
Abstract
The exchange of nutrients among different species is one of the most complex, intriguing, and important factors that favored the development of certain microorganisms in evolutionary systems. As a direct result of this, microorganisms in natural system often co-exist with many others in an organized manner. From an engineering approach, the benefits of having multiple microorganisms in a bioprocess may be beneficial to a number of reasons, including favoring the preferred metabolic pathways of one organism, simultaneously by promoting other additional effects by another. Among the many examples of microbial communities or consortia, one of the most studied from a biological point of view are known as lichens—a combination of fungi and algae and/or cyanobacteria. Adding to the fact that over one fifth of all fungal species today known to be in its lichenized form, it is crucial to understand the hidden benefits that such combination can offer to biosystems engineers. Fungal-algal symbiotic relationships often offer significant and efficient exchange of gases, promoting a healthy growth, and can be directly applied towards the solution of a wide range of issues faced today by biorefineries. This chapter summarizes some of the applications of this novel, yet promising, concept, including algal harvesting, lipid accumulation, and bioremediation of wastewater, presenting also a brief history and perspectives on the concept.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
16.1 Introduction
The interaction between different species is a key driver for evolutionary purposes (Shong et al. 2012). The exchange of nutrientes among symbiotic organisms is one of the most important and most studied factors in microbial communities (Sabra et al. 2010). Despite innumerous advances over the past centuries, understanding how a species collaborates with another has been one of the hardest tasks in the biological realm of studies, and many reasons and mechanisms remain nuclear. Among the many types of mutual relationship between species, one of the most intriguing and omnipresente is found in what we know today as lichens. Lichens exist as a dynamic system between fungi and algae or cyanobacteria, and are widespread on the most diverse habitats on the Earth crust mainly for reasons of nutrient exchange, and therefore, for mutual survival (Rajendran and Hu 2016). Today, about 25% of all fungi species are known to be in their lichen form (Lutzoni et al. 2001), and not only they represent part of the geographic history of the world on their own, but they may be part of the next wave of discoveries in biological engineering.
The proper mechanism of lichenization is species-dependent (Chen et al. 2000; Lutzoni et al. 2001). The association of cyanobacteria or algae with fungi represent a successful nutritional mode in which a heterotrophic fungal species partner with a carbon dioxide-consuming and oxygen-emitting algae (Rajendran and Hu 2016). Such association allows both species to co-exist without the need to rely on “external” sources of energy, e.g., sunlight and other carbon sources. The overall evoluationary success of lichens, today estimated to cover roughly 10% of the terrestrial ecosystem, is hypothesized to play an important role in early development of terrestrial ecosystems, and have been reported to correlate properly with paleological events, such as global cooling (Quirk et al. 2015). Due to evolutionary and adaptation to different habitats, lichens have adapted themselves to a wide range of growth forms and habitat preferences. Lichens can be found as small crusts adhered to leaf surfaces in the tropics to large turfs in the Arctic (Park et al. 2015). Ascomycota, the group to which most lichenized fungi belong to, has been reported to have gained and lost lichenization preferences over the geological eras (Blanco et al. 2006). With recent developments in microalgal and fungal research and their potential applications in bioprocessing, the field of artificial, or laboratory-made lichens has become reality.
The recent boom in microalgae research over the past two decades has answered many questions over the many promising properties of microalgae in bioprocessing. A common bottleneck that still represents an enormous task for their use relies on the economic feasibility of the harvesting step (Reis et al. 2014). Though microalgae brings the potential to become a key driver to a bio-renewable economy, the operation facilities of microalgae production are still hampered by the energy-intensive or by the application of hazardous and recalcitrant chemicals in the harvesting process (Mata et al. 2010). Significant challenges to be overcome are also related to the water and nutrient requirements, and the ecological implications of massive invasion to the local biota. The harvesting process of microalgae remains a challenging task mainly due to the morphological properties of a microalgae cell. Differently from multi-cellular organisms, as fungi, or even large unicellular yeast cells, microalgae cells are small and have specific gravity values close to those of their culture medium (Pragya et al. 2013). Microalgae cells are usually cultured in suspension. The naturally occurring negative charge of a microalgae cell surface usually brings stability for the cells (Pragya et al. 2013). Microalgae cells are usually cultivated in dilute conditions, which generally yields a poor ratio of biomass to liquid, increasing not only the water footprint of such process, but also the downstream requirements. Conventional algae harvesting operations are dependent on centrifugation, flocculation, flotation, or filtration methods (Mata et al. 2010). Though there have been tremendous advances in the field of algae harvesting, this chapter will focus only on the processes related to the combination of algae and fungi, which will be better described in the further sections. As previously mentioned, other species, as fungi, are relatively easier to harvest than microalgae. With the proper combination of fungi and microalgae, proper cell attachment can occur, leaving a fungal biomass rich in microalgae cells on its surface, and a clear culture medium (Gultom and Hu 2013). The application of such concept is relatively new and requires further understanding in order to fully optimize the process. As of today, the process of co-culture, or co-harvesting, is limited to a few microalgal species (Zamalloa et al. 2017). As the mechanism of attachment is still unclear, research has pointed difficulties in obtaining totality of cell aggregation and in scaling up such process.
