
Abstract
Risks of human psychiatric conditions have evolved, and their symptoms represent perturbations to adaptive cognitive and affective systems. Evolutionary considerations are useful in this context because they direct us to the identification of specific human adaptations that become dysregulated in disease, though either underdevelopment or overdevelopment. Autism is thus conceptualized in terms of underdeveloped social cognition, such that the highly elaborated human social brain does not complete its usual developmental trajectory. Psychotic affective conditions, mainly schizophrenia, bipolar disorder, and depression, are, in contrast to autism, conceptualized in terms of dysfunctionally overdeveloped aspects of social cognition, such that they are caused by opposite neural system alterations to those producing autism. The hypothesis that autism and psychotic affective conditions represent diametric disorders is supported by a wide range of convergent evidence from genetics, development, neuroscience, psychology, and cognitive science. The diametric model provides for reciprocal illumination of the causes of these conditions and makes specific recommendations for research strategies and the development of novel treatments.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
FormalPara Lay SummaryThe mental and behavioural traits that have evolved in humans, such as complex sociality, make us vulnerable to corresponding mental disorders, such as disorders that involve too little, or too much, social thinking. Autism can be considered as a disorder where complex sociality does not develop, while schizophrenia, bipolar disorder, and depression can be considered as the opposite: pathologically overdeveloped social thought and behaviour, as seen, for example, in paranoia and hearing voices. Evolutionary biology is fundamentally important in understanding, defining, and treating mental disorders because it helps us to determine what the brain has evolved to do, which informs us about the different ways that brain functions can become dysregulated in disease.
1 Introduction
1.1 The Standard Medical Model and the Reification of Psychiatric Disorders
The standard medical model for understanding and treating disease focuses on determining its proximate physiological and developmental causes, in terms of how functional systems have become dysregulated [1]. High blood glucose levels, for example, may be due to type 1 diabetes, which results from specific, well-characterized physiological and molecular biological causes and, as a result, can be unambiguously diagnosed. Understanding the normal functioning of blood glucose regulation, or any other physiological system, thus represents a key precondition for determining aetiology and effective treatments.
How can the standard medical model be applied to psychiatric disorders? The medical model assumes that illness can be objectively and unequivocally quantified. By contrast, the causes and patterns of brain functions that underlie psychiatric disorders are only dimly understood. Psychiatric disorders are, instead, abstract, heuristic, descriptive constructs that are more or less useful for guiding research, diagnoses, and treatments. The clearest evidence for such artificiality is the Diagnostic and Statistical Manual of Mental Disorders (DSM) criteria for diagnosing psychiatric disorders, which comprise detailed lists of symptoms, some set of which are considered necessary and sufficient to infer the presence of disease.
Despite these considerations, it is commonplace for psychiatric conditions to be reified—that is—considered as real, for research, medical, and societal purposes [2]. Such pragmatic reification can be considered as innocuous, but it is not: it constrains and biases how researchers think about mental disorders and their associated research agendas and leads to misconceptions of psychiatric disorders as objectively defined, purely pathological ‘diseases’ that people ‘have’ comparable in some fundamental way to diseases like diabetes, cancer, or atherosclerosis that can be objectively and physiologically quantified in terms of their causes and effects.
Under current paradigms, determining the ‘causes’ of mental disorders often becomes conflated with characterizing mental pathologies or deficits, at levels from genes, to neurodevelopment and function, to cognitive functions, and to deleterious environments. By contrast, according to the standard medical model, mental disorders should instead be conceptualized and analysed, in terms of what functional mental systems have become dysregulated and what forms such dysregulations take. In this regard, for example, to better understand autism, we must also better understand the development of neurotypical social cognition, and to understand bipolar disorder and depression, we must also understand the adaptive functions of normal, contextual variation in mood.
1.2 The Evolution of Mental Adaptations
Adaptive functions of the human mind and brain, like those of glucose regulation, have, of course, evolved. Most generally, this meaning of ‘adaptive’ means that such systems show, and have for many, many past generations shown, genetically based variation among individuals that has influenced survival and reproduction. Such variation has thus been subject to natural selection, which leads, across generations, to increases in, or maintenance of, the adaptive ‘fit’ or ‘match’ between organismal phenotypes and aspects of their environments. For example, the beaks of Darwin’s finches are ‘fit’ in their sizes and shapes for different food sources. Similarly, specific regions of the human neocortex adaptively function to recognize individual faces (the fusiform gyrus), or to infer the thoughts and intentions of other humans (the medial prefrontal cortex). Specific mental adaptations, like the insulin pathway, are real and quantifiable and the subject of intense interest in disciplines such as cognitive neuroscience.
Natural selection of human physiology and morphology is expected, under basic evolutionary considerations, to have led to the maximization of functional robustness, homoeostatic ability, and efficiency, as well as optimal flexibility under variable circumstances, all in the service of survival and reproduction. But what, then, is natural selection—the driver of adaptation—expected to maximize with regard to human cognition, emotion, and behaviour? We usually think of mental disorders as centrally involving unhappiness of the subject as well as their social circle, which motivates the seeking of help from the medical community. However, natural selection is by no means expected to maximize happiness, simply because increased happiness is by no means a primary means or route to increased survival and reproduction [3]. Instead, natural selection is predicted, by basic theory, to maximize condition-dependent human striving for the goals that have led, across many past generations in relevant environments, to high survival and reproduction, relative to other humans.
In the context of striving, human emotional systems have evolved to motivate and modulate goal-seeking, dynamically across different circumstances. Such motivation is mediated by the human ‘liking’ and ‘wanting’ reward systems, as well as by unhappiness or dissatisfaction with current situations. Human cognitive systems, by contrast, represent sets of evolved mechanisms for information processing, causal thinking, and decision-making that subserve identification of appropriate goals and tactics for reaching them. Both emotional and cognitive systems develop across infancy, childhood, and adolescence, whereby genes, environments, and gene-by-environment interactions mediate neurodevelopment. To understand human psychiatric disorders from an evolutionary perspective, it thus becomes necessary to connect these psychological trajectories and adaptations with their corresponding maladaptations (lacks of fit of phenotypes to the environment), expressed as developmental, emotional, and cognitive dysfunctions that revolve around human striving and cognition. What adaptations, then, are dysregulated in major human mental disorders and how?
Evolutionary biology is useful in medicine for two main reasons: (1) it teaches us how to think about human medically relevant phenotypes and diagnoses, in novel, productive ways, and (2) it indicates specific new data to collect and new approaches for therapies. In this chapter, I focus on the evolutionary biology of psychiatric disorders centrally involving social cognition, affect, and development. I first describe the primary types of causes of mental disorders, from evolutionary medical thinking. Next, I describe autism spectrum disorders and psychotic affective spectrum disorders, in the context of these causes, with reference to recent findings in genetics, neuroscience, and psychology, and in the context of which human-evolved adaptations have been subject to what forms of alteration in each case. Third, I describe and evaluate hypotheses for the relationships of these disorders with one another—relationships that define evolved axes of human development, affect, and cognition that structure variation in adaptive and maladaptive human mental functioning. Finally, I make specific suggestions for research and clinical therapies that follow directly from these considerations.
2 Research Findings
2.1 Evolutionary Causes of Mental Disorders
The evolutionary causes of psychiatric disorders represent the ‘ultimate’ sources of these conditions, which indicate why, given their evolutionary history, humans exhibit particular forms of mental disorders with particular symptoms and severities. Each of the six main causes described below centres on explanations for deviations from mental adaptation and health, in the context of how maladaptations can arise, and be maintained, in populations.
2.1.1 Deleterious Alleles
Mutations generate novel alleles that usually cause reduced genetic function, because the perturbations randomly alter a system that would otherwise develop reasonably well. Highly penetrant mutations, with large effects, are especially likely to be highly deleterious, and considerable evidence attests to important roles for de novo, deleterious mutations, such as copy number variants or changes to highly conserve amino acid residues, in the causes of mental illness (e.g. [4]). Highly deleterious alleles that are associated with relatively severe mental illnesses include monogenic causes of autism or schizophrenia that evolve under mutation–selection balance: rare mutations arise and are selected against because their bearers exhibit greatly reduced reproduction.
Rare, deleterious alleles such as copy number variants have been estimated to account for a small percentage of cases of major mental illness [5]. Most inferred ‘risk alleles’ for mental disorders, such as those identified with genomewide association studies are, however, relatively common (at frequencies above 1 % or 5 %) and have small effects on risk through one dimension of their multifaceted impacts on neurodevelopment, neuronal function, and other systems. The degree to which such alleles can be considered as deleterious to health overall—given all of their effects—remains an open question; for example, neurodegenerative disease risk trades off with cancer risk, such that higher risks in one domain of disease may commonly entail lower risks in another [6]. Presumably, if psychiatric risk alleles were purely deleterious, they would indeed not be common in populations. Risk alleles may also exhibit positive effects, on health and reproduction, when expressed in genetic relatives of individuals with mental illness [7]; these findings indicate that ‘risk’ alleles do not simply confer increased risk of disease, but may, depending on the context, confer benefits as well. Such considerations can help to explain the high heritabilities of psychiatric conditions including autism, bipolar disorder, and schizophrenia, on the order of 50–80 % (e.g. [8]).
