
Abstract
Mass spectrometry (MS) is one of several molecular-level technologies that have revolutionized the practice of clinical microbiology for accurate microorganism identification. Several different MS protocols and assays are being developed and implemented for screening and detection of drug-resistant strains of microorganisms. Functional assays that involve the combination of MS and stable-isotope labeling for establishing drug resistance are reviewed here. These assays include global or local labeling of growth media with C, N, or H stable isotopes in abundance ratios differing from the natural-isotope abundances of these elements. Drug resistance is determined by observing characteristic mass shifts of one or more microorganism-specific biomarkers. A related approach involves the amplification of organism-specific bacteriophages in targeted microorganisms. In this approach, the shift in biomarker masses for phages, initially proliferated in isotopically manipulated growth medium and subsequently grown in microorganisms in growth medium with elements of natural abundance ratios, is monitored and is an indication of drug resistance. Advantages of these assays as well as tools for automating the data analysis are also briefly discussed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
The emergence of drug-resistant microorganisms (“superbugs”), unaffected by available pharmaceuticals, has been recognized as an acute worldwide health problem (Choffnes et al. 2010) recognized by the World Health Organization (WHO 2015). Rapid determination of the responses of a pathogenic microorganism to antibiotics is very important for taking, for example, timely countermeasures in response to a bioterrorism attack or for efficient curbing of the spread of infections in a hospital setting. A US national strategy for combating drug resistance , formulated in the fall of 2014, envisions complex measures to mitigate the emergence and spread of antibiotic resistance (National Strategy on Combating Antibiotic-Resistant Bacteria 2014) . Classical microbiology techniques, including broth dilution or disk diffusion, have been used for decades to determine drug resistance (Coudron et al. 1986; Tang and Stratton 2006) . These techniques infer organism proliferation in the presence of a drug (i.e., resistance) by, for example, monitoring changes in optical density (turbidity) of culture suspensions. However, such classical assays for drug resistance are not rapid, typically taking between 24 and 48 h. These delays reduce markedly the efficacy of efforts to curb disease spread or mitigate effects of bioterrorism activities. Time is of the essence in identifying, treating, or eradicating, particularly, virulent and unknown pathogens. In addition, existing classical tests with higher false-positive or false-negative rates can result in additional complications, wasting valuable resources in improper and inefficient treatments.
Mass spectrometry (MS) , a molecular-level biophysical technology, offers several advantages for pathogen detection, including speed, sensitivity, and specificity, and it rapidly revolutionizes the practice of infectious disease diagnostics (Wilkins et al. 2005; Demirev and Fenselau 2008a, b; Shah and Gharbia 2010; Cliff et al. 2011; Ho and Reddy 2010; Fenselau and Demirev 2011; Havlicek et al. 2013; Sandrin et al. 2013) . A number of MS approaches for direct detection of drug-resistant bacterial strains have been introduced recently (Hrabak et al. 2013; Demirev 2014; Kostrzewa et al. 2013) . For example, matrix-assisted laser desorption/ionization (MALDI) MS generated biomarker profiles differentiate between methicillin-susceptible and methicillin-resistant Staphylococcus aureus (MRSA) strains (Edwards-Jones et al. 2000; Jackson et al. 2005; Du et al. 2002; Wolters et al. 2011; Shah et al. 2011; Burckhardt and Zimmermann 2011; Muroi et al. 2012) . Direct detection of drug metabolites (e.g., degradation products) in cultures grown in the presence of drugs is another MS approach for establishing drug resistance (Hrabak et al. 2013) . In it, the lower m/z (less than 600) range of a mass spectrum is monitored for the presence of peak characteristic of the drug and/or its metabolites. This approach relies on knowledge of the degradation products, arising as a result of the known enzyme-facilitated resistance mechanisms.
