
Abstract
Despite high biodiversity and its importance in provisioning ecosystem services (Corbera et al. 2007; Vörösmarty et al. 2010), Neotropical lotic ecosystems rank among the most threatened in the world (Barlow et al. 2018). The conversion of native land cover, typically forest but also grassland, to human-centered land use represents a primary driver of Neotropical biodiversity loss due to a disproportionately large degree of species richness and land conversion (Barlow et al. 2018).
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

3.1 Introduction
Despite high biodiversity and its importance in provisioning ecosystem services (Corbera et al. 2007; Vörösmarty et al. 2010), Neotropical lotic ecosystems rank among the most threatened in the world (Barlow et al. 2018). The conversion of native land cover, typically forest but also grassland, to human-centered land use represents a primary driver of Neotropical biodiversity loss due to a disproportionately large degree of species richness and land conversion (Barlow et al. 2018).
The loss of native land cover especially in riparian zones exerts a strong impact on lotic ecosystem structure and function (Haddad et al. 2015) due to energetic coupling of streams and rivers with terrestrial vegetation (Webster and Benfield 1986; Marcarelli et al. 2011). Resource subsidies from riparian vegetation provides an important organic matter base to aquatic food webs and physical substrate for benthic aquatic macroinvertebrates. Physical and chemical effects result as elevated solar radiation exposure following vegetation loss increases water temperature (Webster and Benfield 1986; Martins et al. 2017) on lotic macroinvertebrates. Sediment and nutrient concentrations also often increase following vegetation conversion and both environmental stressors typically reduce macroinvertebrate biodiversity (Ferreira et al. 2015). Although the effects of agricultural expansion on the physical, chemical, and macroinvertebrate community properties of stream ecosystems are now well-documented, most studies are concentrated in temperate climates. Here, we review studies exploring the effects of native terrestrial ecosystem conversion to agriculture in the Neotropics with an emphasis on lotic macroinvertebrates.
3.2 Review: Agricultural Expansion and Benthic Macroinvertebrates in Neotropical Streams
Loss of natural cover to agricultural expansion is particularly acute in tropical and subtropical ecosystems due to an increased demand for agricultural products coupled with population growth and advancing economies (Millennium Ecosystem Assessment 2005; Laurance et al. 2014). In the Neotropics, these transformations in land use for agricultural expansion have been extensive. In Brazil, the Colombia Andes, and the Ecuadorian Andes, for example, vegetation loss to agriculture remains a critical threat to lotic ecosystems and loss of megadiversity (Iniguez-Armijos et al. 2014; Chará-Serna et al. 2015; Strassburg et al. 2017).
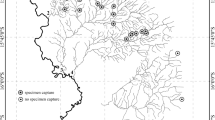
In this chapter, we conducted a qualitative review of studies investigating how agricultural expansion impacted benthic macroinvertebrate communities in headwater Neotropical streams between 23.5° N and 23.5° S. Our review included published studies that quantified entire communities, multimetric indices, and/or those with a focus on focal taxa, most commonly aquatic insects such as Ephemeroptera, Plecoptera, Trichoptera, Odonata, and Diptera - EPTOD that were identified to various taxonomic levels (order, family, genera) (see Table 3.1). We analyzed 34 papers that met geographic and scope criteria. The survey revealed that most studies reflect recent efforts, with the most papers published between 2014 and 2019. Furthermore, studies were unevenly distributed geographically, with study sites primarily located in: Brazil (n = 25) and Ecuador (n = 5) (Figs. 3.1 and Table 3.1). Most Brazilian studies were focused on the southeastern region of the country, where universities and biodiversity researchers are concentrated (Clarivate Analytics, Research in Brazil 2017). Large Neotropical countries, such as Mexico and Peru, apparently lack recent investigations of how agricultural land use conversion impacts lotic macroinvertebrates.

Map showing the distribution of the studies in the Neotropical region
Such geographic disparities also reflect ecological concentrations of effort. Among ecosystems covered, Atlantic tropical moist forest (= 11) and the Brazilian savanna (Cerrado biome) (=10) constituted the study sites for over half of those surveyed. The Atlantic Forest and Cerrado are considered biodiversity hotspots with high endemism (Myers et al. 2000) and rank among the most threatened ecosystems globally (Beuchle et al. 2015; Hunke et al. 2015). Agriculture and pasture cattle grazing converted 88% and 80% of native vegetation in the Atlantic Forest and Cerrado, respectively (Ribeiro et al. 2009; Ferreira et al. 2012; Strassburg et al. 2017). The large number of studies may be related to the need for knowledge about the effects of landscape conversion on streams in these ecosystems (Roque et al. 2021; Albuquerque et al. 2021).