2.1.2 Mismatched Environments
Populations and individuals are always adapted to past environments, and if environments change more rapidly than they can be tracked by selection and genetic response to selection, then populations will be maladapted. Human environments have changed radically over the past few hundred years, which is expected to lead to higher risk of psychiatric disorders to the extent that the novel environments include risk factors such as increased social stress and isolation, or toxins such as lead and mercury that degrade neurodevelopment. For example, some of the highest rates of schizophrenia are found among visible-minority (e.g. different skin colour) immigrants, who appear to be subject to relatively severe psychosocial stresses due to their novel, challenging environments [9].
2.1.3 Extremes of Adaptations
Some psychiatric conditions, such as generalized anxiety disorder, or some manifestations of obsessive–compulsive disorder such as excessive hygienic behaviour, clearly represent extremes of normally adaptive behaviour: anxiety functions to modulate arousal and attention under challenging conditions [10], and hygiene reduces risks of infection [11]. This conceptual framework has been generalized to connect normal personality variation along a spectrum to personality disorders and to severe psychiatric disorders, by demonstrating which aspects of personality are amplified, reduced, or otherwise distorted to generate mental dysfunction [12]. This approach has successfully described continua in personality traits from normal to maladaptive extremes, although the adaptive significance, in terms of fitness-related benefits and costs of personality variation among normal individuals, remains largely unstudied. Maladaptive extremes can also be considered more directly in the context of human evolutionary history, in that the evolution of human-specific traits, such as large brain size and language, has generated potential and scope for loss of these specific traits, as in microcephaly and specific language impairment, as well as potential and scope for dysfunctional overdevelopment, as in macrocephaly and the disordered and exaggerated components of speech in schizophrenia [13, 14].
2.1.4 Trade-Offs
Trade-offs have been well characterized for developmental and physiological phenotypes, whereby, for example, increased resource allocation in one domain takes away from another. For neurological and psychological phenotypes, however, conceptual paradigms based on trade-offs have yet to be developed, despite evidence for trade-offs of verbal–social with visual–spatial skills [15], empathic with systemizing (rule-based) interests and abilities [16], neural flexibility with stability [17], as well as trade-offs between neural activation of the internally, self-directed default mode network, and the outwardly focused task-positive network [18]. Cognitive and emotional trade-offs are important because they structure the brain’s functional architecture and generate coincidences of relative strengths with relative deficits; for example, Kravariti et al. [19] found that having closer relatives with schizophrenia was strongly associated with better verbal skills relative to visual–spatial skills. Trade-offs are stronger under resource-related constraints, which may commonly follow from dysfunctional neurodevelopment, and their extremes are expected to characterize some psychiatric conditions. Autism, for example, has been strongly associated with a combination of high systemizing and low empathizing, whereas some combination of dysfunctionally high empathizing and low systemizing appears to characterize some psychotic affective conditions [20], especially borderline personality disorder and depression [21].
2.1.5 Conflicts
Genetically based conflicts, whereby two parties exhibit different optima for some genetically based phenotype, generate risk of maladaptation because one party may more or less ‘lose’ the conflict, resources are wasted on conflictual interactions, and conflict mechanisms generate novel targets for dysregulation and disease [22]. The forms of evolutionary genetic conflict most salient to psychiatric conditions include parent–offspring conflict (e.g. [23]), genomic imprinting conflict [24, 25], and sexual conflict [26]. Dysregulated genomic imprinting, for example, underlies the expression of Prader–Willi syndrome, one of the strongest genetic causes of psychosis [27], and this syndrome represents only an extreme case of such psychiatric effects [13]. Similarly, a recent epidemiological study of over two million individuals demonstrated that unaffected sisters (but not brothers) of individuals with schizophrenia and bipolar disorder exhibit higher fertility than controls, a pattern that is uniquely predicted by a hypothesis of ‘sexually antagonistic’ alleles that impose costs on males but benefit females [7].
2.1.6 Defences Mistaken as Symptoms
This last ‘cause’ of disease is only apparent: some psychiatric symptoms represent conditionally adaptive defences for alleviating problematic conditions, rather than deleterious manifestations of disease. Thus, in the same way that fever represents a conditionally adaptive bodily response to infection, with health benefits that usually outweigh its costs, some psychiatric symptoms can be interpreted as conferring benefits, relative to their absence or reduction. Examples of such phenomena include the following: (a) repetitive behaviour in autism, which serves to dampen excessively high levels of autonomic and sensory arousal [28], (b) dissociation, as a psychological mechanism to reduce deleterious effects of trauma [29], (c) delusion formation in psychosis, as a means to mentally cope with the exaggerated and disordered perceptions of salience (causal meaning) [30], and (d) mild depression (low mood), as a conditionally adaptive response to circumstances that favour disengagement from failing or unreachable goals—which escalates to full depression if useless goal-seeking persists [31]. The danger of conceptualizing defences, like fever, as purely deleterious symptoms is that treating them is expected to make the situation specifically worse unless the underlying cause of the disorder (and defence) is addressed, such as the sensory hypersensitivity in autism, the trauma in dissociation, or the challenging life events and personal motivational structure that underlie liability to low mood and depression.
These six causes of aetiology and symptoms of psychiatric conditions converge in their emphases on determining what evolved genetic, developmental, neural, cognitive and emotional systems are altered, and how they are altered, in psychiatric conditions. These causes also provide our framework for determining how nominal, DSM-designated psychiatric conditions are related to one another in their causes, as independent and separate, partially overlapping, or diametric to one another in the same general way as the development or activity of any biological system or pathway can be altered in two opposite directions.
2.2 Autism Spectrum Conditions
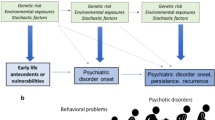
Autism is defined, and commonly reified, as a combination of deficits in social reciprocity and communication with high levels of restricted interests and repetitive behaviour (Fig. 20.1). The degree to which this combination represents a cohesive syndrome, with causally shared rather than independent symptoms and causal factors, remains unclear [32]. Beyond these two commonalities, autism presents diverse features, with overall intellectual abilities varying from very low to above average, cognitive enhancements (above neurotypical) in sensory and visual–spatial abilities in a substantial fraction of individuals and a sex ratio that is highly male-biased overall but much less so among more severely affected individuals [33].

Phenotypes that describe a the autism spectrum and b the psychotic affective spectrum, based on DSM-V diagnoses, evolutionary considerations, and the hypothesized relationships between the two sets of disorders
The most straightforward connection between the major features of autism, and human evolution, is that our evolutionary history has been characterized by elaboration of the ‘social brain’: the distributed, integrated set of neural systems that subserve the acquisition, processing, and use of social information. It is these social brain phenotypes that are specifically underdeveloped in autism. As such, autism can be conceptualized as the expression of maladaptive extremes of social brain underdevelopment, which, in principle, may be caused in a proximate way by reduction or loss of any of the myriad systems that are necessary or sufficient for human social brain development. Autism thus exhibits many single-gene, syndromic causes due to deleterious mutation, but it is also commonly underlain by combined effects from the hundreds or thousands of genes bearing alleles that affect social brain development [34]. As such, there can be no primary, proximate physiologically based cause of autism (as there may be, for example, for type 1 diabetes), and the search for causes becomes a differential characterization, subdivision, and prioritization of the diverse genetic, epigenetic, and environmental influences that converge on underdevelopment of the social brain.
As social cognition is underdeveloped under all psychologically based theories for autism, it can also be conceptualized, and studied, in terms of developmental heterochrony, whereby child cognitive development is not completed in autism, and childhood characteristics, including reduced social cognition, are retained into adulthood [35, 36]. In this context, other human-elaborated traits including highly developed, regulated social striving and goal-seeking, guided by perceived reward-associated or cost-associated (aversive) salience (inferred, causative meaning) of social stimuli, remain underdeveloped as well on the autism spectrum. External stimuli may thus have salience predominantly in terms of perceived sensations, or specific, highly restricted non-social interests, especially foci of highly selective attention [37]. Frith [38] indeed sees a weak drive to discern meaning in the world as epitomizing the weak central coherence theory of autism, which has been supported by a wide range of evidence.
A central, unresolved question in the study of autism is whether a single, central, psychological, or cognitive-level factor can explain the apparently inexplicable combination of reduced sociality with restricted interests and repetitive behaviour. In the context of social brain underdevelopment, increased restricted interests and repetitive behaviours, and sensory, visual–spatial, and mechanistic cognition enhancements in autism, can be explained by several hypotheses.
