
Abstract
Along the Scheldt estuary, intertidal areas such as intertidal flats and intertidal marshes have important functions including flood water storage, water quality regulation and provision of important habitats. The functioning of intertidal marshes is determined by short-term geomorphic processes: the marshes are regularly flooded and sediments are deposited on the marsh surface, a process through which marshes gain elevation. The spatial patterns and rates of sedimentation on marshes are mainly governed by temporal and spatial variations in tidal characteristics and flow patterns, which are strongly influenced by marsh vegetation, leading to a micro-topography typical for tidal marshes. Strong human modifications of the Scheldt estuary by embankments, channelization and dredging have been increasing the tidal range, especially in the more upstream regions. As part of an integrated management plan for the Scheldt estuary—the so-called Sigmaplan in Flanders, new intertidal areas have been created and more than 2000 ha are planned to be ready by 2030. This intertidal flat and marsh creation is realized in some cases as flood control areas with a reduced tidal exchange between the estuary and the flood control area, where the geomorphologic processes are comparable to natural marshes, although we note some important differences.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
17.1 Introduction
The riverbanks of the Scheldt estuary are occupied by intertidal flats and marshes (Fig. 17.1). These are the areas that periodically flood and drain during flood and ebb tides, respectively.

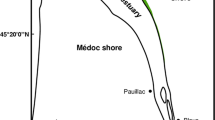
a Situation of the Scheldt estuary within NW-Europe, b overview of the Scheldt estuary with all the major intertidal flats, marshes and field sites and place names mentioned in the text
The lower areas, called intertidal flats, are flooded every semi-diurnal high tide. These areas are too dynamic (strong tidal currents and waves, intense sediment dynamics) or the inundation time is too long to support vegetation establishment. Higher areas have shorter hydroperiods (shorter inundation time and higher frequency) and are characterized by a dense vegetation cover of flood-tolerant plant species (Fig. 17.2). These areas are called intertidal marshes. The lower intertidal marshes are flooded every tide, while the highest marshes are only inundated during spring tides, i.e. the highest water levels of the fortnightly spring-neap tidal cycle (Fig. 17.3). Intertidal marshes are flooded and drained by channels, also called tidal creeks or tidal channels. The intertidal flats and marshes have important functions: they serve as natural flood control areas, they are sinks for nutrients, organic matter and sediments and sources of oxygen and they provide habitats for many species. These environments are in a geomorphic view very dynamic, with several processes even measurable during one single tide.

a View of the Paulina salt marsh at low water, b aerial picture of the Saeftinghe brackish marsh (https://beeldbank.rws.nl, Rijkswaterstaat/Joop van Houdt) and c view of intertidal flats and freshwater marshes from the Sea Scheldt close to Lippenbroek

a 14-day spring-neap tidal cycle as measured in Antwerp in 2015 (data: www.waterinfo.be), with schematic representation of marsh elevations in green and b a single semi-diurnal tide with different tidal phases
The historical evolution and especially anthropogenic influences such as construction of large-scale embankments and dredging have been altering the Scheldt estuary, leading to a rise of the tidal range (i.e. the difference between low and high tide water level). As a safety measure, several large flood control areas with new intertidal habitats have been (or are currently being) installed, which offer unique possibilities to study the geomorphological processes in intertidal areas that are in an early stage of geomorphological and ecological development. The Scheldt estuary, although nowadays strictly confined by historical land reclamation and dike building, still accommodates remains of a scenic natural habitat surrounded by a highly populated and built-up area of Belgium and the Netherlands.
In this chapter, we will discuss the current knowledge of geomorphic processes in the natural and newly created intertidal areas, especially the intertidal marshes, together with their effect on the functioning of these intertidal landscapes along the Scheldt estuary.
17.2 Geographical Setting
The estuary of the river Scheldt extends from the mouth at the North Sea near Vlissingen (NL) (0 km) to Ghent (160 km from the estuary mouth), where sluices impede further inland propagation of the tidal wave (Meire et al. 2005) (Fig. 17.1). Along this stretch, the salinity gradually decreases in landward direction, and therefore the intertidal marshes can be differentiated into salt, brackish and freshwater marshes (Fig. 17.1b, see Meire et al. (2005) for more information). The Saeftinghe brackish marsh, with an area of around 3000 ha is the biggest intertidal marsh area of the estuary and the southern North Sea coastal area (Fig. 17.2b). The other remaining marshes are small and scattered fringing the riverbanks. The Western Scheldt, the Dutch part of the Scheldt estuary near the mouth of the Scheldt River, is a multichannel system with extensive intertidal flats bordering the channels (Fig. 17.1b). On the Belgian territory, the tidal part is called the Sea Scheldt and consists of a single channel river with intertidal flats next to the subtidal channel.
17.3 Functions of Intertidal Flats and Marshes
17.3.1 Attenuation of Floods, Waves and Erosion
Intertidal areas and especially intertidal marshes play an important role in the water management of the estuary. They act as natural flood control areas, creating additional water storage capacity at extreme high water levels, thereby reducing peak water levels within the estuary and reducing the flood risks for nearby urban areas, such as Antwerp. The flood storage capacity of marshes is especially important at the Saeftinghe marsh and the Belgian part of the tidal river. Model analysis shows for example that the flood storage capacity of the Saeftinghe marsh decreased the water level by almost 20 cm in Antwerp during a storm tide in 2013 (Smolders et al. 2015). Furthermore, marshes strongly attenuate waves and tidal currents (Möller and Spencer 2002; Widdows and Brinsley 2002; Bouma et al. 2005a, b; Möller 2006; Van der Wal et al. 2008; Möller et al. 2014). As such they protect against or reduce erosion (Widdows and Brinsley 2002; Neumeier and Ciavola 2004; Neumeier and Amos 2006; Van der Wal et al. 2008; Möller et al. 2014). Dikes, artificially constructed levees for flood protection, need less maintenance if they are protected by marshes at the riverside and are less prone to dike breaches during storm floods (Möller 2006; Möller et al. 2014).
17.3.2 Filter Function
Marshes have an important filter function for the Scheldt water. The river water is heavily loaded with sediment, nutrients (e.g. nitrogen) and pollutants (e.g. heavy metals), originating from soil erosion, agricultural and industrial activities in the catchment of the Scheldt River, and from landward tidal pumping of sediments that are as such trapped within the estuary. High nutrient loads result in eutrophication with episodic anoxic water conditions, strongly limiting biological activity. Luckily, the anoxic episodes have become less common the last 10 years. Marshes, on the contrary, are sources of oxygen and silica and they trap nutrients (Gribsholt et al. 2005; Struyf et al. 2007), pollutants (Du Laing et al. 2007; Teuchies et al. 2013) and sediments (Temmerman et al. 2004b; Van Damme et al. 2009). Silica is an important building block of phytoplankton (diatoms), the primary producers that form the foundation of the estuarine food web. Hence the marshes can be considered as natural filters for the Scheldt River, which may contribute to reducing the costs for dredging and removing nutrients and pollutants. A similar filter functioning is also expected in realignment areas (i.e. de-embankments, with a landward retreat of the dike) along the Scheldt, as well as in the flood control areas with a reduced tide (areas with a sluice through the dike, that allows controlled tidal flooding of the land).
