier published data (Haynes et al. 2000; Sambhara et al. 2001; van Duin et al. 2007a). Unpublished findings from our laboratory indicate a substantial delay in the mobilization of DCs and macrophages into lungs and regional draining lymph nodes following infection with influenza virus in aged compared to younger animals, suggesting an altered microenvironment. A decline in the expression of pathogen sensors (specifically TLRs) and the secretion of cytokines and chemokines was also observed that may influence the migration, activation, differentiation, and function of macrophages and dendritic cells. Indeed, a recently published study demonstrated poor induction of costimulatory molecules on monocytes of older adults following TLR stimulation, consistent with our observations in the murine model (Renshaw et al. 2002; van Duin and Shaw 2007). Using adjuvants that stimulate the innate immune system, providing costimulation, or supplementing with a cocktail of cytokines along with antigen at the time of immunization significantly improved adaptive immune responses by stimulating antigen-presenting cells (APCs) (Sambhara et al. 1998, 2001; Haynes et al. 2004). Hence, it is logical to formulate vaccines for older adults with adjuvants to induce an optimal priming environment for adaptive immune responses.

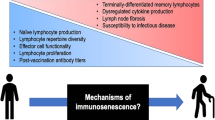
Immune dysfunction in older adults. The characteristics of the alterations observed in both the innate and adaptive immune compartments
2.3 Adaptive Immunity
Hematopoietic stem cells (HSC) in the bone marrow give rise to both myeloid- and lymphoid-committed stem cells. While the myeloid lineage gives rise to monocytes, macrophages, and dendritic cells, lymphoid-committed stem cells give rise to T lymphocytes and B lymphocytes which go through “education and selection” in the thymus (in the case of T cells) and bone marrow (in the case of B cells), a process that removes potentially autoreactive clones. These cells are further educated in the periphery to be tolerant to self-antigens. Although there is some indirect evidence that HSC numbers decline with age, detailed studies are yet to be done to determine if HSC numbers, function or migration alter with age (Wang et al. 1995; Lamberts et al. 1997; de Haan and Van Zant 1999). Changes in the cellularity of bone marrow with aging have been clearly documented, and may alter the local cytokine milieu, thus affecting the proliferation, differentiation, and seeding of secondary lymphoid organs by lineage-specific stem cells (Liang et al. 2005).
2.3.1 Humoral Immunity in Aging
Antigen-specific adaptive immune responses against influenza virus infection or vaccination are mediated by B lymphocytes and T lymphocytes; both contribute to humoral and cellular immunity to influenza. T helper cells secrete cytokines for B lymphocyte differentiation and class switching. Following the recognition of antigens with their surface immunoglobulin receptors, B lymphocytes undergo differentiation to become plasma cells that secrete antibody. Antibodies against the major surface glycoprotein of influenza viruses, the hemagglutinin (HA), neutralize the virus by binding to conformational determinants on HA, and prevent infection. Antibodies directed against the second major surface glycoprotein, the neuraminidase (NA), can limit virus release from an infected cell and can therefore reduce virus replication. The functionality of anti-HA antibodies is usually determined by the hemagglutination-inhibition (HAI) test, and in some cases by virus-neutralization tests. A HAI titer of ≥1:40 is correlated with a 50% protection rate in a population against influenza viral infections (Wood et al. 1997). Due to the high mutation rate of this RNA virus and the selection pressure of pre-existing antibody in humans that acts on circulating viruses, influenza viruses accumulate mutations in HA and NA genes, leading to antigenic drift, which requires that the strains of influenza contained in the vaccine must be updated every year to antigenically match the circulating strains. In general, it is known that humoral immune responses induced by influenza vaccination decline with age. However, humoral immune responses as measured by HAI titers in community-dwelling “healthy older adults” and centenarians are similar to those observed in younger adults, indicating that aging alone does not affect antibody responses against influenza vaccination. Other contributing factors to the decline in antibody responses include comorbid conditions such as chronic diseases and frailty, as well as poor nutrition, stress, and limited physical activity. Pre-existing humoral immunity due to annual vaccination of older adults does not appear to impact the antibody responses to subsequent vaccinations and does not explain the poor vaccine efficacy. One possibility is that the quality and duration, rather than the magnitude of the antibody response, may be affected; however, results from a recent study indicate that this may not be the case (de Bruijn et al. 1999; Gardner et al. 2001; Iorio et al. 2007). Hence, additional markers of the immune response may be needed to predict vaccine efficacy in the older adult population. Earlier studies from our laboratory have shown that serum antibody titers did not correlate with the susceptibility to influenza virus infection among older adults, suggesting that both antibodies and cellular immunity contribute to clinical protection against influenza illness. Although the antibody responses are strain-specific within a subtype, they do provide cross-protection against viruses of the same subtype via antibody-dependent cell-mediated cytotoxicity carried out by NK cells or macrophages.
