
Abstract
Living systems, besides acquiring information ontologically, i.e., as an inherent part of their nature, have the special ability to constantly incorporate the available data on their environment through learning (cognition). Therefore, cognition is one of the basic properties that characterize living beings, since even lesser organisms display some kind of learning. This chapter shares the opinion that “humans are a cultural species,” because much of human behavior is determined by an information system acquired via social transmission. It is assumed that humans are characterized by the display of behaviors derived from information processing of three distinct natures: ontological, those acquired by individual experience, and those acquired by social transmission. It is recognized that culture can evolve following Darwinian assumptions, and that knowledge transmission is one of the elementary processes in cultural evolution and in the relation between people and their understanding of nature. This chapter now turns to the three bases description of human information, and then it turns its attention to social knowledge transfer from an evolutionary perspective. Finally, we discuss how information transmission has been analyzed by ethnobiological studies and how it can be used to understand the evolutionary dynamics of social-ecological systems, drawing upon standard ethnobiological works.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
7.1 Introduction
It is certainly possible to affirm, even in general, that all living beings are essentially structures capable of storing and processing information. It does not mean that this property defines life itself, as other physical systems are capable of storing and processing information as well. What distinguishes living systems from physical systems? The difference is based on their origin and nature. Living systems, besides acquiring information ontologically, i.e., as an inherent part of their nature, have the special ability to constantly incorporate the available data on their environment through learning (cognition). The classical learning concept considers any change in animal behavior that comes from the processing of new information, acquired through experience at a given moment in time (T 1) and which can be detected at a later moment (T 2) (Heyes 1994). Therefore, cognition is one of the basic properties that characterize living beings, since even lesser organisms display some kind of learning process (Maturana and Varela 2007).
But if acquired information distinguishes living systems from physical systems, what distinguishes humans from other living beings? This chapter shares the opinion of Mesoudi (2011), who stated categorically that “humans are a cultural species,” because much of human behavior is determined by an information system acquired via social transmission, through specific processes such as imitation, teaching and language. In this sense, the concept of social transmission equals the concept of culture (Richerson and Boyd 2005; Mesoudi 2007; Mesoudi 2011). In summary, it is assumed that humans are characterized by the display of behaviors derived from information processing of three distinct natures: ontological, those acquired by individual experience, and those acquired by social transmission.
To investigate knowledge through social transmission is essentially to analyze the peculiar nature of human beings, i.e., their culture. Furthermore, it is recognized that culture can evolve following Darwinian assumptions, and that knowledge transmission is one of the elementary processes in cultural evolution and in the relation between people and their understanding of nature. This chapter now turns to the three bases description of human information, and then it turns its attention to social knowledge transfer from an evolutionary perspective. Finally, we discuss how information transmission has been analyzed by ethnobiological studies and how it can be used to understand the evolutionary dynamics of social-ecological systems, drawing upon standard ethnobiological works.
7.2 Genetic Basis Information
Some kinds of human behavior are the result of information processing stored in genetic material. These behaviors are characterized by not being influenced by peers during the acquisition process (Mesoudi and Whiten 2008). Genetic inheritance is not related to a trait of specific knowledge, such as knowing a plant is used for a particular purpose, but by determining individual characteristics that favor or not this knowledge construction, such as the capacity and speed of learning (Reyes-Garcia et al. 2009). However, there are few studies within ethnobiology that explicitly discuss inherited or genetically predisposed behaviors. Thus, little is known about the cultural aspects that are influenced a priori by an individual biological basis, i.e., inherent to an individual.
7.3 Individual Knowledge Production
Individual knowledge production is the process by which the individual builds, mostly through trial and without any social influence (Rendell et al. 2009), new information that may spread and settle in wider culture or not. This cognitive process is also named “individual learning” (Heyes 1994) or “asocial production” (Laland 2004). However, the use of the term “individual learning production” rather than the term “asocial learning” is defended, as it is believed that no knowledge is, in its entirety, “asocial”. Even when knowledge is produced by a single individual, its necessity and the reality in which it is produced are the result of a context and of social relations.
From an evolutionary point of view, individual learning is important for the following reasons: (a) it demands time and energy to access information; (b) in special situations it may cause risk to the individual, for example, when developing knowledge about medicinal plants; (c) although costly, new information will always adapt to the context in which it was conceived, even in unstable environments; and (d) being analogous to the mutation process in biological evolution, it diversifies traits and behaviors to be selected in a culture. Based on the considerations above, considering trade off, Footnote 1 energy expenditure and adaptability, individual knowledge production is favored under environmental instability conditions, because although costly, innovations will always be adapted to the new context (Cavali-Sforza and Feldman 1981; Hewlett and Cavali-Sforza 1986; McElreath and Strimling 2008).
