
Abstract
This paper describes the discovery of new, exceptionally well preserved Late Jurassic (Late Tithonian) horseshoe crabs belonging to the genus Limulus, which are considered to be the oldest known representatives of the genus. The three-dimensionally preserved fossil horseshoe crabs, belonging to two species: Limulus darwini and Crenatolimulus sp. nov., recently were discovered in the Owadów-Brzezinki Quarry in Central Poland. The state of preservation of the fossil horseshoe crabs is described and the evolutionary lineage of both extinct and extant limulids is discussed. The palaeoenvironment inhabited by the new species and the trophic relationships of living horseshoe crabs are investigated in an attempt to determine the potential relationships between Mesozoic and recent xiphosurans.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
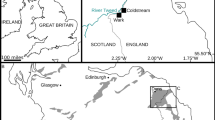
The recently discovered paleontological site of the Sławno limestones at the Owadów-Brzezinki Quarry in Central Poland (Fig. 1.1) has yielded exceptionally well-preserved fossils of Late Jurassic horseshoe crabs. The discovery of new, more or less three-dimensionally preserved Late Jurassic Xiphosurida arthropods adds significantly to our understanding of a group that has a stratigraphic range throughout almost the entire Phanerozoic period.

Road map with the location of Owadów-Brzezinki Quarry and its proximity to Tomaszów Mazowiecki in Central Poland
The uniqueness of these new finds lies in their very close stratigraphical and geological relationship to one of the most famous Fossil-Lagerstätte localities in the world – Solnhofen-Eichstätt and Nusplingen, in southern Germany, where the best-known genus of the Jurassic horseshoe crab, Mesolimulus, is found. Horseshoe crabs lived during the Late Jurassic at Nusplingen (Late Kimmeridgian, Beckeri Zone), Solnhofen-Eichstätt (Early Tithonian, Hybonotum Zone) and in Owadów-Brzezinki (Late Tithonian = Middle Volgian, Zarajskensis Subzone of the Scythicus Zone), under very similar environmental conditions. As such, it presents an unrivalled opportunity for studying their evolution and rate of speciation.
2 Geological and Paleontological Settings
The exposed carbonate sequence of the Sławno limestones belongs to the Kcynia Formation (Kutek 1994) and is divided into four successive units. In general, first three units (I, II and III) appear to represent a continuous transition from an offshore to nearshore, perhaps lagoonal settings, whereas unit IV shows evidence of a return to more open marine conditions. The Kcynia Formation is underlain by yellowish marls and marly clays of the Pałuki Formation.
The lowermost part of the unit III (Fig. 1.2) is very fossiliferous. Numerous specimens of horseshoe crabs were found in association with an enormously rich assemblage of the soft-shelled bivalves Corbulomima sp., remains of various fish and marine reptiles, rare ammonites, decapod crustaceans (Feldmann et al. 2015), land insects (dragonflies, beetles, grasshoppers) and pterosaurs (Kin and Błażejowski 2012; Kin et al. 2012, 2013). There are many similarities between the Owadów-Brzezinki and Solnhofen-Nusplingen faunas, which allow for comparative paleontological studies at a previously unattainable level of taxonomic resolution. The Owadów-Brzezinki Quarry may be regarded as a new ‘taphonomic window’ into the living world of the latest Jurassic, being the first near-contemporary paleontological ‘supplement’ to a previously known Fossil-Lagerstätten.

Panoramic view of the highest level of exploitation in Owadów-Brzezinki Quarry, Poland (i.e. unit III and most fossiliferous ‘Corbulomima horizon’ occurring in the middle of the quarry wall)
3 Materials and Methods
The studied fossils of horseshoe crabs were collected by the author during field work in the Owadów-Brzezinki Quarry (lat. 51°37.4238′, lon. 20°13.6343′) in 2012–2014. The collected materials are housed at the Institute of Paleobiology, Polish Academy of Science in Warsaw (ZPAL X.1/O-B).
