
Abstract
The higher cognitive processes cannot be explained by processes of sequential neural activation and new systemic approaches to the study of electroencephalographic correlates of cognition are presently proposed. As the mental activity appears to be the result of a systemic integration of different neural network activities (brain networks, BNet), the associated mutual interaction between different frequency bands generates the phenomenon of cross-frequency modulation (CfM) that supports the synchronization and the coordination of the high-frequency activity of distant brain areas. Within this paper we present two kinds of evidence supporting the key role of CfM in allowing the above mentioned integration, one coming from experimental EEG data concerning human subjects, and the other coming from computer simulations of models of brain network activity. As concerns the experimental evidence in 8 subjects, aged between 20 and 51 years old, not suffering of any psychopathology or neurological disorders, EEG activity was recorded during the execution of a cognitive task in which they were asked to identify the non-previously known criteria used by the computer to classify some presented images. The study of the Event Related Potentials (ERPs) showed a longer latency in responses following an error. Independent EEG components with a strong fit with a dipole model were identified and CfM of these components was computed either independently from the stimulus and related to the stimulus and to the subject response. The obtained results were consistent with the role of CfM in the functional activation of BNets and with the subject cognitive performance. Some computer simulations of the activity of networks of Integrate-and-fire neurons were also carried out. The latter showed that the presence of synchronization between neuron activities was facilitated by the presence of long-range synaptic interactions, periodicity of the stimulus inputs, adaptation of the neuronal threshold and presence of inhibitory synapses. We found also evidence of the key role played by network connectivity structure. Namely, both theoretical arguments and simulation results agreed in showing that, in the case of external discontinuous stimuli the scale-free networks are more likely to manifest a high frequency resonance phenomena coupled with persistent low frequency oscillations. All these data are supporting the role of CfM in the functional integration of complex networks and suggest that this phenomenon could be typical of a large number of other complex systems such as, e.g., the social networks.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 The Systemic Approach to the Study of Brain Activity
A consistent body of neurophysiological research, performed in the last 20 years (and particularly in the last decade) has shown that the higher cognitive processes can not be explained by processes of sequential neural processing. As well known, past neurophysiologic studies based on the technique of evoked potentials were based on this assumption. However, the actually available faster, portable and powerful computational tools allowed the development of far more sophisticated signal processing algorithms. This circumstance led to the availability of the technical and methodological background needed for the study of electroencephalographic correlates of the higher cognitive processes, adopting a non-linear and more systemic approach. Moreover, the integration of neurophysiological studies with brain-imaging studies (fMRI, PET) marked the beginning of a new approach to neuroscience, commonly known as Systems Neuroscience (see, for instance, [23]). Within this context the obtained experimental data allowed us to hypothesize that the mental activity may be the result of a systemic integration of different network activities, not necessarily alternative, based on at least four functional wide range nets (brain networks, BNet) (see, for instance, [3, 12, 16, 18, 19]). These four BNet handle respectively the inner thoughts and memories of the management, evaluation of external stimuli and response behaviors, the level of attention and awareness and the assessment of stimulus meaning. The related neurophysiological underlying phenomena are the expression of a systemic coordination of the activities of a large numbers of neurons. This coordination can be detected in the scalp EEG signal under the form of a phase coherence observed in different frequency bands on more or less extensive areas of the cortex. Some authors attribute to phenomena of phase synchronization at low frequencies also the classical perturbations of EEG waves observed during the first 600 ms after the presentation of a stimulus [8, 10, 14]. These bands seem to correlate with different functions, such as the analysis of the formal characteristics of the stimuli (which occurs locally and which correlate with EEG activity on higher frequency bands), or the processes of long-distance communication, through which meaningful information results in the activation of many different functional areas, more or less extensive, of different networks [12, 20, 24, 25]. Different bands thus play different roles [17] and broader synchronized activations can generate different system phenomena, such as the emergence of conscious thought processes [13, 21]. The mutual interaction between different frequency bands generates the phenomenon of cross-frequency modulation (CfM), that not only supports the synchronization and the coordination of the high-frequency activity of areas even very far, but may also represent a generalized mode of systems communication and integration [6].