Streams in the Atlantic Forest are under a long and historic degradation due to landscape conversion and many fish and aquatic macroinvertebrates have been largely reduced in this biome (Dala-Corte et al. 2020). Likewise, the Cerrado is the second largest biome in Brazil (after the Amazon) and represents about 23% of the country (Ratter et al. 1997) and has been strongly altered by pasture and crop agriculture (Hunke et al. 2015). It harbors many important large rivers, and its network of headwater streams contain a large diversity of species and ecosystem services (Strassburg et al. 2017).
The impacts of agriculture on Neotropical streams have also been documented in studies in the Brazilian Amazon, montane rainforest, and the tropical moist forest in the Ecuadorian Choco (Chará-Serna et al. 2015; Brito et al. 2019; Morabowen et al. 2019; Lima et al. 2020). In the Brazilian Amazon, for example, agriculture expansion has resulted in the loss of about 18% of the native forest ecosystems (Ferreira et al. 2012) and had considerable impacts on the habitat structure and community composition of many small Amazon streams (Leal et al. 2016).
Similar to related efforts in other regions, a strong majority of studies in Neotropical streams indicated that the loss of native vegetation associated with agricultural practices consistently impacts the structure and composition of aquatic invertebrate communities (Table 3.1). As many studies from temperate zones have demonstrated (e.g., Lammert and Allan 1999; Sponseller et al. 2001; Sweeney et al. 2004), multimetric indices, diversity, and richness tend to decline as agricultural land cover increases in Neotropical watersheds (i.e., Nessimian et al. 2008; Siegloch et al. 2014; Chará-Serna et al. 2015). Community structure also tends to shift toward more homogeneous assemblages dominated by tolerant individuals (Macedo et al. 2014; Brito et al. 2019; Firmiano et al. 2019). Such findings are consistent with works conducted elsewhere, suggesting that the ecological impacts of converting natural cover to agriculture are globally consistent. We also found that in many Neotropical streams, total macroinvertebrate abundance often increases following forest cover loss (Buss et al. 2004). Most studies attributed greater abundance to an increase in light that elevated primary production, allowing grazers to thrive (Bojsen and Jacobsen 2003). Unlike in temperate regions, primary production in Neotropical streams in most cases is not limited by low temperatures and seasonal fluctuations in light.
Although agricultural land use conversion consistently alters macroinvertebrate assemblages in Neotropical streams, a notable minority of studies did not detect changes in taxonomic composition or richness along changes in spatial and temporal gradients of agricultural cover. Examples span diverse ecological settings, such as karstic forest streams in Brazil (Corrêa et al. 2018, 2019), high-elevation montane forests of Ecuador (Ordóñez 2011; Bücker et al. 2010), and the coffee-growing region of Colombia (Chará et al. 2007). Although works that do detect richness and diversity loss are more common, findings of resilience are encouraging because they suggest that biodiversity can persist despite land cover conversion. Many watersheds (e.g., karstic streams) or ecological attributes could help to diminish the effects of land use conversion (Omernik and Bailey 1997).
Scientific recommendations have been made to conserve lotic ecosystems and to reduce threats to biodiversity (Azevedo-Santos et al. 2021). Given the high degree of landscape change by agriculture, pasture, and other land uses in the Neotropical region. Such recommendations have emphasized the need to create stream conservation strategies and the rehabilitation of disturbed areas, particularly those altered by agricultural activities and ranching. Studies emphasize that governmental agencies need to enforce environmental laws to ensure that riparian vegetation is preserved, maintenance and rehabilitation of the ecological condition of water bodies, and the conservation of aquatic biodiversity (Noss 1999) as well as national and international efforts to safeguard the freshwater life of this hyperdiverse region.
3.3 Case Study: Bodoquena Plateau, Brazil
To illustrate how stream ecosystem responses can be context-specific and nuanced, we present a brief case study of macroinvertebrate community changes in a karstic watershed of tropical Central-West of Brazil. Study sites typify severe land use change in the region, yet we detected limited community structure changes in macroinvertebrates even within streams with radically altered watersheds. Although the effort was part of a larger study designed to explore how macroconsumers such as fish and shrimp induce trophic cascades (see Correa et al. 2018, 2019), the design also allows for comparisons of macroinvertebrate communities along a gradient of native vegetation in riparian buffers.
