
Abstract
Muscle atrophy is the hallmark of several catabolic conditions. Whatever the cause, the skeletal muscle loss is associated with comorbidities and poor survival. The mass of the skeletal muscle is maintained normally by equilibrium between protein synthesis and breakdown. Rate of synthesis in particular is positively regulated by nutrition and exercise. Anabolic resistance can be defined as a situation where the skeletal muscle is unable to respond appropriately to these anabolic stimuli by stimulating protein synthesis. Anabolic resistance contributes to muscle mass loss in elderly, during immobilization as well as in response to inflammation and cancer. The mechanisms responsible for this blunted response to anabolic stimuli are still under investigation. Several strategies may serve to compensate for anabolic resistance. Optimization of protein intake, resistance exercise, and anti-inflammatory agents appear promising to override this anabolic resistance and mitigate its consequence, the skeletal muscle mass loss.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Background
Muscle atrophy is the hallmark of several catabolic conditions. Whatever the cause, the skeletal muscle loss is associated with comorbidities and poor survival [1].
Muscle proteins are in dynamic equilibrium between their respective rates of synthesis and breakdown. A net gain of muscle mass is only possible if muscle protein synthesis [MPS] exceeds muscle protein breakdown [MPB]. Rate of MPS is positively regulated by nutrient availability and physical activity and negatively regulated by disuse, aging, and muscle wasting-related diseases [2].
The mass of the skeletal muscle is maintained normally by ingestion of protein-containing meals, which results in systemic hyperaminoacidemia. With feeding, MPS is stimulated, and, to a lesser extent, MPB is decreased, causing the protein balance to become positive. During the postprandial period, the balance of muscle protein becomes negative, due to decreased MPS and increased MPB. Therefore, postprandial stimulation of MPS is an important factor in muscle mass maintenance [3].
The stimulation of MPS by protein intake is transient and dose dependent. Indeed, increase in MPS after feeding is of finite duration despite enduring substrate availability. MPS increases after an oral protein bolus with a peak at 2 h before returning to baseline levels, despite elevated plasma amino acid (AA) concentrations [4]. Thus, the muscle becomes refractory to persistent elevations of circulating AA concentrations (“muscle full” concept). The relationship between the dose of protein intake and its resulting stimulation of MPS is also well described. With increasing the dose of protein, MPS increases fast to reach a plateau at approximately 20 g of high-quality protein, with higher-protein intake stimulating AA oxidation rather than MPS. In contrast, MPB decreases slightly but continuously with increasing protein intake. Therefore, protein intake higher than known to stimulate MPS maximally might lead to a greater anabolic response via suppression of MPB [5, 6].
While most AAs are contained in proteins, it appears only to be the essential or indispensable AAs (EAAs), which represent 40–50 % of the total AAs of a high-quality protein, that stimulate MPS. Of the EAAs, the branched chain AAs (BCAAs) are primarily responsible for stimulating MPS [7]. In this regard, leucine is important as it comprises about one-fifth of EAAs needs, and apart from serving as a substrate, it directly activates the signaling pathway stimulating protein synthesis. Leucine on its own appears to stimulate MPS to nearly the same level as a mixture of AAs [8].
Although insulin is of great importance in inhibiting MPB, it plays only a permissive role in stimulating MPS [9]. Although the postprandial rise in circulating insulin might modulate the MPS response through its vasodilatory properties resulting in greater nutritive blood flow to muscle tissue, the basal levels of circulating insulin seem sufficient for maximally stimulating MPS in humans [10]. Systemic administration of insulin alone causes hypoaminoacidemia by inhibiting protein degradation which may in turn inhibit MPS [11]. However, when hypoaminoacidemia is prevented by exogenous AAs replacement, insulin co-administration effectively increases MPS, but this effect is due to the hyperaminoacidemia [12]. The primary effect of insulin on the skeletal muscle is therefore to decrease MPB without a significant effect on MPS [13].
Besides protein intake, resistance exercise is a potent stimulator of MPS and appears to synergistically enhance the gain stimulated by feeding. Even though the rate of MPB increases following exercise, the corresponding increase in MPS is three- to fivefold greater [14]. While the feeding-mediated increase in MPS lasts only few hours at most, the resistance exercise-induced MPS is sustained for about 24–48 h. The synergistic effect of resistance exercise and protein ingestion on muscle protein anabolism is well established. Indeed, the effect of protein intake on muscle protein accretion can further be stimulated by prior exercise training [15], thereby permitting muscle hypertrophy when practiced frequently over time. This effect appears to be mediated by an exercise-induced improvement in nutrient-stimulated vasodilatation and nutrient delivery to muscle rather than potentiation of insulin signaling [16]. Food combined to exercise elevates net muscle protein balance for many hours after exercise [17].
2 Anabolic Resistance Concept
In many situations, MPS in response to feeding or exercise is blunted. This abnormal response of the skeletal muscle to previously well-established anabolic stimuli is known as anabolic resistance [18, 19]. In these situations, the MPS is refractory to EAA provision, irrespective of the availability of insulin. Therefore, anabolic resistance can be defined as a situation where the skeletal muscle is unable to respond appropriately to anabolic stimuli by stimulating protein synthesis and less importantly by inhibiting proteolysis. Indeed, MPB has a minor role in the protein anabolic response to EAAs [20]. The mechanisms responsible for this blunted response of MPS to anabolic stimuli are still under investigation. However, it is highly probable that alterations in protein digestion, AA absorption, hormonal response, microvascular blood flow, AA uptake into the muscle, and intramuscular signaling may play a role.
3 Methodological Considerations
Although the skeletal muscle accounts for 40 % of the body weight and constitutes between 50 and 75 % of all proteins, it represents only 30 % of whole-body protein synthesis. Furthermore, muscle proteins turn over at a rate that is significantly less [about 20-fold] than splanchnic or plasma proteins. Therefore, whole-body protein turnover does not reflect changes which take place into the skeletal muscle [21].