It is important to know in ethnobiological studies that the evolutionary assumptions outlined above are not always true, because depending on how it occurs, individual learning will have different evolutionary implications. As stated, the theoretical accumulation of individual learning through evolutionary implications assumes that the production of new information, or innovations, will always be costly and in some situations will present risks to the individual, such as trying new plants for the treatment of a serious illness. However, it is possible to discover an innovation without energy expenditure and without risk to the individual, a point that is generally disregarded by evolutionary approaches. Considering that planting is an action determined by rain stimulus, for example, a farmer needs to know the ideal time to perform it. Thus, through his/her perception, but without any landscape management or planning experience, he/she realizes that a given plant, such as a “ipê rosaFootnote 2,” for example, flourishes days before the rain. The farmer then determines the “plant” action to the “flourish” stimulus offered by the plant. In this case, the new knowledge (behavior), “it is good to plant when the ‘ipê rosa’ flourishes”, is the result of an event that did not require additional time or energy for its production. If this hypothesis is true, it is necessary to relativize discussions on cultural evolution guided by the premise that, although highly adaptive, individual knowledge production demands surplus energy expenditure.
7.4 Information Transmission or Social Learning
Social learning is the process by which information is transmitted in a social group through the interaction between peers, or of those with interaction products (Boyd and Richerson 2005; Mesoudi and Whiten 2008). For many authors, culture is nothing but shared knowledge, i.e., the information set acquired by social transmission through mechanisms such as imitation, teaching and language (Rogers 1988; Boyd and Richerson 2005; Mesoudi 2011).
One of the most fruitful discussions, especially in comparative psychology, is the one that seeks to understand the social learning process, i.e., whether this culture is exclusive to human beings or if it also exists in nonhuman animals. On the one hand, some studies suggest that certain animals, especially chimpanzees, are able to develop their own language mechanisms and behavioral traditions, which are transmitted through many generations (Boesh and Tomasello 1998). Such evidence suggests that human cultural systems have evolutionary roots prior to Homo sapiens. So, what are the unique features of human cultural systems that distinguish them from other cultural animals? Boesh and Tomasello (1998) suggested two: (a) the use of a complex language (b) the accumulation of cultural changes. Concerning the first feature, the authors stated that developed language allows further dissemination of cultural traits both in time and space and, possibly, the transmission of different types of information. Concerning the second, it is considered that humans have a considerable capacity to accumulate information and its modifications, making the human cultural system unique because it is progressively more complex (Boesh and Tomasello 1998; Mesoudi 2011). It is improbable that a single child or a group of isolated children would create a cultural system as complex as human society, because human cultures are historical products built over many generations.
7.5 Social Transmission and Cultural Evolution
From an evolutionary perspective, it is believed that the cognitive structure of the human mind was selected to permit cultural groups, thus making culture a human adaptation (Rogers 1988; Boyd and Richerson 2005; Mesoudi and Whiten 2008). Many arguments support this belief, such as humankind’s ecological success in different environments (Cavali-Sforza and Feldman 1981; Boyd and Richerson 1995; Richerson and Boyd 2005) and the capacity to accumulate adaptive information without individual production costs, increasing the fitness of the population (Rogers 1988; Boyd and Richerson 1995).
Historically, one of the most important studies on the role of social transmission in cultural evolution, which determined the future of this scientific field of investigation, was the work of Rogers (1988). His first major contribution was to evaluate the adaptability of information transmission and, therefore, its role in cultural evolution. Rogers (1988) assumed that natural selection tends to increase the average population fitness, otherwise there is no reason to believe that selection results in adaptation. In this way, if culture is adaptive, a population that transmits its information among its peers (recalling that the concept of culture equals the concept of social knowledge transmission) should have a better average fitness than a population with poor social transmission. In a variable environment, Rogers mathematically modeled a population in which individuals take two behavior types: (a) produce their own information (producers) or (b) copy it from their peers (social learners). In both situations, individuals will take trade off implications: energy consumption during knowledge production versus produced knowledge adaptability. Producers expend a lot of energy but the knowledge produced is highly adaptable to environmental changes, while social learners get their information at no additional cost but are susceptible to acquiring outdated or inappropriate information for a changing environment (Rendell et al. 2009).
