
Abstract
Long-term agricultural research sites like the Wisconsin Integrated Cropping Systems Trial (WICST), with annual and perennial cropping systems growing alongside rotationally grazed pasture and native prairie plantings, provide invaluable insight into the carbon sequestration potential of agriculture in the North Central USA. Analysis of soil organic C (SOC) stocks during the first 20 years of the WICST study showed that annual row crop agriculture lost 5.5 Mg C ha−1, while rotationally grazed pasture was a significant sink to 0–60 cm depth (5.1 Mg ha−1). While SOC changes for the Prairie, Conservation Reserve Program planting (CRP), and forage systems were not significantly different from zero when considered independently of one another, differences between systems suggest a trend of SOC accumulation in Prairie and loss in the CRP and forage systems. Carbon stabilization and accretion on the fine textured Mollisols of the North Central U.S. may not be possible, even under agricultural best management practices, without the inclusion of perennial grasses.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
Introduction
Agricultural lands have been promoted as promising and economically viable sinks for atmospheric C (Conant et al. 2007; Lal 2008). Lal (2004) suggested that the carbon sequestration potential of agricultural soils for the near future (25–50 years) would be modest (50–100 Pg C) but important as it is a functional reservoir for CO2. Practices such as no-till farming (NT) (West and Post 2002; Huggins et al. 2007), nitrogen fertilization (Huggins et al. 1998; Nafziger and Dunker 2011), animal and green manures (Ogle et al. 2005), and increased use of perennials crops (Huggins et al. 1998; West and Post 2002) have been suggested as ways to manage agricultural lands for carbon storage. There is general agreement that such practices have beneficial effects on soil carbon status, but long-term soil organic carbon (SOC) stabilization is dependent on soil texture, mineralogy, and the environment (Kögel-Knabner et al. 2008; Fierer et al. 2009; Schmidt et al. 2011) making it unlikely that a standard set of farming practices will have the same beneficial effects when adopted under diverse edaphic conditions.
There is a growing body of scientific literature that offers contradictory results related to SOC sequestration under agronomic best management practices (Huggins et al. 2007; Luo et al. 2010; Sanford et al. 2012). Huggins et al. (2007), in a study of tillage and crop rotations effects on SOC in southern Minnesota, report losses of 3.7 and 1.6 Mg SOC ha−1 for low productivity or aggressively tilled systems and high productivity or minimally tilled systems respectively. They concluded that under the best case scenario of highly productive continuous corn and reduced tillage, stabilization of initial SOC would require reducing decay rates by over 50 % or doubling C inputs. These findings are supported by those of Sanford et al. (2012) who showed 20 year losses of SOC in six common Wisconsin agroecosystems despite the use of agricultural best management practices (NT, perennials, manure). Luo et al. (2010) in a meta-analysis of 69 paired experiments found that conversion from conventionally tilled to no-till farming practices affected the distribution, but not the concentration of SOC (>40 cm) under similar crop rotations.
The Mollisols of the North Central USA account for 85 % of national corn production, 25 % of the nation’s SOC stocks (Guo et al. 2006; USDA 2013), and figure prominently in efforts to re-claim historically depleted SOC reserves. A potential management strategy for SOC accretion, that does not rely upon annual row crops or perennial forage rotations, is the re-establishment of grasslands and native prairie vegetation for agricultural production (forage and biomass). This strategy is gaining traction as the benefits of rotational grazing are increasingly recognized (Follett and Reed 2010; Oates et al. 2011) and perennial grasses such as switchgrass (Panicum virgatum L.), big bluestem (Andropogon gerardii Vitman) and giant miscanthus (Miscanthus x giganteus Greef & Deuter ex Hodkinson & Renvoize) are promoted for bioenergy production (Heaton et al. 2008; Hong et al. 2013).