In terms of mechanisms underlying atrophy, the data on rodent and human muscles are difficult to compare for several reasons. Among many, it is worth noting that most of the rodent work has been carried out with young and not adult animals. Moreover, while muscle atrophy in humans mainly involves reduced MPS, increased MPB seems the predominant feature in rodents [22, 23].
Studies focusing on acute changes in MPS cannot predict the net changes in muscle protein balance as opposite changes in MPB may damper the muscle anabolic effect expected based on changes in MPS, especially when long-term changes on the skeletal muscle mass are considered.
Finally, from a functional perspective, changes in muscle function are probably more important than changes in the skeletal muscle mass and should be the most clinically relevant end point to study. This point has not been investigated however in the context of anabolic resistance.
4 Clinical Situations of Anabolic Resistance
4.1 Aging
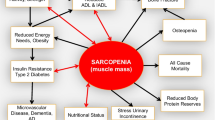
Skeletal muscle mass begins to decline in the fourth or fifth decade of life, at a rate of 0.5–1.5 %/year between 50 and 80 years old. The debilitating effects of muscle loss include declines in physical function and quality of life and increases in morbidity and mortality. Loss of muscle mass with aging is thus a major public health concern. The age-related muscle mass wasting is known as sarcopenia. The loss of muscle mass with aging is probably not due to chronic changes in MPS or MPB but to a blunted feeding-induced rise in MPS [24]. Indeed, muscles of the elderly are resistant to normally robust anabolic stimuli. This anabolic resistance may be a key factor in the development and progression of sarcopenia. Elderly are characterized by a resistance to the three main anabolic stimuli (exercise, AAs, and insulin). Indeed, aging reduces the anabolic response to resistance exercise [25, 26], despite equal circulating and muscle AA concentrations. Furthermore, the stimulation of the muscle anabolic signaling pathways and MPS by AA infusion is significantly blunted in elderly [27, 28]. Finally, aging is associated with reduced inhibition of proteolysis in response to insulin [29]. This anabolic resistance may result from gradual decline in physical activity or to low-grade inflammation (cf. infra).
4.2 Immobilization
During disuse, the skeletal muscle loss occurs at a rate of 0.5 % of muscle mass per day, which translates in 150 g of muscle tissue lost per day [30]. Prolonged disuse (more than 10 days) leads to a decline in basal and postprandial rates of MPS without apparent changes in muscle MPB, except maybe at the early stage (−5 days) [30, 31]. Protein ingestion or AA infusion, even at high rate, increased MPS less in the immobilized leg than in the control one [30, 32]. Therefore, this anabolic resistance can account for the immobilization-induced muscle atrophy. Older adults are more susceptible than young persons to muscle loss after short-term bed rest [23].
4.3 Inflammation
The inflammation associated with sepsis is often responsible for a state of anabolic resistance. In animal models, sepsis blunts the MPS response caused by leucine, exercise, and insulin, the three main anabolic stimuli [33, 34]. The inability of these anabolic stimuli to stimulate muscle protein synthesis during sepsis seems to be related to a defect in signaling to step in translation initiation [34, 35]. This sepsis-induced anabolic resistance in the skeletal muscle results from the cooperative interaction of both cytokines such as TNFα and glucocorticoids [36]. Anabolic resistance in aging may also be related to inflammation, as suggested by the increase in muscle NFkB activity. Indeed, low-grade inflammation as observed in old animals impairs the MPS response to feeding [37].
4.4 Cancer
Cancer cachexia, a metabolic condition caused by tumor burden, is characterized by muscle atrophy and sometimes fat loss. It is observed in 85 % of cancer patients and is implicated in 25 % of deaths. Cancer patients display reduced postprandial MPS compatible with a state of anabolic resistance. Interestingly, tumor resection is able to restore the normal postprandial MPS response [38]. However, a state of anabolic resistance does not seem to be always observed in cancer patients. Several authors showed indeed that cancer patients and controls show comparable protein anabolism during feeding or AAs ingestion, at least at high dose [39–42]. These observations suggest that a window of anabolic potential exists early on in cachexia development [43]. However, in some of these studies, protein anabolism was assessed at the whole-body level and not at the skeletal muscle level.
4.5 Obesity
The MPS in response to AAs appears to be negatively correlated to whole-body fat mass and insulin sensitivity [44]. This suggests that insulin resistance caused by lipid excess due to intracellular accumulation of lipid in muscle or to increased free fatty acid availability may lead to anabolic resistance. In animals, chronic lipid accumulation in muscles as observed in high-fat diet-induced obesity is associated with a concomitant reduction of MPS in response to feeding [45]. Muscle ectopic fat deposition may contribute to anabolic resistance through increase in muscle ceramides. Intramuscular accumulation of ceramides has been previously involved in the insulin resistance for glucose metabolism observed in obesity. Interestingly, ceramides are also able to reduce intracellular AAs availability and protein synthesis in muscle cells [46]. The MPS response to exercise is also blunted in obese animals compared to the lean ones [47]. In humans, lipid infusion to cause reduced whole-body glucose disposal impairs the MPS in response to AAs ingestion [48]. This anabolic resistance to AAs caused by lipid infusion is associated with repression of translation initiation. In obese subjects, the net protein anabolic response to insulin was also blunted compared to healthy subjects [49]. Therefore, excess lipid availability can induce insulin resistance of the skeletal muscle glucose metabolism but also anabolic resistance of AA metabolism. This is not unexpected as many conditions of anabolic resistance such as aging, disuse, and critical illness are characterized by the inability of the skeletal glucose metabolism to respond adequately to insulin.
4.6 Chronic Kidney Disease
In animal models, acute uremia causes a severe resistance to leucine-induced activation of the MPS [50]. Metabolic acidosis by itself impairs leucine-stimulated MPS [51]. Therefore, acidosis may account for the anabolic resistance and ultimately contribute to the muscle wasting which develops in uremia. In human subjects, besides blunting muscle glucose uptake in response to insulin, chronic renal failure and acidosis interfere with the normal suppression of MPB in response to insulin [52]. This state of insulin resistance is also observed in dialysis patients where it is associated with increased MPB [53].
