
Abstract
Small evergreen trees or shrubs; branchlets opposite, with conspicuous leaf scars, often pubescent when young, sometimes pendulous with aerial roots; plants dioecious. Leaves simple, pinnately veined, opposite, without stipules, stoutly petiolate; blade thick leathery to thick chartaceous, dark green and shiny above, light green below, sometimes variegated, pubescent or glabrous; margin serrate or glandular serrate, rarely entire. Flowers small, unisexual, epigynous, actinomorphic. Both male and female flowers arranged in terminal thyrsoid compound inflorescences; female inflorescences usually smaller than male ones. Male flowers subtended by one small, lanceolate and pubescent bracteole; calyx minute, 4-dentate; petals 4, appearing free, valvate in bud, oblong or ovate, 3–4 mm long, purple red to yellow or greenish, floral disk fleshy, slightly 4-lobed; stamens 4, alternating with petals; filaments conical, short; anthers yellow, dorsifixed, bithecate , rarely unithecate, dehiscing with longitudinal slits; female flowers similar to male ones but pedicel articulate at the base of the ovary and subtended by one or two lanceolate bracteoles; style simple, short and thick; stigma capitate, straight or bent to one side, slightly 2–4-lobed; ovary unicarpellate, cylindric or ovoid, inferior, 1-locular; ovule 1, pendant, anatropous with dorsal raphe, unitegmic, crassinucellate. Fruit a fleshy 1-seeded drupe, cylindric or ovate, red when mature, black when dry, crowned by persistent calyx teeth, style, and stigma. Endosperm copious; embryo minute.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Small evergreen trees or shrubs; branchlets opposite, with conspicuous leaf scars, often pubescent when young, sometimes pendulous with aerial roots; plants dioecious. Leaves simple, pinnately veined, opposite, without stipules, stoutly petiolate; blade thick leathery to thick chartaceous, dark green and shiny above, light green below, sometimes variegated, pubescent or glabrous; margin serrate or glandular serrate, rarely entire. Flowers small, unisexual, epigynous, actinomorphic. Both male and female flowers arranged in terminal thyrsoid compound inflorescences; female inflorescences usually smaller than male ones. Male flowers subtended by one small, lanceolate and pubescent bracteole; calyx minute, 4-dentate; petals 4, appearing free, valvate in bud, oblong or ovate, 3–4 mm long, purple red to yellow or greenish, floral disk fleshy, slightly 4-lobed; stamens 4, alternating with petals; filaments conical, short; anthers yellow, dorsifixed, bithecate , rarely unithecate, dehiscing with longitudinal slits; female flowers similar to male ones but pedicel articulate at the base of the ovary and subtended by one or two lanceolate bracteoles; style simple, short and thick; stigma capitate, straight or bent to one side, slightly 2–4-lobed; ovary unicarpellate, cylindric or ovoid, inferior, 1-locular; ovule 1, pendant, anatropous with dorsal raphe, unitegmic, crassinucellate. Fruit a fleshy 1-seeded drupe, cylindric or ovate, red when mature, black when dry, crowned by persistent calyx teeth, style, and stigma. Endosperm copious; embryo minute.
A monogeneric family consisting of only Aucuba with 8–11 species distributed in eastern Asia (Japan, China, Vietnam, to eastern Himalayas).
Vegetative Morphology. The shoot of Aucuba japonica shows a typical dichasial branching system and a decussate phyllotaxis that is maintained from the vegetative to the reproductive phase (Hara 1980). The sequence of initiation of successive foliar appendages is very precise in its order. A majority of the shoot apices changes into the reproductive phase to form a terminal inflorescence in the mature plants. Some shoot meristems continue their vegetative growth when they are not transformed into the reproductive phase. Aerial roots may develop on branchlets (Konta 1994).
Vegetative Anatomy. Stomata are of the anomocytic type and mainly confined to the lower leaf surface. The wood is diffuse-porous with a tendency to being semi-ring-porous, showing distinct (e.g., A. japonica) or indistinct (A. chinensis) growth rings (Noshiro and Baas 1998). Vessels are mostly solitary or rarely in multiples or clusters of 2 or 3; perforation scalariform with 27–100 bars. In A. chinensis bar number is less variable, ranging from 27 to 35. Intervessel wall pitting uncommon; if present, it is often opposite or rarely scalariform, and without vestures. Average length of vessel elements ranges from 850–1085 μm, and average vessel diameter from 35–39 μm. Helical thickenings are present in both vessels and vascular tracheids. Fibers have minutely bordered pits in their radial walls. Axial parenchyma is present and mostly scanty paratracheal, occasionally diffuse. Rays consist of a mixture of procumbent, square, and upright cells (Adams 1949; Li and Chao 1954; Noshiro and Baas 1998).