Long-term agricultural research sites like the Wisconsin Integrated Cropping Systems Trial (WICST), with annual and perennial cropping systems growing alongside rotationally grazed pasture and native prairie plantings, provide insight into the carbon sequestration potential of diverse agroecosystems. Recent work at the WICST demonstrated the importance of pasture grasses in stabilizing SOC in the 0–30 cm soil depth and the general loss of SOC in well managed annual and perennial crop rotations (Sanford et al. 2012). To better understand the importance of perennial grass systems in the SOC dynamics of fine textured Mollisols, two native species plantings were compared to the three general land cover groups (LCGs) of the WICST core trial (grain, forage, and pasture). These native plantings included a native prairie restoration (Prairie) typical of the vegetation historically found in southern Wisconsin, and a lower diversity native plant mixture promoted through the USDA’s Conservation Reserve Program (CRP) for the stabilization of highly erodible land.
Methods
The Wisconsin Integrated Cropping System Trial (WICST) was initiated in 1989 at the University of Wisconsin Arlington Agricultural Research Station (43°18′N, 89°20′W), Arlington, WI, USA. Six cropping systems are present in the core trial: three cash grain, two crop based dairy forage rotations, and one pasture based dairy system (Posner et al. 1995). In 1989 a cool season grass mix (Kentucky Bluegrass [Poa pratensis L.], Quackgrass [Elymus repens L. Gould], Smooth Bromegrass [Bromus inermis Leyss.]) was planted at WICST in 1.2 ha not occupied by the core trial. In 1998, the cool season grasses were plowed under and the site was planted to soybeans. The following growing season two native perennial systems were established. The two systems were (1) a native prairie planting (Prairie: 25 species mix) and (2) a planting mix typical of those used in the Conservation Reserve Program (CRP: 6 species mix). Further establishment details can be found in Simonsen et al. 2002.
Soil sampling and SOC determination (g SOC [kg soil]−1 and Mg SOC ha−1) followed the same protocols as those outlined in Sanford et al. (2012), as follows: two time points [1989 & 2009]; 4 soil depths [0–15, 15–30, 30–60, and 60–90 cm]; and six cores/depth/plot including calculations of equivalent soils mass (Lee et al. 2009). For the two native plantings (Prairie & CRP) an additional set of soil samples were collected (0–15 and 15–30 cm only) and analyzed at the time of system establishment (1999).
Analyses were conducted to compare changes in SOC (g kg−1 and Mg ha−1) between the three LCGs from the WICST core trial (grain, forage, pasture) and the two native species plantings (CRP, prairie). The five LCGs analyzed were therefore: Grain (CS1, CS2, and CS3), Forage (CS4, CS5), Pasture (CS6), CRP, and Prairie. All statistical analyses were conducted using linear mixed effects models in SAS v 9.3. (see Sanford et al. 2012 for greater detail on statistical methodology).
Results and Discussion
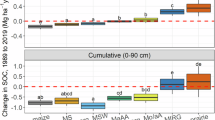
Analysis of changes in SOC (g kg−1) revealed significant differences (p < 0.05) between grain, forage, and pasture for each of the four soil profiles analyzed. This trend held in the analysis of ΔSOC Mg ha−1. When looking at the grain, forage, and pasture side by side (Figs. 29.1 and 29.2), with a Δ = 0 reference line, the pasture and grain differed. The pasture was not only a significant sink of atmospheric CO2 in the surface 0–60 cm profile (Fig. 29.2), when the entire soil profile was considered it was statistically indistinguishable from zero (Figs. 29.1 and 29.2) indicating that the Pasture has effectively stabilized SOC over 20 years at WICST. These findings are similar to those reported by Sanford et al. (2012) who showed that pasture C accumulation was significant in the 0–15-cm horizon. Sequestration of SOC in the surface horizons of the pastures was likely the result of both the quantity and quality of belowground net primary production. In perennial grass systems like the rotationally grazed pasture, 80–90 % of the belowground biomass is concentrated in the surface 30 cm of the soil and is dominated by fine roots (0–2 mm) (Jackson et al. 1996; Rasse et al. 2005).

