
Abstract
Very little is known about human occupation of the East African Coast prior to the Swahili period. Only a handful of pre-Swahili archaeological sites from this region have been investigated in any detail. The Holocene archaeology of the coastal region requires research attention if the role of indigenous African populations in later ethnic mosaics is to be understood. Pollen , fungal spores, and charcoal remains from a sediment core derived from Mtwapa Creek are presented with the goal of better understanding mid-to-late Holocene (c. 7500–4000 cal yr BP) occupations of the coast. Evidence for increased local burning, herbivore activity, and overall landscape disturbance possibly derived from cultivation activities characterizes the period between c. 6200–4000 cal yr BP. Intensive exploitation of riverine habitats was practiced in the neighboring hinterland during this period, and it is proposed that similar adaptations may have been at play in coastal drainages at Mtwapa Creek and elsewhere contemporaneously. A variety of investigative methods are proposed that may yield additional information on human activities during this time period.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others
Keywords
Introduction
In contrast to the well known Swahili cultural phenomenon of the past two millennia, the archaeology of the pre-Swahili East African coast has been largely ignored. Indeed, the scope of human activities in this region prior to the Late Holocene is almost completely unstudied. Exceptions to this trend are few and far between (Omi 1982, 1984, 1986, 1991; Helm 2000; Helm et al. 2012; Shipton et al. 2018), yet this evidence suggests that the East African Coast was rife with human activity through, and perhaps previous to, the Holocene . As little archaeological detail dating to the Early and Middle Holocene is yet available for this region, a landscape-scale approach is employed here to render any significant changes to ancient coastal ecologies due to human activities visible. In an effort to clarify the nature of pre-Swahili human environmental impacts, I offer a paleoecological reconstruction of Mtwapa Creek Estuary and the surrounding environs during the Middle Holocene (c. 7500–4000 cal yr BP), using pollen , microscopic charcoal , and fungal spore data from a sediment core spanning this period. This data is then integrated with known archaeological knowledge of Holocene human occupation of the East African coast. First, a brief overview of integrated palynological and fungal research in East Africa is presented, followed by a review of pertinent archaeological data from the East African coast. Next, nested microbotanical data from a 5 m sediment core collected at Mtwapa Creek are discussed and interpreted. I use these interpretations to hypothesize the nature of human occupation of this area during the Middle Holocene , and advocate for the usefulness of interpreting multi-proxy environmental records in light of local archaeological evidence as a potential means of dealing with equifinal data sets. Finally, suggestions for additional topical research are put forth.
Multi-proxy paleoecological data sets have aided in defining environmental and cultural chronologies in numerous regions in East Africa (e.g. Cohen et al. 2005; Gelorini et al. 2011, 2012a, b; Lejju et al. 2005; Palacios-Fest et al. 2005a, b; Rucina et al. 2009; van Geel et al. 2011; Wooller et al. 2003; and many others). Lejju et al. (2005) have reconciled several lines of paleoecological evidence, including pollen , phytoliths , and fungal spores, with archaeological and historical evidence at Munsa, Uganda to better understand site use and occupation. Further South, past ecological change at Lake Tanganyika has been documented in a set of very detailed environmental records derived from numerous proxies by Cohen et al. (2005) in a special issue of the Journal of Paleolimnology (Palacios-Fest et al. 2005b). Data from fungal spores is emphasized more specifically by Gelorini et al. (2011, 2012a, b) and van Geel et al. (2011). Van Geel et al. (2011) analyzed an assemblage of fungal “non-pollen palynomorphs” (NPP’s) from Lake Challa in southeastern Kenya, and found that more than two thirds of typed NPP specimens could be identified, or “linked,” to known taxa of fungi . As study of microscopic African fungal bodies is still in its relative infancy, many specimens which could not be reliably assigned to taxonomic groups were “typed” according to shared morphological affinities. Nevertheless, the authors found that diagnostic spores could often be assigned to known genera or species, yielding (in these cases) a level of taxonomic resolution similar to that possible with pollen grains. Of these diagnostic types, a variety are substrate-or-environment-specific indicators. Sporormiella, for example, has been documented as an obligate coprophile, growing exclusively on dung substrates (Davis1987; Davis and Shafer 2006; Raper and Bush 2009; Parker and Williams 2011; Baker et al. 2013).
Local Environment and Sedimentary Context
The surrounding landscape of Mtwapa Creek today is agrarian to semi-urban in nature. Agricultural activities in the vicinity are largely coconut, mango, and maize farming. Small-scale husbandry of cattle, goats, and sheep is also practiced. The coring location is situated approximately 4 km West of the town center of Mtwapa on a tidal mudflat off the northern shore of the Mtwapa Creek estuary. A band of small agricultural homesteads shields the coring site from the small commercial area surrounding the B8 highway, the major (and only) North-South highway serving the coastal region . Moisture availability in the near-coastal plain is greater than in the more arid hinterland. Rainfall in the region varies between 800–1400 mm per annum (Oosterum 1988).
Recent coastal sediment deposits in this region, dating to the post-Pleistocene, have been described by Thompson (1956), Williams (1962), and later by Oosterum (1988), who synthesized descriptions from these and other earlier studies, and whose designations I employ here. The intertidal mudflats from which core material was extracted belong to the aptly named recent tidal-flat deposits identified by these authors, being derived primarily from fluvial sedimentation processes. Sedimentary material comprising these deposits originates primarily from surrounding land terraces eroded via rainfall and mechanical disturbance, as well as suspended particulate matter of small size transported into mudflats at high tide. These mudflat deposits occur frequently in mangrove environments throughout the East African littoral, and have been characterized as Thionic Fluvisols (Michieka et al. 1978; Boxem et al. 1987). These sediments are comprised of dark grey-to-olive green clays and clays with varying proportions of sand in their matrix. These deposits have been identified as Holocene and Late Pleistocene in age, with only very gentle gradation in sediment characteristics being detectable over this timespan.
Mangrove communities are by far the predominant vegetation along the estuary margins and undeveloped parts of the coast. Eight of the nine recognized species of East African mangroves are native to Kenya, including Rhizophora mucronata (Lam.), Ceriops tagal (C. B. Rob.), Bruguiera gymnorrhiza (L.), Avicennia marina (Forsk.), Sonneratia alba (Sm.), Heritiera littoralis (Aiton.), Xylocarpus granatum (Koenig.), and Lumnitzera racemosa (Willd.). The ninth species, Avicennia officinalis, is today found only in coastal Madagascar. Local coastal communities frequently rely on mangrove forests for building timber, craftworking (e.g. construction of fish traps), and as a fuel source (Taylor et al. 2003). According to Taylor et al. (2003) mangroves are a lynchpin of estuarine ecologies, with leaf litter from mangrove forests responsible for a large portion of organic inputs into these environments. Mangrove pollen is well represented throughout the sediments examined here.