4.7 Other Conditions of Anabolic Resistance
Normobaric hypoxia blunts the increased MPS response to acute resistance exercise [54]. This may contribute to muscle atrophy and blunt the hypertrophic response to resistance exercise in hypoxic conditions.
Glucocorticoids cause the skeletal muscle atrophy by themselves but also by contributing to muscle atrophy observed in many catabolic conditions such as cancer, diabetes, and acute inflammation [55]. Furthermore, glucocorticoids amplify the muscle atrophy caused by immobilization [56].
Most patients who are critically ill combine several causes of anabolic resistance. In addition to the procatabolic hormonal and cytokine milieu, age, immobilization, and hypoxia contribute to a blunted MPS response [18]. Poor delivery of hormones and nutrients caused by compromised microvascular blood flow may also play a role.
5 Cellular Mechanisms
Protein synthesis in the skeletal muscle is regulated through a number of signaling pathways that control the mRNA translation. The protein kinase mTORC1 [mechanistic target of rapamycin complex 1] plays a crucial role in this process by serving as a critical point of integration of most of the anabolic stimuli for the skeletal muscle [57]. Furthermore, blocking mTORC1 activity with the rapamycin drug blocks the contraction and the EAAs-mediated increase in MPS [58, 59].
In response to anabolic stimuli, mTORC1 targets and activates downstream kinases such as S6kinase1 (S6K1) or binding proteins like eIF4E-binding protein1 (4E-BP1). The action of mTORC1 and its downstream mediators heightens the efficiency of ribosomal biogenesis and ultimately translation. Indeed, phosphorylation of 4E-BP1 by mTORC1 prevents its association with eIF4E, allowing eIF4E to bind eIF4G to form the active mRNA cap-binding complex, eIF4F. Thus, mTORC1 stimulates mRNA joining to the 43S preinitiation complex [60].
Growth factors such as insulin and IGF-I activate mTORC1 by stimulating the PI3kinase-Akt/PKB pathway. AAs promote mTORC1 signaling independently of changes in growth factors or mechanic load. The mechanism of activation of mTORC1 by AAs, in particular leucine, involves several molecular actors, in particular a family of GTPases called Rags which act as AA sensors. The lysosomal membrane represents the site at which the AA- and growth factor-sensing machineries converge to stimulate mTORC1. Compared with the regulation of mTORC1 signaling by growth factors and AAs, much less is known about the pathway through which resistance exercise stimulates mTORC1 activity. A specific form of PGC-1α, PGC-1α4, which results from alternative promoter usage and splicing of the primary transcript PGC-1α, is preferentially induced in mouse and human muscle during resistance exercise. This isoform specifically induces IGF-I and represses myostatin [61]. Therefore, PGC1α4 may be a major player in the muscle hypertrophy induced by exercise. However, how this factor stimulates MPS activity is still unknown [62].
REDD1 has been recently identified as an inhibitor of mTORC1. Therefore, REDD1 induction by immobilization [63], glucocorticoids [55], hypoxia, or sepsis [33] may limit MPS by inhibiting mTORC1 signaling and contribute to the anabolic resistance observed in these conditions [64]. AMP-activated kinase (AMPK) functions as a fuel sensor. This enzyme plays an important role in regulating the muscle response to negative energy balance. More specifically, it inhibits mTORC1 signaling when cellular ATP levels are decreased and AMP levels increase in response to limited energy availability. Therefore, insufficient energy availability may limit MPS by inhibiting mTORC1 through activation of AMPK.
Many observations support the role of alterations in the mTOR pathway and its downstream effectors in the muscle resistance to anabolic stimuli.
In elderly, the phosphorylation of mTORC1 and its downstream targets, S6K1 and 4E-BP1, is dampened in the skeletal muscle as compared with the young in response to AAs infusion and exercise [25, 28, 65, 66]. Interestingly, this decline in MPS in response to AAs occurs in association with a reduction of AA transporter content [67].
Interestingly, immobilization which causes a decrease in the global rates of MPS has been associated with a decrease [68] as well an increase in mTOR [69] signaling. In the latter case, the mTOR activation might help to alleviate the immobilization-induced decrease in MPS and muscle mass, as blocking mTOR signaling with rapamycin exacerbates the decline in MPS and muscle mass [69]. Nevertheless, in human immobilization studies, decreases in mTOR have not been observed [10].
The inability of the major anabolic stimuli to stimulate MPS during sepsis seems to be related to a defect in signaling to step in translation initiation, with the redistribution of eIF4E from the active eIF4E.eIF4G complex to the inactive eIF4E.4E-BP1 complex [34, 70], impairing the formation of eIF4F.
Muscle ectopic fat deposition may contribute to anabolic resistance through increase in eIF2α activation [71]. The anabolic resistance to AAs caused by lipid infusion is associated with repression of translation initiation at the level of 4E-BP1 [48].
In animal models, acute uremia causes a severe resistance to leucine-induced activation of the MPS [51] and the mTOR anabolic signaling pathway [50]. Metabolic acidosis impairs leucine-stimulated MPS and activation of the signaling downstream of mTOR at the level of S6K1.
6 Tissular Mechanisms
6.1 Splanchnic Sequestration
About 95 % of the AAs are absorbed through the intestine and released into the portal vein or used by the gut, with only 50–60 % of the ingested dietary protein being released into the systemic circulation. An increase in hepatic/splanchnic uptake of AAs will reduce the amount of AAs available to stimulate the MPS and may blunt the MPS response to protein feeding [72, 73]. This mechanism may therefore contribute to anabolic resistance.
6.2 Microvascular Blood Flow Alterations
Nutritive blood flow is a very important determinant of the anabolic response to food. Poor delivery of nutrients at the sites of microvascular perfusion may contribute to anabolic resistance. Changes in MPS induced by insulin are correlated with changes in AA delivery and blood flow. Alterations in microvascular blood flow associated with reduced AA delivery are observed in critically ill patients, after immobilization, and in elderly. This mechanism may therefore contribute to anabolic resistance. However, although resistance exercise enhances muscle microvascular blood flow in older individuals, it does not restore muscle anabolic response to nutrition [74].