Inflorescence and Flower Morphology. Both male and female inflorescences are basically a thyrsoid type of monotelic synflorescence in which the paracladia are dichasia (Hara 1980). Larger numbers of flowers are produced in male inflorescences (Hara 1980). Reidt and Leins (1994) observed the corolla originating from a low ring-wall, the resulting sympetalous part being so small, however, that it is easily overlooked in adult flowers.
Embryology. Embryo sac development usually is of the Polygonum type with some exceptions. In some ovules, an eight-nucleate mature embryo sac is formed by the fusion of two four-nucleate embryo sacs (Sato 1971). Such ovules have 4–5 parietal layers, differing from those containing the typical Polygonum type embryo sac with 6–7 parietal layers (Sato 1971). This unusual type of embryo sac development occurs at a much higher rate in A. japonica var. borealis than in A. japonica var. japonica (Sato 1976). Typically, each ovule contains one mature embryo sac.
Pollen Morphology. Pollen grains are spheroidal or subprolate in shape and 36–64 (polar) × 29–55 (equatorial) μm in size, 3-colporate, and radially symmetrical. Endoapertures are simple-diffuse and without lamellation of the endexine, indistinct, more or less circular or oblong. Exine 2–3.5 μm thick. Sexine thicker than nexine, intectate, psilate. Psila composed of columella and expanded head. There is little variation in pollen morphology among the three species examined (A. chinensis, A. japonica, and A. himalaica; Ferguson 1977; Ferguson and Hideux 1978).
Karyology. Chromosome numbers were reported for three species: A. japonica: 2n = 16, 32; A. chinensis and A. himalaica: 2n = 16. Plants of A. japonica from north and central Japan (var. japonica and var. borealis) are tetraploids, and those from southern Japan are diploids (var. australis). Tetraploid A. japonica is more vigorous than the diploid material (Viinikka 1970; Kurosawa 1971; see Hara 1972).
Reproductive Biology. The flowers are visited by a variety of insects, including small bees, flies, and beetles. Effects of floral display size and flowering phenology on fruit set was investigated in A. japonica by Abe (2001). The results indicated that display size did not affect fruit set; the proportion of flowers setting fruit remained nearly constant independent of the total number of flowers per inflorescence, the total number of inflorescences per plant, and the total number of flowers per plant. However, flowering phenology did affect fruit set. Fruit set was most abundant when male and female flowering was most abundant. Female flowers remained receptive until they received pollen, and fruit set was generally not pollen-limited unless male flowers were scarce (Abe 2001). Asexual reproduction via pendulous branchlets bearing aerial roots was observed in A. japonica (Konta 1994). New plantlets can grow from these branchlets when they have contact with soil.
Phytochemistry. Fruits of A. japonica contain 2.75 % lipids including 0.45 % fatty acids of their dry weight. Of the fatty acids, 26.37 % are C16, 71.02 % C18, and 2.62 % C20. Among the C18 fatty acids, petroselinic acid is dominant (33.78 %). Other C18 fatty acids include stearic acid (2.38 %), oleic acid (26.99 %), linoleic acid (17.98 %), and linolenic acid (24.09 %; Breuer et al. 1987). Iridoids (aucubin) and several flavonols and flavone glycosides were found in Aucuba (Bate-Smith et al. 1975; Bernini et al. 1984; Iwashina et al. 1997). Two anthocyanins were isolated from the fruits of A. japonica (Ishikura 1971). Tannins, procyanidins, and cyanogenic compounds were not detected in the genus (Bate-Smith et al. 1975).
Affinities. The affinities of Aucuba have long been controversial. The genus has commonly been treated either as a member of Cornaceae (e.g., Harms 1898; Wangerin 1910; Cronquist 1981, 1988), or as a monotypic family (e.g., Airy-Shaw 1973; Takhtajan 1980, 1987) allied to Cornaceae. Molecular sequence data suggested that Aucuba is clearly not a close relative of Cornus, but sister of the western American Garrya, and both are closely related to Eucommiaceae (Xiang et al. 1993; Xiang and Soltis 1998; Soltis et al. 2000). Aucuba and Garrya are also similar in phytochemistry (Bate-Smith et al. 1975; Kleiman and Spencer 1982; see Eyde 1988). Both genera contain the decarboxylated iridoid aucubin, petroselinic acid in their fruit oils, and no tannins. Aucuba and Garrya were grouped together as Garryaceae in the APG III classification. The family was placed in Garryales that also include Eucommiaceae (APG III 2009).