Pre-Swahili Archaeological Research
Available data on Holocene human habitation of the East African coast is very scanty in comparison with what is known of the later Swahili occupations of the region. Linguistic and ecological evidence has been cited in argument that southwest Asian colonizers definitely encountered significant populations of indigenous African peoples occupying the coast (Schoenbrun 1993; Ehret 1998). However, only a few studies containing primary data relating to ‘stone age’ coastal occupants have been published, spanning from present day Mozambique (Morais 1988; Adamowicz 1990) north through Kenya and Tanzania (Soper 1966; Chami and Kessy 1995; Kessy 1997; Chami 1998; Omi 1982, 1984, 1986, 1991; Shipton et al. 2018), and into Somalia (Coltorti and Mussi 1984). A succinct overview of these studies is provided by Helm (2000), but the findings of Omi (1982, 1984, 1986, 1988, 1991) will be the primary sources discussed further here, as lithics pre-dating the Early Iron Age are detailed there. The very recent findings of Shipton and colleagues (2018) are not discussed here.
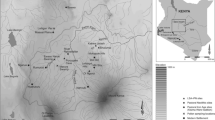
Omi (1982, 1984, 1986, 1991), and contributors to these reports, describe multiple coastal archaeological sites in varying degrees of detail (See Fig. 1), focusing primarily on the geomorphic and stone tool content of each. The main site of focus in these studies however, is the large, multi-component hillside site from where these volumes get their name, Mtongwe. This site, southwest of Mombasa City, across the large creek channel, is dominated by three separate hills: Sassoon Hill, Omar Hill, and the more southerly Ali Abubakar Hill. Excavations were performed at each of these hill site areas of Mtongwe. The authors provide detailed maps of the sedimentary strata encountered in various excavations at Mtongwe, but the only artifactual evidence presented is recovered lithic material.

Location map of the study area.
Considerable variation exists between the lithic assemblages recovered at each hill excavation. Following this, Omi (1984) argues that these assemblages reflect human activity spanning the “… Earlier Stone Age to the Later Stone Age (Omi 1984, p. 22; authors emphasis).” Unfortunately, Omi and colleagues do not employ lithic typologies specific to East African stone tool industries, assigning given finds to “Acheulean,” “Mousterian,” or “Fauresmith” industries. Nevertheless, detailed drawings and measurements, and even photographic plates in later volumes of this series, are provided of recovered lithic implements, allowing independent analysis of these assemblages. The exact identities of lithic assemblages analyzed and recovered during these excavations are beyond the scope of this chapter. It suffices to say that variation in lithic assemblages recovered from Mtongwe and other coastal sites such as Panga ya Saidi (Shipton et al. 2018) suggests that human occupation of the Kenyan coast prior to the advent of the Swahili period persisted for some time.
The few published radiocarbon dates from this project (Omi 1991), derived from materials excavated at Ali Abubakar Hill, support the notion of human coastal occupation from at least the Late Pleistocene into, and likely through, the Holocene and proto-Swahili period, slightly predating the second millennium BP. Four radiocarbon assays were made on charcoal recovered from a bed of reddish orange sand with calcareous particles in the matrix. The resultant dates varied in age considerably, ranging from 17,680 ± 270 BP [21,325 ± 367 cal BP] to 1,090 ± 90 BP [953 ± 186 cal BP]. Calibrated dates reported here were obtained via Calib 7.1, using the SHcal13 calibration curve (Stuiver et al. 2017). Alternate calibrations using the mixed Marine & NH Atmosphere curve in this program (Stuiver et al. 2017), with hypothetical marine carbon percentages of 1–5% resulted in only small departures from calibration with the SHcal13 curve. Furthermore, the youngest dates were derived from materials in deeper sedimentary levels than older dates. Excavation depth measurements recorded in Omi’s 1991 volume are reported in meters above sea level, rather than the more traditional centimeters below surface/datum. Thus, higher measurement values indicate shallower sedimentary contexts from the excavation reported therein. Sediment admixture and/or the nature of recovery techniques may be responsible for this juxtaposition. A large discrete charcoal block with provenience at 41.70 masl (meters above sea level; original measurement value) yielded a date of 9,050 ± 150 BP [10103 ± 214 cal BP] (again, see Omi 1991, p. 33 for clarification of depth measurements). The authors note that the remaining three dating assays were performed on collections of small charcoal fragments collected from throughout each additional sedimentary level. Contamination by modern plant roots and insect bioturbation, they argue, resulted in influx of young carbon into lower stratigraphic levels. Therefore, the date derived from 41.70 masl likely represents the most secure date from this site. Regardless of overall stratigraphic integrity at this site, the dates indicate a long history of occupation, well preceding the last several thousand years. While Omi and colleagues posit an age for the lithic assemblages at Ali Abubakar Hill much older than these radiocarbon dates suggest, this conclusion is based upon now outdated lithic temporal associations. In fact, the lithic assemblages from numerous coastal sites documented by Omi and co-workers closely resemble Eburran lithics from the Central Rift Valley, detailed by Ambrose (1984, 1998) and others (Wilshaw 2012). Hypothesizing a cultural or evolutionary connection between coastal and Rift Valley lithics would obviously be impossible at this point, however, as lithic technologies are known to display high inter-site and inter-regional variability. Despite this, general trends in dimensional reduction over the last 10,000–15,000 years, documented by Ambrose (1984), suggest that coastal lithics from sites such as Ngutatu and Mto Panga may potentially be of Late Pleistocene or Holocene origin. Thus, coastal occupations and activity sites detailed in the works of Omi (1982, 1982, 1984, 1986, 1991) may actually be considerably younger than argued to be in these volumes. Comparison of these finds with lithic data recently available from the site of Panga ya Saidi (Shipton et al. 2018) will undoubtedly aid in contextualizing these materials.
Methods
A five meter sediment core was extracted from the intertidal zone of Mtwapa Creek using a modified piston corer. Sediments were extruded and sampled every 5 cm along the entire length of the core. Approximately 15–20 ml of sediment were excised for each sample. Sampled sediments were sealed in whirl-paks© and returned to the Department of Anthropology at Washington State University for processing and analysis in the Palynology and Paleoenvironments Laboratory (now the Archaeobotany Laboratory) housed there. Nineteen sediment samples were selected for analysis. Sediment processing was performed using the following procedure: quantification of 2 ml of sediment for each sample, addition of two commercially produced Lycopodium tablets to allow calculation of pollen and spore concentrations, carbonate removal with 10% Hydrochloric Acid, screening through 150 μm wire mesh, overnight immersion in 65% Hydrofluoric Acid to remove silicates, 20” treatment with 1% Potassium Hydroxide to dissolve humates, dehydration in 99% Glacial Acetic Acid, acetolysis treatment as outlined by Erdtman (1960), heavy density separation with Sodium Polytungstate at 2.34 specific gravity, curation with glycerin in glass vials. Each chemical treatment step was followed by repeated rinsing in deionized water and centrifugation at >2,000 rpm.