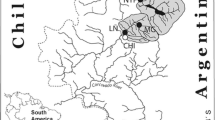
The Bodoquena Plateau (19° 45′ e 22° 15′ S, 57° 30′ e 56° 15′ W) has an area of 890,68 km2 and is located in the transition between Cerrado and Atlantic Forest. The hydrography is partially underground with sinks, calcareous tuff, and cave formations, resulting in clear water with high levels of dissolved calcium and bicarbonate. These karst aquatic systems are extensive in Brazil (Sallun-Filho and Karmann 2007). The rivers of the Bodoquena Plateau drain the karst watersheds into the Miranda River, a subunit of the Upper Rio Paraguay Basin, which includes the Pantanal; one the most biodiverse wetlands in the world (Junk et al. 2011). Native vegetation has been rapidly fragmented by cattle ranching and agriculture with predominance of grain monoculture (soybeans and maize), resulting in a mosaic of land cover that ranges from dense riparian vegetation to deforested pasture (Roque et al. 2016).
In May 2015, we selected seven streams within a gradient of remaining forest cover (20–100% of the remnant forest cover) and were distributed across the Bodoquena region. To investigate potential effects of native vegetation loss on benthic aquatic macroinvertebrates, we deployed ten substrate baskets (20 × 20 × 10 cm with mesh opening of 50 mm) in a single pool along gradients of elevation and vegetation cover among seven karst streams with similar substrate composition. Baskets were filled with limestone to promote colonization.
Most of the chemical water parameters, such as temperature (21.0–22.8 °C) (p = 0.43), pH (7.2–8.1) (p = 0.04), dissolved oxygen (60.0–77.0 mg l−1) (p = 0.140) were similar between the sampled sites. Total nitrogen (1.87–0.85 lg/l) (p = 0.38), total phosphorous (0.179–0.102 lg/l) (p = 0.47), and turbidity (80–100 NTU) (p = 0.63) were also not correlated with the vegetation cover gradient.
We found a total of 2465 benthic macroinvertebrates comprising 25 families in the stream sampling along the gradient of vegetation cover. Macroinvertebrate abundance varied more than twofold across sites, with a minimum of 224 individuals found at the Taquaral stream (100% vegetation cover) to a maximum of 600 at the Serra stream (79% vegetation cover). Leptophlebiidae (with 934 individuals), Chironomidae (682), Hydrobiidae (177), and Elmidae (124) were the most abundant families along the gradient cover vegetation. Collector-gatherers-scrapers comprised 56% of total invertebrate abundance, followed by collectors-filterers-gatherers (33%) and predators (11%). Permutational multivariate analyses of variance (PERMANOVA) suggested that benthic macroinvertebrate community composition was unrelated to the vegetation gradient in Bodoquena Plateau.
Our results showed that the vegetational loss gradient had no influence on the macroinvertebrate community composition (Fig. 3.2), in contrast with the results found in other studies involving Neotropical streams (Moretti et al. 2007; Silva-Araujo et al. 2020). Similar studies of multitaxa communities in Bodoquena Plateau streams also did not find detectable responses between terrestrial and aquatic insects across a riparian cover gradient (Roque et al. 2017), and comparisons of fish communities along of the land cover gradient also demonstrated weak or no effects (Casatti et al. 2010). The lack of effect on macroinvertebrates could be attributed to mosaics of vegetation in the Bodoquena Plateau, which may maintain functional connectivity despite land conversion. In karstic systems, the watershed or forest cover delimitation is particularly challenging because surface water does not drain directly into surficial channels. Such a hydrogeomorphic settings likely could decouple the effects of landscape degradation from ecological processes in receiving waters.

Site-specific ln-response ratio (LRR) of total invertebrates’ abundance with respect to the forest cover gradient. Shown are the means of treatment divided by control values ±1 standard error. Sites are ordered from least to most forest cover
Findings from our case study plus collective findings from studies in our qualitative survey suggest that while land use conversion can be expected to impact macroinvertebrate communities in Neotropical streams, observed impacts are likely to be context-specific. Landuse conversion that increases light to stream channels can increase productivity, resulting in elevated macroinvertebrate abundance for grazing taxa (Casatti et al. 2010; Silva-Araújo et al. 2020). Tropical streams are likely to consistently show such patterns because they experience high light and warm temperatures year-round (Boulton et al. 2008). Local hydrogeologic attributes might buffer macroinvertebrate communities from becoming severely degraded, a concept that likely extends beyond tropical regions. Finally, another potential mitigation strategy for minimizing land use effects in streams is maintaining sufficiently wide riparian buffer zones (Azevedo-Santos et al. 2021).