7 How to Override Anabolic Resistance?
7.1 Optimization of Protein Intake
Several nutrition-based strategies may serve to compensate for anabolic resistance. Optimization of protein intake in particular can be applied to maximize the skeletal muscle protein anabolism. The MPS in response to protein intake may be improved by controlling the amount of dietary protein, the nature of the protein, the content in specific AAs, the timing of administration, and the co-ingested macronutrients [3].
In contrast to young adults, in whom postexercise rates of MPS are saturated with 20 g of protein, elderly subjects may need as much as 35–40 g to maximally stimulate MPS [75–79]. Therefore, the elderly may require more protein to acutely increase rates of MPS than the young. This fits with the hypothesis of an increased AA threshold that must be surpassed after protein ingestion to stimulate MPS above rest in elderly [24]. It remains to be determined whether more protein is required to maximize MPS in other situations of anabolic resistance such immobilization and inflammation.
Besides the amount of protein, the nature of the protein ingested may also determine the degree of MPS. The MPS response to a specific protein depends on its digestibility and absorption as well as its AA composition. Both milk and beef ingestion augments the postexercise MPS response, with a stronger stimulation of MPS during the early postprandial stage after milk ingestion [80]. Dairy proteins seem to offer some advantage for muscle anabolism over other protein sources, in particular plant-based soy protein. The two major types of milk protein, casein and whey, markedly stimulate MPS. The more rapidly the protein is digested and absorbed, the greater the postprandial MPS is. Keeping with this, hydrolyzed casein is more potent than intact or micellar casein to stimulate MPS [81]. Whey, a fast digestible soluble protein, increases MPS even more than casein [82]. While casein is converted to a solid clot in the gastric acid environment, whey remains soluble in the stomach, allowing rapid digestion. Differences in anabolic properties of various protein sources are also attributable to differences in AA composition. The higher leucine content of whey protein versus casein may contribute to the greater anabolic properties of whey protein compared to casein. Indeed, there is a good correlation between the rise in circulating leucine concentrations and the postprandial MPS after whey and casein ingestion [83].
Supplementation with BCAAs has been shown to attenuate the loss of muscle mass caused by disuse [84–86] or aging [87]. Supplementation of a suboptimal protein intake with leucine is as effective as a complete protein intake in stimulating postprandial MPS [88]. Leucine-enriched EAAs ingestion after resistance exercise seems to prolong the anabolic response of the skeletal muscle to AAs in older adults [88]. Leucine-rich AA mixtures or proteins appear more efficient than leucine alone to improve muscle mass and performance. However, until now, there is no evidence that chronic free leucine supplementation promotes muscle mass or prevents protein loss during states of anabolic resistance [89, 90]. This may be due to desynchronization between leucine signal and the rise in all AAs, decline in other circulating BCAAs valine, and isoleucine or parallel stimulation of MPB [91].
Finally, the timing of protein intake is an important parameter to consider for optimizing the anabolic response of AAs. The ingestion of protein in the hours just after exercise enhances the MPS. It even seems that this “anabolic window” lasts for at least 24 h following exercise. To reach a near-maximal postprandial MPS response, it is advised to provide 20–25 g dietary protein every 4–5 h and a further 25–40 g protein prior to sleep [30, 92]. Repeated ingestion of 20 g of protein every 3 h during 12 h after a bout of resistance exercise is superior for stimulating MPS than two bolus of 40 g or eight pulses of 10 g [93]. Similarly, emerging evidence suggests that the elderly may need to distribute protein intake evenly throughout the day across three or more daily meals [24], as long as each meal provides enough protein to reach the anabolic threshold (at least 20 g).
It could be suggested that the stimulation of insulin secretion by CHO may enhance the MPS response to protein intake. However, ingestion of CHO with sufficient amounts of protein does not further increase MPS following exercise [94]. Indeed, circulating insulin is more permissive than stimulatory on MPS response [10].
In critically ill patients, there is a lack of data on protein requirements necessary to override the anabolic resistance [95].
7.2 Resistance Exercise
Exercise in general appears to depress MPS, whereas MPB is probably increased, causing negative net muscle protein balance. However, the inhibition of MPS that occurs during muscle contraction is rapidly reversed during postexercise recovery. Therefore, positive net balance may be achieved after the exercise when AA availability is increased, thereby raising MPS markedly. Postexercise-increased AA availability is less crucial than insulin for inhibiting MPB.
Resistance exercise is an important countermeasure to disuse atrophy and to age-related declines in the skeletal muscle mass. What is less well understood is how the intensity and the volume of the resistance exercise stimulus are sufficient to result in rises in MPS. Evidence suggests that minimal resistance exercise preserves MPS throughout bed rest [96]. In older adults, frequent high-intensity weight lifting or alternatively low-intensity high-volume weight lifting increases muscle mass.
The synergistic anabolic effect of resistance exercise in combination with EAAs ingestion has been well documented in particular in the elderly. It appears that protein ingestion at doses at least 20 g and perhaps as high as 30–40 g in close proximity to resistance exercise may be able to elicit an anabolic response in the elderly. Resistance training has been shown to sensitize the skeletal muscle to feeding for up to 24 h [15, 97]. Therefore, resistance exercise training combined with appropriately timed protein (likely leucine-rich) ingestion may represent a highly effective treatment strategy to counteract the sarcopenia.
Older adults are more susceptible than young persons to muscle loss after short-term bed rest. Interestingly, exercise rehabilitation has been shown to restore bed rest-induced deficit in lean mass, strength, and nutrient-induced anabolism in older subjects [23].
There are substantial evidence that resistance training increases muscle mass and strength in older adults [98] with improvement in function and performance of activities of daily living [99].
