
Abstract
The biomineralization process is genetically controlled, and it is the result of the metabolic activity of different organisms . Microorganisms , plants and animals produce calcium biomineralizations , calcium oxalates and carbonates being the most representative. The Quaternary pedosedimentary sequences of the southeastern coast of Buenos Aires province evolved from bioclastic and loess sediments, reworked by water and/or wind action . Calcium biomineralizations play an important role in the development of soils and contribute to differentiate sedimentary levels affected by pedogenesis . This work aims to characterize calcium biomineralizations in bioclastic , loessic and fluvio-eolian pedosedimentary sequences , typical of coastal environments of the southeastern Buenos Aires province . Modal soil profiles were defined in pedosedimentary sequences of the fluvio-eolian and coastal plains , in which disturbed and undisturbed samples were analyzed. Samples were analyzed at different scales of resolution: mesoscopic , microscopic and submicroscopic , using optical microscopy and scanning electron microscopy/energy-dispersive X-ray spectroscopy (SEM/EDX) . Organic matter content , pH , particle size distribution , mineralogy and calcium content were also quantified. Three geopedological units were differentiated in a sequence developed on loess mantle deposits: dunes and interdunes , coastal lagoon/paleo-coastal lagoon and shell ridges . In soils with incipient development, calcium biomineralizations are scarce, mainly associated with isolated bioclasts . In soils with more pedological development , bioclasts are affected by bioerosion through microorganism action (fungi and algae ), and subsequently calcium re-precipitated as secondary oxalates and carbonates (biomineralizations ). These biomineralizations also weakly add or bind skeletal components , incorporating themselves into the matrix of soils and sediments . The type and diversity of calcium biomineralizations increase directly in relation with time and pedogenetic evolution ; so, these biomineralizations have been determinant in the origin, evolution and resistance to natural and anthropic degradation of the late Quaternary pedosedimentary sequences of southeastern Buenos Aires province , Argentina .
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The biomineralization process is genetically controlled; it generates paracrystalline or crystalline biominerals and amorphous compounds , produced by the metabolic activity of different organisms . Biomineralizations have accompanied the evolution of life from the earliest appearance of biota on the planet, thus being present at all levels of the biosphere (Lowenstan 1981; Osterrieth 2004; Coe et al. 2014). Biomineralizations give a selective advantage to organisms that produce them and favor several biological functions ; for example, biominerals and amorphous compounds act as a sink of ions that can be reutilized when necessary (Simkiss and Wilbur 1989). Calcium , iron and silicon biomineralizations are the most common in the planet, and among them, calcium carbonate polymorphisms are the most widely found (Mann 2001).
Calcium biomineralizations are produced by microorganisms (bacteria , algae , fungi , actinomycetes ), plants and animals (Graustein et al. 1977; Osterrieth and Oyarbide 1998; Osterrieth et al. 1998). Calcite , vaterite and aragonite are the major minerals in animals , whereas in plants , calcium oxalate —both whewellite (dihydrated, CaC2O4.2H2O) and weddellite (polyhydrated, CaC2O4.(2+x)H2O)—is the most representative (Arnott 1982). These biomineralizations originate through processes mediated by an organic matrix , structure or mold: organisms constitute a structure or mold where the ions are introduced and induced to precipitate and crystallize. In fungi , from the first identification by Schmidt in 1847 (Horner et al. 1983), the presence of calcium biomineralization has been widely studied in vegetative and reproductive structures (Osterrieth et al. 1998). The biomineralization process occurs intra- and/or extracellularly, and it is associated with vegetative growth , degradation of organic resource and the activity of mycorrhizal species. The latter produces calcium oxalate salts on the walls of the hyphae by the reaction of excreted oxalic acid and calcium available in the environment (Verrecchia et al. 1993). Species producing calcium oxalate crystals include mycorrhizal basidiomycetes : Hysterangium crassum, Paxillus involutus (Lapeyrie et al. 1990); ascomycetes : Dasyscypha capitate; and mucorales : Mucor mucedo, M. phimbesis, Cunninghamella echinulata (Horner et al. 1983; Cromack et al. 1979) and Trichoderma koningii recently included among the fungi that produce calcium biomineralizations (Osterrieth et al. 1998; Oyarbide et al. 2001).
Calcium biomineralizations generate structures with a variable structural complexity ranging from single crystals to aggregates (Mann 2001). The aggregated units are usually placed in an order, and the crystallographic axes are partially or entirely aligned. These ordered structures are generally differentiated in a number of microarchitectural units , each of which is wrapped by a film, coating or organic membrane called biofilm (Lowenstam 1981; Jahren 1996; Addadi et al. 2003).
In the last thirty years, calcium biomineralizations have been widely reported in sedimentary sequences of different environments in the world (James 1972; Klappa 1980; Verrecchia et al. 1993, 1996). Research has also increased on the role in the pedological processes of Calcimorphic and Rendzina soils , among others (Verecchia et al. 2006; Tuason et al. 2009; Aragno et al. 2010; Dhami et al. 2013) and in the genesis and degradation of calcretes (Verrecchia and Verrecchia 1994; Verrecchia 1990, 1994; Verrechia et al. 1995). The latter processes are the result of a biochemical sequence that starts as calcium oxalate—whewellite (CaC2O4.H2O) and/or weddellite (CaC2O4.2H2O)—and ends as calcite (CaCO3) in a stable and definitive form (Verrecchia and Verrecchia 1994; Verrecchia 1990, 1994; Verrechia et al. 1993; Loysi et al. 1999; Osterrieth 2005).
Bioerosion is another process associated with biomineralization . It occurs due to the dissolution or erosion of the surface of minerals , fissures and the drilling of minerals and/or biominerals . This process has been underestimated by scientists, in spite of its high impact on all environments, especially in calcium biomineralizations developed in aqueous environments (Warme 1975). These processes are of great interest in the research on corrosion of both natural and artificial materials (archaeological and paleontological materials , artwork , monuments , metal alloys , steels , etc.).
The Quaternary pedosedimentary sequences of the southeastern coast of Buenos Aires province evolved from bioclastic and loess sediments, reworked by water and/or wind action . They have been affected by a succession of marine and terrestrial actions, associated with sea level changes , which have generated landforms and microlandforms in a changing water regime . During the last sixty years, they have been studied from geological , sedimentary , paleontological and paleoecological perspectives (Ameghino 1908; Frenguelli 1935, 1950; Fidalgo et al. 1973, 1991; Violante 1992; Violante et al. 2001; Isla 1989; Schnack et al. 1982, 1984; Schnack and Gardenal 1979; Tricart 1973; Osterrieth and Martínez 1993; Osterrieth and Schnack 1984; Osterrieth 2005; Bortolus et al. 2001; Espinosa 1994, etc.).
Closely associated with coastal landforms , different soil types are developed, in which calcium biomineralizations play an important role. Soil is open, complex, pluri-structural and multifunctional, with numerous definitions according to the discipline involved in its analysis. In summary, “soil is a natural resource that varies continuously in space and time,” whose structure and function are little known, and whose properties are transient and change in space and time (Jenny 1941; Simonson 1959; Ibáñez et al. 2000).
