
Abstract
The ubiquitous adenine nucleoside adenosine (Ado), which plays an important role in cellular energetics, is released from cells under physiologic and pathophysiologic conditions. Another source of extracellular Ado is rapid degradation of extracellular adenosine 5′-triphosphate (ATP) by ectoenzymes. Extracellular Ado acts as an autocrine and paracrine agent by the activation of G protein-coupled cell surface receptors (GPCRs), designated as A1, A2A, A2B, and A3. Almost four decades ago, published data have indicated that Ado could play a role in immune-mediated histamine release from pulmonary mast cells. Since then, numerous studies have indicated that Ado’s signal transductions are involved in various pulmonary pathologies including asthma and COPD. This chapter is a succinct review of recent studies in this field.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
18.1 Introduction
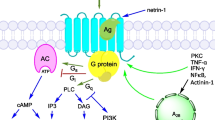
The ubiquitous adenine nucleoside adenosine (Ado) plays a major role in cellular metabolism and energetics. The levels of extracellular Ado are determined by its release from cells under physiologic and pathophysiologic conditions as well as the degradation of extracellular adenosine 5′-triphosphate (ATP) by ectoenzymes, CD39 and CD73 in particular (Zimmermann 2000; Zimmermann et al. 2012). Extracellular Ado is eliminated from the extracellular space by ecto-adenosine deaminase (ADA) (Franco et al. 1997) and active Ado transporters that transport Ado into cells (Thorn and Jarvis 1996). Extracellular Ado acts as an autocrine and paracrine agent, the effects of which are mediated by four different G protein-coupled receptors: A1, A2a, A2b, and A3 (A1AdoR, A2aAdoR, A2bAdoR, and A3AdoR, respectively). These receptors, which are expressed to various degrees by different cell types throughout the body including the lungs (Burnstock et al. 2012; Zhou et al. 2009), manifest variable affinity to Ado (Fredholm et al. 2001). AdoR are coupled to cascades of intracellular cellular-response pathways, mainly through the activation or inhibition of adenylate cyclase and the subsequent alteration of intracellular cyclic adenosine monophosphate (cAMP) levels (Karmouty-Quintana et al. 2013). The activation of AdoR can lead to both pro-inflammatory and anti-inflammatory effects, depending on the type of receptor and the experimental setting (Schepp and Reutershan 2008).
In the early 1980s, Holgate and his colleagues have shown for the first time that inhaled Ado induces bronchoconstriction in asthmatic but not healthy human subjects and that this action was mediated mainly by histamine released from lung mast cells and not by a pulmonary-pulmonary central vagal reflex triggered by Ado (Cushley et al. 1983; Mann et al. 1985; Church and Holgate 1993). Since then, data obtained in numerous studies have shown that Ado plays important roles in pulmonary physiology and pathophysiology (Caruso et al. 2006; Polosa and Blackburn 2009; Caruso et al. 2009). One characteristic of acute lung injury (ALI) is elevated Ado levels in the lung, which activate anti-inflammatory and tissue-protective mechanisms (Zhou et al. 2009; Karmouty-Quintana et al. 2013; Gonzales et al. 2014). Specifically, initial activation of AdoR blunts the production of multiple cytokines, decreases inflammatory cell infiltration, and preserves pulmonary vascular barrier function (Eckle et al. 2009). Similarly, Ado exerted a protective anti-inflammatory effect in an in vitro model of lung transplantation/ischemia-reperfusion model (Smail et al. 2016). In contrast, sustained AdoR activation results in pro-inflammatory effects, e.g., increased cytokine production and inflammatory cell infiltration (Zhou et al. 2009). This conundrum of the Ado’s dual nature has stymied the development of new drugs aimed at the modulation of AdoR signal transductions.
This chapter is a succinct review of recent studies in this field. The large number of relevant publications and the limited scope of this review inevitably impose selective articles’ citation. We thus apologize if our selection of articles does not include article(s) that others may feel should have been cited.