Functional Assays for Establishing Drug Resistance
Stable-Isotope Biomarker Labeling During Growth in Isotopically Manipulated Culture Medium (Metabolic Labeling)
The labeling of biomolecules with stable (nonradioactive) isotopes during microorganism growth was introduced almost 60 years ago (Meselson and Stahl 1958) . In the past 20 years, improved sensitivity and signal-to-noise ratio in MS of biomolecules derived from microorganisms have been achieved by growth media manipulation—enrichment or depletion of minor stable isotopes, such as 13C and 15N—and metabolic biomolecule labeling (Oda et al. 1999; Mann 2006; Marshall et al. 1997; Stump et al. 2003) . This has resulted in improved and more accurate microorganism identification (ID; Stump et al. 2003) . A rapid functional method to determine drug resistance in microorganisms has been demonstrated recently (Demirev et al. 2013) . It is based on microorganism growth in stable-isotope-labeled media that also contain target drugs (Sparbier et al. 2013; Jung et al. 2014) . Mass spectra obtained from microorganisms grown under such conditions are compared to control spectra obtained from microorganisms grown in non-labeled media without the drug. Isotope-labeled nutrients are consumed and metabolized during the continued microorganism growth if the organism is not susceptible to the specific drug(s) present. Drug resistance is inferred by observing characteristic mass shifts of one or more microorganism biomarkers. These characteristic mass shifts are a result of the isotopic label(s) being incorporated into the biomarker molecules. The shifts thus indicate that the microorganism is viable even in the presence of the drug. The shifts can be determined experimentally and can also be derived by bioinformatics algorithms. For example, they can be predicted from the isotope composition and the stable-isotope ratios and the actual or estimated biomarker elemental composition. All C-containing molecules consist of approximately one 13C-atom for every 99 12C-atoms at natural-isotope abundance. When a molecule containing n C-atoms is enriched 100 % in 13C (i.e., all 12C-atoms are replaced by 13C) its molecular weight will be shifted by approximately n Da, compared to the molecular weight upon natural-isotope abundance.
Two approaches for isotope labeling of the growth medium have been demonstrated. One approach utilizes globally labeled medium with all growth medium molecules labeled at a predefined isotope ratio of, for example, 13C–12C atoms (Demirev et al. 2013) . A partially labeled (“locally labeled”) medium contains a particular component that is labeled, for example, 13C- and/or 15N-labeled-specific amino acids. The second approach has been introduced more than 10 years ago as a major step in MS-based quantitative proteomics approaches. Stable isotope labeling by amino acids in cell cultures (SILAC; Mann 2006) is the such a major protocol, and it has been used extensively for identification and quantitative evaluation of the expression levels of individual cellular proteins under various conditions.
The first approach has been illustrated with intact Escherichia coli, grown in control (unlabeled) and 13C-labeled media, and analyzed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (Demirev et al. 2013) . This approach is nearly universal: it can be applied to both known and unknown microorganisms (e.g., with unsequenced genomes and/or newly emerging organisms). If the organism’s genome is available, simultaneously with testing for drug resistance the microorganism can also be identified by bioinformatics tools (Pineda et al. 2000; Demirev et al. 2004) . On the other hand, neither prior identification of the microorganism being tested nor prior availability of a reference mass spectrum for that organism is required. Depending on the prior knowledge, one or more algorithms for data analysis can be utilized either individually or in parallel (Fig. 13.1). For example, if the organism’s genome is known, mapping of observed microorganism protein biomarkers to their respective amino acid sequences is possible (Jung et al. 2014). Thus, direct “counting” of all C- or N-atoms in an observed protein would provide the expected mass shift upon substitution of 12C with 13C (14N with 15N), respectively.