The coastal pedosedimentary sequences developed during the late Quaternary have acquired diagnostic features and experienced environmental changes of varying intensity in short time intervals, resulting in a succession of specific soil bodies (Osterrieth and Schnack 1984; Osterrieth and Martínez 1993; Osterrieth 1998). Differentiating sedimentary levels affected by pedogenesis is not always easy: it depends on the intensity with which it disturbed the parent materials , and on whether there were erosional processes , diagenetic calcification or decalcification , saturation conditions , syn- or post-pedogenetic processes, etc. Few studies use tools originated in pedosedimentary studies based on interdisciplinary biomineralogical studies. Our approach is part of the challenge to pursue studies in a field that is still weak in Argentina .
This work aims to characterize calcium biomineralizations in bioclastic , pedosedimentary sequences , typical of coastal environments of southeastern Buenos Aires province .
2 Materials and Methods
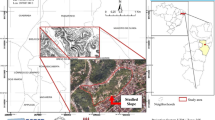
The site under study is located in the coastal areas of Mar Chiquita and General Pueyrredón counties , Buenos Aires province (34°–39°S and 57°–63°W) (Fig. 1). After detailed preliminary field studies, modal soil profiles were defined in pedosedimentary sequences of the fluvio-eolian plain and coastal plain (Schnack et al. 1982). Across a toposequence , the horizons of the modal profiles, disturbed and undisturbed samples were described according to Soil Taxonomy (1996) (Figs. 1 and 2).

Map of the study area. Location of toposequence and geopedological units (UGIA, UGIB, UGIC), and profiles studied (1–5)

Toposequence , geopedological units , pedosedimentary and modal profiles studied (modified of Schnack et al. 1982)
For each sample, pH in 1:1 paste was measured with a digital Orion Research 501 pH meter , the organic matter content was calculated (Walkley 1965; Black 1965) and the particle size distribution was assessed with sieve and pipette analysis (Ingram 1971; Galehouse 1971). Calcium content was determined according to Dawis and Freitas (1970).
In all the disturbed samples, loose grains, peds , skeletal fraction and matrix were analyzed at different scales of resolution: mesoscopic (binocular magnifying glass ), microscopic (polarization microscope , PM, Olimpus BX51P) and submicroscopic (scanning electron microscopy—SEM —JEOL JSM 6460 LV), and semi-quantitative studies of elementary components through microdispersive analysis with X-ray spectrometer (EDXS) , operating between 15 and 25 kV. Mineralogical studies were carried out concentrating light and heavy minerals with sodium polytungstate at a density of 2.89 g/cm3.
Undisturbed samples of 30 × 30 mm and 1 × 1 mm were analyzed with polarized and scanning electron microscopy according to Brewer (1964) and Bullock et al. (1985). The identification and description of calcium oxalates and calcium carbonates were carried out according to Verrecchia et al. (1995) and Verrecchia and Verrecchia (1994), respectively. The characterization of calcified filaments was carried out based on the definition of Klappa (1979).
Dating by 14C was performed at LATYR (Universidad Nacional de La Plata, Argentina) on bioclastic material of Heleobia sp., Tagelus plebeius and Mactra isabelliana.
3 Results and Discussion
The study area is located in the coastal lagoon of Mar Chiquita and is part of the Pampa , as defined by Frenguelli (1950). The study site includes the following geomorphological zones : coastal dunes , marginal plains , shell ridges and continental deposits (Schnack and Gardenal 1979) (Figs. 1 and 2). The sedimentary characteristics vary in relation to the climate , the water body and the alternation of transgressive–regressive coasts from the Late Pleistocene to the present. The studied sequences evolved from ancient coastal deposits linked to the last transgressive–regressive cycle of the Late Pleistocene and Holocene . Discordant deposits of the Mar Chiquita Formation are located over the Santa Clara Formation (Schnack et al. 1982), composed by Pampean loess and silts (Frenguelli 1950). The Mar Chiquita Formation is composed of both marine beach facies , formed by ridges of an age of approximately 6000 years BP (Schnack et al. 1982) and coastal lagoon facies formed by estuarine deposits with abundant whole and partially fragmented bioclasts . These mollusk shells include Heleobia sp., radiocarbon dated at 1670 ± 70 year BP, T. plebeius , dated between 1710 ± 60 and 2880 ± 90 year BP, and M. isabelliana , at the bottom of the profile, dated at 2820 ± 80 year BP (Schnack et al. 1982; Osterrieth 1998). This area of study has been closely associated with the evolution of the coastline and/or to the fluctuations of the sea level which took place during the transgressive–regressive cycle of the Holocene (Schnack et al. 1982).
The climate is mesothermic and subhumid , with little or no water deficiency (Burgos and Vidal 1951). The annual precipitation averages 809 mm. The average annual temperature is 13.7 °C; the mean minimum temperature reaches 8.1 °C in June, with an average maximum of 19.8 °C in January (Servicio Meteorológico Nacional of Argentina at the city of Mar del Plata, according to the 1920–1980 records).
The study area is an ecotone with specific biotic characteristics. These environments , broadly characterized as wetlands , are among the richest in the world in terms of their high biological diversity of both vertebrates and invertebrates (Mistch and Gopsselink 1993). Plant communities of coastal lagoons support very complex food chains , the dominant communities being “flechillar ,” wet meadows , salty meadows , “Espartillar ,” “Duraznillar ,” “Hunquillar ” and dune vegetation (Vervoorst 1967; Isacch 2001).
The dominant soil assemblages in the study area are composed of Udipsamments on coastal dunes , calcimorphic soils and Rendzinas on shell ridges , saline and calcimorphic soils , Endoaquolls , Calciaquolls and Hapludolls on estuarine sediments of the low and flooded areas surrounding the Mar Chiquita lagoon .
Such geopedological characteristics have conditioned the evolution of this complex coastal system , where pedological processes associated with calcic biomineralizations have furthered the understanding of their genesis. The work was made on five modal profiles across a toposequence (12 km), in the coastal and mixed typical geopedological units (Figs. 1 and 2).
4 Mixed Coastal Environment
Three geopedological units were differentiated in a sequence developed on loess mantle -shaped deposits: dunes and interdunes (UGIA) (Profiles 1 and 2), coastal lagoon/paleo-coastal lagoon in the Mar Chiquita lagoon marginal floodplain (UGIB) (profiles 3 and 4) and shell ridges (UGIC) (Profile 5).
5 Geopedological Unit of Dunes and Interdunes (UGIA)
The studied soils are located in the dune barrier extending from the Mar Chiquita lagoon to Punta Rasa (Fasano et al. 1982) (Figs. 1 and 2).
Soils in dune crests (Profile 1) are subject to strong wind and marine activities (Fig. 3a). They are sandy Mineral soils (typic Udipsamment ) with textural unimodal distribution, and the medium and fine sands being the main fractions (Table 1). Soils are light gray (l0YR 7/2), exhibiting little evidence of pedogenic development , with no organic matter . They are colonized by scarce vegetation (Spartina ciliata, Panicum racemosum), except in those areas undergoing fixing and afforestation . They are excessively drained and highly permeable soils , and they are impacted by bioturbations of both vertebrates and invertebrates (Fig. 3c). They have a higher content of calcium carbonate associated with the presence of abundant shell remains, which, added to the influence of sea spray , generates a moderately alkaline pH (Table 1).

