
Abstract
Acute energy crisis and vulnerable climatic conditions are the major factors behind adoption of efficient and ecofriendly fuel source such as bioalkanes/bioalkenes which are produced using biomass as the feedstock while alkanes/alkenes on the contrary are produced through chemical means. In this context, lignocellulosic biomass presents a ray of hope since its usage will make the process ecofriendly, renewable, and sustainable. Against this backdrop, this chapter presents a detailed overview of the possible conversion technologies viz., chemical and microbial to obtain bioalkanes/bioalkenes and also unravels the possible routes through the intervention of genetic engineering approaches in order to make the process more efficient. It also discusses in details the types of hydrocarbons produced by the microorganisms and their physiological roles so that the biosynthetic pathways can be tapped to the maximum of their potential.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others



Keywords
1 Introduction
The advancement of new technologies for the production of energy and chemicals from renewable resources has encouraged extensive research on biomass valorization (Huber et al. 2006; Corma et al. 2007). Biomass research has now focused more towards the production of innovative candidate fuels over ethanol, diesel, and methane. Target-oriented upgradation of biomass to high-valued fuels such as alkanes and alkenes appear to be an attractive route. These fuels are convenient to the present transportation infrastructures such as pipeline systems for liquid fuels and gas stations for gaseous fuels. Biomass such as lignocellulosics consists of a complex heterogeneous mixture of carbohydrate polymers namely cellulose, hemicellulose, and lignin. Biotransformation of these energy rich molecules (cellulose, hemicellulose, and lignin) into liquid hydrocarbon fuels such as bioalkanes and bioalkenes appears to be an innovative approach for advanced biofuels production (Xia et al. 2016). Thus, utilization of lignocellulosic waste such as agricultural and forest residues for fuel production helps to combat the food versus fuel issue while at the same time attaining international energy security along with reduction of greenhouse gas emissions.
Till date the worldwide supply of liquid fuels for transportation heavily depends on petroleum (more than 95%). The production of petroleum and its derivatives such as short/medium chain hydrocarbons has become more expensive and results into political and military conflicts. Its production process is not only costly but also adds large amount of CO2 in the atmosphere ultimately contributing to global warming. Hong et al. (2013) studied the environmental impact of bio-jet fuels (bio-paraffins 1 (Bio-P1) and bio-jet paraffins 2 (Bio-JP2)) and observed that CO2 lifecycle can be reduced by 76 and 81% respectively for Bio-P1 and Bio-JP2 respectively with respect to fossil jet fuel. Similar kind of study has also been carried out by Jiménez-Díaz et al. (2016) who reported that emissions due to the utilization of bio-jet fuels lead to lower amounts of greenhouse gas emissions such as SO x , NO x , and particulate matter in comparison to conventional jet fuel. Therefore, significant development regarding the raw material selection and processing technologies are indeed necessary, and high-valued fuels produced preferably from the renewable raw biomass will be needed to complement or substitute the present petroleum- and diesel-based fuels. Thus the present chapter discusses the conversion strategies of biomass to bioalkanes/bioalkenes in details with special emphasis on metabolic engineering approaches along with the bottlenecks which hinder their commercial scale production as well as the possible solutions to the impending hurdles.
2 Bioalkanes and Bioalkenes as a Futuristic Source of Fuel
Bioalkanes and bioalkenes are renewable chemicals which have diverse chemical structures and likewise can be used for a number of applications such as fuels, starting compounds for the synthesis of aromatic molecules and as lubricants (Deneyer et al. 2015). The term “bioalkanes and bioalkenes” has a distinct difference with “alkanes and alkenes” in terms of the substrate used for production where biomass is used for production of bioalkanes/bioalkenes while chemical precursors are generally used for producing alkanes/alkenes. Bioalkanes having applications as transportation fuels majorly comprises of hydrocarbons having carbon numbers ranging from C6 to C15 (Xin et al. 2014) and it encompasses fuels like gasoline, diesel, and jet fuel. Bioalkanes are considered to be superior fuels since its fuel properties are better than its counterparts viz., ethanol and butanol. Energy density of alkanes lies in the range of 42–47 MJ/kg while for others it lies in the range of 27 MJ/kg (ethanol)—36 MJ/kg (iso-butanol) (Wang et al. 2016). A comparison of the physical, chemical, and application aspects of bioalkanes over ethanol and butanol is shown in Table 1 (Wang et al. 2016).
Bioalkanes can mainly be produced using renewable carbohydrate sources such as lignocellulosic biomass and also by lipids. Lignocellulosic biomass majorly comprises of lignin, cellulose, and hemicellulose, all of which can be converted to alkanes through different approaches. Lignin is a phenolic polymer linked through ether and C–C interlinkages and thus decoupling reactions of these molecules will lead to phenolic monomers (C6–C11) and dimmers (C12–C22). These phenolic molecules can be converted to cyclic alkanes via deoxygenation and hydrogenation. On the contrary, cellulose and hemicellulose, carbohydrate polymers can be converted to alkanes via decoupling of the polymer to yield individual monomers followed by dehydrogenation to form alkanes. Lipid-rich biomass presents alternative source for alkane production since it is mainly composed of triglycerides from which if the glycerol backbone is removed by hydrolysis, β-elimination or hydrogenolysis then a successive hydrogenation and oxygen-removal steps such as hydrodeoxygenation, decarbonylation, and decarboxylation will convert the fatty acids to alkanes (Deneyer et al. 2015; Zhang et al. 2014). Apart from chemical techniques, biological routes viz., microbial production and application of genetic engineering techniques have also been reported for production of bioalkanes/bioalkenes (Jankowski and ZoBell 1944; Schirmer et al. 2010; Stone and ZoBell 1952; Wang et al. 2013; Kageyama et al. 2015; Gehret-Mccarthy et al. 2012).