4 The Finds of Late Jurassic Limulus Fossils
After 6 years of field work, limestones of this unit III have yielded more than 150 well-preserved specimens of horseshoe crabs, including 11 almost completely articulated exoskeletons (Figs. 1.3 and 1.4). Tables of biometric measurements of selected specimens Limulus darwini are given in Kin and Błażejowski (2014). The preservation of ventral side in some specimens (Fig. 1.5) is one of the most promising aspects of the limuline material from Owadów-Brzezinki. This material is currently the subject of detailed study and will undoubtedly provide new information on the evolution of the horseshoe crabs.

(a) Late Jurassic horseshoe crab species Limulus darwini, Owadów-Brzezinki, Poland (holotype ZPAL X.1/O-B/XA 1, Kin and Błażejowski 2014), (b) juvenile specimen of the extant Atlantic horseshoe crab species Limulus polyphemus, USA (ZPAL X.1/JB-NY)

Three-dimensionally preserved representatives of Late Jurassic horseshoe crabs from Owadów-Brzezinki, Poland. Crenatolimulus sp. nov. (a) Negative (external mold) and (b) positive (rock slab with imprint) (ZPAL X.1/O-B/14.1), (c) Crenatolimulus sp. nov. (ZPAL X.1/O-B/XA 13.B), (d) Limulus darwini prosoma with exceptionally well-preserved left compound eye (ZPAL X.1/O-B/14.2), (e) nearly complete exuvia of Crenatolimulus sp. nov. (ZPAL X.1/O-B/XAC 5n) (f) Crenatolimulus sp. nov. (ZPAL X.1/O-B/XAC 2), no telson preserved, exhibits depressed and beaded occipital band

The ventral side of the late Jurassic horseshoe crab form Owadów-Brzezinki, Poland (ZPAL X.1/O-B/XA 14.4)
The ventral presentation of the fossil limuline provides an opportunity to study the appendages and mouthparts, and comparison of these extant forms will probably allow us to determine the evolutionary pathways of these features in far more detail than previously.
The vast majority of these finds are incomplete; they are preserved as isolated fragments of the prosoma and opisthosoma, and represent all four stages of disarticulation (Fig. 1.6) as observed by Babcock et al. (2000). Specimens of L. darwini (Kin and Błażejowski 2014) constitute the first Late Jurassic representatives of the order Xiphosurida to be recorded from Poland. Previously, the only known limulid was a single specimen of lower Pliensbachian Limulitella cf. liasokeuperinus (Braun 1860) from a brackish-marine stratum of the Gielniów Formation at Skarżysko-Kamienna, south-central Poland (Karaszewski 1975). This particular specimen was lost during World War II. Because extinct horseshoe crabs are extremely rare in the fossil record, it is virtually impossible to establish an exact phylogenetic relationship between extinct and extant families and genera.

Late Jurassic horseshoe crabs from Owadów-Brzezinki, Poland; different stages of disarticulation (ZPAL X.1/O-B/XAC 8.3)
After a detailed analysis of three-dimensionally preserved Late Jurassic limulids from Owadów-Brzezinki, Kin and Błażejowski (2014) showed that there are no significant morphological differences between specimens from Owadów-Brzezinki and extant juvenile individuals of the genus Limulus (Müller 1785). Clearly, modern horseshoe crabs of that genus are more closely related to Late Jurassic forms than previously thought (Riek and Gill 1971; Sekiguchi and Shuster 2009) (Fig. 1.3). Indeed, the morphological features of the opisthosoma of Late Jurassic Limulus from Poland are very similar to the holotype of Limulus coffini (Reeside and Harris 1952) from the Upper Cretaceous of Colorado. This clearly indicates a close relationship, while L. darwini is almost certainly an early link both with L. coffini and the extant Limulus polyphemus (Linnaeus 1758). Thus, the genus Limulus existed at least 148 million years ago and has survived to the present day in an almost unchanged form (Kin and Błażejowski 2014).
5 Comparison of Late Jurassic Limulus with Extinct and Extant Limulines
Much of the general paleontological background to the horseshoe crabs may be found in several earlier publications, especially in Anderson and Selden (1997), Anderson and Shuster (2003), and Rudkin and Young (2009), and numerous references cited therein.