In summary, the application of microbial consortia, as is the case of artificial lichens, compared to monocultures can be advantageous from different perspectives from a biological engineering standpoint. Microbial consortia allow different species to share different synthetic functions, i.e., it does not necessarily exploit the potential of a single species to its maximum, but rather, divide the issues among different species. In addition, microbial consortia can aid the fine-tuning of different functionalities of a biorefinery. It does poses additional difficulties, however, as many interactions are not predictable from single-strain experiments. Thus, in order to fully understand the potential of microbial consortia in a biorefinery setting, one must understand the benefits and challenges of a given species and how such species collaborates with another one. This chapter introduces some of the applications of artificial lichens—one of the many groups defined as microbial consortia, and their potential in bioprocessing.
16.2 Characteristics and Formation of Fungal-Algal Systems
Naturally occuring algal biofilms are composed of three-dimensional, multispecies and multilayered structures, involving a complex consortia of heterotrophic and autotrophic organisms (Gauslaa 2014). The photoautotrophic microorganisms may often include unicellular macro- and microalgae in natural systems, and these organisms are characterizaed by their pigmentation, i.e., chlorophyll presence in their cells, which is able to capture solar energy and promote cellular functions (Campbell et al. 1998). The heterotrophic portion of these complex bodies can often include protoza, bacteria, and fungi, which colonize different zones within the biofilm structure aiming at the most suitable conditions for cell growth (McCune et al. 1997). The microorganisms present in these complex ecosystems are usually held together by matrices of EPS, which functionalize adhesion, retention of water and nutrients, diffusion, protection, and aggregation (Rajendran and Hu 2016). EPS can account for up to 90% of the dry biofilm mass and is mainly constituted by linear or branched heteropolysaccharides, which monomers include hexoses, pentoses, uronic acids, and deoxy-sugars (Nithyanand et al. 2015).
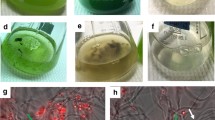
The cell surface of a biofilm material, such as the case of artificial lichens, is usually soft and covered with hydrated flexible macromolecules that facilitate adhesion between cells and from cells to surfaces (Gultom and Hu 2013). Algal and fungal cells produce surface macromolecules, which are often bridged. It has been proposed that specifically in the case of a fungal-algal biofilm, the formation of such complex structure is given by the following steps: (i) adhesion of fungal spores or germinating fungal spores and algae to a matrix, (ii) proliferation of mononucleated cells over the surface or the matrix, (iii) induction of fungal hyphal formation, inducing algal attachment on mycelia (Rajendran and Hu 2016). Another important morphological aspect is that fungal spores present crystalline-like layers, which prevent aggregation within themselves and with other strains, in such a way that the growing hyphae acts as the major promoter of bridge interactions between germinating spores and other species. Microscopically, lectin-carbohydrate is an important factor in the binding of algae and fungi. Particularly involving the role of lectin-carbohydrate is the presence of calcium ions, which act as a link between negatively-charged cell and extracellular nucleic acids, which enable cell-to-cell communication through electrostatic interactions (Zamalloa et al. 2017). Through unclear mechanisms to date, the weak binding of lectin-carbohydrate forms stable cell arrangements and increases the shear resistance.
The use of artificial matrices has been tested in the formation of lichen-type biofilms. Using a polypropylene spun and tape matrix, (Rajendran and Hu 2016) described the four major steps in the formation of such biofilm on a M. circinelloides and C. vulgaris setting: (i) preferential attachment of strains: fungal spores are known to be highly hydrophobic, thus, the preferential attachment is found on the tape yarn of the matrix within a few hours of the innoculation. It has also been observed that algae cells attach to fungal cells within moments after the spore germination. Within such system, it has been observed that the polypropylene fraction acts more as a mechanical support than a growing support. The surface characteristics of the hydrophobic material also have effects on the rate and on the extent of attachment by fungal spores, on a sense that rougher and more hydrophobic materials will likely develop biofilms more rapidly. (ii) germination of fungal spores: in which the fungal spores attached to the polypropylene matrix propagate along its length, and algae cells attach to their surface. Since M. circinelloides is a dimorphic fungus, it can either grow as yeast or filamentous. It is unclear, though, what type of dimorphism M. circinelloides presents at the initial stages of growth. Microscopic evidence shows that algal cells are covered by fungal cells, with little algal growth. (iii) elongation and branching: after the fungal cells have filled in the space in-between the perpendicular rows of polypropylene spun and tape yarn by its hyphae form, algal cells start to autotrophically develop in the liquid phase. It is hypothesized that the lichen formation of M. circinelloides is similar to the one reported by Candida albicans, in which yeast and hyphal cells comprise the fungal network (Finkel and Mitchell 2011). (iv) mature lichen biofilm: as the thickness of the biofilm increases, algal cells attach to the fungal mycelia. EPS concentrations increase and as pointed above, may significantly contribute to the cell cohesion to the surface.