Rogers’ (1988) model states that: (a) individuals who rely on their own knowledge fitness depend only on its production cost and benefits. They have no relation to their peers’ behavior which is, therefore, continuous; (b) when individuals who copy information are rare in a population, most pairs will behave as producers, which will offer low-cost, constantly adapted knowledge to social learners. In this situation, the average fitness of learners is greater than that of producers. When learners are the majority, little of the information available for copying will take into account recent environmental changes (i.e., adaptive changes). In this population configuration, the average fitness of producers will be higher. However, the most important point is that, regardless of the two strategies’ population frequency, the existing trade off between learners and producers creates a stable balance between their frequencies when the average fitness of these behaviors is the same. The first striking conclusion of Rogers (1988) is that a population with social learning does not have a higher average fitness than that of a population without social learning. Therefore, from an evolutionary point of view, cultural systems should not be expected if culture is understood only as social knowledge transmission. These notes do not say that culture is inherently adaptive; they only suggests that, in cases where it is, it must be because of properties not considered in the evaluated model. Due to the impact of Rogers’ work on the academic community, the author’s notes are known in scientific literature as “Rogers’ Paradox”.
Boyd and Richerson (1995) questioned whether the paradox was an artifact, due to the simplicity of Rogers’ (1988) basic model design. After all, the modeling considers only two behaviors (strategies): individual production (producers) and social knowledge transmission (learners), trying to adapt them closer to reality. The first model alteration, made by Boyd and Richerson (1995) was to model a population of individuals inserted into an environment that varies spatially. The second alteration allowed the modeled population of individuals to assume more than two behaviors. Finally, Boyd and Richerson (1995) adapted the model to include the possibility of individual producers that devised false or ill-adapted information. Despite these modifications, which made the model more realistic, Boyd and Richerson’s (1995) results corroborated Rogers’ model. Thus, inserting (a) an environmental variation and (b) other learning strategies, or (c) the existence of unprofitable information from an evolutionary point of view did not explain the existence of cultural systems, because, as had been seen in Rogers (1988), there was no increase in the average fitness of individuals who copy information from their peers.
In an attempt to find evolutionary explanations for culture, Boyd and Richerson (1995) constructed a mathematical model in which information transfer does not occur randomly, but selectively, and where individuals from the modeled group assess the behavior of peers and choose the one that appears to be the best. This type of transmission was named “biased transmission” and it happens when knowledge transfer follows certain trends, i.e., depending on some variable. This modification to the model, proposed by Boyd and Richerson (1995), assumes that if individuals are able to copy those peers recognized as the “best”, with the most advantageous information, the biased transmission would justify the existence of cultural systems because it would favor adaptive knowledge. However, this model analysis confirmed that this assumption is false, and therefore does not explain, from an evolutionary point of view, the advent of culture. Based on the above, Boyd and Richerson (1995) concluded that the ideas proposed by Rogers (1988) are mathematically robust.
How to resolve Rogers’ paradox then? How can information transfer, i.e., culture, be adaptive? It can be stated that culture increases the average fitness of a population and has an evolutionary justification if it increases the aptitude of learners and producers. As already revealed, when modeled by Rogers (1988), the presence of social transmission only optimized the average fitness of the individuals copying behavior, but not of the producers, whose fitness remained constant. When Richerson and Boyd (1995) modeled a situation in which the average fitness of individuals who produce their own knowledge increased as the population frequency of individuals who copy their peers increased, Roger’s paradox disappeared. In this sense, culture is adaptive only if (a) it makes individual learning less costly and (b) that learning becomes more accurate.
And how does culture perform these two elemental functions? First, the existence of social learning as a behavioral alternative to individual production allows producers to be selective. In this case, individuals can learn opportunistically, i.e., producing their own information when this strategy is more accurate and less costly, or in situations less favorable to production, choose to learn socially. The central question of this weighting is behavioral versatility, to what extent individuals are able to condition their behavior towards the reality in which they live. Second, culture allows information to be accumulated from generation to generation. Learning in small steps is less costly and more accurate than learning in large steps because, during complex knowledge construction, each required a small investment of energy . In every innovation event, new information is selected, creating adapted knowledge. Given the above, it is recognized that culture can play an adaptive role in the occurrence of additional processes, which are able to make social transmission more accurate and less costly.
However, from an evolutionary point of view an important question deserves an explanation: is knowledge transmission always accurate and does it favor useful traits? Certain authors believe the answer is “no,” stating that culture can set maladaptive traits (e.g., Boyd and Richerson 2005). Tanaka et al. (2009), mathematically modeled medicinal plants transmission and showed that a maladaptive character, in this case the absence of effective therapeutic resources, can remain in the population, because people may use it many times even though it is inefficient, allowing peers to copy and disseminate this behavior.
In summary, one can say that culture itself is not adaptive; in some situations it may even produce maladaptation. However, as stated earlier, there are procedures that make knowledge transmission an effective process from an evolutionary point of view. Enquist et al. (2007) stated that culture, to be adaptive, needs (a) information transfer to be faithful enough and (b) biases that favor adaptive traits and inhibit the perpetuation of maladaptive traits.