First, increases in asocial phenotypes may pre-empt the development of social phenotypes, such as by directing perceived salience, interests, and brain specializations along asocial paths. Such effects, which are notably represented by a theory for autism aetiology based on enhanced perceptual functioning [39], may be mediated by overdevelopments of sensory perception and mechanistic, systemizing cognition [33].
Second, increased asocial cognition may itself be a direct result of reduced social cognition, as a compensatory or trade-off-based neurodevelopmental mechanism akin to the overdevelopment of non-visual senses among the blind.
Third, some such asocial cognition and behaviour, and phenotypes such as insistence on sameness and stimulus over-selectivity, may, as noted above, represent defences that aid in coping with challenging symptoms such as increased perceptual sensitivity or avoidance of stress from dealing with inexplicable social cognitive tasks.
Finally, one possible resolution, based on reduced expression of a phenotype virtually unique to humans, is that autism is, in part, underpinned psychologically by underdeveloped imagination, defined as ‘the faculty or action of forming new ideas, or images or concepts of external objects not present to the senses’. This hypothesis, originally described by Rutter [40], and Wing and Gould [41], can, in principle, jointly explain social and asocial alterations in autism, including reduced pretend play, reduced social imagination as expressed in theory of mind, restriction of interests and repetition of behaviour, and insistence on sameness.
Determining the degree to which these hypotheses are correct, in general or for any particular individual, is crucially important to autism therapy, especially to prevent enhancements or conditionally adaptive defences of autistic individuals from being treated as deleterious symptoms.
2.3 Psychotic Affective Spectrum Conditions
Psychotic affective spectrum conditions include a set of DSM disorders, mainly schizophrenia, bipolar disorder, and depression, which broadly overlap in their symptoms, neurological and psychological correlates, and genetic and environmental risk factors [42] (Fig. 20.1). All of these conditions exhibit substantial genetic components and mediation in part by rare, penetrant risk factors, although most genetic risk appears to be underlain by many alleles each with small effect.
Schizophrenia, as well as other conditions that involve psychosis, can be understood most directly and simply in terms of dysfunction of the human adaptive system for assigning salience (causal meaning) to external, and internally generated, stimuli [30, 43]. Salience assignment, which is underpinned by a dedicated neural system involving the anterior cingulate cortex and insula, is fundamental to cognition, behaviour, and goal-seeking, in that it mediates subjective causal understanding of perceptual inputs. Psychosis thus involves overdeveloped and inappropriate salience, usually in the contexts of social interactions, agency, intentionality, self-other associations, and other aspects of mentalistic (social and mind-related) thought, apparently due to the primacy of social cognition in human goal-directed behaviour [24]. Paradigmatic manifestations of psychosis thus involve paranoia, other social delusions, megalomania, belief that events always refer to the self, alterations to self-other distinctions, and assignment of mind, agency, and intentions to inappropriate subjects and inanimate objects. Such reality distortions are mediated by top-down cognitive processes, and they can be considered as attempts to ‘make sense’ of the excessively high and inappropriate salience assignment, for external stimuli, that is driven by hyperdopaminergic neurotransmission [30, 44, 45]. Hallucinations, in turn, can be understood as misinterpreted and exaggerated internal perceptions, mediated by overdeveloped salience of internal representations, such that given certain neurophysiological alterations, thought, inner speech, and imagination come to be considered as external percepts. Like delusions, hallucinations are usually expressed as social phenomena, especially auditory hallucinations.
Schizophrenia is predominantly considered as a disorder of cognition, whereby the causal meanings that guide striving become overdeveloped and dysfunctionally overmentalistic. Bipolar disorder and depression, by contrast, represent mainly disorders of emotion, the set of neural and hormonal systems that motivate and modulate striving and goal-seeking across different contexts. Understanding such mood disorders requires consideration of the adaptive significance of condition-dependent variation in human emotions, especially with regard to the social interactions that permeate human thought and behaviour [46]. In this context, considerable evidence indicates that low mood is normally adaptive in situations where individuals benefit by disengaging from unreachable or unprofitable goals, as it facilitates such disengagement and motivates alternative behavioural patterns of goal-seeking that should be more advantageous [47]. High, positive mood, in comparison, represents an emotional mechanism whereby human reward systems motivate continuation of beneficial behaviour, because one’s goals are being reached. Depression, then, can be conceptualized and studied as overly low and overly stable mood, a maladaptive extreme of an adaptation, whereby individuals fail to disengage from deleterious thought patterns and striving [3, 47]. Conversely, mania represents an emotional opposite to depression, as the expression of inability to emotionally restrain high mood and intensity of striving, even if and when its consequences become detrimental [48, 49]. Behaviours associated with mania and hypomania can, moreover, be directly interpreted in the context of extreme striving for social dominance, power, and influence, which, if successful, leads to substantial benefits [50, 51]. This evolutionary perspective can explain shifts between mania and depression in bipolar disorder, in that mania is expected to foster pursuit of goals that become more and more risky, unreachable or unsuccessful, eventually prompting the generation of mixed states and descent into depression.
In bipolar disorder, then, cognitive salience systems, and choices of goals, commonly remain functional, but the homoeostatic regulation of the emotions that underlie goal pursuit becomes dysregulated, towards overly low or overly high moods and their sequelae. Moreover, like schizophrenia, mania and depression both centrally involve extremes of social, mentalistic thought and behaviour, here in the context of guilt, shame, embarrassment, perceived social defeat, and social rumination in depression, and social dominance pursuit and pride in mania. Affective psychoses, which comprise psychosis with alterations of mood, may thus be mediated by self-punishment-driven, or reward-driven, overattributions of social salience, in the context of emotionality that becomes sufficiently strong to dysregulate salience. These considerations can help to explain well-documented, otherwise-inexplicable associations of bipolar disorder with high social motivation and achievement [49, 52, 53]. Moreover, bipolar disorder, as well as schizophrenia and schizotypy, have been associated across a wide diversity of studies with increased social imagination, divergent thinking, creativity, and goal attainment, especially in the arts and humanities [54–59]. Imagination can indeed be considered, under Bayesian models of cognition and learning, as directly associated with causal cognition and inference of meaning, such that salience, causal thinking, and imagination should tend to increase, or decrease, in concert with one another [60].
2.4 The Relationship Between Autism Spectrum and Psychotic Affective Spectrum Disorders
Bleuler invented the term ‘autism’ to describe withdrawal from reality and social interactions in schizophrenia, but Kanner was careful to point out that his conceptualization of autism referred to children who had never participated in social life [61]. The relationship between autism and schizophrenia, and psychotic affective disorders more generally, has since been considered in terms of two main hypotheses: (1) partial overlap, with some degree of shared social cognitive deficits and genetic risk factors; (2) a diametric (opposite) relationship, based, at a psychological level, on underdevelopment of social cognition and affect in autism, normality at the centre, and dysfunctional forms of their overdevelopment in psychotic affective conditions [62] (Fig. 20.2). The partial overlap hypothesis is data-driven and motivated primarily by the prominence of social deficits especially in autism and schizophrenia. By contrast, the diametric hypothesis follows directly from evolutionary and neurodevelopmental considerations, under the premises that human evolution has been characterized primarily by elaboration of social cognition (generating increased scope for altered development of specific phenotypes) and that the neurodevelopmental systems that underlie it, like all biological systems, can vary and be perturbed in two opposite directions towards lower or higher expression (Fig. 20.2).

The autism spectrum and the psychotic affective spectrum can be conceptualized as diametric disorders, with regard to the direction of alterations in uniquely human or human-elaborated phenotypes that comprise their core features
A central prediction of the diametric hypothesis is that autism and psychotic affective conditions (especially schizophrenia, for which most of the relevant data are available) should exhibit opposite phenotypes and genetic risk factors. A suite of such evidence is described in Table 20.1, which provides support for the diametric hypothesis from diverse and independent sources of data. The partial overlap hypothesis is consistent with the sharing of deficits, especially in social cognition, between autism and schizophrenia, but such deficits can also be considered as deriving from opposite alterations both of which reduce performance on standard tests. Genetic risk factors, such as some genomic copy number variants and some SNPs, have also been associated with both autism spectrum disorders and schizophrenia [62]. Such findings, however, are subject to the caveat that premorbidity to schizophrenia in children and young adolescents, in the form of social deficits and associated developmental problems, can be realistically diagnosed only as autism spectrum since there is not (and never has been) a diagnostic category for schizophrenia premorbidity [63]. This structural limitation in the DSM is expected to lead to a non-negligible incidence of false-positive diagnoses of autism among children who are actually premorbid for schizophrenia, especially among individuals harbouring relatively penetrant genetic risk factors such as copy number variants. Patterns of diagnoses for well-studied CNVs indeed fit with expectations from such false-positive diagnoses [63].
