
Abstract
Since life evolved on our planet there have been subtle interplays between biology and Earth System Components (atmosphere-lithosphere-ocean-interior). Life, for example, can impact weathering rates which, in turn, influence climate stabilizing feedback cycles on Earth. Photosynthesis is ultimately responsible for our oxygen-rich atmosphere, which favours the formation of the protective ozone layer. The recent rise of exoplanetary science has led to a re-examination of such feedbacks and their main drivers under different planetary conditions. In this work we present a brief overview of potential biosignatures (indicators of life) and review knowledge of the main processes, which influence them in an exoplanetary context. Biosignature methods can be broadly split into two areas, namely “in-situ” and “remote”. Criteria employed to detect biosignatures are diverse and include fossil morphology, isotope ratios, patterns in the chemical constituents of cells, degree of chirality, shifts from thermal or redox equilibrium, and changes in the abundance of atmospheric species. For the purposes of this review, our main focus lies upon gas-phase species present in Earth-like atmospheres, which could be detected remotely by spectroscopy. We summarize current knowledge based on the modern (and early) Earth and the Solar System then review atmospheric model studies for Earth-like planets, which predict climate, photochemistry and potential spectral signals of biosignature species.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

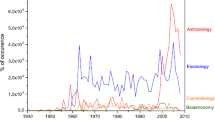

Keywords
- Exoplanet Science
- Potential Biosignatures
- Earth-like Planets
- Atmospheric Biosignatures
- CHaracterising ExOPlanet Satellite (CHEOPS)
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
The search for life beyond the Earth has been a fascination, which can be traced back to the dawn of human reasoning. The atomist school of Democritus in ancient Greece, for example, postulated the existence of many worlds—both like and unlike the Earth (see discussion in Dick 1984). In modern times, the dawning of the exploration of the Solar System and, more recently, the onset of the burgeoning exoplanet era have marked exciting times in which the age-old question “Are we alone?” is now moving into the realm of modern science.
This chapter reviews principles and techniques discussed in the exoplanetary literature for the remote detection of potential atmospheric biosignatures. We do not focus here on in-situ biosignatures involving microbial morphology, molecular biology etc., although these are fascinating and expanding fields (see Horneck et al. 2016 and references therein). Although exoplanetary science has made great headway since its inception over 20 years ago, relatively little is known about the atmospheric properties (such as mass and composition) of potentially habitable, rocky exoplanets. Therefore, a common approach in the study of potential exoplanetary biosignatures is to apply numerical atmospheric models developed from Earth’s present and past and then to vary boundary conditions, such as the spectral class of the central star, the planet-star distance etc., in order to investigate the response of planetary climate and potential biosignatures. These models are mostly constrained by knowledge gained from the modern (and Early) Earth and the Solar System. Therefore, we also briefly review knowledge of Solar System atmospheres and how this expertise can be applied in an exoplanet context. In this chapter we do not focus on the Search for Extraterrestrial Intelligence (SETI) Program nor discuss in detail the concept of “technosignatures” (signs of advanced life) but instead refer the reader to some of the main studies, such as Lin et al. (2014); Stevens et al. (2016); Griffith et al. (2015) and Korpela et al. (2015).
A comprehensive introduction to the literature is provided by five detailed review papers in remote biosignature science which recently appeared as part of the NASA Nexus for Exoplanet System Science (NExSS) and Astrobiology Program. First, Schwieterman et al. (2018) review remotely detectable potential signs of life. Second, Meadows et al. (2018) discuss atmospheric molecular oxygen in the context of its environment. Third, Catling et al. (2018) propose a framework for assessing potential biosignature signals based upon Bayesian statistics. Fourth, Walker et al. (2018) discuss a flexible framework to guide future biosignature search programs. Finally, Fujii et al. (2018) summarize future observational prospects. The present work compliments these reviews by focusing mainly on the photochemical and climate responses of atmospheric biosignatures. Note that Grenfell (2017) also summarizes the topic of exoplanetary biosignatures but with less focus on biological responses and more emphasis on physical aspects.
This chapter is organized as follows. Section 11.2 discusses briefly the challenge of defining life and its needs. Section 11.3 describes the process of preparing suitable planetary targets for biosignature assessment and the importance of understanding habitability. Section 11.4 describes currently proposed exoplanetary biosignature techniques. Section 11.5 discusses spectroscopic exoplanetary biosignatures, including climate, photochemical and spectral responses of gas-phase biosignature species in the context of modern and Early Earth, the Solar System and Earth-like exoplanets. Section 11.6 describes the “dead Earth ” as a benchmark to compare against when investigating biosignature candidates. Section 11.7 presents a brief discussion and conclusions.
2 Challenges of Defining Life and Its Needs
Commonly quoted is the NASA-based definition (Joyce et al. 1994), which proposes that life is a “self-sustaining chemical system capable of Darwinian evolution.” Benner (2010) discuss the challenges of defining a system undergoing Darwinian evolution and its general applicability beyond the Earth. Smith (2016) provides a recent summary of the philosophical challenges of defining life. A common issue is to construct a universal definition—for example the animal kingdom features hybrid crosses which are clearly alive but which cannot reproduce. Walker et al. (2018) summarize theories for defining life including the idea of biosignatures based on network theory of chemical reactions in planetary atmospheres. Life-as-we-know it has basic requirements in order to establish itself and thrive. These are, firstly, an energy source; secondly, a liquid water solvent and thirdly, a suitable supply of the chemical elements CHNOPS, as reviewed in Cockell (2016).
3 Preparing a Planetary Target List for Biosignature Assessment
3.1 Steps to Forming an Earth-Like Planet
We define two basic steps, which are necessary to form an Earth-like planet, which can host life. The first step is for a rocky planet to form with suitable water, organic and nutrient inventory. Earlier numerical simulations of volatile delivery during planetary formation (see for example Raymond et al. 2007) suggest that Earth-like planets with initially large water inventories could be rather common in nature, although this issue is still debated. The second step is for the planet to possess an atmosphere with surface climate conditions suitable to maintain liquid water over a sufficiently long period such that life can develop and thrive. The extent to which Earth-like planets can maintain mantle convection and outgas significant atmospheres (see for example Stamenkovic et al. 2012; Tackley et al. 2013; Tosi et al. 2017; Noack et al. 2017) is currently debated. Also, whether such atmospheres can be maintained against loss processes depends upon the incoming stellar wind and the planetary mass (see for example Kislyakova et al. 2014). The “eta-Earth” (ζearth) parameter (see for example Gaidos 2013) designates the fraction of stars, which possess an Earth-like planet lying in the Habitable Zone (see Sect. 11.3.2). Haghighipor (2015) define eta-Earth as an occurrence rate i.e. the mean number of rocky planets with 1–2 Earth radii per star. Better constraining eta-Earth is one of the major challenges in exoplanetary science. Traub (2015) discuss steps towards estimating ζearth using Kepler data. Estimates of ζearth have been performed for Earth-size planets orbiting cooler (M-dwarf) stars using Kepler data. Results are in the range (0.2–0.8) (see Morton and Swift 2014; Dressing and Charbonneau 2015) depending on the extent of the HZ assumed. Kane et al. (2016) discuss the challenges involved.
3.2 Habitability and the Habitable Zone (HZ)
Habitability refers to the potential of an environment to sustain life (see review by Cockell 2016). Note that it does not refer to the potential for life to emerge or to whether life is actually present.
3.2.1 Classical HZ
The classical HZ (Huang 1959; Kasting et al. 1993) is defined to be the annulus region around a star where liquid water can exist on a rocky planet’s surface. Numerous works have estimated the HZ boundaries by applying radiative-convective atmospheric models (Kaltenegger and Sasselov 2011; Yang et al. 2013; Kopparapu et al. 2014; Godolt et al. 2016; Kitzmann 2016) as well as the climate and photochemical responses of planetary atmospheres across the HZ (Segura et al. 2003; Grenfell et al. 2007; Kaltenegger and Sasselov 2011). The inner and outer boundaries depend on factors, such as the stellar type, the planetary atmosphere, and on climate feedbacks involving, for example, cloud formation. Recent estimations (see e.g. Kane et al. 2016, their Fig. 1) suggest that, for a Sun-like star, the HZ boundaries range from about 1.8FEarth (inner HZ) out to about 0.3FEarth (outer HZ) (where “FEarth“ denotes the mean solar flux received by the modern Earth) and possibly even further out (Pierrehumbert and Gaidos 2011).
3.2.2 Non-Classical HZ
Considering solvents other than water (e.g. water-ammonia, water-sulphuric acid mixtures) in the HZ definition can clearly extend the HZ boundaries (see for example Ludwig et al. 2016). Also, considering possible sub-surface oceans on moons, which lie beyond the snow-line (Vogel 1999) can extend the classical HZ outwards (although the chances of detecting potential sub-surface life remotely on such worlds seem rather remote). Such oceans have been suggested to be associated with heat sources arising either from long-term radioactive elements or/and from tidal forces (see for example Tyler 2008) and maintained by thermal blanketing provided by water-ice sheets. These issues are of particular interest in the context of the forthcoming JUICE (JUpiter ICy moons Explorer) mission (see for example Grasset et al. 2012). Heller and Barnes (2013) suggested potential habitability due to tidal heating of hypothetical moons orbiting known giant exoplanets at ~10 planetary radii. Lammer et al. (2009) proposed four classes of planetary habitability—ranging from Earth-like planets lying in the classical HZ out to icy moons possessing sub-surface oceans. The four classes are based on stellar, orbital and geophysical criteria. Also, atmospheres rich in molecular hydrogen can lead to additional greenhouse warming hence to an extension of the HZ outwards (see for example Pierrehumbert and Gaidos 2011; Ramirez et al. 2014).