Given these observations, a better understanding of how to achieve advantages by copying information is necessary, assuming the possible risks of this learning strategy. Many studies have sought this understanding, and, according to Rendell et al. (2010), advances in this research area are due mainly to the union of theoretical and empirical studies. These advances have attempted to answer four basic questions: (a) “what is copied?” i.e., whether or not some information is favored during transmission; (b) “when is it copied?” i.e., what are the situations where individuals must choose to copy information; (c) “how is it copied?” which questions the copy, observation, education, and language mechanisms; and (d) “who to copy?” i.e., whether there is any trait that makes an individual a model to be copied (Laland 2004; Mesoudi and Whiten 2008). These questions are now discussed.
7.5.1 “Who to Copy?” Questions
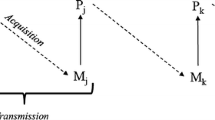
When copying, individuals must choose some of their peers as models and sources of knowledge. Strategies that identify the “who” show the characteristics of the selection process. Cavali-Sforza and Feldman (1981) initiated “who to copy” investigations, establishing a classification that became classic in cultural evolution studies, based on the degree of relatedness between model and learner. According to this classification, knowledge can be transferred culturally: (a) from parents to children (vertical type); (b) between same generation (horizontal type); (c) between generations, but only when young people copy adults who are not their parents (oblique type) (Fig. 7.1).

Social information transmission types. The large sphere represents a cultural system, such as a traditional community composed of different individuals (small spheres). Some of them are special because they are recognized as containing great knowledge and/or social prestige (centered white spheres). These are usually elders, healers, teachers, doctors and nurses. The tone distinction in the largest sphere seeks to represent age differences in individuals, i.e., distributing them in generations. Vertical transmission occurs within a family unit, while horizontal transmission occurs between unrelated pairs belonging to the same age group. When elders are the model to be copied, the transmission is called a “many-to-one”. In this case, the information always passes over a generation or more. The “one-to-many” transmission is modeled on a teacher, doctor or other agent with enough social prestige to influence many people
Hewlett and Cavali-Sforza (1986) re-structured the original Cavali-Sforza and Feldman (1981) classification, subdividing the “oblique” category into two types: (a) that of a teacher, leader or medium, such as radio or television directed towards many individuals in a group (one-to-many), which are generally pupils or learners; or (b) from older to new members in a social group (many-to-one or concerted) (Fig. 7.1). Although not explicit in Hewlett and Cavali-Sforza’s (1986) text, fission seems to be justified because these two new categories have distinct evolutionary implications that, when united in the “oblique” category, were indistinguishable.
Vertical transmission promotes a high knowledge variation between individuals in a group and between different groups. Parents-to-child transmission is highly conservative because it hinders the diffusion of innovation. As a result, the cultural evolution of a group that supports this type of strategy is rather slow (Fig. 7.2). On the other hand, systems that present knowledge transmission horizontally find that innovations are easily diffused and well developed. In these groups, knowledge variation between individuals and groups can be high and cultural evolution is very fast. “One-to-many” and “many-to-one” strategies have distinct evolutionary implications. In the first case, innovation diffusion occurs very easily. As a result of the knowledge homogenization process, knowledge between individuals in the group is very similar, although variation between groups is occasionally high. As in horizontal transmission, cultural evolution in groups using the “one-to-many” strategy is pretty quick. The second case, “many-to-one” type, is rather conservative, as elders’ values, traits and skills are transmitted on a large scale to other group members. This transmission form creates a situation that inhibits innovation diffusion and knowledge diversification within the group, as well as cultural evolution (Fig. 7.2).

The evolutionary implications of different social transmission strategies, adapted from Hewlett and Cavali-Sforza (1986)
The frequencies of each transfer pathway in a cultural system are not random and depend on many factors, especially the environmental and social background (McElreath and Strimling 2008; Reyes-Garcia et al 2009.). McElreath and Strimling (2008) concluded that copying parents is the most favorable situation for vertical type transmission. Using mathematical models, the authors stated that this social learning type should be more common in situations where the behavior is associated with fertility and care of their children and grandchildren than survival itself. Vertical transmission is favored in stable environments, as discussed earlier, and in situations where evolutionary pressures are very strong (McElreath and Strimling 2008). In contrast, horizontal transmission pathways and “one-to-many” strategies, which result in less conservative and more diffusive processes, are stimulated in varying environments (McElreath and Strimling 2008; Reyes-Garcia et al 2009).