16.3 Applications in Microalgae Harvesting
The cultivation of autotrophic algae is highly promising towards the development of novel bioprocesses (Reis et al. 2014). The ability to utilize carbon dioxide as carbon source is renowned as a ‘win-win’ situation, in which effluent gases, e.g. flue gas, can be directly used as feedstock for growing microalgae, at the same time reducing the treatment costs for such effluent. However, in order to fully establish an autotrophic microalgae biorefinery, the cell harvesting step remains as the most significant obstacle on the downstream processing. Autotrophic microalgae cells usually have low density, small size, and quasi-colloidal properties (Zhang et al. 2010). A number of different harvesting techniques have been applied, such as gravity sedimentation, centrifugation, flocculation and flotation, ultrasonic aggregation, and filtration, and combinations of the aforementioned operations (Reis et al. 2014). It has been reported that the utilization of most of these approaches are feasible when it comes to the extraction of high-value chemicals from microalgal biomass, however, for the production of commodity and fuel chemicals, harvesting hinders the economic feasibility. Harvesting fungi, on the other hand, is an easier operation when compared to microalgae. Filamentous fungi grow from a hyphae structure, i.e., a polarized pattern in the form of filaments (Xia et al. 2011). The elements present in a fungal hyphae outgrow of single cells or spores as multinucleate tubes, which moves within a hypha towards the hyphal tip (Balmant et al. 2015). The hyphal tip is the only place where the hyphae grows. From the hyphal tip, which is place responsible for additional growth, diversification of hyphal growth is given, forming a network of hyphae, or also known as mycelia. Depending on the conditions applied to a given submerged fungal growth, mycelia can form pellets, which are spherical or elipsoidal masses of hyphae. Such pellets can vary significantly from loose to compact structures, which form a clear phase separated from the submerged growth, easily harvested after the fermentation process (Xia et al. 2011).
A method to harvest microalgae has been disclosed based on the utilization of fungal pellets as adsorbent matrix and has been reproduced within different studies (Gultom et al. 2014). The fungal-algal co-pelletization process can be considered as a type of co-culture, in which the fungi co-exist with microalgae cells. In the cases involving autotrophic microalgae, without any external carbon source to the system, the symbiotic relationship of algae and fungi has been reported to be similar to the one found in natural lichens (Gultom et al. 2014). The fungal-algal co-pelletization is reported to be similar to a flocculation process, in which the hyphae acts as flocculants to the microalgae cells (Ummalyma et al. 2017). Though the mechanism of such attraction remains unclear to date, it can possibly be explained through the understanding of zeta potential at each cell surface (Zamalloa et al. 2017). It is known that the surface cell charge and size of microalgae are the main factors for its suspension stability. Under neutral pH, microalgae is usually characterized to have a negative charge due to the presence of carboxylic, phosphoric, and hydroxyl groups on the proteic cell surface (Zamalloa et al. 2017). The zeta potential is usually within the range of −10 to −35 mV (Henderson et al. 2008). Fungi, on the other hand, if grown on acid conditions, usually possess positive zeta potentials (Holder et al. 2007). It has been reported for Aspergillus flavus measurements of +46.1 mV, and for Beauveria bassiana, +22 mV (Holder et al. 2007). Therefore, it is hypothesized that an electrostatic attraction between fungi and algae is a major factor for this phenomenon to happen. Such phenomena is similar to the one found in bioflocculation, a concept studied for decades, using bacteria and algae. The principles of bioflocculation are linked to the production of bioflocculant chemicals on the cell surface of different bacteria species.
It has been reported for a system of A. niger and C. vulgaris that within the best culturing conditions, more than 90% of microalgal cells can easily be harvested by using pelletized fungal cells (Gultom et al. 2014). In such system, an organic carbon source was necessary in order to sustain the fungal growth, and it has been observed that such concept can be applied to C. vulgaris under hetero- or photoautotrophic modes. Regarding the external carbon source, glycerol and acetate were reported to be successfully used, both which were functional, but with lower results in terms of harvesting efficiency than glucose. Competition was clearly observed regarding the nutrients in heterotrophic conditions, but the symbiotic results indicated greater biomass accumulations than the axenic cultures (Gao et al. 2015; Gultom et al. 2014). Yields no higher than 75% of attachment have been reported on large scale operations, and the most successful application of such concept in the literature is related to a biofilm membrane photobioreactor, utilizing a submerged membrane, which was able to harvest Chlorella vulgaris at an efficiency of 72.4% (Gao et al. 2015).
16.4 Lipid Accumulation
The production of lipids by oleaginous fungi is a promising route to reduce the need of oil feedstock for fuel and chemical applications (Rivaldi et al. 2017). Similarly to oleaginous fungi, several algae strains are also known to accumulate high contents of lipids, which have been reported to be successfully used as feedstock for biodiesel and other applications (Reis et al. 2014). Both fungi and algae can accumulate lipids with high content of fatty acids, thus, being considered a substitute for plant-based oils for biodiesel production. Fungi and algae have the ability to grow rapidly in bioreactors, and are able to utilize a wide range of lignocellulosic waste material as nutrient source, as nitrogen and phosphorus. Though the cell wall of fungi can represent additional challenges to the extraction of lipids, especially due to their extensively cross-linked chitin composition, the purpose of this chapter remains within the scope of presenting the opportunities from utilizing lichens, and technical challenges of single-species cultures will not be thoroughly discussed. Therefore, the beneficial physical and physiological interactions among different microbial communities can represent a promising strategy for next-generation lipid production. It may seem counter-intuitive to try to consolidate multiple microorganisms into a single processing if one thinks about consolidated bioprocessing (CBP) approaches. CBP is often described as the engineering behind the consolidation of multiple pathways and operations to the least amount of processing possible, usually into a single organism (Olson et al. 2012). Among the drawbacks CBP is known for, the metabolic bottlenecks stands as the most critical ones (Banerjee et al. 2010). Consequently, in applications like lipid production, a mixed culture may be a solution for the selection of microalgae and fungi species and strains that are more suitable to perform one task of the overall process, instead of many. Therefore, specially in lipid producing and accumulating strains, the application of microbial consortia can therefore deviate the burden of introducing new functionalities to strains, and rather simply improving naturally-occurring metabolic pathways.