3.3 HZ Around M-Dwarf Stars
M-dwarf stars are a central focus in exoplanetary science, firstly because they are numerous in the Solar Neighborhood, secondly because their relatively weak luminosity leads to favourable (planet/star) contrast ratios and, thirdly because the close-in HZ means a more favourable geometrical transit probability and suggests a fast planetary orbital period hence faster data collection. The close proximity of the HZ to the star however, suggests that planets lying in this region could be (a) tidally-locked i.e. having a constant day and night-side and (b) strongly bombarded by cosmic rays. The habitability of planets in the HZ of M-dwarf stars was reviewed by Scalo et al. (2007) and more recently by Shields et al. (2016). The effect of cosmic rays upon biosignatures was investigated by Grenfell et al. (2012) and Tabataba-Vakili et al. (2016).
When preparing a target list for biosignature assessment an important criterion is that the planet should lie in the HZ. The value of the ζηearth parameter will therefore depend on our knowledge of the extent of the HZ.
3.4 Exoplanet Missions Providing Planetary Targets
Forthcoming missions such as the Transiting Exoplanet Space Survey (TESS) (Ricker et al. 2014) and the CHaracterising ExOPlanets Satellite (CHEOPS) (Fortier et al. 2014) will expand our knowledge of hot Jupiters, mini gas planets and hot super-Earths. The PLATO 2.0 (Rauer et al. 2014) mission will provide accurate age and mean planetary density for a considerable sample of terrestrial planets lying in the HZ. The James Webb Space Telescope (JWST) (Lightsey et al. 2012) will carry out atmospheric spectroscopy on mini gas planets and super-Earths at a hitherto unprecedented level of resolution and accuracy—although finding atmospheric biosignatures on Earth-like planets will present a major challenge even for this mission. Planned ground-based telescopes, such as the European Extremely Large Telescope (see for example Snellen 2014) will perform high resolution spectroscopy of Earth-like exoplanets, although detecting atmospheric biosignatures is anticipated to be challenging also for this mission. Further afield, the Large Aperture UV-Optical-Infrared (LUVOIR) NASA spaced-based telescope (Bolcar et al. 2016) plans to survey atmospheric biosignatures on Earth-like planets. The Habitable Planet Imaging (HabEx) mission (Mennesson et al. 2016) is currently being considered and plans to hunt spectroscopically for signs of life on nearby cool rocky worlds and has a suggested launch date in the 2030s. The mission will operate with a >3.5 m optical mirror for direct imaging and spectroscopy in order to search for exoplanetary atmospheric biosignatures. In general, knowledge of the planetary environment and its evolution, for example by measuring the stellar spectrum etc., is desirable in order to better assess candidate biosignature detections. It has been proposed that the detection of ocean “glint” could also indicate whether such targeted worlds feature liquid oceans (see for example Williams and Gaidos 2008) although achieving such detection is very challenging (Cowan et al. 2012).
In summary, the pathway to compiling a suitable target list of planets suitable for biosignature assessment involves the detection of rocky Earth-like planets lying in the HZ of their central star. Horneck et al. (2016) discuss an astrobiological European roadmap in which a key goal is to define the pathway towards remote detection of life outside the Solar System.
4 Remote Biosignature Techniques
Transit spectrophotometry was discussed in an Earth-like exoplanetary context by Schneider (1994) and by numerous theoretical works since (see for example Rauer et al. 2011). Spectropolarimetry (see for example Stam 2008; Sterzik et al. 2012) is also commonly discussed in the exoplanetary literature as a possible technique to search for remote exoplanetary biosignatures. Retrieving planetary conditions, such as atmospheric composition and climate, from the spectroscopic data can be associated with degeneracies (see for example Benneke and Seager 2012) and other processes (e.g. band overlap, false positives etc. as we will discuss), which have to be considered and discounted. Spectroscopic-based topics in the context of remote biosignatures include the ‘vegetation red edge’, ‘isotopic signals’ and ‘atmospheric gas-phase species’ which we will now discuss.
5 Spectroscopic Exoplanetary Biosignatures
5.1 Reflectivity Increase with Wavelength
The “vegetation red edge” refers to the abrupt increase in reflectance of electromagnetic radiation in the wavelength range ~700–750 nm arising due to vegetation. This phenomenon is related to the marked difference between, on the one hand, strong absorption of chlorophyll in the red wavelength region and, on the other hand, high internal scattering which leads to strong reflectivity (R) by leaves in the near infrared (see for example Horler et al. 1983; Seager et al. 2005). The Horler et al. (1983) study discussed the dependence of the spectral form of the red edge, as determined by remote-sensing, on the modern Earth upon, for example species-type, developmental stage, chlorophyll content and vertical leaf-stacking. They noted in particular the dependence of λmax (i.e. the wavelength where dR/dλ attains its maximum). They also discussed some of the main challenges of performing and interpreting Earth-based remote sensing of the red-edge, such as the relatively high spectral resolution required and possible pitfalls when interpreting the spectra due to, for example, different forms of chlorophyll.
For exoplanets, Seager et al. (2005) summarized current theories regarding the potential for Earth-like planets to develop O2-producing photosynthesis. Several studies, such as Seager et al. (2005), Pallè et al. (2009), Kaltenegger et al. (2012), have discussed the vegetation red edge in an exoplanet context, including the challenge of detectability and potential false positives due to phenomena such as surface mineral reflectance. Numerous works (such as Woolf et al. 2002; Arnold et al. 2002; Sterzik et al. 2012) discussed the red edge spectral feature in modern Earthshine spectra and the challenges of applying this technique in an exoplanetary framework.
Analogous to the ‘red edge’ but shifted to weaker energies is the spectral feature related to purple bacteria (Sanroma et al. 2014). These could have been present during the Archaean Eon on the Early Earth. They are non-oxygenic phototrophs that gain energy by cleaving H2 or H2S (instead of H2O for the oxygenic case), hence their photosynthetically-active wavelength region extends out into the infra-red range as far as ~1025 nm. Applying a similar principle as the red edge, Schwieterman et al. (2015) considered non-photosynthetic pigments in general as remote biosignatures.
5.2 Isotopic Ratios
Biochemical processes can be highly specific and tend to favor the lighter isotope (cf. Chap. 3). Biological isotopic fractionation takes place for chemical elements such as C, N, S, Fe, Cu and Zn. It is hypothesized that passive diffusion across the pores of semi-permeable membranes during photosynthesis occurs more quickly for the lighter isotope. Biofractionation (which is relevant for all metabolisms, not just photosynthesis) in geochemical and biological environments has been studied in detail in works by Schidlowski (1988) (who discusses the isotopic carbon record in sedimentary rock); Boschker and Middleburg (2002) (who study stable isotope biomarker signals in the context of microbial ecology) and Fujii et al. (2014) (who investigate biomarkers related to transition metal isotope signals).
In an exoplanet context, however we will focus here only on the lighter elements from the above list because these could, in theory, accumulate in planetary atmospheres and hence be detected remotely via spectroscopy. Oxygen, whose atmospheric isotopic ratios are influenced by a rather wide range of both abiotic and biotic processes, is nevertheless a particular focus in an exoplanetary context since its isotopic distribution in Earth’s (oxygen-rich) atmosphere has been measured remotely by high resolution spectroscopy of Earthshine (Yan et al. 2015). There is no clear consensus about the extent to which life influences the atmospheric oxygen isotope ratio. Addressing this question requires consistent Earth system models featuring abiotic processes (such as ocean-atmosphere exchange, temperature, photochemistry etc.) and biotic processes (e.g. oxygen exchange within chloroplasts) in the carbon-oxygen-nitrogen cycles. Farquhar and Lloyd (1993), for example, discuss the influence of vegetation effects upon the isotopic composition of oxygen in atmospheric CO2. Abiotic effects have been established for some time, for example Kroopnick and Craig (1972) discuss the temperature-dependence of oxygen isotopic fractionation in seawater.
Hedelt et al. (2011) showed in a proof-of-concept that isotopes in Venus’ CO2 atmosphere could be distinguished by IR spectroscopy performed from the ground on Earth . The challenges inherent in detecting gas-phase species and their isotopes via infrared spectroscopy in exoplanetary atmospheres (mostly hot Jupiters) are discussed in works, such as Encrenaz (2014). In an exoplanet context however, it is very challenging to detect isotope fractionations which arise due to potential biotic activity (Snellen 2014; Holmen 1992) using spectrophotometry. A discussion of possible atmospheric isotopic ratios for a “dead Earth” in an exoplanet context is provided in Sect. 11.6.
5.3 Gas-Phase Species as Biosignatures
Here we present the main focus of this chapter—a review of photochemical and climate processes affecting key atmospheric species, such as O2, O3, and N2O, in an exoplanetary context. These molecules can be interpreted to be remote biosignatures if they occur in an exoplanetary atmosphere at an abundance (or feature a time-dependence), which is inexplicable in the context of the exoplanetary environment and its evolution without invoking biology. There are two principal of criteria that a gas-phase species must fulfil to qualify as an atmospheric biosignature: it should be uniquely attributable to life (i.e. lacking abiotic sources) and it should feature a strong and unique (i.e. without band overlap) spectral signal which is straightforward to retrieve (see e.g. Meadows et al. 2018).
Examples of potential biosignatures include, for example, the Earth’s rich-oxygen-atmosphere, or its strong ozone shield. In each case, potential abiotic sources (so-called false positives) should be discounted taking into account the planet’s environmental evolution, as we will discuss. A review of atmospheric biosignatures was provided by Seager et al. (2013, 2016). Those works grouped atmospheric biosignatures into those produced by metabolism and those produced by stress or signalling. A challenge of the approach (noted in those works) is that biomass on our planet emits a wide range of relatively simple molecules which have both biotic and non-biotic sources.
Due to the lack of knowledge (in for example mass, composition) of potentially habitable Earth-like atmospheres in exoplanetary science, a common approach is therefore to investigate responses of proposed biosignatures based on modern Earth, Early Earth, Solar System and then to extrapolate the knowledge gained (using, for example, atmospheric model studies) to Earth-like exoplanets . The following sections therefore discuss potential gas-phase biosignatures species on modern Earth, on Early Earth, in the Solar System and on Earth-like exoplanets.