Remember that information provided by the parental route has exactly the same nature of genetic transmission, because it shares the same common ancestor. Therefore, would it be possible to use the most modern tool of evolutionary biology, i.e., phylogeny, to investigate cultural evolution? Undoubtedly, yes, as argued by Boyd et al. (1997) and Mace (2005). Phylogeny, in short, seeks to understand evolutionary history and to establish historical relations between species, families and subfamilies. To do so it uses homologous characters, analyzing a set of shared characteristics among the organisms. Each of these has the same embryological origin and is present in all descendants of the same ancestral line. All phylogeny and evolutionary hypotheses on knowledge construction, often represented by phylogenetic trees or cladograms, are based solely on homologous characters. To have the same kind of genetic transmission, i.e., to share a common ancestor, the information transmitted by the parental route is recognized as homologous. In this sense, cultural systems where vertical transmission predominates should, therefore, adapt and be fully explained by phylogenetic models. In such cases, it is believed that cultural system evolution is determined by phylogeny events, i.e., conservative transmission (Fig. 7.3).

Schematic representation of two hypothetical social groups and their cladistics knowledge analyzes. In the first case, vertical transfer dominates (phylogeny events), and in the second case horizontal transfer, “many-to-one” and “one-to-many” are prominent (ethnogenesis events)
However, as stated by Mesoudi (2011), transmission types that diffuse horizontally, using “many-to-one” and “one-to-many” information do not present correspondents in biological evolution. These transmissions diffuse information beyond parental logic and in this way produce homoplastic similarities, i.e., shared characteristics. Unlike homologues they do not have a common ancestry. As phylogenic order is based on ancestry, homoplasy, by reflecting similarities of a nonhomologous nature, should invalidate phylogenetic tools used for cultural systems (Mesoudi et al 2006; Greenhill et al. 2009). However, despite this contrast, some studies show that horizontal transmission does not invalidate this evolutionary approach and its tools. For example, Greenhill et al. (2009) constructed a mathematical model to determine different diffusion level effects on the accuracy of phylogenetic estimates. The authors concluded that phylogenetic inferences are quite robust, even in situations influenced by high horizontal transmission levels. Collard et al. (2006) compared phylogenetic trees produced from biological and cultural information with trees models and concluded that these two databases produce similar patterns.
However, systems dominated by processes that spread information horizontally do not fit into phylogenetic models and, as a result, trees originating from these cultural systems do not exhibit the typical bifurcation structure (Fig. 7.3). Similarities between people and cultural groups that do not share a common ancestor are called, in the context of cultural evolution, ethnogenesis, which blurs phylogenetic analyzes (Fig. 7.3). Therefore, it is possible to evaluate the processes that underlie cultural evolution by assessing cultural information adequacy in a phylogenetic tree.
7.5.2 “When to Copy?” Questions
Laland (2004) made an ecological reading of social learning strategies and concluded that there are three ways for an individual to behave in a cultural system: (a) to ignore information (cultural traits); (b) to imitate their peers and copy the desired information; and finally, (c) to produce their own knowledge. In the first case, for example, the author considered the feeding behaviors of an animal that had no knowledge of the ways to obtain food. In this case, the animal was unaware of the necessary information to obtain the desired resource, but met his demands by depending on or looting his peers. In a second strategy, the animal acquired knowledge by observing or copying others from his group (cultural transmission). In the third and last option, self-production, where animals are seen as information producers, they construct new knowledge individually through their own methods, such as “trial and error”.
Laland (2004) constructed a hierarchical model of these strategies (Fig. 7.4) and argued that human and nonhuman animals preferably opt for the less costly strategy, following this order: not knowing, copy and produce. Therefore, behavior selection is determined by the strategy cost-benefit ratio, which in turn is determined by the particular environment in which the group is inserted (greatly or little productive) and the frequency of each strategy in the group (Laland 2004). The author assumes that individuals benefit by copying information, because by doing so, they will acquire valid information in a quick manner, freeing up knowledge and individual production costs. However, simply copying information is not the key to success, since this strategy requires that individuals feed the group with new information. In a group where nobody produces knowledge, copying information cannot be a profitable strategy (Laland 2004).

Hierarchical control of social learning strategies. Adapted from Laland (2004)
Using the ecological cost-benefit ratio, Laland (2004) tried to understand “when to copy” social strategies, seeking to specify circumstances that favor information diffusion. The author considered that individuals must first absorb knowledge from their peers when the lack of such information is unproductive, i.e., does not provide the necessary benefits. Otherwise, when the environment or social context is productive, individuals will preferentially adopt the most convenient strategy, unlearn information and rely on their peers for their basic needs. Seen in these terms, keeping in mind the context of traditional medicine, individuals will likely invest in knowledge production when there are no less costly alternatives to curing a disease available, for example, using traditional medicines prepared by others. According to Laland (2004), individuals from a social group will preferentially copy the information from their peers when the production of new knowledge is highly costly or dangerous. Producing knowledge that aims to solve problematic new situations demands a high investment of energy and thus individuals have a lower risk and little to lose when guided by others (Laland 2004).