18.2 A1AdoR
A1AdoR , which has the highest affinity to Ado, is expressed at low levels in the lung; relatively higher levels were detected in the newborn mice (Metsola et al. 2014). The activation of A1AdoR reduces adenylate cyclase’s activity via the activation of pertussis toxin-sensitive Gi and Go proteins, stimulates phospholipase C via Gβγ subunits, and activates pertussis toxin-sensitive K+ channels (Pelleg et al. 1996) and KATP channels (Schepp and Reutershan 2008). Activation of the A1AdoR could be pro- or anti-inflammatory depending on the specific pathophysiologic conditions and the levels of extracellular Ado.
Early studies have shown that ischemia-induced constriction of the pulmonary vasculature is mediated by the activation of A1AdoR, which results in thromboxane release (Neely et al. 1991) as well as pulmonary ischemia-reperfusion injury (Neely and Keith 1995). Subsequent in vivo studies using a mouse model have shown that physiologic doses of Ado decrease alveolar fluid clearance (AFC), probably by means of an A1AdoR-dependent mechanism that causes Cl− efflux through CFTR, whereas lower doses increased AFC via the A2aR and/or A3R (Factor et al. 2007). Similarly, alcohol decreases alveolar fluid clearance and impairs survival from acute lung injury. Alcohol induced increases Ado levels in the lung, which may be responsible for reduction in AFC and associated worsening of lung injury (Dada et al. 2012).
A1AdoR expression is increased significantly during LPS-induced injury thereby suggesting that extracellular Ado levels and Ado receptors may play a role in LPS-induced toxicity (Metsola et al. 2014). Indeed, A1AdoR has been implicated in polymorphonuclear cell trafficking and alterations in microvascular permeability in lipopolysaccharide (LPS)-induced lung injury model (Ngamsri et al. 2010). A more recent study in a mouse model has shown that A1AdoR activation improves lung function and decreases inflammation, edema, and neutrophil chemotaxis after ischemia-reperfusion (Fernandez et al. 2013a). This protective effect of A1AdoR’s activation agrees with the finding that pulmonary injury was exacerbated in ADA double-knockout mice, which is also deficient in the expression of A1AdoR (Sun et al. 2005). Similarly, an A1AdoR agonist significantly increased lung compliance and oxygenation and decreased pulmonary artery pressure, decreased neutrophil infiltration by myeloperoxidase activity and edema, and reduced tumor necrosis factor-alpha production in an isolated, ventilated, blood-perfused rabbit lung model of ischemia-reperfusion (Gazoni et al. 2010).
The expression of A1AdoR in the airways of asthmatic subjects is upregulated (Wilson et al. 2009). Ado-induced bronchoconstriction in human subjects is indirect (see above); however, in rodents this action is mediated by A1AdoR and a central vagal reflex (Wilson et al. 2009). Although A1AdoR activation affects “different cell types to produce bronchoconstriction, inflammation, mucous gland hyperplasia, angiogenesis, and fibrosis, all of which are important in the pathophysiology of human asthma” (Wilson et al. 2009), the exact role of Ado and A1AdoR in asthma has not been fully delineated, and accordingly none of the drug candidates targeting A1AdoR signal transduction has been approved by the FDA (see, e.g., Gottlieb et al. 2011).
Adenosine activation of leukocyte A1AdoR plays a significant role in their recruitment to lungs infected with an influenza virus and thereby contributes to influenza pathogenesis (Aeffner et al. 2014).
18.3 A2AAdoR
A2AAdoR is expressed by various inflammatory cell types, and its activation results in broad anti-inflammatory effects (Hasko and Pacher 2008). A2AAdoR signaling attenuates acute lung injury by enhancing alveolar fluid clearance in mice (Eckle et al. 2008). The expression of A2AAdoR was decreased significantly by an allergen challenge in a mouse model of asthma suggesting that A2AAdoR deficiency leads to airway inflammation and airway hyperresponsiveness in this setting (Nadeem et al. 2007).