ΔSOC concentration (1989–2009) comparison of Prairie and CRP plantings to the three land cover groups in the core WICST trial (Pasture, Forage, & Grain). ΔSOC values for each system represent the average change in SOC over the soil horizon analyzed. Error bars represent 90 % confidence limits to test difference of system mean from zero. Letters represent significant differences between systems within a given depth (Fishers LSD, α = 0.1)

ΔSOC mass comparison of Prairie and CRP plantings (1989–2009) to the three land cover groups in the core WICST trial (Pasture, Forage, & Grain). ΔSOC values for each system represent the average change in SOC over the soil horizon analyzed. Error bars represent 90 % confidence limits to test difference of system mean from zero. Letters represent significant differences between systems within a given depth increment (Fishers LSD, α = 0.1)
The opposite trend was observed for grain, where mean SOC losses were significant. This was likely the result of limited below ground carbon inputs (DeLuca and Zabinski 2011), historical SOC losses following the cessation of manure application to these fields at the start of WICST (Sanford et al. 2012), and mineralized SOC loss from tillage (Lal 2008). The pasture and grain were significantly different from one another in each of the eight profiles studied (Figs. 29.1 and 29.2).
Statistical differences between the prairie, CRP, forage, and pasture LCG’s were less pronounced and highlight the importance of grassland systems in stabilizing SOC. The prairie system was similar to pasture followed by the forage LCG. The CRP was comparable to the grain LCG in all soil profiles considered. Although mean ΔSOC values in CRP suggest carbon losses similar to those observed in the grain LCG this was not statistically significant.
The beneficial effects of a perennial system on SOC accretion are in part related to the advantages of reduced tillage as well as the fact that perennial plants allocate greater C resources belowground than annual crops. The influence of belowground biomass on SOC dynamics cannot be understated, playing a greater role in the long term stabilization of soil C than inputs from aboveground biomass (Rasse et al. 2005; Katterer et al. 2011). While aboveground productivity in tallgrass prairies may not differ from that of annual cropping systems, belowground productivity is far greater than that of annual systems (Guzman and Al-Kaisi 2010; DeLuca and Zabinski 2011). DeLuca and Zabinski (2011) reported that under tallgrass prairie, belowground net primary production can range from 8 to 15 Mg ha−1 year−1 with an estimated 3–5 Mg ha−1 year−1 retained in the soil.
The differences observed between Prairie and CRP suggest that species diversity has played a role in SOC stabilization and accretion at WICST. In the case of both the prairie and CRP plantings, the plots were in cool season grasses between 1989 and 1997 at which point they were tilled and planted to soybeans for a year prior to initiation of the native plantings (Simonsen et al. 2002). The SOC values for these systems therefore represent an 8-year period of C3 grass followed by a period of soil disturbance, and then a 10-year period of native grasslands. Evaluation of g SOC (kg soil)−1 data from the three time points (1989, 1999, 2009) shows a steady increase in SOC between 1989 and 1999 in the 0–15 cm horizon, and a steady decrease in SOC during the same time period in the 15–30 cm horizon (Fig. 29.3). Following tillage, and upon conversion to prairie and CRP, SOC in the prairie system stabilized between 0 and 15 cm and increased between 15 and 30 cm depth. The trend for CRP was different, with SOC loss occurring between 0 and 15 cm, and the rate o loss increasing in the 15–30 cm layer. Between 1999 and 2009, the CRP system changed from being comparable to the prairie to having significantly lower SOC levels in both the 0–15 and 15–30 cm soil layers (P = 0.002 and 0.017, respectively).