References
Albuquerque LG, Roque FO, Valente-Neto F, Koroiva R, Buss DF, Baptista DF, Hepp LU, Kuhlmann ML, Sundar S, Covich AP, Pinto JOP (2021) Large-scale prediction of tropical stream water quality using rough sets theory. Eco Inform 61:101226
Azevedo-Santos VM, Rodrigues-Filho JL, Fearnside PM, Lovejoy TE, Brito MFG (2021) Conservation of Brazilian freshwater biodiversity: thinking about the next 10 years and beyond. Biodivers Conserv 30:235–241
Barlow J, França F, Gardner TA, Hicks CC, Lennox GD, Berenguer E, Graham NAJ (2018) The future of hyperdiverse tropical ecosystems. Nature 559:517–526
Beuchle R, Grecchi RC, Shimabukuro YE, Seliger R, Eva HD, Sano E, Achard F (2015) Land cover changes in the Brazilian Cerrado and caatinga biomes from 1990 to 2010 based on a systematic remote sensing sampling approach. Appl Geogr 58:116–127
Bojsen BH, Jacobsen D (2003) Effects of deforestation on macroinvertebrate diversity and assemblage structure in Ecuadorian Amazon streams. Archiv fur Hydrobiologie 158:317–342
Boulton AJ, Boyero L, Covich AP, Dobson M, Lake S, Pearson R (2008) Are tropical streams ecologically different from temperate streams? Chapter 9. In: Dudgeon D (ed) Tropical stream ecology. Academic, San Diego, pp 257–284
Brito JG, Roque FO, Martins RT, Nessimian JL, Oliveira VC, Hughes RM, de Paula FR, Ferraz SFB, Hamada N (2019) Small forest losses degrade stream macroinvertebrate assemblages in the eastern Brazilian Amazon. Biol Conserv 241:108263
Bücker A, Sondermann M, Frede HG, Breuer L (2010) The influence of land-use on macroinvertebrate communities in montane tropical streams - a case study from Ecuador. Fundam Appl Limnol 177:267–282
Buss DF, Baptista DF, Nessimian JL, Egler M (2004) Substrate specificity, environmental degradation and disturbance structuring macroinvertebrates assemblages in neotropical streams. Hydrobiologia 518:179–188
Casatti L, Romero RM, Teresa FB, Sabino J, Langeani F (2010) Fish community structure along a conservation gradient in Bodoquena Plateau streams, Central West of Brazil. Acta Limnol Bras 22:50–59
Chará J, Pedraza G, Giraldo L, Hincapié D (2007) Efecto de los corredores ribereños sobre el estado de quebradas em la zona ganadera del río La Vieja, Colombia. Agroforestería em las Américas 45:72–78
Chará-Serna AM, Chará J, Giraldo LP, Zúñiga MC, Allan JD (2015) Understanding the impacts of agriculture on Andean stream ecosystems of Colombia: a causal analysis using aquatic macroinvertebrates as indicators of biological integrity. Freshw Sci 34:727–740
Clarivate Analytics (2017) Research in Brazil. pp 1–73. http://www.sibi.usp.br. Accessed 28 Sept 2021
Corbera E, Kosoy N, Martinez-Tuna M (2007) Equity implications of marketing ecosystem services in protected areas and rural communities: case studies from Meso-America. Glob Environ Chang 17:365–380
Corbi JJ, Trivinho-Strixino S (2006) Influence of taxonomic resolution of stream macroinvertebrate communities on the evaluation of different land uses. Acta Limnol Bras 18:469–475
Cornejo A, Tonin AM, Checa B, Tuñon AR, Pérez D, Coronado E, González S, Rios T, Macchi P, Araneda FC, Boyero L (2019) Effects of multiple stressors associated with agriculture on stream macroinvertebrate communities in a tropical catchment. PLoS One. https://doi.org/10.1371/journal.pone.0220528
Corrêa EC, Roque FO, Utz RM, Correa JS, Souza FL, Covich AP (2018) Effects of macroconsumers on benthic communities: rapid increases in dry-season accrual of calcium in a tropical karst stream. PLoS One 13:e0209102
Corrêa EC, Roque FO, Utz RM, da Silva WM, Severo-Neto F, Gido KB (2019) Effects of macroconsumers on benthic communities across a gradient of vegetation loss in tropical karst streams. Hydrobiologia 836:21–34