Biomass-based sustainable fuel production has been gaining increasing attention due to global climatic condition and future energy security. Hydrocarbons, mainly bioalkanes and bioalkenes, are of special interest because of their potential to be utilized as an advanced biofuel (Lennen et al. 2010) and till date only few works have been focused on the bio-based production of bioalkanes or bioalkenes (Schirmer et al. 2010). Microbial production (300 mg/L) of bioalkanes and bioalkenes was reported by using an engineered strain of E. coli which contained an alkane biosynthesis operon obtained from cyanobacterial genes encoding for aldehyde decarbonylase and acyl-ACP reductase (Schirmer et al. 2010). Slightly different approach was adopted by Harger et al. (2013) who reported that by expressing FabH2 gene of Bacillus subtilis in E. coli, 10 folds increase in the production of alkanes was observed. It was also reported that in absence of FabH2 gene, the bioalkanes produced mainly consisted of n-alkanes (chain length 13, 15, and 17) while its incorporation into E. coli lead to the production of C14 and C16 n-alkanes due to the broad specificity of FabH2 for fatty acid initiation in comparison to the native FabH gene of E. coli.
Bioalkanes are prevalent in a variety of microorganisms such as in cyanobacteria (Winters et al. 1969; McInnes et al. 1980; Dembitsky and Srebnik 2002) which are phylogenetically uniform, 50 of which have been sequenced completely and the data is available in the scientific domain. Among these, 10 cyanobacterial strains having sequenced genome have the potential to produce bioalkanes (Table 2) (Schirmer et al. 2010). These findings further broaden the research on the production of alkanes.
Advancement in metabolic engineering, systems and synthetic biology have paved a way to engineer the microbes to produce high-valued fuels such as bioalkanes and bioalkenes (Keasling 2010; Nielsen and Keasling 2011). Many factors should be taken care of before developing an organism to produce bioalkanes and bioalkenes such as engine type (compression or spark ignition), quality of combustion or ignition delay, cloud point, toxicity, stability, viscosity, lubricity, energy content, water miscibility, volatility and odor (Lee et al. 2008; Peralta-Yahya and Keasling 2010). The main hurdle in exploiting native hosts for the conversion of feedstocks into advanced high-valued biofuels such as bioalkanes and bioalkenes is to gain control over the endogenous regulation of pathways leading to biofuels production with high yields (Peralta-Yahya and Keasling 2010).
Recently, advances in the field of utilizing different non-edible feedstocks and innovative engineering of metabolic pathways for advanced biofuels production have been witnessed. The viability of these engineered metabolic pathways leading to advanced biofuels production has been revealed by the efficient conversion of CO2, syngas, switchgrass and algal hydrolysate into free fatty acids, higher alcohols, and biofuels derived from isoprenoid compounds (Peralta-Yahya and Keasling 2010). In the current energy crisis situation, scientific community is eagerly seeking for feedstocks that can produce advanced biofuels cost-effectively with high yield. Genetic engineering of the biofuel producing pathways along with the choice of an appropriate substrate are the major challenges confronted by today’s researchers for an optimal production of bioalkanes/bioalkenes.
3 Conventional Routes for the Production of Bioalkanes and Alkenes
In order to accommodate a future transition to renewable carbon sources, development of new process technologies is crucial. Potential biomass feedstocks viz., glycerides, lignocellulosics, platform compounds from biomass such as sugar alcohols, hydoxymethyl furfural, furfurals, levulinic acid, and monofunctional hydrocarbons (Deneyer et al. 2015; Zhang et al. 2014) can satisfy this transition. Bioalkane/bioalkene production is one such futuristic fuel which can be produced using these biomasses. Among these biomasses, non-edible/grazable lignocellulosics have the utmost potential in terms of their rich biochemical composition (15–30% lignin, 35–50% cellulose, and 25–30% hemicellulose) and the capability of eradicating the food, fodder versus fuel controversy. All these three biomolecules comprising the biomass have the potential to be converted to bioalkanes/bioalkenes. Lignin is a biopolymer of phenylpropanoid subunits and has high energy content as compared to holocelluloses. It has phenolic molecules such as p-coumaryl, coniferyl, and sinapyl alcohol along with their derivatives. Thus in order to transform lignin and its derivatives to bioalkane, deoxygenation of these molecules has to be conducted. Deoxygenation of lignin is generally carried out by using sulfide (cobalt or nickel-doped molybdenum sulfides) and nonsulfide catalysts (Pt/C, Pd/C, Ni, zeolite-supported nanocatalysts and ionic liquid-stabilized nanocatalysts). Sulfide catalysts are mostly used but they have an innate disadvantage of contaminating the product. Hydrogenolysis of biomass produce monomers (50%) and dimmers (15%) which can be further converted via deoxygenation and hydrogenation to short (C6–C9) and mid-range (C12–C18) cyclic alkane products (Zhang et al. 2014). An alternate one step process for producing mid-range alkanes is the coupling of phenolic monomers followed by conversion to bicycloalkanes (C12–C18). In addition to lignin, cellulose and hemicellulose, carbohydrate polymers also have the potential to produce alkanes/alkenes. Cellulose is a homopolymer of D-glucose and hemicellulose is a heteropolymer of C5 and C6 sugars. Hemicellulose is amorphous and has branched structure which makes it more accessible than cellulose. In order to convert these polysaccharides to alkanes, decoupling step is required. Among these two polymers, hemicellulose is more potent owing to is amorphous nature and thus more accessibility. For decoupling of these polymers, acidic pH and high temperature is maintained to convert them to their respective sugar monomers followed by deoxgenation to produce alkanes. Short and linear chain alkanes can be produced by combining the decoupling step along with the dehydration/hydrogenation step while multistep process is needed for branched or longer bioalkanes. During the conversion, initial step is the conversion of sugars into platform molecules such as γ-valerolactone, angelica lactone or hydroxymethyl furfural. Second step is coupling through aldol condensation, radical reaction, oligomerization or alkylation for long chain formation. Final step is the removal of functional group to obtain alkanes with various lengths (C6–C31), with or without cyclic and/or branched structures (Zhang et al. 2015). A schematic representation of the chemical processing of lignocellulosic biomass for alkane production is shown in Fig. 1.