a Panoramic view of UGBI, and plant communities . b Detail of oxyaquic Udipsamments (P2). c Detail of bioturbations in present soils . d Mesoscopic view of apedal microstructure in AC horizon (P2), bar: 100 μm. e SEM image of an oxalate crystal to fungal activity in isolated grain (P1), bar: 20 μm. f Microscopic microstructure (PM) view with bioclasts (white arrow) of AC, P2. bar: 100 μm. g SEM image of filaments and tubes calcified (AC, P2), bar: 100 μm. h SEM image of filaments and tubes associated with fungal hyphae , in AC, P2, bar: 50 μm. i SEM image of oxalate crystals in the fungal hyphae filaments , bar: 5 μm. j SEM image interface in shell/Ca biomineralizations , bar: 100 μm
Light and heavy minerals in the medium and fine sand fractions indicate higher levels of rounded shells , as most of the observed minerals. Due to coastal dynamics (Teruggi 1959), the maximum concentration of heavy minerals was found in the fine sand fraction, where pyroxene , epidote , amphibol , opaque minerals , biotite , garnet and tourmaline prevail. Lighter minerals are characterized by quartz , plagioclase , lithic fragments , volcanic glass and abundant bioclasts (Table 1; Fig. 3f).
The bioclasts or shell fragments are fibrous and rounded, granular and/or partially weathered. At the surface, there are few fibrous calcium biomineralizations , with crystal morphologies (filaments and rods or rollers) of varying sizes, ranging from 0.5 to 10 μm (Fig. 3e, g–j).
Soils in the interdune area (Profile 2) have slight development (oxyaquic Udipsamment ) with AC and C horizons; they are of a yellowish brown color (10YR 5/2), and present abundant shell remains and an important density of Bioturbations by both invertebrates (carabids ) and vertebrates ( Ctenomys talarum ) (Antenuchi and Busch 1992) (Fig. 3b, c). The physico-chemical characteristics are similar to the dune crest profile, but most vegetation cover (S. ciliata, Androtrichum trygynum, Cakile maritima) generates an increase of 0.6% in organic matter content. The reaction is slightly alkaline and carbonate content is highest in the AC pedogenetic horizon (25%) (Table 1). They are sandy soils with a predominance of very fine, fine and medium sand fractions . The textural distribution is irregular, with polymodal design in pedological horizons , showing an increase in the fine silts reaching up to 16% (Table 1).
The light minerals are predominant and are composed of quartz , plagioclase , lithic fragments , volcanic glass and abundant bioclasts (Fig. 3f). Heavy minerals do not exceed 17% in the AC/C horizons , including pyroxene , epidote , amphibol , opaque minerals , biotite and abundant minerals covered by iron oxides (Table 1, Fig. 3d).
The microstructure of the AC horizon is apedal (with some instances of very weak pedality ); the loose grain is slightly connected by low mineralized organic matter , amorphous iron and/or little biogenic oxalates and calcium carbonate . It has a great number and variety of holes where bioturbation structures filled with materials enriched with MOS and/or iron oxides are common (Fig. 3d, f).
The microdispersive analyses (EDXS) with beryllium window (unable to detect C and O) showed that the matrix is predominantly composed of carbon , calcium , sodium and chlorine . In the coating of mineral grains , calcium , iron and sodium predominate, whereas in the coating of bioclasts there is a predominance of calcium oxalates and carbonates , followed by sodium and magnesium .
6 Calcium Biomineralizations
The shells are always present, with maximum values in the C horizon . They are rounded, with a cloudy appearance, and have abundant calcium biomineralizations associated with remnants of shells , forming an intricate network of very small calcified organic filaments (Fig. 3e, g–j). The term “filament ” describes any tubular and/or rod-like organic structure that is not, or is slightly, calcified. The term “calcified filament ” describes a structure where biogenic calcium carbonate replaces the filament and has precipitated on the shells (incrustation ) or inside the shells (impregnation ) (Klappa 1979). International studies on these biomineralizations are numerous (Arnott 1982; Verrecchia et al. 1993, 1995). These calcified filaments , on the bioclastic material of the AC horizon , conform an intricate network of elongated tubes of several tens of microns long and 3–8 µm wide; they are dichotomous, with a central hollow channel of 0.5–4 µm. Solid elongated rods of 20 µm of length and 0.5–1 µm of width are also present (Fig. 3g–j). The shells have a fibrous and porous appearance, and the depressions on their surface are very common, perhaps due to bioerosion processes (Warme 1975; Jahren 1996) (Fig. 3j). There are few variations in the surface elemental composition of the tubes and shells ; calcium predominates, whereas iron is abundant in the wall of the shell but almost unnoticeable in the calcified filament . In some calcified filaments , secondary calcite and some stylets typical of calcium oxalate of fungal origin (Verrecchia and Verrecchia 1994) were observed, in oxyaquic Udipsamment soils predominantly (Fig. 3g–j).
In summary, calcium biomineralizations are scarce in typic Udipsamments (crest of the dunes ), associated mainly with isolated bioclasts with different sand sizes. In the AC/C pedological horizon of the interdune soils , there is a greater number of bioclasts impacted by bioerosion . These dissolution processes are generated by the action of microorganisms (fungi and algae ), which subsequently re-precipitated as secondary oxalates and calcium carbonates (biomineralization ), coating the bioclasts and other mineral grains. Moreover, they weakly add or bind skeletal components , incorporating themselves into the matrix of soils and sediments . In general, they are fibrous and have various crystal morphologies such as filaments , rods and tubes (Fig. 3e, g–j).
7 Geopedological Unit of Coastal Lagoon/Paleo-Coastal Lagoon (UGIB)
The pedosedimentary sequences have developed in the marginal plain of the Mar Chiquita lagoon , from estuarine sediments linked to the last transgressive–regressive cycle of the Holocene (Schnack et al. 1982). The relief is flat and irregular, characterized by a microrelief associated with the presence of marshes , old tidal channels and reworked bioclastic deposits . Whole and partially fragmented individuals of Heleobia sp., T. plebeius , and Mactra sp. are abundant, with 14C ages between 1670 ± 70 and 2820 ± 80 (Schnack et al. 1982; Osterrieth 1998). In the area, grassland “espartillares ” predominate, mostly composed by Spartina densiflora and S. alterniflora (Vervoorst 1967; Isacch 2001). Soils are moderately developed (without the full solum ): mainly Endoaquolls , Natraquolls , Calciaquolls and Hapludolls ; they are conditioned by hydrological variations related to the dynamics of the coastal lagoon .
Profile 3 developed from reworked bioclastic sediments , located between the Mar Chiquita lagoon and the dune barrier , and it is impacted by hydrological changes and episodic sea entries (Figs. 1 and 2).
The vegetation covers the soils completely, and it consists of Cortaderia selloana, P. racemosum, Juncus sp., etc. (Fig. 4a).

a Panoramic view of UGBI, and plant communities in profile 3. b Detail of typic Endoacuoll . c Mesoscopic view of “lublinite ” in ACk Hz, bar: 1000 μm. d Microstructure (PM) view with bioclasts (white arrow) of ACk, bar: 100 μm. e Eroded bioclast. f–n Different habits , according morphologies described by Verrecchia and Verrecchia (1994). f–g Monocrystals M, bar: 2 μm. g–h M and MA, paired rods , bar: 20 μm. i–j MA serrated plates, MAB: intermediate forms from the smooth coupled rods , bar: 2 μm. j–k General view of M, MAB, bar: 10 μm. l–m General view of M, MAB MB3 and Polycrystalline chains P2, l: bar 10 μm. n bar 20 μm
The soils have epipedons of a length of more than 20 cm, composed of surface horizons A-AC. Bioclasts are abundant and increase toward the base of the profile. The colors range from very dark grayish brown (10YR 3/1) to pale gray (10Y 7/2). They are imperfectly drained soils with slow permeability due to the presence of secondary calcium carbonate that hardened bioclastic levels. Concretions and iron coatings evidence hydromorphism conditions , which allow us to classify them as typic Endoaquolls (Fig. 4b).