7.3 Anti-inflammatory Agents
Inflammation has been shown to blunt the anabolic effect of feeding on MPS [100]. Reduction of low-grade inflammation with NSAID restores blunting of postprandial muscle anabolism in old rats and potentiates the exercise-induced increase in muscle mass and strength in humans [101, 102]. Similarly, omega or n-3 fatty acids which exert anti-inflammatory action enhance the sensitivity of MPS to co-infusion of AAs and insulin in older adults [103]. This effect was associated with increased phosphorylation of mTORC1 and S6K1. These observations fit with the hypothesis of an increased anabolic threshold caused by inflammation and an attenuation of anabolic resistance by n-3 fatty acids. It is also possible that n-3 FA have some intrinsic muscle protein anabolic effect, as this stimulation of MPS occurs to the same extent in healthy young subjects where inflammation is probably absent. However, it is not known whether the decrease in the anabolic threshold obtained with anti-inflammatory agents is large enough to preserve muscle mass in the long term in humans.
References
Cohen S, Nathan JA, Goldberg AL (2015) Muscle wasting in disease: molecular mechanisms and promising therapies. Nat Rev Drug Discov 14(1):58–74
Phillips BE, Hill DS, Atherton PJ (2012) Regulation of muscle protein synthesis in humans. Curr Opin Clin Nutr Metab Care 15(1):58–63
Gorissen SH, Remond D, Van Loon LJ (2015) The muscle protein synthetic response to food ingestion. Meat Sci 109:96–100
Atherton PJ, Etheridge T, Watt PW, Wilkinson D, Selby A, Rankin D et al (2010) Muscle full effect after oral protein: time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am J Clin Nutr 92(5):1080–1088
Dideriksen K, Reitelseder S, Holm L (2013) Influence of amino acids, dietary protein, and physical activity on muscle mass development in humans. Nutrients 5(3):852–876
Deutz NE, Wolfe RR (2013) Is there a maximal anabolic response to protein intake with a meal? Clin Nutr 32(2):309–313
Kimball SR, Jefferson LS (2006) Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J Nutr 136(1):227S–231S
Ham DJ, Caldow MK, Lynch GS, Koopman R (2014) Leucine as a treatment for muscle wasting: a critical review. Clin Nutr 33(6):937–945
Svanberg E, Jefferson LS, Lundholm K, Kimball SR (1997) Postprandial stimulation of muscle protein synthesis is independent of changes in insulin. Am J Physiol 272(5 Pt 1):E841–E847
Greenhaff PL, Karagounis LG, Peirce N, Simpson EJ, Hazell M, Layfield R et al (2008) Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am J Physiol Endocrinol Metab 295(3):E595–E604
Barazzoni R, Short KR, Asmann Y, Coenen-Schimke JM, Robinson MM, Nair KS (2012) Insulin fails to enhance mTOR phosphorylation, mitochondrial protein synthesis, and ATP production in human skeletal muscle without amino acid replacement. Am J Physiol Endocrinol Metab 303(9):E1117–E1125
Trommelen J, Groen BB, Hamer HM, de Groot LC, Van Loon LJ (2015) Mechanisms in Endocrinology: exogenous insulin does not increase muscle protein synthesis rate when administered systemically: a systematic review. Eur J Endocrinol 173(1):R25–R34
Heslin MJ, Newman E, Wolf RF, Pisters PW, Brennan MF (1992) Effect of hyperinsulinemia on whole body and skeletal muscle leucine carbon kinetics in humans. Am J Physiol 262(6 Pt 1):E911–E918
Phillips SM, Tipton KD, Aarsland A, Wolf SE, Wolfe RR (1997) Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am J Physiol 273(1 Pt 1):E99–E107
Biolo G, Tipton KD, Klein S, Wolfe RR (1997) An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol Endocrinol Metab 273(1):E122–E129
Timmerman KL, Dhanani S, Glynn EL, Fry CS, Drummond MJ, Jennings K et al (2012) A moderate acute increase in physical activity enhances nutritive flow and the muscle protein anabolic response to mixed nutrient intake in older adults. Am J Clin Nutr 95(6):1403–1412
Haran PH, Rivas DA, Fielding RA (2012) Role and potential mechanisms of anabolic resistance in sarcopenia. J Cachexia Sarcopenia Muscle 3(3):157–162
Rennie MJ (2009) Anabolic resistance in critically ill patients. Crit Care Med 37(10 Suppl):S398–S399
Rennie MJ, Selby A, Atherton P, Smith K, Kumar V, Glover EL et al (2010) Facts, noise and wishful thinking: muscle protein turnover in aging and human disuse atrophy. Scand J Med Sci Sports 20(1):5–9
Glynn EL, Fry CS, Drummond MJ, Dreyer HC, Dhanani S, Volpi E et al (2010) Muscle protein breakdown has a minor role in the protein anabolic response to essential amino acid and carbohydrate intake following resistance exercise. Am J Physiol Regul Integr Comp Physiol 299(2):R533–R540
Guillet C, Boirie Y, Walrand S (2004) An integrative approach to in-vivo protein synthesis measurement: from whole tissue to specific proteins. Curr Opin Clin Nutr Metab Care 7(5):531–538
Bodine SC (2013) Disuse-induced muscle wasting. Int J Biochem Cell Biol 45(10):2200–2208
Tanner RE, Brunker LB, Agergaard J, Barrows KM, Briggs RA, Kwon OS et al (2015) Age-related differences in lean mass, protein synthesis and skeletal muscle markers of proteolysis after bed rest and exercise rehabilitation. J Physiol 593(18):4259–4273
Breen L, Phillips SM (2011) Skeletal muscle protein metabolism in the elderly: interventions to counteract the ‘anabolic resistance’ of ageing. Nutr Metab [Lond] 8:68
Kumar V, Selby A, Rankin D, Patel R, Atherton P, Hildebrandt W et al (2009) Age-related differences in the dose-response relationship of muscle protein synthesis to resistance exercise in young and old men. J Physiol 587(Pt 1):211–217