A2AAdoR activation stimulates the formation of cAMP (Ongini and Fredholm 1996); the latter can significantly upregulate SOCS-3 protein expression (Sands et al. 2006). In vitro and in vivo studies have shown that hypoxia can induce pulmonary artery smooth muscle cell A2AAdoR expression (Qian et al. 2013; Fan et al. 2016). A2AAdoR upregulates SOCS-3 protein, which inhibits pulmonary vascular remodeling in lung tissue of hypoxic pulmonary hypertension rats (Fan et al. 2016). In addition, a recent study utilizing a hypoxia-induced pulmonary hypertension (HPH) mouse model has shown that hypoxia enhanced the expression of A2AAdoR, the activation of which attenuated the release of specific inflammatory cytokines in the lung (Huang et al. 2017). The latter was associated with reduced thickening of pulmonary arterioles and improved hypoxemia (Huang et al. 2017).
An A2AAdoR agonist significantly increased lung compliance and oxygenation and decreased pulmonary artery pressure, decreased neutrophil infiltration by myeloperoxidase activity and edema, and reduced tumor necrosis factor-alpha production in an isolated, ventilated, blood-perfused rabbit lung model of ischemia-reperfusion (Gazoni et al. 2010).
In contrast, it was recently shown that the activation of A2AAdoR modulated the activation of fresh human alveolar inflammatory cells in patients with interstitial lung disease (Alfaro et al. 2017). A2AAdoR plays a role in the regulation of different stages of immune responses, including antigen presentation, T-cell activation, expansion, survival, and memory (Ohta and Sitkovsky 2001). Specifically, Ado suppresses the immune response by activation of A2AAdoR expressed by T-cells. Indeed, a selective antagonist of A2AAdoR reduced the tumor burden in a mouse lung cancer in vivo model and restored immune responsiveness ex vivo (Mediavilla-Varela et al. 2017).
Chronic lung inflammation is associated with fibroblasts proliferation, beginning to proliferate, the formation of new blood vessels, and the increase in extracellular matrix resulting in the development of pulmonary fibrosis (Della Latta et al. 2013). Genetic knockout of A2AAdoR in mice significantly exacerbates, while activation of A2AAdoR attenuates the progression of pulmonary fibrosis induced by bleomycin (a chemotherapeutic agent used to treat several neoplastic diseases and widely used ones for the induction of lung fibrosis (Della Latta et al. 2015; Chen et al. 2017). The beneficial effects of A2AAdoR activation in this setting are mediated at least partially via the stromal cell-derived factor-1 (SDF-1)/C-X-C chemokine receptor type 4 (CXCR4) pathway, the inhibition of which protects the lungs from fibrogenesis in BLM-exposed mice (Chen et al. 2017).
18.4 A2BAdoR
Ado and its cell surface receptors play an important role in the regulation of inflammation following acute lung injury (Karmouty-Quintana et al. 2013). A2BAdoR are expressed by mast cells , bronchial smooth muscle cells, and lung fibroblasts; their activation increases the release of various inflammatory cytokines and promotes differentiation of lung fibroblasts into myofibroblasts, typical of the fibrotic events (Della Latta et al. 2013).
