
Abstract
Habitat loss and human-caused mortality have led to an approximate 50% reduction of the distribution of the jaguar (Panthera onca). The large contraction in the jaguar’s occurrence points to a need to understand its population size and habitat preferences to apply to the species’ conservation. Typically, jaguar densities are estimated with capture–recapture modeling of photographic captures of individually identifiable individuals, while habitat selection is estimated from telemetry data. However, advances in spatial capture-recapture modeling now permit the simultaneous estimation of density and habitat selection based solely upon photographic detection data from camera-trapping grids. Here, we used data from 356 double camera-trap stations across five sites in the Paraguayan Dry Chaco to simultaneously estimate jaguar density and resource selection. We found that jaguar densities ranged from 0.58 to 1.39 individuals/100 km2. At the spatial scale of our analysis, jaguars showed a strong preference for forest cover, while space use was not affected by the Human Footprint Index. Our density estimates were consistent with previous estimates based upon a subset of our data, as well as with estimates for jaguar populations in other dryland ecosystems. Furthermore, the strong selection for forest was also consistent with range-wide patterns in jaguar space use and habitat selection derived from telemetry data. Due to extensive and ongoing deforestation in the Dry Chaco, combined with high human-caused mortality, the jaguar is critically endangered in Paraguay. Although we show that jaguars can persist in anthropogenically altered landscapes in Paraguay, their long-term survival at the national level is strongly dependent upon the effective enforcement of the national jaguar conservation law, and application of the national jaguar management plan, to mitigate negative population effects from habitat loss and human-caused mortality.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Apex predators are often used as conservation surrogates as they may play important roles in affecting ecosystem function and serve as bioindicators (Sergio et al. 2008). In the Neotropics, the occurrence of the jaguar (Panthera onca), the largest felid in the Americas, is associated with high levels of biodiversity, the provision of multiple ecosystem services, and consequently serves as an indicator of the conservation value of landscapes (Thornton et al. 2016; Burke et al. 2019). Since the jaguar is a forest-associated species throughout its range and across ecosystems types, and is sensitive to landscape-scale anthropogenic factors (Morato et al. 2018; Alvarenga et al. 2021; Thompson et al. 2021), it is a logical species to be used to examine the ecological effects of forest loss and other landscape-scale anthropogenic factors. However, the jaguar’s distribution has been reduced by about 50% range-wide and by 80% outside of Amazonia due to habitat loss and human–jaguar conflict driven persecution (de la Torre et al. 2018). This reduction in distribution indicates an important need to understand the effects of habitat loss and degradation on jaguar populations, the implications for their conservation, and in turn the ecosystem services with which they are associated.
Within the jaguar’s distribution, habitat conversion from expanding agricultural production has been acute during the last 20 years in dry forest and savanna systems in southern South America, particularly in the Dry Chaco of Argentina, Bolivia, and Paraguay which has undergone some of the highest rates of forest loss in the world (Curtis et al. 2018; Zalles et al. 2021; Da Ponte et al. 2021; Buchadas et al. 2022). Despite its high levels of habitat loss and often high levels of biodiversity, the Dry Chaco receives relatively little attention from the global conservation community due to a strong focus upon tropical humid systems (Redford et al. 1990; Kuemmerle et al. 2017; Qin et al. 2022). This pattern was discussed by Redford et al. (1990) who, in referring to the Dry Chaco, pointed out that “the attention to rainforest has acted like blinders” and “The concentration on rainforests, and the rhetoric that accompanies it, has led to the neglect of other severely threatened ecosystems.”
The statements of Redford et al. (1990) were prescient as they foresaw the forthcoming deforestation in the Dry Chaco and its implications for conservation that were to occur during the twenty-first century. However, in recent years, there has been an increasing amount of research on the effects of deforestation on multiple taxa and communities in the Dry Chaco (Periago et al. 2014; Cartes et al. 2015; Núñez-Regueiro et al. 2015; Semper-Pascual et al. 2018, 2020; Camino et al. 2020, 2022; de la Sancha et al. 2021), including the jaguar (Fig. 10.1) (Altrichter et al. 2006; Noss et al. 2012; Quiroga et al. 2014; Thompson and Velilla 2017; McBride and Thompson 2018; Romero-Muñoz et al. 2019; Thompson et al. 2020, 2021, 2022). Importantly, for the jaguar, although forest availability is key, it is not the sole factor determining jaguar occurrence in the Dry Chaco. Anthropogenic factors have a disproportionately strong negative influence on jaguar occurrence despite sufficient habitat and prey availability (Quiroga et al. 2014; McBride and Thompson 2018; Romero-Muñoz et al. 2019, 2020; Thompson et al. 2020). Consequently, evaluating the relative importance of both environmental and anthropogenic factors on jaguar density and habitat use is a necessary precursor to understand how deforestation in the Dry Chaco is affecting the jaguar population.