The fossil record of the basic xiphosurid horseshoe crab body plan certainly extends back as far as the Ordovician, which indicates an origin which lies outside of the paraphyletic ‘synziphosurine’ (Rudkin and Young 2009). Thus Xiphosurans have existed for some 480 Mya (Lamsdell 2013), with the earliest unequivocal representatives found in the Upper Ordovician of Manitoba, Canada (Rudkin et al. 2008), followed by further Xiphosurida finds from the Lower Ordovician of Morocco (Van Roy et al. 2010). Fossil species from the earliest Palaeozoic (Cambro-Silurian) have a segmented opisthosoma characteristic of the order Synziphosurida, whereas, for the Xiphosurida, the midpart of the body was essentially a solid piece of fused segments.
Of all known extinct Mesozoic and Cenozoic horseshoe crabs (Størmer 1952; Kin and Błażejowski 2014) Limulus darwini seems to be almost identical with the previously mentioned Late Jurassic limuline from Germany (Hauschke and Wilde 1991) and unusually well preserved early Cretaceous limulines from the Glen Rose Formation, north-central Texas (Feldmann et al. 2011).
Certainly the new species of Late Jurassic horseshoe crabs presented here shows some features in common with representatives of the genus Mesolimulus, in particular with M. walchi (Desmarest 1822) from the late Kimmeridgian pelitic limestones of Brunn (Röper et al. 1996) and Nusplingen (Schweigert and Dietl 2002; Briggs et al. 2005), and the early Tithonian lithographic limestones of Solnhofen (Münster 1840; Frickhinger 1994). A narrower furrow along the inner margin of the genal angles on the prosoma and the presence of short articular processes with an absence of distinct dorsal furrows on the opisthosoma are the main features that distinguish Limulus darwini from M. walchi (Kin and Błażejowski 2014). However, the material from Germany gives no clues as to whether these differences are merely a function of the state of preservation of the significantly less compressed L. darwini specimens. In respect to some similarities to the extant genus Tachypleus, the Jurassic Mesolimulus walchi is referenced by many researchers as Mesotachypleus (Anderson and Shuster 2003).
The most important morphological features that distinguish the Lower Cretaceous (Albian) Crenatolimulus paluxyensis (Feldman et al. 2011) from Limulus darwini (Kin and Błażejowski 2014) and all other post-Palaeozoic horseshoe crabs are the beaded rims of the posterior region on the prosoma and flanks with two prominent ridges defining a tripartite surface on the opisthosoma. The other exoskeleton features are generally similar to L. darwini, with the particular exception of the proportionally much longer genal spines on the prosoma and the clearly pronounced axial ridge on the opisthosoma (Kin and Błażejowski 2014). Significantly, as well as L. darwini, other specimens belonging to the genus Crenatolimulus sp. nov. (Figs. 1.4 and 1.6) have recently been found. These are the first records of the genus in Europe. They obviously represent a species new to science and are the subject of ongoing studies.
With respect to the abdominal morphology, it is important to compare L. darwini with another extinct horseshoe crab Limulus coffini, which has a three-dimensionally preserved complete opisthosoma. Both show similar morphological characteristics. Among the most important features are six fixed spines terminated posteriorly by two large marginal spines occurring on both sides of the relatively deep and wide posterior margin, six distinct apodomes on either side of the axis and large muscle scars separating anterior and posterior opisthosomal parts. The morphological features that distinguish L. darwini from L. coffini are the lack of a pronounced axial ridge, the occurrence of three (not two) tubercles along the axial ridge and slightly larger articular processes (Kin and Błażejowski 2014).
The separation of the larger anterior part and abdominal posterior part of the opisthosoma in L. darwini is similar in comparably sized individuals, i.e. juveniles of L. polyphemus (compare Babcock et al. 2000). Other similar morphological features include the median cardiac node and the shape of the occipital bands on the prosoma and the size and shape of the articular processes on the opisthosoma (Kin and Błażejowski 2014).
Unfortunately, there are no specimens comparable in size with mature individuals of L. polyphemus in the recent collection from Owadów-Brzezinki, and the absence of fully-grown exoskeletons known from the late Jurassic remains unexplained.