5.3.1 Modern Earth
Earth’s atmosphere influences the biosphere by modifying climate, protecting the surface from harmful radiation and cosmic rays and enables the existence of surface liquid water. Conversely, life has modified Earth’s atmospheric composition over time, for example via photosynthetic and biomass emissions. Over geological time, life could have significantly influenced weathering rates and hence mantle hydration and plate tectonics (Höning et al. 2014). Life may have strongly influenced the Earth’s nitrogen cycle (e.g. Krissansen-Totton et al. 2016).
Molecular Oxygen (O2)
Earth’s rich mantle of atmospheric O2(g) (volume mixing ratio (vmr), ξo2 = 0.21; total atmospheric mass ~1.2 × 1021 g) is mainly of biotic origin. Atmospheric abundances are mostly constant in latitude and longitude and in altitude up to ~80 km above which molecular oxygen starts to photolyse into its constituent atoms. Regeneration of the resulting O2(g) is associated with the oxygen airglow (a phenomenon which is also seen in the atmospheres of Venus and Mars) (Slanger and Copeland 2003).
The main source of molecular oxygen is via photosynthesis regulated by geological burial and amounts to ~320 Tg/year (see e.g. Lasaga and Ohmoto 2002) with a smaller source which arises due to photolysis of water vapor followed by atmospheric escape of the resulting hydrogen atoms—this process constitutes therefore a net source for atmospheric oxygen. Non-biological sources of O2(g) in modern Earth’s atmosphere are, by comparison, negligible. The main sinks are reaction with reducing gases (such as CH4 and H2S) in the atmosphere (Catling and Claire 2005) and surface weathering (Holland 2002).
Ozone (O3)
Earth’s ozone layer forms a protective shield against harmful incoming solar radiation and high-energy particles. Ozone is formed in Earth’s atmosphere starting with photolysis of molecular oxygen (see below), therefore it can be interpreted as a potential biosignature for Earth-like planets, if abiotic sources on the planet are demonstrated to be weak.
Stratosphere
On Earth the O3(g) layer peaks in the mid-stratosphere reaching ~10 parts per million (ppm) at ~30 km and with a total atmospheric mass ~3 × 1015 g O3(g). Ozone is formed by the so-called Chapman mechanism (Chapman 1930):
The main sinks for O3(g) are via gas-phase catalytic cycles split according to so-called chemical “families”, for example via hydrogen-oxides (HOx) (in the mid to upper atmosphere, Bates and Nicolet 1950), ClOx (mostly occurring in the upper stratosphere; Stolarski and Cicerone 1974), and NOx (mostly in the lower stratosphere; Crutzen 1970). An overview of these cycles and how they impact ozone on the modern Earth is provided by Wayne (1993).
Ozone is affected mainly by transport in the lower stratosphere and by photochemistry in the upper stratosphere where its lifetime is shorter (World Meteorological Organization (WMO) Report 1995). Its main formation region is in the Earth’s tropics from where it is transported polewards via the Brewer-Dobson circulation in the middle atmosphere. The weakened photolytic sink of ozone in winter leads to a characteristic annual variation with an amplitude in the ozone column of (15–20%). In addition to protecting the surface from UV, ozone heating leads to a temperature inversion from ~20 to 30 km, which is the origin of the stability of air parcels against mixing processes in the stratosphere. A wide range of studies exists which have investigated the interactions of ozone with climate change (see e.g. the Intergovernmental Panel on Climate Change (IPCC) Fourth Assessment Report 2007).
Troposphere
In this region (from the surface up to the region between 10 and 20 km) ozone is formed mainly via the so-called “smog-mechanism” (Haagen-Smit 1952), which proceeds via the NOx catalysed oxidation of volatile organic compounds in the presence of UV. Production via this mechanism accounts for ~10% of the ozone on the modern Earth.
Mesosphere
In this region (from ~50 to 80 km) ozone is removed in the atmosphere mainly via photolysis and by destructive catalytic cycles involving principally HOx, whose rates are influenced by the abundance of water vapour. Hence, there is a link between ozone, water and dynamical processes. Ozone features a secondary maximum in the upper mesosphere (see for example Smith and Marsh 2005), possibly associated with low temperatures there which slow its photochemical sink(s).
Nitrous Oxide (N2O)
N2O(g) (“laughing gas”) is produced mainly by (de)nitrifying bacteria as part of the nitrogen cycle. It is an important greenhouse gas (in Earth’s modern atmosphere it has an abundance, ξN2O = 3.3 × 10−7 vmr (IPCC Fourth Assessment Report 2007) with a total mass in Earth’s atmosphere of ~2.6 × 1015 g) and is a dynamical tracer of air motions in the lower atmosphere due to its long lifetime of several hundred years against chemical destruction . It is removed in-situ in Earth’s atmosphere mainly by the following gas-phase reactions (see for example McElroy and McConnell 1971):
where O* denotes electronically-excited oxygen atoms. Global production and loss are reviewed in Syakila and Kroeze (2011). Non-biological sources are essentially negligible on the modern Earth. Suggested abiotic sources include production by lightning (Levine et al. 1979; Levine and Shaw 1983) and reaction of dissolved nitrite on iron-containing minerals (Samarkin et al. 2010).
Methane (CH4)
About 90% of modern Earth’s CH4(g) (ξCH4 ~1.8 × 10−6 vmr with a total mass of ~5.1 × 1015 g) source (amounting to about 500 Tg/year) is produced biologically via methanogenic bacteria. The remaining, abiotic contributions (Schoell 1988 provides an overview) arise from volcanic emissions or other geological processes. Atmospheric methane is destroyed mainly in the troposphere by gas-phase reaction with the hydroxyl (OH) radical:
This is the first step in a rather complex sequence of reactions which ultimately lead to methane being oxidized into water and carbon dioxide. Atmospheric OH is generated mainly via the reaction:
where O* mainly arises via photolysis of ozone. As discussed for N2O(g) above, CH4(g) is also a tracer of dynamical motions and has a lifetime against chemical loss of ~8 years (IPCC Fourth Assessment Report 2007).
5.3.2 Early Earth
During its 4.56 billion years (Ga) history, Earth’s atmosphere has undergone considerable changes—evolving from the hot, neutral to slightly reducing conditions of the Hadean up to the present-day oxidizing atmosphere. The evolution of Earth’s atmosphere over geological periods is commonly applied in exoplanetary science as a proxy for investigating the development of Earth-like atmospheres over time.
Oxygen
A wide range of geological and atmospheric proxy data (Holland 2006) suggest that Earth’s atmosphere was marked by two oxidation events in its history, the first (“Great Oxidation Event ”, GOE, Kopp et al. 2005; Catling and Claire 2005) occurred at ~2.5 Ga ago and atmospheric oxygen levels rose to at least ~1% of the modern day abundance. During the “Second Oxidation Event ” (SOE, see for example Campbell and Squire 2010) at ~0.6 Ga ago oxygen levels rose to ~10–100% of the modern day abundance. Kump et al. (2011) suggested the GOE may have been related to an increase in the burial rate of organic carbon at the ocean floor possibly associated with changes in continental distribution. Gaillard et al. (2011) suggested it could have been related to a change in the oxidation state of volcanically-emitted sulphur species. Sverjensky and Lee (2010) proposed that oxidation of Earth’s atmosphere could have led to a rapid increase in mineral diversification on the surface. The study by Anbar et al. (2007) suggested small, possibly localized excursions in atmospheric oxygen could have occurred in some regions on the Early Earth even before the GOE. Changing oxygen in the atmosphere can affect the abundance of greenhouse gases, such as methane, and, hence, impact climate. The ‘Faint Young Sun” (FYS) problem (or paradox) refers to the inconsistency between proxy (paleosol) data (which suggests habitable conditions on the Early Earth surface) and model simulations (which suggest a globally frozen surface or “Snowball Earth”). Feulner (2012) provide a review of the FYS. The study by Gebauer et al. (2017) applied a diagnostic package (“pathway analysis program”) to the output of a coupled climate-biogeochemical column model of the Early Earth atmosphere, whereas Gebauer et al (2018) performed a similar study but for the atmospheric evolution of an Earth-like planet orbiting in the HZ of an M-dwarf star. Those works identified and quantified the main chemical pathways influencing O2(g). Results suggest that oxygen is mainly destroyed by rather complex oxidation pathways in the lower atmosphere, whereas in the mid to upper stratosphere, O2(g) can be produced via photolysis of CO2.
Ozone
Model estimates suggest that the protective shield of ozone was mostly in place in its present form after the Great Oxidation Event (see for example Gebauer et al. 2017). Ozone abundances are relatively stable against changes in atmospheric oxygen increases. Why is this? An initial increase in oxygen would first favour an increase in ozone via the Chapman mechanism as shown above. The increased ozone however blocks UV at lower atmospheric levels where less oxygen is photolysed and so less ozone is formed. As this opposes the original change in ozone, a negative feedback is said to be operating.
Nitrous Oxide
Although a relatively minor greenhouse in modern Earth’s atmosphere, on the Early Earth nitrous oxide could have played a major role in affecting climate as suggested by Buick (2007) (see also the atmospheric model studies of Grenfell et al. 2011; Roberson et al. 2011). Recent work (Airapetian et al. 2016) has suggested that strong stellar activity could lead to non-biological formation of nitrous oxide in Earth-like atmospheres.
Methane
Methane has been proposed as a greenhouse gas resulting in a relatively warm the Early Earth (e.g. Pavlov et al. 2000) with methane abundance up to several orders of magnitude more than the present day atmosphere. High methane abundances were likely favoured due to neutral to slightly reducing conditions on Early Earth where methanogenic bacteria could thrive. Also, volcanic emissions of methane were likely enhanced on the Early Earth (Kasting and Catling 2003). Haqq-Misra et al. (2009) noted, however, that methane’s warming potential could be limited by haze formation, which would lead to an opposing , cooling effect.