Durham WJ, Casperson SL, Dillon EL, Keske MA, Paddon-Jones D, Sanford AP et al (2010) Age-related anabolic resistance after endurance-type exercise in healthy humans. FASEB J 24(10):4117–4127
Volpi E, Mittendorfer B, Rasmussen BB, Wolfe RR (2000) The response of muscle protein anabolism to combined hyperaminoacidemia and glucose-induced hyperinsulinemia is impaired in the elderly. J Clin Endocrinol Metab 85(12):4481–4490
Cuthbertson D, Smith K, Babraj J, Leese G, Waddell T, Atherton P et al (2005) Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J 19(3):422–424
Wilkes EA, Selby AL, Atherton PJ, Patel R, Rankin D, Smith K et al (2009) Blunting of insulin inhibition of proteolysis in legs of older subjects may contribute to age-related sarcopenia. Am J Clin Nutr 90(5):1343–1350
Wall BT, Snijders T, Senden JM, Ottenbros CL, Gijsen AP, Verdijk LB et al (2013) Disuse impairs the muscle protein synthetic response to protein ingestion in healthy men. J Clin Endocrinol Metab 98(12):4872–4881
Breen L, Stokes KA, Churchward-Venne TA, Moore DR, Baker SK, Smith K et al (2013) Two weeks of reduced activity decreases leg lean mass and induces “anabolic resistance” of myofibrillar protein synthesis in healthy elderly. J Clin Endocrinol Metab 98(6):2604–2612
Glover EI, Phillips SM, Oates BR, Tang JE, Tarnopolsky MA, Selby A et al (2008) Immobilization induces anabolic resistance in human myofibrillar protein synthesis with low and high dose amino acid infusion. J Physiol 586(Pt 24):6049–6061
Steiner JL, Lang CH (2015) Sepsis attenuates the anabolic response to skeletal muscle contraction. Shock 43(4):344–351
Vary TC, Jefferson LS, Kimball SR (2001) Insulin fails to stimulate muscle protein synthesis in sepsis despite unimpaired signaling to 4E-BP1 and S6K1. Am J Physiol Endocrinol Metab 281(5):E1045–E1053
Lang CH, Frost RA (2004) Differential effect of sepsis on ability of leucine and IGF-I to stimulate muscle translation initiation. Am J Physiol Endocrinol Metab 287(4):E721–E730
Lang CH, Frost RA (2006) Glucocorticoids and TNFalpha interact cooperatively to mediate sepsis-induced leucine resistance in skeletal muscle. Mol Med 12(11–12):291–299
Balage M, Averous J, Remond D, Bos C, Pujos-Guillot E, Papet I et al (2010) Presence of low-grade inflammation impaired postprandial stimulation of muscle protein synthesis in old rats. J Nutr Biochem 21(4):325–331
Williams JP, Phillips BE, Smith K, Atherton PJ, Rankin D, Selby AL et al (2012) Effect of tumor burden and subsequent surgical resection on skeletal muscle mass and protein turnover in colorectal cancer patients. Am J Clin Nutr 96(5):1064–1070
Deutz NE, Safar A, Schutzler S, Memelink R, Ferrando A, Spencer H et al (2011) Muscle protein synthesis in cancer patients can be stimulated with a specially formulated medical food. Clin Nutr 30(6):759–768
Winter A, Macadams J, Chevalier S (2012) Normal protein anabolic response to hyperaminoacidemia in insulin-resistant patients with lung cancer cachexia. Clin Nutr 31(5):765–773
Engelen MP, Safar AM, Bartter T, Koeman F, Deutz NE (2015) High anabolic potential of essential amino acid mixtures in advanced nonsmall cell lung cancer. Ann Oncol 26(9):1960–1966
van Dijk DP, van de Poll MC, Moses AG, Preston T, Olde Damink SW, Rensen SS et al (2015) Effects of oral meal feeding on whole body protein breakdown and protein synthesis in cachectic pancreatic cancer patients. J Cachexia Sarcopenia Muscle 6(3):212–221
Prado CM, Sawyer MB, Ghosh S, Lieffers JR, Esfandiari N, Antoun S et al (2013) Central tenet of cancer cachexia therapy: do patients with advanced cancer have exploitable anabolic potential? Am J Clin Nutr 98(4):1012–1019
Guillet C, Delcourt I, Rance M, Giraudet C, Walrand S, Bedu M et al (2009) Changes in basal and insulin and amino acid response of whole body and skeletal muscle proteins in obese men. J Clin Endocrinol Metab 94(8):3044–3050
Masgrau A, Mishellany-Dutour A, Murakami H, Beaufrere AM, Walrand S, Giraudet C et al (2012) Time-course changes of muscle protein synthesis associated with obesity-induced lipotoxicity. J Physiol 590(Pt 20):5199–5210
Hyde R, Hajduch E, Powell DJ, Taylor PM, Hundal HS (2005) Ceramide down-regulates System A amino acid transport and protein synthesis in rat skeletal muscle cells. FASEB J 19(3):461–463
Nilsson MI, Dobson JP, Greene NP, Wiggs MP, Shimkus KL, Wudeck EV et al (2013) Abnormal protein turnover and anabolic resistance to exercise in sarcopenic obesity. FASEB J 27(10):3905–3916
Stephens FB, Chee C, Wall BT, Murton AJ, Shannon CE, Van Loon LJ et al (2015) Lipid-induced insulin resistance is associated with an impaired skeletal muscle protein synthetic response to amino acid ingestion in healthy young men. Diabetes 64(5):1615–1620
Murphy J, Chevalier S, Gougeon R, Goulet ED, Morais JA (2015) Effect of obesity and type 2 diabetes on protein anabolic response to insulin in elderly women. Exp Gerontol 69:20–26
McIntire KL, Chen Y, Sood S, Rabkin R (2014) Acute uremia suppresses leucine-induced signal transduction in skeletal muscle. Kidney Int 85(2):374–382
Sood S, Chen Y, McIntire K, Rabkin R (2014) Acute acidosis attenuates leucine stimulated signal transduction and protein synthesis in rat skeletal muscle. Am J Nephrol 40(4):362–370