Camera-trap record of Jaguar (Panthera onca) in the Paraguayan Medanos
Advances in spatial capture–recapture (SCR) modeling allow for the simultaneous estimation of density, habitat selection, and connectivity (Royle et al. 2013b; Sutherland et al. 2015; Morin et al. 2017), providing a powerful framework to integrate population and landscape ecology (Royle et al. 2018). We previously took advantage of this framework and found that jaguar density and landscape connectivity in the Paraguayan Dry Chaco were negatively affected by increases in deforestation within landscapes (Thompson et al. 2022), which was consistent with expectations based upon patterns in jaguar space use (Thompson et al. 2021). Furthermore, we illustrated the importance and value of accounting for potential bias stemming from habitat-driven space use by jaguars when estimating density in a spatial capture–recapture modeling framework (Thompson et al. 2022).
Historically, the movements of animals within their home range (third-order habitat selection; Johnson 1980), and the drivers of those movements (Manly et al. 2002), have typically been estimated using telemetry data. However, SCR modeling has been extended to estimate resource selection by taking advantage of the spatial information provided by spatial recaptures of individuals (Royle et al. 2013b; Sollmann et al. 2016; Linden et al. 2018). This advance consequently allows for the estimation of habitat use within activity areas using Resource Selection Functions (RSF; Manly et al. 2002) based solely upon spatial capture-recapture data, while allowing to simultaneously estimate density (Royle et al. 2013b).
We took advantage of the capabilities of SCR modeling to include RSFs to simultaneously estimate habitat selection within activity areas and density of jaguars in the Paraguayan Dry Chaco. This allowed us to explore how jaguar space use was affected by forest availability and landscape-scale anthropogenic factors across landscapes with differing intensities of forest conversion. Our sampling areas included landscapes with varying levels of deforestation, as well as the Medanos, a mixed matorral – savanna – forest system within the Bolivian–Paraguayan Dry Chaco, where in Paraguay jaguars have yet to be studied. Recognizing that jaguars demonstrate a general preference for forest cover, avoidance of pastures, reduced occurrence in response to anthropogenic factors, and in the Paraguayan Dry Chaco reduced densities in relation to deforestation (Morato et al. 2018; Thompson et al. 2020, 2021, 2022; Alvarenga et al. 2021), we expected that (1) within their activity areas, jaguars will select forest regardless if the heterogeneity in forest availability was natural or due to deforestation, while (2) areas with higher anthropogenic impacts will be avoided. Furthermore, we intended to show the capabilities of SCR modeling to expand its utility beyond density estimation to address key conservation questions, such as resource selection, solely from camera-trap-based capture–recapture data.
2 Methods
2.1 Study Site
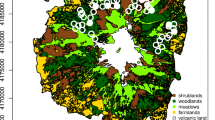
The Dry Chaco extends across ~787,000 km2 in parts of Argentina (62%), Paraguay (22%), and Bolivia (16%) and is the largest subtropical dry forest system in the world (Olson et al. 2001). The Dry Chaco is a global deforestation hotspot, whereby ranching and row crop agriculture have driven the deforestation of about 27% of the ecoregion’s forest (Caldas et al. 2015; Da Ponte et al. 2021; Hansen et al. 2013; Vallejos et al. 2015). In Paraguay, 64,000–77,000 km2 (~45%) of its Dry Chaco forest area was lost by 2020, mostly due to pasture creation for cattle raising, equating to an annual forest transformation rate of about 4% or ~ 500 ha/day/year since 2000 (Fig. 10.2; Hansen et al. 2013; Caldas et al. 2015; Vallejos et al. 2015; Baumann et al. 2017; Da Ponte et al. 2021). Importantly, from a conservation perspective, the forest loss during the past two decades has occurred in wilderness areas of high conservation value, so that since 2000 the area of intact forest landscapes in the Paraguayan Dry Chaco has decreased by 76% (Watson et al. 2018; Williams et al. 2020).

Location of the Dry Chaco within South America and land cover and protected areas in the Paraguayan Dry Chaco for the years 2000 and 2021. The graph shows the cumulative annual forest loss and cattle population from 1990 to 2021 for the Paraguayan departments of Alto Paraguay and Boquerón, which encompass the large majority of the Dry Chaco in Paraguay. (Data from: https://www.senacsa.gov.py/index.php/informacion-publica/estadistica-pecuaria)
2.2 Camera-Trap Sampling
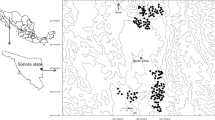
During the austral fall to winter (May–September) 2016–2018, we employed camera-trap sampling at four sites with semideciduous xeric forest in the Dry Chaco (Mereles 2005; Mereles et al. 2019) along a gradient of deforestation intensity ranging from 17% to 51% deforested. We employed 335 double camera-trap stations located on roads, firebreaks, or along fence lines (Fig. 10.3, Table 10.1; Thompson et al. 2022). Additionally, during 2018 we also sampled a site in the Medanos within the Dry Chaco, a matorral – savanna system interspersed with xeric forest (Mereles 2005; Mereles et al. 2019) which previously had not been sampled for jaguars in Paraguay (Fig. 10.3, Table 10.1). The initial four sampling sites were all located on private ranchlands and the site in the Medanos was on a private protected area and adjoining indigenous land.