17.3.3 Habitat Function
Intertidal areas comprise important habitats for the ecosystem of the Scheldt estuary. Their nature value is acknowledged by several national and international directives (e.g. EU habitat and bird directives). For example, the Scheldt estuary is an important area for water birds in NW-Europe. Situated along the East Atlantic Flyway, 80 different species of waterfowl and waders are observed (Ysebaert et al. 2000; Van Den Bergh et al. 2005). The tidal wave reaches 160 km upstream making the Scheldt estuary one of the few remaining estuaries in Europe with a full salinity gradient from saltwater over brackish to freshwater marshes and associated plants and fauna (Van den Bergh et al. 2001). Biodiversity decreases with salinity. The salt marshes consist of a limited number of halophytes, plant species adapted to tolerate salinity such as glasswort (Salicornia sp.) and common cordgrass (Spartina anglica). These species are low herbs and grasses with a maximum vegetation height of 0.8 m (Temmerman et al. 2003b). Brackish marshes have a higher biodiversity, but freshwater marshes host the most diverse species composition, from herbs up to different trees such as willow (Salix sp.). Biodiversity and species composition not only change with salinity, but also with flooding frequency. Pioneer species may colonize the higher parts of the intertidal flat whereas the high marsh zones are vegetated by climax species (Van den Bergh et al. 2001). In salt marshes, glasswort (Salicornia sp.) and common cordgrass (S. anglica) are pioneer species, whereas Elytrigia atherica is the climax vegetation that is present on the high salt marshes and on higher channel-bordering levees, but absent on lower elevations. In brackish marshes, seaside bulrush (Bolboschoenus maritimus) occurs on the lower elevations, while common reed (Phragmites australis) is less salt tolerant and only present on the higher elevated areas. Schoenoplectus sp. are the pioneer species of the freshwater marshes. At higher elevations, common reed (P. australis) can reach exceptional heights of up to 4 m. Willow (Salix sp.) is the climax species in the freshwater zone of the Scheldt estuary.
Also small variations in the micro-topography of the marsh surface lead to distinct plant communities. These species patterns are also visible on Fig. 17.2b, where distinct patches of darker and lighter green vegetation are present, as a result of different vegetation communities (i.e. different species compositions). Hence the geomorphological changes influence habitat shifts in marshes. The geomorphic processes play an important role in the sustainable ecological management of the Scheldt estuary and affect the functioning of the marshes (see Sect. 17.7.4).
17.4 Pleistocene and Holocene Changes in the Scheldt River
During the Saale glacial period, the period of maximum ice sheet extent in North Europe and lowest sea level that lasted until ca. 130 ka BP, the main rivers in Flanders followed a northwest direction past Ghent, eroding a deep valley, called ‘The Flemish Valley’ (see Chap. 18). During the Weichselian, the last glacial period that lasted until ca. 12 ka BP, the Flemish Valley was gradually filled by mostly sandy deposits originating from a braided river system and covered with eolian sediments (De Moor 1983; Bogemans 1993). This resulted in a relatively flat landscape, only dissected in certain zones by the rivers (Bogemans et al. 2012). Near the end of the Weichselian, a sand ridge was formed that blocked the drainage of the rivers in the Flemish Valley to the northwest, forcing the main rivers to shift their course more eastwards, through the ‘Hoboken gap’, cutting through the cuesta of resistant Oligocene clay, towards the north (Kiden 1991, 2006). This is visible in Fig. 17.1b between the Rupel confluence and Hoboken where the Scheldt River follows a straight course towards the north. Near the end of the Weichselian the Scheldt River shifted also from a braiding to a meandering river, forming extensive fossil meanders, such as at Overmere Donk (Fig. 17.1b). The size of the fossil meanders indicates that discharges were 3–5 times larger than those of the contemporary Scheldt river (Kiden 1991). During the Late Dryas fluvial processes interacted with eolian processes, resulting in local dune formation in the river floodplain (Bogemans et al. 2012).
The first period of the Holocene was characterized by diminishing fluvial activity. It was a remarkable period of river stability with (almost) neither new river channel formation nor lateral erosion (Kiden 1991; Bogemans et al. 2012). The paleochannels and floodplains were gradually filled with gyttja, peat and organic to peaty clay (Kiden 1991). The tidal influence began ca. after 400 AD at the Belgian border. Brackish water intrusion stopped peat growth and clay was deposited. In Antwerp, peat could continue to grow until 500–600 AD, but the tidal influence in the more inland part of the river was still very limited until 1100 AD (Kiden 2006; Coen 2008). A nice example of this is an old meandering part of the Scheldt that was cut off in the thirteenth century AD. This cut-off part, located on the right bank between the Durme tributary and Temse (Fig. 17.1b), called the ‘Oude Schelde’ (Old Scheldt), has an average width of 75 m, a dimension that indicates limited or no tidal influence yet in the thirteenth century AD (Kiden 2006).
The tidal range and local high water level in the Scheldt River increased rapidly since about 1000–1100 AD (Fig. 17.4), by natural processes such as the development, widening and deepening of a new, shorter tidal connection to the North Sea (via the Western Scheldt, whereas before that period the connection to the North Sea was via the Eastern Scheldt) and elevation gain by sediment accretion of the intertidal areas, as well as by human impact, namely large-scale embankments of intertidal areas to create agricultural land. Since the middle of the 20th century, land reclamation has been driven by industrial and urban developments (Meire et al. 2005). The land reclamation in the 20th century resulted in a 16% loss of the total surface area of the estuary, mainly reducing the area of mature marshes.

In order to provide safe access to the port of Antwerp and to enable shipping traffic in more upstream areas of the Scheldt, several modifications altered the river system. In parts of the freshwater zone, the river channel was historically shortened and straightened, thereby reducing the sinuosity of the river (Van Braeckel et al. 2006). In the brackish and salt water zones downstream of the port of Antwerp, maritime access routes are being dredged on a large scale. Also upstream Antwerp the river is dredged for inland shipping. The dredging activities are concentrated on deepening the sills, i.e. shallow areas of the navigation channel. As a consequence, the bottom friction of the channel decreases and the tidal wave can propagate further upstream (Vroon et al. 1997). The average tidal range now increases from Vlissingen (3.8 m) to Antwerp (5.25 m). The maximal tidal range is observed upstream of Antwerp at Temse (5.41 m) and decreases to 2 m around Ghent (data 2001–2010, www.waterinfo.be, and Fig. 17.4). The continued increase of tidal range, tidal discharge and ebb and flood velocities enhanced erosion of the Scheldt river bed, making the river wider and deeper over the last centuries. This led also to an increase of high-dynamic areas. The increase of tidal range raised the high water levels. At the Durme tributary for example, the mean high water level rose 150 cm since 1860 (Van Braeckel et al. 2006) and the inundation risk increased accordingly.
To reduce the flood risk after severe storm floods in 1953 and 1976, the Delta Works and Sigmaplan were initiated along the Dutch (Western Scheldt) and Belgian sides (Sea Scheldt) of the Scheldt River, respectively (Meire et al. 2014). The Dutch Works consisted of conventional engineering solutions such as the cut-off of estuaries by large-scale dams or storm surge barriers (for example the Haringvliet sluices and the Eastern Scheldt storm surge barrier respectively) and by heightening the existing dikes. The more recent Belgian Sigma plan consisted of heightening of the dikes, as well as increasing the water storage capacity of the river by creating new intertidal areas and flood control areas (Table 17.1). This novel approach was adopted to guarantee the safety along with the naturalness and the functioning of the estuarine system. Some of the new intertidal areas are created by managed retreat or realignment of the dikes. Flood control areas (FCA’s), on the other hand, are low-lying areas, next to the estuary, which are surrounded by a high dike (Fig. 17.5). The dike at the river side is lowered (the overflow dike, see Fig. 17.5) so that water can flow over the dike during a storm tide, dissipating the flood wave energy and storing a large volume of water. As a result, the water level of the river lowers, reducing the flood risk for inhabited areas. In some FCA’s, a controlled reduced tide (CRT) will be introduced: a limited tidal water exchange can enter and exit the FCA-CRT through different sluices (Fig. 17.5). In the FCA-CRT’s we expect significant elevation changes, comparable with sedimentation and erosional processes in natural marshes (Cox et al. 2006; Maris et al. 2007).

Functioning of a flood control area with controlled reduced tides (FCA-CRT). At flood tide, water enters the area through inlet sluices and at ebb, the water flows back to the estuary through outlet sluices. In the FCA, the tidal range is much reduced compared to the estuary itself. On the right, overview of pilot area Lippenbroek (location see Fig. 17.1b) (Photo Vildaphoto/Yves Adams) with inlet sluice (inset) after Temmerman et al. (2013)
In total, almost 3000 ha of flood control area (without tides) will be installed and more than 2000 ha of new intertidal areas will be created by 2030 (Table 17.1). The new intertidal areas are either compensation for reclaimed marshes or needed to reach the favourable state of conservation according to the EU Habitat directive.