2 The Cross-Frequency Modulation
As well known, the phenomenon of CfM typically concerns the communication between two different brain networks—here called, for simplicity, the sender and the receiver—and consists in the modulation of the amplitude of high-frequency oscillations of the receiver by the phase of low-frequency oscillations of the sender. Such a phenomenon, however, has been sometimes detected also within single networks. The CfM appear as characterized by features more or less shared by normal subjects, while pathological subjects (like schizophrenic ones) are characterized by different feature patterns (see, for instance, [2]). Thus, while CfM seems to be a robust organizational principle of systemic integration, it could be used also as a diagnostic tool.
Many authors have argued that EEG oscillations in the high-frequency gamma band are local expression analysis of the specific properties of the stimulus, while the low-frequency oscillations are primarily an expression of information processes integration in large functional networks. In this way the integration of the details of the stimulus in more complex psychological entities would be done. At the same time the theoretical data and the simulation data lead us to hypothesize in a similar way that the low-frequency oscillations allow the transmission of information to a wide distance, with higher speeds than the information integrated by local high-frequency activities. This would allow the coordinated analysis on a large scale macro system. These data have an anatomical correspondence in the relation between the white matter myelinated pathways and the gray matter pathways of the brain. The phenomenon of CfM, therefore, it is likely to be a key element of the brain integration [9], featuring the learning process [22], the higher cognitive functions as well as the emergence of consciousness [13, 21]. At the same time it could represent also a basic mode in which the information is propagated in scale-free networks, where clusters of the type “small world” are complemented by hub nodes [7]. This type of model finds in the thalamic-cortical-striatal-thalamic circuits their anatomical and functional correspondence [26]. In this perspective it has to be also reminded the heterogeneous structure of the thalamus, in which nuclei, with small-world architecture, are interconnected and separated by areas containing hub neurons (intra-laminar nuclei).
3 An Experimental Study of CfM
A preliminary study was conducted on 8 subjects, 4 males + 4 females, right handed, aged between 20 and 51 years old (m.: 32. 3 ± 10. 3), not suffering of any psychopathology or neurological disorders. The experimental setting consisted in a cognitive task to be completed while EEG signal was recording. EEG signal was recorded at 250 Hz with 256 channels caps by EGI Geodesic equipment. The cognitive task (Cognitive Stress Paradigm—CSP) is a visual paradigm presented on the screen at 60 cm of distance. The CSP is very similar to the Wisconsin Card Sort Test where the geometric pictures presented in the 64 cards are substituted by 64 cartoons. The cartoons concern pictures of landscapes, animals, buildings or plants presented on one card in groups of 1 to 4 elements with four different background colors (light blue, orange, yellow, green). The subject had to match these \(4 \times 4 \times 4 = 64\) cards, presented one per time, with four example cards displayed in the top row of the screen. The matching criteria were the background color, the number of the elements inside the picture and the class of picture presented (landscapes, animals, buildings or plants). The computer chose randomly the matching criteria (avoiding to get two equal criteria one after the other) which were blind to the subject and were stable till a specific number of correct responses was reached. This number varied randomly from 8 to 12. The subject had to infer the criteria adopted in that moment by the computer which just gave a feedback to the subject if the response was correct or not. The response had to be given within 5 s. If this time limit was overcome, the response was considered as dropped. The task was stopped when the limit of \(64 \times 2 = 128\) presented cards was reached or the subject completed 6 consecutive criteria.