SOC concentration over time in the WICST Prairie and CRP plantings. Between 1989 and 1999 the systems were planted to C3 grasses, prairie and CRP were established in 1999. Error bars show ±1 s.e., NS = non-significant
There have been some reports on the enhanced SOC sequestration capabilities of diverse native systems compared to less diverse systems (e.g. Fornara and Tilman 2008; De Deyn et al. 2011). As species diversity increases, plants make greater use of the resources in an ecosystem and net productivity (above- and below-ground) increases (Hector et al. 1999; Spehn et al. 2005; Tilman et al. 2006).
Conclusions
Long-term agricultural research sites provide invaluable insight into the carbon sequestration potential of diverse agroecosystems. Analysis of SOC stocks during the first 20 years of WICST showed that annual row crop agriculture lost a significant amount of C (−5.5 Mg ha−1) while rotationally grazed pasture was a significant sink (5.1 Mg ha−1). Prairie, CRP, and forage systems were not significantly different from one another, but followed a trend suggesting that the prairie system was sequestering SOC, while the CRP and forage systems were losing SOC. Differences between SOC stock in prairie and CRP following establishment in 1999 (p < 0.05) suggest a link between SOC accumulation and plant functional diversity in native grasslands. Results from this and other similar investigations highlight the importance of perennial grass systems for maintaining and building SOC stocks. Carbon stabilization and accretion on the fine textured Mollisols of the North Central U.S. may not be possible, even under agricultural best management practices, without the inclusion of perennial grasses.
References
Conant RT, Easter M, Paustian K, Swan A, Williams S (2007) Impacts of periodic tillage on soil C stocks: a synthesis. Soil Tillage Res 95:1–10
De Deyn GB, Shiel RS, Ostle NJ, McNamara NP, Oakley S et al (2011) Additional carbon sequestration benefits of grassland diversity restoration. J Appl Ecol 48:600–608
DeLuca TH, Zabinski CA (2011) Prairie ecosystems and the carbon problem. Front Ecol Environ 9:407–413
Fierer N, Grandy SA, Six J, Paul EA (2009) Searching for unifying principles in soil ecology. Soil Biol Biochem 41:2249–2256
Follett RF, Reed DA (2010) Soil carbon sequestration in grazing lands: societal benefits and policy implications. Rangel Ecol Manag 63:4–15
Fornara DA, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96:314–322
Guo YY, Amundson R, Gong P, Yu Q (2006) Quantity and spatial variability of soil carbon in the conterminous United States. Soil Sci Soc Am J 70:590–600
Guzman JG, Al-Kaisi MM (2010) Soil carbon dynamics and carbon budget of newly reconstructed tall-grass prairies in south central Iowa. J Environ Qual 39:136–146
Heaton EA, Dohleman FG, Long SP (2008) Meeting US biofuel goals with less land: the potential of Miscanthus. Glob Chang Biol 14:2000–2014
Hector A, Schmid B, Beierkuhnlein C, Caldeira CM, Diemer M (1999) Plant diversity and productivity experiments in European grasslands. Science 286:1123–1127
Hong CO, Owen VN, Lee DK, Boe A (2013) Switchgrass, big bluestem, and indiangrass monocultures and their two- and three-way mixtures for bioenergy in the northern Great Plains. Bioenerg Res 6:229–239
Huggins DR, Buyanovsky GA, Wagner GH, Brown JR, Darmody RG, Peck TR, Lesoing GW, Vanotti MB, Bundy LG (1998) Soil organic C in the tallgrass prairie-derived region of the corn belt: effects of long-term crop management. Soil Tillage Res 47:219–234
Huggins DR, Allmaras RR, Clapp CE, Lamb JA, Randall GW (2007) Corn-soybean sequence and tillage effects on soil carbon dynamics and storage. Soil Sci Soc Am J 71:145–154