Dala-Corte R, Melo AS, Siqueira T, Bini LM, Martins RT, Cunico AM, Pes AM, Magalhães ALB et al (2020) Thresholds of freshwater biodiversity in response to riparian vegetation loss in the Neotropical region. J Appl Ecol. https://doi.org/10.1111/1365-2664.13657
De Castro DMP, de Carvalho DR, Pompeu PS, Moreira MZ, Nardoto GB, Callisto M (2016) Land use influences niche size and the assimilation of resources by benthic macroinvertebrates in tropical headwater streams. PLoS One. https://doi.org/10.1371/journal.pone.0150527
De Castro DMP, Dolédec S, Callisto M (2018) Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol Indic 84:573–582
De Oliveira-Junior JMB, Shimano Y, Gardner TA, Hughes RM, Júnior PDM, Juen L (2015) Neotropical dragonflies (Insecta: Odonata) as indicators of ecological condition of small streams in the eastern Amazon. Austral Ecol 40:733–744
Encalada AC, Calles J, Ferreira V, Canhoto CM, Graca MAS (2010) Riparian land use and the relationship between the benthos and litter decomposition in tropical montane streams. Freshw Biol 55:1719–1733
Ferreira J, Pardini R, Metzger JP, Fonseca CR, Pompeu PS, Sparovek G, Louzada J (2012) Towards environmentally sustainable agriculture in Brazil: challenges and opportunities for applied ecological research. J Appl Ecol 49:535–541
Ferreira V, Castagneyrol B, Koricheva J, Gulis V, Chauvet E, Graça MAS (2015) A meta-analysis of the effects of nutrient enrichment on litter decomposition in streams. Biol Rev 90:669–688
Ferreira-Peruquetti P, De Marco P Jr (2002) Efeito da alteração ambiental sobre comunidades de Odonata em riachos de Mata atlântica de Minas Gerais, Brasil. Revista Brasileira de Zoologia 19:317–327
Firmiano KR, Ligeiro R, Macedo DR, Juen L, Hughes RM, Callisto M (2017) Mayfly bioindicator thresholds for several anthropogenic disturbances in neotropical savanna streams. Ecol Indic 74:276–284
Firmiano KR, Cañedo-Argüelles M, Gutiérrez-Cánovas C, Macedo DR, Linares MS, Bonada N, Callisto M (2019) Land use and local environment affect macroinvertebrate metacommunity organization in Neotropical stream networks. J Biogeogr:1–13
Guimarães-Souto RM, Facure KG, Jacobucci GB (2021) Do tropical riparian forests in the Cerrado Biome act as a buffer against the impacts of agriculture and livestock on benthic macroinvertebrate communities? Limnetica 40:329–342
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song DX, Townshend JR (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:e1500052
Hunke P, Mueller EN, Schröder B, Zeilhofer P (2015) The Brazilian Cerrado: assessment of water and soil degradation in catchments under intensive agricultural use. Ecohydrology 8:1154–1180
Iñiguez–Armijos C, Leiva A, Frede HG, Hampel H, Breuer L (2014) Deforestation and benthic indicators: how much vegetation cover is needed to sustain healthy Andean streams? Plos one 9:e105869
Junk WJ, da Silva CJ, Nunes da Cunha C, Wantzen KM (2011) The Pantanal: ecology, biodiversity and sustainable management of a large Neotropical seasonal wetland. Pensoft Publishers, Sofia, p 870
Lammert M, Allan JD (1999) Assessing biotic integrity of streams: effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates. Environ Manag 23:257–270
Laurance WF, Sayer J, Cassman KG (2014) Agricultural expansion and its impacts on tropical nature. Trend Ecol Evol 29:107–116
Leal CG, Pompeu PS, Gardner TA, Leitão RP, Hughes RM, Kaufmann PR, Zuanon J, de Paula FR, Ferraz SFB, Thomson JR, Mac Nally R, Ferreira J, Barlow J (2016) Multi-scale assessment of human-induced changes to Amazonian instream habitats. Landsc Ecol. https://doi.org/10.1007/s10980-016-0358-x
Ligeiro R, Hughes RM, Kaufmann PR, Macedo DR, Firmiano KR, Ferreira WR, Oliveira D, Melo AS, Callisto M (2013) Defining quantitative stream disturbance gradients and the additive role of habitat variation to explain macroinvertebrate taxa richness. Ecol Indic 25:45–57