The diametric hypothesis for autism and psychotic affective disorders is novel and controversial and has just begun to be subject to systematic, large-scale testing of its predictions (e.g. [64]). However, to the extent that it is correct, the study of human disorders involving social cognition should be revolutionized and provided its first solid grounding in basic evolutionary principles.
3 Implications for Policy and Practice
Risks for human mental disorders have evolved. Evolutionary conceptualizations of autism and psychotic affective disorders lead directly to novel, specific implications for understanding, studying, and treating these conditions.
First, autism, schizophrenia, bipolar disorder, and depression cannot justifiably be considered as ‘diseases’ under standard medical models of disease, because the neural system adaptations subject to maladaptive alteration in each case remain inadequately understood. Instead, these conditions currently represent broad-scale, heuristic descriptions for suites of related psychological and behavioural problems, none of which has currently specifiable genetic or neurological causes in the same way as do diseases like cancer or diabetes, and all of which grade smoothly in their symptoms into normality. As such, schizophrenia and related psychotic and affective disorders can best be considered as ‘syndromes’: groups of symptom dimensions that cluster in different combinations across different individuals [65]. Risks and symptoms for these psychiatric conditions have, however, evolved in close conjunction with the evolution of complex human social cognition, affect, and behaviour, which provides the basis for an ultimate understanding, and nosology, of psychiatric maladaptations. In this context, DSM descriptions of autism or a psychotic affective condition should represent starting points for differential diagnosis of their genetic, neurological, social, and environmental causes, for each specific individual. Such causes are expected to involve some combination of effects from deleterious mutations, evolutionarily novel environments, extremes of adaptations, trade-offs, genomic conflicts, and evolved defences.
Second, autism can be considered, from an evolutionary perspective, in terms of underdevelopment of social cognition and affect, centrally involving some combination, and causal conjunction, of reduced social development with increased non-social perception, attention, and cognition. Such social and non-social alterations may have diverse proximate causes, but they appear to commonly converge, psychologically, on reductions in imagination, which can explain both lower levels of sociality and increases in restricted interests and repetitive behaviour. This conceptualization of autism is fully compatible with previously developed psychological models founded on reduced central coherence [66], lower empathizing and higher systemizing [33], and enhanced perceptual function [39].
Third, psychotic affective disorders can be considered as centrally involving dysfunctionally overdeveloped social cognition, affect, and behaviour, expressed as social hypersalience in aspects of psychosis, dysregulated social goal motivation and dominance-seeking in mania, and extremes of negative social emotionality in depression. Each of these disorders, which grade into one another, can best be understood in the individual-level contexts of the developmental causes of negatively valenced and imaginative social salience, and the motivational structure of one’s past, current, and future imagined life goals, especially regarding regulation of, and impediments to, success in striving. This framework is fully compatible with current psychological, neurological, cognitive-science-level accounts of psychotic affective conditions (e.g. [30, 43, 49, 51]), but grounds them in evolutionary considerations and in their relationship to the autism spectrum.
Fourth, autism and psychotic affective conditions can be considered and analysed as diametric (opposite) disorders with regard to social development, cognition, affect, and behaviour. This diametric model provides for comprehensive, reciprocal illumination of the diagnoses, causes, and treatments of these disorders, such that insights derived from studying one set of disorders can be applied directly to the other. Most generally, cognitive behavioural treatments for autism should especially focus on enhancing phenotypes that are overdeveloped in psychotic affective conditions, including social imagination, flexible and social salience, and social motivation and goal-seeking. By contrast, treatments for psychotic affective conditions, in addition to focusing more directly on the adaptive, dynamic regulation of social cognitive salience and mood-directed striving, should involve therapies to make perception, cognition, affect, and behaviour relatively ‘more autistic’. Similar considerations apply to pharmacological effects: for example, valproate during foetal development represents a well-established human cause, and animal model, of autism [67], but valproate is also used to treat bipolar disorder and schizophrenia [68]; comparably, mGlur5 pathway antagonists are being used to treat fragile X syndrome and autism [69], whereas mGlur5 agonists are being developed to treat schizophrenia [70].
The findings and inferences described here emphasize that evolutionary approaches in medicine, and psychiatry, can offer specific, well-rationalized hypotheses and can help to direct research and treatments along novel and promising paths. Such progress should lead, eventually, to the integration of psychiatry with the standard medical model of disease, as dovetailing evolutionary and proximate approaches to the study of brain development and function uncover the adaptive significance of psychological, cognitive, and affective phenotypes and their neurological and genetic foundations.
References
Nesse RM, Stein DJ (2012) Towards a genuinely medical model for psychiatric nosology. BMC Med 10:5. doi:10.1186/1741-7015-10-5
Crespi B (2011) One hundred years of insanity: genomic, psychological and evolutionary models of autism in relation to schizophrenia. In: Ritsner M (ed) Handbook of schizophrenia-spectrum disorders, vol I. Springer, Netherlands, pp 163–185
Nesse RM (2004) Natural selection and the elusiveness of happiness. Philos Trans R Soc Lond B Biol Sci 359(1449):1333–1347
Malhotra D, Sebat J (2012) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148(6):1223–1241. doi:10.1016/j.cell.2012.02.039
Escudero I, Johnstone M (2014) Genetics of schizophrenia. Curr Psychiatry Rep 16(11):502. doi:10.1007/s11920-014-0502-8
Plun-Favreau H, Lewis PA, Hardy J et al (2010) Cancer and neurodegeneration: between the devil and the deep blue sea. PLoS Genet 6(12):e1001257. doi:10.1371/journal.pgen.1001257
Power RA, Kyaga S, Uher R et al (2013) Fecundity of patients with schizophrenia, autism, bipolar disorder, depression, anorexia nervosa, or substance abuse vs their unaffected siblings. JAMA Psychiatry 70(1):22–30. doi:10.1001/jamapsychiatry.2013.268
Singh S, Kumar A, Agarwal S et al (2014) Genetic insight of schizophrenia: past and future perspectives. Gene 535(2):97–100. doi:10.1016/j.gene.2013.09.110
Bourque F, van der Ven E, Fusar-Poli P et al (2012) Immigration, social environment and onset of psychotic disorders. Curr Pharm Des 18(4):518–526
Stein DJ (2013) What is a mental disorder? A perspective from cognitive-affective science. Can J Psychiatry 58(12):656–662
Curtis VA (2014) Infection-avoidance behaviour in humans and other animals. Trends Immunol 35(10):457–464. doi:10.1016/j.it.2014.08.006
Trull TJ, Widiger TA (2013) Dimensional models of personality: the five-factor model and the DSM-5. Dialogues Clin Neurosci 15(2):135–146
Crespi B (2008) Language unbound: genomic conflict and psychosis in the origin of modern humans. In: Hughes D, D’Ettorre P (eds) Sociobiology of communication: an interdisciplinary perspective. Oxford Universtiy Press, Oxford, pp 225–248
Crespi B, Leach E (2015) The evolutionary biology of human neurodevelopment: evo-neuro-devo comes of age. In: Boughner J, Rolian C (eds) Evolutionary developmental anthropology. Wiley, New York (in press)