Samples material was mounted on glass microscope slides to undergo microscopy. Counting of pollen , spores, and charcoal was performed under a light microscope at 100–400x magnification, with higher magnifications of up to 1000x being used when necessary. Vertical traverses of each slide were made until at least 400 pollen grains were counted for each sample. This procedure was repeated for counting of fungal spores and of charcoal fragments. Identifications were made using published micrographs and available reginal pollen (Caratini and Guinet 1974; Bonnefille and Riollet 1980) and spore (Jarzen and Elsik 1986; Gelorini et al. 2011; van Geel et al. 2003, 2011) keys, as well as online resources such as the African Pollen Database. Charcoal was categorized into size ranges [small-sized 3–25 µm, medium-sized 26–75 µm, large-sized 76–150 µm], and counted following the procedure outlined by Rucina et al. (2009), with identifications proceeding according to the recommendations of Waddington (1969) and other authors (Patterson et al. 1987; Pitkanen and Huttunen 1999; Clark 1988). Graphical rendering was performed on Tilia 2.0 (Grimm 1991). Pollen , spore types, and charcoal size classes are rendered as percentages of the overall assemblage in each sample.
Results
Radiocarbon Dates
Six bulk carbon dates were obtained via Direct AMS, which indicate that this sediment core captures the majority of the Middle Holocene (See Table 1). All dates were calibrated using CALIB 7.1 (Stuiver et al. 2017) under the MARINE13 calibration curve to account for marine reservoir effects. Results confirmed that the period between c. 7500–4000 cal yr BP is reflected in these sediments, comprising the majority of the Middle Holocene (8200–4200 cal yr BP), and likely verging into the earliest part of the Late Holocene (4200 cal yr BP-forward) as defined by Walker et al. (2012).
After graphical rendering of results, sum of squares analysis suggested the existence of two major analytical zones: a younger zone defined by sediments above 175 cm below ground level, and an older zone comprised of all deeper sediments. The older zone is from here on called Zone 1, and the younger zone, Zone 2. This phenomenon was captured independently in pollen , fungi , and charcoal datasets. Association between dates suggests rapid sediment accumulation during the mid-8th millennium BP and during the mid-to-late 7th millennium BP. During each of these periods, over 100 cm of sediment accumulated over very short timeframes. During the earlier of these two periods, diversity of pollen and fungal spores is relatively high (See Figs. 2 and 3). High rates of sediment accumulation such as witnessed at these times are likely the result of precipitation and/or erosion rates. As will be discussed later, climate models indicate that wetter conditions than present predominated during and through these timeframes. Especially wet periods during this extended humid phase may be responsible for these brief periods of extreme sediment deposition. Erosion due to landscape disturbance is another possibility which may account for high sedimentation, but charcoal frequencies and other palynological indicators of disturbance present conflicting signals with regard to forces affecting landscape alteration, leaving this interpretation dubious, but not able to be ruled out.

Pollen representation at Mtwapa Creek.

Fungal representation at Mtwapa Creek.
Pollen
The pollen assemblage from Mtwapa Creek was found to be of relatively low diversity, being overwhelmingly composed of the pollen of various mangrove types (See Fig. 2). The majority of grains identified capture an antagonistic relationship between Rhizophora mucronata (always dominant) and the less diagnostic grains of both Bruguiera gymnorrhiza and Ceriops tagal, which cannot be reliably distinguished at the relatively low levels of magnification used in this study. Other mangroves, including Sonneratia alba, Avicennia marina, Lumnitzera racemosa, and Xylocarpus granatum are present in lower concentrations. This likely reflects fluctuation in tidal heights and/or salinity conditions within the local mudflat over time, and corresponding movement of groves of high, medium, and low tide adapted mangroves. Research related to the dynamics of mangrove ecology has been recently reviewed by Punwong et al. (2013a, b), and is not rehashed here (see also, Gastaldo 2012).
Sedge and grass pollen also are present in considerable concentrations. In the oldest sediments, Cyperaceae and Poaceae percentages seem to mirror each other. Yet in the youngest sediments, an inverse relationship between these types develops. Closer examination of Cyperaceae species composition in this assemblage may yield pertinent information on the ecologies of present sedge types, shedding light on the nature of the relationship between grasses and sedges in this environment. Overall grass pollen concentrations remain relatively high and constant through the core stratigraphy, with fluctuations and low percentages characteristic of depths between 100–200 cm. Low concentrations of large-sized Poaceae pollen (“large-sized” being defined as >37 µm in maximum diameter) appear in the youngest sediments, along with trace percentages of Heliotropium steudneri and other assorted herbaceous taxa, present in very low amounts. Asteraceae percentages generally parallel those of grass pollen .
Patterns in arboreal pollen representation are variable. Podocarpus and Celtis pollen (presumably not of local origin) are a constant, low-level presence in the oldest sediments, but then vanish and reappear intermittently through the youngest sediments. Low levels of Hyphaene thebaica and Steganotaenia arialiaceae are present in older sediments in very low numbers, while pollen of Mimosaceae presents in trace amounts in the youngest sediments.
Fungi
Fungal diversity was found to be considerably higher than pollen diversity (See Fig. 3). Encountered spores were grouped based on environmental and substrate-mediated conditions which each fungal type has been found to be associated (see Fig. 3; fungal representation), with unidentified and indeterminate types being grouped according to spore morphology. Type UG 1109, as-yet unnamed, has been posited to be associated with xeric conditions by Gelorini et al. (2011). Concentrations of these spores closely follow Poaceae pollen percentages. Wood and vegetation decay commensals were found in moderate diversity through all sediments. Concentrations of individual types vary over time, but remain relatively constant as a percentage of overall spore content throughout core stratigraphy.
Spores commensal with dung and soil isolates were very diverse in all levels. The strongest presence is that of Glomus-type spores, a soil erosion indicator with concentrations which vary through time (especially in the youngest sediments), yet remain a relative constant. Many saphrotrophic and coprophilous fungal types present in low concentrations throughout the core, while others, including Apiosordaria, Ascodesmis, and Podospora appear only in the top 1–2 m of sediment.
Spores which are often found in association with particular plants or types of vegetation were also recovered. Curvularia spp. have been found to grow on grasses, maize, wheats, and rice (Ellis 1971). Various species of Fusarium have also been reported on grasses, but more frequently on stored sorghum and millet (Onyike and Nelson 1992; Penugonda et al. 2011). Overall spore diversity, to include unidentified and non-diagnostic spores, increased in sediments shallower than 200 cm below ground level.