Lima DVM, Please LPM, Silva IHL (2020) Effects of land use on the community of benthic macroinvertebrates in streams of the Iquiri River basin (Acre, Brazil). South Am J 7:160–175
Lorion CM, Kennedy BP (2009) Relationships between deforestation, riparian forest buffers and benthic macroinvertebrates in neotropical headwater streams. Freshw Biol 54:165–180
Macedo DR, Hughes RM, Ligeiro R, Ferreira WR, Castro MA, Junqueira NT, ... & Callisto M (2014) The relative influence of catchment and site variables on fish and macroinvertebrate richness in cerrado biome streams. Landsc Ecol 29(6):1001–1016. https://doi.org/10.1007/s10980-014-0036-9
Marcarelli AM, Baxter CV, Mineau MM, Hall RO (2011) Quantity and quality: unifying food web and ecosystem perspectives on the role of resource subsidies in freshwaters. Ecology 92(6): 1215–1225. https://doi.org/10.1890/10-2240.1
Martins RT, Couceiro SRM, Melo AS, Moreira MP, Hamada N (2017) Effects of urbanization on stream benthic invertebrate communities in Central Amazon. Ecol Indic 73:480–491
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island Press, Washington, p 137
Morabowen A, Crespo-Pérez V, Ríos-Touma B (2019) Effects of agricultural landscapes and land uses in highly biodiverse tropical streams of the Ecuadorian Choco. Inland Waters. https://doi.org/10.1080/20442041.2018.1527597
Moraes AB, Wilhelm AE, Boelter T, Stenert C, Schulz UH, Maltchik L (2014) Reduced riparian zone width compromises aquatic macroinvertebrate communities in streams of southern Brazil. Environ Monit Assess 186:7063–7074
Moretti MS, Gonçalves JF Jr, Ligeiro R, Callisto M (2007) Invertebrates colonization on native tree leaves in a neotropical stream (Brazil). Int Rev Hydrobiol 92:199–210
Myers N, Mittermeyer RA, Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Nessimian JL, Venticinque EM, Zuanon J, Marco P Jr, Gordo M, Fidelis L, Batista JD, Juen L (2008) Land use, habitat integrity, and aquatic insect assemblages in Central Amazonian streams. Hydrobiologia 614:117–131
Noss RF (1999) Assessing and monitoring forest biodiversity: a suggested framework and indicators. For Ecol Manag 115:135–146
Omernik JM, Bailey RG (1997) Distinguishing between watersheds and ecoregions. J Am Water Resour Assoc 33:935–949
Ometo JPHB, Martinelli LA, Ballester MV, Gessner A, Krusche AV, Victoria RL, Williams M (2000) Effects of land use on water chemistry and macroinvertebrates in two streams of the Piracicaba river basin, south-east Brazil. Freshw Biol 44:327–337
Ongaratto R, Loureiro RC, Restello RM, Hepp LU (2018) Effects of land use and limnological variables on the dissimilarity of common and rare aquatic insects in Atlantic Forest streams. Rev Biol Trop 66:1223–1231
Ordóñez MV (2011) Influencia del uso de suelo y la cobertura vegetal natural en la integridad ecológica de los ríos altoandinos al noreste del Ecuador. MS Thesis, Universidad San Francisco de Quito, Quito
Ratter J, Ribeiro J, Bridgewater S (1997) The Brazilian cerrado vegetation and threats to its biodiversity. Ann Bot 80:223–230
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni F, Hirota MM (2009) Brazilian Atlantic forest: how much is left and how is the remaining forest distributed? Implications for conservation. Biol Conserv 142:1141–1153
Rodrigues ME, Roque FO, Quintero JMO, Pena JCC, Sousa DC, Junior PM (2016) Nonlinear responses in damselfly community along a gradient of habitat loss in a Savanna landscape. Biol Conserv 194:113–120
Roque FO, Ochoa-Quintero J, Ribeiro DB, LSM S, Costa-Pereira R, Lourival R, Bino G (2016) Upland habitat loss as a threat to Pantanal wetlands. Conserv Biol 30:1131–1134