Distribution and Habitats. Aucuba, as the only genus of the family, contains 8–11 species distributed in eastern Asia, from Japan to the Himalayas. All species occur in areas south of the Yellow River in China. Most species are confined to China. Only A. japonica extends its distribution to Japan and Korea, and A. himalaica to the Himalayan region. Plants of Aucuba grow in subalpine coniferous forests below 2800 m and in broad-leaved evergreen or broad-leaved evergreen-deciduous mixed forests, favoring habitats with rich soil and abundant moisture (Hu and Soong 1990).
Economic Importance. Some species of Aucuba are used in folk medicine; wood of a few species is used for carving, cigarette pipes, walking canes, etc. Aucuba japonica is commonly cultivated as an ornamental because of its bright red showy fruits in the winter season, and shiny coriaceous and yellow variegated evergreen leaves (Hara 1972; Hu and Soong 1990).
Only one genus:
-
Aucuba Thunb. Fig. 4
Fig. 4. 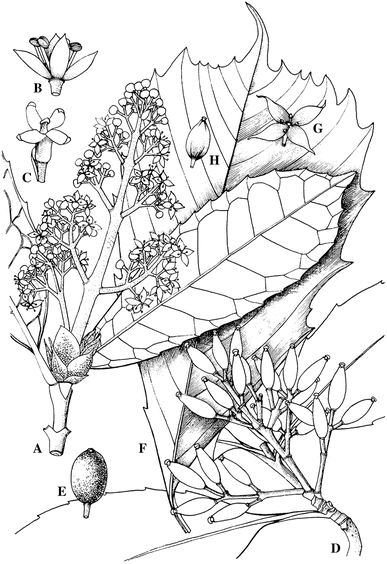
Aucubaceae. A–E Aucuba chinensis var. chinensis. A Male flowering branch. B Male flower. C Female flower. D Fruiting branch. E Fruit. F–H A. obcordata. F Leaf. G Male flower. H Fruit. (from Xiang and Boufford 2006, with permission from Missouri Bot. Garden Press, St. Louis and Science Press, Beijing)
Aucuba Thunb., Nov. Gen. 3: 61 (1783); Fl. Jap. 4, t. 12 & 13 (1784); Xiang, Boufford in Wu and Raven, Flora of China 14: 222–226 (2005).

Description as for family.
Aucuba chinensis Benth., A. himalaica Hook. f. & Thomson, and A. japonica Thunb. are three long-recognized species. Several new species and varieties were added to the genus from the flora of China mainly in the last two decades.
Flowering Jan.–Feb., fruit maturing in the following Feb.
Selected Bibliography
Abe, T. 2001. Flowering phenology, display size, and fruit set in an understory dioecious shrub, Aucuba japonica (Cornaceae). Amer. J. Bot. 88: 455–461.
Adams, J.E. 1949. Studies in the comparative anatomy of the Cornaceae. J. Elisha Mitchell Sci. Soc. 65: 218–244.
Airy-Shaw, H.K. 1973. A dictionary of the flowering plants and ferns, 8th ed. Cambridge: Cambridge Univ. Press.
APG III (Angiosperm Phylogeny Group) 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161: 105–121.
Bate-Smith, E.C., Ferguson, I.K., Hutson, K., Nielsen, B.J., Swain, T. 1975. Phytochemical interrelationships in the Cornaceae. Biochem. Syst. Ecol. 3: 79–89.
Bernini, R.C., Iavarone, C., Trogolo, C. 1984. 1-O-B-D-Glucopyranosyleucommiol, an iridoid glucoside from Aucuba japonica. Phytochemistry 23: 1431–1433.
Breuer, B., Stuhlfauth, T., Fock, H., Huber, H. 1987. Fatty acids of some Cornaceae, Hydrangeaceae, Aquifoliaceae, Hamamelidaceae and Styracaceae. Phytochemistry 26: 1441–1445.
Cronquist, A. 1981. An integrated system of classification of flowering plants. New York: Columbia University Press.
Cronquist, A. 1988. The evolution and classification of flowering plants, 2nd ed. New York: New York Botanical Garden.
Eyde, R.H. 1964. Inferior ovary and generic affinities of Garrya. Amer. J. Bot. 51: 1083–1092.
Eyde, R.H. 1988. Comprehending Cornus: puzzles and progress in the systematics of the dogwoods. Bot. Rev. 54: 233–351.
Ferguson, I.K. 1977. World pollen and spore flora 6: Cornaceae Dum. Stockholm: Almquist and Wiksell.
Ferguson, I.K., Hideux, M.J. 1978. Some aspects of the pollen morphology and its taxonomic significance in Cornaceae sens. lat. Proc. IV Int. Palynol. Conf., Lucknow 1: 240–249 (printed in 1980).
Hara, H. 1972. Corresponding taxa in North America, Japan and the Himalayas. In: Valentine, D.H. (ed.) Taxonomy, phytogeography and evolution. London, New York: Academic Press, pp. 61–72.