Lipid accumulation in microorganisms is regulated by starvation of a nutrient, usually N (Ratledge and Wynn 2002). Experiments have been performed to demonstrate that nitrogen depletion started earlier in a co-culture system when compared to axenic cultures. In such case, various other metabolites have also been enhanced, such as peptides, amino acids, and nitric oxide. The strategy of early nitrogen depletion can also be thought of being an engineering approach, as it shortens the culture time. Systems like these tend to increase the complexity of a given study, and understanding the competition effects may be necessary. In a competitive system of Monoraphidium sp. and Chlorella sp., the microalgae cells produced more C18 fatty acids that inhibited the growth of Monoraphidium cells (Zhao et al. 2014). The ‘survival’ strategy adopted by Chlorella cells is in accordance to other studies that claim that C. vulgaris is able to release inhibitory compounds when co-cultivated with other microorganisms (Cheirsilp et al. 2011). In addition, the utilization of mixed cultures may be beneficial in systems with nutrient sources derived from lignocellulosic feedstock. As an example, as R. toruloides is known to be a poor producer of inulases and amylases, it was grown with Saccharomycopsis fibuligera, a known producer of both enzymes, in a cassava starch medium (Dostálek and Häggström 1983). The result was an efficient and low-cost method of producing single cell oil, achieving 64.9% of dry cell weight as lipids and cell concentrations as high as 20.1 kg m−3. Similarly, the use of Rhodotorula mucilaginosa and P. guilliermondii was tested in a 2.0%-inulin medium, achieving lipid concentrations as high as 53.16% on a weight basis (Zhao et al. 2011).
In order to establish a lipid producing system based on a microbial consortia, one must take in consideration that the strains selected should have high lipid productivities, with similar nutrient profiles, preferably obtained from complementary niches, which may help to reduce the competition between species. In a study with A. fumigatus and different microalgae strains, it was concluded that lipid production in the fungal-algal pellets showed complex profiles, to which three main factors are attributed: (i) total biomass production, (ii) lipid concentrations in fungal and algal cells before and during cultivation process, and (iii) harvesting efficiency of A. fumigatus pellets (Wrede et al. 2014). It has also been reported that after 48 h of co-culturing A. fumigatus with oleaginous microalgae, which were defined with lipid concentrations greater than 10% of biomass dry weight, lipid concentration in the formed pellets were lower than in mono-cultured algae, but similar or greater than those observed in mono-cultured A. fumigatus. Regarding fatty acid composition, the fungal contribution led to high concentrations of C16:0, C18:0, C18:1, and C18:2 (Wrede et al. 2014).
It is important to understand the feasibility of using fungal-algal based lipids in the production chain of biofuels. In this case, considering biodiesel as the main product from microbial lipids, the fungal-algal biomass can be used as feedstock usually through extraction of lipids and followed by transesterification, or by in situ transesterification followed by extraction of biodiesel from biomass (Li et al. 2008). Extraction processes are generally energy intensive and costly, since they involve the use of solvents in traditional processes and significant consumption of electricity. The Department of Energy (US DOE) proposed the following benchmark for biofuel production from algae, which could be extrapolated to the use of fungal-algal biomass: the extraction should consume no more than 10% of the total energy produced on a daily basis (Liu et al. 2012). Some microalgal species have additional morphological barriers to energy-efficient extraction processes, as is the case of Nannochloropsis occulata, which have very tough cell walls (Borges et al. 2011). Similarly to N. occulata, the production of fungal-algal platforms involves an increase in EPS, which as discussed in other parts of this chapter, increase significantly the toughness of the overall structure, posing additional difficulties to the extraction process. The transesterification process per se can be considered less challenging than the extraction steps, as conversion yields in the literature are usually close to totality in acid-, base-, and enzyme-catalyzed systems (Meng et al. 2009).
16.5 Bioremediation Applications
The use of algal-based systems in wastewater treatment has been extensively studied over the past decades (Mallick 2002), and has been successfully applied in a number of large industrial scale operations (Craggs et al. 2012). Despite the advances, there are technical limitations, which limit the application of algal-based systems. As of today, applications as reverse osmosis are being used worldwide in order to meet demand for water reclamation in intensive urban systems. Reverse osmosis, however, generates saline concentrate streams, which contain almost all of the nutrients and contaminants at elevated concentrations (Dialynas et al. 2008). The growing need to explore cost-effective treatment operations is an opportunity for biological-based systems to be fully explored in order to reduce environmental and health risks that traditional methods, as reverse osmosis, can often bring. The concept of utilizing wastewater treatment methods based on algae-based biofilms is that they offer a simple, yet energy-efficient technology for sorption of key nutrients, as N and P, and offer an easy and robust separation from the wastewater bulk (Mehta and Gaur 2005). The use of co-culture to treat various types of wastewater is a developing field of study. As mentioned in the previous sections, the combination of different microorganisms in a complex media may significantly increase the biotechnological effect of a given culture. Recently, the use of fungal-algal pellets has been used on reducing the toxicity of different effluents. Fungal-algal pellets of Aspergillus sp. and C. vulgaris have been used to treat centrate (Wrede et al. 2014), i.e. a liquid stream generated through centrifugation of activated sludge, and diluted swine manure. The first one contained about 50 mg L−1 of ammonia and phosphates, while the later, a higher concentration of ammonia (89 mg L−1) and lower concentration of phosphates (1.8 mg L−1). The researchers observed that in both conditions, the pellets were able to significantly decrease N and P, as well as color, producing a practically colorless suspension in about 24 h (Wrede et al. 2014). Similar results were observed by the utilization of A. fumigatus with C. protothecoides and Tetraselmis suecica using digestate from swine manure, with concentration uptakes of as high as 73.9% of ammonia and 55.6% of phosphates. It has also been observed that the uptake of key nutrients led up to 1.7 fold increase in biomass production after 48 h of treatment. The use of A. fumigatus with T. chuii has also been tested with diluted swine wastewater, diverted from a lagoon system, in which up to 86% of ammonia and 69% of phosphate were uptaken in just 48 h of treatment (Wrede et al. 2014). In addition, pellets from A. fumigatus and Synechocystis sp. were grown on digestate wastewater from a municipal treatment facility (Miranda et al. 2015). For the latter study, it has been reported that in relatively concentrated suspensions (25% of original wastewater concentration), pellets were able to reduce ammonia-N to levels close to 90% (164.3 to 18.2 mg L−1) and phosphate-P to levels as high as 75% (38.7 to 9.8 mg L−1). Interestingly, the synergystic effect was higher than in the axenic cultures, which achieved results no higher than 52% for ammonia-N, and 45% for phosphate-P. Still regarding this study, biomass yields were increased to 2.3-fold compared to the control system, and lipids were increased by 1.5-fold (Miranda et al. 2015).