Soil reaction ranges from moderately alkaline in the surface to alkaline at the base of the sequence. The organic matter content is moderate in the A horizon , descending sharply toward the base of the profile. The calcium carbonate content , associated with bioclastic materials, increases toward the lower horizons (Table 1).
The sequence is texturally homogeneous, the fine and very fine sand fractions being dominant. Distribution is mostly unimodal, except in the pedological horizons : this is due to a slight increase in clays and silts.
The lighter minerals dominate the entire sequence, composed of bioclasts (Fig. 4e), feldspar , lithoclasts , volcanic glass , minerals coated by brown to reddish oxides or amorphous , quartz and muscovite . Heavy minerals are composed of pyroxene , opaque minerals , covered minerals , amphibol , epidote , tourmaline , garnet and zircon , in order of importance. In the silt fraction , other amorphous silica biomineralizations , like silicophytoliths and diatoms , predominate (Table 1).
The microstructure of the pedological horizons (A, ACk) is weak pedality ; the grain is connected by mineralized organic matter , amorphous iron , and common biogenic oxalates and calcium carbonate (Fig. 4d). The coarse minerals (or pedological horizon skeleton) consist of fine sand-silty, with a predominance of lighter and heavy minerals abundant shells and remains of shells (Fig. 4d).
8 Calcium Biomineralizations
Although bioclasts are present in all horizons , they are more relevant in the ACk horizons . The bioclastic material is affected by dissolution and re-precipitation of calcium carbonates and oxalates of biogenic origin . The re-precipitation process is relevant in bioclasts and in the matrix between grains and bioclasts , as noted in Profile 2. The matrix is fibrous and very white and could be defined as “lublinite ” (Ivanoff 1906), or as calcite with a particular habit (Stoops 1976) (Fig. 4c). Peds have dense crystalline forms: calcified filaments and tubes , rods , and short and long needles with sharp or straight ends (Fig. 4f–n). All these morphologies have been precisely described in different types of calcretes (Verrecchia and Verrecchia 1994) (Fig. 4f–n).
-
(a)
Monocrystals M: individual microrods of 0.3 µm width and 1 µm length (Fig. 4f–h).
-
(b)
Large monocrystalline rods MA: 0.5–2 µm width and < 100 µm length (Fig. 4f–k).
-
MA 1: paired rods that have a cross section in the shape of an “eight” figure
-
MA 2: three or more pairs of rods that form planes (Fig. 4f–k).
-
MA 3: two pairs of rods that have a cross section in the shape of an X (Fig. 4k).
-
MA 4: two pairs of rods that have a cross section in the shape of a bar bell.
-
-
(c)
Complex forms MB: paired monocrystalline rods with serrated plates along their length. There are three different habits , according to the number of plates and their orientation relative to the rod (Fig. 4i, h, n).
-
MB 1: two plates are situated at the top and bottom of the “8” formed by the crystal in the cross section (Fig. 4j–l).
-
MB 2: two plates are oriented at 90° to those in MB 1.
-
MB 3: four plates form an X in cross section (Fig. 4k–n).
-
MAB: intermediate forms from the smooth coupled rods to rods with serrated edges (Fig. 4i–n).
-
-
(d)
Polycrystalline chains P: P1, P2 and P3 morphologies (Fig. 4l–m).
The MA, MB and MAB habits are most often found and are formed by cementation and precipitation of calcium carbonate caused by biogeochemical processes (Fig. 4c–n), (Verrecchia et al. 1993). Some of them have been defined as raphides and styloids of whewellite (monohydrate) or weddellite (polyhydrate) (Osterrieth et al. 1998; Oyarbide and Osterrieth 2000), typical of pedogenic calcium oxalates mediated by fungal biomineralization (Verrecchia et al. 1993). Thus, in general, all morphologies are associated with fungal hyphae , forming an intricate mesh of needles : filaments , rods and tubes that coated peds , pores and the matrix of bioclastic–matrix interface (Fig. 4h, n).
Profile 4 is representative of a polycyclic pedosedimentary sequence , located in the W-NW sector of the marginal floodplain of the Mar Chiquita lagoon , in a flat relief with a 0.5% slope. Soils have evolved from sediments of an ancient coastal lagoon , linked to the last transgressive–ingressive Holocene cycle. Vegetation cover is complete, composed of S. densiflora, Panicum sp., Distichlis spicata, Sarcocornia sp. and other grasses. Soil drainage and permeability are moderate (Fig. 5a).

a View of UGBI, profile 4, acuic Hapludoll . b Crabs bioturbations . c View of pedal microstructure with chambers bioturbation (white arrow), in A horizon bar: 100 μm. d SEM image of styloids of weddellite crystal associated with the fungal mycelia , bar: 20 μm. e SEM image filaments and bacteria (white arrow), in 2ACk Hz, bar: 2 μm. f SEM image algae and fungus covering on the shells in 3ACgk Hz, bar: 10 μm. g–i Different habits , according morphologies described by Verrecchia and contributors (1993), g, j SEM image of weddellite pseudo-rombohedral crystals bar: 2 μm. h, k SEM image of styloids Ca oxalates surrounding the exterior of the filaments , bar: 5 μm. i, l SEM image of calcite surrounding the hollow fungal filaments , bar: 5 μm
These soils have a moderate development, with a 40-cm-thick solum composed of a mollic epipedon of 25 cm and dark color (10YR 2/1), and a weak endopedion Bw, classified according to the USDA Soil Taxonomy (1996) as acuic Hapludolls . Then, the 2ACk horizon has abundant and well-preserved Heleobia sp. that has lived 1670 ± 70 year BP. This soil is based on a 3Cgk sedimentary level, of gray color (5Y 5/1), with sandy loam, where T. plebeius and M. isabelliana have lived there around 2040 ± 40 year BP and 2820 ± 80 year BP, respectively. The sequence is based on clayey silt sediments of the Santa Clara Formation , with colors ranging from gray to bluish gray and gray light brown (10YR 6/2), partially to fully calcretized in the top and very bioturbated by crabs (Schnack et al. 1982) (Fig. 5b).
The organic matter content is high on the A horizon and moderate on the 2ACgk horizon ; the pH is slightly to moderately alkaline . Carbonates are present as whole or fragmented shells , scattered in the soil mass from Bw horizon, with higher values in the bioclastic levels (Table 1).
The distribution of the different particle sizes is variable. In the superficial horizons , muddy sediment values reach 50%, in addition to fine and very fine sands. In the 2ACk horizon , a discordance is evident due to substantial increase in sand and bioclastic fractions. The distribution is bimodal in the pedological horizons and polymodal in the two lower sedimentary levels (Table 1).
The mineralogy is recurrent with 92–98% of light minerals and 8–2% of heavy ones, and is characterized by the same minerals as Profile 3. In addition, silicobioliths reach values exceeding 15% in the pedological horizons , with a common presence of diatoms and their remains in 3 and 4Cgk horizons .