5.3.3 Solar System
Oxygen
CO2-dominated atmospheres , such as those found on Mars and Venus, lead to the formation of low abundances of O2(g) (ξo2 = 1.4 × 10−3 vmr on Venus and ξo2 = 3 × 10−7 vmr on Mars (see Yung and DeMore 1999). O2(g) in these environments is formed by photolysis of CO2(g) into CO+O followed by the combination of two O atoms (a so-called “third body” is also required to be present e.g. N2 or O2 in order to remove excess vibrational energy of the reactants). The amount of abiotic O2 formed depends, therefore, on the rate of CO2(g) photolysis and on the fate of the resulting O-atoms. Potentially complex, catalytic cycles involving HOx (formed e.g. from water vapor) and NOx (formed e.g. from lightning and cosmic rays in the presence of N2(g)-O2(g)) control how efficiently CO and O are recycled back into CO2(g). If such cycles operate quickly then the formation of abiotic O2(g) is slowed. Although abiotic formation of O2(g) on Mars and Venus is weak compared with the biological sources on Earth, it has been suggested that this process can proceed much more quickly in exoplanet conditions if the UV-environment is favourable (see below). Small amounts of O2(g) can also be formed abiotically by the action of high energy particles on rocky or icy surfaces . This mechanism has been suggested to operate on Mercury (Hunten 1988) and the moons Europa (Hall et al. 1995), Ganymede (Hall et al. 1998) and Rhea (Teolis et al. 2010).
Ozone
As discussed above, the atmospheres of Mars and Venus feature small amounts of O2(g) and O(g). These species can react together in a three-body reaction to form O3(g), albeit in low amounts. On Venus the ozone column (Montmessin et al. 2011) is estimated to be ~150 times weaker than on Earth, whereas on Mars it is (100–1000) times weaker (Perrier et al. 2006). Weak ozone signals have also been detected in the outer solar system on some of the Galilean and Saturnian moons (see for example Noll et al. 1997). Its formation is proposed to occur via a mechanism involving high energy particles impinging upon icy surfaces.
Nitrous Oxide
Abiotic sources of N2O(g) could have operated on the Early Earth and early Venus and Mars. The mechanism involves high energy particles breaking up N2(g) to form reactive nitrogen radicals in the presence of oxygen-containing species (see papers such as Naa Mvondo et al. 2001).
Methane
Measurements by Formisano et al. (2004) triggered a debate as to the existence of methane in the Martian atmosphere (see e.g. overview presented in Zahnle et al. 2011). Recent studies which suggested up to a couple of ppbv CH4(g) as detected by the Rover Curiosity (Webster et al. 2015), have added confidence to the argument that CH4(g) is indeed present in the Martian atmosphere. The origin (Atreya et al. 2007) and the lifetime against chemical loss (Lefèvre and Forget 2009) of Martian methane, however, are not well understood (see Chap. 12).
5.3.4 Earth-Like Exoplanets
To estimate potential gas-phase biosignatures (and their false positives) on Earth-like planets, a common approach (given the paucity of data) is to extrapolate knowledge gained from the (Early) Earth and Solar System to an Earth-like exoplanet context, by performing studies with coupled convection-climate-photochemistry models. Assuming the Earth’s development and biomass, one typically varies key parameters, such as the stellar spectrum, the orbital parameters etc., and calculates the resulting atmospheric and climate responses, hence the potential effect upon atmospheric spectral bands (see for example Grenfell et al. 2007; Segura et al. 2003; Rauer et al. 2011). In an alternative approach, which is somewhat less Earth-based, models simulating planetary formation and water delivery are applied (as studied in Raymond et al. 2007) in order to learn about the occurrence and nature of rocky planets in the Habitable Zone . Other potentially important factors affecting habitability include whether the planet has a large moon (Laskar et al. 1993) or the presence of gas giants (Horner and Jones 2008). We now discuss some of the main results for proposed atmospheric biosignatures in an Earth-like exoplanetary context.
Oxygen
The case of O2(g) has illustrated the importance of elucidating the planetary environment (for example, stellar insolation etc.), primarily in order to disregard false positives when assessing potential biosignatures. A potentially important false positive for O2(g) involves H2O(g) photolysis followed by escape of atomic hydrogen. This process operates efficiently for low mass exoplanets subject to strong EUV input from their central star (which can be especially active in the pre-main sequence phase), as discussed in e.g. Luger and Barnes (2015). Wordsworth and Pierrehumbert (2014) noted that the rate of O2(g) abiotic formation for rocky planets in the HZ depends on the ability of the atmosphere’s “cold trap” (the temperature minimum at the tropopause) to freeze-out and retain water in the troposphere and, hence, prevent it photolysing. This ability is, in turn, related to the atmosphere’s inventory of non-condensable gases (such as N2(g)). Another potentially significant false positive for O2(g) involves CO2(g) photolysis followed by self-reaction of the resulting oxygen atoms, as investigated by Tian et al. (2014) and Domagal-Goldman et al. (2014). The resulting abiotic O2(g) source on Earth-like planets is sensitive to the incoming NUV from the star and could, under certain conditions, even exceed the abundance of O2(g) on the Earth. Since CO2(g) photolysis leads to CO(g) formation, the presence of this species has been suggested to be an “anti-biosignature”, i.e. an indication of abiotic oxygen formation (Wang et al. 2016; see also Schwieterman et al. 2016). The propensity to form abiotic O2(g) could be related to the elemental composition of the planet, which is mainly determined during formation (Hu and Seager 2014).
Regarding potential biological sources of O2(g) on Earth-like planets, possible photosynthesis analogues for rocky worlds orbiting in the HZ of M-dwarf stars were discussed in Kiang et al. (2007). Their work implied up to half of Earth’s photosynthetic activity occurring in visible wavelengths but possibly stronger than Earth’s activity in the IR range. Note that the release of biological O2 into Earth’s atmosphere depends upon geological processes, such as burial of organic material, which is efficient near continental shelves. Therefore, in an exoplanet context, the abundance of atmospheric O2(g) could be influenced by continental distribution and subduction—parameters which are poorly constrained for Earth-like planets. Regarding sinks of O2(g) on Earth-like planets, reaction with reducing gases in the planet’s atmosphere are estimated to be potentially important, for example some studies (Segura et al. 2005; Grenfell et al. 2012) have suggested elevated abundances of atmospheric methane (up to 1000 times those on Earth) for rocky planets orbiting in the HZ of M-dwarf stars. This is due to weaker UV output of the star, which leads to reduced production of OH, an important sink for methane. With respect to changes in the O2(g) biosignature over geological lifetimes, studying the Early Earth suggests that Earth-like atmospheres could feature strong variations in O2(g) as the planetary atmosphere evolves. Feedbacks between atmospheric evolution, climate and the development of life were discussed in Grenfell et al. (2010). Theoretical spectra for Earth-like planets calculated at different geological eons were presented and discussed in Schindler and Kasting (2000) and Kaltenegger et al. (2007).
O2 Atmospheric Spectra
Often discussed in an exoplanetary context is the rather thin O2(g) absorption band which occurs at 0.76 microns in the visible spectrum. Detecting such spectral features in the atmospheres of Earth-like planets is expected to be extremely challenging—even for bright targets in the Solar neighborhood using next generation instruments (see e.g. Snellen 2014). Des Marais et al. (2002) presented a seminal study, which investigated theoretical spectral features of Earth-like atmospheres. Rodler and López-Morales (2014) suggested that detecting the 0.76 micron band would require several tens of transits assuming an Earth-twin orbiting a bright M-star with medium (M3–M6) spectral class. The study by Kawahara et al. (2012) however suggested instead to focus on the 1.27 microns band due to more straightforward adaptive optics from the ground at these wavelengths. Misra et al. (2014) suggested to look for O2-O2 dimer band features in the IR as a proxy for detecting O2(g).
Ozone
On Earth-like planets the ozone abundance will be determined firstly by the incoming UV from the central star, which regulates the Chapman cycle forming ozone and, secondly, by trace species such as hydrogen oxides (formed for example from photolysis of water vapor) and nitrogen oxides (formed from processes such as lightning and cosmic rays) which regulate the catalytic ozone loss cycles (see above). Since ozone strongly absorbs UV it also indirectly controls the abundance of other atmospheric biosignatures, such as nitrous oxide, which are efficiently removed by photolysis in the UV. Earth-like planets orbiting M-dwarf stars are important targets in exoplanetary science as already mentioned above. A key issue is whether the close-in habitable zone, together with the planet’s possibly slowed rotation rate (hence, potentially weaker magnetosphere) due to tidal-locking, could lead to strong perturbations of the planet’s atmosphere by cosmic rays and stellar flares. The effect of these upon atmospheric biosignatures has been demonstrated by model studies to be potentially important (see Grenfell et al. 2012; Tabataba-Vakili et al. 2016; Segura et al. 2010). Regarding the evolution of atmospheric ozone in time, several studies (for example Kaltenegger et al. 2007 and Des Marais et al. 2002; Rugheimer et al. 2015) have investigated this effect by calculating theoretical atmospheric spectra.
Ozone Atmospheric Spectra
Ozone features spectral bands in the IR at 9.6 microns as well as in the visible and UV in the Chappuis and Hartley regions. The strength of certain spectral features depends not only on the ozone abundance in the atmosphere but also on the difference in temperature between the troposphere and stratosphere. Furthermore, since ozone can be produced abiotically in steam atmospheres and in carbon dioxide-dominated atmospheres, Selsis et al. (2002) put forward the concept of the “triple signature”. This proposes to search for the simultaneous presence of ozone, water and carbon dioxide (and not only ozone) as a more reliable potential biosignature. The reasoning is as follows: finding ozone in the presence of water (which can photolyse into HOx, see above, which efficiently destroys ozone) is a clue that such an ozone signal must be strongly produced in order to overcome the strong HOx sink. Atmospheres with large carbon dioxide amounts produce ozone abiotically and would tend to mask the ozone band. Retrieving atmospheric information from spectral observations is a challenging process. The study by von Paris et al. (2013) highlighted the difficulties of extracting temperature and biosignature information for nearby Earth-like planets. For ozone, a potential false positive which must be considered is band overlap with CO2 (von Paris et al. 2011) and interfering effects due to the presence of clouds (Kitzmann et al. 2011). Rauer et al. (2011) and Hedelt et al. (2013) discussed the detection signals possible for next generation missions for atmospheric CO2 and O3 for such worlds considering photon noise. Barstow and Irwin (2016) assumed an Earth-like ozone layer for the exoplanets TRAPPIST-1c and 1d and suggested that this species could be detected by averaging several tens of transits with the James Web Space Telescope.