Johnson W, Bouchard TJ Jr (2007) Sex differences in mental abilities: g masks the dimensions on which they lie. Intelligence 35:23–39
Nettle D (2007) Empathizing and systemizing: what are they, and what do they contribute to our understanding of psychological sex differences? Br J Psychol 98:237–255
Liljenström H (2003) Neural stability and flexibility: a computational approach. Neuropsychopharmacology 28(Suppl 1):S64–S73
Jack AI, Dawson AJ, Begany KL et al (2013) fMRI reveals reciprocal inhibition between social and physical cognitive domains. Neuroimage 66C:385–401
Kravariti E, Toulopoulou T, Mapua-Filbey F et al (2006) Intellectual asymmetry and genetic liability in first-degree relatives of probands with schizophrenia. Br J Psychiatry 188:186–187
Brosnan M, Ashwin C, Walker I et al (2010) Can an ‘Extreme Female Brain’ be characterised in terms of psychosis? Pers Indiv Diff 49(7):738–742
Dinsdale N, Crespi BJ (2013) The borderline empathy paradox: evidence and conceptual models for empathic enhancements in borderline personality disorder. J Pers Disord 27(2):172–195
Crespi B, Foster K, Úbeda F (2014) First principles of Hamiltonian medicine. Philos Trans R Soc Lond B Biol Sci 369:20130366
Crespi B (2010) The strategies of the genes: genomic conflicts, attachment theory and development of the social brain. In: Petronis A, Mill J (eds) Brain, behaviour and epigenetics. Springer, Berlin, pp 143–167
Crespi B, Badcock C (2008) Psychosis and autism as diametrical disorders of the social brain. Behav Brain Sci 31(3):241–261; discussion 261–320
Haig D (2014) Coadaptation and conflict, misconception and muddle, in the evolution of genomic imprinting. Heredity (Edinb) 113(2):96–103
Haig D, Ubeda F, Patten MM (2014) Specialists and generalists: the sexual ecology of the genome. Cold Spring Harb Perspect Biol 6(9):a017525. doi:10.1101/cshperspect.a017525, pii:a017525
Soni S, Whittington J, Holland AJ et al (2008) The phenomenology and diagnosis of psychiatric illness in people with Prader-Willi syndrome. Psychol Med 38(10):1505–1514
Hirstein W, Iversen P, Ramachandran VS (2001) Autonomic responses of autistic children to people and objects. Proc Biol Sci 268(1479):1883–1888
Russo DA, Stochl J, Painter M et al (2014) Trauma history characteristics associated with mental states at clinical high risk for psychosis. Psychiatry Res. doi:10.1016/j.psychres.2014.08.028, pii:S0165-1781(14)00725-2
Kapur S (2003) Psychosis as a state of aberrant salience: a framework linking biology, phenomenology, and pharmacology in schizophrenia. Am J Psychiatry 160(1):13–23
Nesse RM, Jackson ED (2011) Evolutionary foundations for psychiatric diagnosis: making DSM-V valid. In: De Block A, Adriaens P (eds) Maladapting minds: philosophy, psychiatry, and evolutionary theory. Oxford University Press, Oxford, pp 167–191
Brunsdon VE, Happé F (2014) Exploring the ‘fractionation’ of autism at the cognitive level. Autism 18(1):17–30. doi:10.1177/1362361313499456
Baron-Cohen S, Lombardo MV, Auyeung B et al (2011) Why are autism spectrum conditions more prevalent in males? PLoS Biol 9(6):e1001081. doi:10.1371/journal.pbio.1001081
Heil KM, Schaaf CP (2013) The genetics of Autism Spectrum Disorders–a guide for clinicians. Curr Psychiatry Rep 15(1):334. doi:10.1007/s11920-012-0334-3
Woodard CR, Van Reet J (2011) Object identification and imagination: an alternative to the meta-representational explanation of autism. J Autism Dev Disord 41(2):213–226
Crespi B (2013) Developmental heterochrony and the evolution of autistic perception, cognition and behaviour. BMC Med 11:119
Ploog BO (2010) Stimulus overselectivity four decades later: a review of the literature and its implications for current research in autism spectrum disorder. J Autism Dev Disord 40(11):1332–1349
Frith U (2012) Why we need cognitive explanations of autism. Q J Exp Psychol (Hove) 65(11):2073–2092. doi:10.1080/17470218.2012.697178
Mottron L, Dawson M, Soulières I et al (2006) Enhanced perceptual functioning in autism: an update, and eight principles of autistic perception. J Autism Dev Disord 36(1):27–43
Rutter M (1972) Childhood schizophrenia reconsidered. J Autism Dev Disord 2:315–337
Wing L, Gould J (1979) Severe impairments of social interaction and associated abnormalities in children: epidemiology and classification. J Autism Dev Disord 9:11–29
Doherty JL, Owen MJ (2014) Genomic insights into the overlap between psychiatric disorders: implications for research and clinical practice. Genome Med 6(4):29. doi:10.1186/gm546
Winton-Brown TT, Fusar-Poli P, Ungless MA et al (2014) Dopaminergic basis of salience dysregulation in psychosis. Trends Neurosci 37(2):85–94. doi:10.1016/j.tins.2013.11.003
Cook J, Barbalat G, Blakemore SJ (2012) Top-down modulation of the perception of other people in schizophrenia and autism. Front Hum Neurosci 6:175. doi:10.3389/fnhum.2012.00175
Howes OD, Murray RM (2014) Schizophrenia: an integrated sociodevelopmental-cognitive model. Lancet 383(9929):1677–1687. doi:10.1016/S0140-6736(13)62036-X
Nesse RM, Ellsworth PC (2009) Evolution, emotions, and emotional disorders. Am Psychol 64(2):129–139
Keller MC, Nesse RM (2005) Is low mood an adaptation? Evidence for subtypes with symptoms that match precipitants. J Affect Disord 86(1):27–35
Johnson SL (2005) Mania and dysregulation in goal pursuit: a review. Clin Psychol Rev 25(2):241–262
Johnson SL, Fulford D, Carver CS (2012) The double-edged sword of goal engagement: consequences of goal pursuit in bipolar disorder. Clin Psychol Psychother 19(4):352–362
Johnson SL, Carver CS (2012) The dominance behavioural system and manic temperament: motivation for dominance, self-perceptions of power, and socially dominant behaviours. J Affect Disord 142(1–3):275–282
Johnson SL, Leedom LJ, Muhtadie L (2012) The dominance behavioural system and psychopathology: evidence from self-report, observational, and biological studies. Psychol Bull 138(4):692–743. doi:10.1037/a0027503
Coryell W, Endicott J, Keller M et al (1989) Bipolar affective disorder and high achievement: a familial association. Am J Psychiatry 146(8):983–988
Higier RG, Jimenez AM, Hultman CM et al (2014) Enhanced neurocognitive functioning and positive temperament in twins discordant for bipolar disorder. Am J Psychiatry. doi:10.1176/appi.ajp.2014.13121683
Nettle D (2001) Strong imagination: madness, creativity and human nature. Oxford University Press, Oxford
Nettle D (2006) Schizotypy and mental health amongst poets, visual artists, and mathematicians. J Res Pers 40(6):876–890
Burns JK (2004) An evolutionary theory of schizophrenia: cortical connectivity, metarepresentation, and the social brain. Behav Brain Sci 27(6):831–855; discussion 855–885
Simeonova DI, Chang KD, Strong C et al (2005) Creativity in familial bipolar disorder. J Psychiatr Res 39(6):623–631
Carson SH (2011) Creativity and psychopathology: a shared vulnerability model. Can J Psychiatry 56(3):144–153
Bilder RM, Knudsen KS (2014) Creative cognition and systems biology on the edge of chaos. Front Psychol 5:1104. doi:10.3389/fpsyg.2014.01104
Walker CM, Gopnik A (2013) Causality and imagination. In: Taylor M (ed) The development of imagination. Oxford University Press, New York, pp 342–358
Kanner L (1965) Infantile autism and the schizophrenias. Behav Sci 10(4):412–420
Crespi B, Stead P, Elliot M (2010) Comparative genomics of autism and schizophrenia. Proc Natl Acad Sci USA 107(Suppl 1):1736–1741
Crespi B, Crofts HJ (2012) Association testing of copy number variants in schizophrenia and autism spectrum disorders. J Neurodev Disord 4(1):15. doi:10.1186/1866-1955-4-15
Byars SG, Stearns SC, Boomsma JJ (2014) Opposite risk patterns for autism and schizophrenia are associated with normal variation in birth size: phenotypic support for hypothesized diametric gene-dosage effects. Proc Biol Sci 281(1794):20140604
van Os J (2009) ‘Salience syndrome’ replaces ‘schizophrenia’ in DSM-V and ICD-11: psychiatry’s evidence-based entry into the 21st century? Acta Psychiatr Scand 120(5):363–372. doi:10.1111/j.1600-0447.2009.01456.x
Happé F, Frith U (2006) The weak coherence account: detail-focused cognitive style in autism spectrum disorders. J Autism Dev Disord 36(1):5–25
Markram K, Markram H (2010) The intense world theory—a unifying theory of the neurobiology of autism. Front Hum Neurosci 4:224