17.5 Intertidal Flats
Intertidal flats are the intertidal areas that flood every tide, positioned higher in the tidal frame than the subtidal river channel and lower than the vegetated intertidal marshes (Fig. 17.3). Although intertidal flats may look like uniform slopes next to the river channel, surprisingly different morphological features can be distinguished in these dynamic systems.
However, as the Scheldt River is continuously modified by maintenance dredging and sand extraction, the morphology of the tidal river is continuously shifting towards new equilibria. These dynamics, together with natural variations in, e.g. discharge and suspended sediment concentrations, change sedimentation or erosion patterns constantly. This is also evident in monitoring campaigns (e.g. Van Braeckel et al. 2014), where the substrate of the intertidal flat changes from field visit to field visit, and sedimentation or erosion patterns are not consistent between years, places or geomorphologic places (e.g. inner or outer river bend). This uncertainty makes prediction and generalization of the intertidal flat morphological processes very difficult, so we will only give a brief overview of intertidal flat features in the next sections.
17.5.1 Mudflat Versus Sandflat
The first difference that is obvious between different intertidal flats is the texture of the sediment, there are sandy and muddy intertidal flats, also called sandflats and mudflats.
The hydrodynamic exposure of a particular place determines the texture of the sediment that is deposited on or transported to the intertidal flats. Areas that experience strong ebb or flood currents or even waves have a sandy texture. Currents and waves keep small particles in suspension, while coarser particles settle, or are transported as bedload. In hydrodynamically sheltered areas, only fine-grained suspended mud (silt and clay sized particles) accumulates.
The differential deposition of sediment texture classes can be observed along an elevation gradient from the lowest intertidal flat areas, close to the subtidal channel to the highest flats at the marsh edge: the lower areas of the intertidal flat experience flooding during almost the whole tidal cycle and hence are generally subjected to stronger flood and ebb tidal currents compared to higher, mudflat areas that only flood at slack high water. This may result in finer sediments deposited at the higher portions of the intertidal flat. Such small changes in sediment texture can be observed at several locations along the Scheldt River (Van Braeckel et al. 2014), although at many locations the difference is negligible.
There is in general also a textural difference evident along the estuarine gradient: the more downstream, the sandier the texture of the intertidal flats. But also here there are many local exceptions in this overall tendency.
The repetition of flood currents, high water slack and ebb currents may lead to the deposition of alternating layers of coarse and fine material on the intertidal flats and marshes, called ‘tidal bedding’. During each ebb and flood phase with high flow velocities, coarse sediments are deposited, while during the high water phase, when the flow velocity decreases and changes direction, finer particles are deposited. In the same way, also spring and neap cycles or seasonal cycles may be recorded in the sediment. However, the abovementioned tidal bedding will only be observed in areas with continued sedimentation, rather than erosion. Such fine-layered tidal bedding is often visible in intertidal flat or marsh sediments, which can get exposed by vertical cliffs (ranging from a few dm to sometimes more than 1 m in height) originating from wave- and current-induced erosion (Fig. 17.6a).

Different morphological structures of intertidal flats. a Tidal flat with tidal bedding visible in the cliff (white arrow) and microripples in front of the cliff, b Microcliff and c meseta morphology caused by Vaucheria sp. (Photos (c) Research Institute for Nature and Forest (INBO))
17.5.2 Erosion on Intertidal Flats
Where erosion is the dominant process on the intertidal flats, different erosional processes can be observed.
When waves or currents erode the intertidal flat deposits or underlying hard substrates, such as clay or peat layers, a variety of geomorphological forms can be observed (De Smedt 1969; Van Braeckel et al. 2014), resulting from differential erosion between harder and softer substrates (Fig. 17.6):
-
Microcliffs: small cliffs facing the low water line.
-
Micromeseta: small plateaus with eroding edges at both sides.
-
Microcuesta: the escarpment facing the marsh edge, the backslope inclining towards the low water line.
During the dry period, water seeping out of the marsh on the mudflat can erode rills and small channels. In general, steeper slopes of the mudflat result in straight, single channels, while meandering, dentritic channels form on gentle mudflat slopes (De Smedt 1969; Van Braeckel et al. 2009). On sandy flats, the steep slopes do not show any geomorphological features such as channels, since water can infiltrate the sandy substrate. On the flat surfaces, however, micro-, meso- or macroripples are often visible, indicators of tidal currents and waves (Fig. 17.6a). The slope of the intertidal flats generally decreases downstream in the estuary, with average slopes >15% towards Ghent to <5% at the Dutch border (Van Braeckel et al. 2014). This corresponds to the decrease of the average width of the river. In the gap of Hoboken, where the river cuts into resistant Oligocene clay (see Sect. 17.4), the slopes are steeper than expected.
A special geomorphological pattern on the intertidal flats may arise by diatoms. Diatoms may accumulate on the intertidal flats, forming a visible, smooth thick brown biofilm on top of the sediment, inhibiting erosion and increasing sedimentation of fine-grained particles (Weerman et al. 2010; van de Koppel et al. 2012). If the diatoms form discontinuous films, erosion of the lower unprotected areas results in a regular pattern with higher diatom-covered hummocks and lower depressions. The same process is also visible when Vaucheria sp., a kind of green algae, is present on the intertidal flats (Fig. 17.6c).
It is important to note that only 10% of the riverbanks of the Sea Scheldt are still natural. All the other parts of the Scheldt river banks are reinforced by riprap, on the intertidal flat or at the intertidal marsh edge (Van Braeckel et al. 2009). These structures prevent erosion of the underlying intertidal flats or the marsh cliff (see next section).
17.6 Intertidal Flat—Marsh Edge
The transition from intertidal flat to marshes can be gradual or very abruptly marked by a cliff. While marshes occur on a (gentle) slope towards the mudflat, pioneer species can expand to the lower mudflats, as does for instance the brackish pioneer species B. maritimus at the Dutch-Flemish border. In the freshwater marshes, common reed (Pragmites australis) is often found in the lower parts of a gentle marsh slope (Van Braeckel et al. 2014). Both species can, under favourable circumstances, laterally expand at rates >2 m/year (Van Braeckel et al. 2014).
Cliffs are characterized by a steep elevation difference between the low intertidal flat and the higher marsh platform and are observed along different salt (van de Koppel et al. 2005; Van der Wal et al. 2008), brackish (Van der Wal et al. 2008) and freshwater marshes (Brys et al. 2005) in the Scheldt estuary. Cliff formation is promoted by differential sedimentation rates on the marsh and the adjacent mudflat, steepening the marsh front and hence making it more vulnerable to wave-induced erosion (Allen 2000; Van de Koppel et al. 2005). At the edge of marsh plateaus, lateral cliff retreat through undercutting can result in marsh rejuvenation (Van de Koppel et al. 2005; Van der Wal et al. 2008) when the hydrodynamic conditions and the elevation of the intertidal flat in front of the cliff are favourable for pioneer species re-colonization. As the marsh further expands over the intertidal flat in front of the cliff, the abandoned cliff may remain visible within the vegetated marsh. Active cliffs and marsh rejuvenation are visible at several marshes along the Western Scheldt such as the Paulina marsh (van de Koppel et al. 2005; Van der Wal et al. 2008). At the Sea Scheldt, however, more than 90% of the river banks are reinforced with riprap (Van Braeckel et al. 2009) and this hinders cliff erosion and marsh rejuvenation.