Strategies for chemical processing of lignocellulosic biomass to alkanes. Cracking reagent and conditions: Catalyst- HZSM-5, 500 °C (Zhang et al. 2015); Coupling reagent and conditions: Catalyst- Pd, 473–523 K (Zhao et al. 2012); Decoupling reagent and conditions: Catalyst-Ru/C, Tungstosilicic acid (TSA) hydrate, water, 483 K (de Beeck et al. 2015)
4 Biological Route: A Novel Approach for the Production of Bioalkanes and Bioalkenes
The genesis of bioalkane/bioalkene production from biological sources emanated from a simple question about the role of microorganisms in producing fossil fuels and organic substances comprising the sediments. From this point onwards, a plethora of knowledge started arriving on the research arena concerning intracellular and extracellular hydrocarbon content of the microorganisms and a range of studies were conducted on deciphering the biosynthesis of these hydrocarbons within the microbes (Bird and Lynch 1974; Albro 1976; Weete 1976). These studies were highly complemented by the advent of gas/liquid chromatographic techniques which enabled the detection of hydrocarbons with high amount of precision.
4.1 Microbial Synthesis of Bioalkanes and Bioalkenes
Hydrocarbon production by microorganisms can be broadly categorized into internal and external depending upon their site of production. Internal hydrocarbons are widely found in a variety of microorganisms viz., bacteria, fungi and yeast where they serve the physiological role of cell membrane permeability but the yield of accumulation is low (0.005–2.69% in bacteria and 0.01–1.6% in yeast). The first report on the microbial hydrocarbon production dates back to 1944 when Jankowski and ZoBell (1994) reported the production of C10–C25 aliphatic hydrocarbons by sulfate reducing bacteria maintained in a growth media containing seawater along with fatty acids. Stone and ZoBell (1952) reported the production of hydrocarbons (0.03 and 0.25% of the biomass) by Serratia marinorubrum and Vibrio ponticus respectively when cultured in seawater peptone media. The type of hydrocarbons produced is the signature of the microorganisms as photosynthetic cyanobacteria has a unique ability to produce branched C18 hydrocarbons (7-methylheptadecane and 8-methylheptadecane) in a ratio of 1:1 (Ladygina et al. 2006). Studies have also shown that alkanes (chain length 17–20 carbon atoms) serve the purpose of biomarkers and indicate the contribution of cyanobacteria towards the formation of organic matter in the sediments (Brocks et al. 2003). The distinguishing quality of anaerobic phototrophic bacteria (Rhodopseudomonas sp., Chlorobium sp. etc.,) is the fact that they produce isoprenoids viz., pristine and phytane. Clostridium sp. produces hydrocarbons having a chain length of C11–C35 among which the dominant ones are middle chain (C18–C27) or long chain (C25–C35) n-alkanes while Desulfovibrio desulfuricans produces alkanes of chain length C11–C35 (Ladygina et al. 2006). Facultatively anaerobic bacteria of the genus Vibrio stores peculiar amount of n-heptadecane as 80% of the biomass is constituted by the hydrocarbons (Ladygina et al. 2006). Aerobic bacteria mainly represented by Micrococcus and Sarcina produces hydrocarbons (C23–C30) which are mainly monounsaturated and have a double bond at the centre of the chain while the chain endings are frequently marked with iso-methyl or anteiso-methyl branches (Ladygina et al. 2006).
Studies were also conducted on yeast accumulating hydrocarbons and it was observed that they accumulated nearly 0.01–1.6% of the cell dry weight under aerobic conditions and if grown anaerobically, Saccharomyces sp. has shown to accumulate up to 10.2% hydrocarbon by its dry cell weight (Ladygina et al. 2006). The hydrocarbons produced have a chain length of C10–C34 and includes straight chain saturated, unsaturated, and branched hydrocarbons. The effect of carbon source on the type of hydrocarbon produced is significant as middle chain alkanes (C16–C19) are mainly produced by C. tropicalis when cultured in glucose, whereas long chain alkanes (C22–C25) are majorly produced when acetate or glycerol are used as the carbon source (Ladygina et al. 2006).