Algorithmic approaches for establishing drug resistance in microorganisms by MS, after growth in an isotopically labeled medium. Each algorithm can be applied individually or in parallel, depending on the available prior information. (Reprinted from Demirev et al. (2013) , with permission from Springer)
The local labeling approach based on the incorporation of specific isotopically labeled amino acids has been demonstrated by Kostrewa and coworkers using methicillin-resistant S. aureus (Sparbier et al. 2013) and Pseudomonas aeruginosa (Jung et al. 2014) . In both the studies, either lysine at natural-isotope abundance or 13C- and 15N-labeled lysine has been added to the growth medium. In the second study, the susceptibility of P. aeruginosa to meropenem, tobramycin, and ciprofloxacin (antibiotics with differing mechanisms of action) has been evaluated utilizing a semiautomated algorithm . As expected, a shift of organism-specific biomarkers is observed for drug-resistant strains upon growth in labeled medium (Fig. 13.2). The observed shifts in biomarker masses in the case of locally isotope-labeled medium (i.e., individual amino acids) are typically at least an order of magnitude smaller when compared to shifts, observed for globally labeled medium. Thus, local labeling places more stringent requirements for experimental mass resolution than global labeling. In addition, since in many cases the labeled amino acid may be nonessential (i.e., some can be synthesized de novo during cell development) both unlabeled and labeled biomarker peaks are observed (Fig. 13.2). More sophisticated algorithms, preferably combined with knowledge of the amino acid sequence of the protein biomarker, are required for data deconvolution and semiquantitative analysis when using local isotope labeling protocols. On the other hand, considering the availability of isotope-labeled components of growth media [35], local labeling protocols can be developed more readily from currently existing culture protocols than protocols for global growth medium labeling. Both labeling protocols can be combined with direct MALDI-TOF MS analysis of intact cells or their protein extracts as well as with bottom-up or top-down approaches using liquid chromatography/ESI tandem MS instrumentation. The advantages of using MALDI-TOF MS are relative simplicity of the analysis, combined with microorganism ID, speed, wide instrument availability, and cost effectiveness. Almost all sample preparation and analysis stages can be automated and multiplexed. Isotope-labeling strategies for establishing drug resistance can be particularly useful for classes of anaerobes, fastidious bacteria, and other slow-growing bacteria (Biswas and Rolain 2013) .

Mass (m/z) shifts in MALDI-TOF mass spectra of Pseudomonas aeruginosa. a Control, spectrum for bacteria grown in a medium containing lysine at natural-isotope abundance. b P. aeruginosa grown for 2.5 h in a medium containing 13C- and 15N-labeled lysine. Lysine has six C- and two N-atoms, thus 13C- and 15N-labeled lysine will be 8 Da heavier than its unlabeled counterpart. Incorporation of four and five labeled lysines in the (most probably) ribosomal protein biomarker at m/z 5212 can be inferred from the mass shifts in spectrum (b). (Reprinted from Jung et al. (2014) , with permission from Springer)
Bacteriophage-Based Amplification for Drug Resistance Detection
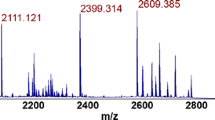
Phage amplification detection (PAD) of bacteria via MS relies on detecting bacteriophages specific to their target host organism (Rees and Voorhees 2005) . Phages self-replicate and proliferate only in metabolically active host cells, have extensive shelf lives, and are inexpensive. Targeted microorganisms are identified through detection by MS of secondary biomarkers originating from organism-specific bacteriophages after their amplification in the target cells. Only proteins indicative of progeny phages, that are bacteria specific, are detected. For example, when both MS2 and MPSS-1 are mixed with E. coli, only an MS2 biomarker protein is detected by MALDI-TOF MS. Phage-based amplification has been successfully expanded into a functional assay for drug resistance screening of targeted bacteria (Pierce et al. 2011, 2012; Rees et al. 2015) . In this process, the shift in characteristic biomarker masses for phages, initially proliferated in stable-isotope-manipulated growth medium, for example, 15N-enriched, is monitored as an indication of successful phage proliferation. High 15N-labeled phage titers (above the MS instrument’s detection limits) are used to spike the sample. The isotope-labeled phages can be readily distinguished from phages subsequently proliferating in bacteria in media with natural-isotope abundance by the observed (downward) shifts in biomarker masses (Fig. 13.3). Monitoring phage growth in drug-containing and control culture media in parallel results in an assay for establishing drug susceptibility —detection of phage-specific biomarkers in samples with the drug will