Nitrous Oxide
An important factor influencing the biosignature nitrous oxide is the UV output from the central star. This is expected to represent the main atmospheric sink for this species, whereas the main source on Earth is through biological activity. Theoretical studies applying atmospheric models with coupled climate-photochemistry (see for example Segura et al. 2005; Grenfell et al. 2014) suggest that Earth-like planets in weak UV environments (i.e. orbiting cooler stars) could feature a significant build-up in nitrous oxide in the planetary atmosphere and, therefore, an enhanced detectability in its spectral signature. On Earth, nitrous oxide is a tracer of dynamical motions due to its long lifetime against chemical removal processes in the atmosphere. Responses in the global atmospheric circulation on Earth-like exoplanets has been investigated by several 3D model studies, such as Merlis and Schneider (2010); Godolt et al. (2015). Regarding the evolution of atmospheric nitrous oxide on Earth-like planets with time, studies from the Earth’s Proterozoic (such as Buick 2007, see above) suggest that this species could have been elevated due to enhanced biological activity. For planets that experience enhanced input of high energy particles, the study by Airapetian et al. (2016) suggested a significant abiotic source of nitrous oxide in nitrogen-oxygen atmospheres. This suggests a trade-off between UV from super-flares (destroying nitrous oxide directly via photolysis) and high-energy particles associated with flares producing nitrous oxide abiotically, which requires further investigation. Regarding spectral features, nitrous oxide absorbs rather weakly in the IR at 7.8, 4.5 and 3.7 microns (Muller 2013). In addition to its spectral features being sensitive to UV, the study by Grenfell et al. (2014) also noted that reducing methane biomass emissions led to a cooling in the middle atmosphere due to less shortwave absorption, which produced an enhancement in the spectral features of nitrous oxide.
Methane
Sources and sinks for CH4 on Earth-like exoplanets are not known. One can, however perform studies varying those processes which are known to be important on Earth, such as surface biomass emissions and abiotic sources from geological processes. In the atmosphere, methane can be produced by degradation of higher hydrocarbons and destroyed mainly (on Earth) by reaction with hydroxyl, whose concentration is sensitive to the incoming UV and water vapour abundance. For Earth-like planets orbiting in the habitable zone of M-dwarf stars, an important effect suggested by model studies (e.g. Segura et al. 2005; Rauer et al. 2011; Grenfell et al. 2014) is that weak incoming UV can lead to reduced OH in the atmosphere, which enables methane to build-up to values of up to ×1000 the methane abundance on modern Earth. Guzmán-Marmolejo et al. (2013) suggested that abiotic methane sources in rocky exoplanets from geological processes and volcanism could produce up to 10 ppmv i.e. about ×5 more than the abundance on the modern Earth. Elevated methane (and other hydrocarbons) can form hazes the properties of which could be interpreted as a biosignature , as proposed by the Arney et al. (2016) study. Theoretical studies (such as that by Schindler and Kasting 2000) suggested that atmospheric methane with abundance in excess of about ×50 that of modern Earth could be detected on nearby Earth-like exoplanets by future space missions. Regarding its spectral properties, CH4(g) features rotational-vibrational absorption bands in the IR at ~3.4 μm and ~7.7 μm (see for example Rauer et al. 2011; Werner et al. 2016). At wavelength resolutions (R = λ/δλ) coarser than R = 20, the methane bands could become indistinguishable from water absorption bands (des Marais et al. 2002).
5.4 Atmospheric Redox Disequilibrium as a Biosignature
Up to now this section has focused on the role of individual gas-phase species as atmospheric biosignatures. Alternatively, one can search for combinations of species, which are present in redox disequilibrium, i.e. the simultaneous detection of an oxidizing and a reducing chemical species whose relative abundance cannot be accounted for by abiotic processes alone. This “redox disequilibrium (or imbalance)” approach offers a new avenue to explore. It is based on the principle that strongly oxidizing and reducing species in atmospheres are not compatible and would tend to chemically react unless they are re-supplied as metabolic by-products from life. The essential idea is based on earlier works by Lovelock (1965) and Lederberg (1965). They suggested that the abundances of methane (a reducer) and oxygen (an oxidizer) in Earth’s atmosphere could suggest the presence of life. Without methanogenic bacteria supplying methane and cyanobacteria supplying oxygen, these two species would react within a few thousand years to reach abundances several orders of magnitude lower than their present day atmospheric values. The idea was discussed further in the context of the Galileo spacecraft mission in a well-known paper by Sagan et al. (1993). Simoncini et al. (2013) calculated that the CH4-O2 redox imbalance on modern Earth needs an input of ~0.7 TW from life on our planet.
The above ideas were extended further by calculations in Krissansen-Totton et al. (2016). Their work calculated redox disequilibrium for the Earth and solar system planets. Their results suggested that (N2-O2) in the presence of liquid water is an important indication of chemical disequilibrium in Earth’s atmosphere since, without life, atmospheric nitrogen would be transformed by lightning into nitrogen oxides, which would eventually be washed out and exist in the chemical stable form of nitrate in the ocean (although some molecular nitrogen could be returned to the atmosphere via mid ocean ridges). When assessing biosignature candidates using redox equilibrium, several false positives have been suggested, for example from ablating micrometeorites (Court and Sephton 2012) and the presence of a moon (Rein et al. 2014).
5.5 Additional Atmospheric Biosignatures
5.5.1 Species Containing Sulphur
Species such as dimethyl sulphide (CH3SCH3) have been suggested to build up as atmospheric biosignatures (Domagal-Goldman et al. 2011; Pilcher 2004) especially for Earth-like planets orbiting M-dwarf stars, which are weak emitters of UV. Global estimates of the atmospheric sources and sinks of sulphur compounds based on the modern Earth have been estimated (see for example Seinfeld and Pandis 2016).
5.5.2 Chloromethane
This species has been investigated as a biosignature (Segura et al. 2005; Grenfell et al. 2014). In the atmosphere this rather reactive species has an abundance of ~0.6 ppbv and is removed mainly by reaction with OH and by photolysis with a half-life of 1–2 years (IPCC Fourth Assessment Report 2007). Recently, Agúndez (2017) suggested an abiotic source of CH3Cl via comet delivery. Regarding spectral absorption bands , chloromethane absorbs rather weakly in the IR, for example at ~13.7 microns.
6 Earth-Like Planets Without Life (‘Dead Earths’)
When investigating potential biosignature candidates it is clearly useful as a benchmark for comparison to calculate atmospheric spectra for planets where life did not evolve but which otherwise have similar properties (such as mass, radius, central star, orbit etc.) to the Earth—such worlds are sometimes referred to as “dead Earths”. There are different approaches (Margulis and Lovelock 1974). First, starting with the modern Earth, one can switch off the biomass fluxes and calculate the response in atmospheric composition and climate. Second, one can estimate the evolution of an “Earth” atmosphere where life never evolved. The second approach is more challenging than the first but has the advantage that it includes the feedback that life had upon Earth’s evolution.
Margulis and Lovelock (1974) estimated the change in Earth’s atmospheric composition by removing life on Earth. Atmospheric oxygen is removed mainly via weathering of surface minerals whereas atmospheric nitrogen is removed as follows. First it undergoes molecular dissociation via lightning or cosmic rays to form atomic nitrogen which quickly forms NOx in the atmosphere. NOx is then oxidized in-situ mainly into nitric acid, which is removed for example by rainout to form nitrate in the ocean. Molecular nitrogen can be returned into the atmosphere via outgassing at mid ocean ridges. The atmospheric CO2(g) abundance in the atmosphere of a Dead Earth is challenging to estimate. The study by Margulis and Lovelock (1974) suggested 0.3–1000 mb at the surface. More recent studies however (see for example Morrison and Owen 2003) suggested that all the Earth’s CO2 (their study suggested ~69 bars which on the modern Earth resides mainly in the lithosphere) would, instead, reside in the atmosphere in the case of a dead Earth. In order to constrain these large uncertainties, improved information regarding the flux responses in Earth’s carbon cycle are required. In a somewhat different approach, the study by O’Malley-James et al. (2014) investigated the future Earth where the biosphere dies at ~2.8 Gr as the luminosity of the sun increases. Their work suggested an intensification in the hydrological cycle with photosynthesis stopping when carbon dioxide levels in the atmosphere decrease below 10 ppmv, i.e. about ×40 times lower than the present day values. It is desirable in the biosignature literature to re-investigate such issues with the modern suite of 1D and 3D coupled atmospheric models.
The massive extinction event at the end of the Permian 251 million years ago led to the loss of ~95% of Earth’s species. Geochemical data (see for example Benton and Twitchet 2003) suggest a decrease of ~6 parts per thousand in δ18O as well as a negative excursion in δ13C from +2 to +4 down to −2 parts per thousand in the ocean (with possibly a similar change in the lower atmosphere) (see also Kump 1991). It is not clear, however to what extent these signals could have arisen due to the loss of life (via a reduction in the global organic carbon burial fluxes) or, due to other abiotic processes, for example, possibly large releases of methane from gas hydrates at the time. In an exoplanet context, however, such small isotopic signals are not likely to be measured remote spectroscopy within the next few decades. An additional major challenge is to determine which isotopic background to compare against when searching for signs of life.
7 Discussion and Conclusions
The search for signs of life beyond the Earth is one of the greatest challenges of our times. Scientific developments in exoplanetary science have been particularly rapid over the last few decades. Significant progress has already been made in observing the occurrence and nature of the hotter, gassy planets. For the cooler, rocky planets the determination of their occurrence and characterization is still in a relatively early stage. Regarding theoretical studies of the potential distribution of life beyond the Earth there is still much headway to be made. Some of the key issues are to investigate the factors which affect how life is initiated and better constrain conditions under which it can thrive, survive and possibly even spread beyond its planet of origin. In a more concrete sense, these issues are related to the distribution of habitable environments in the Universe, which are likely linked with the occurrence of rocky planets and the width of the habitable zone. Such issues are active areas of research in exoplanetary science.