An experimental setup was also established using corn to ethanol co-products as culture medium (Rajendran et al. 2017). The major difference between corn to ethanol co-products, in particular, stillage water, and agricultural effluents is the concentration of nutrients (Reis et al. 2017). Particularly speaking, Condensed Distillers Solubles (CDS) has at least one order of magnitude on concentrations of P and N than diluted swine manure and other hydrolysates used by (Miranda et al. 2015). In a study with diluted CDS, a biofilm composed by Mucor sp. and C. vulgaris was tested, and results have shown that the microbial biomass achieved concentrations as high as 9.358 g L−1 at a 12-day culture, being mostly composed by fungal cells (97.2% of the mature biofilm) (Rajendran et al. 2017). Bioremediation of viscous materials like CDS should also take in consideration the presence of solids attached to the biofilm. Particularly in the aforementioned study, the attached solids decreased from 6.304 to 3.349 g L−1 at 4 days and 12 days of culture respectively. Nutrient wise, the biofilm was able to retain 55.7 and 74% of the original concentrations of P (818 mg L−1) and N (924 mg L−1), respectively. Results also show a decrease in Chemical Oxygen Demand (COD) levels up to 65.6% (Rajendran et al. 2017). As other studies have also shown, the combination of different cultures achieved higher biomass concentrations than the axenic cultures under all the conditions tested.
It is important to note that most of these applications of co-culturing microorganisms are relatively new, though the concept of applying lignocellulosic waste as nutrient source to fungi and algae has been a key topic of research for decades (Sánchez 2009). Fungal treatments are known for their ability to digest lignocellulosic biomass, especially by the production of active enzymes including cellulases and hemicellulases, pectinases, phytases, laccases and manganese peroxidases. Understanding the fungal role in producing such enzymes may be an important factor when upscaling bioremediation and processes involving accumulation of commodity biomass. For example, in order to potentially decrease the costs of fungal-assisted bioflocculation using A. fumigatus as the fungal species, rice straw could succesfully be used as partial replacement of nutrients for the microorganisms involved due to the extensive production of endo- and exo-glucanases, beta-glucosidases, laminarinases, lichenases, xylanases, and pectin lyases, which are able to hydrolyze rice straw and provide a healthy system for microbial growth (Zhang et al. 2011). It has been observed in the cited study that in a media containing wheat straw as the sole source of carbon, fungal pellets showed similar growth rates and sizes to the control experiments, i.e., those grown on glucose (Muradov et al. 2015). One reported drawback, however, was that fungal pellets when grown on wheat straw biomass release chemicals that are inhibitory for algal growth (Muradov et al. 2015). Therefore, the application of lignocellulosic materials for fungal-assisted algal flocculation needs to be further studied in order to reduce the production of anti-algal chemicals.
16.6 Perspectives on the Development of Lichen-Based Bioprocesses
Though the application of microbial consortia in the reported literature results mostly considers the results of a ‘single’ organism, i.e., as if the mutualistic organisms acted as they were one, it is important to understand that the characteristics of a given biomass material generated from artificial lichens can be differed according to the composing species. It is true that mixed cultures generate a single biomass type, which is close to being homogeneously distributed across its volume, except in cases with gradient conditions of growth, e.g. light and nutrient. The resulting biomass can be roughly estimated as the weighted average of the composing organisms contribution (Wrede et al. 2014). It has been reported that particularly in lichens, the fungal weight contribution exceeds greatly the algal composition (Rajendran et al. 2017; Rajendran and Hu 2016); therefore, one must take in account that the fungal composition may largely exceed the algal contribution to the overall biomass. Understanding this concept is particularly of interest in cases in which a compound is either fungal or algal-based, and a co-culture is used to produce it. Another important factor to be taken in consideration is that co-culture operations lead to more robust systems, which may reduce the energy required for sterilization (Muradov et al. 2015). Though single-cell cultures can be preferred due to their reproducibility, an ideal co-culture system should be able to provide support to the development of natural biological characteristics of each organism in order to enhance their cellular interactions.