Another situation where knowledge production can be highly costly occurs in unstable environments or when new situations are presented to the group (Laland 2004). In these situations, individuals may display conservative or indifferent behavior, depending on their peers’ action and knowledge, or they may rely on their own knowledge. For the latter alternative, there is the example of emergency foods (famine foods) where an information set about food plants is put into practice in adverse situations, such as when the environment presents some limiting situation (Nascimento et al. 2012).
Another context in which environmental change can play a key role in knowledge structuring is in instances of migration. With each new event the social group has new experiences, especially environmental knowledge. Individuals, therefore, will preferably ignore information or rely on their past knowledge of medicinal plants. But how does this second option take place, bearing in mind that, when migrating a new set of sources is available? The alternative would be to invest in knowledge of widely distributed cosmopolitan plants. Thus, migration events can influence the richness of known species in relation to their biogeographic origin (see Medeiros et al. 2012).
7.5.3 “What to Copy?” Questions
What kind or set of information must be copied into a cultural system? Possibly the most parsimonious strategy is to copy the majority, i.e., the most widespread behavior and knowledge in the social group (Laland 2004). This strategy is dependent on information distribution because the probability of an individual acquiring knowledge depends on his frequency within the cultural system (Henrich and Boyd 1998; Laland 2004). However, information copying may not only depend on the frequency but also on the content of the information. Nairne et al. (2008), for example, found that information related to survival is favored during memorization and, therefore, in information reproduction, what is known as adaptive bias. The human mind is predisposed to recall information related to subsistence because it confers greater adaptive advantage. Such predisposition is one of the best storage and decoding procedures identified in the human mind (Nairne et al. 2008).
7.5.4 “How to Copy?” Questions
According to Mesoudi and Whiten (2008), few studies have been concerned with understanding “how” information is transmitted within a group, especially when comparing different social learning processes. Available data suggests that botanical knowledge in a rural community is acquired through practice, as individuals perform daily activities (Zarger and Stepp 2004; Lozada et al 2006; Mesoudi and Whiten 2008; Reyes-Garcia et al 2009). Many authors claim that popular knowledge is transmitted mainly orally. However, it is necessary to investigate other transfer forms. Cordel literature, for example, is a widely known and widespread literary style in northeastern Brazil. Poems are presented in small booklets sold at fairs and streets, always fixed by strings (hence the “cordel” name). The booklets contain popular poems that record knowledge, practices, values, myths and legends of the popular universe. They are a prime example of knowledge transfer in writing. It is worth quoting verses from João Martins de Athayde, from in the booklet “Gypsy Esmeralda’s Testament”: “dream of peppermint, is a vain dream, it means that there are women, with voluminous hair, to arrest men, in a perfumed tie”.
7.6 Knowledge Transmission in Ethnobiology Studies
Most of the papers previously discussed address knowledge transmission from mathematical models or from empirical data of different types. However, social learning has also been the target of ethnobiological investigations (Frazão-Moreira 1997; Ladio and Lozada 2004; Garcia 2006; Lozada et al. 2006; Reyes-Garcia et al. 2009; Tehrani and Collard 2009). What are these studies’ contributions to our understanding of cultural evolution? Some studies simply focus on the description of a process, usually answering questions like “when,” “how” and from “whom” do you learn? Lozada et al. (2006), for example, described the transmission of knowledge about medicinal and food plants, focusing on the following aspects: when and where the informant-initiated plant resources were used, who taught him this knowledge, and where and how knowledge was taught. Other research, as well as describing knowledge transmission, tested hypotheses about transmission mechanisms. In two communities, one in Thailand and another in Argentina, Ladio and Lozada (2004) and Srithi et al. (2009) respectively tested whether knowledge erosion was due to information transmission failures. In principle, there are two approaches in transmission studies: process descriptions or process descriptions with hypothesis testing.
However, there are few examples of ethnobiological investigations that discuss their positions cultural evolution using the existing theoretical approach. A prime example is Garcia’s (2006) investigation of knowledge transmission and the use of wild foods among the Paniya. The author concluded that there is both positive and negative feedback in the transfer of this knowledge. Mothers teach why they think wild plants are “good” and “healthy foods,” but at the same time they attach a pejorative value to the value of these resources because, locally, they are associated with poverty. This is a trade off example, showing the conflict between biology and cultural spheres, which can be the cornerstone of a great evolutionary debate. What are the evolutionary dynamics of this knowledge that is both biologically basic (plants for food) and culturally disposable? Is there a tendency for this information to be perpetuated or will it be lost over time? Which sphere is most crucial in the transmission process, the biological demand or cultural values? However, despite the importance of feedback in knowledge transmission, no discussions were made from an evolutionary perspective.