The EEG tracks were processed by means of MatLab and by EEGlab package. The EEG records were reduced from 256 to 71 channels, and cleaned for muscle and EOG artifacts. Then an Independent Component Analysis (ICA) was performed and a dipole best fit computed for each Independent Component (IC). The ICs, with the best fit dipole inside the brain volume and a residual variance of less than 10 %, were considered for further analysis. This ICs pool was segmented in epochs with a time lag ranging from − 200 to 800 ms (Time 0 = card presentation start time). Taking into account the subject performance to the previous and to the present trial of CSP, the epochs were labeled in four different groups: Correct Response after Correct One, Correct Response after an Error, Error Response after Correct, and Error Response after an Error. The error after the criteria change was excluded by the error responses. The pool of the selected ICs of all subjects was grouped by means of principal component analysis (PCA) and cluster analysis (CA). The statistical tests non executed by MatLab were done by means of the statistical package SPSS.
The observed parameters have been:
-
1.
behavioral parameters describing the performance to the CSP as it is in the original WCST: time to complete task, average trial time, number of completed categories, failures to maintain the set, number of total errors, number of perseverant errors, drops, number of correct responses, total number of presented cards;
-
2.
electrophysiological parameters describing the event related EEG perturbation occurring within 800 ms after the stimulus: the usual ERP waves (N e , N 1, N 2, P 2, P 3a , P 3b );
-
3.
system integration indices (see [6]): the Cross Frequency Modulation Index (CfMI), the Modulation Phase Angle (MPA) and the Resultant Vector Length (MVL) and the residual variance of the associated dipole of the selected ICs (DIP rvcm ).
4 Behavioral Data
The mean time to complete the task was 12. 0 ± 2. 5 min, and completed categories were 4. 5 ± 1. 7 on average, with a mean time of 1. 52 ± 0. 14 min/category when the criterion of 6 completed categories was reached. Mean perseverant errors were 7. 15 ± 3. 3, mean drops 2. 18 ± 2. 4, mean failures to maintain the set 1. 5 ± 1. 8. When six categories were completed a mean of 84. 7 ± 7. 5 cards were shown. Mean response time was 1901 ± 523 ms, with significant differences (F 3, 166 = 25. 33; p < 0. 001) between response conditions (Correct on Correct: 1489 ± 344 ms; Error on Correct: 1816 ± 507 ms, Correct on Error: 1996 ± 428 ms; Error on Error: 2296 ± 451 ms).
5 Brain Areas Involved in the Event Related Response to CSP
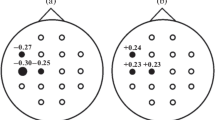
Globally within the 8 subjects considered in this study, 81 ICs were selected and they were aggregated by PCA and CA in 9 clusters: only 4 of these 9 clusters (ACC—anterior cingulate cortex, l-OFC—left orbito frontal cortex, visual cortex and lPOC—left occipito parietal cortex) presented significant difference in ERSP in relation to the task performances.
6 Correlation Study About CfM Effect Relationship with Neurophysiology and Task Performance
In the whole group of nine clusters the CfM parameters were related to the task performance and neurophysiologic parameters by a correlation study (Tables 2.1, 2.2, 2.3, and 2.4).
7 Modulation Delay (Phase Angle)
The parameter with the highest number of significant correlations has been the Phase Angle computed on continuous data (MPA cd ), which appears to be higher (this could mean to represent a more delayed CfM effect) when the attempts to 1st category are lower (\(R = -0.228\); p = 0. 003), the cards number is higher (\(R = -0.223\); p = 0. 004), the time to complete the task is shorter (\(R = -0.262\); p = 0. 001), but also when errors (\(R = -0.230\); p = 0. 003) and perseverant errors (\(R = -0.243\); p = 0. 001) are higher. Theses data can be interpreted as indicating that the presence of errors leads to a delay in modulation per modulating frequency, as the number of structures and the length of involved pathways increase.