Techniques to search for potential exoplanetary life include hunting for spectral signals of vegetation’s strong reflection in visible wavelengths (the “red edge”) and the luminescence of, for example, biopigments. Another technique involves searching for atmospheric species associated with life in planetary atmospheres via spectrophotometry and spectropolarimetry. Regarding atmospheres there are two main possibilities—either one can search for individual gas-phase species (such as ozone, nitrous oxide etc.) or one can search for combinations of reducing-oxidizing gas-phase species with abundances which are incompatible without life (“redox disequilibrium” method).
Our work has focused mainly on gas-phase atmospheric species and their potential to act as biosignatures. Ozone is a key biosignature species because it protects the lower atmosphere from UV hence control to a large extent whether other atmospheric biosignatures can survive. Oxygen has received considerable attention lately, whereby various works discuss possible abiotic sources and which emphasize the role of the environment when assessing atmospheric biosignatures. Nitrous oxide is usually acknowledged to arise almost exclusively from biology, although caveats to this idea have recently appeared (see for example Airapetian et al. 2016) which suggest abiotic production due to cosmic rays associated with stellar flares.
Hedelt et al. (2013) calculated theoretical transit depth signals estimated for nearby Earth-like planets as well as atmospheric absorption features for e.g. climate relevant species such as the broad CO2 feature (at 15 microns) and potential biosignatures such as ozone (at 9.6 microns).
Regarding future exoplanetary missions, TESS and CHEOPS will improve understanding of gas planets and hot Super-Earths. PLATO 2.0 will provide a target list of habitable, rocky worlds in the HZ for follow-up spectroscopy. It is possible that the JWST and E-ELT could deliver the first spectroscopic observations of atmospheric biosignatures if a list of suitable targets (for example rocky planets in the HZ of nearby, M-dwarf stars) is available. Missions planned for farther into the future such as LUVOIR and HabEx aim to observe atmospheric biosignatures for a wider sample of cool, rocky planets.
As a final note we should keep in mind that the first exoplanetary biosignatures to be detected will likely be those with clear spectral signals on planets where strong biomass emissions have globally modified the atmospheric composition. A lack of detection of such signals, however, does not necessarily mean that the planet is not inhabited.
We live in exciting times—where age-old debates move into the realm of modern science.
References
Agúndez M (2017) Organohalogens in space. Nat Astron 1:655–656
Airapetian VS, Glocer A, Gronoff G et al (2016) Prebiotic chemistry and atmospheric warming of early Earth by an active young sun. Nat Geosci 9:452–455
Anbar AD, Duan Y, Lyons TW et al (2007) A whiff of oxygen before the great oxidizing event. Nature 317:1903–1906
Arney G, Domagal-Goldman S, Meadows VS et al (2016) The pale orange dot: the spectrum and habitability of Hazy Archean Earth. Astrobiology 16:873–899
Arnold L, Gillet S, Lardière et al (2002) A test for the search for life on extrasolar planets. Looking for the terrestrial vegetation signature in Earthshine spectrum. Astron Astrophys 92:231–237
Atreya SK, Mahaffy PR, Wong AS (2007) Methane and related trace species on Mars: origin, loss, implications for life and habitability. Planet Space Sci 55:358–369
Barstow J, Irwin PGJ (2016) Habitable worlds with JWST: transit spectroscopy of the TRAPPIST-1 system? MNRAS 461:L92–L96
Bates DR, Nicolet M (1950) The photochemistry of atmospheric water vapor. J Geophys Res 55:301–327
Benneke B, Seager S (2012) Atmospheric retrieval for Super-Earths. Astrophys J 753:2
Benner SA (2010) Defining life. Astrobiology 10:1021–2030
Benton MJ, Twitchet RJ (2003) How to kill (almost) all life: the end-Permian extinction event. Trends Ecol Evol 18:358–365
Bolcar MR, Balasubramanian K, Crooke J et al (2016) Technology gap assessment for a future large-aperture ultraviolet-optical infrared space telescope. Astron Telesc Instrum Syst 2:041209
Boschker HTS, Middleburg JJ (2002) Stable isotopes and biomarkers in microbial ecology. FEMS Microbiol 40:85–95
Buick R (2007) Did the Proterozoic ‘Canfield ocean’ cause a laughing gas greenhouse? Geobiology 5:97–100
Campbell H, Squire RJ (2010) The mountains that triggered the Late Neoproterozoic increase in oxygen: the second great oxidation event. Geochim Cosmochim Acta 74:4187–4206
Catling DC, Claire MW (2005) How Earth’s atmosphere evolved to an oxic state: a status report. Earth Planet Sci Lett 237:1–20
Catling DC, Krissansen-Totton J, Kiang NY, Crisp D, Robinson TD et al (2018) Exoplanet biosignatures: a framework for their assessment. Astrobiology 18:709–738
Chapman S (1930) On ozone and atomic oxygen in the upper atmosphere. The Lond Edin Dub Philps Mag J Sci 10:369–383
Cockell C (2016) Habitability: a review. Astrobiology 16:89–117
Court RW, Sephton MA (2012) Extrasolar planets and false atmospheric biosignatures: the role of micrometeoroids. Planet Space Sci 73:233–242
Cowan NB, Abbot DS, Voigt A (2012) A false positive for ocean glint on exoplanets: the latitude-albedo effect. Astrophys J 752:L3
Crutzen PJ (1970) The influence of nitrogen oxides upon the atmospheric ozone content. Q J R Met S 96:320–325
Des Marais DJ, Harwit MO, Jucks KW (2002) Remote sensing of planetary properties and biosignatures on extrasolar terrestrial planets. Astrobiology 2:153–181
Dick SJ (1984) The plurality of worlds: the extra-terrestrial life debate from Democritus to Kant. Cambridge University Press, Cambridge, UK
Domagal-Goldman S, Meadows VS, Claire MW (2011) Using biogenic sulfur gases as remotely detectable biosignatures on anoxic planets. Astrobiology 11:419–441
Domagal-Goldman S, Segura A, Claire MW (2014) Abiotic ozone and oxygen in atmospheres similar to prebiotic Earth. Astrphys J Lett 787:2
Dressing C, Charbonneau D (2015) The occurrence of potentially habitable planets orbiting M dwarfs estimated from the full Kepler dataset and an empirical measurement of the detection sensitivity. Astrophys J 807:45
Encrenaz T (2014) Infrared spectroscopy of exoplanets: observational constraints. Philos Trans R Soc A 372:20130083
Farquhar GD, Lloyd J (1993) Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial planets and the atmosphere. In: Ehleringer JR, Hall AE, Farquhar GD (eds) Stable isotopes and plant carbon-water relations. Elsevier, New York, pp 47–70
Feulner G (2012) The faint young Sun problem. Rev Geophys 50:1–29
Formisano V, Atreya S, Encrenaz T (2004) Detection of methane in the atmosphere of Mars. Nature 306:1758–1761
Fortier A, Beck T, Benz W (2014) CHEOPS: a space telescope for ultra-high precision photometry of exoplanet transits. J Astron Telesc Instrum Syst 9143
Fujii T, Moynier F, Blichert-Toft J et al (2014) Density functional theory estimation of isotope fractionation of Fe, Ni, Cu, and Zn among species relevant to geochemical and biological environments. Geochim Cosmochim Acta 140:553–576
Fujii Y, Angerhausen D, Deitrick R et al (2018) Exoplanet biosignatures: observational prospects. Astrobiology 18(6). doi: https://doi.org/10.1089/ast.2017.1733
Gaidos E (2013) Candidate planets in the habitable zones of Kepler stars. Astophys J 770:2
Gaillard F, Scaillet B, Arndt NT (2011) Atmospheric oxygenation caused by a change in volcanic degassing pressure. Nature 478:229–232
Gebauer S, Grenfell JL, Stock JW et al (2017) Evolution of Earth-like extrasolar planetary atmospheres. Astrobiology 17:27–54
Gebauer S, Grenfell JL, Lehmann R, Rauer H (2018) Evolution of Earth-like planetary atmospheres around M Dwarf Stars: assessing the atmospheres and biospheres with a coupled atmosphere biogeochemical model. Astrobiology 18:856–872
Godolt M, Grenfell JL, Hamann-Reinus A et al (2015) 3D climate modeling of Earth-like extrasolar planets orbiting different types of host stars. Planet Space Sci 111:62–76
Godolt M, Grenfell JL, Kitzmann D et al (2016) Assessing the habitability of planets with Earth-like atmospheres with 1D and 3D climate modeling. Astron Astrophys 592:A36
Grasset O, Dougherty MK, Coustenis A et al (2012) JUpiter ICy moons Explorer (JUICE): an ESA mission to orbit Ganymede and to characterise the Jupiter system. Planet Space Sci 78:1–21
Grenfell JL (2017) A review of exoplanetary biosignatures. Phys. Rep. 713:1–17
Grenfell JL, Stracke B, von Paris P et al (2007) The response of atmospheric chemistry on earthlike planets around F, G and K stars to small variations in orbital distance. Planet Space Sci 55:661–671
Grenfell JL, Rauer H, Selsis F et al (2010) Co-evolution of atmospheres, life and climate. Astrobiology 10:77–88
Grenfell JL, Gebuaer S, von Paris P et al (2011) Sensitivity of biomarkers to changes in chemical emissions in Earth’s Proterozoic atmosphere. Icarus 211:81–88