Investigations were also focused towards fungi as the source for hydrocarbon production and it was observed that it produces hydrocarbons majorly in its mycelia and spores and the content differed drastically between these two parts. In mycelia, about 0.06–0.7% of the dry biomass was the hydrocarbon content and mainly consisted of n-alkanes having a chain length of C15–C36 while spores accumulated about 0.004–0.015% of the dry biomass and contained mainly odd numbered n-alkanes (C27, C29, and C35). A summary of some of the important studies conducted in this area have been tabulated in Table 3.
From Table 3, it can be inferred that the yield of intracellular hydrocarbons is less and the only exception to the trend is V. furnissii which produces about 60% bioalkanes of its dry weight. Considering the low yield of internal hydrocarbons, their commercial viability as a transportation fuel is questionable considering the costly downstream processing step. Extracellular hydrocarbon production received prime importance owing to the possibility of the process to be commercialized. Bacteria (Desulfovibrio and Clostridium) majorly produce extracellular hydrocarbons where the distinguishing feature with its intracellular counterpart is the low chain length, C19–C21 and C16–C18 respectively in case of Clostridium pasteurianum and Desulfovibrio sulfuricans (Bagaeva and Zinurova 2004). The exchange of hydrocarbons from the interior to the exterior of the cell through the cell membrane is attributed to the short chain length of the hydrocarbons (Bagaeva and Zinurova 2004). The mode of production was mostly anaerobic in nature along with the presence of essential mineral salts and calcium lactate. Belyaeva et al. 1995 reported the production of short chain aliphatic hydrocarbons (C14–C25) under anaerobic conditions while using mineral salts and calcium lactate as the medium composition.
4.1.1 Metabolic Pathways for Hydrocarbon Production
Cyanobacteria have the capability of producing hydrocarbons through natural process but the yield is low, as only 0.12% of the biomass can be converted to bioalkanes/bioalkenes (Coates et al. 2014; Winters et al. 1969). Thus it can be inferred that through natural route, bioalkane/bioalkene production is not a viable option thus genetic engineering should be adopted for maximizing the yield of bioalkanes. Generally hydrocarbons are produced via fatty acids and the processing involves deformylation and decarboxylation of fatty aldehydes. Some prokaryotes viz., Feotgalicoccus sp. synthesize bioalkane through cytochrome P450 mediated decarboxylation of fatty acids. Apart from these steps, alkanes can also be produced by head-to-head condensation of fatty acids, organisms which undergo this procedure include Micrococcus luteus ATCC 4698.
Within cyanobacteria two distinct metabolic pathways have been deciphered for the production of hydrocarbons from fatty acids. One pathway involves the production of bioalkane from fatty acyl-ACPs via its conversion to fatty acyl aldehydes by fatty acyl-ACP reductase (FAAR) which is then converted to an odd chain length saturated alkane by the action of aldehyde deformylating oxygenase. A host of systems viz., plants, eukaryotes, prokaryotes, and humans also contain this catalytic machinery (FAAR). For this reaction to take place, NADPH is required since it provides a hydride for attacking the carbonyl of the fatty acid thioester. A schematic overview of the pathway is shown in Fig. 2 (Coates et al. 2014).

FAAR/ADO pathway. Asterisk Heptadecane, 8-Heptadecene, 7-Methylheptadecane
The second pathway is known as the olefin biosynthetic (OLS) pathway which was initially found in Synechococcus sp. and Prochloron didemni. This pathway was further modified by Mendez-Perez et al. (2011) where an alternate promoter was used to improve the productivity of the process. Gehret-Mccarthy et al. (2012) deciphered the structure of sulfotransferases (STs) in Moorea producens and Synechococcus sp. and observed that they had a similar structure and both the pathways lead to the production of bioalkanes. This pathway distinctly utilizes fatty acid as the substrate and produces odd numbered hydrocarbons with terminal olefinic bonds. Mendez-Perez et al. (2011) proposed that the OLS pathway includes ATP consuming fatty acyl ACP ligase (FAAL) which is responsible for transferring the fatty acid to OLS acyl carrier protein. The substrate bound to ACP then undergoes elongation by an extension module to add two carbons from malonyl-CoA thus reducing the β keto group. Gehret-McCarthy et al. (2012) observed that the final step of the OLS pathway involves sulfotransferase and thioesterase which transfers sulfonate from 3-phosphoadenosine-5-phosphosulfate to the β-hydroxyacyl substrate. This will activate the intermediate for catalysis using thioesterase (TE) to undergo hydrolysis from the enzyme surface followed by decarboxylation and desulfation to form terminal alkene product. The formation of terminal double bond is a unique structural signature of the OLS pathway compared to the FAAR/ADO pathway. A schematic overview of the OLS pathway is shown in Fig. 3.

OLS pathway for the production of terminal alkene
Some of the eukaryotic organisms also produce hydrocarbons via fatty acid decarboxylation pathways. Examples of these organisms include Botryococcus braunii race A which utilize an aldehyde decarbonylase for the production of bioalkanes, Gomphonema parvulum (hormosirene, a pheromone) utilizes hydroperoxide lyase, lepidoteran insects (7-methyl heptadecane, a pheromone) utilize P450 cytochrome to conduct fatty acid decarbonylase reaction ultimately leading to the formation of bioalkanes. However the biosynthetic pathways in case of eukaryotic organisms are completely different in comparison to the cyanobacterial pathway.