8 Number of Brain Resources Involved (Dipole Residual Variance)
However the mean cluster residual variance of the matched IC dipoles (DIP rvcm ) has the strongest relationships with the task performance and a high DIP rvcm is correlated with a higher number of completed categories (\(R = +0.391\); p < 0. 001), with a lower number of drops (\(R = -0.554\); p < 0. 001) and attempts to complete the 1st category (\(R = -0.567\); p < 0. 001), with a lower frequency of failure to maintain the set (\(R = +0.292\); p < 0. 001), and a shorter time to complete task (\(R = -0.604\); p < 0. 001). This means that the single dipole model for those cluster ICs was less probable in spite of a multiple dipole model when the best performances are observed.
The presence of a higher number of completed categories was related also to a shorter latency of P 2 wave of ERPs (\(R = -0.213\); p = 0. 005) (Table 2.2), which in turn is related to a higher DIP rvc m (\(R = -0.157\); p < 0. 040), (Table 2.3). The presence of a higher DIP rvcm relates with a lower amplitude of P 3a and P 3b , as well as a longer latency of N 2 and P 3a and a shorter latency of P 2. This means that the lag N 2 − P 2 is shorter and the lag P 2 − P 3a longer, and can be interpreted as the combined effect of more than one modulating frequency band, a hypothesis that it is coherent with the involvement of many dipoles.
9 Activation of Salience Network (Event Related Modulation Index and N 1-P 2-N 2 Triplet)
The CfM Index computed on continuous data (CfMI cd ) resulted to be higher when the perseverant errors were lower (\(R = -0.286\); p < 0. 001) and the mean response time is shorter (\(R = -0.600\); p < 0. 001). On the other hand the presence of higher frequency of perseverant errors appeared to be linked to a lower value of CfMI cd (Table 2.1), while the same index computed on segmented data (CfMI sd ) gave no significant results (Table 2.1). The same condition of low frequency of perseverant errors was linked also with a lower response on the N 1 wave (\(R = +0.215\); p = 0. 005) (less deep negativity) and P 2 wave (\(R = -0.296\); p < 0. 001) (less high positivity) on ERP profile (Table 2.2). The presence of a higher CfMI cd and CfMI sd was related to an enhancing effect on N 1, P 2 and N 2 waves (Table 2.3) (CfMI cd : N 1, \(R = -0.317\), p < 0. 001; P 2 \(R = +0.179\), p = 0. 019; N 2 \(R = -0.281\); p < 0. 001) (CfMI sd : N 1 \(R = -0.375\), p < 0. 001; P 2 \(R = +0.437\), p < 0. 001; N 2 \(R = -0.367\); p < 0. 001). These data are coherent with a synchronization effect of CfM on early EEG events before 300 ms after the card exposure, which leads to an augmented amplitude of the EEG oscillations described by the triplet N 1-P 2-N 2. This triplet usually is correlated with the orienting reaction to novelty or error and to the salience attribution process. We can see also an increase of N 2 latency (the third peak of the triplet) in relation to both CfMI cd (\(R = +0.166\), p < 0. 031) and CfMI sd (\(R = +0.282\), p < 0. 001), a fact which suggests that this synchronization effect may be related to a decrease in the modulating frequency, which in turn can explain the slowing of response reaction when the C wave oscillation (orienting response) increases (Tables 2.2 and 2.3). The higher Pearson R found with parameters computed on segmented data may suggest that this CfM may be actually related to the new stimulus reaction.
10 Error State Management (Non Event Related Modulation Index and N e )
The N e wave and early negativity are usually related with the error condition. This circumstance (Table 2.2) appears to be less evident (less negative) when correct responses occur (\(R = +0.216\), p = 0. 005) and it appears to be enhanced (more negative) by a modulating effect of CfMI cd computed on continuous data (\(R = -0.343\), p < 0. 001) (Table 2.4). The non significant correlation with the CfMI cd computed on segmented data (Table 2.4) may suggest a stimulus independent dynamics in such a modulation. It remains here open the question if it can be related to the previous feedback received. Differently from what was found with the N 1-P 2-N 2 triplet, when enhanced the N e appears to correlate with shorter response time (Table 2.4).