17.7 Intertidal Marshes
Intertidal marshes are the vegetated areas whose elevation allows flooding only during relatively short periods around high water level (Fig. 17.3). A marsh typically starts with a pioneer species that colonizes the intertidal flat at elevations just below the high water level at neap tides (Fig. 17.3). The vegetation reduces the water flow velocity, thus promoting sedimentation. As the marsh platform accretes, other plant species will also colonize the marshes. The higher the marsh platform rises, the less frequent it will be inundated, reducing accretion, until it only gets flooded by the higher spring tides (Fig. 17.3). However, marshes can also disappear by cliff erosion (see previous section) or by drowning, if they get more and more flooded, for example when relative sea level rise is faster than the sediment accretion rate. In the Scheldt estuary, horizontal expansion or retreat of the marshes is limited to those in the Western Scheldt because most edges at the Sea Scheldt are reinforced with riprap. The geomorphological and sedimentary processes within marshes and newly created flood control areas will be discussed in detail in the next paragraphs. We first focus on vertical sediment accretion on the marsh surface.
17.7.1 Sedimentation in Time and Space
The suspended sediment concentration (SCC), i.e. the mass of dry sediment per volume of water (in g/l), determines the maximum sediment deposition rate that can occur on a marsh surface. The SSC is determined by the incoming river/sea water, although local scouring can entrain sediment from the creek bed and banks. The SSC strongly fluctuates during a spring-neap tidal cycle and throughout the year (Fettweis et al. 1998; Temmerman et al. 2003b). For example, SSC measurements at the Notelaar, a freshwater marsh just downstream Temse (Fig. 17.1b), showed a variation between 10 and 300 mg/l (Temmerman et al. 2003b). In general, the sediment concentration is higher when the marsh is flooded by higher tides (spring tides), due to higher water velocities (Bouma et al. 2005a); additionally, this relationship between SSC and high tide level has a steeper slope in winter, i.e. for a similar high tide level the SSC is higher in winter than in summer (Fettweis et al. 1998; Temmerman et al. 2003b). The total sediment deposited on a marsh surface will follow the tidal and seasonal variations in incoming SCC. As a result, most sedimentation is occurring during the highest spring tides in winter (Temmerman et al. 2003b).
Sedimentation on the marsh surface shows a clear spatial pattern (Fig. 17.7a). A corresponding digital elevation model is shown in Fig. 17.7b. Most of the sediment is deposited in the low-elevated marshes, close to the marsh edge. The lower the marsh elevation, the higher the inundation frequency and hence the higher the sediment deposition rate (Stoddart et al. 1989; Cahoon and Reed 1995; Temmerman et al. 2003b). On the elevated marshes further away from the marsh edge, less sedimentation occurs, the variation being also determined by the distance to the nearest tidal creek. A greater distance from a tidal creek results in lower deposition rates (Reed et al. 1999; Allen 2000; Christiansen et al. 2000; Temmerman et al. 2003b). Sedimentation over marsh platforms is dominated by tidal advective processes (Allen 2000; Temmerman et al. 2003b), hence the spatial patterns of sediment deposition can be explained by the water flow over the marsh. The tidal inundation starts at the lower marsh edge. These are the lowest areas and flooded first. As the water level rises, the tidal channels are filled. Once the tidal channels are full, the water flows from the channels over the surrounding marsh platform. Once the water level exceeds the elevation of the marsh platform, the tidal flow will also directly flow from the Scheldt River over the platform. The sediment-laden water loses progressively more suspended sediment along its flow path from the marsh edge to the vegetated marsh platform (Christiansen et al. 2000; Friedrichs and Perry 2001; Moskalski and Sommerfield 2012). Consequently the largest, heaviest particles (fine sand) are deposited close to the marsh edge and the tidal channels. The remaining fine-grained sediment (silt and clay) is deposited further away on the marsh platform (Allen 2000; Temmerman et al. 2003b, 2005a). When the water level rises higher than the vegetation canopy and the vegetation is deeply submerged, the water will flow as sheet flow from the river over the marsh platform (Temmerman et al. 2005a; Vandenbruwaene et al. 2013).

a Aerial image of Paulina marsh (see Fig. 17.1 for location). The bars represent cumulative sedimentation during a spring-neap tidal cycle (ca. 14 days) on 50 locations. Reddish colours represent marsh vegetation, greenish colours the intertidal flat and channels. b Elevation map of the same area (green to red represent low to high elevation). White boxes correspond to the modelling areas of Fig. 17.8. After Temmerman et al. (2006)
In FCA-CRT’s, the water volume that flows in and out every tide is set by the tidal height in the estuary and sluice configuration, and contrary to natural intertidal marshes, independent of the site elevation. Classical feedback mechanisms, which slow down sedimentation rates when marshes grow higher in elevation, function therefore differently in a CRT (Vandenbruwaene et al. 2011a). A small amount of sedimentation is wanted for the geomorphic development to a marsh in FCA-CRT’s, but too much sedimentation will affect the storage capacity (see Sect. 17.7.4). Measurements reveal that the initial sedimentation rate is strongly related to the inundation depth and inundation time, with rates similar to those of natural marshes (Vandenbruwaene et al. 2011a). As sedimentation is related to tidal characteristics, which are regulated by the sluice system, sedimentation rates can be controlled in an FCA-CRT.
17.7.2 Crucial Impact of Vegetation
Marshes have a dense vegetation cover (e.g. Fig. 17.2). In general, if water flows through dense vegetation, the flow velocity is strongly reduced due to the friction exerted by the vegetation canopy on the water flow, thereby promoting sedimentation (Allen 2000; Leonard and Reed 2002; Bouma et al. 2005a; Leonard and Croft 2006; Neumeier and Amos 2006). In intertidal marshes, however, the vegetation has a more complex effect that is crucial for erosion and sedimentation. This is illustrated by a computer model (Delft3D) of Paulina marsh (see location in Fig. 17.1b) that incorporates the physical laws of water flow and sedimentation over a marsh surface with vegetation (Temmerman et al. 2005b, 2007). To demonstrate the impact of vegetation, we simulated flow velocities and sedimentation on the same marsh topography, but without vegetation (Temmerman et al. 2012). In other words, we simulated the flow and sedimentation patterns that we should observe if we mowed all the vegetation on an existing marsh platform. The objective of these model simulations is to show the impact of vegetation on the flow and sedimentation patterns on an intertidal marsh by comparing the simulated patterns with presence and absence of vegetation. Of particular interest are the areas on the marsh edge (Fig. 17.8a–d) and along an existing creek (Fig. 17.8e–h):

(a–d) Results of model simulations at the marsh edge (upper white box in Fig. 17.7b): flow pattern with (a) and without (b) marsh vegetation; sedimentation pattern with (c) and without (d) marsh vegetation. (e–h) Results of the model simulations at the elevated marsh (lower white box in Fig. 17.7b): flow pattern with (e) and without (f) marsh vegetation; sedimentation pattern with (g) and without (h) marsh vegetation. Figure after Temmerman et al. (2005b) uv: flow velocity in m s-1; sr: sedimentation rate in g m-1
-
At the transition between the unvegetated intertidal flats and vegetated marshes, the flow velocities are strongly reduced within the vegetation by the friction exerted by the vegetation canopy (Fig. 17.8a). In between vegetated areas, however, the flow converges and velocities are enhanced. Apart from the model simulations, this effect is also empirically observed (Vandenbruwaene et al. 2011b Bouma et al. 2013). As a consequence of this flow pattern, the model shows that sediment is deposited within the vegetation, while in between the vegetation, very low sedimentation rates or even erosion can take place (Fig. 17.8c). This effect of vegetation patches on sedimentation within and erosion around the patches has also been empirically observed (Bouma et al. 2007).
-
On the mature high marsh with deep tidal creeks, the simulated water velocities are the highest in the creeks (Fig. 17.8e): here the friction is low in absence of vegetation. Once the water level exceeds the level of the creek banks, the creek water starts to flood the high vegetated marsh platform where the flow velocities quickly drop. These simulations are in accordance with field measurements of flow velocity patterns (Temmerman et al. 2012).