Garibotto G, Sofia A, Russo R, Paoletti E, Bonanni A, Parodi EL et al (2015) Insulin sensitivity of muscle protein metabolism is altered in patients with chronic kidney disease and metabolic acidosis. Kidney Int 88(6):1419–1426
Siew ED, Ikizler TA (2010) Insulin resistance and protein energy metabolism in patients with advanced chronic kidney disease. Semin Dial 23(4):378–382
Etheridge T, Atherton PJ, Wilkinson D, Selby A, Rankin D, Webborn N et al (2011) Effects of hypoxia on muscle protein synthesis and anabolic signaling at rest and in response to acute resistance exercise. Am J Physiol Endocrinol Metab 301(4):E697–E702
Schakman O, Kalista S, Barbe C, Loumaye A, Thissen JP (2013) Glucocorticoid-induced skeletal muscle atrophy. Int J Biochem Cell Biol 45(10):2163–2172
Paddon-Jones D, Sheffield-Moore M, Cree MG, Hewlings SJ, Aarsland A, Wolfe RR et al (2006) Atrophy and impaired muscle protein synthesis during prolonged inactivity and stress. J Clin Endocrinol Metab 91(12):4836–4841
Zheng X, Liang Y, He Q, Yao R, Bao W, Bao L et al (2014) Current models of mammalian target of rapamycin complex 1 [mTORC1] activation by growth factors and amino acids. Int J Mol Sci 15(11):20753–20769
Drummond MJ, Dreyer HC, Fry CS, Glynn EL, Rasmussen BB (2009) Nutritional and contractile regulation of human skeletal muscle protein synthesis and mTORC1 signaling. J Appl Physiol [1985] 106(4):1374–1384
Dickinson JM, Fry CS, Drummond MJ, Gundermann DM, Walker DK, Glynn EL et al (2011) Mammalian target of rapamycin complex 1 activation is required for the stimulation of human skeletal muscle protein synthesis by essential amino acids. J Nutr 141(5):856–862
Kimball SR, Jefferson LS (2010) Control of translation initiation through integration of signals generated by hormones, nutrients, and exercise. J Biol Chem 285(38):29027–29032
Ruas JL, White JP, Rao RR, Kleiner S, Brannan KT, Harrison BC et al (2012) A PGC-1alpha isoform induced by resistance training regulates skeletal muscle hypertrophy. Cell 151(6):1319–1331
Marcotte GR, West DW, Baar K (2015) The molecular basis for load-induced skeletal muscle hypertrophy. Calcif Tissue Int 96(3):196–210
Kelleher AR, Kimball SR, Dennis MD, Schilder RJ, Jefferson LS (2013) The mTORC1 signaling repressors REDD1/2 are rapidly induced and activation of p70S6K1 by leucine is defective in skeletal muscle of an immobilized rat hindlimb. Am J Physiol Endocrinol Metab 304(2):E229–E236
Gordon BS, Williamson DL, Lang CH, Jefferson LS, Kimball SR (2015) Nutrient-induced stimulation of protein synthesis in mouse skeletal muscle is limited by the mTORC1 repressor REDD1. J Nutr 145(4):708–713
Guillet C, Zangarelli A, Mishellany A, Rousset P, Sornet C, Dardevet D et al (2004) Mitochondrial and sarcoplasmic proteins, but not myosin heavy chain, are sensitive to leucine supplementation in old rat skeletal muscle. Exp Gerontol 39(5):745–751
Rivas DA, Morris EP, Haran PH, Pasha EP, Morais MS, Dolnikowski GG et al (2012) Increased ceramide content and NFkappaB signaling may contribute to the attenuation of anabolic signaling after resistance exercise in aged males. J Appl Physiol [1985] 113(11):1727–1736
Drummond MJ, Dickinson JM, Fry CS, Walker DK, Gundermann DM, Reidy PT et al (2012) Bed rest impairs skeletal muscle amino acid transporter expression, mTORC1 signaling, and protein synthesis in response to essential amino acids in older adults. Am J Physiol Endocrinol Metab 302(9):E1113–E1122
Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R et al (2001) Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol 3(11):1014–1019
You JS, Anderson GB, Dooley MS, Hornberger TA (2015) The role of mTOR signaling in the regulation of protein synthesis and muscle mass during immobilization in mice. Dis Model Mech 8(9):1059–1069
Lang CH, Frost RA, Vary TC (2007) Regulation of muscle protein synthesis during sepsis and inflammation. Am J Physiol Endocrinol Metab 293(2):E453–E459
Tardif N, Salles J, Guillet C, Tordjman J, Reggio S, Landrier JF et al (2014) Muscle ectopic fat deposition contributes to anabolic resistance in obese sarcopenic old rats through eIF2alpha activation. Aging Cell 13(6):1001–1011
Boirie Y, Gachon P, Beaufrere B (1997) Splanchnic and whole-body leucine kinetics in young and elderly men. Am J Clin Nutr 65(2):489–495
Gorissen SH, Burd NA, Hamer HM, Gijsen AP, Groen BB, Van Loon LJ (2014) Carbohydrate coingestion delays dietary protein digestion and absorption but does not modulate postprandial muscle protein accretion. J Clin Endocrinol Metab 99(6):2250–2258
Phillips BE, Atherton PJ, Varadhan K, Limb MC, Wilkinson DJ, Sjoberg KA et al (2015) The effects of resistance exercise training on macro- and micro-circulatory responses to feeding and skeletal muscle protein anabolism in older men. J Physiol 593(12):2721–2734
Pennings B, Groen B, de Lange A, Gijsen AP, Zorenc AH, Senden JM et al (2012) Amino acid absorption and subsequent muscle protein accretion following graded intakes of whey protein in elderly men. Am J Physiol Endocrinol Metab 302(8):E992–E999
Yang Y, Churchward-Venne TA, Burd NA, Breen L, Tarnopolsky MA, Phillips SM (2012) Myofibrillar protein synthesis following ingestion of soy protein isolate at rest and after resistance exercise in elderly men. Nutr Metab [Lond] 9(1):57
Yang Y, Breen L, Burd NA, Hector AJ, Churchward-Venne TA, Josse AR et al (2012) Resistance exercise enhances myofibrillar protein synthesis with graded intakes of whey protein in older men. Br J Nutr 108(10):1780–1788