A2BAdoR mediates the relaxing effects of Ado on guinea pig airways (Breschi et al. 2007). Hypoxia is associated with increased soluble CD73 activity, which contributed to hypoxia-induced increase in Ado’s plasma level and Ado-mediated erythrocyte A2BAdoR activation inducing 2,3-BPG production and triggering O2 release that prevents multiple tissue hypoxia, inflammation, and pulmonary vascular leakage (Liu et al. 2016). Extracellular Ado promote dermal fibrosis (Fernandez et al. 2013b), and the activation of A2BAdoR promoted renal fibrosis in both mice infused with angiotensin II and mice subjected to unilateral ureteral obstruction (Dai et al. 2011). A study in a mouse model of bleomycin-induced injury and whole-lung lysate from patients without and with idiopathic pulmonary fibrosis has shown that inhibition of hypoxia-inducible factor 1-α (HIF1α) attenuated pulmonary fibrosis in association with reductions in A2BAdoR expression in alternatively activated macrophages (Philip et al. 2017). These data were interpreted to suggest that hypoxia, through HIF1α, contributes to the development and progression of pulmonary fibrosis through its regulation of A2BAdoR expression on alternatively activated macrophages (AAMs), cell differentiation, and production of profibrotic mediators (Philip et al. 2017).
In contrast, oxygenation may weaken local tissue hypoxia-dependent Ado and A2AAdoR-mediated anti-inflammatory mechanism and thereby further exacerbating lung injury in the setting of acute respiratory distress syndrome (ARDS) (Thiel et al. 2005; Aggarwal et al. 2013). That notwithstanding, exposure to a hyperoxic environment causes lung injury associated with an increase of Ado levels, which protects vascular barrier function in hyperoxic lung injury through the A2BAdoR-dependent preservation of the endothelial cellular adhesion protein occluding (Davies et al. 2014). In addition, the activation of A2BAdoR reduced endotoxin-induced acute lung injury in a murine model (Schingnitz et al. 2010).
A2BAdoR was found to play an important role in mediating lung inflammation after an ischemia-reperfusion challenge by stimulating cytokine production and neutrophil chemotaxis in a mouse model in vivo (Anvari et al. 2010). In contrast, several studies have shown that Ado and A2BAdoR play an important role in the resolution of pulmonary edema and inflammation during ALI (Eckle et al. 2009). For example, in a mouse model of ALI (intratracheal LPS treatment followed by injurious mechanical ventilation), it was found that alveolar epithelial A2bAdoR signaling contributes to lung protection (Hoegl et al. 2015).
Stromal cell-derived factor (SDF)-1 is a chemokine that regulates the release of neutrophils from the bone marrow into the circulation; during inflammation, the concentration of SDF-1 in the bone marrow decreases, and polymorphonuclear neutrophils enter the circulation from where they can migrate to the lungs during ALI (Konrad et al. 2017). SDF-1 is expressed in the human lung during acute lung injury (Petty et al. 2007). The effects of SDF-1 are mediated by two receptors: CXCR4 and CXCR7 (Rath et al. 2014), the inhibition of which results in an anti-inflammatory effect during ALI (Konrad et al. 2017). Using a mouse model of ALI and an in vitro preparation of human epithelium/endothelium, it was found that the anti-inflammatory effects of CXCR4 and CXCR7 antagonism in terms of PMN migration, chemokine release, and microvascular permeability are linked to adenosine A2BAdoR signaling in hematopoietic cells (Konrad et al. 2017).
18.5 A3AdoR
Significant amounts of A3AdoR are expressed in the rat lung (Haeusler et al. 2015). A3AdoR levels have been found to be elevated in subjects with chronic lung disease and in different pulmonary pathological conditions (Della Latta et al. 2013). The selective Ado A3AdoR agonist 2-chloro-N6-(3-iodobenzyl)adenosine-5′-N-methylcarboxamide (2-Cl-IB-MECA) significantly increased lung compliance and oxygenation and decreased pulmonary artery pressure, decreased neutrophil infiltration by myeloperoxidase activity and edema, and reduced tumor necrosis factor-alpha production in an isolated, ventilated, blood-perfused rabbit lung model of ischemia-reperfusion (Gazoni et al. 2010). Similarly, 2-Cl-IB-MECA attenuated lung dysfunction, inflammation, and neutrophil infiltration in a mouse model of lung ischemia-reperfusion (Mulloy et al. 2013).