Innovative ways of co-cultivating microorganisms can lead to outcomes that are close to unachievable in non-gene modified single-cell cultures. Examples regarding lipid accumulation are: (i) increase in release of free fatty acids into the extracellular medium; (ii) production of enzymes that are beneficial to the other species, such as inulases and amylases in a Saccharomycopsis fibuligera and R. toruloides in a cassava starch medium (Gen et al. 2014). In this regard, the choice of a consortium of lipid-producing strains may also pose challenges on the rheology of submerged cultures and lipogenic capacities. From an engineering perspective, the enhancement of mass transfer coefficients is a major challenge improving production yields (Boopathy 2000). Though it may seem counter-intuitive to think that aeration may be necessary in systems with algae, usually fungal consumption of oxygen is higher than production by algae, therefore, oxygenation and oxygen mass transfer is necessary for an efficient growth and production of desired metabolites. In high-cell density cultures, growth is dependent on the mass transfer rate. In such sense, diffusion limitations results in lower productivities. Therefore, an adequate reactor system that can provide high mass transfer rates is usually necessary in order to establish a bioprocess like the one discussed (Boopathy 2000). Even though high mass transfer rates are needed, one also must take in account the shear and the stress suffered by the cells inside a bioreactor. As a bottomline, these and other issues should be taken in consideration when addressing the commercial viability of a fungal-algal medium.
As is the case with many other emerging technologies, many questions are still unanswered. Usually experiments in co-culture systems are carried out by having two or more controls using monocultures in a pure culture system, in which the strains are submitted to identical conditions, e.g. operating conditions, substrate concentration, medium type, etc., in order to mimic the biological environment and their reactions to the growth. Co-culture assays, however, may impose competitiveness among different cultures, and results may not be easy to interpret and obtain ‘logical’ data from. Competition may benefit one species in detriment of the other, or may harm both. A clear example is found in the case of lipid-producing strains, in which competition can clearly impact the lipogenesis process. In order to overcome such difficult comprehension, a handful of techniques can be applied. One of them is molecular biology and genetics, in order to establish the metabolic guidelines and function of genes involved in microbial lipid assimilation, accumulation, and processing. Therefore, thoroughly understanding the strain abilities, kinetics, and biological mechanisms has become a focus for researchers, not only to fundamentally develop mechanisms for enhancement of processes, but also to avoid the misinterpretation of results. Among the techniques and mechanisms researchers have used to better understand the cell-to-cell communication are cross feeding and metabolite exchange. Metagenomic sequencing is another proeminent tool for steering the functionalities of microbial communities, especially by describing cellular regulations at metabolic levels and by drawing a simplified microenvironment of a given mixed culture (Morfopoulou and Plagnol 2015).
Finally, additional challenges are often found not only in the fermentation itself, but also in the downstream process. Downstream processes are usually considered high-cost and challenging depending on the cultivation process. For example, continuous centrifugation to separate lipid-producing cells from the fermentation broth is one of the most considered alternatives to the separation of biomass from the fermentation broth. However, such process is highly intensive and often present technical challenges. Other options, as is the case of flocculation and filtration may also present cost inhibition to an overall biotechnological process (Milledge and Heaven 2013). In the case of intracellular metabolites, as is usually the case for lipid bodies, following the cell separation steps, cell lysis and solvent extraction are also a suggested route. Both processes are well understood and reviewed in the literature, however as it is pointed out, chemical treatment can also pose environmental risks. In addition, many extraction processes rely on dry biomass, which pose additional energy requirements for drying the cells prior to extraction.
16.7 Conclusion and Future Perspectives
This chapter comes in the context that in a world with growing population, increasing urban densities, and increased footprint on natural resources, alternatives must be developed in order to ensure a sustainable future. Artificial lichens can be a promising alternative to some of the challenging tasks of the nearby future. The extensive use of plantable lands and the growing consumption of freshwater by energy crops have been an incentive for next generation bioenergy and a bio-based economy. As pointed by (Langeveld et al. 2010), the selection criteria for suitable alternatives in a bio-based economy must be as follows: (i) strains with high growth rates and increased biomass production, (ii) strains that are able to produce high concentrations of energy-dense molecules, e.g., lipids, (iii) high harvesting index and ease of harvesting, (iv) capacity to grow on marginal and low-value lands, with lack of competition with existing agricultural crops, (v) low dependence on freshwater, (vi) low costs for growth and harvest, (vii) production of value-added products. Generally speaking, despite being an early concept, novel tools for analysis and engineering of microbial communities have been developed, including those present in synthetic biology and metabolic engineering. A common concept that is sometimes overlooked is the process of obtaining a reliable community behavior and reproducible results. Studies in the area should take advantage of the innumerous advances in the intersection of molecular biology and biosystems engineering in order to fully develop solutions for the problems in bioprocessing, bioremediation, and even biosensing, and other many applications in which answers are urgely needed to meet the demands of a sustainable future.