Reyes-Garcia et al. (2009) moved in this direction, as they used existing theoretical approach to predict the cultural evolution of the group they studied. In that text, the authors showed that oblique transmission is very important among the Tsimane of the Bolivian Amazon. They commented: “… research suggests that oblique transmission, involving many demonstrators for a learner, tend to generate a high uniformity within a social group, while allowing cultural changes across generations. If, as data suggested, Tsimane favor oblique ways for cultural knowledge transmission, then a uniform change in Tsimane society is expected. Moreover, this growing oblique transmission dependence produces faster innovation diffusion in ethnobotanical knowledge [than]if vertical transmission was favored”. It is possible to do an exercise to recognize Reyes-Garcia et al.’s (2009) findings in Fig. 7.2. Another example is the Tanaka et al. (2009) study that, despite being merely mathematical, used transmission dynamics to discover why maladaptive traits remain in cultural systems. In summary, Tanaka et al. (2009) used prediction and transmission models to answer a real question with strong evolutionary implications.
It is not intended to disparage or discredit currently existing studies here. The intention is to point out that a big discussion about cultural evolution has not yet been incorporated into ethnobiological investigations. A theoretical approach, which goes beyond the knowledge transmission routes description, is necessary. An approach that includes, for example, the evolutionary implications of different learning strategies is needed.
7.7 Final Considerations
This chapter’s main objective was to characterize humans as beings who handle information from three different sources: genetic, individual learning and social transmission. Social transmission allows the establishment of a cultural system that, in humans, is established only through cognitive processes and achieves a greater degree of complexity. Therefore information transfer is a fundamental process to understanding human nature and cultural evolution, because this process allows traits diffusion and fixation through cultural selection.
By focusing on local knowledge as one a main area of study, ethnobiology can greatly help to clarify the dynamics of socialization and information, and consequently, of cultural evolution. However, as was noted above, this theoretical approach is still in its infancy. Today the existing research, despite describing the process, lacks a strong approximation of theories that explain the spatial and temporal variation of cultural traits. A program that guides investigative efforts in ethnobiology, building generalizations and theories about local knowledge transmission and its evolution is now necessary.
Notes
- 1.
Trade off, in an evolutionary context, refers to dilemma situations, and a beneficial alternative entails inherent losses. For example, it is important that a feline has the strength to hunt its food. But strength requires body mass. The bigger its body mass is, the greater the animal’s weight and, of course, the lower its ability to run fast. In this case, there is a trade off between strength and speed.
- 2.
Tabebuia roseoalba (Ridl.) Sandwith.
References
Boesh C, Tomasello M (1998) Chimpanzee and human cultures. Curr Anthropol 39:591–601
Boyd R, Richerson PJ (1995) Why does culture increase human adaptability? Ethol Sociobiol 16:125–143
Boyd R, Richerson PJ (2005) The origins and evolution of human culture. Oxford University Press, Oxford
Boyd R, Richerson PJ, Borgerhoff-Mulder M, Durham WH (1997) Are cultural phylogenies possible? In: Weingart P, Richerson PJ, Mitchell SD, Maasen S (eds) Human by nature, between biology and the social sciences. Lawrence Erlbaum Associates, New Jersey, pp 355–386
Cavali-Sforza LL, Feldman M (1981) Cultural transmission and evolution: a quantitative approach. Princeton University Press, Princeton
Collard M, Shennanb SJ, Tehrani JJ (2006) Branching, blending, and the evolution of cultural similarities and differences among human populations. Evol Hum Behav 27:169–184
Enquist M, Eriksson K, Ghirlanda S (2007) Critical social learning: a solution to Rogers’ Paradox of non adaptive culture. Am Anthropol 109:727–734
Frazão-Moreira A (1997) Meninos entre árvores e lianas – aprendizagem do mundo e das plantas pelas crianças Nalu (Guiné-Bissau). Educ Soc Cult 7:75–108
Garcia GSC (2006) The mother–child nexus: knowledge and valuation of wild food plants in Wayanad, Western Ghats, India. J Ethnobiol Ethnomed 2:39–45