Hara, N. 1980. Shoot development of Aucuba japonica I. Morphological study. Bot. Mag. Tokyo 93: 101–116.
Harms, H. 1898. Cornaceae. In: Nat. Pflanzenfam. III, 8. Leipzig: Engelmann, pp. 250–270.
Hu, W. K., Soong, T.P. 1990. Cornaceae. In: Fang, W.P., Hu, W.K. (eds.) Flora reipublicae popularis sinicae 56: 3–108. Beijing: Science Press.
Ishikura, N. 1971. Pelargonidin glycosides in fruits. Experientia 27: 1006.
Iwashina, T.K., Kamenosono, K., Hatta, H. 1997. Flavonoid glycosides from leaves of Aucuba japonica and Helwingia japonica (Cornaceae): Phytochemical relationship with the genus Cornus. J. Japan. Bot. 72: 337–346.
Kleiman, R., Spencer, G.F. 1982. Search for new industrial oils: XVI. Umbelliflorae - Seed oils rich in petroselinic acid. J. Amer. Oil Chemist’s Soc. 59: 29–38.
Konta, F. 1994. Development of aerial roots and asexual propagation in Orixa japonica (Rutaceae) and Aucuba japonica (Cornaceae). Bull. Natn. Sci. Mus. (Tokyo) Ser. B 20(4): 157–161.
Kurosawa, S. 1971. Cytotaxonomical studies on the genus Aucuba. J. Jap. Bot. 46: 231–238.
Li, H.-L., Chao, C.-Y. 1954. Comparative anatomy of the woods of the Cornaceae and allies. Quat. J. Taiwan Mus. 7: 119–136.
Noshiro, S., Baas, P. 1998. Systematic wood anatomy of Cornaceae and allies. IAWA J. 19: 43–97.
Reidt, G., Leins, P. 1994. Das Initialstadium der sympetalen Krone bei Sambucus racemosa L. und Viburnum farreri Stearn. Bot. Jahrb. Syst. 116: 1–9.
Sato, Y. 1971. Embryological study in Aucuba japonica Thunb., with special reference to the unusual development of the embryo sac. Sci. Rep. Tohoku Univ. Ser. IV (Biol.), 35: 201–206.
Sato., Y. 1976. Embryological studies of some cornaceous plants. Sci. Rep. Tohoku Univ. Ser. IV (Biol.) 37: 117–130.
Soltis, D.E., Soltis, P.S., Chase, M.W., Mort, M.E., Albach, D.C., Zanis, M., Savolainen, V., Hahn, W.H., Hoot, S.B., Fay, M.F., Axtell, M., Swensen, S.M., Price, L.M., Kress, W.H., Nixon, K.C., Farris, J.S. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133: 381–461.
Takhtajan, A.L. 1980. Outline of the classification of flowering plants (Magnoliophyta). Bot. Rev. 46: 225–359.
Takhtajan, A.L. 1987. Sistema magnoliofitov. Leningrad: NAUKA.
Viinikka, Y. 1970. A comparative study of mitotic and meiotic chromosomes of Aucuba japonica Thunb. Ann. Bot. Fennici 7: 203–211.
Wangerin, W. 1910. Cornaceae. In: Engler, A. (ed.) Das Pflanzenreich IV, 229, Heft 4: 1–101. Leipzig: W. Engelmann.
Xiang, Q.-Y., Boufford, D.E. 2005 (Text), 2006 (Illustrations). Aucubaceae. In: Wu, Z.Y., Raven, P.H., Hong, D.Y. (eds.) Flora of China, vol. 14. St. Louis: Missouri Bot. Garden Press, Beijing: Science Press, pp. 222–226, fig. 410.
Xiang, Q.-Y., Soltis, D.E. 1998. RbcL sequence data define a cornaceous clade and clarify relationships of Cornaceae sensu lato. In: Boufford, D. E., Ohba, H. (eds.) Sino-Japanese flora - its characteristics and diversification. Univ. Mus. Bull. 37: 123–137.
Xiang, Q.-Y., Soltis, D.E., Morgan, D.R., Soltis, P.S. 1993. Phylogenetic relationships of Cornus L. sensu lato and putative relatives inferred from rbcL sequence data. Ann. Missouri Bot. Gard. 80: 723–734.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Xiang, QY. (2016). Aucubaceae. In: Kadereit, J., Bittrich, V. (eds) Flowering Plants. Eudicots. The Families and Genera of Vascular Plants, vol 14. Springer, Cham. https://doi.org/10.1007/978-3-319-28534-4_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-28534-4_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-28532-0
Online ISBN: 978-3-319-28534-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)