References
Balmant W, Sugai-Guérios MH, Coradin JH, Krieger N, Junior AF, Mitchell DA (2015) A model for growth of a single fungal hypha based on well-mixed tanks in series: simulation of nutrient and vesicle transport in aerial reproductive hyphae. PLoS ONE 10(3):e0120307
Banerjee S, Mudliar S, Sen R, Giri B, Satpute D, Chakrabarti T, Pandey RA (2010) Commercializing lignocellulosic bioethanol: technology bottlenecks and possible remedies. Biofuels, Bioprod Biorefin 4(1):77–93
Blanco O, Crespo A, Ree RH, Lumbsch HT (2006) Major clades of parmelioid lichens (Parmeliaceae, Ascomycota) and the evolution of their morphological and chemical diversity. Mol Phylogenet Evol 39(1):52–69
Boopathy R (2000) Factors limiting bioremediation technologies. Biores Technol 74(1):63–67
Borges L, Morón-Villarreyes JA, D’Oca MGM, Abreu PC (2011) Effects of flocculants on lipid extraction and fatty acid composition of the microalgae Nannochloropsis oculata and Thalassiosira weissflogii. Biomass Bioenerg 35(10):4449–4454
Campbell D, Hurry V, Clarke AK, Gustafsson P, Öquist G (1998) Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol Mol Biol Rev 62(3):667–683
Cheirsilp B, Suwannarat W, Niyomdecha R (2011) Mixed culture of oleaginous yeast Rhodotorula glutinis and microalga Chlorella vulgaris for lipid production from industrial wastes and its use as biodiesel feedstock. New Biotechnol 28(4):362–368
Chen J, Blume H-P, Beyer L (2000) Weathering of rocks induced by lichen colonization—a review. CATENA 39(2):121–146
Craggs R, Sutherland D, Campbell H (2012) Hectare-scale demonstration of high rate algal ponds for enhanced wastewater treatment and biofuel production. J Appl Phycol 24(3):329–337
Dialynas E, Mantzavinos D, Diamadopoulos E (2008) Advanced treatment of the reverse osmosis concentrate produced during reclamation of municipal wastewater. Water Res 42(18):4603–4608
Dostálek M, Häggström MH (1983) Mixed culture of Saccharomycopsis fibuliger and Zymomonas mobilis on starch-use of oxygen as a regulator. Appl Microbiol Biotechnol 17(5):269–274
Finkel JS, Mitchell AP (2011) Genetic control of Candida albicans biofilm development. Nat Rev Microbiol 9(2):109
Gao F, Yang ZH, Li C, Zeng GM, Ma DH, Zhou L (2015) A novel algal biofilm membrane photobioreactor for attached microalgae growth and nutrients removal from secondary effluent. Bioresour Technol, 179. https://doi.org/10.1016/j.biortech.2014.11.108
Gauslaa Y (2014) Rain, dew, and humid air as drivers of morphology, function and spatial distribution in epiphytic lichens. The Lichenologist 46(1):1–16
Gen Q, Wang Q, Chi Z-M (2014) Direct conversion of cassava starch into single cell oil by co-cultures of the oleaginous yeast Rhodosporidium toruloides and immobilized amylases-producing yeast Saccharomycopsis fibuligera. Renewable Energy 62:522–526
Gultom SO, Hu B (2013) Review of microalgae harvesting via co-pelletization with filamentous fungus. Energies 6(11):5921–5939
Gultom SO, Zamalloa C, Hu B (2014) Microalgae harvest through fungal pelletization—co-culture of Chlorella vulgaris and Aspergillus niger. Energies 7(7):4417–4429
Henderson RK, Parsons SA, Jefferson B (2008) Successful removal of algae through the control of zeta potential. Sep Sci Technol 43(7):1653–1666
Holder DJ, Kirkland BH, Lewis MW, Keyhani NO (2007) Surface characteristics of the entomopathogenic fungus Beauveria (Cordyceps) bassiana. Microbiology 153(10):3448–3457
Langeveld JWA, Dixon J, Jaworski JF (2010) Development perspectives of the biobased economy: a review. Crop Sci 50(Suppl1):S-142–S-151
Li Q, Du W, Liu D (2008) Perspectives of microbial oils for biodiesel production. Appl Microbiol Biotechnol 80(5):749–756
Liu X, Clarens AF, Colosi LM (2012) Algae biodiesel has potential despite inconclusive results to date. Biores Technol 104:803–806
Lutzoni F, Pagel M, Reeb V (2001) Major fungal lineages are derived from lichen symbiotic ancestors. Nature 411(6840):937
Mallick N (2002) Biotechnological potential of immobilized algae for wastewater N, P and metal removal: a review. Biometals 15(4):377–390
Mata TM, Martins AA, Caetano NS (2010) Microalgae for biodiesel production and other applications: a review. Renew Sustain Energy Rev 14(1):217–232
McCune B, Dey J, Peck J, Heiman K, Will-Wolf S (1997). Regional gradients in lichen communities of the southeast United States. Bryologist, 145–158
Mehta SK, Gaur JP (2005) Use of algae for removing heavy metal ions from wastewater: progress and prospects. Crit Rev Biotechnol 25(3):113–152
Meng X, Yang J, Xu X, Zhang L, Nie Q, Xian M (2009) Biodiesel production from oleaginous microorganisms. Renewable Energy 34(1):1–5
Milledge JJ, Heaven S (2013) A review of the harvesting of micro-algae for biofuel production. Rev Environ Sci Biotechnol 12(2):165–178
Miranda AF, Taha M, Wrede D, Morrison P, Ball AS, Stevenson T, Mouradov A (2015) Lipid production in association of filamentous fungi with genetically modified cyanobacterial cells. Biotechnol Biofuels 8(1):179