Witard OC, Jackman SR, Breen L, Smith K, Selby A, Tipton KD (2014) Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am J Clin Nutr 99(1):86–95
Witard OC, Cocke TL, Ferrando AA, Wolfe RR, Tipton KD (2014) Increased net muscle protein balance in response to simultaneous and separate ingestion of carbohydrate and essential amino acids following resistance exercise. Appl Physiol Nutr Metab 39(3):329–339
Burd NA, Gorissen SH, van Vliet S, Snijders T, Van Loon LJ (2015) Differences in postprandial protein handling after beef compared with milk ingestion during postexercise recovery: a randomized controlled trial. Am J Clin Nutr 102(4):828–836
Koopman R, Crombach N, Gijsen AP, Walrand S, Fauquant J, Kies AK et al (2009) Ingestion of a protein hydrolysate is accompanied by an accelerated in vivo digestion and absorption rate when compared with its intact protein. Am J Clin Nutr 90(1):106–115
Walrand S, Gryson C, Salles J, Giraudet C, Migne C, Bonhomme C et al (2015) Fast-digestive protein supplement for ten days overcomes muscle anabolic resistance in healthy elderly men. Clin Nutr (in press)
Pennings B, Boirie Y, Senden JM, Gijsen AP, Kuipers H, Van Loon LJ (2011) Whey protein stimulates postprandial muscle protein accretion more effectively than do casein and casein hydrolysate in older men. Am J Clin Nutr 93(5):997–1005
Stein TP, Schluter MD, Leskiw MJ, Boden G (1999) Attenuation of the protein wasting associated with bed rest by branched-chain amino acids. Nutrition 15(9):656–660
Paddon-Jones D, Sheffield-Moore M, Urban RJ, Sanford AP, Aarsland A, Wolfe RR et al (2004) Essential amino acid and carbohydrate supplementation ameliorates muscle protein loss in humans during 28 days bedrest. J Clin Endocrinol Metab 89(9):4351–4358
Paddon-Jones D (2006) Interplay of stress and physical inactivity on muscle loss: nutritional countermeasures. J Nutr 136(8):2123–2126
Fiatarone MA, O’Neill EF, Ryan ND, Clements KM, Solares GR, Nelson ME et al (1994) Exercise training and nutritional supplementation for physical frailty in very elderly people. N Engl J Med 330(25):1769–1775
Ferrando AA, Paddon-Jones D, Wolfe RR (2006) Bed rest and myopathies. Curr Opin Clin Nutr Metab Care 9(4):410–415
Verhoeven S, Vanschoonbeek K, Verdijk LB, Koopman R, Wodzig WK, Dendale P et al (2009) Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men. Am J Clin Nutr 89(5):1468–1475
Balage M, Dardevet D (2010) Long-term effects of leucine supplementation on body composition. Curr Opin Clin Nutr Metab Care 13(3):265–270
Dardevet D, Remond D, Peyron MA, Papet I, Savary-Auzeloux I, Mosoni L (2012) Muscle wasting and resistance of muscle anabolism: the “anabolic threshold concept” for adapted nutritional strategies during sarcopenia. ScientificWorldJournal 2012:269531
Groen BB, Res PT, Pennings B, Hertle E, Senden JM, Saris WH et al (2012) Intragastric protein administration stimulates overnight muscle protein synthesis in elderly men. Am J Physiol Endocrinol Metab 302(1):E52–E60
Areta JL, Burke LM, Ross ML, Camera DM, West DW, Broad EM et al (2013) Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J Physiol 591(Pt 9):2319–2331
Koopman R, Beelen M, Stellingwerff T, Pennings B, Saris WH, Kies AK et al (2007) Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am J Physiol Endocrinol Metab 293(3):E833–E842
Weijs PJ, Cynober L, Delegge M, Kreymann G, Wernerman J, Wolfe RR (2014) Proteins and amino acids are fundamental to optimal nutrition support in critically ill patients. Crit Care 18(6):591
Ferrando AA, Paddon-Jones D, Wolfe RR (2002) Alterations in protein metabolism during space flight and inactivity. Nutrition 18(10):837–841
Devries MC, Phillips SM (2015) Supplemental protein in support of muscle mass and health: advantage whey. J Food Sci 80(Suppl 1):A8–A15
Hagerman FC, Walsh SJ, Staron RS, Hikida RS, Gilders RM, Murray TF et al (2000) Effects of high-intensity resistance training on untrained older men. I. Strength, cardiovascular, and metabolic responses. J Gerontol A Biol Sci Med Sci 55(7):B336–B346
Little JP, Phillips SM (2009) Resistance exercise and nutrition to counteract muscle wasting. Appl Physiol Nutr Metab 34(5):817–828
Costamagna D, Costelli P, Sampaolesi M, Penna F (2015) Role of inflammation in muscle homeostasis and myogenesis. Mediators Inflamm 2015:805172
Rieu I, Magne H, Savary-Auzeloux I, Averous J, Bos C, Peyron MA et al (2009) Reduction of low grade inflammation restores blunting of postprandial muscle anabolism and limits sarcopenia in old rats. J Physiol 587(Pt 22):5483–5492
Trappe TA, Carroll CC, Dickinson JM, LeMoine JK, Haus JM, Sullivan BE et al (2011) Influence of acetaminophen and ibuprofen on skeletal muscle adaptations to resistance exercise in older adults. Am J Physiol Regul Integr Comp Physiol 300(3):R655–R662
Smith GI, Atherton P, Reeds DN, Mohammed BS, Rankin D, Rennie MJ et al (2011) Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: a randomized controlled trial. Am J Clin Nutr 93(2):402–412
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Thissen, JP. (2016). Anabolic Resistance. In: Preiser, JC. (eds) The Stress Response of Critical Illness: Metabolic and Hormonal Aspects. Springer, Cham. https://doi.org/10.1007/978-3-319-27687-8_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-27687-8_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-27685-4
Online ISBN: 978-3-319-27687-8
eBook Packages: MedicineMedicine (R0)