-
The model simulations show that in absence of vegetation (and hence friction) on the marsh platform, the water will flow easily from the Scheldt river over the unvegetated marsh platform instead of flowing first through the creeks (Fig. 17.8b, F). Therefore the flow velocities in the creeks will be lower. Hence, the flow patterns on vegetated marshes lead to limited sedimentation or erosion in the tidal creeks. In the absence of vegetation, lower velocities in the creeks could lead to creek infilling and lower sedimentation rates on the marsh platform (Fig. 17.8d, h). This simulated effect of presence or absence of marsh vegetation on flow velocities in the tidal creeks dissecting the marsh are also confirmed by field measurements, where marsh vegetation was experimentally mowed (Temmerman et al. 2012).
These simulations and measurements show that interactions between vegetation, flow hydrodynamics and sedimentation-erosion dynamics play an essential role in the long-term geomorphic development of the intertidal marsh platform, leading to the erosion of tidal creeks in between vegetation patches (D’Alpaos et al. 2006; Temmerman et al. 2007; Vandenbruwaene et al. 2013) and to the development of a levee-basin topography on the vegetated marsh platform (Temmerman et al. 2004a, 2005b; Mudd et al. 2013; Vandenbruwaene et al. 2013, 2015): along the tidal creeks, elevated levees form that are typically 20–30 cm higher than the lower basins further from the tidal creeks. If vegetation is absent such as on intertidal flats, levees are generally not developed along the tidal channels that dissect the surface, while they are typically present along the tidal channels present in vegetated marshes (Vandenbruwaene et al. 2015). This vegetation-induced micro-topography can, in turn, enhance the flow concentration towards the channels (Vandenbruwaene et al. 2015).
In FCA-CRT’s, spontaneous channel formation already took place after one year of tidal action, but vegetation did not yet have a significant influence on the channel extension (Vandenbruwaene et al. 2012). Here, channel formation is influenced by the initial topography and the underlying high erosion-resistant polder clay (Vandenbruwaene et al. 2012).
17.7.3 Long Term Effect of Sedimentation
In the previous paragraphs we mentioned that sedimentation on the short time scale of single tides is strongly governed by the inundation frequency, inundation depth and inundation period. On the long term (decades to centuries), the flooding dynamics will change. Marshes build up by sedimentation, but the high water levels also change, e.g. due to relative sea-level rise or construction of embankments (see Sect. 17.4). Relative sea-level rise (RSLR) can cause marshes to become more and more flooded, ultimately drowning them and leading to marsh die-off on a large scale (for example in the Mississippi delta (Penland et al. 2000), Chesapeake Bay marshes (Kearney et al. 1988; Schepers et al. 2017), Venice Lagoon (Carniello et al. 2009)). In other estuaries, however, marshes elevate with a rate similar to RSLR (van der Wal and Pye 2004; Kirwan and Megonigal 2013). It is clear that these different evolutions of marshes with respect to RSLR have important implications for their functioning.
How do the marshes of the Scheldt estuary evolve with changing high water levels? Measurements and coring of an old (>200 year) and a young (ca. 50 year) freshwater marsh (The Notelaar, just downstream of Temse, see Fig. 17.1b) give insight in long-term sedimentation rates representative for the Scheldt River (Temmerman et al. 2003a) (Fig. 17.9):

Sedimentation of an old marsh (dark blue, dotted) and a young marsh (light blue, dashed) of the Notelaar (location just downstream of Temse, Fig. 17.1b) during the last 60 years, dated by sediment cores (after Temmerman et al. 2003a). The black line shows the evolution of the mean high water level at the same site
-
Young, low-elevated marshes are frequently flooded by the tides, with a long inundation time and depth, so that the initial sedimentation rate is high (e.g. 6–8 cm year−1 between 1950 and 1965). Similar high elevation gains, up to 1.5 m in 15–30 year were also measured in Saeftinghe in the period 1931–1963 (Wang and Temmerman 2013) (see location in Fig. 17.1b). As a consequence of this elevation gain relative to the mean high water level (MHWL), the flooding frequency, duration and depth decrease gradually. Therefore, the sedimentation rate also decreases. This negative feedback between sedimentation and inundation frequency, inundation time and inundation depth leads to vertical accretion of marsh surfaces up to an equilibrium level, around 10–20 cm above MHWL in the Scheldt estuary (Temmerman et al. 2004a; Wang and Temmerman 2013). As MHWL rises due to RSLR and human modifications to the estuary such as embankment building, the marshes build up and follow the MHWL rise.
-
At older, high-elevated marshes (already at the equilibrium level), sedimentation rates are comparable to the rates of MHWL rise (Fig. 17.8). At the Notelaar, this is 1.2–1.8 cm year−1.
This explains the generally flat topography of intertidal marshes. However, there is still a micro-topography of levees and basins related to channel systems (see Sect. 17.7.2). Research has shown that once the micro-topography of marshes has established, accumulation on the higher levees and in the lower marsh basins occurs at the same rate, in equilibrium with the rate of MHWL rise (Temmerman et al. 2004a), so the micro-topography of the marshes is preserved with long-term accretion.
These morphodynamics do not only occur in the marshes of the Notelaar but also apply very similarly to other marshes along the Scheldt river (Temmerman et al. 2004b). However, there are also regional differences: young marshes elevate faster in the more inland Sea Scheldt than in the more seaward Western Scheldt, due to higher suspended sediment concentrations in the inland part. Also older marshes rise faster in the Sea Scheldt by a faster MHWL rise rate.
As for the response to increased sea level rise, model results show that the marshes of the Scheldt estuary could keep up with MHWL rise even if MHWL should rise 1.5 times faster in the future than during the last 70 years (Temmerman et al. 2004b). However, if the sediment concentration is halved in this scenario of accelerated MHWL rise, the marshes are no longer able to keep up with it and they will be increasingly flooded, leading to important vegetation changes and maybe even vegetation die-off. Note that in these analyses we do not take into account lateral erosion of the marshes (i.e. cliff erosion).
In realignment areas such as Sieperda marsh (bordering on the east side to Saeftinghe, Fig. 17.1b), sedimentation rates measured close to the Scheldt reach similar high values (up to 3–5 cm year−1) as in young marshes, due to the low elevation. This will continue until the high water level is reached (Verbeek and Storm 2001). In the FCA-CRT’s, we already mentioned that the sedimentation is related to the inundation depth and inundation time, determined by the initial topography before introduction of the tide. So the lower areas fill up faster than higher elevated marshes that inundate less frequently, leading to a levelling of the topography (not taking into account the formation of tidal creeks and levee-basin micro-topography). In the long term, model analyses show that the marsh elevation–sedimentation feedback (i.e. rising marsh elevation resulting in decreasing inundation depths and hence reduced rate of elevation increase) is absent in FCA-CRT’s (Maris et al. 2007; Vandenbruwaene et al. 2011a) because the water volume that is exchanged every tide is not determined by the marsh elevation of the FCA-CRT compared to the water level (as in natural marshes) but depends on (adjustable) sluice dimensions. Hence, in the long term, FCA-CRT’s could eventually build up to an equilibrium elevation close to the MHWL of the Scheldt.
17.7.4 Impacts of Geomorphological Processes on the Functioning of Marshes and Flood Control Areas
The water control capacity of intertidal marshes and flood control areas might decline by the natural process of sedimentation and subsequent rise of the marsh elevation. The long-term effect of sedimentation to the water control capacity is dependent on the type of intertidal or flood control area. (i) Natural, low-elevated marshes or realignments elevate rapidly by high, frequent and long inundations. Examples are the young marshes of the Notelaar downstream of Temse, ca. 50 cm in 11 year (1947–1958) (Temmerman et al. 2003a) and Saeftinghe, ca. 150 cm in 32 year (1931–1963) (Wang and Temmerman 2013). This might affect the water storage capacity on natural marshes. However, not only the water storage capacity but also the elevation of intertidal marshes is important for storm tide attenuation, whereby higher marsh elevations provide more storm tide attenuation compared to lower marsh elevations (Smolders et al. 2015). (ii) Flood control areas with a reduced tide store flood water that flows over the overflow dike (Fig. 17.5). Observations from Lippenbroek, an installed FCA-CRT (Fig. 17.5), show that in the lower elevated areas, initial sedimentation rates in the first year after installation reached 13.4 cm year−1 (Vandenbruwaene et al. 2011a). As the lowest areas fill up, these high sedimentation rates decrease (currently 6.3 cm year−1). Averaged over the whole area, sedimentation rate in the Lippenbroek FCA-CRT is now 2.35 cm year−1, comparable with natural marshes. Although sedimentation will continue in the FCA-CRT’s, the amount of water (and the amount of sediment) that enters the FCA-CRT can be controlled by the inlet sluice dimensions. In this way, the sluice dimensions can be regulated to keep the flood water storage capacity as high as possible, while letting in enough water for the development of an estuarine ecosystem. (iii) In FCA’s without a reduced tide, only during the highest storm tides (one or two times a year) will flood water enter the FCA, depositing very little sediment with no impact on the flood control capacity of the area.