Discussion
The often equifinal nature of anthropogenic and climatic environmental signatures poses obvious problems in the investigation of ancient ecologies. This is particularly true for archaeologists interested in human landscape impacts. As discussed earlier, multiple lines of proxy evidence have been used to approach this issue with great affect in East Africa and elsewhere. Paleoenvironmental records from East Africa suggest that persistent humid conditions progressively gave way to a more xeric climate between the Early Holocene and c. 5700 cal yr BP, after which dry conditions seem to have persisted until well after 4000 cal yr BP. A warming trend seems to have ensued after c. 4500 cal yr BP, although conditions remained dry (Richardson 1972; Nicholson 1996; Bergner et al. 2003).
Zone 1: c. 7500–6300 cal yr BP
If human impacts indeed shaped the Middle Holocene ecology of the Mtwapa Creek catchment, data patterns should differ from what would be predicted by climate models for the region. Of particular importance is the period after c. 6200 cal yr BP, which seems to capture much environmental fluctuation. If ecological changes captured in this core are solely climatologically mediated, proxy data should point toward a relatively humid, yet drying, period between c. 7000–5700 BP, and a dry period between c. 5700–4000 BP, when core chronology terminates.
Climate models predict a relatively humid phase prior to 5700 cal yr BP in this region (van Zinderen Bakker 1966; Thompson et al. 2002; Kiage and Liu 2006). Mangrove pollen ratios remain relatively stable during this timeframe, while grass pollen percentages are relatively high, as are Asteraceae and Podocarpus percentages. While these patterns would not necessarily be expected under conditions of relative humidity, moisture availability may not have been great enough to favor the development of forest cover that could outcompete grassland environments. Based on floristic analysis of numerous grid squares surrounding a mangrove coring site, Punwong et al. (2013a) contend that Poaceae representation in sediment cores from mangrove environments may not accurately reflect the actual presence of these environs in the immediate landscape. However, based on the presence of considerable amounts of grass-dominated landscape near the Mtwapa Creek coring site, there does not seem to be sufficient reason to discount the importance of grasses in this analysis. Podocarpus presence is likely a relic of regional pollen rain, which may have been preferentially introduced during precipitation events. Sedges, many of which favor less saline environments, also experience an extended period of stasis during this early period. Based on these pollen data, there does not seem to be much justification for discounting climatic patterns as a primary mediator of environmental conditions during the first approximately 1000 years captured in sediments from Mtwapa Creek.
Fungal data from the oldest sediments unfortunately do not much aid in further clarifying the source(s) of ecological mediation during this period. Spores of both Glomus sp. and Glomus-type, which are taken as indicators of soil erosion, display elevated concentrations in sediments dating between c. 7500–6200 cal yr BP. The means by which erosion may have been affected, however, are still unclear. Precipitation and/or disturbance potentially related to dessication, mechanical disturbance by human activity or animal traffic, or grazing by herbivores may all result in similar erosional signals. Cercophora sp. and Cercophora-type spores also show slightly elevated concentrations in these sediments. While often found on dung, Cercophora also occurs on decayed wood and/or plant material (Bell 1983). Canalisporium spp. is commensal with decaying and inundated wood, and is found in elevated percentages in the oldest sediments. Wood decay likely would be favored by moist conditions, as predicted by climate data. Gelorini et al. (2011) identified Type UG-1109 as a possible indicator of locally dry conditions based on the similarity of this type to a European spore type which indicated such conditions. Elevated percentages of Type UG-1109 during the likely relatively wet period between 7500–6000 BP cast this relationship into doubt. The existence of human-created dry microhabitats in this vicinity which might have played host to xeric-commensal fungi should not be ruled out, however. Thusfar though, the most likely scenario favors the prevailing climatic regime as the primary shaper of the Mtwapa Creek ecology prior to c. 6200 BP.
Zone 2: c. 6200–4000 BP
In sediments shallower than approximately 175 cm below ground level, relatively rapid fluctuations in pollen and fungal representation occur. Climate models predict the existence of very dry conditions during this period, with a notable warming trend beginning slightly prior to 4500 BP. Punwong et al. (2013a) appear to capture the predicted cool and dry period in the Rufiji Delta of Tanzania between c. 5600 and 4600 cal yr BP as a seaward movement of mangroves during this period, indicating a sea level regression. After this time, sea levels appear to rise again, enabling a coincident mangrove resurgence. If this phenomenon occurred at Mtwapa Creek, it does not appear readily apparent in the pollen data. Indeed, both Rhizophora mucronata and Sonneratia alba present with relatively high values during this period. This would not be expected if sea levels were subsiding. It may be that sea levels dropped in this vicinity, yet that tidal levels did not drop sufficiently to render the immediate mudflat environment unsuitable for Rhizophora mucronata. Tidal levels may have dropped enough, however, to create favorable nearby conditions for Xylocarpus granatum, which prefers much less frequent inundation.
In the terminal sediments of Zone 2, Rhizophora percentages drop, while percentages of Ceriops tagal and Bruguiera gymnorrhiza increase. This may reflect selective mangrove harvesting for building materials and other uses, as has been documented in later Swahili communities (Kusimba 1999). This could indicate sea level subsidence during the period shortly preceding 4000 cal yr BP. Sedge percentages also increase during this latest period. These patterns, as well as the fact that no further significant sediment accumulation apparently took place, may be due to the absence of sufficient suspended sediment in tidal events to affect a depositional environment. Fluvial transport inputs from landward freshwater runoff likely were not powerful enough to reach mid-mudflat contexts, leading to the effective termination of sedimentation at the coring location.
The near disappearance (and subsequent reappearance) of grass pollen during the earliest part of Zone 2 is anomalous. Rhizophora pollen may have simply been too numerous for many grass pollen grains to be captured during microscopy. Cool and dry conditions are not a likely culprit responsible for very low grass pollen values, as sedge percentages are elevated at this point in the Mtwapa Creek chronology. A potential scenario which could account for both low Poaceae and high mangrove percentages would be burning of grasslands. Large-sized (>75μm) charcoal fragment concentrations peak during this time, indicating increased local burning of vegetation (see Fig. 4). This would also explain the elevated sedge percentages seen at this time, as a waterlogged environment would provide a measure of protection against land fires. Heliotropium steudneri and other herbaceous taxa appear in low numbers coincident with the lowest observed grass pollen values. Asteraceae percentages, at all times, seem to parallel grass pollen percentages, albeit in much lower numbers. It may be that select herbaceous taxa were able to colonize (if only temporarily) recently disturbed or burned areas prior to the resurgence of grasslands after fire events.

Integrated charcoal and palynomorph indicator representation at Mtwapa Creek.