Grenfell JL, Griessmeier J-M, von Paris P et al (2012) Response of atmospheric biomarkers to NOx-induced photochemistry generated by stellar cosmic rays for Earth-like planets in the habitable zone of M dwarf stars. Astrobiology 12:1109–1122
Grenfell JL, Gebauer S, von Paris et al (2014) Sensitivity of biosignatures on Earth-like planets orbiting in the habitable zone of cool M-dwarf stars to varying stellar UV radiation and surface biomass emissions. Planet Space Sci 98:66–76
Griffith RL, Wright JT, Maldonado J et al (2015) The G infrared search for extraterrestrial civilizations with large energy supplies. Astrophys J 217(2)
Guzmán-Marmolejo A, Segura A, Escobar-Briones E (2013) Abiotic production of methane in terrestrial planets. Astrobiology 13:550–559
Haagen-Smit AJ (1952) Chemistry and physiology of Los Angeles smog. Ind Eng Chem 44:1342–1346
Haghighipor N (2015) Eta-Earth. Encyclopedia of astrobiology. Springer, Heidelberg
Hall DT, Strobel DF, Feldman PD et al (1995) Detection of an oxygen atmosphere on Jupiter’s moon Europa. Nature 373:677–681
Hall DT, Feldman PD, McGrath MIA et al (1998) The far-ultraviolet oxygen airglow of Europa and Ganymede. Astrophys J 449:475–481
Haqq-Misra JD, Domagal-Goldman SD, Kasting PJ et al (2009) A revised, hazy methane greenhouse for the Archean Earth. Astrobiology 8:1127–1137
Hedelt P, Alonso R, Brown T et al (2011) Venus transit 2004: illustrating the capacity of exoplanet transmission spectroscopy. Astron Astrophys 533:A136
Hedelt P, von Paris P, Godolt M et al (2013) Spectral features of Earth-like planets and their detectability at different orbital distances around F, G, and K-type stars. Astron Astrophys 553:A9
Heller R, Barnes R (2013) Exomoon habitability constrained by illumination and tidal heating. Astrobiology 13:18–46
Holland HD (2002) Volcanic gases, black smokers and the great oxidation event. Geochim Cosmochim Acta 66:3811–3826
Holland HD (2006) The oxygenation of the atmosphere and oceans. Philos Trans R Soc Lond Biol Sci 361:903–915
Holmen K (1992) The global carbon cycle. London Academic Press, London, pp 237–262
Höning D, Hansen-Goos H, Airo A (2014) Biotic vs. abiotic Earth: a model for mantle hydration and continental coverage. Planet Space Sci 98:5–13
Horler DNH, Dockray M, Barber J (1983) The red edge of plant leaf reflectance. Int J Remote Sens 4:273–288
Horneck G, Walter N, Westall F et al (2016) AstRoMap European Astrobiology Roadmap. Astrobiology 16:201–243
Horner J, Jones BW (2008) Jupiter – friend or foe? I: the asteroids. Int J Astrobiol 7:251–261
Hu R, Seager S (2014) Photochemistry in terrestrial planet atmospheres III. ApJ 784:1
Huang S (1959) Occurrence of life in the universe. Am Sci 47:397–402
Hunten DM (1988) Mercury. University of Arizona Press, Tucson, AZ
International Panel on Climate Change (IPCC) Climate Change (2007) In: Solomon S et al (eds) The physical basis. IPCC, Geneva
Joyce G, Deamer DW, Fleischaker GR (1994) In: Deamer DW, Fleichacker GR (eds) Origins of life: the central concepts. Jones and Bartlett, Boston, pp xi–xii
Kaltenegger L, Sasselov D (2011) Exploring the habitable zone for Kepler planetary candidates. ApJ 736:2
Kaltenegger L, Traub WA, Jucks KW et al (2007) Spectral evolution of an Earth-like planet. ApJ 658:1
Kaltenegger L, Miguel Y, Rugheimer S (2012) Rocky exoplanet characterization and atmospheres. Int J Astrobiol 11:297–307
Kane SR, Hill ML, Kasting JF et al (2016) A catalogue of Kepler habitable zone exoplanet candidates. ApJ 830(1)
Kasting JF, Catling DC (2003) Evolution of a habitable planet. Annu Rev Astron Astrophys 41:429–463
Kasting JF, Whitmire DP, Reynolds RT (1993) Habitable zones around main sequence stars. Icarus 101:108–128
Kawahara H, Matsuo T, Takami M et al (2012) Can ground-based telescopes detect the 1.27 micron absorption feature as a biomarker in exoplanets? ApJ 758:1
Kiang NY, Segura A, Tinetti G et al (2007) Spectral signatures of photosynthesis. II. Coevolution with other stars and the atmosphere on extrasolar worlds. Astrobiology. 7:252–274
Kislyakova KG, Johnstone CP, Odert P et al (2014) Stellar wind interaction and pick-up ion escape of the Kepler-11 “super-Earths”. Astron Astrophys 562:A116
Kitzmann D (2016) Revisiting the scattering greenhouse effect of CO2 ice clouds. ApJL 817:2
Kitzmann D, Patzer ABC, von Paris P et al (2011) Clouds in the atmospheres of extrasolar planets. Astron Astrophys 531:A62
Kopp RE, Kirschvink JL, Hilburn IA et al (2005) The Paleoproterozoic snowball Earth: a climate disaster triggered by the evolution of oxygenic photosynthesis. Proc Natl Acad Sci USA 102:11131–11136
Kopparapu RK, Ramses M, Schttelkotte J et al (2014) Habitable zones around main sequence stars: dependence upon planetary mass. ApJL 787:2
Korpela EJ, Sallmen SM, Greene DL (2015) Modeling indications of technology in planetary transit light curves – dark-side illumination. ApJ 809:2
Krissansen-Totton J, Bergsman DS, Catling DC (2016) On detecting biosignatures from chemical thermodynamic disequilibrium in planetary atmospheres. Astrobiology 16:39–67
Kroopnick P, Craig H (1972) Atmospheric oxygen: isotopic composition and solubility fractionation. Science 175:54–55
Kump LR (1991) Interpreting carbon-isotope excursions: Strangelove oceans. Geology 19:299–302
Kump LR, Junium C, Arthur MC et al (2011) Isotopic evidence for massive oxidation of organic matter following the Great Oxidation Event. Science 334:1694–1696
Lammer H, Bredehöft JH, Coustenis A et al (2009) What makes a planet habitable? Astron Astrophys Rev 17:181–189
Lasaga AC, Ohmoto H (2002) The oxygen geochemical cycle: dynamics and stability. Geochim Cosmochim Acta 66:361–381
Laskar J, Joutel F, Roboutal P et al (1993) Stabilization of the Earth’s obliquity by the Moon. Nature 361:615–617
Lederberg J (1965) Signs of life. Nature 207:9–13
Lefèvre F, Forget F (2009) Observed variations of methane on Mars unexplained by known atmospheric chemistry and physics. Nature 460:720–723
Levine JS, Shaw EF (1983) In situ aircraft measurements of enhanced levels of N2O associated with thunderstorm lightning. Nature 303:312–314
Levine JS, Hughes RE, Chameides WL et al (1979) N2O and CO production by electric discharge: atmospheric implications. Geophys Res Lett 6:557–559
Lightsey PA, Atkinson CB, Clampin MC et al (2012) James Webb Space Telescope: large deployable telescope in space. Opt Eng 51:1
Lin HW, Abad GG, Loeb A (2014) Detecting industrial pollution in the atmospheres of Earth-planets. Astrophys J Lett 791:1
Lovelock JE (1965) A physical basis for life detection experiments. Nature 207:568–570
Ludwig W, Eggl S, Neubauer D et al (2016) Effective stellar flux calculations for limits of life-supporting zones of exoplanets. MNRAS 458:3752–3759
Luger R, Barnes R (2015) Extreme water loss and abiotic O2 buildup on planets throughout the habitable zone on M-dwarfs. Astrobiology 15:119–143
Margulis LM, Lovelock JE (1974) Biological modulation of the Earth’s atmosphere. Icarus 21:471–489
McElroy MB, McConnell JC (1971) Nitrous oxide: a natural source of NO. Am Met Soc 28:1095–1098
Meadows VS, Reinhard CT, Arney GN et al (2018) Exoplanet biosignatures: understanding oxygen as a biosignature in the context of its environment. Astrobiology 18(6):630–662
Merlis TM, Schneider T (2010) Atmospheric dynamics of Earth-like tidally-locked aquaplanets. J Adv Mod Earth Sys 2:13
Mennesson B, Gaudi S, Seager S et al (2016) The Habitable Exoplanet (HabEx) Imaging Mission: preliminary science drivers and technical requirements. J Astron Telesc Instrum Syst 9904
Misra A, Meadows VS, Claire MW et al (2014) Using dimers to measure biosignatures and atmospheric pressure for terrestrial exoplanets. Astrobiology 14:67–86
Montmessin F, Bertaux JL, Lefèvre F et al (2011) A layer of ozone detected in the nightside upper atmosphere of Venus. Icarus 216:82–85
Morrison D, Owen T (2003) The planetary system, 3rd edn. Addison-Wesley, Reading, MA
Morton TD, Swift J (2014) The radius distribution of planets around cool stars. Astrophys J 791:10
Muller C (2013) N2O as a biomarker: from the Earth and solar system to exoplanets. Astrophys Spa Sci Proc 35:99–106
Naa Mvondo D, Navarro-Gonzalez R, McKay CP et al (2001) The production of nitrogen oxides by lightning and coronal discharges in simulated early Earth, Venus and Mars environments. Adv Space Res 27:217–223
Noack L, Rivoldini A, Van Hoolst T (2017) Volcanism and outgassing of stagnant-lid planets: implications for the habitable zone. PEP 269:40–57
Noll KS, Roush TL, Cruikshank DP et al (1997) Detection of ozone on Saturn’s satellites Rhea and Dione. Nature 388:45–47
O’Malley-James JT, Greaves JS, Raven JA et al (2014) Swansong Biospheres II: the final signs of life on terrestrial exoplanets near the end of their habitable lifetimes. Int J Astrobiol 13:229–243
Pallé E, Osorio MRZ, Barena R et al (2009) Earth’s transmission spectrum from lunar eclipse measurements. Nature 459:814–816
Pavlov AA, Kasting JF, Brown LL et al (2000) Greenhouse warming by CH4 in the atmosphere of Early Earth. J Geophys Res 105:11,981–11,990
Perrier S, Bertaux JL, Lefèvre F et al (2006) Global distribution of total ozone on Mars from SPCAM/MEX UV measurements. J Geophys Res 111:E9