2.3.2 Cellular Immunity in Aging
Unlike B lymphocytes, T lymphocytes recognize peptide fragments derived from the antigens that are presented with major histocompatibility complex molecules by professional antigen-presenting cells such as dendritic cells. T lymphocytes consist of CD4 T helper cells and CD8 cytotoxic T cells. While CD4 T lymphocytes recognize peptides that are processed from exogenous antigens (e.g., killed virus) presented with class II MHC molecules, CD8 T lymphocytes recognize peptide fragments derived from endogenous antigens (e.g., peptides derived from virus replicating inside the cell) that are presented with class I MHC molecules. Depending on the pattern of cytokines they secrete, CD4 T lymphocytes are further classified as T helper 1 (Th1), T helper 2 (Th2), T helper 3 (Th3), and T helper 17 (Th17) cells. T lymphocytes recognize peptide fragments derived from both surface glycoproteins and internal proteins. While surface glycoproteins (HA and NA) vary due to antigenic drift or shift, the internal proteins, namely the nucleoprotein, matrix protein and others, are fairly conserved within the subtype of influenza viruses. It has been shown that although T lymphocytes will not prevent infection, cytotoxic T lymphocytes kill virus-infected cells and aid in viral clearance, thus contributing to clinical protection against influenza illness (Yap et al. 1978). Hence, the activation of both CD4 and CD8 T lymphocytes will provide cross-protection against variant viruses within a subtype. Virus infection induces robust T lymphocyte responses, which persist for a very long time and provide cross-protection in mice. The magnitude and durability of T lymphocyte responses depend on the route of infection/immunization, whether or not the vaccine is formulated to induce or recall especially CD8 T lymphocyte responses. The current inactivated split-virus influenza vaccines provide only exogenous antigens for stimulation of T lymphocytes and thus are poor inducers of CD8 T lymphocyte responses. Activating or recalling CD8 T lymphocyte responses by formulating vaccines with adjuvants which will stimulate antigen-presenting cells creates an optimal priming environment and activates T lymphocytes to provide broader protection against serologically distinct viruses. CD4 T helper cells provide growth factors for B and CD8 T lymphocytes, thereby occupying a central role in the induction of humoral and cellular immune responses. Th1 and Th2 cells were defined based on the secretion of IFN-γ. While Th1 cells secrete IFN-γ following stimulation by IL-12, Th2 cells stimulated by IL-4 secrete IL-4, IL-5, and IL-13. The decline of naïve T cells in the repertoire due to thymic involution and accumulation of dysfunctional memory T cells is well established, but the mechanism for these observations goes beyond that which can be explained by thymic involution alone. Interleukin 7 appears to play an important role in T cell survival in thymic recombination events, and in expanding positively selected thymocytes (Hare et al. 2000; Huang et al. 2001). An age-related reduction in production of IL-7 within the thymus may be responsible for the age-related decline in thymic output of naïve T cells (Andrew and Aspinall 2002; Ortman et al. 2002). In humans, accumulation of an anergic CD28− T cell population with age, especially among the CD8 T cell subset, has been documented (Boucher et al. 1998; Sansoni et al. 2008). The molecular mechanisms leading to the loss of CD28 are not known (Boucher et al. 1998; Sansoni et al. 2008). The CD28− T cells are anergic to stimulation with antigen or mitogen. In murine studies, it has been clearly shown that the clonal expansion and function of naïve CD4 or CD8 T cells is significantly reduced when compared to their younger counterparts (Plowden et al. 2004a,b; Jiang et al. 2007).
In addition to T helper and cytotoxic T cells, Th3 or Treg cells that are CD4+CD25+Fox3+ have been shown to play an important role in regulating immune responses (Dejaco et al. 2006; Hill et al. 2007). A recently published report and our unpublished findings show a significant increase in the Treg population and function with age, which may be contributing to poor adaptive immune responses (Zhao et al. 2007). However, a direct demonstration of the role of Tregs in the decline in immune responsiveness with aging is lacking, although our preliminary results indicate that depleting the Treg subset prior to immunization or infection with A/PR/8/34 virus enhanced both humoral and cellular immune responses in aged mice when compared to the control aged mice. An increased number of Tregs with age may aid in controlling the initiation of autoimmune disorders, but may come at the cost of reducing effective immune responses against infectious agents. The evolutionary significance of this finding is not clear. The functionality of CD4+ Th17 cells is beginning to be elucidated in mice, and very limited information is available on their role in humans and the impact of aging on the function of this subset (Bi et al. 2007; Nakae et al. 2007; Chen and O’Shea 2008).