Through the majority of Zone 2, paralleling both grass and Asteraceae values, large-sized Poaceae pollen (>37 μ in diameter) appear in low numbers. All large-sized Poaceae in this core fall into a size range between 37–42 μ in diameter. This closely matches the size range of the pollen of domesticated sorghum ( Sorghum bicolor, (L.) Moench), an important traditional East African cultigen (Schmidt and Bothman 2005). While this is reason for pause, numerous uncultivated wild grasses have been found to share overlapping (and larger) size ranges with large-sized Poaceae pollen from Mtwapa Creek (Mercuri et al. 1999; Mercuri and Trevisan Grandi 2001; Mercuri 2008). Though it cannot thus be proven that the sudden appearance of relatively large-sized Poaceae pollen in the Mtwapa Creek record represents cultivation activity, the possibility merits further exploration.
Alongside large-sized Poaceae pollen and later peaks in Asteraceae percentages, a fungal type identified as cf. (conferatur) Fusarium sp. appears. Numerous species within the genus Fusarium have been found strongly associated with stored grains (Onyike and Nelson 1992; Chala et al. 2014). While filamentous grass-commensal fungi are often found in association with grassland environments, Fusarium has been found to predominate in stored-grain contexts. While, again, this does not represent an evidentiary “smoking gun” for grain storage or domesticated grains, cultivation and collection of wild grains by foraging populations may be responsible for this particular fungal signature. Also a possibility is that a particular type(s) of wild grass (as yet unidentified specifically) producing relatively large pollen grains found an amenable habitat niche during the increasingly arid phase captured in Zone 2. Concurrently, fungal types including species of Fusarium may have found the same.
Also of particular note, unique increases in the representation of coprophilous fungi occur within Zone 2. Ascodesmis sp. and Delitschia spp. fungal spores appear only during Zone 2, concurrent with, or slightly later than evidence for large-sized Poaceae, Fusarium, and increased percentages of large-sized charcoal , indicating more localized burning. Ascodesmis spores have been reported on various types of dung, while Delitschia spp. have been identified as distinct indicators of large herbivore activity by Gelorini et al. (2012b). Sporormiella-type spores, while present in very low concentrations in Zone 1, experience peak percentages in Zone 2. Non-obligate coprophiles including Apiosordaria-type and Podospora-type also only appear in Zone 2.
The representation of Gelasinospora-type spores also deserves attention. The presence of Gelasinospora in sediments has been shown to be strongly associated with burning. Indeed, the initial appearance of Gelasinospora-type in Mtwapa Creek sediments occurs immediately after a posited fire event captured in the oldest sediments of Zone 1, evidenced by increased occurrence of charcoal in the size range of 26–75 μ. While a long period of relative stasis characterizes both larger sized charcoal and Gelasinospora-type concentrations, in Zone 2 sediments percentages of both are elevated. Furthermore, concentration increases in both of these (and other) proxies are episodic and irregular, rather than progressively increasing. An already dry environment experiencing progressive warming after c. 4500 cal yr BP, as predicted by climate models (Karlen et al. 1999; Kiage and Liu 2006), might be expected to likewise undergo progressively more (and more frequent) burning events. This is apparently not the case, as large-sized charcoal concentrations level off during the period of the expected heatwave. Overall charcoal data patterns support the idea that episodic burning was characteristic of the period between c. 6200–4000 BP. With the exception of the previously mentioned small spike in larger-sized charcoal fragment concentrations after a putative burning event early in Zone 1 sediments, charcoal in the largest two size classes only appears in any number during the period represented by Zone 2.
Middle Holocene Landscape Use in the Coastal Hinter
Multi-proxy research is an important analytical strategy for investigating Middle Holocene coastal occupations in East Africa, as archaeological information dating to this period is chronically underrepresented, not only in the coastal region , but in hinterland regions as well (Kusimba et al. 2005; Wright 2007; Wright et al. 2007). Kusimba et al. (2005) surveyed and excavated numerous sites from the Tsavo region, with a minority of sites from the Bungule complex containing occupations dating to the mid-and-earliest Holocene . Wright (2005), and later, Wright et al. (2007) provided optically stimulated luminescence (OSL) dates from several sites along the Galana River in Tsavo which placed early floodplain occupations in the region at c. 6000 BP. Wright (2007) and Wright et al. (2007) argue that the Galana River floodplain provided an attractive living environment to hunter-gatherers exploiting mostly local resources for over 5000 years, from roughly 6000–900 BP. This more intensive exploitation of riverine environments would have been a potential means of dealing with increasingly xeric conditions. Wright et al. (2007) also contend that hunter-gatherers living along the Galana River adopted intensive local resource exploitation strategies after c. 6000 BP, relying on locally available game and lithic materials, mainly quartz. A similar lithic material procurement strategy seems to be characteristic of coastal occupations described by Omi. Local slate was utilized primarily, with quartzite being of secondary importance.
Evidence for human activity on the East African Coast during this period also appears at inland sites in the form of marine shell (Leakey 1966; Nelson 1993; Wright 2005), indicating the existence of geographically extensive resource exchange networks better evidenced by lithic source materials (Kusimba 2003). Unfortunately, these occupations are located hundreds of kilometers away from Mtwapa Creek, and thus cannot be considered part of the “local” coastal archaeological record. Lithic evidence described by Omi (1982, 1984, 1986, 1988, 1991), attributed to Early-to-Late Stone Age peoples, provides the only described record of potential Middle Holocene human occupation of actual coastal environments in present-day Kenya.
Of the sites and materials described by Omi, the most proximal to the coring site is the site if Ngutatu, roughly seven kilometers southwest of the mudflat coring location at Mtwapa Creek. According to Omi (1982):
At Ngutatu A, seven crescent-type stone tools, four micro-blades, seven cores, fifty-three flakes and one small chopper were collected. All implements are made of shale, except for one chopper and one flake made of quartzite. At Ngutatu B, four side-scrapers, one end-scraper, one micro-blade, thirty-one cores and seventy-one flakes and chips were collected, all implements being made of shale. (Omi 1982, p. 145).
Ambrose (1984) identifies progressive reduction in microlith sizes as a useful means of inferring the age of lithic assemblages from the Central Rift Valley. While exact numerical metrics for lithics from Ngutatu are not given, microlith dimensions reconstructed from scaled illustrations of Ngutatu microliths indicate mean length and width measurements that fall within the range of Ambrose’s Eburran 4 and 5, corresponding to a time range between c. 8500 BP and c. 2000 BP. A thorough overview of Eburran chronology is provided by Wilshaw (2012). As this estimate uses comparison with a non-local lithic typology, this is obviously a very unsatisfactory means of inferring the age of the Ngutatu occupation. However, considering metric and technological similarities between the Ngutatu lithic assemblage and lithic types from the Holocene of central Kenya (including Eburran assemblages), as well as marine shell remains found in these contexts, it is not outside the realm of possibility that microlith size reduction trends identified by Ambrose (1984) might be in evidence during the Middle Holocene in the coastal region of Kenya as well. Although the existence of a relationship between coastal and Rift Valley lithic technologies remains pure speculation at this point, I argue that it is worthwhile to consider the possibility that similar lithic technologies and manufacturing trends were practiced during the Holocene in these two very different regions.