The elevation gain also affects the filter function of intertidal marshes and FCA’s. As sedimentation decreases with increasing elevation, also the rate at which sediments are removed from the river water decreases. So we can expect that in a short term, the realignments of marshes and the installation of FCA-CRT’s will have a positive effect (due to high sedimentation rates), but eventually this effect will decline.
The filter function of nutrients and pollutants is somewhat more complex and still poorly understood on a longer timescale, since biogeochemical cycling is controlled by sediment structure and sub-surface hydrology (Spencer and Harvey 2012). Nevertheless, marshes and FCA-CRT’s remove up to 1 kg of dissolved inorganic nitrogen per tide per ha from the water column, as is observed in the freshwater marshes of Tielrode (at the mouth of the Durme tributary, see Fig. 17.1b) (Gribsholt et al. 2005) and the FCA-CRT of Lippenbroek.
Finally, also the ecological functioning of marshes will be affected by sedimentation processes. Fast build-up of marsh elevation drives a fast vegetation succession from pioneer species to a climax vegetation (Temmerman et al. 2003a). This is also one of the reasons why low-elevated marshes with pioneer vegetation are scarce in the estuary. Large elevation differences are rapidly levelled out by the inundation–sedimentation feedback, inevitably leading to a loss in habitat diversity and biodiversity. In FCA-CRT’s, where the inundation–sedimentation feedback is not present, the inundation frequency and hence the species composition will be determined by the elevation distribution within the area and the sluice dimensions.
17.8 Synthesis and Conclusion
We showed that the geomorphology of intertidal flats and marshes along the Scheldt estuary is very dynamic. The precise processes leading to sedimentation or erosion on intertidal flats are complex, producing a diversity of geomorphic features. We also detailed the morphological developments of intertidal marshes. On a short term and a local scale, the sedimentary processes are determined by the tidal characteristics (flooding frequency, depth and period) and the flow patterns over the marshes, in turn strongly influenced by the marsh vegetation. The resulting micro-topography drives plant species composition, with different species on the slightly higher levees compared to the inner marsh basins.
Extensive embankments decimated the former marsh areas of the Scheldt estuary and, together with dredging and other anthropogenic modifications or natural changes, altered the tidal characteristics. In general, the tides rose significantly in the estuary (Fig. 17.4). In the long term, marshes build up relatively rapidly to an equilibrium elevation around mean high water level. This process led to the few remaining mature marshes that we still see along the river banks of the Scheldt, now usually protected from cliff erosion and marsh rejuvenation by ripraps.
Increased flood risk led to the creation of more than 500 ha of new intertidal areas as an ecosystem-based contribution to flood defense foreseen in the so-called Sigmaplan. In restored marshes with controlled reduced tides, we observed similar processes as in the natural marshes, though some processes are governed by the sluice configuration rather than the elevation of the site. Since the natural process of sedimentation reduces some functions of marshes and flood control areas, continued monitoring is needed in order to follow up and adjust the sluice configuration in order to guarantee an optimal combination of the different marsh functions (flood safety, filtering of the water and valuable ecological habitats).
This research highlights that understanding of present-day geomorphic processes is of utmost importance for a sustainable management of the Scheldt estuary. Hence, a good cooperation between scientists and policy makers is needed to safeguard the estuary as a navigation channel, as a safe work and residential area and as a valuable natural area.
References
Allen JRL (2000) Morphodynamics of Holocene salt marshes: a review sketch from the Atlantic and Southern North Sea coasts of Europe. Quat Sci Rev 19:1155–1231. doi:10.1016/S0277-3791(99)00034-7
Bogemans F (1993) Quaternary geological mapping on basis of sedimentary properties in the eastern branch of the Flemisch Valley. Mem Geol Surv Belgium 35
Bogemans F, Meylemans E, Jacops J, Perdaen Y, Storme A, Verdurmen I, Deforce K (2012) The evolution of the sedimentary environment in the lower River Scheldt valley (Belgium) during the last 13,000 a BP. Geol Belgica 15:105–112
Bouma TJ, De Vries MB, Low E, Kusters L, Herman PMJ, Tánczos IC, Temmerman S, Hesselink A, Meire P, van Regenmortel S (2005a) Flow hydrodynamics on a mudflat and in salt marsh vegetation: identifying general relationships for habitat characterisations. Hydrobiologia 540:259–274. doi:10.1007/s10750-004-7149-0
Bouma TJ, De Vries MB, Low E, Peralta G, Tánczos IC, Van De Koppel J, Herman PMJ (2005b) Trade-offs related to ecosystem engineering: a case study on stiffness of emerging macrophytes. Ecology 86:2187–2199. doi:10.1890/04-1588
Bouma TJ, van Duren LA, Temmerman S, Claverie T, Blanco-Garcia A, Ysebaert T, Herman PMJ (2007) Spatial flow and sedimentation patterns within patches of epibenthic structures: Combining field, flume and modelling experiments. Cont Shelf Res 27:1020–1045. doi:10.1016/j.csr.2005.12.019
Bouma TJ, Temmerman S, van Duren LA, Martini E, Vandenbruwaene W, Callaghan DP, Balke T, Biermans G, Klaassen PC, van Steeg P, Dekker F, van de Koppel J, de Vries MB, Herman PMJ (2013) Organism traits determine the strength of scale-dependent bio-geomorphic feedbacks: a flume study on three intertidal plant species. Geomorphology 180–181:57–65. doi:10.1016/j.geomorph.2012.09.005
Brys R, Ysebaert T, Escaravage V, Van Damme S, Van Braeckel A, Vandevoorde B, Van Den Bergh E (2005) Afstemmen van referentiecondities en evaluatiesystemen in functie van de KRW : afleiden en beschrijven van typespecifieke referentieomstandigheden en/of MEP in elk Vlaams overgangswatertype vanuit de—overeenkomstig de KRW—ontwikkelde beoordelingssystemen voor biologische kwaliteitselementen. Report of the Research Institute for Nature and Forest (IN.O.2005.7).
Cahoon D, Reed D (1995) Relationships among marsh surface topography, hydroperiod, and soil accretion in a deteriorating Louisiana salt marsh. J Coast Res 11:357–369.