CTL activity has been shown to be important for recovery from influenza virus infection in the absence of seroprotective antibodies to the infecting virus strain (McElhaney et al. 2006). CD8+ T lymphocytes recognize peptide fragments derived from viral proteins that are bound to class I MHC molecules and lyse the influenza virus-infected cells. The lysis of target cells can be mediated by perforin or by granule-mediated or Fas-mediated mechanisms (Apasov et al. 1993). CD8 T cell cytolytic activity is normally measured by labeling the MHC-compatible target cells (which are either pulsed with relevant peptides or infected with virus) with 51Cr and determining the amount of 51Cr released into the medium 4–5 h after the addition of CD8 T cells (Martz et al. 1974). Another assay to assess CTL activity is the measurement of granzyme B activity in lysates of influenza virus-stimulated PBMC; low levels of granzyme B have been correlated with risk for influenza illness in older adults (McElhaney et al. 1996, 2006).
3 Influenza Vaccine Efficacy in the Older Adult Population
Annually, influenza epidemics cause three to five million cases of severe illness with about 250,000–500,000 deaths worldwide (World Health Organization 2008). In an average year in the United States, complications from influenza infections result in about 250,000 hospitalizations and 36,000 deaths, with the majority of the fatalities occurring among the elderly population (Thompson et al. 2004; Simonsen et al. 2005). Complications from influenza viral infections resulting in hospitalizations and death are greatest among older adults, people with chronic medical conditions or immunological disorders, and infants and young children (i.e., ≤2 years of age) whose immune systems are still maturing (Fiore et al. 2007). Vaccination is the primary strategy for reducing the morbidity and mortality associated with human influenza. An inactivated detergent-split trivalent influenza vaccine (TIV) containing two influenza A viruses (H1N1 and H3N2) and a type B virus as well as a live-attenuated nasal influenza vaccine containing all three components are marketed in the USA. While injectable vaccine is recommended for people at risk, including persons aged 50 years and older, live influenza vaccine is only recommended for persons 2–49 years of age (FDA 2007). Because older adults are a high-risk group for influenza-related deaths, the goal is to vaccinate 90% of this population (DHHS 2000). However, recent vaccination rates are stagnant and coverage still hovers around 65% (National Center for Health Statistics 2003). In healthy, younger adults, the vaccine may be 70–90% effective in preventing influenza-like illness if the vaccine antigen is antigenically closely matched with the circulating epidemic strain (Gross 2002). However, vaccine efficacy is substantially reduced to 30–50% in preventing complications from influenza infections among older adults (Nichol et al. 2007). The mortality benefits from influenza vaccination of older adults is a hotly debated topic (Simonsen et al. 2007). A meta-analysis of 18 cohorts of older adults in one HMO comprising data for ten seasons from 1990–1991 through 1999–2000 indicates that vaccination resulted in a 27% reduction in the risk for hospitalization due to influenza and a 48% reduction in the risk for death (Vu et al. 2002). However, the outcomes used for these studies included hospitalizations for pneumonia or influenza and death from any cause, which are not influenza-specific. Despite increased vaccination coverage of older adults since 1980, there was no decrease in influenza-related excess mortality rates among older adults in the USA (Thompson et al. 2003; Simonsen et al. 2005). Similarly, the results from studies of Netherlands and Italian groups suggest that vaccination did not result in a reduction in excess mortality due to influenza-like illnesses, although there was not enough statistical power to generalize those findings (Govaert et al. 1994; Rizzo et al. 2006). Ideally, a randomized placebo-controlled clinical trial with clearly defined clinical outcomes such as culture-positive influenza illnesses rather than influenza-like illness and pneumonia and all-cause mortality is required to evaluate the benefit of vaccination of older adults. However, policy decisions regarding the vaccination of all older adults make a placebo-controlled study ethically unacceptable to investigate the mortality benefits of influenza vaccination (Smith and Shay 2006). It has been shown previously that influenza vaccination is 49% and 32% effective in preventing hospitalizations from pneumonia or influenza and 55% and 64% effective in preventing death from any cause among older adults at low or intermediate risk, respectively. However, among older adults who are at high risk due to comorbid conditions, vaccination is 29% and 49% effective in preventing hospitalization and death, respectively. Furthermore, when efficacy and effectiveness of vaccination among older adults are stratified by age, a different picture emerges. The efficacy of vaccination in preventing illness and hospitalization decreases with advancing age and when associated with comorbid conditions, suggesting that old older adults do not mount optimal protective immune response to vaccination. Factors that impact vaccine efficacy are presented in Fig. 3, and immunosenescence is discussed above along with other factors that influence the outcome.