Pierrehumbert R, Gaidos E (2011) Hydrogen greenhouse planets beyond the habitable zone. Astrophys J Lett 734:L13
Pilcher CB (2004) Biosignatures of Early Earths. Astrobiology 3:471–486
Ramirez RM, Kopparapu R, Zugger ME et al (2014) Warming early Mars with CO2 and H2. Nat Geosci 7:59–63
Rauer H, Gebauer S, von Paris P et al (2011) Potential biosignatures in super-Earth atmospheres. I. Spectral appearance of super-Earths around M dwarfs. Astron Astrophys 529:A8
Rauer H, Catala C, Aerts C et al (2014) The PLATO 2.0 Mission. Exp Astron 38:249–330
Raymond SN, Quinn T, Lunine JI (2007) High-resolution simulations of the final assembly of Earth-like planets. 2. Water delivery and planetary habitability. Astrobiology 7:66–84
Rein H, Fujii Y, Spiegel DS (2014) Some inconvenient truths about biosignatures involving two chemical species on Earth-like exoplanets. Proc Natl Acad Sci USA 111:6871–6875
Ricker GR, Winn JN, Vanderspeck R et al (2014) Transiting exoplanet survey satellite. J Astron Telesc Instrum Syst 1:014003
Roberson AL, Roadt J, Halevy I et al (2011) Greenhouse warming by nitrous oxide and methane in the Proterozoic eon. Geobiology 9:313–320
Rodler F, López-Morales M (2014) Feasibility studies for the detection of O2 in an Earth-like exoplanet. Astrophys J 781:1
Rugheimer S, Kaltenegger L, Segura A et al (2015) Effect of UV on the spectral fingerprints of Earth-like planets orbiting M-stars. Astrobiology 809:1–16
Sagan C, Thompson WR, Carlson R et al (1993) A search for life on Earth from the Galileo spacecraft. Nature 365:375–377
Samarkin VA, Madigan MT, Bowles MW et al (2010) Abiotic nitrous oxide emission from the hypersaline Don Juan Pond in Antarctica. Nat Geophys 3:341–344
Sanroma E, Palle E, Parenteau MN et al (2014) Characterizing the purple Earth: measuring the globally-integrated spectral variability of the Archaean Earth. Astrophys J 780(1)
Scalo J, Segura A, Fridlund M et al (2007) M stars as targets for terrestrial exoplanet searches and biosignature detection. Astrobiology 7:85–166
Schidlowski M (1988) A 3800 million-year isotopic record of life from carbon in sedimentary rocks. Nature. 333:313–318
Schindler TL, Kasting JF (2000) Synthetic spectra of simulated terrestrial atmospheres containing possible biomarker gases. Icarus 145:262–271
Schneider J (1994) On the search for O2 in extrasolar planets. Astrophys Space Sci 212:321–325
Schoell M (1988) Multiple origins of methane in the Earth. Chem Geol 71:1–10
Schwieterman E et al (2015) Non photosynthetic pigments as potential biosignatures. Astrobiology 15:341–361
Schwieterman EW, Cockell CS, Meadows VS et al (2016) Identifying planetary biosignature imposters: spectral features of CO and O4 resulting from O2/O3 production. Astrophys J 819(1)
Schwieterman EW, Kiang NY, Parenteau MN, Harman CE, DasSarma S et al (2018) Exoplanet biosignatures: a review of remotely detectable signs of life. Astrobiology 18:663–708
Seager S, Turner E, Schafer LJ et al (2005) Vegetation’s red edge: a possible spectroscopic biosignature of extraterrestrial plants. Astrobiology 5:372–390
Seager S, Bains W, Hu R (2013) Biosignature gases in H2-dominated atmospheres on rocky planets. Astrophys J 777:2
Seager S, Bains W, Petkowski JJ (2016) Toward a list of molecules as potential biosignature gases for the search for life on exoplanets and applications to terrestrial biochemistry. Astrobiology 16:465–485
Segura A, Krelove K, Kasting JF et al (2003) Ozone concentrations and ultraviolet fluxes on Earth-like planets around other stars. Astrobiology 3:689–708
Segura A, Kasting JF, Meadows VS et al (2005) Biosignatures from Earth-like planets around M-stars. Astrobiology 5:706–725
Segura A, Walkowicz MVS et al (2010) The effect of a strong stellar flare on the atmospheric chemistry of an Earth-like planet orbiting an M-dwarf. Astrobiology 10:751–771
Seinfeld JH, Pandis SN (2016) From air pollution to climate change. Wiley, Hoboken, NJ
Selsis F, Despoit D, Parisot J-P et al (2002) Signature of life on exoplanets: can Darwin produce false positive detections? Astron Astrophys 388:985–1003
Shields AL, Ballard S, Johnson JA (2016) The habitability of planets orbiting M-dwarf stars. Phys Res 663:1–38
Simoncini E, Virgo N, Kleidon A et al (2013) Quantifying drivers of chemical disequilibrium: theory and application to methane in Earth’s atmosphere. Earth Syst Dyn 4:317–331
Slanger TG, Copeland RA (2003) Energetic oxygen in the upper atmosphere and the laboratory. Chem Rev 103:4731–4766
Smith KC (2016) Life is hard: countering definitional pessimism concerning the definition of life. Int J Astrobiol 15:277–289
Smith AK, Marsh DR (2005) Processes that account for the ozone maximum at the mesopause. J Geophys Res 110:D23
Snellen I (2014) High-dispersion spectroscopy of extrasolar planets: from CO in hot Jupiters to O2 in exo-Earths. Philos Trans R Soc A 372:20130075
Stam DM (2008) Spectropolarimetric signatures of Earth-like extrasolar planets. Astron Astrophys 482:989–1007
Stamenkovic V, Noack L, Breuer D et al (2012) The influence of pressure-dependent viscosity on the thermal evolution of Super-Earths. Astrophys J 748:1
Sterzik MF, Bagnul S, Palle E (2012) Biosignatures as revealed by spectropolarimetry of Earthshine. Nature 483:64–66
Stevens A, Forgan D, James JOM (2016) Observational signatures of self-destructive civilizations. Int J Astrobiol 15:33–44
Stolarski RJ, Cicerone RS (1974) Stratospheric chlorine: a possible sink for ozone. Can J Chem 52:1610–1615
Sverjensky DA, Lee N (2010) The great oxidation event and mineral diversification. Elements 6:31–36
Syakila A, Kroeze C (2011) The global nitrous oxide budget revisited. Greenhouse Gas Meas Manag 1:17–26
Tabataba-Vakili F, Grenfell JL, Griessmeier J-M et al (2016) Atmospheric effects of stellar cosmic rays on Earth-like exoplanets orbiting M-dwarfs. Astron Astrophys 585:A96
Tackley PJ, Ammann M, Brodholt JP (2013) Mantle dynamics in super-Earths: post-perovskite rheology and self-regulation of viscosity. Icarus 225:50–61
Teolis BD, Jones GH, Miles PF (2010) Cassini finds an oxygen-carbon dioxide atmosphere at Saturn’s icy moon Rhea. Science 333:6012
Tian F, France K, Linsky JL et al (2014) High stellar FUV/NUV ratio and oxygen contents in the atmospheres of potentially habitable planets. Earth Planet Sci 385:22–27
Tosi N, Godolt M, Stracke B et al (2017) The habitability of a stagnant-lid Earth. Astron Astrophys 605:A71
Traub WA (2015) Steps towards eta-Earth from Kepler data. Int J Astrobiol 14:359–363
Tyler RH (2008) Strong ocean tidal flow and heating on moons of the outer planets. Nature 456:770–772
Vogel G (1999) Expanding the habitable zone. Science 286:70–71
von Paris P, Cabrera J, Godolt M et al (2011) Spectroscopic characterization of the atmospheres of potentially habitable planets: Gl581d as a model case study. Astron Astrophys 534:A26
von Paris P, Hedelt P, Selsis F et al (2013) Characterization of potentially habitable planets: retrieval of atmospheric and planetary properties from emission spectra. Astron Astrophys 551:A120
Walker SI, Bains W, Cronin L et al (2018) Exoplanet biosignatures: future directions. Astrobiology 18(6):779–824
Wang Y, Tian F, Li T et al (2016) On the detection of carbon monoxide as an anti-biosignature in exoplanetary atmospheres. Icarus 266:15–23
Wayne RP (1993) Chemistry of atmospheres, 2nd edn. Oxford University Press, Oxford
Webster CR, Mahaffy P, Atreya SK (2015) Mars methane detection and variability at gale crater. Science 412:415
Werner MW, Swain MR, Vasisht G et al (2016) Extension of ATLAST/LUVOIR’s capabilities to 5 μm or beyond. J Astron Telesc Instrum Syst 2:041205
Williams DM, Gaidos E (2008) Detecting the glint of starlight on the oceans of distant planets. Icarus 195:927–937
Woolf NJ, Smith PS, Traub WA et al (2002) The spectrum of Earthshine: a pale blue dot observed from the ground. Astrophys J 574:430–433
Wordsworth R, Pierrehumbert R (2014) Abiotic oxygen-dominated atmospheres on terrestrial habitable zone planets. Astrophys J Lett 785:1–4
World Meteorological Organization (WMO) (1995) Scientific assessment of ozone depletion: 1994. Report Number 37. WMO, Geneva
Yan F, Fosbury RAE, Petr-Gotzens MG et al (2015) High-resolution transmission spectrum of the Earth’s atmosphere-seeing Earth as an exoplanet using a lunar eclipse. Int J Astrobiol 14:255–266
Yang J, Cowan NB, Abbot DS (2013) Stabilising cloud feedback dramatically expands the habitable zone of tidally-locked planets. ApJL 771:2
Yung YL, DeMore WB (1999) Photochemistry of planetary atmospheres. Oxford University Press, Oxford
Zahnle K, Freedman RS, Catling DC (2011) Is there methane on Mars? Icarus 212:493–503
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Grenfell, J.L. (2019). Exoplanetary Biosignatures for Astrobiology. In: Cavalazzi, B., Westall, F. (eds) Biosignatures for Astrobiology. Advances in Astrobiology and Biogeophysics. Springer, Cham. https://doi.org/10.1007/978-3-319-96175-0_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-96175-0_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-96174-3
Online ISBN: 978-3-319-96175-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)