Locations of camera-trap grids and their 25 km buffers in the Paraguayan Dry Chaco (Olson et al. 2001) in relation to forest cover in 2017 (Hansen et al. 2013). Shaded areas are outside the limit of the Dry Chaco. Inset shows the location of the Dry Chaco within South America
Sampling periods ranged from 98 to 279 days, with a mean of 100 operational days per station per site (Table 10.1). The site with the longest sampling period (Site 4) resulted from its sampling grid being divided into two temporally overlapping sections (48 and 27 stations). The long sampling periods were justifiable to increase detections (Dupont et al. 2019; Tobler and Powell 2013), while we see no reason to expect any effect on density as a function of sampling year as there were no abnormal differences in temperature or precipitation among the 2016–2018 sampling periods (www.meteorologia.gov.py). Moreover, in our analysis we accounted for potential site-specific differences in detectability and space use (see Data analysis).
Minimum convex polygons of our trapping grids ranged between 80 and 282 km2 and, as discussed by Thompson et al. (2022), were sufficiently large related to jaguar activity areas to minimize bias in density estimates (Sollmann et al. 2011; Tobler and Powell 2013). We defined our modeling state space with a 25-km buffer around our trapping grids (see Data analysis), with our initial four sampling sites having a proportional area deforested ranging from 17% to 51% (Hansen et al. 2013). At the fifth site in the Medanos, the majority of unforested land was naturally occurring matorral with some deforested areas, so 18% of the modeled state space did not include forest (Fig. 10.3, Table 10.1).
2.3 Data Analysis
We used spatial capture-recapture modeling framework incorporating resource selection (hereafter SCR–RS) to explore the effect of forest cover and anthropogenic pressures on jaguar resource selection while simultaneously estimating jaguar densities (Royle et al. 2013b; Sollmann et al. 2016). The SCR–RS modeling framework accommodates resource selection models by incorporating the resource selection information spatially inherent in camera-trapping data. This permits the estimation of resource selection parameters while relaxing the assumption of symmetrical distance in the SCR detection function, allowing for space use to be a function of differential habitat preferences. Apart from being able to estimate third-order habitat use directly from camera-trap data, accounting for habitat-driven differences in space use reduces bias in detectability when estimating density (Royle et al. 2013b). We considered this important given the preferred use of the forest by jaguars (Alvarenga et al. 2021; Morato et al. 2018; Thompson et al. 2021), and previous modeling of a subset of our data which demonstrated the importance of accounting for asymmetrical space use when estimating jaguar density in fragmented Dry Chaco forest (Thompson et al. 2022).
Using the photographic captures from our sampling we developed individual daily encounter histories for jaguars by assigning jaguar photographs to each sampling station by date and individual. Jaguars were identified by their spot patterns and sexed by genitalia, with juveniles accompanied by their mother excluded from the data set. Using these spatial encounter histories, we estimated densities for each site in a multisite (multisession) model in R (R Core Team 2019) using the oSCR package (Sutherland et al. 2015) with a state space delimited by a 25-km buffer from traps at a resolution of 1 km. Although in previous modeling of a subset of our data a resolution of 2 km was used (Thompson et al. 2022), we found that estimates were sensitive to resolutions greater than 1 km, which we attribute to the fine-grain nature of habitat heterogeneity in the additional site from the Medanos. Since jaguars in the Paraguayan Dry Chaco can move more than 14 km per day on average (McBride and Thompson 2018; Thompson et al. 2021) a 1 km resolution is consistent with the scale of jaguar space use in our study areas.
As in Thompson et al. (2022), we used the mean proportional area of forest as a biologically relevant metric to characterize forest availability. We developed maps of forest occurrence from 1 arc-second spatial data on forest canopy density corresponding to the year of our sampling (Hansen et al. 2013), classifying all areas with a forest canopy density of ≥15% as forest (based upon ground truthing and our familiarity with the study system), and then deriving the mean proportional forest area for each grid cell of our 1 km resolution state space. To quantify anthropogenic influences within our state spaces we used the Human Footprint Index (HFI) (Sanderson et al. 2002; Venter et al. 2016; wcshumanfootprint.org), taking the average value at the 1 km resolution for the year corresponding to the beginning of sampling for each site.
We tested a set of models that evaluated the probability of space use as a function of the proportional forest area and the HFI, additively or separately. Models always included a site (session) effect on density, to meet closure assumptions, and on σ (scale parameter of decline in p with distance) and detection (p) to account for potential differences in space use and detection, respectively among sites. The scale parameter σ describes the rate that the probability of detection (p) decreases with distance from the estimated activity center of individuals, whereby p0 is the baseline detection probability at an individual’s activity center. We also evaluated the effect of sex on σ and p0. Model selection was based on rankings of Akaike Information Criteria (AIC) weights (Burnham and Anderson 2002). Since AIC has a tendency to select models with uninformative parameters with no ecological effects that do not reduce AIC but increase model complexity (Burnham and Anderson 2002; Arnold 2010), we excluded models with uninformative parameters from the final model set (Arnold 2010). A parameter was considered uninformative when its significance level was <85% since the effect of such parameters is insufficient to warrant their inclusion in a model (Arnold 2010).
Similar to Sollmann et al. (2016), we used linear discriminant analysis (Manly and Alberto 2016) to evaluate how modeling state spaces differ in anthropogenic and environmental characteristics to potentially explain the difference in jaguar densities among sites. We examined differences in proportional forest area, HFI, estimated net primary productivity (Zhao et al. 2005), and estimated mean annual precipitation along with its coefficient of variation (Fick and Hijmans 2017), using the mean values at the 1 km resolution of the modeling state spaces. We evaluated correlations among covariates from the discriminant functions that explained at least 95% of the between-group variance. We considered a strong correlation between covariables where coefficients were ≥ 0.7 and a p-value <0.05. The linear discriminant analysis was undertaken in R using the MASS package (Venables and Ripley 2002).