Factors contributing to poor or suboptimal vaccine effectiveness in seasonal influenza vaccination of older adults
4 Active and Passive Immunization Strategies for Older Adults
A number of strategies to induce protective immune responses against influenza are presented in Fig. 4. Although passive antibody for influenza has not been considered a potential approach for both preventive and therapeutic needs, this approach has its own merits, especially when older adults who are at high risk or frail older adults who exhibit severe immune dysfunction are the target group. In addition, if the infection is caused by drug-resistant strains of influenza or a pandemic strain, passive therapy with human polyclonal antibodies offers a potential therapeutic benefit (Traggiai et al. 2004; Lanzavecchia et al. 2007; Simmons et al. 2007). Currently, transgenic animals that carry human immunoglobulin genes make human polyclonal immunoglobulins when immunized with antigens from infectious disease agents are available and these animals can serve as a potential tool to generate influenza strain-specific human antibodies for passive transfer (Fishwild et al. 1996; Tomizuka et al. 2000; Kuroiwa et al. 2002; Buelow and van Schooten 2006).

Passive and active immunization strategies for older adults against influenza
It is clear that influenza vaccine needs to be formulated differently for older adults to overcome the age-related decline in immune function and enhance the immunogenicity and protective levels of both humoral and cellular immune responses. Although MF59-adjuvanted TIV vaccine is marketed in Europe for older adults and has been shown to be safe and immunogenic, it is not yet approved in the USA (Podda and Del Giudice 2003). Newer adjuvant systems such as ASO3 have been shown to enhance the immunogenicity of H5N1 virus vaccines, and may provide a potential benefit to the older adult population if formulated with seasonal vaccines. (Treanor et al. 2006, 2007; Leroux-Roels et al. 2007; Sambhara and Poland 2007). Increasing the vaccine dose from 15 μg of HA of each vaccine component of a TIV vaccine is a potential option for enhancing the levels of protective antibodies. In a recent multisite, phase II, randomized, double-blind clinical study, older adults who received 60 μg of HA of each component were shown to generate higher levels of HAI and neutralizing antibodies when compared to those who received the standard vaccine dose of 15 μg of HA of each of the components. However, the vaccinees who received the higher dose of HA experienced more local and systemic reactions than those who received the standard vaccine dose (Keitel et al. 2006; Couch et al. 2007). This result is consistent with earlier studies and the concern that increasing the vaccine dose produces unacceptable local reactions, an effect that may be overcome with an adjuvanted vaccine to improve responses in older adults. Another possibility is to vaccinate older individuals more than once during the influenza season in order to boost antibody responses. In a small clinical study, revaccination of older adults twelve weeks later did not enhance HI titers, suggesting that such an approach may not be a viable alternative (Buxton et al. 2001). However, one of the caveats of this study was the lack of baseline titers of the vaccinees who received the second dose of the vaccine. Hence, additional studies are needed to evaluate if such an approach is a viable strategy to enhance the levels of protective antibodies in the older adult population.
5 Summary and Conclusions
Influenza is a vaccine-preventable disease, and the benefits of vaccination in preventing infection and complications arising from infection among adults are clearly documented. It is clear that the immune response declines with age due to alterations in innate and adaptive immune functions and that the vaccine does not provide adequate protection in this population. Hence, for older adults—a major target population for annual influenza vaccination, vaccine efficacy at preventing infection is low and the risk of serious complications from these infections is compounded by increasing age and comorbid conditions. Thus, efforts should be directed at formulating vaccines with adjuvants specifically for older adults to overcome immunosenescence, and passive immunization strategies with human polyclonal antibodies should be considered. In addition to conventional serological assays in the selection of vaccine candidates, other parameters namely, induction of cellular immune responses as well as activation of the innate immune system to facilitate an optimal microenvironment for the mobilization, activation, differentiation, maturation, and migration of antigen-presenting cells should be considered.