signal the presence of live drug-resistant bacterial strains. The initial input inoculum can be readily distinguished from phages successfully proliferating in the bacteria, which eliminates the possibility of false-positive results. Isotope labeling leads to improved selectivity, high initial phage titers, and sensitivity—only organism-specific phages proliferate. The overall time for organism ID as well as drug susceptibility testing is markedly reduced. The method, combined with MALDI-TOF MS, has been demonstrated for rapid detection of MRSA strains (Pierce et al. 2011) . In this method, a S. aureus-specific phage is initially labeled by proliferating in organisms in 15N-labeled culture medium. Subsequently, the presence of S. aureus is confirmed by detecting a 14N-labeled bacteriophage capsid protein signal after 90-min phage incubation in a sample in a growth medium at natural-isotope abundance. The assay has been combined with LC/ESI/tandem MS for multiple reaction monitoring (MRM) of phage-specific tryptic peptides for rapid and accurate quantitation of viable S. aureus (Pierce et al. 2012) . After spiking the sample with 15N-labeled phages, and following 2-h incubation, the sample is rapidly digested with trypsin. Target tryptic peptides unique to both the 15N-labeled input and 14N progeny capsid proteins are analyzed in MRM. The peptides from a known number of 15N-metabolically labeled phages (109 PFU) are used as an internal standard for quantitation. A linear S. aureus response is achieved in the range from 5.0 × 104 to 2.0 × 106 CFU/mL. Since different antibiotics have different mechanisms of action, there may be a latency period during which the drug might be effective but there will still be phage amplification . To avoid such false positives of purported microbial resistance against, for example, β-lactam antibiotics, a modified phage amplification protocol for simultaneous ID and susceptibility testing to multiple drugs has been implemented (Rees et al. 2015) . Delayed 15N-labeled phage K infection is tested when testing for resistance against slower-acting drugs, including cefoxitin, allowing for effects of the drug to take action (Fig. 13.4). The end-to-end assay can be performed in less than 8 h. This is at least a factor of 3 faster than clinical microbiology assays currently used to detect MRSA.

Overlaid MALDI-TOF mass spectra of initial Staphylococcus bacteriophage 53 inoculum (2 × 108 PFU mL−1), grown in 15N-labeled medium—green trace, and after its propagation for 2 h in S. aureus culture (6.7 × 107 CFU mL−1)—blue trace. The downward shift in mass of the observed biomarkers before and after amplification is due to replacement of the 15N isotope of the approximately 420 N atoms in the phage capsid protein with 14N isotopes. (Adapted with permission from Pierce et al. (2011) , copyright 2011, American Chemical Society)

Workflow for a phage-amplification assay using stable-isotope labeled phages to simultaneously determine the presence of Staphylococcus aureus in a sample, as well as the susceptibility of microorganism to two antibiotics—clindamycin and cefoxitin. (Reprinted with permission from Rees et al. (2015) , copyright 2015, American Chemical Society)
Conclusion
The emergence of multidrug-resistant microorganism strains requires novel methods for antimicrobial resistance and virulence testing. The functional assays for establishing drug resistance , reviewed here, are based on MS and stable isotopes for manipulating culture media. Drug resistance can be inferred from characteristic differences in the masses of primary (microorganism-derived) or secondary (phage-derived) biomarkers appearing upon microorganism growth in the presence of a drug. These assays have a number of advantages. They are much more rapid than classical microbiology assays. Also, they are broadly applicable to a variety of drug/ microorganism types since no prior knowledge of the type of drug and its mechanism of action is required. The assays have potential in high-throughput mass screening against known or unknown pathogens. In a clinical sample, pathogenic microorganism ID/confirmation and drug susceptibility testing can be performed in parallel, thus reducing the time for accurate diagnosis and subsequent treatment. These assays can be multiplexed for simultaneous analysis of mixtures of drugs and organisms (samples), potentially reducing cost. Bioinformatics/statistical methods can be implemented for automated data analysis
References
Choffnes ER, Relman DA, Mack A, Rapporteurs, Forum on Microbial Threats, Institute of Medicine. Antibiotic resistance: implications for global health and novel intervention strategies. Washington, DC: National Academy of Sciences; 2010.
Biswas S, Rolain J-M. Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture. J Microbiol Methods. 2013;92:14–24.
Burckhardt I, Zimmermann S. Using matrix-assisted laser desorption ionization-time of flight mass spectrometry to detect carbapenem resistance within 1-2.5 hours. J Clin Microbiol. 2011;49:3321–4.