11 Considerations About Experimental Data
When interpreting the previously presented data, we have to take into account the fact that the present study has considered parameters relative to the whole epoch of 5000 ms of inter stimulus time-lag (Cross frequency Modulation and Behavioral Parameters) and parameters relative to the time-lag 0–800 ms after the card onset. Subjects are exposed (during the 5 s of between-cards time lag) to two important stimuli: the card onset, with the consequent orienting, salience and response processing, and the computer reward, occurring 300 ms after the subject response. Responses occurred on average about 1900 ms after the card onset, with a response mean time varying significantly in respect to the context of the previously received reward and to the quality of present response.
Thus our results (especially those linked to neurophysiologic data) are focused mainly on orienting reaction and salience attribution (concerning the salience network activities), and just the early phases only of the response processing. Moreover CfM parameters may represent two kind of conditions: one condition may be linked to the card presentation, grouped by previous response and present performance, and the other may be linked to a general subject attitude toward error risk management, independent from card onset.
Taking into account this framework, the error management as an attitude to expect to deal with an error risk (N e ) seems related more to a stimulus independent attitude then to an event related reactivity. Moreover the presence of an error context seems to correlate with a delay in cross frequency modulation effect alike if, in such a context, more resources and longer pathway could be involved. The involvement of more brain resources as a prerequisite to best performances has a correlate with the increase of residual variance value when best task performances are observed. This attributes to this parameter a meaning supplementing its more obvious one: i.e. in spite of the fact that a lower value of it is considered as a good match for the IC associated dipole, we have, however, that in experimental settings the residual variance can be influenced by the number of dipoles generating that IC. In other words such a hypothesis implies that an EEG IC may be not concerning just only one dipole but it can be produced by the integrated action of many neighboring dipoles. This result is congruent with other experimental evidences, which support the theory that more complex cognitive actions imply the activation and synchronization of many brain structures. Moreover in such conditions, it is possible that the CfM activity may act through different EEG bands (Delta, Theta, Alfa) on many EEG high frequency bands.
Finally the focusing of neurophysiologic parameters on ERPs occurring in the first 500 ms implies that the action of the salience network is more evidenced then the activity of other BNets, as it could be shown by the involvement of the N 1-P 2-N 2 triplet and by the modulation of such activity more likely done by a new stimulus related CfM.
In any case these results confirm the role of CfM in Brain Network activation and modulation. At the same time they highlight the fact that this phenomenon has an inner complexity and modulation in relation to the type of action and to the complexity of processing.
12 Computer Simulations
The existence of a tight interconnection between theory and experiments, which characterizes the systems neuroscience, induced us to resort to computer simulations of neural network activity in order to support the validity of the hypotheses concerning phenomena such as the CfM. In these simulations we used the Integrate-and-Fire neuron models, which are computationally simple enough while, at the same time, being endowed with a good level of plausibility. In logical terms, the dynamical rule of this kind of neurons can be written under the form:
Here v(t) is the membrane potential, s a threshold, A the spike amplitude, and c the resting potential. As shown by a number of authors (see [4, 5]) networks made by these neurons exhibit, in presence of a suitable percentage of excitatory and inhibitory connections, globally synchronized oscillations and multistability phenomena.
Our simulations, differently from what can be generally found in literature, deal with networks in which each neuron is characterized by a “personalized” set of parameter values, initially chosen in a random way. The global network behavior, however, is influenced not so much by the inhomogeneity of the neurons, but rather by the nature of their interconnections, chiefly by the presence or absence of long-range interconnections. In this regard, an important feature of the connections is given by the degree of each unit, identified with the number of connections (or links) related to the unit itself. We must remind that the scale-free networks, already quoted above, are characterized (see [1]) by the fact that the probability distribution of the degree k of their units P(k) depends on k through a law of the form:
These networks, as already mentioned, are characterized by the presence of a small number of hubs, that is units with high degree, and a large number of units with small degrees. Some researchers (cfr. [9]) suggested that the phenomenon of CfM is a direct consequence of the scale-free character of the biological networks, because the typically large distances between the hubs should favor an information transmission based on large wavelengths (therefore long-ranged) and small frequencies, whose phase should necessarily modulate the short-range high-frequency signals. But the main question is: how the neural networks acquire a scale-free nature?