References
Alberti S, Cevenini E et al (2006) Age-dependent modifications of Type 1 and Type 2 cytokines within virgin and memory CD4+ T cells in humans. Mech Ageing Dev 127(6):560–566
Andrew D, Aspinall R (2002) Age-associated thymic atrophy is linked to a decline in IL-7 production. Exp Gerontol 37(2–3):455–463
AoA. (2007) A profile of older Americans: 2007. Administration on Aging, US Department of Health and Human Services. http://www.aoa.gov/prof/Statistics/profile/2007/2007profile.pdf. Accessed 28 Feb 2008
Apasov S, Redegeld F et al (1993) Cell-mediated cytotoxicity: contact and secreted factors. Curr Opin Immunol 5(3):404–410
Aw D, Silva AB et al (2007) Immunosenescence: emerging challenges for an ageing population. Immunology 120(4):435–446
Bi Y, Liu G et al (2007) Th17 cell induction and immune regulatory effects. J Cell Physiol 211(2):273–278
Bingle CD, Craven CJ (2002) PLUNC: a novel family of candidate host defence proteins expressed in the upper airways and nasopharynx. Hum Mol Genet 11(8):937–943
Boucher N, Dufeu-Duchesne T et al (1998) CD28 expression in T cell aging and human longevity. Exp Gerontol 33(3):267–282
Brown GD (2006) Dectin-1: a signalling non-TLR pattern-recognition receptor. Nat Rev Immunol 6(1):33–43
Buelow R, van Schooten W (2006) The future of antibody therapy. Ernst Schering Found Symp Proc 4:83–106
Buxton JA, Skowronski DM et al (2001) Influenza revaccination of elderly travelers: antibody response to single influenza vaccination and revaccination at 12 weeks. J Infect Dis 184(2):188–191
Chen Z, O’Shea JJ (2008) Regulation of IL-17 production in human lymphocytes. Cytokine 41(2):71–78
Couch RB, Winokur P et al (2007) Safety and immunogenicity of a high dosage trivalent influenza vaccine among elderly subjects. Vaccine 25(44):7656–7663
de Bruijn IA, Remarque EJ et al (1999) Quality and quantity of the humoral immune response in healthy elderly and young subjects after annually repeated influenza vaccination. J Infect Dis 179(1):31–36
de Haan G, Van Zant G (1999) Dynamic changes in mouse hematopoietic stem cell numbers during aging. Blood 93(10):3294–3301
Dejaco C, Duftner C et al (2006) Are regulatory T cells linked with aging? Exp Gerontol 41(4):339–345
DHHS (2000) Understanding and improving health and objectives for improving health. US Department of Health and Human Services, Washington DC. http://www.healthypeople.gov/Document/tableofcontents.htm#under. Accessed on 10 March 2008
Ely KH, Roberts AD et al (2007) Aging and CD8+ T cell immunity to respiratory virus infections. Exp Gerontol 42(5):427–431
FDA. (2007) FDA approves nasal influenza vaccine for use in younger children. http://www.fda.gov/bbs/topics/NEWS/2007/NEW01705.html. Accessed 12 Feb 2008
Fiore AE, Shay DK et al (2007) Prevention and control of influenza. Recommendations of the Advisory Committee on Immunization Practices (ACIP), 2007. MMWR Recomm Rep 56(RR-6):1–54
Fishwild DM, O’Donnell SL et al (1996) High-avidity human IgG kappa monoclonal antibodies from a novel strain of minilocus transgenic mice. Nat Biotechnol 14(7):845–851
Franceschi C (2007) Inflammaging as a major characteristic of old people: can it be prevented or cured? Nutr Rev 65(12 Pt 2):S173–S176
Franceschi C, Bonafe M et al (2000) Inflammaging. An evolutionary perspective on immunosenescence. Ann N Y Acad Sci 908:244–254
Ganz T (2002) Epithelia: not just physical barriers. Proc Natl Acad Sci USA 99(6):3357–3358
Ganz T, Oren A et al (1992) Defensins: microbicidal and cytotoxic peptides of mammalian host defense cells. Med Microbiol Immunol (Berl) 181(2):99–105
Gardner EM, Bernstein ED et al (2001) Characterization of antibody responses to annual influenza vaccination over four years in a healthy elderly population. Vaccine 19(32):4610–4617
Gardner EM, Gonzalez EW et al (2006) Age-related changes in the immune response to influenza vaccination in a racially diverse, healthy elderly population. Vaccine 24(10):1609–1614
Ginaldi L, Loreto MF et al (2001) Immunosenescence and infectious diseases. Microbes Infect 3(10):851–857
Gosain A, DiPietro LA (2004) Aging and wound healing. World J Surg 28(3):321–326