Previous studies have shown that extracellular purine nucleosides and nucleotides are potent modulators of human lung mast cell (HLMC) degranulation and histamine release (Pelleg and Schulman 2002; Polosa et al. 1995). Low and high concentrations of Ado enhanced and suppressed, respectively, the release of histamine from FcεRI-stimulated HLMC; this action was mediated by A3AdoR (Gomez et al. 2011). The dose-dependent contrasting effects of Ado on histamine release associated with an allergic reaction were confirmed in a subsequent study using cultured HLMC (Nishi et al. 2016). Rudich et al. examined whether the activation of A3AdoR modulates rather than mediates the effects of Ado in human mast cells, presumably at a transcriptional level (Rudich et al. 2015). Using the HMC-1 cell line, Rudich et al. found that A3AdoR activation represses the expression of genes involved in tissue remodeling and that is coupled to downregulation of the receptor expression, both at protein and mRNA levels (Rudich et al. 2015). Furthermore, since in this study, dexamethasone, a commonly prescribed asthma medication, exerted a synergistic signal that increased Cl-IB-MECA induced gene upregulation and facilitated cytokine secretion, it was speculated that this interaction might underlie the resistance to corticosteroids that is experienced in severe refractory asthma (Rudich et al. 2015).
18.6 Conclusions
Although the prevailing school of thought is that the production of Ado and its activation of AdoR play largely beneficial roles in acute pathophysiologic condition and sustained elevated Ado levels can become detrimental by activating pathways that promote tissue injury and fibrosis (Karmouty-Quintana et al. 2013), many studies challenge this concept. Specifically, it seems that even in the acute phase of injury, the resulting effects of elevated extracellular Ado levels critically depend on the targeted cell type and the localized Ado level at the receptors’ sites. This could explain why, until now, neither selective agonists nor antagonists of AdoR subtypes have been approved by the FDA for the treatment of either acute or chronic lung injury and inflammation (Borea et al. 2016).
References
Aeffner F, Woods PS, Davis IC (2014) Activation of A1-adenosine receptors promotes leukocyte recruitment to the lung and attenuates acute lung injury in mice infected with influenza a/Wsn/33 (H1n1). Virus J Virol 88:10214–10227
Aggarwal NR, D’alessio FR, Eto Y et al (2013) Macrophage A2a Adenosinergic receptor modulates oxygen-induced augmentation of murine lung injury. Am J Respir Cell Mol Biol 48:635–646
Alfaro TM, Rodrigues DI, Tome AR et al (2017) Adenosine A2a receptors are up-regulated and control the activation of human alveolar macrophages. Pulm Pharmacol Ther 45:90–94
Anvari F, Sharma AK, Fernandez LG et al (2010) Tissue-derived Proinflammatory effect of adenosine A2b receptor in lung ischemia-reperfusion injury. J Thorac Cardiovasc Surg 140:871–877
Borea PA, Gessi S, Merighi S et al (2016) Adenosine as a multi-Signalling Guardian angel in human diseases: when where and how does it exert its protective effects? Trends Pharmacol Sci 37:419–434
Breschi MC, Blandizzi C, Fogli S et al (2007) In vivo adenosine a(2b) receptor desensitization in Guinea-pig airway smooth muscle: implications for asthma. Eur J Pharmacol 575:149–157
Burnstock G, Brouns I, Adriaensen D et al (2012) Purinergic signaling in the airways. Pharmacol Rev 64:834–868