Conclusion
Even taken together, microbotanical, settlement, and lithic evidence presented here in support of human modification of East African coastal landscapes during the Middle Holocene is hardly irrefutable, and based at times on little more than speculation. Having said this, the list of coincident data patterns with uncertain origins presented here should not be ignored. Fungal and palynological data suggesting that large herbivore activity may have increased between c. 6200–4000 BP co-occur with increased evidence for intermittent localized vegetation burning at Mtwapa Creek. Novel pollen types including large-sized Poaceae, and fungal types found associated with grain storage (e.g. cf. Fusarium sp.) also appear and persist during this period, suggesting that cultivation of plant foods may have occurred at an early date. Macro-botanical evidence will obviously be needed to confirm this. Even if these patterns can potentially be interpreted as merely the result of fluctuations in the local climatic and/or precipitation regimes and floral and faunal adaptations to them, human adaptations in the bordering Tsavo region after c. 6000 BP reflect settlement and resource use strategies designed to exploit virtually the same conditions that environmental models predict existed at Mtwapa Creek during this period. Less extensive and more intensive occupation and use of more circumscribed environments would presumably lead to less ephemeral exploitation of such landscapes, leading to more distinctive environmental signatures of land and resource use. Even if human occupation of the East African Coast was of relatively low intensity prior to the last few millennia comprising the Swahili and proto-Swahili periods, these ancient human populations merit the archaeological attention that has historically been denied them. It is rarely demonstrated directly or indirectly (Morris et al. 2015) that colonizing populations from southwest Asia and elsewhere around the Indian Ocean littoral encountered indigenous African populations upon arrival. These populations are often given little more than lip-service acknowledgement as undoubtedly an “important” part of the multi-ethnic mosaic that came to comprise later East African coastal populations. Cultural traditions documented to have been introduced from afar are often emphasized in the archaeological literature, while it is implicitly assumed that indigenous cultural adaptations must have filled the “vacuum” left unexplained in the creation of the Swahili mosaic. I argue that these indigenous populations deserve our renewed future attention.
Several researchers, in addition to Omi and colleagues, have attempted to address these pre-Swahili contexts in recent years (Crowther 2012; Helm et al. 2012; Shipton et al. 2018). Building on this work, several lines of inquiry may be useful in future investigation of the history of coastal occupation in East Africa. More finely resolved records of sea levels through the Holocene will shed light on the significance of mangrove pollen fluctuations over time. Marine terrace records detailed by Oosterum (1988) and others are effectively geologic in time scale resolution. Century, or at least millennial scale knowledge of ancient sea levels would be more appropriate for making inferences concerning past environments. Some research has already been performed in this area (Woodroffe et al. 2015). Mangrove pollen data will undoubtedly be part of this investigation.
Metric data on pollen of coastal grasses will shed light on the nature of relatively large-sized Poaceae pollen recovered in Zone 2. Ecological information on endemic grasses producing pollen of this size will render a clearer picture of grassland dynamics through the Holocene . Sampling of fungal communities commensal with specific grassland contexts will further enlighten this investigation, allowing for comparison of modern and ancient grassland fungal communities. Additionally, modern fungal samples gathered from coastal livestock grazing plots will elucidate the potential value of coprophilous fungi as indicators of herbivore density. In sum, the constellation of proxy signals presented here is sufficient to suggest that human occupation of the East African coast during the Middle Holocene was sufficient to impact coastal ecologies considerably.
Finally, renewed archaeological investigation of known archaeological sites predating the Swahili period will undoubtedly provide new insight into the nature of Holocene coastal occupations. Focus on other aspects of human behavior, aside from lithic technology, that may be preserved at these sites will be a valuable source of information. Additional survey of coastal riparian and hinterland environments for evidence of pre-Swahili sites should provide a clearer picture of the task ahead.
References
Adamowicz L (1990) Newly discovered Stone Age cultures in Nampula province, Northern Mozambique. In: Sinclair P, Pwiti G (eds) Urban origins in eastern Africa: project proposals and workshop summaries. Paper No. 6. Stockholm, Sweden. The Swedish Central Board of National Antiquities, pp 137–197
Ambrose SH (1984) The introduction of pastoral adaptations to the highlands of East Africa. In: Clark D, Brandt SA (eds) From hunters to farmers: the causes and consequences of food production in Africa. University of California Press, Berkeley
Ambrose SH (1998) Chronology of the Later Stone Age and food production in East Africa. J Arch Sci 25(4):377–39
Ambrose SH, DeNiro MJ (1989) Climate and habitat reconstruction using stable carbon and nitrogen isotope ratios of collagen in prehistoric herbivore teeth from Kenya. Quat Res:407–422
Ambrose SH, Sikes NE (1991) Soil carbon isotope evidence for Holocene habitat change in the Kenya Rift Valley. Science 253:1402–1405
Baker AG, Bhagwat SA, Willis KJ (2013) Do dung fungal spores make a good proxy for past distribution of large herbivores? Quat Sci Rev 62:21–31
Bell A (1983) Dung fungi, an illustrated guide to the coprophilous fungi in New Zealand. Victoria University Press, Wellington
Bergner AGN, Trauth MH, Bookhagen B (2003) Paleoprecipitation estimates for the Lake Naivasha basin (Kenya) during the last 175 k.y. using a lake-balance model. Global Planet Change 36:117–136
Bonnefille R, Riollet G (1980) Pollens des Savanes d’Afrique Orientale. Centre National de la Recherche Sientifique, Paris, France