Govaert TM, Thijs CT et al (1994) The efficacy of influenza vaccination in elderly individuals. A randomized double-blind placebo-controlled trial. JAMA 272(21):1661–1665
Gross PA (2002) Review: inactivated vaccines provide the greatest protection against influenza in healthy persons. ACP J Club 136(3):103
Hare KJ, Jenkinson EJ et al (2000) An essential role for the IL-7 receptor during intrathymic expansion of the positively selected neonatal T cell repertoire. J Immunol 165(5):2410–2414
Hartman M, Catlin A et al (2008) US health spending by age, selected years through 2004. Health Affairs 27(1):w1–w12
Haynes L, Eaton SM et al (2000) The defects in effector generation associated with aging can be reversed by addition of IL-2 but not other related gamma(c)-receptor binding cytokines. Vaccine 18(16):1649–1653
Haynes L, Eaton SM et al (2004) Inflammatory cytokines overcome age-related defects in CD4 T cell responses in vivo. J Immunol 172(9):5194–5199
Hill JA, Benoist C et al (2007) Treg cells: guardians for life. Nat Immunol 8(2):124–125
Hoffmann JA, Kafatos FC et al (1999) Phylogenetic perspectives in innate immunity. Science 284(5418):1313–1318
Holmskov U, Thiel S et al (2003) Collectins and ficolins: Humoral lectins of the innate immune defense. Annu Rev Immunol 21(1):547–578
Htwe TH, Mushtaq A et al (2007) Infection in the elderly. Infect Dis Clin North Am 21(3):711–743
Huang J, Durum SK et al (2001) Cutting edge: histone acetylation and recombination at the TCR gamma locus follows IL-7 induction. J Immunol 167(11):6073–6077
Imler JL, Hoffmann JA (2001) Toll receptors in innate immunity. Trends Cell Biol 11(7):304–311
Iorio AM, Camilloni B et al (2007) Effects of repeated annual influenza vaccination on antibody responses against unchanged vaccine antigens in elderly frail institutionalized volunteers. Gerontology 53(6):411–418
Jiang J, Gross D et al (2007) Aging affects initiation and continuation of T cell proliferation. Mech Ageing Dev 128(4):332–339
Kang DC, Gopalkrishnan RV et al (2002) Mda-5: an interferon-inducible putative RNA helicase with double-stranded RNA-dependent ATPase activity and melanoma growth-suppressive properties. Proc Natl Acad Sci USA 99(2):637–642
Keitel WA, Atmar RL et al (2006) Safety of high doses of influenza vaccine and effect on antibody responses in elderly persons. Arch Intern Med 166(10):1121–1127
Kovaiou RD, Herndler-Brandstetter D et al (2007) Age-related changes in immunity: implications for vaccination in the elderly. Expert Rev Mol Med 9(3):1–17
Kuroiwa Y, Kasinathan P et al (2002) Cloned transchromosomic calves producing human immunoglobulin. Nat Biotechnol 20(9):889–894
Lamberts SW, van den Beld AW et al (1997) The endocrinology of aging. Science 278(5337):419–424
Lanzavecchia A, Corti D et al (2007) Human monoclonal antibodies by immortalization of memory B cells 88. Curr Opin Biotechnol. 18(6):523–528
Laube S (2004) Skin infections and ageing. Ageing Res Rev 3(1):69–89
Leroux-Roels I, Borkowski A et al (2007) Antigen sparing and cross-reactive immunity with an adjuvanted rH5N1 prototype pandemic influenza vaccine: a randomised controlled trial. Lancet 370(9587):580–589
Liang Y, Van G Zant et al (2005) Effects of aging on the homing and engraftment of murine hematopoietic stem and progenitor cells. Blood 106(4):1479–1487
Linton PJ, Li SP et al (2005) Intrinsic versus environmental influences on T cell responses in aging. Immunol Rev 205(1):207–219
Lu J, Teh C et al (2002) Collectins and ficolins: sugar pattern recognition molecules of the mammalian innate immune system. Biochim Biophys Acta 1572(2–3):387–400
Lung TL, Saurwein-Teissl M et al (2000) Unimpaired dendritic cells can be derived from monocytes in old age and can mobilize residual function in senescent T cells. Vaccine 18(16):1606–1612
Martinon F, Tschopp J (2005) NLRs join TLRs as innate sensors of pathogens. Trends Immunol 26(8):447–454
Martz E, Burakoff SJ et al (1974) Interruption of the sequential release of small and large molecules from tumor cells by low temperature during cytolysis mediated by immune T cells or complement. Proc Natl Acad Sci USA 71(1):177–181