3 Results
We detected between 6 and 13 individual jaguars per site. One GPS collared individual from site 3 was censored because it was killed after moving outside our sampling area. The total number of recaptures per site ranged from 73 to 197 and the total spatial recaptures per site ranged from 38 to 116 (Table 10.1). Of the models evaluated, three contained 100% of the model weights, with the model including the effect of forest cover on resource selection, and no sex effect on detection probability (p0) or on the spatial scale factor (σ); being the highest ranked supported by 53% of model weights (Table 10.2). This model indicated a preference by jaguars for areas with increasing proportional forest cover (α = 1.31, SE = 0.2; Table 10.3). The other two models included an uninformative parameter for the sex (−0.18, SE = 0.21) on detection probability (p0) or HFI (α = 0.06, SE = 0.29) on resource selection and consequently were excluded. Based upon the best supported model, density estimates ranged from 0.58 to 1.39 individuals/100 km2 and fully forested areas were estimated to be used 3.7 times more than completely deforested areas (Fig. 10.4). Baseline detectability at the mean forest cover value from each site was estimated to be between 62% and 80% (Table 10.4). Estimates of σ ranged from 3.3 to 5.6 km and sex ratio estimates had high uncertainty, but the estimated probability of being a male varied from 33% to 67% across sites (Table 10.4).

Prediction of relative probability of site use by jaguars as a function of proportional forest area. Shaded area represents the 95% confidence interval
The first two discriminant functions of the habitat analysis explained 98.19% of the between-group variance among the five state spaces. The first discriminant function of the habitat analysis explained 84.07% of the between-group variance and was strongly negatively correlated with the coefficient of variance of annual precipitation and strongly positively correlated with mean annual precipitation (Table 10.5). The second discriminant function explained 14.12% of the variance between sites, with no strong correlations with covariates; however, net primary productivity showed a moderate correlation (Table 10.5). The effect of mean annual precipitation and its coefficient of variation grouped Sites 1–4 together, albeit with some among-site differences, while Site 5 from the Medanos was distinctly separated from the other sites (Fig. 10.5).

Plot of the first and second linear discriminant functions for the habitat covariables from the spatial capture-recapture modeling state spaces. The first discriminant function was strongly negatively correlated with the coefficient of variation of annual precipitation and strongly positively correlated with mean annual precipitation. The second discriminant function only had a moderate positive correlation with net primary productivity
4 Discussion
By using the capacity of the SCR–RS model to estimate resource selection we further confirm the importance of forest cover for jaguars documented both range-wide (Morato et al. 2018; Thompson et al. 2021) and in the Dry Chaco (Romero-Muñoz et al. 2019; Thompson et al. 2022). We found strong third-order habitat selection for forested areas by jaguars in the Paraguayan Dry Chaco. Importantly, this relationship was evident in areas with forest fragmented through deforestation or naturally heterogeneous as in the matorral – savanna – forest system of the Medanos. At the same time, we found no avoidance of anthropogenic factors represented by the HFI. Given the known negative relationship of anthropogenic factors on the occurrence and density of jaguars in the Dry Chaco (McBride and Thompson 2018; Romero-Muñoz et al. 2019; Thompson et al. 2020, 2022) this was unexpected and is likely due to the spatial scale of the HFI, or the information comprising the HFI, not capturing the relevant effects of jaguar space use.
Previously, in four of our five sites a negative relationship of deforestation on jaguar density was shown between the most and least deforested sites, as well as a strong negative effect of deforestation on population connectivity (Thompson et al. 2022). However, with the inclusion of the naturally heterogeneous site from the Medanos (Site 5), this relationship was not evident as its estimated density was similar to the most deforested of our sites (Sites 3 and 4). Nor did the linear discriminant analysis find a strong difference in net primary productivity among sites, which has been shown to decrease jaguar spatial needs, and in turn lead to increased density (Thompson et al. 2021). However, the analysis did show that the site from the Medanos (Site 5) received less, and much more variable, precipitation than the other sites. This suggests that the intra- and interannual variability of water is an important driver in limiting jaguar density in the Medanos despite ample forest cover and relatively low anthropogenic pressures. As our study areas are at the arid extreme of the jaguar habitat, and our density estimates are consistent with those from protected areas in other dryland systems with high variability in precipitation (Sollmann et al. 2011, 2013; Noss et al. 2012; Gutiérrez-González et al. 2015; Finnegan et al. 2020), our results provide valuable inferences on habitat needs for the conservation of jaguar in arid systems throughout its range and in support of potential reintroduction efforts for jaguars in arid systems such as in the southwestern United States (Sanderson et al. 2021, 2022).
We found no sex effect on either detectability or σ which was consistent with previous modeling that included our data (Thompson et al. 2022), but unexpected compared to general patterns in estimates of jaguar space use from spatial capture–recapture modeling and telemetry data (Sollmann et al. 2011; Tobler et al. 2013, 2018; Boron et al. 2016; Morato et al. 2016; Alvarenga et al. 2021; Thompson et al. 2021). Differences in sex ratios among sites may cause abundance related effects on detectability by sex which collectively offset the effect of sex on detection since both males and females were detected more frequently dependent upon the site (Table 10.1).