Haddad PM, Das A, Ashfaq M et al (2009) A review of valproate in psychiatric practice. Expert Opin Drug Metab Toxicol 5(5):539–551. doi:10.1517/17425250902911455
Lozano R, Hare EB, Hagerman RJ (2014) Modulation of the GABAergic pathway for the treatment of fragile X syndrome. Neuropsychiatr Dis Treat 10:1769–1779
Matosin N, Newell KA (2013) Metabotropic glutamate receptor 5 in the pathology and treatment of schizophrenia. Neurosci Biobehav Rev 37(3):256–268
Rees E, Walters JT, Georgieva L et al (2014) Analysis of copy number variations at 15 schizophrenia-associated loci. Br J Psychiatry 204(2):108–114
Rees E, Kirov G, Sanders A et al (2014) Evidence that duplications of 22q11.2 protect against schizophrenia. Mol Psychiatry 19(1):37–40
Brunetti-Pierri N, Berg JS, Scaglia F et al (2008) Recurrent reciprocal 1q21.1 deletions and duplications associated with microcephaly or macrocephaly and developmental and behavioural abnormalities. Nat Genet 40(12):1466–1471
Qureshi AY, Mueller S, Snyder AZ et al (2014) Opposing brain differences in 16p11.2 deletion and duplication carriers. J Neurosci 34(34):11199–11211. doi:10.1523/JNEUROSCI.1366-14.2014
Chaste P, Sanders SJ, Mohan KN et al (2014) Modest impact on risk for autism spectrum disorder of rare copy number variants at 15q11.2, specifically breakpoints 1 to 2. Autism Res 7(3):355–362. doi:10.1002/aur.1378
Pathania M, Davenport EC, Muir J et al (2014) The autism and schizophrenia associated gene CYFIP1 is critical for the maintenance of dendritic complexity and the stabilization of mature spines. Transl Psychiatry 4:e374. doi:10.1038/tp.2014.16
Courchesne E, Mouton PR, Calhoun ME et al (2011) Neuron number and size in prefrontal cortex of children with autism. JAMA 306(18):2001–2010
Baribeau DA, Anagnostou E (2013) A comparison of neuroimaging findings in childhood onset schizophrenia and autism spectrum disorder: a review of the literature. Front Psychiatry 4:175. doi:10.3389/fpsyt.2013.00175
Haijma SV, Van Haren N, Cahn W et al (2013) Brain volumes in schizophrenia: a meta-analysis in over 18,000 subjects. Schizophr Bull 39(5):1129–1138. doi:10.1093/schbul/sbs118
Hobson RP, Bishop M (2003) The pathogenesis of autism: insights from congenital blindness. Philos Trans R Soc Lond B Biol Sci 358(1430):335–344
Ek U, Fernell E, Jacobson L (2005) Cognitive and behavioural characteristics in blind children with bilateral optic nerve hypoplasia. Acta Paediatr 94(10):1421–1426
Landgraf S, Osterheider M (2013) To see or not to see: that is the question. The “Protection-Against-Schizophrenia” (PaSZ) model: evidence from congenital blindness and visuo-cognitive aberrations. Front Psychol 4:352
Silverstein SM, Wang Y, Keane BP (2013) Cognitive and neuroplasticity mechanisms by which congenital or early blindness may confer a protective effect against schizophrenia. Front Psychol 3:624
Brown WA, Cammuso K, Sachs H et al (2003) Autism-related language, personality, and cognition in people with absolute pitch: results of a preliminary study. J Autism Dev Disord 33(2):163–167
Mottron L, Bouvet L, Bonnel A et al (2013) Veridical mapping in the development of exceptional autistic abilities. Neurosci Biobehav Rev 37(2):209–228
Heaton P, Hudry K, Ludlow A et al (2008) Superior discrimination of speech picth and its relationship to verbal ability in autism spectrum disorders. Cogn Neuropsychol 25:771–782
Heaton P, Williams K, Cummins O et al (2008) Autism and pitch processing splinter skills: a group and subgroup analysis. Autism 12(2):203–219
Dohn A, Garza-Villarreal EA, Heaton P et al (2012) Do musicians with perfect pitch have more autism traits than musicians without perfect pitch? An empirical study. PloS One 7(5):e37961
Falter CM, Braeutigam S, Nathan R et al (2013) Enhanced access to early visual processing of perceptual simultaneity in autism spectrum disorders. J Autism Dev Disord 43(8):1857–1866. doi:10.1007/s10803-012-1735-1
Tavassoli T, Miller LJ, Schoen SA et al (2014) Sensory over-responsivity in adults with autism spectrum conditions. Autism 18(4):428–432
Bates TC (2005) The panmodal sensory imprecision hypothesis of schizophrenia: reduced auditory precision in schizotypy. Pers Indiv Diff 38(2):437–449
Leitman DI, Foxe JJ, Butler PD et al (2005) Sensory contributions to impaired prosodic processing in schizophrenia. Biol Psychiatry 58(1):56–61
Leitman DI, Sehatpour P, Higgins BA et al (2010) Sensory deficits and distributed hierarchical dysfunction in schizophrenia. Am J Psychiatry 167(7):818–827
Force RB, Venables NC, Sponheim SR (2008) An auditory processing abnormality specific to liability for schizophrenia. Schizophr Res 103(1):298–310
Javitt DC (2009) Sensory processing in schizophrenia: neither simple nor intact. Schizophr Bull 35(6):1059–1064
Javitt DC (2009) When doors of perception close: bottom-up models of disrupted cognition in schizophrenia. Annu Rev Clin Psychol 5:249–275
Mason OJ, Brady F (2009) The psychotomimetic effects of short-term sensory deprivation. J Nerv Ment Dis 197(10):783–785. doi:10.1097/NMD.0b013e3181b9760b
Daniel C, Lovatt A, Mason OJ (2014) Psychotic-like experiences and their cognitive appraisal under short-term sensory deprivation. Front Psychiatry 5:106. doi:10.3389/fpsyt.2014.00106
Kohl S, Wolters C, Gruendler TO et al (2014) Prepulse inhibition of the acoustic startle reflex in high functioning autism. PLoS One 9(3):e92372. doi:10.1371/journal.pone.0092372
Madsen GF, Bilenberg N, Cantio C et al (2014) Increased prepulse inhibition and sensitization of the startle reflex in autistic children. Autism Res 7(1):94–103. doi:10.1002/aur.1337
Swerdlow NR, Light GA, Sprock J et al (2014) Deficient prepulse inhibition in schizophrenia detected by the multi-site COGS. Schizophr Res 152(2–3):503–512
Orekhova EV, Stroganova TA (2014) Arousal and attention re-orienting in autism spectrum disorders: evidence from auditory event-related potentials. Front Hum Neurosci 8:34
Nagai T, Tada M, Kirihara K et al (2013) Mismatch negativity as a “translatable” brain marker toward early intervention for psychosis: a review. Front Psychiatry 4:115
Todd J, Harms L, Schall U et al (2013) Mismatch negativity: translating the potential. Front Psychiatry 4:171
Oberman LM, Hubbard EM, McCleery JP et al (2005) EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Brain Res Cogn Brain Res 24(2):190–198
Kana RK, Wadsworth HM, Travers BG (2011) A systems level analysis of the mirror neuron hypothesis and imitation impairments in autism spectrum disorders. Neurosci Biobehav Rev 35(3):894–902. doi:10.1016/j.neubiorev.2010.10.007
McCormick LM, Brumm MC, Beadle JN et al (2012) Mirror neuron function, psychosis, and empathy in schizophrenia. Psychiatry Res 201(3):233–239. doi:10.1016/j.pscychresns.2012.01.004
Mehta UM, Thirthalli J, Basavaraju R et al (2014) Reduced mirror neuron activity in schizophrenia and its association with theory of mind deficits: evidence from a transcranial magnetic stimulation study. Schizophr Bull 40(5):1083–1094. doi:10.1093/schbul/sbt155
Kennedy DP, Redcay E, Courchesne E (2006) Failing to deactivate: resting functional abnormalities in autism. Proc Natl Acad Sci USA 103:8275–8280
Buckner RL, Andrews-Hanna JR, Schacter DL (2008) The brain’s default network: anatomy, function, and relevance to disease. In: Kingstone A, Miller MB (eds) The year in cognitive neuroscience. Ann NY Acad Sci, New York, pp 1–38. doi:10.1196/annals.1440.011
Iacoboni M (2006) Failure to deactivate in autism: the coconstitution of self and other. Trends Cogn Sci 10:431–433
Kennedy DP, Courchesne E (2008) Functional abnormalities of the default network during self- and other-reflection in autism. Soc Cogn Affect Neurosci 3(2):177–190
Landin-Romero R, McKenna PJ, Salgado-Pineda P et al (2014) Failure of deactivation in the default mode network: a trait marker for schizophrenia? Psychol Med 21:1–11
Immordino-Yang MH, Christodoulou JA, Singh V (2012) Rest is not idleness implications of the brain’s default mode for human development and education. Perspect Psychol Sci 7(4):352–364
von dem Hagen EA, Stoyanova RS, Baron-Cohen S et al (2013) Reduced functional connectivity within and between ‘social’ resting state networks in autism spectrum conditions. Soc Cogn Affect Neurosci 8(6):694–701