Morfopoulou S, Plagnol V (2015) Bayesian mixture analysis for metagenomic community profiling. Bioinformatics 31(18):2930–2938
Muradov N, Taha M, Miranda AF, Wrede D, Kadali K, Gujar A, Stevenson T, Ball AS, Mouradov A (2015) Fungal-assisted algal flocculation: application in wastewater treatment and biofuel production. Biotechnol Biofuels 8(1):24
Nithyanand P, Shafreen RMB, Muthamil S, Pandian SK (2015) Usnic acid inhibits biofilm formation and virulent morphological traits of Candida albicans. Microbiol Res 179:20–28
Olson DG, McBride JE, Shaw AJ, Lynd LR (2012) Recent progress in consolidated bioprocessing. Curr Opin Biotechnol 23(3):396–405
Park CH, Kim KM, Elvebakk A, Kim OS, Jeong G, Hong SG (2015) Algal and fungal diversity in Antarctic lichens. J Eukaryot Microbiol 62(2):196–205
Pragya N, Pandey KK, Sahoo PK (2013) A review on harvesting, oil extraction and biofuels production technologies from microalgae. Renew Sustain Energy Rev 24:159–171
Quirk J, Leake JR, Johnson DA, Taylor LL, Saccone L, Beerling DJ (2015) Constraining the role of early land plants in Palaeozoic weathering and global cooling. Proc R Soc B Biol Sci 282:1813. https://doi.org/10.1098/rspb.2015.1115
Rajendran A, Hu B (2016). Mycoalgae biofilm: development of a novel platform technology using algae and fungal cultures. Biotechnol Biofuels 9(1):112. https://doi.org/10.1186/s13068-016-0533-y
Rajendran A, Fox T, Hu B (2017) Nutrient recovery from ethanol co‐products by a novel mycoalgae biofilm: attached cultures of symbiotic fungi and algae. J Chem Technol Biotechnol 92(7):1766–1776
Ratledge C, Wynn JP (2002) The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv Appl Microbiol 51:1–52
Reis CER, de Souza Amaral M, Loures CCA, da Rós PCM, Hu B, Izário Filho HJ, de Castro HF, Gianesella SMF, Silva MB (2014). Microalgal feedstock for bioenergy: opportunities and challenges. In: Biofuels in Brazil, Springer, pp 367–392
Reis CER, Rajendran A, Hu B (2017) New technologies in value addition to the thin stillage from corn-to-ethanol process. Rev Environ Sci Biotechnol 16(1):175–206
Rivaldi JD, Carvalho AKF, da Conceição LRV, de Castro HF (2017) Assessing the potential of fatty acids produced by filamentous fungi as feedstock for biodiesel production. Prep Biochem Biotechnol 47(10):970–976
Sabra W, Dietz D, Tjahjasari D, Zeng AP (2010) Biosystems analysis and engineering of microbial consortia for industrial biotechnology. Eng Life Sci 10(5):407–421
Sánchez C (2009) Lignocellulosic residues: biodegradation and bioconversion by fungi. Biotechnol Adv 27(2):185–194
Shong J, Diaz MRJ, Collins CH (2012) Towards synthetic microbial consortia for bioprocessing. Curr Opin Biotechnol 23(5):798–802
Ummalyma SB, Gnansounou E, Sukumaran RK, Sindhu R, Pandey A, Sahoo D (2017) Bioflocculation: an alternative strategy for harvesting of microalgae–an overview. Bioresour Technol
Wrede D, Taha M, Miranda AF, Kadali K, Stevenson T, Ball AS, Mouradov A (2014) Co-cultivation of fungal and microalgal cells as an efficient system for harvesting microalgal cells, lipid production and wastewater treatment. PLoS ONE 9(11):e113497
Xia C, Zhang J, Zhang W, Hu B (2011) A new cultivation method for microbial oil production: cell pelletization and lipid accumulation by Mucor circinelloides. Biotechnol Biofuels 4(1):15
Zamalloa C, Gultom SO, Rajendran A, Hu B (2017) Ionic effects on microalgae harvest via microalgae-fungi co-pelletization. Biocatal Agric Biotechnol 9:145–155
Zhang X, Hu Q, Sommerfeld M, Puruhito E, Chen Y (2010) Harvesting algal biomass for biofuels using ultrafiltration membranes. Biores Technol 101(14):5297–5304
Zhang F, Rodriguez S, Keasling JD (2011) Metabolic engineering of microbial pathways for advanced biofuels production. Curr Opin Biotechnol 22(6):775–783
Zhao C-H, Chi Z, Zhang F, Guo F-J, Li M, Song W-B, Chi Z-M (2011) Direct conversion of inulin and extract of tubers of Jerusalem artichoke into single cell oil by co-cultures of Rhodotorula mucilaginosa TJY15a and immobilized inulinase-producing yeast cells. Biores Technol 102(10):6128–6133
Zhao P, Yu X, Li J, Tang X, Huang Z (2014) Enhancing lipid productivity by co-cultivation of Chlorella sp. U4341 and Monoraphidium sp. FXY-10. J Biosci Bioeng 118(1):72–77
Acknowledgements
Authors are grateful to the financial support provided by MnDRIVE, LCCMR, CAPES (Process Number 13252/13-5) and to Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP (Process Numbers 17/12908-8 and 16/10636-8).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Rodrigues Reis, C.E., Rajendran, A., Silva, M.B., Hu, B., de Castro, H.F. (2018). The Application of Microbial Consortia in a Biorefinery Context: Understanding the Importance of Artificial Lichens. In: Singh, O., Chandel, A. (eds) Sustainable Biotechnology- Enzymatic Resources of Renewable Energy. Springer, Cham. https://doi.org/10.1007/978-3-319-95480-6_16
Download citation
DOI: https://doi.org/10.1007/978-3-319-95480-6_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-95479-0
Online ISBN: 978-3-319-95480-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)