According to Barthel (1974), all specimens of M. walchi were presumably exoskeletons (molts) of only young animals. The finds of giant limulid trackways (i.e. Kouphichnium lithographicum, Oppel 1862) in the Upper Jurassic of Germany (Kolb 1963; Frickhinger 1994; Schweigert 1998) and France (Gaillard 2010), which are attributed to much larger individuals, seem to support an assumption that only juveniles can be preserved in the fossil state. This puzzling feature could possibly be explained by the fact that subadult horseshoe crabs molted less often than juveniles, as is the case in modern limulids (Barthel 1974; Carmichael et al. 2003; Chiu and Morton 2004). Shuster and Sekiguchi (2003) suggest that Limulus polyphemus takes 9–11 years and 19 or fewer molts to reach maturity. The shallow water in which the Owadów-Brzezinki sediments accumulated may thus be interpreted a “limulid nursery,” where young individuals spend their first years rapidly increasing its size, prior to venturing out into deeper waters (Carmichael et al. 2003; Chiu and Morton 2004).
The poor fossil record is, of course, the main obstacle for clear understanding of how exactly the living horseshoe crabs derived from Mesozoic lineages (Fisher 1982, 1984). Molecular studies on extant population (Avise et al. 1994; Xia 2000) have provided some clues to their genealogical relationships, but many unsolved questions still concern their patchy distribution.
6 Summary and Conclusions
Because Xiphosurida arthropods are extremely rare in the fossil record, the recent finds have an exceptional importance. It is, indeed, particularly fortunate that the horseshoe crabs are found in such an excellent state of preservation. Furthermore, L. darwini from Owadów-Brzezinki is regarded as the oldest known representative of the genus (Kin and Błażejowski 2014). Possibly, these new discoveries have only a limited bearing on genealogical relationships within the group, but they certainly throw a great deal of new light on several aspects of evolution and adaptation processes. Evidently, this extraordinary group of organisms has survived more than one major extinction event and global biotic crises (Raup and Sepkoski 1982) in the history of our planet, including P/Tr (Permian–Triassic) when trilobites became extinct, and K/T (Cretaceous–Tertiary) which 65 million years ago turned dinosaurs and ammonites into museum exhibits. All horseshoe crabs found in Owadów-Brzezinki were located in strata with abundant soft-shelled bivalves, within so-called ‘Corbulomima horizon’, what may be related to their potential feeding habits. The diet of modern Atlantic Limulus polyphemus is highly diverse and consists of variety of small marine fauna including bivalves (Smith 1953; Botton 1984). According to Botton and Ropes (1989), bivalves of the genus Corbula form part of the diet of L. polyphemus, and the prolific Corbulomima sp. at Owadów-Brzezinki acted as possible food attractor for L. darwini. Thus the depositional area of ‘Corbulomima horizon’ could have been both “nursery” and feeding zone for L. darwini.
Kin and Błażejowski (2014) suggested that the relative evolutionary conservatism noticeable in Limulus (and, presumably, all known representatives of Xiphosurida), may be related to a unique formula of adaptative strategy. In order to systematize this phenomenon, Kin and Błażejowski (2014) introduced the term ‘stabilomorphism’, which is a relative morphological stability of organisms in time and spatial distribution, the taxonomic status of which does not exceed genus level. The definition refers exclusively to genera that have survived at least one of the great mass extinctions. By this definition, the resulting morphological structure of the presumed stabilomorph must have been virtually unchanged for more than 65 million years (e.g. Limulus). A high effectiveness of adaptation significantly reduces the need for differentiated phenotypic variants in response to environmental changes and provides for long-term evolutionary success (Kin and Błażejowski 2014).