To answer we performed a number of simulations including not only synaptic interconnections between neurons but also gap junctions. Without entering into technical details (see [15]), we mention that in our simulations, following the method of Lewis and Rinzel [11], the gap junctions act on a neuron through two influences: by directly sending the spikes and by sending a signal dependent on the difference between the membrane potentials of the two involved neurons. Concerning the synaptic efficacies, we postulated an Hebbian law of variation, dependent on spike correlations, including also a spontaneous decay and a saturation effect. The dynamical contributions of the synapses and gap junctions were both weighted, but with different coefficients, so as to allow to investigate the effect on network dynamics of strong, weak, and absent gap junctions. Without showing the results of all simulations performed, we will limit here to present the snapshots of average network activities in three different simulations, all performed on networks of 100 neurons, whose initial connections were given at random.
As it is possible to see from Fig. 2.1, a strong influence of gap junctions leads towards a stable attractor, with a constant value of average activity. However after 100 time steps the network achieved a scale-free structure with γ = 1. 28055. In the case of absence of gap junctions we see from Fig. 2.2 a seemingly noisy average activity, characterized, however, by the presence of different frequency components. In this case the network achieved a strongly scale-free structure with γ = 1. 847489. Finally Fig. 2.3 shows that, in presence of a weak effect of gap junctions, a globally oscillatory state characterizes the network dynamics, while the scale-free features practically disappeared (we obtained a modest γ = 0. 3550416).

Strong gap junctions (activation vs time)

Absence of gap junctions (activation vs time)

Weak gap junctions (activation vs time)
These results seem to evidence that the gap junctions could act as modulators of the network structure and, therefore, of the nature of global activity it can support. It remains to be found what are the physiological processes implementing such a modulation activity. Answering this question would be equivalent to understand in what circumstances we could expect a normal operation of CfM.
13 Conclusions
In principle, it would be possible that a network, with synaptic weights varying as a function of the inner activity according to an Hebbian-like law, could reach a scale-free structure, if supported by a suitable influence of gap junctions. This would open the way to a conception of the CfM as an almost universal systemic feature of the normal operation of most systems (not only of neural networks). If this would be the case, then this model could be exported also to more complex systems such as social networks, in which micro-social units with interpersonal high density relationships similar to “small world” architectures can be integrated in macro-social structures in which the small world elements are mutually functionally integrated by individuals with the ability to “hubbing” through low frequency collective activities.
References
Albert, R., & Barábasi, A.-L. (2002). Statistical mechanics of complex networks. Reviews of Modern Physics, 74, 47–97.
Allen, E. A., Liu, J., Kiehl, K. A., Gelemter, J., Pearlson G. D., Perrone-Bizzozero, N. I., & Calhoun, V. D. (2011). Components of cross-frequency modulation in health and disease. Frontiers in Systems Neurosciences, 5, 1–16.
Bressler, S. L., & Menon, V. (2010). Large-scale brain networks in cognition: Emerging methods and principles. Trends in Cognitive Sciences, 14, 277–290.
Brunel, N. (2000). Dynamics of sparsely connected networks of excitatory and inhibitory spiking neurons. Journal of Computational Neuroscience, 8, 183–208.
Brunel, N., & Hakim, V. (2008). Sparsely synchronized neuronal oscillations. Chaos, 18, 015113.
Canolty, R. T., & Knight, R. T. (2010). The functional role of cross-frequency coupling. Trends in Cognitive Sciences, 14(11), 506–515.
Da Rocha, E. L., & da Cunha, C. R. (2011). The transition from fracton to phonon states in a Sierpinski triangle lattice. Chaos, Solitons & Fractals, 44, 241–247.