Jung M, Kosaka H, Saito DN et al (2014) Default mode network in young male adults with autism spectrum disorder: relationship with autism spectrum traits. Mol Autism 5:35
Whitfield-Gabrieli S, Thermenos HW, Milanovic S et al (2009) Hyperactivity and hyperconnectivity of the default network in schizophrenia and in first-degree relatives of persons with schizophrenia. Proc Natl Acad Sci USA 106(4):1279–1284
Tang J, Liao Y, Song M et al (2013) Aberrant default mode functional connectivity in early onset schizophrenia. PLoS One 8(7):e71061. doi:10.1371/journal.pone.0071061
Li M, Deng W, He Z, Wang Q, Huang C, Jiang L, Gong Q, Ziedonis DM, King JA, Ma X, Zhang N, Li T (2015) A splitting brain: Imbalanced neural networks in schizophrenia. Psychiatry Res 232(2):145–153. doi:10.1016/j.pscychresns.2015.03.001
Broyd SJ, Demanuele C, Debener S et al (2009) Default-mode brain dysfunction in mental disorders: a systematic review. Neurosci Biobehav Rev 33(3):279–296. doi:10.1016/j.neubiorev.2008.09.002
Karbasforoushan H, Woodward ND (2012) Resting-state networks in schizophrenia. Curr Top Med Chem 12(21):2404–2414
Lombardo MV, Chakrabarti B, Bullmore ET et al (2011) Specialization of right temporo-parietal junction for mentalizing and its relation to social impairments in autism. Neuroimage 56(3):1832–1838
Kana RK, Libero LE, Hu CP et al (2014) Functional brain networks and white matter underlying theory-of-mind in autism. Soc Cogn Affect Neurosci 9(1):98–105
Wible CG (2012) Hippocampal temporal-parietal junction interaction in the production of psychotic symptoms: a framework for understanding the schizophrenic syndrome. Front Hum Neurosci 6:180
Chevallier C, Kohls G, Troiani V et al (2012) The social motivation theory of autism. Trends Cogn Sci 16(4):231–239
Baron-Cohen S (2010) Empathizing, systemizing, and the extreme male brain theory of autism. Prog Brain Res 186:167–175. doi:10.1016/B978-0-444-53630-3.00011-7
Harkness KL, Washburn D, Theriault JE et al (2011) Maternal history of depression is associated with enhanced theory of mind in depressed and nondepressed adult women. Psychiatry Res 189(1):91–96
Baez S, Herrera E, Villarin L et al (2013) Contextual social cognition impairments in schizophrenia and bipolar disorder. PLoS One 8(3):e57664
Konstantakopoulos G, Oulis P, Ploumpidis D et al (2014) Self-rated and performance-based empathy in schizophrenia: the impact of cognitive deficits. Soc Neurosci 9(6):590–600
Kasari C, Chamberlain B, Bauminger N (2001) Social emotions and social relationships: can children with autism compensate? In: Burack JA, Charman T, Yirmiya N, Zelazo PR (eds) The development of autism: perspectives from theory and research. Lawrence Erlbaum Associates Publishers, Mahwah NJ USA, pp 309–323
Kim S, Thibodeau R, Jorgensen RS (2011) Shame, guilt, and depressive symptoms: a meta-analytic review. Psychol Bull 137(1):68–96. doi:10.1037/a0021466
Swettenham J, Remington A, Murphy P et al (2014) Seeing the unseen: autism involves reduced susceptibility to inattentional blindness. Neuropsychology 28(4):563–570. doi:10.1037/neu0000042
Hanslmayr S, Backes H, Straub S et al (2012) Enhanced resting-state oscillations in schizophrenia are associated with decreased synchronization during inattentional blindness. Hum Brain Mapp 34(9):2266–2275. doi:10.1002/hbm.22064
Reed P, McCarthy J (2012) Cross-modal attention-switching is impaired in autism spectrum disorders. J Autism Dev Disord 42(6):947–953. doi:10.1007/s10803-011-1324-8
Morris R, Griffiths O, Le Pelley ME et al (2013) Attention to irrelevant cues is related to positive symptoms in schizophrenia. Schizophr Bull 39(3):575–582. doi:10.1093/schbul/sbr192
Granger KT, Prados J, Young AM (2012) Disruption of overshadowing and latent inhibition in high schizotypy individuals. Behav Brain Res 233(1):201–208. doi:10.1016/j.bbr.2012.05.003
Adams NC, Jarrold C (2009) Inhibition and the validity of the Stroop task for children with autism. J Autism Dev Disord 39(8):1112–1121
Westerhausen R, Kompus K, Hugdahl K (2011) Impaired cognitive inhibition in schizophrenia: a meta-analysis of the Stroop interference effect. Schizophr Res 133(1–3):172–181. doi:10.1016/j.schres.2011.08.025
South M, Chamberlain PD, Wigham S, Newton T, Le Couteur A, McConachie H, Gray L, Freeston M, Parr J, Kirwan CB, Rodgers J (2014) Enhanced decision making and risk avoidance in high-functioning autism spectrum disorder. Neuropsychology 28(2):222–228
Adida M, Maurel M, Kaladjian A, Fakra E, Lazerges P, Da Fonseca D, Belzeaux R, Cermolacce M, Azorin JM (2011) Decision-making and schizophrenia. Encephale 37(Suppl 2):S110–S116
Cascio CJ, Foss-Feig JH, Burnette CP et al (2012) The rubber hand illusion in children with autism spectrum disorders: delayed influence of combined tactile and visual input on proprioception. Autism 16(4):406–419. doi:10.1177/1362361311430404
Paton B, Hohwy J, Enticott PG (2012) The rubber hand illusion reveals proprioceptive and sensorimotor differences in autism spectrum disorders. J Autism Dev Disord 42(9):1870–1883
Palmer CJ, Paton B, Hohwy J et al (2013) Movement under uncertainty: the effects of the rubber-hand illusion vary along the nonclinical autism spectrum. Neuropsychologia 51(10):1942–1951
Park S, Nasrallah HA (2014) The varieties of anomalous self experiences in schizophrenia: splitting of the mind at a crossroad. Schizophr Res 152(1):1–4. doi:10.1016/j.schres.2013.11.036
Chance SA (2014) The cortical microstructural basis of lateralized cognition: a review. Front Psychol 5:820
Beversdorf DQ, Smith BW, Crucian GP et al (2000) Increased discrimination of “false memories” in autism spectrum disorder. Proc Natl Acad Sci USA 97(15):8734–8737
Hillier A, Campbell H, Keillor J et al (2007) Decreased false memory for visually presented shapes and symbols among adults on the autism spectrum. J Clin Exp Neuropsychol 29(6):610–616
Corlett PR, Simons JS, Pigott JS et al (2009) Illusions and delusions: relating experimentally-induced false memories to anomalous experiences and ideas. Front Behav Neurosci 3:53. doi:10.3389/neuro.08.053.2009
Kanemoto M, Asai T, Sugimori E et al (2013) External misattribution of internal thoughts and proneness to auditory hallucinations: the effect of emotional valence in the Deese-Roediger-McDermott paradigm. Front Hum Neurosci 7:351. doi:10.3389/fnhum.2013.00351
Grant P, Balser M, Munk AJ et al (2014) A false-positive detection bias as a function of state and trait schizotypy in interaction with intelligence. Front Psychiatry 5:135
Faust M, Kenett YN (2014) Rigidity, chaos and integration: hemispheric interaction and individual differences in metaphor comprehension. Front Hum Neurosci 8:511
Kercood S, Grskovic JA, Banda D et al (2014) Working memory and autism: a review of the literature. Res Autism Spectrum Dis 8(10):1316–1332
Ruthsatz J, Urbach JB (2012) Child prodigy: a novel cognitive profile places elevated general intelligence, exceptional working memory and attention to detail at the root of prodigiousness. Intelligence 40(5):419–426
Lee J, Park S (2005) Working memory impairments in schizophrenia: a meta-analysis. J Abnorm Psychol 114(4):599–611
Silver H, Feldman P, Bilker W et al (2003) Working memory deficit as a core neuropsychological dysfunction in schizophrenia. Am J Psychiatry 160(10):1809–1816
Treffert DA (2009) The savant syndrome: an extraordinary condition. A synopsis: past, present, future. Philos Trans R Soc Lond B Biol Sci 364(1522):1351–1357. doi:10.1098/rstb.2008.0326
Cardoso-Martins C, da Silva JR (2010) Cognitive and language correlates of hyperlexia: evidence from children with autism spectrum disorders. Read Writ 23(2):129–145
Samson F, Mottron L, Soulières I et al (2012) Enhanced visual functioning in autism: an ALE meta-analysis. Hum Brain Mapp 33(7):1553–1581. doi:10.1002/hbm.21307
Revheim N, Butler PD, Schechter I et al (2006) Reading impairment and visual processing deficits in schizophrenia. Schizophr Res 87(1–3):238–245
Revheim N, Corcoran CM, Dias E et al (2014) Reading deficits in schizophrenia and individuals at high clinical risk: relationship to sensory function, course of illness, and psychosocial outcome. Am J Psychiatry 171(9):949–959
Arnott W, Sali L, Copland D (2011) Impaired reading comprehension in schizophrenia: evidence for underlying phonological processing deficits. Psychiatry Res 187(1–2):6–10