Caruso M, Holgate ST, Polosa R (2006) Adenosine Signalling in airways. Curr Opin Pharmacol 6:251–256
Caruso M, Varani K, Tringali G et al (2009) Adenosine and adenosine receptors: their contribution to airway inflammation and therapeutic potential in asthma. Curr Med Chem 16:3875–3885
Chen Y, Yu X, He Y et al (2017) Activation of A2ar attenuates Bleomycin-induced pulmonary fibrosis via the Sdf-1/Cxcr4 Axis-related pathway. Am J Transl Res 9:4125–4136
Church MK, Holgate ST (1993) Adenosine-induced bronchoconstriction and its inhibition by Nedocromil sodium. J Allergy Clin Immunol 92:190–194
Cushley MJ, Tattersfield AE, Holgate ST (1983) Inhaled adenosine and Guanosine on airway resistance in normal and asthmatic subjects. Br J Clin Pharmacol 15:161–165
Dada L, Gonzalez AR, Urich D et al (2012) Alcohol worsens acute lung injury by inhibiting alveolar sodium transport through the adenosine A1 receptor. PLoS One 7:E30448
Dai Y, Zhang W, Wen J et al (2011) A2b adenosine receptor-mediated induction of Il-6 promotes Ckd. J Am Soc Nephrol 22:890–901
Davies J, Karmouty-Quintana H, Le TT et al (2014) Adenosine promotes vascular barrier function in Hyperoxic lung injury. Physiol Rep 2:e12155
Della Latta V, Cabiati M, Rocchiccioli S et al (2013) The role of the Adenosinergic system in lung fibrosis. Pharmacol Res 76:182–189
Della Latta V, Cecchettini A, Del Ry S et al (2015) Bleomycin in the setting of lung fibrosis induction: from biological mechanisms to counteractions. Pharmacol Res 97:122–130
Eckle T, Grenz A, Laucher S et al (2008) A2b adenosine receptor signaling attenuates acute lung injury by enhancing alveolar fluid clearance in mice. J Clin Invest 118:3301–3315
Eckle T, Koeppen M, Eltzschig HK (2009) Role of extracellular adenosine in acute lung injury. Physiology (Bethesda) 24:298–306
Factor P, Mutlu GM, Chen et al (2007) Adenosine regulation of alveolar fluid clearance. Proc Natl Acad Sci U S A 104:4083–4088
Fan R, Huang XY, Du KY et al (2016) The regulative Effcets of A2a adenosine receptor on expression of Socs-3 in rats of hypoxic pulmonary hypertension. Zhonghua Jie He He Hu Xi Za Zhi 39:469–473
Fernandez LG, Sharma AK, Lapar DJ et al (2013a) Adenosine A1 receptor activation attenuates lung ischemia-reperfusion injury. J Thorac Cardiovasc Surg 145:1654–1659
Fernandez P, Perez-Aso M, Smith G et al (2013b) Extracellular generation of adenosine by the Ectonucleotidases Cd39 and Cd73 promotes dermal fibrosis. Am J Pathol 183:1740–1746
Franco R, Casado V, Ciruela F et al (1997) Cell surface adenosine Deaminase: much more than an Ectoenzyme. Prog Neurobiol 52:283–294
Fredholm BB, Ap IJ, Jacobson KA et al (2001) International union of pharmacology xxv nomenclature and classification of adenosine receptors. Pharmacol Rev 53:527–552
Gazoni LM, Walters DM, Unger EB et al (2010) Activation of A1-A2a or A3 adenosine receptors attenuates lung ischemia-reperfusion injury. J Thorac Cardiovasc Surg 140:440–446
Gomez G, Zhao W, Schwartz LB (2011) Disparity in Fcepsilonri-induced degranulation of primary human lung and skin mast cells exposed to adenosine. J Clin Immunol 31:479–487
Gonzales JN, Gorshkov B, Varn MN et al (2014) Protective effect of adenosine receptors against lipopolysaccharide-induced acute lung injury. Am J Physiol Lung Cell Mol Physiol 306:L497–L507
Gottlieb SS, Ticho B, Deykin A et al (2011) Effects of Bg9928-an adenosine a(1) receptor antagonist in patients with congestive heart failure. J Clin Pharmacol 51:899–907