The microstructure is moderately pedal . The solum , in terms of fine mineral components, the factor B (“birefringence ”) is composed of silt and clay grains, organic matter , amorphous iron ; and calcium oxalates and carbonates with an important presence in sub-superficial horizons (Bw to 4Cgk). Also, present matrix with weak crystic fabric and scare calcitans . The organic components of the basal mass are formed by decomposed Organic materials , mostly monomorphic , forming a homogeneous mass in color and density. There is a high density of interconnected channels and chambers, with irregular morphologies and varied size and layout, which could be attributed to insect activity (Genise and Poire 1998) (Fig. 5c). Many calcium biomineralization coatings were observed around sand and silt grains, associated with biogenic structures , minerals and/or humified organic matter (Fig. 5c–f).
9 Calcium Biomineralizations
In peds , the bioclastic material is affected by dissolution and re-precipitation of calcium oxalates and carbonates through the fungi and algae action (Fig. 5d–l). The re-precipitation processes are more conspicuous than in previously analyzed profiles, because they become part of a granular matrix with white and fibrous irregular areas. These calcium biomineralizations have variable sizes and morphologies: filaments (5 µm width, 60 µm length), rods (0.6–1.2 µm width and 15 µm length, with straight or pointed ends, and very fine needles , with 0.2 µm width and 10 µm length) and some ovoid structures and composed rods that could be calcified bacteria (Fig. 5d–f) (Verrecchia 1990, 1994; Verrecchia and Verrecchia 1994; Verrecchia et al. 1993, 1995, 1996; Wright 1984, 1986). The presence of algae covering the shells of current mollusks has been described (Fig. 5f). This would suggest that, under saturation conditions and adequate biogeochemistry, algae could also generate calcium biomineralizations . In this regard, there are numerous research studies about the influence of mucus or biofilms , rich in algae and Bacteria , involved in the creation of an environment that enables the precipitation of calcite (Coleman 1993; Jones 1995) (Fig. 5e, f, l).
The study of crystals by EDAX confirms the presence of calcium and variable carbon content according to the stage of development of calcium oxalates (primary nuclei in close association with the hyphae and/or more developed crystals : rosettes or druses ) (Osterrieth et al. 1998). All the described morphologies coincide with the findings of Verrecchia et al. (1993); they explained the transformation of weddellite into whewellite by dehydration , in addition to the evolution to more stable forms, such as calcite (Fig. 5g–i). The most stable forms of calcium oxalate are the styloids of weddellite (Fig. 5d). These could lead to more stable forms by processes of dehydration : the raphides and styloids of whewellite . These biomineralizations can be transformed, in turn, into needles and other forms of calcium carbonate by the action of soil microfauna (Verrecchia et al. 1993). It is reported that microorganisms may form calcite crystals directly and/or act as catalysts by releasing metabolic products (Fig. 5d, k) (Loisy et al. 1999).
Rods and filaments are very common and are composed of sharpened bundles associated with the mycelia that are generated by chitinolytic and cellulolytic processes during the degradation of the organic substrate (Fig. 5d, j, k). These processes show a considerable time sequence, from the formation of an alveolar substructure to the subsequent random arrangement through diagenesis (Fig. 5d, e).
The structures are associated with very small calcified , crystalline , ellipsoidal , spherical bacteria . They are isolated and they coat mineral grains ; they are grouped into chains in the micropores of the matrix , or joined by calcified strands (Fig. 5 e) (Perry 1999).
10 Geopedological Unit of Shell Ridges (UGIC)
The pedosedimentary sequence is represented by Profile 5, a moderately developed soil , well drained and with very rapid permeability , which has evolved over the shell ridges . These ridges form a narrow strip, slightly wavy, parallel to Route 11, which runs along the western margins of the Mar Chiquita lagoon (Figs. 1 and 2). Currently, there are only remnants, as the soil has been used in roadwork. The shell levels are constituted by more than 90% of bioclasts ; they have been studied in the east-northeast of the Buenos Aires province by several authors (Cortelezzi 1977; Fidalgo et al. 1973; Violante 1992) (Fig. 6a–c). The morphological , physico-chemical and mineralogical properties of the soil profile show a pedological cycle from bioclastic materials of ancient regressive deposits that form elevations of 1.5 m in the marginal plain . The vegetation cover is dominated by communities of grass affinity and a modern arboreal stratum . These soils evolved from bioclastics , calcium -enriched sediments called typic Calciudolls .
The content of soil organic matter is high in the A horizon and very low in the underlying horizons . There is a high percentage of carbonates , represented by complete shells , remains of shells , and a massive to powdery carbonate with a strong reaction to HCl . The reaction is alkaline at soil surface, increasing progressively toward the base of the profile.
The particle size of these soils is essentially psephitic, bioclastic and sandy. In the mollic epipedon and transitional horizon , the texture (excluding the psephitic fraction), is sandy loam with polymodal distributions . The largest proportion of heavy minerals is found in the very fine sand fraction of the parent material (Ck), decreasing toward the surface.
The light and heavy minerals are very similar to those previously described, with slight intra-profile variations. At the base of the profile, quartz , tourmaline and zircon are more abundant.
The microstructure is moderately pedal , in the mollic epipedon (Ak and ACk horizons ), with crystic fabrics , calcitans , hipo- and neocalcitans and alveolar micro fabrics (Fig. 6). The matrix is enriched in calcium oxalates and carbonates , organic matter and bioclasts (Fig. 6c, e).
11 Calcium Biomineralizations
Peds have white efflorescences of different morphology. In all horizons, there are shells of Heleobia sp., bivalves , and many bioclastic fragments, all under the effect of dissolution and re-precipitation of calcium oxalates and carbonates (Fig. 6c, d).
These biomineralizations conform dense white mats, of varying sizes, which appear from the surface, reaching their maximum expression from 68 cm onwards. They are common in the matrix of aggregates, where they join clasts, minerals, in addition to amorphous materials , bioclasts and aggregates , as well as aggregates with each other (Fig. 6a–e). Others with a tubular and compact appearance form dense networks of calcified filaments distributed superficially, in some cases forming a compact crust. The presence of these white masses filling the pores is common (Fig. 6e, f).
The morphologies found show a series of specific forms and compositions associated with fungal mats that have been defined as:
-
1.
Calcium oxalate biominerals : whewellite (CaC2O4.H2O) and weddellite (CaC2O4.(2+x)H2O). They are the product of fungal activity favored by the presence of an organic horizon of several cm (Graustein et al. 1977; Cromack et al. 1979; Osterrieth et al. 1998). Crystals of weddellite occur in all stages of development as pseudo-rhombohedral crystals or pyramids (Horner et al. 1983; Osterrieth et al. 1998): “primary nuclei ” (1–4 μm), “rosettes ” (2–5 μm) and “druses ” (8–12 μm), a stage of greater complexity (Fig. 6g, h). In all cases, the close relationship between the crystals and the fungal cell wall is evident: the “primary nuclei ” are located within the hyphae (Figs. 3i, 4f, 5j); the “rosettes ” are closely associated with the wall, but in an external position (Fig. 6g), and the “druses ” have little visible relationship to the hyphae (Fig. 6h) (Arnott 1982; Horner et al. 1983; Verrecchia et al. 1993; Osterrieth et al. 1998).
-
2.