Carniello L, Defina A, D’Alpaos L (2009) Morphological evolution of the Venice lagoon: evidence from the past and trend for the future. J Geophys Res Earth Surf 114:1–10. doi:10.1029/2008JF001157
Christiansen T, Wiberg PLL, Milligan TGG (2000) Flow and sediment transport on a tidal salt marsh. Estuar Coast Shelf Sci 50:315–331. doi:10.1006/ecss.2000.0548
Coen I (2008) De eeuwige Schelde? Ontstaan en ontwikkeling van de Schelde. Report of Flanders Hydraulics Research. Borgerhout
Cox T, Maris T, De Vleeschauwer P, De Mulder T, Soetaert K, Meire P (2006) Flood control areas as an opportunity to restore estuarine habitat. Ecol Eng 28:55–63. doi:10.1016/j.ecoleng.2006.04.001
D’Alpaos A, Lanzoni S, Mudd SM, Fagherazzi S (2006) Modeling the influence of hydroperiod and vegetation on the cross-sectional formation of tidal channels. Estuar Coast Shelf Sci 69:311–324. doi:10.1016/j.ecss.2006.05.002
De Moor G (1983) Cryogenic structures in the Weichselian deposits of Northern Belgium and their significance. Polarforschung 53:79–86
De Smedt P (1969) Geomorfologie van slikken en schorren langsheen het Schelde-estuarium op Belgisch grondgebied. Acta Geogr Lovan 7:49–63
Du Laing G, Vandecasteele B, De Grauwe P, Moors W, Lesage E, Meers E, Tack FMG, Verloo MG (2007) Factors affecting metal concentrations in the upper sediment layer of intertidal reedbeds along the river Scheldt. J Environ Monit 9:449. doi:10.1039/b618772b
Fettweis M, Sas M, Monbaliu J (1998) Seasonal, neap-spring and tidal variation of cohesive sediment concentration in the Scheldt Estuary, Belgium. Estuar Coast Shelf Sci 47:21–36. doi:10.1006/ecss.1998.0338
Friedrichs CT, Perry JE (2001) Tidal salt marsh morphodynamics: a synthesis. J Coast Res SI 27:7–37
Gribsholt BTH, Boschker S, Struyf E, Andersson M, Tramper A, De Brabandere L, van Damme S, Brion N, Meire P, Dehairs F, Middelburg JJ, Heip CHR (2005) Nitrogen processing in a tidal freshwater marsh: a whole-ecosystem 15 N labeling study. Limnol Oceanogr 50:1945–1959. doi:10.4319/lo.2005.50.6.1945
Kearney MS, Grace RE, Stevenson JC (1988) Marsh loss in Nanticoke Estuary, Chesapeake Bay. Geogr Rev 78:205. doi:10.2307/214178
Kiden P (1991) The lateglacial and holocene evolution of the middle and lower River Scheldt, Belgium. In: Starkel L, Gregory KJ, Thornes JB (eds) Temperate palaeohydrology. Fluvial processes in the temperate zone during the Last 15,000 Years, pp 283–299
Kiden P (2006) Schelde in Belgie en Zuidwest-Nederland na de laatste ijstijd. Belgeo 3:279–294
Kirwan ML, Megonigal JP (2013) Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504:53–60. doi:10.1038/nature12856
Leonard LA, Reed DJ (2002) Hydrodynamics and sediment transport through tidal marsh canopies. J Coast Res 36:459–469
Leonard LA, Croft AL (2006) The effect of standing biomass on flow velocity and turbulence in Spartina alterniflora canopies. Estuar Coast Shelf Sci 69:325–336. doi:10.1016/j.ecss.2006.05.004
Maris T, Cox T, Temmerman S, De Vleeschauwer P, Van Damme S, De Mulder T, Van den Bergh E, Meire P (2007) Tuning the tide: creating ecological conditions for tidal marsh development in a flood control area. Hydrobiologia 588:31–43. doi:10.1007/s10750-007-0650-5
Meire P, Ysebaert T, Van Damme S, Van Den Bergh E, Maris T, Struyf E (2005) The Scheldt estuary: a description of a changing ecosystem. Hydrobiologia 540:1–11. doi:10.1007/s10750-005-0896-8
Meire P, Dauwe W, Maris T, Peeters P, Coen L, Deschamps M, Rutten J, Temmerman S (2014) Estuaries in focus sigma plan proves efficiency: the recent “Saint-Nicholas” storm surge in the Scheldt estuary: the Sigma plan proves its efficiency! ECSA Bull 62:19–23
Möller I (2006) Quantifying saltmarsh vegetation and its effect on wave height dissipation: results from a UK East coast saltmarsh. Estuar Coast Shelf Sci 69:337–351. doi:10.1016/j.ecss.2006.05.003
Möller I, Spencer T (2002) Wave dissipation over macro-tidal saltmarshes: effects of marsh edge typology and vegetation change. J Coast Res 521:506–521. ISSN:0749-0208
Möller I, Kudella M, Rupprecht F, Spencer T, Paul M, van Wesenbeeck BK, Wolters G, Jensen K, Bouma TJ, Miranda-Lange M, Schimmels S (2014) Wave attenuation over coastal salt marshes under storm surge conditions. Geosci, Nat. doi:10.1038/ngeo2251
Moskalski SM, Sommerfield CK (2012) Suspended sediment deposition and trapping efficiency in a Delaware salt marsh. Geomorphology 139–140:195–204. doi:10.1016/j.geomorph.2011.10.018
Mudd SM, Fagherazzi S, Morris JT, Furbish DJ (2013) Flow, sedimentation, and biomass production on a vegetated salt marsh in South Carolina: toward a predictive model of marsh morphologic and ecologic evolution. In: The ecogeomorphology of tidal marshes. American Geophysical Union, pp 165–188. doi:10.1029/CE059p0165
Neumeier U, Ciavola P (2004) Flow resistance and associated sedimentary processes in a Spartina maritima salt-marsh. J Coast Res 202:435–447
Neumeier U, Amos CL (2006) The influence of vegetation on turbulence and flow velocities in European salt-marshes. Sedimentology 53:259–277. doi:10.1111/j.1365-3091.2006.00772.x
Penland S, Wayne L, Britsch D, Williams SJ, Beall AD, Butterworth V (2000) Geomorphic classification of coastal land loss between 1932 and 1990 in the Mississippi river delta plain, Southeastern Louisiana. USGS Open File Report 00-417
Reed DJ, Spencer T, Murray AL, French JR, Leonard L (1999) Marsh surface sediment deposition and the role of tidal creeks: implications for created and managed coastal marshes. J Coast Conserv 5:81–90. doi:10.1007/BF02802742
Schepers L, Kirwan M, Guntenspergen G, Temmerman S (2017) Spatio-temporal development of vegetation die-off in a submerging coastal marsh. Limnology and Oceanography 62:137–150. doi: 10.1002/lno.10381
Smolders S, Plancke Y, Ides S, Meire P, Temmerman S (2015) Role of intertidal wetlands for tidal and storm tide attenuation along a confined estuary: a model study. Nat Hazards Earth Syst Sci 15:1659–1675. doi:10.5194/nhess-15-1659-2015
Spencer KL, Harvey GL (2012) Understanding system disturbance and ecosystem services in restored saltmarshes: Integrating physical and biogeochemical processes. Estuar Coast Shelf Sci 106:23–32. doi: 10.1016/j.ecss.2012.04.020
Stoddart DR, Reed DJ, French JR (1989) Understanding salt-marsh accretion, Scolt Head Island, Norfolk. England. Estuaries 12:228. doi:10.2307/1351902
Struyf E, Temmerman S, Meire P (2007) Dynamics of biogenic Si in freshwater tidal marshes: Si regeneration and retention in marsh sediments (Scheldt estuary). Biogeochemistry 82:41–53. doi:10.1007/s10533-006-9051-5
Temmerman S, Govers G, Meire P, Wartel S (2003a) Modelling long-term tidal marsh growth under changing tidal conditions and suspended sediment concentrations, Scheldt estuary, Belgium. Mar Geol 193:151–169. doi:10.1016/S0025-3227(02)00642-4
Temmerman S, Govers G, Wartel S, Meire P (2003b) Spatial and temporal factors controlling short-term sedimentation in a salt and freshwater tidal marsh, scheldt estuary, Belgium, SW Netherlands. Earth Surf Process Landforms 28:739–755. doi:10.1002/esp.495