Till now the biosynthetic pathways discussed concerned the intracellular hydrocarbon production. Apart from these pathways, metabolic pathways also exist for the production of extracellular hydrocarbons. The pathway initiates with the formation of acetate and formate from carbon dioxide, followed by their reduction to aldehydes and then to bioalkanes via aldol condensation. A schematic representation of the metabolic pathway is shown in Fig. 4 (Ladygina et al. 2006).

Biochemical pathway for hydrocarbon synthesis by sulfate reducing bacteria
Since the yield of the production of hydrocarbons from microorganisms is not encouraging, therefore no experimental works have been conducted so far on bioconversion of biomass to bioalkanes/bioalkenes. Therefore, sophisticated biotechnological techniques should be explored for improving the yield of bioalkane/bioalkene production.
4.2 Genetic Engineering Approach
The drive for adopting genetic engineering techniques to modify microbes stems from the fact that naturally the yield obtained is insufficient to excite investors for putting their interest in this process. A number of studies have been conducted on Synechococcus sp. for improving the yield of bioalkanes/bioalkenes.
As per a study by Wang et al. 2013, genes responsible for biosynthesis of bioalkanes (sll0208 and sll0209) were simultaneously over expressed and the corresponding bioalkane production got improved. Wang et al. (2013) also suggested that Synechococcus sp. mutants over expressing two copies of sll0208 and sll0209 in both the loci slr0168 and slr1556 lead to higher alkane yield in comparison to the over expression of the genes at a single site (slr0168). The final yield obtained was 0.44 and 0.5 g/L for wild type and mutant strain (multiple site) respectively. In another approach, genes for alkane biosynthesis in Synechococcus elongatus PCC7942 (orf1593 and orf1594) and Nostoc punctiforme (npun1710 and npun1711) were expressed in Synechococcus sp. PCC6803 (Wang et al. 2013; Ladygina et al. 2006). It was observed that the activity of AAR and ADO got increased thus converting more amount of acyl-ACP to alkane/alkene.
Kageyama et al. (2015) studied the improvement in bioalkane production by transforming Anabena sp. with the bioalkane synthetic genes from A. halophytica and it was observed that the desired transformation lead to enhanced bioalkane production. Buijs et al. (2015) reported that with the deletion of hexadecenal dehydrogenase Hfd1 along with the expression of the gene responsible for bioalkane synthesis lead to the synthesis of long chain alkanes. Although a number of attempts have been made on improving the efficiency of alkane production through biological route, still the yield of bioalkane production is not encouraging. This issue can be tackled if the alkane production biosynthetic pathway is introduced into a strain having the capability of producing high quantities of free fatty acids. Some of the techniques developed for improving the free fatty acid production are as follows (Ruffing et al. 2013)
-
a.
Gene knock out of the free fatty acid recycling enzyme, acyl-ACP synthetase (aas): Rationale behind this strategy is to stop the natural mechanism of free fatty acid (FFA) consumption as well as recycling. The mechanism of recycling is mainly done through acyl-ACP synthetase gene (aas). The gene “aas” is homologous to acyl-CoA synthetase and is mainly responsible for the consumption of FFA through β-oxidation pathway. Thus the strategy adopted was to knock out this gene so that both consumption and recycling will be stopped and thus the FFA’s will be considered for bioalkane production. Using this strategy, Ruffing et al. (2013) successfully produced >40 mg/L of free fatty acids (FFAs) during the later stages of the stationary phase (after 200 h of incubation) for Synechococcus elongatus PCC7942 while the wild strain on the contrary accumulated negligible amount of FFA. Similar study was also conducted by Kaczmarzyk and Fulda (2010) on “aas” deficient Synechocystis sp. and Synechococcus sp. and observed free fatty acid concentration of 6.4 and 8.4 nmol mL−1 OD −1750 respectively.
-
b.
Insertion of thioesterase for releasing fatty acids from the acyl carrier protein: This strategy was adopted to detach fatty acids from the acyl carrier protein (ACP) so that the FFAs can be accepted as the substrates for bioalkane production. This ability of displacing free fatty acids (FFAs) from ACP is not found innately in Synechococcus elongatus PCC7942 thus, thioesterases from E. coli and Chlamydomonas reinhardtii were introduced into Synechococcus elongatus PCC7942 along with the knockout of “aas” gene (Ruffing et al. 2013). After conducting these modifications it was observed that approximately 30 mg/L of free fatty acids (FFAs) was produced in the S. elongatus PCC7942 but with no significant improvement in the yield in comparison to “aas” knockout. On the contrary Shin et al. (2016) successfully improved the production of FFAs by expressing signal sequence deficient acyl-CoA thioesterase I (TesA) in Escherichia coli cells and the major finding from their work was that the catalytic activity of TesA was more important than its expression for improving the FFA concentration.
-
c.
Enhancing the activity of RuBisCo: RuBisCo is responsible for fixing carbon dioxide and is thought to be the rate determining step of photosynthetic organisms considering the fact that it has low affinity for carbon dioxide and also due to its reversible nature. Thus its over expression may lead to improved production of FFAs. However, when it was over expressed along with “aas” knockout and TE expression, no significant improvement was observed. While the expression of nonnative RuBisCO subunits (rbcLS) in Synechococcus sp. PCC 7002 using psbAI promoter lead to greater than three folds increase in FFA production with a concentration of >130 mg/L (Ruffing 2014).