Gruber, W. R., Klimesch, W., Sauseng, P., & Doppelmayr, M. (2005). Alpha phase synchronization predicts P1 and N1 latency and amplitude size. Cerebral Cortex, 15, 371–377.
He, B. J., Zempel, J. M., Snyder, A. Z., & Raichle, M. E. (2010). The temporal structures and functional significance of scale-free brain activity. Neuron, 66, 353–369.
Klimesch, W., Schackb, B., Schabusa, M., Doppelmayr, M., Gruber, W., & Sauseng, P. (2004). Phase-locked alpha and theta oscillations generate the P1-N1 complex and are related to memory performance. Cognitive Brain Research, 19, 302–316.
Lewis, T. J., & Rinzel, J. (2003). Dynamics of spiking neurons connected by both inhibitory and electrical coupling. Journal of Computational Neuroscience, 14, 283–309.
Meehan, T. P., & Bressler, S. L. (2012). Neurocognitive networks: Findings, models, and theory. Neuroscience and Biobehavioral Reviews, 36, 2232–2247.
Melloni, L., Molina, C., Peña, M., Torres, D., Singer, W., & Rodriguez, E. (2007). Synchronization of neural activity across cortical areas correlates with conscious perception. The Journal of Neuroscience, 27(11), 2858–2865
Muller, V., & Anokhin, A. P. (2012). Neural synchrony during response production and inhibition. PLoS One, 7(6), e38931.
Ostojic, S., Brunel, N. & Hakim, V. (2009). Synchronization properties of networks of electrically coupled neurons in the presence of noise and heterogeneities. Journal of Computational Neuroscience, 26, 369–392.
Park, H.-J., & Friston, K. (2013). Structural and functional brain networks: From connections to cognition. Science, 342, 1238411.
Peña, M., & Melloni, L. (2012). Brain oscillations during spoken sentence processing. Journal of Cognitive Neuroscience, 24(5), 1149–1164.
Raichle, M. E. (2011). The restless brain. Brain Connectivity, 1(1), 3–12.
Sporns, O. (2010). Networks of the brain. MIT Press: Cambridge, MA.
Steinke, G. K., & Galan, R. F. (2011). Brain rhythms reveal a hierarchical network organization. PLoS Computational Biology, 7(10), e1002207.
Tononi, G. (2010). Information integration: Its relevance to brain function and consciousness. Archives Italiennes de Biologie, 148, 299–322.
Tort, A. B., Komorowski, R. W., Manns, J. R., Kopell, N. J., & Eichenbaum, H. (2009). Theta-gamma coupling increases during the learning of item-context associations. Proceedings of the National Academy of Sciences of the United States of America, 106, 20942–20947.
van Hemmen, J. L., & Sejnowski, T. J. (Eds.). (2006). 23 problems in systems neuroscience. Oxford University Press: New York.
von Stein, A., & Sarnthein, J. (2000). Different frequencies for different scales of cortical integration: From local gamma to long range alpha theta synchronization. International Journal of Psychophysiology, 38, 301–313.
Yeung, N., Bogacz, R., Holroyd, C. B., Nieuwenhuis, S., & Cohen, J. D. (2007). Theta phase resetting and the error-related negativity. Psychophysiology, 44, 39–49.
Zamora-Lopez, G., Xhou, C., & Kurths, L. (2011). Exploring brain function from anatomical connectivity. Frontiers in Neurosciences, 5(83), e2011.00083.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Marconi, P.L., Bandinelli, P.L., Penna, M.P., Pessa, E. (2016). Cross-Frequency Modulation, Network Information Integration and Cognitive Performance in Complex Systems. In: Minati, G., Abram, M., Pessa, E. (eds) Towards a Post-Bertalanffy Systemics. Contemporary Systems Thinking. Springer, Cham. https://doi.org/10.1007/978-3-319-24391-7_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-24391-7_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-24389-4
Online ISBN: 978-3-319-24391-7
eBook Packages: Business and ManagementBusiness and Management (R0)