References
Anderson LI, Selden PA (1997) Opisthosomal fusion and phylogeny of Palaeozoic Xiphosura. Lethaia 30:19–31
Anderson LI, Shuster CN Jr (2003) Throughout geologic time: where have they lived? In: Shuster CN, Barlow RB, Brockman HJ (eds) The American horseshoe crab. Harvard University Press, Cambridge, MA, pp 189–224
Avise JC, Nelson WS, Sugita H (1994) A speciational history of “living fossils”: molecular evolutionary patterns in horseshoe crabs. Evolution 48:1986–2001
Babcock LE, Merriam DF, West RR (2000) Paleolimulus, an early limuline (Xiphosurida), from Pennsylvanian—Permian Lagerstatten of Kansas and taphonomic comparison with modern Limulus. Lethaia 33:129–141
Barthel KW (1974) Limulus: a living fossil. Horseshoe crabs and interpretation of an Upper Jurassic environment (Solnhofen). Nature 61(10):428–433
Botton ML (1984) Importance of predation by horseshoe crabs, Limulus polyphemus, to an intertidal sand flat community. J Mar Res 42:139–161
Botton ML, Ropes JW (1989) Feeding ecology of horseshoe crabs on the continental shelf, New Jersey to North Carolina. Bull Mar Sci 45:637–647
Braun CF (1860) Die Thiere in den Pflanzenschiefern der Gegend von Bayreuth. 1771, Zool. Fundamenta
Briggs DEG, Moore RA, Shultz JW et al (2005) Mineralization of soft–part anatomy and invading microbes in the horseshoe crab Mesolimulus from the Upper Jurassic Lagerstatte of Nusplingen, Germany. Proc R Soc B Biol Sci 272:627–632
Carmichael RH, Rutecki D, Valiela I (2003) Abundance and population structure of the Atlantic horseshoe crab Limulus polyphemus in Pleasant Bay, Cape Cod. Mar Ecol Prog Ser 246:225–239
Chiu HMC, Morton B (2004) The behaviour of juvenile horseshoe crabs, Tachypleus tridentatus (Xiphosura), on a nursery beach at Shui Hau Wan, Hong Kong. Hydrobiologia 523(1–3):29–35
Desmarest AG (1822) Les Crustacés prominent dits. In: Brongniart A, Desmarest AG (eds) Historie naturelle des crustacés, sous les rapports zoologiques. Chez F.G. Levrault Libraire, Strasbourg, pp 67–142
Feldmann RM, Schweitzer CE, Dattilo B et al (2011) Remarkable preservation of a new genus and species of limuline horseshoe crab from the Cretaceous of Texas, USA. Palaeontology 54(6):1337–1346
Feldmann RM, Schweitzer CE, Błażejowski B (2015) A new species of lobster (Glypheoidea: Mecochiridae) from the Late Jurassic (late Tithonian) Lagerstätte from central Poland. N Jahrb Geol Paläont Abh 275:107–114
Fisher DC (1982) Phylogenetic and macroevolutionary patterns within the Xiphosurida. Proc N Am Paleontol Convent 1:175–180
Fisher D (1984) The Xiphosurida: archetypes of bradytely? In: Eldredge N, Stanley SM (eds) Living fossils. Springer, New York, pp 196–213
Frickhinger KA (1994) The fossils of Solnhofen. Korb Goldschneck-Verlag, Ostfildern, pp 1–336
Gaillard C (2010) A giant limulid trackway (Kouphichnium lithographicum) from the lithographic limestones of Cerin (Late Kimmeridgian, France): ethological and environmental implications. Swiss J Geosci 103:1–16
Hauschke N, Wilde V (1991) Zur Verbreitung und Okologiemesozoischer Limuliden. N Jahrb Geol Paläon Abh 183:391–411
Karaszewski W (1975) Obecność rzadkiej skamieniałości–limulusa w liasie świętokrzyskim. Prz Geol 23(10):500–501. [in Polish]
Kin A, Błażejowski B (2012) Polskie Solnhofen. Prz Geol 60(7):375–379. [in Polish]
Kin A, Błażejowski B (2014) The horseshoe crab of the genus Limulus: living fossil or stabilomorph? PLoS One 9(10):e108036. doi:10.1371/journal.pone.0108036
Kin A, Błażejowski B, Binkowski M (2012) ‘Polish Solnhofen’: a long-awaited alternative? Geol Today 28(3):91–94
Kin A, Gruszczyński M, Martill D et al (2013) The paleoenvironment and taphonomy of a Late Jurassic (Late Tithonian) Lagerstädtte from central Poland. Lethaia 46:71–81
Kolb A (1963) Riesige Limulus-Fährte aus den lithographischen Schiefern bei Pfalzpaint. Geol Blätter Nordost-Bayern 13:73–78
Kutek J (1994) The Scythicus zone (Middle Volgian) in Poland: its ammonites and biostratigraphic subdivisions. Acta Geol Pol 44:1–33