Boxem HW, de Meester T, Smaling EMA (eds) (1987) Soils of the Kilifi area. Reconnaissance soil survey report No. R11, Kenya Soil Survey, Nairobi and Department of Soil Science and Geology, Agricultural University of Wageningen, Pudoc, Wageningen, The Netherlands
Caratini C, Guinet P (1974) Pollen et Spores d’Afrique Tropicale. Association des Palynologues de Langue Francaise. Ministere de l’Education Nationale, Talence, France
Chala A, Taye W, Ayalew A et al (2014) Multimycotoxin analysis of sorghum (Sorghum bicolor L. Moench) and finger millet (Eleusine coracana L. Garten) from Ethiopia. Food Control 45:29–35
Chami FA (1998) A review of Swahili archaeology. Afr Archaeol Rev 15:199–218
Chami FA, Kessy E (1995) Archaeological work at Kisiju, Tanzania. Nyame Akuma 43:38–45
Clark JS (1988) Particle motion and the theory of stratigraphic charcoal analysis: source area, transport, deposition and sampling. Quat Research 30:67–80
Cohen AS, Palacios-Fest MR, Msaky ES (2005) Paleolimnological investigations of anthropogenic environmental change in Lake Tanganyika: IX. summary of paleorecords of environmental change and catchment deforestation at Lake Tanganyika and impacts on the Lake Tanganyika ecosystem. J Paleolimnol 34(1):125–145
Coltorti M, Mussi M (1984) Late stone age hunter-gatherers of the Juba Valley. Nyame Akuma 28:32–33
Crowther A (2012) Agriculture, subsistence and trade in coastal eastern Africa: archaeobotanical evidence from Kenya, Zanzibar and Tanzania. Paper presented at the 21st Biennial conference of the Society of Africanist Archaeologists, University of Toronto, Ontario, Canada: 20–23 June 2012
Davis OK (1987) Spores of the dung fungus Sporormiella: increased abundance in historic sediments and before Pleistocene megafaunal extinction. Quat Research 28:290–294
Davis OK, Shafer DS (2006) Sporormiella fungal spores, a palynological means of detecting herbivore density. Palaeogeogr Palaeocl 237:40–50
Ehret C (1998) An African classical age. University of Virginia Press, Charlottesville
Ellis MB (1971) Demaiaceous Hyphomycetes. Kew Publishers, Surrey, Commonwealth Mycological Institute
Erdtman G (1960) The Acetolysis method: A revised description. Sven Bot Tidskr 54:561–564
Gastaldo RA (2012) Taphonomic controls on the distribution of palynomorphs in tidally influenced coastal deltaic settings. Palaios 27:798–810
Gelorini V, Verbeken A, van Geel BJ et al (2011) Modern non-pollen palynomorphs from East African lake sediments. Rev Palaeobot Palyno 164:143–173
Gelorini V, Ssemmanda I, Verschuren D (2012a) Validation of non-pollen palynomorphs as paleoenvironmental indicators in tropical Africa: contrasting~200-year paleolimnological records of climate change and human impact. Rev Palaeobot Palyno 186:90–101
Gelorini V, Verbeken A, Lens L et al (2012b) Effects of land use on the fungal spore richness in small crater-lake basins of western Uganda. Fungal Divers 55:125–142
Grimm E (1991) Tilia 1.7. Illinois State Museum, Research and Collection Center, Springfield
Helm R (2000) Conflicting histories: the archaeology of the iron-working, farming communities in the central and southern coast region of Kenya. Dissertation, University of Bristol
Helm R, Crowther A, Shipton C et al (2012) Exploring agriculture, interaction and trade on the eastern African littoral: preliminary results from Kenya. Azania 47:39–63
Jarzen DM, Elsik WC (1986) Fungal palynomorphs recovered from recent river deposits, Luangwa Valley, Zambia. Palynology 10:35–60
Karlen W, Fastook JL, Holmgren K et al (1999) Glacier fluctuations on Mount Kenya since~6000 cal. years BP: implications for Holocene climatic change in Africa. Ambio 28:409–418
Kessy ET (1997) Archaeological sites survey from Kisiju to Dar-es-Salaam. Nyame Akuma 48:57–69
Kiage LM, Liu K (2006) Late Quaternary paleoenvironmental changes in East Africa: a review of multiproxy evidence from palynology, lake sediments, and associated records. Progr Phys Geog 30:633–658
Kusimba CM (1999) The rise and fall of Swahili States. Altamira Press, Walnut Creek
Kusimba SB (2003) African foragers: environment, technology, interactions. Altamira Press, Walnut Creek
Kusimba CM, Kusimba SB, Wright DK (2005) The development and collapse of precolonial ethnic mosaics in Tsavo, Kenya. J Afr Arch 3:345–365
Leakey MD (1966) Excavation of burial mounds in Ngorongoro Crater. Tanzania Notes and Records 66:23–135
Lejju BJ, Taylor D, Robertshaw P (2005) Late-Holocene environmental variability at Munsa archaeological site, Uganda: a multicore, multiproxy approach. Holocene 15(7):1044–1061
Mercuri AM, Trevisan Grandi G, Accorsi CA (1999) Plant exploitation during the early and middle Holocene in the Tadrart Acacus (Central Sahara, Libya): pollen evidence of changes. In: Guarino A (ed) Proceedings of the second science and technology for the safeguard of cultural heritage in the Mediterranean Basin. Elsevier, Paris, pp 1081–1085
Mercuri AM, Trevisan Grandi G (2001) Palynological analyses of the Late Pleistocene, Early Holocene and Middle Holocene layers. In: Garcea EAA (ed) Uan Tabu in the settlement history of the Libyan Sahara. Arid Zone Archaeology, Monographs 2. All’Insegna del Giglio, Firenze, pp 161–188
Mercuri AM (2008) Plant exploitation and ethnopalynological evidence from the Wadi Teshuinat area (Tadrart Acacus, Libyan Sahara). J Archaeol Sci 35:1619–1642
Michieka DO, van der Pouw BJA, Vleeshouwer JJ (1978) Soils of the Kwale-Mombasa-Lungalunga area. Reconnaissance Soil Survey Report No. R3, vol I: Main Report, Draft Edicion, Kenya Soil Survey, Nairobi
Morais J (1988) The early farming communities of southern Mozambique. Maputo, Eduardo Mondulane University and Stockholm, The Swedish Central Board of National Antiquities, Studies in African Archaeology, 3
Morris A, Monge J, Kusimba C et al (2015) Decoding the Swahili: ancient DNA studies on the Kenyan Coast. Paper presented at the 80th meeting of the society for American archaeology, 15–19 April, San Francisco, CA
Msaky ES, Livingstone D, Davis OK (2005) Paleolimnological investigations of anthropogenic environmental change in Lake Tanganyika: V. Palynological Evidence for Deforestation and Increased Erosion. J Paleolimnol 34(1):73–83
Nelson CM (1993) Evidence for early trade between the coast and interior of East Africa. Paper presented at WAC Mombassa intercongress conference volume
Nicholson SE (1996) A review of climate dynamics and climate variability in Eastern Africa. In: Johnson AI (ed) Limnology, climatology and paleoclimatology of the East African Lakes. CRC Press, Boca Raton, pp 25–56
Nirenberg HL, O’Donnell K (1998) New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 90:434–452