Roque FO, Corrêa EC, Valente-Neto F, Stefan G, Schulz G, Souza PRB, Motta CM, Bavutti LLO, Colzani E, Demétrio MF, Escarpinati SC, Silvestre R, Vaz-de- Mello FZ, Siqueira T, Quintero JMO (2017) Idiosyncratic responses of aquatic and terrestrial insects to different levels of environmental integrity in riparian zones in a karst tropical dry forest region. Aust Entomol 56:459–465
Roque FO, Guerra FA, Johnson M, Padovani C, Corbi J, Covich AP, Eaton D, Tomas WM et al (2021) Simulating land use changes, sediment yields, and pesticide use in the Upper Paraguay River Basin: implications for conservation of the Pantanal wetland. Agric Ecosyst Environ 314:107405
Sallun Filho W, Karmann I (2007) Geomorphological map of the Serra da Bodoquena karst, west-central Brazil. J Maps 3:282–295
Sensolo D, Hepp LU, Decian V, Restello RM (2012) Influence of landscape on assemblages of Chironomidae in Neotropical streams. Ann Limnol Int J Limnol 48:391–400
Siegloch AE, Suriano M, Spies M, Fonseca-Gessner A (2014) Effect of land use on mayfly assemblages structure in Neotropical headwater streams. Ann Brazilian Acad Sci 84:1735–1747
Silva-Araújo M, Silva-Junior EF, Neres-Lima V, Feijó-Lima R, Tromboni F, Lourenço-Amorim C, Thomas SA, Moulton TP, Zandonà E (2020) Effects of riparian deforestation on benthic invertebrate community and leaf processing in Atlantic forest streams. Perspect Ecol Conser 18:277–282
Siqueira T, Lacerda CG-LT, Saito VS (2015) How does landscape modification induce biological homogenization in tropical stream metacommunities? Biotropica 47:509–516
Sponseller RA, Benfield EF, Valett HM (2001) Relationships between land use, spatial scale and stream macroinvertebrate communities. Freshw Biol 46:1409–1424
Strassburg BBN, Brooks T, Feltran-Barbieri R, Iribarrem A, Crouzeilles R, Loyola R, Latawiec AE, Oliveira Filho FJB, Scaramuzza de CAM, Scarano FR, Soares-Filho B, Balmford A (2017) Moment of truth for the cerrado hotspot. Nat Ecol Evol 1:99
Suga CM, Tanaka MO (2013) Influence of a forest remnant on macroinvertebrate communities in a degraded tropical stream. Hydrobiologia 703:203–213
Sweeney BW, Bott TL, Jackson JK, Kaplan LA, Newbold JD, Standley LJ, Hession WC, Horwitz RJ (2004) Riparian deforestation, stream narrowing, and loss of stream ecosystem services. Proc Natl Acad Sci 101:14132–14137
Tanaka MO, De Souza ALT, Mischini LE, De Oliveira AK (2016) Influence of watershed land use and riparian characteristics on biological indicators of stream water quality in southeastern Brazil. Agric Ecosyst Environ 216:333–339
Torres PJ, Ramírez A (2014) Land use effects on leaf litter breakdown in low-order streams draining a rapidly developing tropical watershed in Puerto Rico. Rev Biol Trop 62:129–142
Villada-Bedoya S, Ospina-Bautista F, G-Dias L, Varón JVE (2017) Diversidad de insectos acuáticos en quebradas impactadas por agricultura y minería, Caldas, Colombia. Revista de Biologia Tropical 65:1635–1659
Vörösmarty CJ, McIntyre PB, Gessner MO, Dudgeon D, Prusevich A, Green P, Glidden S, Bunn SE, Sullivan CA, Liermann CR, Davies PM (2010) Global threats to human water security and river biodiversity. Nature 46:555–561
Webster JR, Benfield EE (1986) Vascular plant breakdown in freshwater ecosystems. Annu Rev Ecol Evol Syst 17:567–594
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Corrêa, E.C., Utz, R.M. (2023). Effects of Agricultural Expansion on Lotic Benthic Macroinvertebrate Communities: A Review and Case Study from Brazil. In: Myster, R.W. (eds) Neotropical Gradients and Their Analysis. Springer, Cham. https://doi.org/10.1007/978-3-031-22848-3_3
Download citation
DOI: https://doi.org/10.1007/978-3-031-22848-3_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-22847-6
Online ISBN: 978-3-031-22848-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)