We note that our estimates of σ were larger than previous estimates (Thompson et al. 2022) for sites 1–4 which we attribute to differences in the modeling framework, whereby Thompson et al. (2022) explicitly modeled asymmetrical, non-Euclidean space use and connectivity as a function of forest cover, and here we used SCR–RS to estimate habitat driven differences in space use within activity areas. Regardless, both modeling approaches generated similar density estimates, while showing the importance of forest cover for jaguars from different perspectives while reducing potential bias by accounting for asymmetrical space use (Royle et al. 2013a, b; Sutherland et al. 2015).
Although we show a strong selection for forest cover by jaguars, anthropogenic factors negatively affect jaguar survival and occurrence in the Dry Chaco, driving complex source–sink dynamics (McBride and Thompson 2018; Romero-Muñoz et al. 2019, 2020; Thompson et al. 2020). The large movements of jaguars and the maintenance of connectivity in relation to forest loss (McBride and Thompson 2018; Thompson et al. 2021), suggest that the sites with high levels of forest loss are population sinks, with their populations being maintained by immigration (Romero-Muñoz et al. 2019; Thompson et al. 2022). This highlights the importance of considering anthropogenic effects on jaguar occurrence in the Dry Chaco which is further evident in the functional extinction of the jaguar in the Argentine Dry Chaco despite high forest cover and sufficient prey availability (Quiroga et al. 2014; Thompson et al. 2020).
Consequently, our results may not be fully representative of the population trajectory in our study sites. Our sites could be demonstrating extinction debt, whereby individual longevity and immigration are masking negative population trends (Tilman and Lehman 1994; Semper-Pascual et al. 2018; Romero-Muñoz et al. 2019; Thompson et al. 2022). For example, despite similar estimates of density, as well as similarities in anthropogenic influences, the status of the jaguar populations in the more deforested sites (Sites 3 and 4) and in the Medanos (Site 5) is not clear based solely on density estimates. This illustrates the limitations of static estimates of density for conservation evaluations, highlighting the need for long-term studies to quantify survival, recruitment, and immigration to characterize the conservation value of the various landscapes for jaguars (Harihar et al. 2020).
The high rate of forest loss in the Paraguayan Dry Chaco is not only a concern for the conservation of the jaguar, but raises concerns for other forest dependent and endemic species in the Dry Chaco (Romero-Muñoz et al. 2020) such as the Chacoan peccary (Catagonus wagneri), Quebracho crested tinamou (Eudromia formosa), and Chacoan naked-tailed armadillo (Cabassous chacoensis). Furthermore, the forest loss in the Paraguayan Dry Chaco, apart from generating important concerns for biological conservation, has additional cultural and human rights implications as the Paraguayan Dry Chaco is the ancestral lands of multiple indigenous peoples, including the Ayoreo Totobiegosode, the last indigenous peoples outside of the Amazon Basin with members living in isolation (DGEEC 2004).
Considering the jaguar’s effective role as an umbrella and flagship for biodiversity conservation (Thornton et al. 2016), its conservation, which is dependent upon forest conservation and the fostering of coexistence, has significant ecological and cultural implications (United Nations Development Program et al. 2019). However, within Paraguay, the jaguar is listed as critically endangered due to continuing high rates of habitat loss and direct human-caused mortality (Giordano et al. 2017), pointing to the urgent need for the Paraguayan government to better implement the national jaguar conservation plan (Secretaría del Ambiente et al. 2016) and national jaguar conservation law (Congreso de la Nación Paraguaya 2014) to ensure the long-term survival of the species and the services that it provides.
References
Altrichter M, Boaglio G, Perovic P (2006) The decline of jaguars Panthera onca in the Argentine Chaco. Oryx 40:302. https://doi.org/10.1017/S0030605306000731
Alvarenga GC, Chiaverini L, Cushman SA et al (2021) Multi-scale path-level analysis of jaguar habitat use in the Pantanal ecosystem. Biol Conserv 253:108900. https://doi.org/10.1016/j.biocon.2020.108900
Arnold TW (2010) Uninformative parameters and model selection using Akaike’s information criterion. J Wildl Manag 74:1175–1178. https://doi.org/10.1111/j.1937-2817.2010.tb01236.x
Baumann M, Israel C, Piquer-Rodríguez M et al (2017) Deforestation and cattle expansion in the Paraguayan Chaco 1987–2012. Reg Environ Chang 17:1179–1191. https://doi.org/10.1007/s10113-017-1109-5
Boron V, Tzanopoulos J, Gallo J et al (2016) Jaguar densities across human-dominated landscapes in Colombia: the contribution of unprotected areas to long term conservation. PLoS One 11:e0153973. https://doi.org/10.1371/journal.pone.0153973
Buchadas A, Baumann M, Meyfroidt P, Kuemmerle T (2022) Uncovering major types of deforestation frontiers across the world’s tropical dry woodlands. Nat Sustain. https://doi.org/10.1038/s41893-022-00886-9
Burke S, Mulligan M, Stolton S, Dudley N (2019) Ecosystem services provided by the habitat of the Jaguar. United Nations Development Program
Burnham KP, Anderson DR (2002) A practical information-theoretic approach. Model selection and multimodel inference, 2nd edn. Springer, New York
Caldas MM, Goodin D, Sherwood S et al (2015) Land-cover change in the Paraguayan Chaco: 2000–2011. J Land Use Sci 10:1–18. https://doi.org/10.1080/1747423X.2013.807314
Camino M, Thompson J, Andrade L et al (2020) Using local ecological knowledge to improve large terrestrial mammal surveys, build local capacity and increase conservation opportunities. Biol Conserv 244:108450. https://doi.org/10.1016/j.biocon.2020.108450
Camino M, Thompson J, Arriaga Velasco-Aceves P et al (2022) Less than six generations to save the chacoan peccary. Biodivers Conserv 31:413–432. https://doi.org/10.1007/s10531-021-02337-x
Cartes JL, Thompson J, Yanosky A (2015) El Chaco paraguayo como uno de los últimos refugios para los mamíferos amenazados del Cono Sur. Paraquaria Nat 3(2):37–47
Congreso de la Nación Paraguaya Ley N° 5302 de Conservación de la Panthera onca; 2014.