Calcium carbonate biominerals : acicular calcite and elongated needle-fiber monocrystalline crystals of habit type M, MA and MB, of 0.5 × 5 μm average sizes, linked to the fungal cell wall and oxalates that cover it (Fig. 6i). The coexistence of calcite and calcium oxalate biominerals in different proportions is recurrent, but with a marked increase in calcite at the base of the profile (Fig. 6f, i, m). This coincides with the findings of Verrecchia et al. (1993), who argue that there is a chronological sequence beginning with the precipitation of polyhydrated calcium oxalates (weddellite ), then transformed into more stable monohydrated calcium oxalates (whewellite ) by dehydration, which subsequently become calcium carbonate by bacterial activity and diagenesis (Franceschi and Horner 1980; Verrecchia et al. 1993; Cromack et al. 1979).
Water availability and high content of organic matter in the upper levels of the soil profile promote microbial activity and the process of biomineralization . The coexistence of both calcium oxalate and calcite , which increases toward the base, indicates a response to moisture changes that occurred in the environment, favored by the macroporosity of the profile and bioclastic calcareous materials . In addition, we have characterized the genera Penicillium, Trichoderma and Absidia as fungi associated with crystals in the Ak and ACk horizons (Osterrieth et al. 1998, Oyarbide and Osterrieth 2000).
Y-shaped branching and the diameter of the filaments and calcified tubes revealed their fungal nature (Fig. 6f, i, k–n). Unbranched tubes , which could be linked to actinomycetes and algae , were also present, but in a lower proportion (Fig. 6f, j–n).
The crystals are composed almost entirely of calcium , whereas the matrix of the aggregates has a calcium -enriched elemental composition with little to moderate silicon content, and traces of aluminum , sodium chlorine , iron and potassium .
The elemental composition of the crystals in mollic horizons , analyzed without the Be window permitted measurement of light elements (C and O ) in addition to Ca , confirms the presence of calcium and varying carbon contents according to their developmental stage: primary nuclei , rosettes or druses of calcium oxalates (Fig. 6j, a, g, h); and acicular and rombohedral calcite (Fig. 6j, b, i, k–m) (Osterrieth et al. 1998). The detailed characterization of morphologies and their mineralo-chemical study indicate that fungal biomineralization processes , even when they coexist in the same horizons , may have different origins. Filaments and tubes probably associated with species that develop surface crystals , and others that may have formed within ectomycorrhitic pods and then released by decomposition , were differentiated. Some of these needle bundles retain their parallel arrangement to a main longitudinal axis and are alveolar microfactories (Fig. 6e–m). In addition, in these mollic epipedons , the presence of pelletiform calcified structures immersed in calcified mats of filaments and tubes has been recurrent; their shapes and sizes could be associated with the action of fungivore mesofaunan ) (Fig. 6n).

a Panoramic view shell (UGIC). b Detail of typic Calciudolls profile 5. c Microstructure (PM) view calcic matrix with bioclast , in Ak Hz, bar: 1000 μm. d SEM image of Heleobia sp. with marks bioerosion of Ak Hz, bar: 500 μm. e SEM image of microstructure of calcified filaments , on the bioclastic and clastic material of the Ak horizon , bar: 2 μm. f SEM image of filaments and tubes in the ACk matrix bar: 10 μm. g SEM image of “rosettes ” bar: 10 μm. h SEM image of “druses ,” bar: 10 μm. i SEM image of acicular crystals surrounding the exterior of the fungal miscelia . bar: 10 μm. j Spectra of counts registered by the EDXS microprobe of different crystals , filaments , tubes , a calcium oxalates , b calcium carbonates . k SEM image of unbranched calcified tubes in Ak Hz. bar: 10 μm. l SEM image of calcified tubes in matrix (ACk), bar: 20 μm. m SEM image of calcite tubes in matrix of Ck Hz. bar: 4 μm. n SEM image of pelletiform calcified structures (white arrow), immersed in calcified mats of filaments and tubes (Ak Hz), bar: 50 μm
12 Final Considerations and Conclusions
The pedostratigraphic sequences of the current and past coastal plains or marshes have monocyclic and polycyclic soils . These soils are linked to the active coastal morphodynamics , the evolution of the lagoon , and the water changes from the maximum transgressive period , around 7000 year BP to the present. In addition, the calcium biomineralizations associated with bioclastic and silty-sandy deposits have conditioned their pedological evolution .
In these coastal environments , bioclasts and/or their remains, essentially aragonite , are dissolved by the action of microorganisms (fungi , actinomycetes , bacteria and algae ) generating bioerosion . Subsequently, the microflora re-precipitates calcium oxalates and carbonates (biomineralizations ) are incorporated as coatings of clasts and bioclasts , and to the matrix of soils and sediments.
These calcium biomineralizations , present in all the soils studied, become more conspicuous and abundant as one moves from the east to the west of the toposequence . The type and diversity of these biomineralizations increase directly in relation with time and pedogenetic evolution .
Morphologically, biomineralizations are barely visible in soils of dunes and interdunes , but they are evident in the pedosedimentary sequences of the Mar Chiquita lagoon ’s marginal plain and shell ridges , where they reach their maximum expression. They are yellowish-white, pink to white and, in general, they display an earthy and fibrous appearance. Their morphologies are varied: calcified filaments and tubes , rods , short and long needles with sharp or straight ends, among others. Their size also varies from 0.2 to >100 μm.
The microstructure of these sequences ranges from apedal in dune environments to pedal with crystic factories , calcitans , hipo- and neocalcitans and alveolar microfactories . In addition to matrices which are massive or have a filliform appearance, they have the same appearance in faces , pores , chambers and cavities .
The interaction between organic and mineral horizons , rhizosphere fungi–bacteria and present- and paleo-soils , generates gels and biofilms where bacteria which are isolated or joined in chains may calcify and persist in the micro- and mesopores , and in the matrix of the peds . These bacteria , with calcium contents above 85%, are always associated with calcified filaments , rods , etc.
A remarkable variety of dichotomous tubes was found, as clear evidence of fungal origin , in addition to elongated tubes of multiple sizes and diameters, and complex interdigitated crystalline textures of oxalate and calcium carbonates .
The presence of calcified pelletiform structures of various shapes and sizes, present in fibrous and complex matrices, is linked to the activity of fungivore mesofauna , common in mollic epipedons of soils with moderate development.
The chemical composition of calcified structures shows calcium as the main element, in addition to oxygen and Carbon . Mineralo-chemical studies confirm the presence of calcium and varying carbon contents, depending on the stage of development of the crystals : the early stages, in close association with the walls of the hyphae (primary nuclei ), or the more developed stages of crystals , which expand to the environment (rosettes and druses ). Thus, it was possible to define the genetic sequence of calcite via calcium oxalate (weddellite and whewellite ) associated with hyphae , algae , soil bacteria and actinomycetes . Most morphologies match the studies of Verecchia et al. (1995), where the transformation of weddellite into whewellite by dehydration is explained, in addition to the evolution into more stable forms, such as calcite . Based on the elemental composition, a relationship has been established between them, which indicate the loss of carbon and oxygen as calcium oxalate crystals evolve into needles and tubes of calcium carbonates .
Calcium biomineralizations allowed us to characterize and confirm biogenic processes and to define the biological evidence masked by sedimentary and post-pedological processes . Moreover, from the point of view of plant nutrition , they are an important reservoir of calcium in the soil . Given their magnitude, they also play an important role in the calcium biogeochemical cycle , linked to the availability of other fundamental macronutrients for the development of the biota , such as phosphorus (Osterrieth et al. 1998).
While there are still many unanswered questions in this area of multidisciplinary interaction, this research adds important data to the results provided by other evidence and proxies. Calcium biomineralizations are good mineralo-biochemical indicators for a better understanding of both current and past pedological environmental processes.