-
d.
Improving the conversion efficiency of acetyl-CoA to malonyl-CoA: This step is a key regulator where the flux of carbon can be controlled for either FFA production or energy generation and production of key biomolecules required for cell growth. Thus controlling this step will allow the optimum flow of carbon towards FFA production without compromising the cell growth. This step is controlled by the enzyme acetyl-CoA carboxylase (ACC) and if the gene corresponding to this enzyme is expressed then FFA production might improve. Similar quest was tried where four ACC genes (accBCDA) from C. reinhardtii CC-503 were expressed in S. elongatus PCC7942 along with “aas” deletion, TE expression and RuBisCO overexpression and it was observed that a mere 20 mg/L (approx) of FFA was produced (Ruffing et al. 2013). Detailed pathway of alkane biosynthesis along with the key step to control has been highlighted in Fig. 5.
Fig. 5 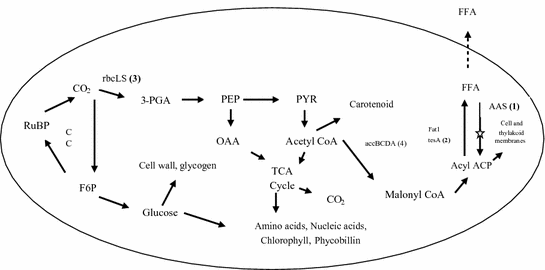
A pictorial view of the metabolic pathways of Synechococcus elongatus PCC7942. 3-PGA: 3-Phosphoglycerate; aas: acyl-ACP synthetase; ACC: Acetyl-CoA carboxylase; accA: ACC subunit, carboxyltransferase α subunit; acein; accC: ACC subunit, biotin carboxylase subunit; accD: ACC subunit, carboxyltransferase β subunit; ACP: Acyl carrier protein; F6P: Fructose-6-Phosphate; fat1: acyl-ACP thioesterase from C. reinhardtii CC-503; FFA: Free fatty acid; CC: Calvin cycle; rbc: RuBisCO; rbcL: RuBisCO large subunit; rbcS: RuBisCO small subunit; RuBisCO: ribulose-1,5-bisphosphate carboxylase/oxygenase; RuBP: ribulose-1,5-bisphosphate; TE: thioesterase; tesA: truncated thioesterase from E. coli; TCA: tricarboxylic acid; PEP: phosphoenolpyruvate; OAA: oxaloacetate; PYR: Pyruvate. The digits (1–4) denote the four strategies for improving the FFA production. denote deletion of the corresponding step

After screening all the available techniques for genetic manipulation it can be inferred that if alkane production machinery is successfully inserted into a high FFA producing organism, then there might be a chance for competitive yield of bioalkane/bioalkene for commercial applications. Thus there is an immense potential for metabolic engineers and molecular biologists to work together for engineering a suitable strain by cloning the required gene and its proper expression for hyperactive yield of bioalkane/bioalkene. Though it is not a very easy task to complete but with continuous efforts and by adopting integrated approach, the purpose of addressing the enigma of finding a potential biofuel can be addressed.
4.3 Application of Novel Enzymes
One of the crucial enzymes involved in alkane production is aldehyde deformylating oxygenase (ADO) since it converts the aldehydes to alkanes in cyanobacteria along with the formation of formic acid. During the reaction, NADPH provides the electrons by ferrodoxin-NADP reductase and ferrodoxin or via phenazine methosulfate. Studies conducted on ADO have suggested that the catalytic turnover of the enzyme is only 3–5 per catalytic site which justifies the low yield of the hydrocarbon production from the microbial source. Even more worrying factor is the inhibition of ADO by hydrogen peroxide which drastically limits its application for an efficiently converting aldehyde to alkane. This issue was tackled ingeniously by Andre et al. (2013) who successfully negated the inhibition caused due to hydrogen peroxide by applying a fusion protein which could encode both catalase and ADO leading to a two folds improvement in the catalytic conversion efficiency of ADO. Catalase was used since it had the capability of converting peroxide to water and oxygen and thus relieving ADO from inhibition.
5 Bottlenecks in the Production of Bioalkanes and Bioalkenes
Biotechnological means of bioalkanes/bioalkenes production is majorly limited by the product toxicity. Alkanes have effect on disorganizing cytoplasmic membrane integrity and its vital functions, such as the loss of ions, metabolites, proteins, and lipids, and the dissipation of the electrical potential and pH gradient. The toxic effect of alkanes (C9 and C10) have been examined with S. cerevisiae using transcriptome analysis by Ling et al. (2013) who suggested that these alkanes influenced various cellular metabolism such as induction of efflux pumps, membrane modification, energy supply, and radical detoxification. Cytosolic alkane toxicity can be overcome by inducing efflux pumps to facilitate alkane secretion from the cell. Heterologous expression of the efflux pumps “SNQ2P and PDR5P” could lower the toxicity of alkanes by reducing intracellular level of alkanes (Ling et al. 2013).
Apart from the product toxicity, limited supply of fatty acid biosynthetic (FAB) precursors and NADPH (cofactors) lowers the yield of alkane production. This problem can be dealt by heterologous expression of NADP+-dependent glyceraldehyde-3-phosphate dehydrogenase (GAPN) which can enhance the NADPH supply and improve the productivity of fatty acid-derived biofuels.