Omi G (1982) Mtongwe 1980: an interim report of the East and Northeast African prehistory research project 1980. Nagoya University, Nagoya
Omi G (1984) Mtongwe 1982: an interim report of the East and Northeast African prehistory research project 1982. Nagoya University, Nagoya
Omi G (1986) Mtongwe 1984: an interim report of the East and Northeast African prehistory research project 1984. Nagoya University, Nagoya
Omi G (1988) Mtongwe and Mgonga 1986: an interim report of the East and Northeast African prehistory research project 1986. Nagoya University, Nagoya
Omi G (1991) An interim report of the East and Northeast African prehistory research project, 1988/1989. Nagoya University, Nagoya
Onyike NBN, Nelson PE (1992) Fusarium species associated with Sorghum grain from Nigeria, Lesotho and Zimbabwe. Mycologia 90:452–458
Oosterum AP (1988) The geomorphology of Southeast Kenya. Dissertation. Agricultural University, Wageningen
Palacios-Fest MR, Cohen AS, Lezzar K (2005a) Paleolimnological investigations of anthropogenic environmental change in Lake Tanganyika: III. Physical Statigraphy and Charcoal Analysis. J Paleolimnol 34(1):31–49
Palacios-Fest MR, Alin SR, Cohen AS (2005b) Paleolimnological investigations of anthropogenic environmental change in Lake Tanganyika: IV. Lacustrine Paleoecology. J Paleolimnol 34(1):51–71
Parker NE, Williams JW (2011) Influences of climate, cattle density, and lake morphology on Sporormiella abundances in modern lake sediments in the US Great Plains. Holocene 22:475–483
Patterson WA III, Edwards KJ, MacGuire DJ (1987) Microscopic charcoal as a fossil indicator of fire. Quat Sci Rev 6:3–23
Penugonda S, Girisham S, Reddy SM (2011) Elaboration of mycotoxins by seed-borne fungi of finger millet (Eleusine coracana L.). Int J Biotechnol 2:198–200
Pitkanen A, Huttunen P (1999) A 1300 year forest-fire history at a site in eastern Finland based on charcoal and pollen records in laminated lake sediment. Holocene 9:311–320
Punwong P, Marchant R, Selby K (2013a) Holocene mangrove dynamics and environmental change in the Rufiji Delta, Tanzania. Veg Hist Archaeobot 22:381–396
Punwong P, Marchant R, Selby K (2013b) Holocene mangrove dynamics in Makoba Bay, Zanzibar. Palaeogeogr Palaeocl 379–380:54–67
Raper D, Bush M (2009) A test of Sporormiella representation as a predictor of megaherbivore presence and abundance. Quat Res 71:490–496
Richardson JL (1972) Paleolimnological records from Rift lakes in central Kenya. Palaeoecol Afr 6:131–136
Rucina SM, Muiruri VM, Kinyanjui RN et al (2009) Late Quaternary vegetation and fire dynamics on Mount Kenya. Palaeogeogr Palaeocl 283:1–14
Schmidt MR, Bothman G (2005) Indications of bee pollination in sorghum and its implications in transgenic biosafety. Int Sorghum Millets Newsl 46:72–75
Schoenbrun DL (1993) We are what we eat: ancient agriculture between the Great Lakes. J Afr Hist 34:1–31
Shipton C, Roberts P, Archer W et al (2018) 78,000-year-old record of Middle and Later stone age innovation in an East African tropical forest. Nat Commun 9(1832):1–8
Soper RC (1966) Archaeological evidence from the coastal hinterland of Kenya and north-east Tanzania. Unpublished paper, Fort Jesus, Mombasa
Stuiver M, Reimer PJ, Reimer RW (2017) CALIB 7.1 [WWW program]. http://calib.org. Accessed 23 Jan 2017
Taylor M, Ravilious C, Green EP (2003) Mangroves of East Africa. UNEP WCMC, Banson, Cambridge
Thompson AO (1956) Geology of the Malindi Area. Report No. 36, Geological Survey of Kenya, Government printer, Nairobi
Thompson LG, Mosley-Thompson E, Davis ME et al (2002) Kilimanjaro ice core records: evidence of Holocene climate change in tropical Africa. Science 298:589–593
van Geel BJ, Buurman O, Brinkkemper J (2003) Environmental Reconstruction of a Romap Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J Arch Sci 30:873–883
van Geel B, Gelorini V, Lyaruu A et al (2011) Diversity and ecology of tropical African fungal spores from a 25,000-year palaeoenvironmental record in southeastern Kenya. Rev Palaeobot Palyno 164:174–190
van Zinderen Bakker EM (ed) (1966) Palaeoecology of Africa and of the surrounding islands and Antarctica. Balkema, Rotterdam
Waddington JCB (1969) A stratigraphic record of the pollen influx to a lake in the Big Woods of Minnesota. Geol S America S 123:263–283
Walker MJC, Berkelhammer M, Bjorck S et al (2012) Formal subdivision of the Holocene series/epoch: a discussion paper by a working group of INTIMATE (Integration of ice-core, marine and terrestrial records) and the Subcommission on Quaternary Stratigraphy (International Commission on Stratigraphy). J Quat Sci 27:649–659
Williams LAJ (1962) Geology of the Hadu-Fundi Isa area North of Malindi. Report No. 52, Geological Survey of Kenya, Government printer, Nairobi
Wilshaw A (2012) An investigation into the LSA of the Nakuru-Naivasha Basin and surrounding, Central Rift Valley, Kenya: technological classifications and population considerations. Dissertation, St. John’s College, Cambridge
Woodroffe SA, Long AJ, Punwong PA et al (2015) Radiocarbon dating of mangrove sediments to constrain Holocene relative sea-level change on Zanzibar in the southwest Indian Ocean. Holocene 25:820–831
Wooller MJ, Swain DL, Ficken KJ et al (2003) Late Quaternary vegetation changes around Lake Rutundu, Mount Kenya, East Africa: evidence from grass cuticles, pollen and stable carbon isotopes. J of Quat Sci 18(1):3–15
Wright DK (2005) New perspectives on early regional interaction networks of East African trade: a view from Tsavo National Park. Kenya. Afr Arch Rev 22(3):111–140
Wright DK (2007) Tethered mobility and riparian resource exploitation among Neolithic hunters and herders in the Galana River basin, Kenyan coastal lowlands. Environ Arch 12:25–47
Wright DK, Forman SL, Kusimba CM et al (2007) Stratigraphic and geochronological context of human habitation along the Galana River. Kenya. Geoarchaeol 22(7):709–728
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Szymanski, R.M. (2018). Middle Holocene Environmental Change at Mtwapa Creek, Kenya: Distinguishing Human Activity from Regional Ecological Processes. In: Mercuri, A., D'Andrea, A., Fornaciari, R., Höhn, A. (eds) Plants and People in the African Past. Springer, Cham. https://doi.org/10.1007/978-3-319-89839-1_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-89839-1_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-89838-4
Online ISBN: 978-3-319-89839-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)