McElhaney JE, Pinkoski MJ et al (1996) The cell-mediated cytotoxic response to influenza vaccination using an assay for granzyme B activity. J Immunol Methods 190(1):11–20
McElhaney JE, Xie D et al (2006) T cell responses are better correlates of vaccine protection in the elderly. J Immunol 176(10):6333–6339
Medzhitov R, Janeway C Jr (2000a) Innate immune recognition: mechanisms and pathways. Immunol Rev 173:89–97
Medzhitov R, Janeway C Jr (2000b) Innate immunity. N Engl J Med 343(5):338–344
Meyer KC (2001) The role of immunity in susceptibility to respiratory infection in the aging lung. Respir Physiol 128(1):23–31
Meyer KC (2004) Lung infections and aging. Ageing Res Rev 3(1):55–67
Meyer KC (2005) Aging. Proc Am Thorac Soc 2(5):433–439
Meyer KC, Ershler W et al (1996) Immune dysregulation in the aging human lung. Am J Respir Crit Care Med 153(3):1072–1079
Meyerson LB (1966) Aging and the skin. Ohio State Med J 62(5):453–456
Miller RA (1996) The aging immune system: primer and prospectus. Science 273(5271):70–74
Miller RA (1997) The aging immune system: subsets, signals, and survival. Aging (Milano) 9(4):23–24
Nakae S, Iwakura Y et al (2007) Phenotypic differences between Th1 and Th17 cells and negative regulation of Th1 cell differentiation by IL-17. J Leukoc Biol 81(5):1258–1268.
National Center for Health Statistics (2003) Early release of selected estimates based on data from the January–September National Health Interview Survey (NHIS). http://www.cdc.gov/nchs/about/major/nhis/released200303.htm#5. Accessed on 10, March 2008
Nichol KL, Nordin JD et al (2007) Effectiveness of influenza vaccine in the community-dwelling elderly. N Engl J Med 357(14):1373–1381
Ortman CL, Dittmar KA et al (2002) Molecular characterization of the mouse involuted thymus: aberrations in expression of transcription regulators in thymocyte and epithelial compartments. Int Immunol 14(7):813–822
Plowden J, Renshaw-Hoelscher M et al (2004a) Innate immunity in aging: impact on macrophage function. Aging Cell 3(4):161–167
Plowden J, Renshaw-Hoelscher M et al (2004b) Impaired antigen-induced CD8(+) T cell clonal expansion in aging is due to defects in antigen presenting cell function. Cell Immunol 229(2):86–92
Podda A, Del Giudice G (2003) MF59-adjuvanted vaccines: increased immunogenicity with an optimal safety profile. Expert Rev Vaccines 2(2):197–203
Reed MJ, Koike T et al (2003) Wound repair in aging. A review. Methods Mol Med 78:217–237
Renshaw M, Rockwell J et al (2002) Cutting edge: impaired Toll-like receptor expression and function in aging. J Immunol 169(9):4697–4701
Rizzo C, Viboud C et al (2006) Influenza-related mortality in the Italian elderly: no decline associated with increasing vaccination coverage. Vaccine 24(42–43):6468–6475
Sambhara S, Kurichh A et al (1998) Enhanced immune responses and resistance against infection in aged mice conferred by Flu-ISCOMs vaccine correlate with up-regulation of costimulatory molecule CD86. Vaccine 16(18):1698–1704
Sambhara S, Kurichh A et al (2001) Severe impairment of primary but not memory responses to influenza viral antigens in aged mice: costimulation in vivo partially reverses impaired primary immune responses. Cell Immunol 210(1):1–4
Sambhara S, Poland GA (2007) Breaking the immunogenicity barrier of bird flu vaccines. Lancet 370(9587):544–545
Sansoni P, Vescovini R et al (2008) The immune system in extreme longevity. Exp Gerontol 43(2):61–65
Schittek B, Hipfel R et al (2001) Dermcidin: a novel human antibiotic peptide secreted by sweat glands. Nat Immunol 2(12):1133–1137
Sebastian C, Espia M et al (2005) MacrophAging: a cellular and molecular review. Immunobiology 210(2–4):121–126
Shay K, Ship JA (1995) The importance of oral health in the older patient. J Am Geriatr Soc 43(12):1414–1422
Simmons CP, Bernasconi NL et al (2007) Prophylactic and therapeutic efficacy of human monoclonal antibodies against H5N1 influenza 362. PLoS Med 4(5):e178
Simonsen L, Reichert TA et al (2005) Impact of influenza vaccination on seasonal mortality in the US elderly population. Arch Intern Med 165(3):265–272
Simonsen L, Taylor RJ et al (2007) Mortality benefits of influenza vaccination in elderly people: an ongoing controversy. Lancet Infect Dis 7(10):658–666