Haeusler D, Grassinger L, Fuchshuber F et al (2015) Hide and seek: a comparative autoradiographic in vitro investigation of the adenosine A3 receptor. Eur J Nucl Med Mol Imaging 42:928–939
Hasko G, Pacher P (2008) A2a receptors in inflammation and injury: lessons learned from transgenic animals. J Leukoc Biol 83:447–455
Hoegl S, Brodsky KS, Blackburn MR et al (2015) Alveolar epithelial A2b adenosine receptors in pulmonary protection during acute lung injury. J Immunol 195:1815–1824
Huang X, Wu P, Huang F et al (2017) Baicalin attenuates chronic hypoxia-induced pulmonary hypertension via adenosine A2a receptor-induced Sdf-1/Cxcr4/Pi3k/Akt signaling. J Biomed Sci 24:52
Karmouty-Quintana H, Xia Y, Blackburn MR (2013) Adenosine signaling during acute and chronic disease states. J Mol Med (Berl) 91:173–181
Konrad FM, Meichssner N, Bury A et al (2017) Inhibition of Sdf-1 receptors Cxcr4 and Cxcr7 attenuates acute pulmonary inflammation via the adenosine A2b-receptor on blood cells. Cell Death Dis 8:E2832
Liu H, Zhang Y, Wu et al (2016) Beneficial role of erythrocyte adenosine A2b receptor-mediated amp-activated protein kinase activation in high-altitude hypoxia. Circulation 134:405–421
Mann JS, Cushley MJ, Holgate ST (1985) Adenosine-induced bronchoconstriction in asthma role of parasympathetic stimulation and adrenergic inhibition. Am Rev Respir Dis 132:1–6
Mediavilla-Varela M, Castro J, Chiappori A et al (2017) A novel antagonist of the immune checkpoint protein adenosine A2a receptor restores tumor-infiltrating lymphocyte activity in the context of the tumor microenvironment. Neoplasia 19:530–536
Metsola J, Maksimow M, Ojaniemi M et al (2014) Postnatal development and Lps responsiveness of pulmonary adenosine receptor expression and of adenosine-metabolizing enzymes in mice. Pediatr Res 76:515–521
Mulloy DP, Sharma AK, Fernandez LG et al (2013) Adenosine A3 receptor activation attenuates lung ischemia-reperfusion injury. Ann Thorac Surg 95:1762–1767
Nadeem A, Fan M, Ansari HR et al (2007) Enhanced airway reactivity and inflammation in A2a adenosine receptor-deficient allergic mice. Am J Physiol Lung Cell Mol Physiol 292:L1335–L1344
Neely CF, Haile DM, Cahill BE et al (1991) Adenosine and Atp produce vasoconstriction in the feline pulmonary vascular bed by different mechanisms. J Pharmacol Exp Ther 258:753–761
Neely CF, Keith IM (1995) A1 adenosine receptor antagonists block ischemia-reperfusion injury of the lung. Am J Phys 268:L1036–L1046
Ngamsri KC, Wagner R, Vollmer I et al (2010) Adenosine receptor A1 regulates Polymorphonuclear cell trafficking and microvascular permeability in lipopolysaccharide-induced lung injury. J Immunol 185:4374–4384
Nishi H, Pelleg A, Schulman ES (2016) Ige receptor-mediated histamine release in human lung mast cells: modulation by Purinergic receptor ligands. Ann Clin Lab Sci 46:463–469
Ohta A, Sitkovsky M (2001) Role of G-protein-coupled adenosine receptors in Downregulation of inflammation and protection from tissue damage. Nature 414:916–920
Ongini E, Fredholm BB (1996) Pharmacology of adenosine A2a receptors. Trends Pharmacol Sci 17:364–372
Pelleg A, Hurt CM, Hewlett EL (1996) Atp shortens atrial action potential duration in the dog: role of adenosine the Vagus nerve and G protein. Can J Physiol Pharmacol 74:15–22
Pelleg A, Schulman ES (2002) Adenosine 5′-triphosphate Axis in obstructive airway diseases. Am J Ther 9:454–464