Cliff JB, Kreuzer HW, Ehrhardt CJ, Wunschel DE, editors. Chemical and physical signatures for microbial forensics. New York: Springer; 2011.
Coudron PE, Jones DL, Dalton HP, Archer GL. Evaluation of laboratory testing for detection of methicillin-resistant Staphylococcus aureus and Staphylococcus epidermidis. J Clin Microbiol. 1986;24:764–9.
Demirev P. Emerging instrumental methods for antimicrobial resistance and virulence testing. In: Havlicek V, Spizek J, editors. Natural products analysis: instrumentation, methods, and applications. New Jersey: Wiley; 2014. p. 25–42.
Demirev P, Fenselau C. Mass spectrometry for rapid characterization of microorganisms. Annu Rev Anal Chem. 2008a;1:71–94.
Demirev P, Fenselau C. Mass spectrometry in biodefense. J Mass Spectrom. 2008b;43:1441–57.
Demirev PA, Feldman AB, Lin JS. Bioinformatics-based strategies for rapid microorganism identification by mass spectrometry. Johns Hopkins APL Tech Dig. 2004;25:27–37.
Demirev PA, Hagan NS, Antoine MD, Lin JS, Feldman AB. Establishing drug resistance in microorganisms by mass spectrometry. J Am Soc Mass Spectrom. 2013;24:1194–201.
Du Z, Yahng AR, Guo Z, Song Y, Wang J. Identification of Staphylococcus aureus and determination of its methicillin resistance by matrix assisted laser desorption ionization time-of-flight mass spectrometry. Anal Chem. 2002;74:487–91.
Edwards-Jones V, Claydon M, Evason D, Walker J, Fox A, Gordon D. Rapid discrimination between methicillin-sensitive and methicillin-resistant Staphylococcus aureus by intact cell mass spectrometry. J Med Microbiol. 2000;49:295–300.
Fenselau C, Demirev PA, editors. Rapid characterization of microorganisms by mass spectrometry. Washington, DC: ACS; 2011.
Havlicek V, Lemr K, Schug KA. Current trends in microbial diagnostics based on mass spectrometry. Anal Chem. 2013;85:790–7.
Ho YP, Reddy PM. Advances in mass spectrometry for the identification of pathogens. Mass Spectrom Rev. 2010;30:1203–24.
Hrabak J, Chudackova E, Walkova R. Matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry for detection of antibiotic resistance mechanisms: from research to routine diagnosis. Clin Microbiol Rev. 2013;26:103–14.
Jackson KA, Edwards-Jones V, Sutton CW, Fox AJ. Optimisation of intact cell MALDI method for fingerprinting of methicillin-resistant Staphylococcus aureus. J Microbiol Methods. 2005;62:273–84.
Jung JS, Eberl T, Sparbier K, Lange C, Kostrzewa M, Schubert S, Wieser A. Rapid detection of antibiotic resistance based on mass spectrometry and stable isotopes. Eur J Clin Microbiol Infect Dis. 2014;33:949–55.
Kostrzewa M, Sparbier K, Maier Th, Schubert S. MALDI-TOF MS: an upcoming tool for rapid detection of antibiotic resistance in microorganisms. Proteomics Clin Appl. 2013;7:767–78.
Mann M. Functional and quantitative proteomics using SILAC. Nat Rev Mol Cell Biol. 2006;7:952–8.
Marshall AG, Senko MW, Li WQ, Li M, Dillon S, Guan SH, Logan TM. Protein molecular mass to 1 Da by C-13, N-15 double depletion and FT-ICR mass spectrometry. J Am Chem Soc. 1997;119:433–4.
Meselson M, Stahl FW. The replication of DNA in Escherichia coli. Proc Natl Acad Sci U S A. 1958;44:671–82.
Muroi M, Shima K, Igarashi M, Nakagawa Y, Tanamoto K. Application of matrix-assisted laser desorption ionization-time of flight mass spectrometry for discrimination of laboratory-derived antibiotic-resistant bacteria. Biol Pharm Bull. 2012;35:1841–5.
National strategy on combating antibiotic-resistant bacteria, White House Executive Order 13676, Washington, DC, 2014.