Smith NM, Shay DK (2006) Influenza vaccination for elderly people and their care workers. Lancet 368(9549):1752–1753
Sobel JD (1997) Pathogenesis of urinary tract infection. Role of host defenses. Infect Dis Clin North Am 11(3):531–549
Sorensen OE, Cowland JB et al (2003) Wound healing and expression of antimicrobial peptides/polypeptides in human keratinocytes, a consequence of common growth factors. J Immunol 170(11):5583–5589
Takaoka A, Wang Z et al (2007) DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 448:501–505
Tesar BM, Walker WE et al (2006) Murine myeloid dendritic cell-dependent toll-like receptor immunity is preserved with aging. Aging Cell 5(6):473–486
World Health Organization (2008) Influenza. http://www.who.int/mediacentre/factsheets/fs211/en/. Accessed 12 Feb 2008
Thomas DR (2001) Age-related changes in wound healing. Drugs Aging 18(8):607–620
Thompson WW, Shay DK et al (2003) Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA 289(2):179–186
Thompson WW, Shay DK et al (2004) Influenza-associated hospitalizations in the United States. JAMA 292(11):1333–1340
Ting JP, Davis BK (2005) CATERPILLER: a novel gene family important in immunity, cell death, and diseases. Annu Rev Immunol 23:387–414
Tomizuka K, Shinohara T et al (2000) Double trans-chromosomic mice: maintenance of two individual human chromosome fragments containing Ig heavy and kappa loci and expression of fully human antibodies. Proc Natl Acad Sci U S A 97(2):722–727
Traggiai E, Becker S et al (2004) An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nat Med 10(8):871–875
Treanor JJ, Schiff GM et al (2006) Dose-related safety and immunogenicity of a trivalent baculovirus-expressed influenza-virus hemagglutinin vaccine in elderly adults. J Infect Dis 193(9):1223–1228
Treanor JJ, Schiff GM et al (2007) Safety and immunogenicity of a baculovirus-expressed hemagglutinin influenza vaccine: a randomized controlled trial. JAMA 297(14):1577–1582
van den Biggelaar AH, Huizinga TW et al (2004) Impaired innate immunity predicts frailty in old age. The Leiden 85-plus study. Exp Gerontol 39(9):1407–1414
van Duin D, Shaw AC (2007) Toll-like receptors in older adults. J Am Geriatr Soc 55(9):1438–1444
van Duin D, Allore HG et al (2007a) Prevaccine determination of the expression of costimulatory B7 molecules in activated monocytes predicts influenza vaccine responses in young and older adults. J Infect Dis 195(11):1590–1597
van Duin D, Mohanty S et al (2007b) Age-associated defect in human TLR-1/2 function. J Immunol 178(2):970–975
Vu T, Farish S et al (2002) A meta-analysis of effectiveness of influenza vaccine in persons aged 65 years and over living in the community. Vaccine 20(13–14):1831–1836
Wang CQ, Udupa KB et al (1995) Effect of age on marrow macrophage number and function. Aging (Milano) 7(5):379–384
Wood J, Schild G et al (1997) Application of an improved single-radial-immunodiffusion technique for the assay of haemagglutinin antigen content of whole virus and subunit influenza vaccines. Dev Biol Stand 39:193–200
Worley CA (2006) Aging skin and wound healing. Dermatol Nurs 18(3):265–266
Yap KL, Ada GL et al (1978) Transfer of specific cytotoxic T lymphocytes protects mice inoculated with influenza virus. Nature 273:413–420
Yoneyama M, Kikuchi M et al (2004) Th.e RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol 5(7):730–737
Zanetti M (2004) Cathelicidins, multifunctional peptides of the innate immunity. J Leukoc Biol 75(1):39–48
Zhao L, Sun L et al (2007) Changes of CD4+CD25+Foxp3+ regulatory T cells in aged Balb/c mice. J Leukoc Biol 81(6):1386–1394
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2009 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Sambhara, S., McElhaney, J.E. (2009). Immunosenescence and Influenza Vaccine Efficacy. In: Compans, R., Orenstein, W. (eds) Vaccines for Pandemic Influenza. Current Topics in Microbiology and Immunology, vol 333. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-92165-3_20
Download citation
DOI: https://doi.org/10.1007/978-3-540-92165-3_20
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-540-92164-6
Online ISBN: 978-3-540-92165-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)