References
Addadi L, Raz S, Weiner S (2003) Taking advantage of disorder: amorphous calcium carbonate and its roles in biomineralization. Adv Mater 15(12):959–970
Ameghino F (1908) Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal. Anales Museo Nacional Buenos Aires 10(3):343–428
Antinuchi CD, Busch C (1992) Burrow structure in the subterranean rodent Ctenomys talarum. Z. Saugetierkunde 57:163–168
Aragno M, Verrecchia E, Job D, Cailleau G, Braissant O, Khammar N, Ferro K, Mota M, Guggiari M, Martin G (2010) Calcium carbonate biomineralization in ferrallitic, tropical soils through the oxalate-carbonate pathway. Bulletin de la Société Suisse de Pédologie 30:127–130
Arnott HJ (1982) Calcium oxalate (weddellite) crystals in forest litter. Scan Elect Microsc 3:1141–1149
Bortolus A (2001) Marismas en el Atlántico sudoccidental. In: Iribarne O (ed) Reserva de la biosfera Mar Chiquita: características físicas, biológicas y ecológicas, pp 83–92
Brewer R (1964) Fabric and mineral analysis of soils. Wiley, New York, p 470
Bullock P, Fedoroff N, Jongerious A, Stoops G, Tursina T, Babel U (1985) Handbook for soil thin section description. Wayne Research Publications, Wolverhampton, p 150
Burgos JJ, Vidal AL (1951) Los climas de la República Argentina, según la nueva clasificación de Tornthwaite. Meteoros 1(1):3–32
Coe HH, Osterrieth M, Fernández Honaine M (2014) Phytoliths and their applications. In: Gomes CH, Osterrieth M (eds) Synthesis of some phytolith studies in South America. Nova Publishers, New York, pp 1–26
Coleman M (1993) Microbial processes: controls on the shape and composition of carbonate concretions. Marine Geol 113:127–140
Cortelezzi CR (1977) Dataciones de las formaciones marinas del Cuaternario en las proximidades de La Plata-Magdalena, Buenos Aires. LEMIT II 342:77–93
Cromack K, Sollins P, Graunstein W, Spiedel K, Todd A, Spycher C, Todd R (1979) Calcium oxalate accumulation and soils weathering in mats of the hypogeous fungus Histerangium crassum. Soil Biochem 11:463–468
Dawis J, Freitas F (1970) Physiological and chemical methods of soil and water analysis. Soil Bull 10:39–51
Dhami NK, Reddy MS, Mukherjee A (2013) Biomineralization of calcium carbonates and their engineered applications: a review. Front Microbiol 4:314. doi:10.3389/fmicb.2013.00314
Espinosa M (1994) Diatom paleoecology of the Mar Chiquita lagoon delta, Argentina. J Paleolimnol 10:17–23
Fasano J, Hernández M, Isla F, Schnack E (1982) Aspectos evolutivos y ambientales de la Laguna Mar Chiquita (provincia de Buenos Aires, Argentina). Oceanologica Acta, Nti SP, pp 285–292
Fidalgo F, Colado U, De Francesco O (1973) Sobre ingresiones marinas cuaternarias en los partidos de Castelli, Chascomús y Magdalena (provincia de Buenos Aires). In: V Congreso Geologico Argentino, Buenos Aires, vol 3, pp 227–240
Fidalgo F, Riggi J, Gentile R, Correa H, Porro N (1991) Los “sedimentos pampeanos” continentales en el ámbito sur bonaerense. Revista Asociación Geológica Argentina 46(3–4):239–256
Franceschi VR, Horner HT Jr (1980) Calcium oxalate crystals in plants. Bot Rev 46:361–427
Frenguelli J (1935) Diatomeas de la Laguna Mar Chiquita al norte de Mar del Plata (Buenos Aires). Notas Museo de La Plata 1:121–140
Frenguelli J (1950) Rasgos generales de la morfología y la geología de la Provincia de Buenos Aires. LEMIT 2(33):72
Galehouse JS (1971) Sedimentation analysis. In: Carver J (ed) Procedures in sedimentary petrology. Wisconsin, Wiley Interscience, USA, pp 69–94
Genise JF, Poire DG (1998) Icnofósiles: estudio y aplicación. Tercera Reunión Argentina de Icnología and Primera Reunión de Icnología del Mercosur. Cuadernillo de apuntes, p 92
Graustein WC, Cromack K, Sollins P (1977) Calcium oxalate: occurrence in soils and effects on nutrient and geochemical cycles. Science 198:1252–1254
Horner H, Tiffani L, Cody A (1983) Formation of oxalate crystal associated with apothecia of the Discomycete Dasycypha capitata. Mycologia 75(3):423–435
Ibáñez J, de Alba S, García Álvarez A (2000) Una disciplina en crisis: bases para un cambio de paradigma en edafología (el suelo, su clasificación e inventario). XVIII Congreso Argentino de la Ciencia del Suelo, Mar del Plata. CD
Ingram RL (1971) Sieve Analysis. In: Carver J (ed) Procedures in sedimentary petrology. Wisconsin, Wiley Interscience, USA, pp 41–68
Isacch JP (2001) Mapa de vegetación de la Reserva Mar Chiquita. In: Iribarne O (eds) Reserva de la biosfera Mar Chiquita: características físicas, biológicas y ecológicas, pp 79–80
Isla F (1989) Holocene sea-level fluctuations in the southern hemisphere. Quat Sci Rev 8:359–368
Iwanoff LL (1906) Ein Wasserhaltiges Calcium Carbonat Aussen Umgebungen von Nowo-Alexandria (guv. Lublin). Annalen der Geologie und Mineralogte der Russland 8:23–25.