Petty JM, Sueblinvong V, Lenox CC et al (2007) Pulmonary stromal-derived Factor-1 expression and effect on neutrophil recruitment during acute lung injury. J Immunol 178:8148–8157
Philip K, Mills TW, Davies J et al (2017) Hif1a up-regulates the Adora2b receptor on alternatively activated macrophages and contributes to pulmonary fibrosis. FASEB J 31:4745–4758
Polosa R, Blackburn MR (2009) Adenosine receptors as targets for therapeutic intervention in asthma and chronic obstructive pulmonary disease. Trends Pharmacol Sci 30:528–535
Polosa R, Ng WH, Crimi N et al (1995) Release of mast-cell-derived mediators after Endobronchial adenosine challenge in asthma. Am J Respir Crit Care Med 151:624–629
Qian G, Cao J, Chen C et al (2013) Paeoniflorin inhibits pulmonary artery smooth muscle cells proliferation via Upregulating A2b adenosine receptor in rat. PLoS One 8:E69141
Rath D, Chatterjee M, Borst O et al (2014) Expression of stromal cell-derived Factor-1 receptors Cxcr4 and Cxcr7 on circulating platelets of patients with acute coronary syndrome and association with left ventricular functional recovery. Eur Heart J 35:386–394
Rudich N, Dekel O, Sagi-Eisenberg R (2015) Down-regulation of the A3 adenosine receptor in human mast cells Upregulates mediators of angiogenesis and remodeling. Mol Immunol 65:25–33
Sands WA, Woolson HD, Milne GR et al (2006) Exchange protein activated by cyclic amp (Epac)-mediated induction of suppressor of cytokine signaling 3 (Socs-3) in vascular endothelial cells. Mol Cell Biol 26:6333–6346
Schepp CP, Reutershan J (2008) Bench-to-bedside review: adenosine receptors--promising targets in acute lung injury? Crit Care 12:226
Schingnitz U, Hartmann K, Macmanus CF et al (2010) Signaling through the A2b adenosine receptor dampens endotoxin-induced acute lung injury. J Immunol 184:5271–5279
Smail H, Baste JM, Gay A et al (2016) Role of inflammatory cells and adenosine in lung ischemia Reoxygenation injury using a model of lung donation after cardiac death. Exp Lung Res 42:131–141
Sun CX, Young HW, Molina JG et al (2005) A protective role for the A1 adenosine receptor in adenosine-dependent pulmonary injury. J Clin Invest 115:35–43
Thiel M, Chouker A, Ohta A et al (2005) Oxygenation inhibits the physiological tissue-protecting mechanism and thereby exacerbates acute inflammatory lung injury. PLoS Biol 3:E174
Thorn JA, Jarvis SM (1996) Adenosine Transporters. Gen Pharmacol 27:613–620
Wilson CN, Nadeem A, Spina D et al (2009) Adenosine receptors and asthma. Handb Exp Pharmacol 329:362
Zhou Y, Schneider DJ, Blackburn MR (2009) Adenosine signaling and the regulation of chronic lung disease. Pharmacol Ther 123:105–116
Zimmermann H (2000) Extracellular metabolism of Atp and other nucleotides. Naunyn Schmiedeberg's Arch Pharmacol 362:299–309
Zimmermann H, Zebisch M, Strater N (2012) Cellular function and molecular structure of Ecto-Nucleotidases. Purinergic Signal 8:437–502
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Pelleg, A., Polosa, R. (2018). Adenosine Receptors in the Lungs. In: Borea, P., Varani, K., Gessi, S., Merighi, S., Vincenzi, F. (eds) The Adenosine Receptors. The Receptors, vol 34. Humana Press, Cham. https://doi.org/10.1007/978-3-319-90808-3_18
Download citation
DOI: https://doi.org/10.1007/978-3-319-90808-3_18
Published:
Publisher Name: Humana Press, Cham
Print ISBN: 978-3-319-90807-6
Online ISBN: 978-3-319-90808-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)