Curtis PG, Slay CM, Harris NL et al (2018) Classifying drivers of global forest loss. Science 361:1108–1111. https://doi.org/10.1126/science.aau3445
Da Ponte E, García-Calabrese M, Kriese J et al (2021) Understanding 34 years of Forest cover dynamics across the Paraguayan Chaco: characterizing annual changes and forest fragmentation levels between 1987 and 2020. Forests 13:25. https://doi.org/10.3390/f13010025
de la Sancha NU, Boyle SA, McIntyre NE et al (2021) The disappearing dry Chaco, one of the last dry forest systems on earth. Landsc Ecol 36:2997–3012. https://doi.org/10.1007/s10980-021-01291-x
de la Torre JA, González-Maya JF, Zarza H et al (2018) The jaguar’s spots are darker than they appear: assessing the global conservation status of the jaguar Panthera onca. Oryx 52:300–315. https://doi.org/10.1017/S0030605316001046
DGEEC (2004) Atlas de las Comunidades Indígenas en el Paraguay. Dirección General de Estadística, Encuestas y Censos
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Finnegan SP, Galvez-Bravo L, Silveira L et al (2020) Reserve size, dispersal and population viability of wide ranging carnivores: the case of jaguars in Emas National Park, Brazil. Anim Conserv acv 12608. https://doi.org/10.1111/acv.12608
Giordano AJ, Giménez D, Martínez V et al (2017) Carnivora. In: Asociación Paraguaya de Mastozoología, Secretaría del Ambiente (eds). Libro Rojo de los Mamíferos del Paraguay: especies amenazadas de extinción, p 137
Gutiérrez-González CE, Gómez-Ramírez MA, López-González CA, Doherty PF (2015) Are private reserves effective for jaguar conservation? PLoS One 10:e0137541. https://doi.org/10.1371/journal.pone.0137541
Hansen MC, Potapov PV, Moore R et al (2013) High-resolution global maps of 21st-century forest cover change. Science 342:850–853. https://doi.org/10.1126/science.1244693
Harihar A, Pandav B, Ghosh-Harihar M, Goodrich J (2020) Demographic and ecological correlates of a recovering tiger (Panthera tigris) population: lessons learnt from 13-years of monitoring. Biol Conserv 252:108848. https://doi.org/10.1016/j.biocon.2020.108848
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71. https://doi.org/10.2307/1937156
Kuemmerle T, Altrichter M, Baldi G et al (2017) Forest conservation: remember Gran Chaco. Science 355:465. https://doi.org/10.1126/science.aal3020
Linden DW, Sirén APK, Pekins PJ (2018) Integrating telemetry data into spatial capture-recapture modifies inferences on multi-scale resource selection. Ecosphere 9:e02203. https://doi.org/10.1002/ecs2.2203
Manly BF, Alberto JAN (2016) Multivariate statistical methods: a primer. Chapman and Hall/CRC
Manly BFJ, McDonald LL, Thomas DL et al (2002) Resource selection by animals: statistical analysis and design for field studies. Kluwer Academic Publishers
McBride RT, Thompson JJ (2018) Space use and movement of jaguar (Panthera onca) in western Paraguay. Mammalia 82:540–549. https://doi.org/10.1515/mammalia-2017-0040
Mereles MF (2005) Una aproximación al conocimiento de las formaciones vegatales del Chaco Boreal, Paraguay. Rojasiana 6:5–48
Mereles MF, Céspedes G, Cartes JL et al (2019) Biological corridors as a connectivity tool in the region of the great American chaco: identification of biodiversity hotspots in the ecoregions of the paraguayan chaco. Restor Ecol 2:20–26. https://doi.org/10.30564/re.v2i1.1324
Morato RG, Stabach JA, Fleming CH et al (2016) Space use and movement of a neotropical top predator: the endangered jaguar. PLoS One 11:e0168176. https://doi.org/10.1371/journal.pone.0168176
Morato RG, Connette GM, Stabach JA et al (2018) Resource selection in an apex predator and variation in response to local landscape characteristics. Biol Conserv 228:233–240. https://doi.org/10.1016/j.biocon.2018.10.022
Morin DJ, Fuller AK, Royle JA, Sutherland C (2017) Model-based estimators of density and connectivity to inform conservation of spatially structured populations. Ecosphere 8:e01623. https://doi.org/10.1002/ecs2.1623
Noss AJ, Gardner B, Maffei L et al (2012) Comparison of density estimation methods for mammal populations with camera traps in the Kaa-Iya del Gran Chaco landscape: density estimation with camera traps in the Chaco. Anim Conserv 15:527–535. https://doi.org/10.1111/j.1469-1795.2012.00545.x
Núñez-Regueiro MM, Branch L, Fletcher RJ et al (2015) Spatial patterns of mammal occurrence in forest strips surrounded by agricultural crops of the Chaco region, Argentina. Biol Conserv 187:19–26. https://doi.org/10.1016/j.biocon.2015.04.001