6 Conclusion
A detailed insight into the research concerning bioalkane/bioalkene synthesis through various means viz., chemical and microbial was conducted in this chapter. From this exercise it can be concluded that a myriad of chemical routes exist which can be efficiently used for bioalkane/bioalkene production. The most effective technique is deoxygenation which can utilize lignin and its derivatives (byproducts of a bioethanol refinery) for bioalkane/bioalkene synthesis. From the review on biotechnological perspective, it can be inferred that a majority of microorganisms have the innate capability of producing hydrocarbons out of which some are produced internally while the other are extracellularly produced. Hydrocarbons produced by the microorganisms serve as the biomarkers which indicate their source of its production. The type of hydrocarbons produced is hugely dependent upon the type of carbon source supplied in the growth media. Naturally the yield of bioalkane/bioalkene production from microorganisms is poor thus limiting its use for commercial applications and hence there is a huge demand for metabolic engineering tools to improve the yield. Studies carried out on metabolic engineering applications for improving bioalkane/bioalkene synthesis have suggested that if the required genetic machinery is expressed in high free fatty acid producing strain then there might be a chance for the commercialization of the process.
References
Albro PW (1976) Bacterial waxes. In: Kolattukudy PE (ed) Chemistry and biochemistry of natural waxes. Elsevier, Amsterdam, pp 419–445
Andre C, Kim SW, Yu XH, Shanklin J (2013) Fusing catalase to an alkane-producing enzyme maintains enzymatic activity by converting the inhibitory byproduct H2O2 to the cosubstrate O2. Proc Natl Acad Sci U S A 110:3191–3196
Bagaeva TV, Zinurova EE (2004) Comparative Characterization of Extracellular and Intracellular Hydrocarbons of Clostridium pasteurianum. Biochem (Moscow) 69:427
Belyaeva MI, Zolotukhina LM, Bagaeva TV (1995) Method for the production of liquid hydrocarbons. Invention Certificate SU2027760
Bird CW, Lynch JM (1974) Formation of hydrocarbons by microorganisms. Chem Soc Rev 3:309–328
Brocks JJ, Buick R, Summons RE, Logan GA (2003) A reconstruction of Archean biological diversity based on molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Hamersley Basin, Western Australia. Geochim Cosmochim Acta 67:4321–4335
Buijs NA, Zhou YJ, Siewers V, Nielsen J (2015) Long chain alkane production by the yeast Saccharomyces cerevisiae. Biotechnol Bioeng 112(6):1275–1279
Coates RC, Podell S, Korobeynikov A, Lapidus A, Pevzner P, Sherman DH, Allen EE, Gerwick L, Gerwick WH (2014) Characterization of cyanobacterial hydrocarbon composition and distribution of biosynthetic pathways. PLoS ONE. doi:10.1371/journal.pone.0085140
Corma A, Iborra S, Velty A (2007) Chemical routes for the transformation of biomass into chemicals. Chem Rev 107(6):2411–2502
de Beeck BO, Dusselier M, Geboers J, Holsbeek J, Morré E, Oswald S, Giebeler L, Sels BF (2015) Direct catalytic conversion of cellulose to liquid straight-chain alkanes. Energy Environ Sci 8:230–240
Dembitsky VM, Srebnik M (2002) Variability of hydrocarbon and fatty acid components in cultures of the filamentous cyanobacterium Scytonema sp. isolated from microbial community “black cover” of limestone walls in Jerusalem. Biochem (Moscow) 67(11):1276–1282
Deneyer A, Renders T, Aelst JV, Van den Bosch S, Gabriels D, Sels BF (2015) Alkane production from biomass: chemo-, bio- and integrated catalytic approaches. Curr Opin Chem Biol 29:40–48
Gehret-McCarthy J, Eisman EB, Kulkarni S, Gerwick L, Gerwick WH, Wipf P, Sherman DH, Smith JL (2012) Structural basis of functional group activation by sulfotransferases in complex metabolic pathways. ACS Chem Biol 7(12):1994–2003
Harger M, Zheng L, Moon A, Ager C, An JH, Choe C, Lai YL, Mo B, Zong D, Smith MD, Egbert RG, Mills JH, Baker D, Pultz IS, Siegel JB (2013) Expanding the product profile of a microbial alkane biosynthetic pathway. ACS Synth Biol 2(1):59–62
Hong S-B, Zalesky A, Cocchi L, Fornito A, Choi E-J, Kim H-H et al (2013) Decreased functional brain connectivity in adolescents with internet addiction. PLoS ONE 8(2):e57831
Huber GW, Iborra S, Corma A (2006) Synthesis of transportation fuels from biomass: chemistry, catalysts, and engineering. Chem Rev 106:4044–4098
Jankowski GJ, ZoBell CE (1944) Hydrocarbon production by sulfate-reducing bacteria. J Bacteriol 47:447
Jiménez-Díaz L, Caballero A, Pérez-Hernández N, Segura A (2016) Microbial alkane production for jet fuel industry: motivation, state of the art and perspectives. Microbiol Biotechnol. doi:10.1111/1751-7915.12423