Lamsdell JC (2013) Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura. Zool J Linn Soc 167:1–27
Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, vol 1, Regnum animale. Editio decima, reformata. Laurentii Salvii, Stockholm
Müller OF (1785) Entomostraca seu Insecta Testacea. Lipsiae et Hafniae, pp 1–135
Münster G (1840) Über die fossilen Arten Limulus in den lithographischen Schiefern von Bayern. In: Münster G (ed) Beiträge zur Petrefacten-Kunde 3:26–27
Oppel A (1862) Ueber Fährten lithographischen Schiefer (Ichnites lithographicus). Paläeontologische Mitteilungen aus dem Museum des Koeniglich Bayerischen Staates 1, pp 121–125
Raup DM, Sepkoski JJ Jr (1982) Mass extinctions in the marine fossil record. Science 215:1501–1503
Reeside JB Jr, Harris DV (1952) A Cretaceous horseshoe crab from Colorado. J Wash Acad Sci 42(6):174–178
Riek EF, Gill ED (1971) A new xiphosuran genus from lower cretaceous freshwater sediments at Koonwarra, Victoria, Australia. Paleontology 14(2):206–210
Röper M, Rothgaenger M, Rothgaenger K (1996) Die Plattenkalke von Brunn (Landkreis Regensburg). Eichendorf Verlag, Eichendorf, pp 1–102
Rudkin DM, Young GA (2009) Horseshoe crabs – an ancient ancestry revealed. In: Tanacredi JT, Botton M, Smith DR (eds) Biology and conservation of horseshoe crabs. Springer, New York, pp 25–44
Rudkin DM, Young GA, Nowlan GS (2008) The oldest horseshoe crab: a new xiphosurid from late Ordovician konservat-lagerstätten deposits, Manitoba, Canada. Palaeontology 51:1–9
Schweigert G (1998) Die Spurenfauna des Nusplinger Plattenkalks (Oberjura, Schwäbische Alb). Stuttg Beitr Nat B 262:1–47
Schweigert G, Dietl G (2002) Miscellanea aus dem Nusplinger Plattenkalk (Ober-Kimmeridgian, Schwäbische Alb), 4. Limuliden Jber Mitt Oberrhein Geol Ver NF 84:323–331
Sekiguchi K, Shuster CN Jr (2009) Limits on the global distribution of horseshoe crab (Limulacea): lessons learned from two lifetimes of observations: Asia and America. In: Tanacredi JT, Botton ML, Smith DR (eds) Biology and conservation of horseshoe crabs. Springer, New York, pp 5–24
Shuster CN Jr, Sekiguchi K (2003) Growing up takes about ten years and eighteen stages. In: Shuster CN Jr, Barlow RB, Brockmann HJ (eds) The American horseshoe crab. Harvard University Press, Cambridge, MA, pp 103–133
Smith OR (1953) Notes on the ability of the horseshoe crab Limulus polyphemus to locate soft-shell clams Mya arenaria. Ecology 34:636–637
Størmer L (1952) Phylogeny and taxonomy of fossil horseshoe crabs. J Paleontol 26:630–640
Van Roy P, Orr PJ, Botting JP et al (2010) Ordovician faunas of Burgess shale type. Nature 465:215–218
Xia X (2000) Phylogenetic relationship among horseshoe crab species: effect of substitution models on phylogenetic analyses. Syst Biol 49:87–100
Acknowledgements
I would like to sincerely thank Peter Walsh (University of Silesia) for many useful suggestions and correction of English. I appreciate the help provided in the field of Robert Siuda (Owadów-Brzezinki Quarry, Nordkalk Company). And last but not least, I wish to acknowledge Kevin Laurie (independent horseshoe crab researcher, Hong Kong), Mark Botton (Department of Natural Sciences, Fordham College at Lincoln Center, New York) and one anonymous reviewer for his critical review and very helpful comments that improved the manuscript. This work was supported by the Polish National Science Centre (grant no.2012/07/B/ST10/04175).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Błażejowski, B. (2015). The Oldest Species of the Genus Limulus from the Late Jurassic of Poland. In: Carmichael, R., Botton, M., Shin, P., Cheung, S. (eds) Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Springer, Cham. https://doi.org/10.1007/978-3-319-19542-1_1
Download citation
DOI: https://doi.org/10.1007/978-3-319-19542-1_1
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-19541-4
Online ISBN: 978-3-319-19542-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)