Williams EL, Casanova MF (2010) Autism and dyslexia: a spectrum of cognitive styles as defined by minicolumnar morphometry. Med Hypotheses 74(1):59–62
Brosnan M, Chapman E, Ashwin C (2014) Adolescents with autism spectrum disorder show a circumspect reasoning bias rather than ‘jumping-to-conclusions’. J Autism Dev Disord 44(3):513–520. doi:10.1007/s10803-013-1897-5
Speechley WJ, Whitman JC, Woodward TS (2010) The contribution of hypersalience to the “jumping to conclusions” bias associated with delusions in schizophrenia. J Psychiatry Neurosci 35(1):7–17
Langdon R, Still M, Connors MH et al (2014) Jumping to delusions in early psychosis. Cogn Neuropsychiatry 19(3):241–256. doi:10.1080/13546805.2013.854198
Pellicano E, Burr D (2012) When the world becomes ‘too real’: a Bayesian explanation of autistic perception. Trends Cogn Sci 16(10):504–510
Lawson RP, Rees G, Friston KJ (2014) An aberrant precision account of autism. Front Hum Neurosci 8:302
Ciaramidaro A, Bölte S, Schlitt S et al (2014) Schizophrenia and autism as contrasting minds: neural evidence for the hypo-hyper-intentionality hypothesis. Schizophr Bull pii:sbu124. [Epub ahead of print]
Backasch B, Straube B, Pyka M et al (2013) Hyperintentionality during automatic perception of naturalistic cooperative behaviour in patients with schizophrenia. Soc Neurosci 8(5):489–504. doi:10.1080/17470919.2013.820666
Moore JW, Pope A (2014) The intentionality bias and schizotypy. Q J Exp Psychol (Hove) 67(11):2218–2224
Bara BG, Ciaramidaro A, Walter H et al (2011) Intentional minds: a philosophical analysis of intention tested through fMRI experiments involving people with schizophrenia, people with autism, and healthy individuals. Front Hum Neurosci 5:7
Blijd-Hoogewys EM, van Geert PL, Serra M et al (2008) Measuring theory of mind in children. Psychometric properties of the ToM Storybooks. J Autism Dev Disord 38(10):1907–1930
Clemmensen L, van Os J, Skovgaard AM et al (2014) Hyper-theory-of-mind in children with psychotic experiences. PLoS One 9(11):e113082
Dziobek I, Fleck S, Kalbe E et al (2006) Introducing MASC: a movie for the assessment of social cognition. J Autism Dev Disord 36(5):623–636
Lahera G, Boada L, Pousa E et al (2014) Movie for the assessment of social cognition (MASC): Spanish validation. J Autism Dev Disord 44(8):1886–1896. doi:10.1007/s10803-014-2061-6
Montag C, Dziobek I, Richter IS et al (2011) Different aspects of theory of mind in paranoid schizophrenia: evidence from a video-based assessment. Psychiatry Res 186(2–3):203–209. doi:10.1016/j.psychres.2010.09.006
Fretland RA, Andersson S, Sundet K et al (2015) Theory of mind in schizophrenia: Error types and associations with symptoms. Schizophr Res. doi:10.1016/j.schres.2015.01.024, pii: S0920-9964(15)00028-6
Sharp C, Pane H, Ha C et al (2011) Theory of mind and emotion regulation difficulties in adolescents with borderline traits. J Am Acad Child Adolesc Psychiatry 50(6):563–573. doi:10.1016/j.jaac.2011.01.017
Bird G, Catmur C, Silani G et al (2006) Attention does not modulate neural responses to social stimuli in autism spectrum disorders. Neuroimage 31(4):1614–1624
Sasson NJ, Touchstone EW (2014) Visual attention to competing social and object images by preschool children with autism spectrum disorder. J Autism Dev Disord 44(3):584–592. doi:10.1007/s10803-013-1910-z
Blake R, Turner LM, Smoski MJ et al (2003) Visual recognition of biological motion is impaired in children with autism. Psychol Sci 14(2):151–157
Kim J, Park S, Blake R (2011) Perception of biological motion in schizophrenia and healthy individuals: a behavioral and FMRI study. PLoS One 6(5):e19971. doi:10.1371/journal.pone.0019971
Almeida RA, Dickinson JE, Maybery MT et al (2013) Visual search targeting either local or global perceptual processes differs as a function of autistic-like traits in the typically developing population. J Autism Dev Disord 43(6):1272–1286
Kana RK, Liu Y, Williams DL et al (2013) The local, global, and neural aspects of visuospatial processing in autism spectrum disorders. Neuropsychologia 51(14):2995–3003
O’Connor JA, Wiffen BD, Reichenberg A et al (2012) Is deterioration of IQ a feature of first episode psychosis and how can we measure it? Schizophr Res 137(1–3):104–109
Russell-Smith SN, Maybery MT, Bayliss DM (2010) Are the autism and positive schizotypy spectra diametrically opposed in local versus global processing? J Autism Dev Disord 40:968–977
Craig J, Baron-Cohen S (1999) Creativity and imagination in autism and Asperger syndrome. J Autism Dev Disord 29(4):319–326
King D, Dockrell J, Stuart M (2014) Constructing fictional stories: a study of story narratives by children with autistic spectrum disorder. Res Dev Disabil 35(10):2438–2449
Jamison KR (1993) Touched with fire: manic-depressive illness and the artistic temperament. Free Press, New York
Nelson B, Rawlings D (2010) Relating schizotypy and personality to the phenomenology of creativity. Schizophr Bull 36(2):388–399
Claridge G, McDonald A (2009) An investigation into the relationships between convergent and divergent thinking, schizotypy, and autistic traits. Pers Indiv Diff 46(8):794–799
Kaufman JC (ed) (2014) Creativity and mental illness. Cambridge University Press, Cambridge
Hobson JA, Hobson RP, Malik S et al (2013) The relation between social engagement and pretend play in autism. Br J Dev Psychol 31(1):114–127. doi:10.1111/j.2044-835X.2012.02083.x
Jarrold C (2003) A review of research into pretend play in autism. Autism 7(4):379–390
Bonne O, Canetti L, Bachar E et al (1999) Childhood imaginary companionship and mental health in adolescence. Child Psychiatry Hum Dev 29(4):277–286
Gleason TR, Jarudi RN, Cheek JM (2003) Imagination, personality, and imaginary companions. Soc Behav Pers 31(7):721–737
McLewin LA, Muller RT (2006) Attachment and social support in the prediction of psychopathology among young adults with and without a history of physical maltreatment. Child Abuse Negl 30(2):171–191
Fernyhough C, Bland K, Meins E et al (2007) Imaginary companions and young children’s responses to ambiguous auditory stimuli: implications for typical and atypical development. J Child Psychol Psychiatry 48(11):1094–1101
Wheelwright S, Baron-Cohen S (2001) The link between autism and skills such as engineering, maths, physics and computing: a reply to Jarrold and Routh. Autism 5(2):223–227
Spek AA, Velderman E (2013) Examining the relationship between Autism spectrum disorders and technical professions in high functioning adults. Res Autism Spectr Disord 7(5):606–612
Dickerson AS, Pearson DA, Loveland KA et al (2014) Role of parental occupation in autism spectrum disorder diagnosis and severity. Res Autism Spectr Disord 8(9):997–1007
Nettle D, Clegg H (2006) Schizotypy, creativity and mating success in humans. Proc Biol Sci 273(1586):611–615
Rawlings D, Locarnini A (2008) Dimensional schizotypy, autism, and unusual word associations in artists and scientists. J Res Pers 42(2):465–471
Campbell BC, Wang SS-H (2012) Familial linkage between neuropsychiatric disorders and intellectual interests. PLoS One 7(1):e30405
Durkin MS, Maenner MJ, Meaney FJ et al (2010) Socioeconomic inequality in the prevalence of autism spectrum disorder: evidence from a U.S. cross-sectional study. PLoS One 5(7):e11551
Leonard H, Glasson E, Nassar N et al (2011) Autism and intellectual disability are differentially related to sociodemographic background at birth. PLoS One 6(3):e17875
Werner S, Malaspina D, Rabinowitz J (2007) Socioeconomic status at birth is associated with risk of schizophrenia: population-based multilevel study. Schizophr Bull 33(6):1373–1378
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Crespi, B.J. (2016). The Evolutionary Etiologies of Autism Spectrum and Psychotic Affective Spectrum Disorders. In: Alvergne, A., Jenkinson, C., Faurie, C. (eds) Evolutionary Thinking in Medicine. Advances in the Evolutionary Analysis of Human Behaviour. Springer, Cham. https://doi.org/10.1007/978-3-319-29716-3_20
Download citation
DOI: https://doi.org/10.1007/978-3-319-29716-3_20
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-29714-9
Online ISBN: 978-3-319-29716-3
eBook Packages: MedicineMedicine (R0)