Greenhill SJ, Currie TE, Gray RD (2009) Does horizontal transmission invalidate cultural phylogenies? Proc Biol Sci B 276:2299–2306
Henrich J, Boyd R (1998) The evolution of conformist transmission and the emergence of between-group differences. Evol Hum Behav 19:215–242
Hewlett BS, Cavali-Sforza LL (1986) Cultural transmission among Aka Pygmies. Am Anthropol 88:922–934
Heyes CM (1994) Social learning in animals: categories and mechanisms. Biol Rev 69:207–231
Ladio A, Lozada M (2004) Patterns of use and knowledge of wild edible plants in distinct ecological environments: a case study of a Mapuche community from northwestern Patagonia. Biodivers Conserv 13:1153–1173
Laland KN (2004) Social learning strategies. Learn Behav 32:4–14
Lozada M, Ladio A, Weigandt M (2006) Cultural transmission of ethnobotanical knowledge in a rural community of northwestern Patagonia, Argentina. Econ Bot 60:374–385
Mace R (2005) Introduction: a phylogenetic approach to the evolution of cultural diversity. In: Mace R, Holden CJ, Shennan S (eds) The evolution of cultural diversity: a phylogenetic approach. Leaf Coast Press, Walnut Creek, pp 1–12
Maturana RH, Varela FJ (2007) A árvore do conhecimento: as bases biológicas da compreensão humana, 6E. Palas Athena, São Paulo
McElreath R, Strimling P (2008) When natural selection favors imitation of parents. Curr Anthropol 49:307–316
Medeiros PM, Soldati GT, Alencar NL, Vandebroek I, Pieroni A, Hanazaki N, Albuquerque UP (2012) The Use of medicinal plants by migrant people: adaptation, maintenance, and replacement. Evid Based Complement Alternat Med 2012:807452
Mesoudi A (2007) A Darwinian theory of cultural evolution can promote an evolutionary synthesis for the social sciences. Biol Theory 2:263–275
Mesoudi A (2011) Cultural evolution: how Darwinian theory can explain human culture & synthesize the social sciences. University Chicago Press, Chicago
Mesoudi A, Whiten A (2008) The multiple roles of cultural transmission experiments in understanding human cultural evolution. Philos Trans R Soc Lond B Biol Sci 363:3489–3501
Mesoudi A, Whiten A, Laland K (2006) Towards a unified science of cultural Evolution. Behav Brain Sci 29:329–383
Nairne JS, Pandeirada JNS, Thompson SR (2008) Adaptive memory the comparative value of survival processing. Psychol Sci 19:176–180
Nascimento VT, Vasconcelos MAS, Maciel MIS, Albuquerque UP (2012) Famine foods of Brazil seasonal dry forests: Ethnobotanical and nutritional aspects. Econ Bot 66:22–34
Rendell L, Boyd R, Cownden D, Enquist M, Eriksson K, Feldman MW, Fogarty L, Ghirlanda ST, Lillicrap T, Laland KN (2009) Why Copy Others? Insights from the social learning strategies tournament. Science 328:1–6
Rendell L, Forgaty L, Laland KN (2010) Rogers’ paradox recast and resolved: population structure and the evolution of social learning strategies. Evolution 64:534–548
Reyes-Garcia V, Molina JL, Broesch J, Calvet L, Fuentes-Pelaez N, McDade TW, Parsa S, Tanner S, Huanca T, Leonard WR, Martinez-Rodriguez MR (2009) Cultural transmission of ethnobotanical knowledge and skills: an empirical analysis from an Amerindian society. Evol Hum Behav 30:1–12
Richerson PJ, Boyd R (2005) Not by genes alone: how culture transformed human evolution. The University of Chicago Press, Chicago
Rogers AR (1988) Does biology constrain culture? Am Anthropol 90:819–831
Srithi K, Balslevb H, Wangpakapattanawonga P, Srisangac P, Trisonth C (2009) Medicinal plant knowledge and its erosion among the Mien (Yao) in northern Thailand. J Ethnopharmacol 123:335–342
Tanaka MM, Kendal JR, Laland KN (2009) From Traditional Medicine to Witchcraft: Why medical treatments are not always efficacious. PLoS One 4:5192–5201
Tehrani JJ, Collard M (2009) On the relationship between interindividual cultural transmission and population-level cultural diversity: a case study of weaving in Iranian tribal populations. Evol Hum Behav 30:286–300
Zarger RK, Stepp JR (2004) Persistence of Botanical Knowledge among Tzeltal Maya Children. Curr Anthropol 45:413–419
Acknowledgements
I thank Dr. Ulysses Paulino Albuquerque for his helpful comments on the several versions of this chapter.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Soldati, G.T. (2015). Knowledge Transmission: The Social Origin of Information and Cultural Evolution. In: Albuquerque, U., De Medeiros, P., Casas, A. (eds) Evolutionary Ethnobiology. Springer, Cham. https://doi.org/10.1007/978-3-319-19917-7_7
Download citation
DOI: https://doi.org/10.1007/978-3-319-19917-7_7
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-19916-0
Online ISBN: 978-3-319-19917-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)