Jackson RB, Canadell J, Ehleringer JR, Mooney HA, Sala OE, Schulze ED (1996) A global analysis of root distributions for terrestrial biomes. Oecologia 108:389–411
Katterer T, Bolinder MA, Andren O, Kirchmann H, Menichetti L (2011) Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric Ecosyst Environ 141:184–192
Kögel-Knabner I, Guggenberger G, Kleber M, Kandeler E, Kalbitz K, Scheu S, Eusterhues K, Leinweber P (2008) Organo-mineral associations in temperate soils: integrating biology, mineraology, and organic matter chemistry. J Plant Nutr Soil Sci 171:61–82
Lal R (2004) Soil carbon sequestration impacts on global climate change and food security. Science 304:1623–1627
Lal R (2008) Carbon Sequestration. Philos Trans R Soc 363:815–830
Lee J, Hopmans JW, Rolston DE, Bear SG, Six J (2009) Determining soil carbon stock changes: simple bulk density corrections fail. Agric Ecosyst Environ 134:251–256
Luo Z, Wang E, Sun OJ (2010) Can no-tillage stimulate carbon sequestration in agricultural soils? A meta-analysis of paired experiments. Agric Ecosyst Environ 139:224–231
Nafziger ED, Dunker RE (2011) Soil organic carbon trends over 100 years in the morrow plots. Agron J 103:261–267
Oates LG, Undersander DJ, Gratton C, Bell MM, Jackson RD (2011) Management-intensive rotational grazing enhances forage production and quality of subhumid cool-season pastures. Crop Sci 51:892–901
Ogle SM, Breidt FJ, Paustian K (2005) Agricultural management impacts on soil organic carbon storage under moist and dry climatic conditions of temperate and tropical regions. Biogeochemistry 72:87–121
Posner JL, Casler MD, Baldock JO (1995) The Wisconsin integrated cropping systems trial: combining agroecology with production agronomy. Am J Altern Agric 10:98–107
Rasse DP, Rumpel C, Dignac MF (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 269:341–356
Sanford GS, Posner JL, Jackson RD, Kucharik CJ, Hedtcke JL, Lin T (2012) Soil carbon lost from Mollisols of the north central U.S.A. with 20 years of agricultural best management practices. Agric Ecosyst Environ 162:68–76
Schmidt MW, Torn MS, Abiven S, Dittmar T, Guggenberger G, Janssens IA, Kleber M, Kogel-Knabner I, Lehmann J, Manning DAC, Nannipieri P, Rasse DP, Weiner S, Trumbore S (2011) Persistence of soil organic matter as an ecosystem property. Nature 478:41–56
Simonsen K, Howell E, Posner JL (2002) Prairie establishment in agricultural soils. In: Hedtcke JL, Posner JL (eds) The Wisconsin integrated cropping systems trial: ninth report. http://wicst.wisc.edu/wp-content/uploads/katie-prairie-91903.pdf. Accessed 20 May 2013
Spehn EM, Hector A, Joshi J, Scherer-Lorenzen M, Scmid M et al (2005) Ecosystem effects of biodiversity manipulation in European grasslands. Ecol Monogr 75:37–63
Tilman D, Reich PB, Knops JMH (2006) Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441:629–632
USDA (2013) Crop production 2012 summary: January 2013. http://usda01.library.cornell.edu/usda/nass/CropProdSu//2010s/2013/CropProdSu-01-11-2013.pdf. Accessed 29 May 2013
West TO, Post WM (2002) Soil organic carbon sequestration rates by tillage and crop rotation: a global data analysis. Soil Sci Soc Am J 66:1930–1946
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Sanford, G.R. (2014). Perennial Grasslands Are Essential for Long Term SOC Storage in the Mollisols of the North Central USA. In: Hartemink, A., McSweeney, K. (eds) Soil Carbon. Progress in Soil Science. Springer, Cham. https://doi.org/10.1007/978-3-319-04084-4_29
Download citation
DOI: https://doi.org/10.1007/978-3-319-04084-4_29
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-04083-7
Online ISBN: 978-3-319-04084-4
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)