Temmerman S, Govers G, Meire P, Wartel S (2004a) Simulating the long-term development of levee-basin topography on tidal marshes. Geomorphology 63:39–55. doi:10.1016/j.geomorph.2004.03.004
Temmerman S, Govers G, Wartel S, Meire P (2004b) Modelling estuarine variations in tidal marsh sedimentation: response to changing sea level and suspended sediment concentrations. Mar Geol 212:1–19. doi:10.1016/j.margeo.2004.10.021
Temmerman S, Bouma TJ, Govers G, Lauwaet D (2005a) Flow paths of water and sediment in a tidal marsh: relations with marsh developmental stage and tidal inundation height. Estuaries 28:338–352. doi:10.1007/BF02693917
Temmerman S, Bouma TJ, Govers G, Wang ZB, De Vries MB, Herman PMJ (2005b) Impact of vegetation on flow routing and sedimentation patterns: three-dimensional modeling for a tidal marsh. J Geophys Res Earth Surf 110:1–18. doi:10.1029/2005JF000301
Temmerman S, Govers G, Bouma TJ, De Vries M, Wartel S, Meire P (2006) Opslibbing van schorren en overstromingsgebieden langs de Schelde: een onvermijdelijk natuurlijk proces. Water 26:1–9
Temmerman S, Bouma TJ, van de Koppel J, van der Wal D, De Vries MB, Herman PMJ (2007) Vegetation causes channel erosion in a tidal landscape. Geology 35:631–634. doi:10.1130/G23502A.1
Temmerman S, Moonen P, Schoelynck J, Govers G, Bouma TJ (2012) Impact of vegetation die-off on spatial flow patterns over a tidal marsh. Geophys Res Lett 39:1–5. doi:10.1029/2011GL050502
Temmerman S, Meire P, Bouma TJ, Herman PMJ, Ysebaert T, De Vriend HJ (2013) Ecosystem-based coastal defence in the face of global change. Nature 504:79–83. doi:10.1038/nature12859
Teuchies J, Vandenbruwaene W, Carpentier R, Bervoets L, Temmerman S, Wang C, Maris T, Cox TJS, Van Braeckel A, Meire P (2013) Estuaries as filters: The role of tidal marshes in trace metal removal. PLoS One 8:e70381. doi:10.1371/journal.pone.0070381
Van Braeckel A, Piesschaert F, Van den Bergh E (2006) Historische analyse van de Zeeschelde en haar getijgebonden zijrivieren. 19e eeuw tot heden. Report of the Research Institute for Nature and Forest (INBO.R.2006.29). Brussel
Van Braeckel A, Mikkelsen JH, Dillen J, Piesschaert F, Van den Bergh E, Coen L, De Mulder T, Ides S, Maximova T, Peeters P, Plancke Y, Mostaert F (2009) Inventarisatie en historische analyse van Zeescheldehabitats Vervolgstudie: resultaten van het tweede jaar. Report of the Research Institute for Nature and Forest (INBO.IR.2009.34)
Van Braeckel A, Elsen R, Van den Bergh E (2014) MONEOS—geomorfologie. Hoogteraaien van slik en schor in de Zeeschelde. Evolutie van toestand tot 2012. Report of the Research Institute for Nature and Forest (INBO.R.2014. 1860252). Brussel
Van Damme S, Frank D, Micky T, Olivier B, Eric S, Britta G, Oswald VC, Patrick M (2009) Tidal exchange between a freshwater tidal marsh and an impacted estuary: the Scheldt estuary, Belgium. Estuar Coast Shelf Sci 85:197–207. doi:10.1016/j.ecss.2009.08.005
van de Koppel J, Van der Wal D, Bakker JP, Herman PMJ (2005) Self-organization and vegetation collapse in salt marsh ecosystems. Am Nat 165:E1–E12. doi:10.1086/426602
van de Koppel J, Bouma TJ, Herman PMJ (2012) The influence of local- and landscape-scale processes on spatial self-organization in estuarine ecosystems. J Exp Biol 215:962–967. doi:10.1242/jeb.060467
Van den Bergh E, Huiskes A, Criel B, Hoffmann M, Meire P (2001) Biodiversiteit op de Scheldeschorren. Levende Nat 102:62–66
Van Den Bergh E, Ysebaert T, Meire P (2005) Water bird communities in the Lower Zeeschelde: long-term changes near an expanding harbour. Hydrobiologia 540:237–258. doi:10.1007/s10750-004-7147-2
van der Wal D, Pye K (2004) Patterns, rates and possible causes of saltmarsh erosion in the Greater Thames area (UK). Geomorphology 61:373–391. doi:10.1016/j.geomorph.2004.02.005
Van der Wal D, Wielemaker-Van den Dool A, Herman PMJ (2008) Spatial patterns, rates and mechanisms of saltmarsh cycles (Westerschelde, The Netherlands). Estuar Coast Shelf Sci 76:357–368. doi:10.1016/j.ecss.2007.07.017
Vandenbruwaene W, Maris T, Cox T, Cahoon DR, Meire P, Temmerman S (2011a) Sedimentation and response to sea-level rise of a restored marsh with reduced tidal exchange: comparison with a natural tidal marsh. Geomorphology 130:115–126. doi:10.1016/j.geomorph.2011.03.004
Vandenbruwaene W, Temmerman S, Bouma TJ, Klaassen PC, De Vries MB, Callaghan DP, Van Steeg P, Dekker F, Van Duren LA, Martini E, Balke T, Biermans G, Schoelynck J, Meire P (2011b) Flow interaction with dynamic vegetation patches: implications for biogeomorphic evolution of a tidal landscape. J Geophys Res Earth Surf 116:1–13. doi:10.1029/2010JF001788
Vandenbruwaene W, Meire P, Temmerman S (2012) Formation and evolution of a tidal channel network within a constructed tidal marsh. Geomorphology 151–152:114–125. doi:10.1016/j.geomorph.2012.01.022
Vandenbruwaene W, Bouma TJ, Meire P, Temmerman S (2013) Bio-geomorphic effects on tidal channel evolution: Impact of vegetation establishment and tidal prism change. Earth Surf Process Landforms 38:122–132. doi:10.1002/esp.3265
Vandenbruwaene W, Schwarz C, Bouma TJ, Meire P, Temmerman S (2015) Landscape-scale flow patterns over a vegetated tidal marsh and an unvegetated tidal flat: implications for the landform properties of the intertidal floodplain. Geomorphology 231:40–52. doi:10.1016/j.geomorph.2014.11.020
Verbeek H, Storm C (2001) Tidal wetland restoration in The Netherlands. J Coast Res SI 27:192–202
Vroon J, Storm C, Coosen J (1997) Westerschelde, stram of struis? Eindrapport van het Project Oostwest, een studie naar de beïnvloeding van fysische en verwante biologische patronen in een estuarium. Report RIKZ, 97.023.
Wang C, Temmerman S (2013) Does biogeomorphic feedback lead to abrupt shifts between alternative landscape states?: an empirical study on intertidal flats and marshes. J Geophys Res Earth Surf 118:229–240. doi:10.1029/2012JF002474
Weerman EJ, van de Koppel J, Eppinga MB, Montserrat F, Liu Q-X, Herman PMJ (2010) Spatial self-organization on intertidal mudflats through biophysical stress divergence. Am Nat 176:E15–E32. doi:10.1086/652991
Widdows J, Brinsley M (2002) Impact of biotic and abiotic processes on sediment dynamics and the consequences to the structure and functioning of the intertidal zone. J Sea Res 48:143–156. doi:10.1016/S1385-1101(02)00148-X
Ysebaert T, Meininger PL, Meire P, Devos K, Berrevoets CM, Strucker RCW, Kuijken E (2000) Waterbird communities along the estuarine salinity gradient of the Schelde estuary, NW-Europe. Biodivers Conserv 9:1275–1296. doi:10.1023/A:1008976306651
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter
Cite this chapter
Schepers, L., Maris, T., Meire, P., Temmerman, S. (2018). The Scheldt Estuary: An Overview of the Morphodynamics of Intertidal Areas. In: Demoulin, A. (eds) Landscapes and Landforms of Belgium and Luxembourg. World Geomorphological Landscapes. Springer, Cham. https://doi.org/10.1007/978-3-319-58239-9_17
Download citation
DOI: https://doi.org/10.1007/978-3-319-58239-9_17
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-58237-5
Online ISBN: 978-3-319-58239-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)