Olson DM, Dinerstein E, Wikramanayake ED et al (2001) Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51:933. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Periago ME, Chillo V, Ojeda RA (2014) Loss of mammalian species from the south American Gran Chaco: empty savanna syndrome?: loss of Chacoan mammals. Mammal Rev. https://doi.org/10.1111/mam.12031
Qin S, Kuemmerle T, Meyfroidt P et al (2022) The geography of international conservation interest in South American deforestation frontiers. Conserv Lett 15. https://doi.org/10.1111/conl.12859
Quiroga VA, Boaglio GI, Noss AJ, Di Bitetti MS (2014) Critical population status of the jaguar Panthera onca in the Argentine Chaco: camera-trap surveys suggest recent collapse and imminent regional extinction. Oryx 48:141–148. https://doi.org/10.1017/S0030605312000944
Redford KH, Taber A, Simonetti JA (1990) There is more to biodiversity than the tropical rain forests. Conserv Biol 4:328–330. https://doi.org/10.1111/j.1523-1739.1990.tb00296.x
Romero-Muñoz A, Torres R, Noss AJ et al (2019) Habitat loss and overhunting synergistically drive the extirpation of jaguars from the Gran Chaco. Divers Distrib 25:176–190. https://doi.org/10.1111/ddi.12843
Romero-Muñoz A, Benítez-López A, Zurell D et al (2020) Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography 43:954–966. https://doi.org/10.1111/ecog.05053
Royle JA, Chandler RB, Gazenski KD, Graves TA (2013a) Spatial capture–recapture models for jointly estimating population density and landscape connectivity. Ecology 94:287–294. https://doi.org/10.1890/12-0413.1
Royle JA, Chandler RB, Sun CC, Fuller AK (2013b) Integrating resource selection information with spatial capture-recapture. Methods Ecol Evol 4:520–530. https://doi.org/10.1111/2041-210X.12039
Royle JA, Fuller AK, Sutherland C (2018) Unifying population and landscape ecology with spatial capture-recapture. Ecography 41:444–456. https://doi.org/10.1111/ecog.03170
Sanderson EW, Jaiteh M, Levy MA et al (2002) The human footprint and the last of the wild. Bioscience 52:891. https://doi.org/10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2
Sanderson EW, Beckmann JP, Beier P et al (2021) The case for reintroduction: the jaguar (Panthera onca) in the United States as a model. Conserv Sci and Prac 3:e392. https://doi.org/10.1111/csp2.392
Sanderson EW, Fisher K, Peters R et al (2022) A systematic review of potential habitat suitability for the jaguar Panthera onca in Central Arizona and New Mexico, USA. Oryx 56:116–127. https://doi.org/10.1017/S0030605320000459
Secretaría del Ambiente, Wildlife Conservation Society Paraguay, Itaipu Binacional (2016) Plan de manejo de la Panthera onca, Paraguay 2017–2026, Asunción
Semper-Pascual A, Macchi L, Sabatini FM et al (2018) Mapping extinction debt highlights conservation opportunities for birds and mammals in the south American Chaco. J Appl Ecol 55:1218–1229. https://doi.org/10.1111/1365-2664.13074
Semper-Pascual A, Decarre J, Baumann M et al (2020) Using occupancy models to assess the direct and indirect impacts of agricultural expansion on species’ populations. Biodivers Conserv 29:3669–3688. https://doi.org/10.1007/s10531-020-02042-1
Sergio F, Caro T, Brown D et al (2008) Top predators as conservation tools: ecological rationale, assumptions, and efficacy. Annu Rev Ecol Evol Syst 39:1–19. https://doi.org/10.1146/annurev.ecolsys.39.110707.173545
Sollmann R, Furtado MM, Gardner B et al (2011) Improving density estimates for elusive carnivores: accounting for sex-specific detection and movements using spatial capture–recapture models for jaguars in Central Brazil. Biol Conserv 144:1017–1024. https://doi.org/10.1016/j.biocon.2010.12.011
Sollmann R, Tôrres NM, Furtado MM et al (2013) Combining camera-trapping and noninvasive genetic data in a spatial capture–recapture framework improves density estimates for the jaguar. Biol Conserv 167:242–247. https://doi.org/10.1016/j.biocon.2013.08.003
Sollmann R, Gardner B, Belant JL et al (2016) Habitat associations in a recolonizing, low-density black bear population. Ecosphere 7:e01406. https://doi.org/10.1002/ecs2.1406
Sutherland C, Fuller AK, Royle JA (2015) Modelling non-Euclidean movement and landscape connectivity in highly structured ecological networks. Methods Ecol Evol 6:169–177. https://doi.org/10.1111/2041-210X.12316