Jahren AH (1996) How and why do phytoliths form? Biomineralization. Phytolitharien Bull 9:2–10
James NP (1972) Holocene and Pleistocene calcareous crust (caliche) profiles: criteria for subaerial exposure. J Sed Petrol 42(4):817–836
Jenny L (1941) Factors of soils formation. McGraw Hill Book Company, New York
Jones B (1995) Processes associated with microbial biofilms in the Twilight zone of caves: example from the Cayman Islands. J Sediment Res 65(3):552–560
Klappa CF (1979) Calcified filaments in Quaternary calcretes: organo-mineral interactions in the subaerial vadose environment. J Sediment Petrol 49(3):955–968
Klappa C (1980) Rhizoliths in terrestrial carbonates: classification, recognition, genesis and significance. Sedimentology 27:613–629
Lapeyrie F, Picatto C, Gerard J, Dexheimer J (1990) TEM study of intracellular and extracellular calcium oxalate accumulation of ectomycorrhizal fungi in pure culture or in association with Eucalyptus seedlings. Symbiosis 9:163–166
Loisy C, Verrecchia E, Dufour P (1999) Microbial origin pedogenic micrite associated with a carbonate paleosol (Champagne, France). Sediment Geol 126:193–204
Lowenstam R (1981) Minerals formed by organisms. Science 211:1126–1131
Mann S (2001) Biomineralization: principles and concepts in bioinorganic materials chemistry. Oxford University Press, New York
Mitsch S, Gosselink J (1993) Wetlands. Van Nostrand Reinhold (eds) New York
Osterrieth ML (1998) Paleosols and their relation to sea level changes during the Late Quaternary in Mar Chiquita, Buenos Aires, Argentina. Quat Int 43–44
Osterrieth ML (2004) Biominerales y Biomineralizaciones. Cristalografía de Suelos. Sociedad Mejicana de Cristalografía (ed), pp 206–218
Osterrieth ML (2005) Biomineralizaciones de hierro y calcio, su rol en procesos biogeoquímicos de secuencias sedimentarias del sudeste bonaerense. XVI Congreso Geológico Argentino 3:255–262
Osterrieth ML, Martínez G (1993) Paleosols on Late Cainozoic Sequences in the Northeastern side of Tandilia Range, Buenos Aires, Argentina. Quat Int 17:57–65
Osterrieth ML, Oyarbide F (1998) Calcium oxalate biominerals generated by fungi in Argiudolls of the Pampean plains, Argentina. A problem? II International Meeting of Phytolith Research 2:43
Osterrieth M, Schnack E (1984) El perfil de Mar de Cobo (partido de Mar Chiquita, provincia de Buenos Aires). Características de sus paleosuelos y posibles correlaciones. Symposium on Oscilaciones del nivel del mar durante el último hemiciclo deglacial en la Argentina, pp 101–117
Osterrieth M, Oyarbide F, Bordas V (1998) Biominerales de oxalato de calcio en suelos de Laguna de Los Padres, Buenos Aires. Revista Argentina Ciencias Suelo 18(1):1–9
Oyarbide RF, Osterrieth M (2000) Presencia y desarrollo de biominerales de calcio de origen fúngico en suelos del sudeste de la provincia de Buenos Aires (Argentina). Congreso Universitario Internacional de Edafología Nicolás Aguilera 1–2:81–88
Oyarbide F, Osterrieth M, Cabello M (2001) Trichoderma koningii as a biomineralizing fungous agent of calcium oxalate crystals in typical Argiudolls of the Los Padres Lake natural reserve (Buenos Aires, Argentina). Microbiol Res 156:113–119
Perry C (1999) Biofilm-related calcification, sediment trapping and constructive micrite envelopes: a criterion for the recognition of ancient grass-bed environments? Sedimentology 46:33–45
Schmidt C (1847) Ueber das Verkommen des oxalsauren Kalks in den einfachsten Zellenpflanzen und dem Secret der Schleimhaute. Ann. Chem. Pharm. (Heidelberg) 61:288–306
Schnack EJ, Gardenal M (1979) Holocene transgressive deposits, Mar Chiquita lagoon coast, Buenos Aires province, Argentina. Proc. Int. Symposium on coastal evolution in the quaternary, pp 419–425
Schnack EJ, Fasano JL, Isla FI (1982) The evolution of Mar Chiquita Lagoon, Province of Buenos Aires, Argentina. In: Colquhom DJ (ed) Holocene sea—level fluctuations: magnitudes and causes. IGCP 61, University of South Carolina, Columbia, pp 143–155
Schnack EJ, Alvarez, J, Cionchi J (1984) El carácter erosivo de la línea de costa entre Mar Chiquita y Miramar, provincia de Buenos Aires. Symposium Oscilaciones del Nivel del Mar durante el último Hemiciclo Deglacial en la Argentina, pp 118–130
Simkiss K, Wilbur K (1989) Biomineralization: cells biology and mineral deposition. Acad. Press Inc., New York, p 327
Simonson RW (1959) Outline of a generalized theory of soil genesis. Soil Sci Soc Am Proc 23:152–156
Soil Survey Staff (1996) Keys to soil taxonomy, 7th edn. Department of Agriculture, USA, p 631
Stoops GJ (1976) On the nature of “lublinite” from Hollanta (Turkey). Am Mineralogist 61:172
Teruggi ME (1959) Las arenas de la costa de la provincia de Buenos Aires entre Cabo San Antonio y Bahía Blanca. LEMIT II, 77, La Plata
Tricart JL (1973) Geomorfología de la pampa deprimida. INTA 12:202
Verrecchia E (1990) Lithodiagenetic implications of the calcium oxalate-carbonate biogeochemical cycle in semi-arid calcretes, Nazareth, Israel. Geomicrobiol J 8:89–101
Tuason MMS, Arocena JM (2009) Calcium oxalate biomineralization by Piloderma fallax in response to various levels of calcium and phosphorus. Appl Environ Microbiol 75(22):7079–7085
Verrecchia E (1994) L’origine biologique et superficielle des croütes zonaires. Bull Soc Geol France 165(6):583–592
Verrecchia E, Verrecchia K (1994) Needle-Fiber calcite: review and classification. J Sediment Res 64(3):650–664
Verrecchia E, Yair A, Ribier J, Kidron G, Rolko-Verrecchia KE (1993) Le role des Cyanobactéries dans la fixation des sols sableux désertiques: un exemple pris dans le désert de Néguev (Nizzana area, Israel). Palynosciences 2:255–266
Verrecchia E, Dumont JL, Verecchia K (1995) Role of calcium oxalate biomineralization by fungi in the formation of calcretes: a case study from Nazareth, Israel. J Sediment Petrol 65:1060–1066
Verrecchia E, Van Grootel G, Guillemet G (1996) Classification of Chitinozoa (Llandoverian, Canada) using image analysis. Microscopy Microanal Microstruct 5–6:461–466
Verrecchia EP, Braissant O, Cailleau G (2006) The oxalate-carbonate pathway in soil carbon storage: the role of fungi and oxalotrophic bacteria. In: Gadd GM (eds) Fungi in biogeochemical cycles. Cambridge University Press, Cambridge, pp 289–310
Vervoorst F (1967) La vegetación de la República Argentina. Vll. Las comunidades vegetales de la depresión del Río Salado, Provincia de Buenos Aires. INTA. Buenos Aires
Violante R (1992) Ambientes sedimentarios asociados a un sistema de barrera litoral del Holoceno en la llanura costera al sur de Villa Gesell, Provincia de Buenos Aires. Revista de la Asociación Geológica Argentina 47(2):201–214
Violante RA, Parker G, Cavallotto JL (2001) Evolución de las llanuras costeras del este bonaerense entre la Bahía Samborombón y la Laguna Mar Chiquita durante el Holoceno. Revista Asociación Geológica Argentina 56(1):51–66
Walkley B (1965) In: Black C (ed) Methods of soil analysis. American Society of Agronomy, pp 1372–1375
Warme J (1975) Boring as trace fossils and the process of marine bioerosion. In: Fray RY (ed) Springer, New York, pp 191–229
Wright VP (1984) The significance of needle-fibre calcite in Lower Carboniferous palaesol. Geol J 19:23–32
Wright VP (1986) Pyrite and the drowning of a paleosol. Geol J 21:139–149
Acknowledgements
This study was financially supported by the Universidad Nacional de Mar del Plata (EXA 741/15), the National Agency for Science and Technology Promotion of Argentina (ANPCyT, BID-PICT N°1583). The authors are especially grateful to Ing. José Vila for their assistance with SEM-EDXS analysis.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Osterrieth, M., Borrelli, N., Frayssinet, C., Frayssinet, L., Cresta, J. (2017). Calcium Biomineralizations Associated with Bioclastic Deposits in Coastal Pedostratigraphic Sequences of the Southeastern Pampean Plain, Argentina. In: Rabassa, J. (eds) Advances in Geomorphology and Quaternary Studies in Argentina. Springer Earth System Sciences. Springer, Cham. https://doi.org/10.1007/978-3-319-54371-0_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-54371-0_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-54370-3
Online ISBN: 978-3-319-54371-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)