Kaczmarzyk D, Fulda M (2010) Fatty acid activation in cyanobacteria mediated by acyl-acyl carrier protein synthetase enables fatty acid recycling. Plant Physiol 152(3):1598–1610
Kageyama H, Waditee-Sirisattha R, Sirisattha S, Tanaka Y, Mahakhant A, Takabe T (2015) Improved alkane production in nitrogen-fixing and halotolerant cyanobacteria via abiotic stresses and genetic manipulation of alkane synthetic genes. Curr Microbiol 71:115–120
Keasling JD (2010) Manufacturing molecules through metabolic engineering. Science 330:1355–1358
Ladygina N, Dedyukhina EG, Vainshtein MB (2006) A review on microbial synthesis of hydrocarbons. Process Biochem 41(5):1001–1014
Lee SK, Chou H, Ham TS, Lee TS, Keasling JD (2008) Metabolic engineering of microorganisms for biofuels production: from bugs to synthetic biology to fuels. Curr Opin Biotechnol 19:556–563
Lennen RM, Braden DJ, West RA, Dumesic JA, Pfleger BF (2010) A process for microbial hydrocarbon synthesis: overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol Bioeng 106:193–202
Ling H, Chen B, Kang A, Lee JM, Chang MW (2013) Transcriptome response to alkane biofuels in Saccharomyces cerevisiae identification of efflux pumps involved in alkane tolerance. Biotechnol Biofuels 6(1):95
McInnes AG, Walter JA, Wright JLC (1980) Biosynthesis of hydrocarbons by algae: decarboxylation of stearic acid to N-heptadecane in Anacystis nidulans determined by13C- and 2H-labeling and13C nuclear magnetic resonance. Lipids 15(9):609–615
Mendez-Perez D, Begemann MB, Pfleger BF (2011) Modular synthase-encoding gene involved in a-olefin biosynthesis in Synechococcus sp. strain PCC 7002. Appl Environ Microbiol 77(12):4264–4267
Nielsen J, Keasling JD (2011) Synergies between synthetic biology and metabolic engineering. Nat Biotechnol 29:693–695
Peralta-Yahya PP, Keasling JD (2010) Advanced biofuel production in microbes. Biotechnol J 5(2):147–162
Ruffing AM (2014) Improved free fatty acid production in cyanobacteria with Synechococcus sp. PCC 7002 as host. Front Bioeng Biotechnol 2:17
Ruffing AM, Trahan CA, Jones HDT (2013) Genetic engineering of cyanobacteria as biodiesel feedstock. SANDIA Report; SAND2013-0627, Sandia National Laboratories, California, USA
Schirmer A, Rude MA, Li X, Popova E, del Cardayre SB (2010) Microbial biosynthesis of alkanes. Science 329(5991):559–562
Shin KS, Kim S, Lee SK (2016) Improvement of free fatty acid production using a mutant acyl-CoA thioesterase I with high specific activity in Escherichia coli. Biotechnol Biofuels 9:208
Stone RW, ZoBell CE (1952) Bacterial aspects of the origin of petroleum. Ind Eng Chem Res 44:2564–2567
Wang W, Liu X, Lu X (2013) Engineering cyanobacteria to improve photosynthetic production of alka(e)nes. Biotechnol Biofuels 6:69
Wang WC, Tao L, Markham J, Zhang Y, Tan E, Batan L, Warner E, Biddy M (2016) Review of Biojet fuel conversion technologies. Technical Report NREL/TP-5100-66291, July 2016
Weete JD (1976) Algae and fungal waxes. In: Kolattukudy PE (ed) Chemistry and biochemistry of natural waxes. Elsevier, Amsterdam, pp 349–417
Winters K, Parker PL, Van BC (1969) Hydrocarbons of blue-green algae: geochemical signfficance. Science 163(3866):467–468
Xia Q, Chen Z, Shao Y, Gong X, Wang H, Liu X, Parker SF, Han X, Yang S, Wang Y (2016) Direct hydrodeoxygenation of raw woody biomass into liquid alkanes. Nat Commun 7:1–10
Xin J, Zhang S, Yan D, Ayodele O, Lua X, Wang J (2014) Formation of C-C bonds for the production of bio-alkanes under mild conditions. Green Chem 16:3589–3595
Zhang W, Chen J, Liu R, Wang S, Chen L, Li K (2014) Hydrodeoxygenation of lignin-derived phenolic monomers and dimers to alkane fuels over bifunctional zeolite-supported metal catalysts. ACS Sustain Chem Eng 2(4):683–691
Zhang Y, Bi P, Wang J, Jiang P, Wu X, Xue H, Liu J, Zhou X, Li Q (2015) Production of jet and diesel biofuels from renewable lignocellulosic biomass. Appl Energy 150:128–137
Zhao C, Camaioni DM, Lercher JA (2012) Selective catalytic hydroalkylation and deoxygenation of substituted phenols to bicycloalkanes. J Catal 288:92–103
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Gujjala, L.K.S., Kumar, S., Rajak, R.C., Banerjee, R. (2018). Bioalkanes and Bioalkenes: An Ecofriendly and Alternate Fuel in Bioenergy Research. In: Kumar, S., Sani, R. (eds) Biorefining of Biomass to Biofuels. Biofuel and Biorefinery Technologies, vol 4. Springer, Cham. https://doi.org/10.1007/978-3-319-67678-4_13
Download citation
DOI: https://doi.org/10.1007/978-3-319-67678-4_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-67677-7
Online ISBN: 978-3-319-67678-4
eBook Packages: EnergyEnergy (R0)