Thompson JJ, Velilla M (2017) Modeling the effects of deforestation on the connectivity of jaguar Panthera onca populations at the southern extent of the species’ range. Endanger Species Res 34:109–121. https://doi.org/10.3354/esr00840
Thompson JJ, Martí CM, Quigley H (2020) Anthropogenic factors disproportionately affect the occurrence and potential population connectivity of the Neotropic’s apex predator: the jaguar at the southwestern extent of its distribution. Glob Ecol Conserv 24:e01356. https://doi.org/10.1016/j.gecco.2020.e01356
Thompson JJ, Morato RG, Niebuhr BB et al (2021) Environmental and anthropogenic factors synergistically affect space use of jaguars. Curr Biol 31:3457–3466.e4. https://doi.org/10.1016/j.cub.2021.06.029
Thompson JJ, Velilla M, Cabral H et al (2022) Jaguar (Panthera onca) population density and landscape connectivity in a deforestation hotspot: the Paraguayan dry chaco as a case study. Perspect Ecol Conserv S2530064422000591. https://doi.org/10.1016/j.pecon.2022.09.001
Thornton D, Zeller K, Rondinini C et al (2016) Assessing the umbrella value of a range-wide conservation network for jaguars (Panthera onca). Ecol Appl 26:1112–1124. https://doi.org/10.1890/15-0602
Tilman D, Lehman CL (1994) Habitat destruction and the extinction debt. Nature 371:65–66
Tobler MW, Carrillo-Percastegui SE, Zúñiga Hartley A, Powell GVN (2013) High jaguar densities and large population sizes in the core habitat of the southwestern Amazon. Biol Conserv 159:375–381. https://doi.org/10.1016/j.biocon.2012.12.012
Tobler MW, Garcia-Anleu R, Carrillo-Percastegui SE et al (2018) Do responsibly managed logging concessions adequately protect jaguars and other large and medium-sized mammals? Two case studies from Guatemala and Peru. Biol Conserv 220:245–253. https://doi.org/10.1016/j.biocon.2018.02.015
United Nations Development Program, Panthera, Wildlife Conservation Society, Worldwide Fund for Nature (2019) Jaguar 2030 Roadmap. Regional plan to save America’s largest cat and its ecosystem.
Vallejos M, Volante JN, Mosciaro MJ et al (2015) Transformation dynamics of the natural cover in the Dry Chaco ecoregion: a plot level geo-database from 1976 to 2012. J Arid Environ 123:3–11. https://doi.org/10.1016/j.jaridenv.2014.11.009
Venables WN, Ripley BD (2002) Modern applied statistics with S, Fourth. Springer, New York
Venter O, Sanderson EW, Magrach A et al (2016) Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat Commun 7. https://doi.org/10.1038/ncomms12558
Watson JEM, Evans T, Venter O et al (2018) The exceptional value of intact forest ecosystems. Nat Ecol Evol 2:599–610. https://doi.org/10.1038/s41559-018-0490-x
Williams BA, Venter O, Allan JR et al (2020) Change in terrestrial human footprint drives continued loss of intact ecosystems. One Earth 3:371–382. https://doi.org/10.1016/j.oneear.2020.08.009
Zalles V, Hansen MC, Potapov PV et al (2021) Rapid expansion of human impact on natural land in South America since 1985. Sci Adv 7:eabg1620. https://doi.org/10.1126/sciadv.abg1620
Zhao M, Heinsch FA, Nemani RR, Running SW (2005) Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens Environ 95:164–176. https://doi.org/10.1016/j.rse.2004.12.011
Acknowledgments
We thank the ranch owners, administrators, and employees who facilitated and made our research possible. This research was funded by the Consejo Nacional de Ciencia y Tecnología (CONACYT) of Paraguay under project 14-INV-060 and by IUCN – Netherlands, in the person of Sander van Andel. JJT, MV, HCB, VR, JMCK, CBR, MFR, and JLC were partially supported by CONACYT through the program PRONII. We thank Esteban Vasconcellos for his cooperation and logistical support, the support of the Pykasu Community of the Pueblo Guaraní Ñandeva, and Ivan Benitez, Victor Robles, Javier Ovelar, Carlos Molinas, Patricia Salinas, Paola Cuevas, Johanna Lopez, Christian Palacios, Magdalena Benitez, Agustin Paviolo, José Aguilera, and José Méndez for assistance with data collection.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Thompson, J.J. et al. (2023). Using Camera-Trap Data to Simultaneously Estimate Jaguar (Panthera onca) Density and Resource Selection in the Paraguayan Dry Chaco. In: Mandujano, S., Naranjo, E.J., Andrade Ponce, G.P. (eds) Neotropical Mammals. Springer, Cham. https://doi.org/10.1007/978-3-031-39566-6_10
Download citation
DOI: https://doi.org/10.1007/978-3-031-39566-6_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-39565-9
Online ISBN: 978-3-031-39566-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)