Oda Y, Huang K, Cross FR, Crowburn D, Chait BT. Accurate quantitation of protein expression and site-specific phosphorylation. Proc Natl Acad Sci U S A. 1999;96:6591–6.
One source of globally isotope-labeled bacterial growth media, as well as individual stable-isotope labeled compounds for locally-labeled or minimal growth media formulations, is Cambridge Isotope Laboratories, Inc. (Andover, MA, USA).
Pierce CL, Rees JC, Fernandez FM, Barr JR. Detection of Staphylococcus aureus using N-15-labeled bacteriophage amplification coupled with matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry. Anal Chem. 2011;83:2286–93.
Pierce CL, Rees JC, Fernandez FM, Barr JR. Viable Staphylococcus aureus quantitation using N-15 metabolically labeled bacteriophage amplification coupled with a multiple reaction monitoring proteomic workflow. Mol Cell Proteomics. 2012;11:M111.012849. doi:10.1074/mcp.M111.012849-5.
Pineda FJ, Lin JS, Fenselau C, Demirev P. Testing the significance of microorganism identification by mass spectrometry and proteome database search. Anal Chem. 2000;72:3739–44.
Rees JC, Voorhees KJ. Simultaneous detection of two bacterial pathogens using bacteriophage amplification coupled with matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun Mass Spectrom. 2005;19:2757–61.
Rees JC, Pierce CL, Schieltz DM, Barr JR. Simultaneous identification and susceptibility determination to multiple antibiotics of Staphylococcus aureus by bacteriophage amplification detection combined with mass spectrometry. Anal Chem. 2015;87:6769–77.
Sandrin TR, Goldstein JE, Schumaker S. MALDI TOF MS profiling of bacteria at the strain level: a review. Mass Spectrom Rev. 2013;32:188–217.
Shah HN, Gharbia SE, editors. Mass spectrometry for microbial proteomic. New York: Wiley-Interscience; 2010.
Shah HN, Rajakaruna L, Ball G, Misra R, Al-Shahib A, Fang M, Gharbia SE. Tracing the transition of methicillin resistance in subpopulations of Staphylococcus aureus, using SELDI-TOF mass spectrometry and artificial neural network analysis. Syst Appl Microbiol. 2011;34:81–6.
Sparbier K, Lange Ch, Jung J, Wieser A, Schubert S, Kostrzewa M. MALDI biotyper-based rapid resistance detection by stable-isotope labeling. J Clin Microbiol. 2013;51:3741–8.
Stump MJ, Jones JJ, Fleming RC, Lay JO, Wilkins CL. Use of double-depleted C-13 and N-15 culture media for analysis of whole cell bacteria by MALDI time-of-flight and fourier transform mass spectrometry. J Am Soc Mass Spectrom. 2003;14:1306–14.
Tang Y-W, Stratton C, editor. Advanced techniques in diagnostic microbiology. New York: Springer; 2006.
Wilkins CL, Jackson O, Lay JO, editors. Identification of microorganisms by mass spectrometry. New York: Wiley; 2005.
Wolters M, Rohde H, Maier T, Belmar-Campos C, Frank G, Scherp S, Aepfelbacher M, Christner M. MALDI-TOF MS fingerprinting allows for discrimination of major methicillin-resistant Staphylococcus aureus lineages. Int J Med Microbiol. 2011;301:64–8.
World Health Organization. Antimicrobial resistance: draft global action plan on antimicrobial resistance, A68/20, March 27, 2015.
Acknowledgments
Preparation of this review was supported in part by a Stuart Janney Sabbatical Fellowship administered by the Johns Hopkins University Applied Physics Laboratory.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Demirev, P. (2016). Stable-Isotope-Based Strategies for Rapid Determination of Drug Resistance by Mass Spectrometry. In: Demirev, P., Sandrin, T. (eds) Applications of Mass Spectrometry in Microbiology. Springer, Cham. https://doi.org/10.1007/978-3-319-26070-9_13
Download citation
DOI: https://doi.org/10.1007/978-3-319-26070-9_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-26068-6
Online ISBN: 978-3-319-26070-9
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)