
Abstract
The history of craniofacial evolution is written on the neck. Here one finds multiple systems communicating between the head and trunk. The invention of the neck was fundamental for the transition to terrestrial life with multiple innovations including a redefinition of head versus trunk, expansion of neurosensory systems for vision and hearing, the capacity to ambulate, and improved systems of prey capture. This chapter is truly interdisciplinary. It is designed to give you not only a neuromeric interpretation of structure but a historical appreciation of how the neck evolved over time. Our learning objective is to achieve an integrated model of a specific component of craniofacial and neck anatomy as part of a single embryologic process. These fundamentals will prove of great value to understand clinical problems that occur in this region.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Hypobranchial muscles
- Trapezius
- Sternocleidomastoid
- Cucullaris
- Strap muscles
- Cervical plexus
- Occipital and cervical somites
- Vertebral artery
- Thyrocervical trunk
- Costocervical trunk
- Branchial plexus
Introduction
A structural analysis of the muscles of “the neck” is somewhat arbitrary; it depends upon one’s concept of the region and encompasses a variety of structures, many with evolutionary changes, incurred with the transition to life on land. Cervical vertebrae were initially three which we mammals have expanded to seven. The larynx and trachea were invented while the primitive gut became the esophagus. Glands such as the thyroid were relocated. Cranial nerve XI resulted from the forward incorporation of a multi-segment peripheral nerve. In discussing the muscles of the neck, we are making a neuromeric transition between muscles arising from somitomeres and those arising from somites. With the important exceptions of sternocleidomastoid and trapezius, all muscles in this chapter are innervated by peripheral nerves. For purposes of this discussion, we will confine ourselves to those muscles supplied by XI and cervical nerves 1–8. We therefore bid adieu to the floor of mouth, the suprahyoid series, and to the muscles of the pharynx and larynx. But there will be obligatory cross-talk regarding SCM and trapezius and their chaperone, the platysma.
This chapter, like its predecessor, makes use of the motor column model to organize muscles into functional groups. But, if we hope to understand the neck in a deeper, more conceptual way, we shall have to give up the segmental comforts of cranial nerves and pharyngeal arches. Neck anatomy is the sine qua non of tetrapod existence. It is all about change and innovation. New systems of respiration and locomotion get their start here. Dermatomes are stretched out into seemingly irrational patterns. Muscles have new roles: some are organized in a garden-variety fashion around the head and spinal column for axial control. Others act as “bridges” to the outboard pectoral girdle. Still another muscle, subcoracoideus, an elevator of coracoid and antagonist of pectoralis, is reincarnated in mammals as the diaphragm.
The evolutionary pressures and mechanisms responsible for this mammalian plan are important to emphasize. Therein, we shall plunge back into the comfortable world of segmentation, based upon the basic vertebrate bauplan: the fish body. We will examine in greater detail how the anatomy of the forefin explains the development of the shoulder girdle. We can gain some insight into the important new role played by lateral plate mesoderm in constructing the trachea and esophagus. Interesting correlations about the twin trajectories of thyroid and thymus will be sought. And, of course, all that is terribly relevant for our study of neck muscles. So … the reader is advised to simply get through this chapter … Subsequently we shall discuss neuromuscular evolution. Subsequently we shall discuss neuromuscular evolution. Feel free to bounce back and forth among the sections of this chapter. In this way, the correlations will leap out at you.
The first section of this chapter will examine the osteology of the region. We will discuss the bones of the occipito¬cervical junction (occipital, proatlas, atlas, and axis), as well as typical cervical vertebrae (C7), paying particular attention to their mechanisms of development. Next, we shall look at the development of the clavicle and scapula. When one combines information regarding the origins of individual muscles from their respective somite (s) with the ossification sequence of the individual bones, the spatial arrangement of muscle binding sites follows a quite predictable neuromeric pattern.
The second section is myology. Five distinct neuromuscular systems are involved: neck to head, neck to neck, neck to trunk, neck to shoulder girdle, and that most unique mammalian invention, the diaphragm. We will organize our discussion about the muscles on the basis of functional groups and motor columns (medial central and lateral). The vital role of the fascia will be emphasized. The origin of the mammalian.
The third section, neurology, reviews the general layout of the cervical spinal cord (emphasizing the motor column model) and the spinal nerves. The cervical plexus will be discussed in detail. Owing to our craniofacial focus (and for the sake of brevity), the brachial plexus will be declared “out of bounds.”
The fourth section, angiology, will explore the rationale behind the arterial supply to the neck. Far from being nonsensical, the vascular system is a logical outcome of segmental developmental processes.
The fifth section, systems analysis, explores important structures that pass through the neck into the chest: trachea, esophagus, endocrine glands, thyroid, and thymus.
As a final note, the overarching theme of the neck is that it represents evolution at work. A craniovertebral joint is created. Vertebral specialization allows for better positioning of the head. Even the length of the neck is dynamic. Central to these changes are shifts in homeotic coding along the craniocaudal axis. These redefine the anatomic contents at various neuromeric levels. Formerly cranial muscles subdivide, with some remaining in place while others are displaced backward into the “new neck.” The brachial plexus relocates and gets larger, that most curious of muscles, the cucullaris, assumes a radical reiteration in mammals, allowing us to oxygenate more efficiently. So keep the hox code in the back of your mind as you enter into the brave new world of the neck.
Drawing the Line: How Do We Define the Neck?
The neck can be conceptualized in various ways. As an expropriation of the rostral trunk, it serves as a simple passageway for pre-existing organ systems between head and trunk. The neck is also a functional entity unto itself. It ensures the positioning of the head and its organs of perception for survival functions such as finding and grabbing a meal…or avoiding becoming someone else’s lunch. Finally, cervical muscles are important for control of the upper extremity: pectoral girdle, arm, forearm, and hand. The list of muscles and bones associated with cervical neuromeres becomes very lengthy. Since our emphasis is craniofacial, where shall we draw the line?
First off, we’ll cover all muscles acting on the neck itself. That means that muscles spanning from the neck up to the head and down to the trunk are fair game. The pectoral girdle is also a target in our crosshairs. It provides support for pharyngeal arch structures (the strap muscles) and connects to the skull. Finally, cervical muscles attach the pectoral girdle to the neck and trunk All remaining structures of the upper extremity are out-of-bounds…with one important exception, the diaphragm, which, as we shall see, represents an evolutionary subversion of a pectoral girdle muscle, subcoracoideus.
Rest assured, all muscles of the upper limb follow the same neuromeric rules: they simply are beyond our scope. One can only hope some reader with a keen interest in the extremity will apply these concepts to better understand the appendicular skeleton and teach the rest of us.
-
In sum: the scope of our inquiry includes all structures associated with cervical neuromeres c1 through c8 with the exception of the upper extremity distal to the clavicle and the scapula.
Organization of Cervical Mesenchyme
At multiple occasions, we have discussed how epiblast cells acquire an identity during gastrulation. Those that ingress through either the lateral aspect of primitive node or the rostral zone of primitive streak will become committed to the somitic cell line. After traversing the streak these cells maintain contact with the basal laminae of both the overlying epiblast and underlying hypoblast. Recall our previous discussion in which cell movements in gastrulation are liken to the flow of toothpaste. The cells nearest the streak dedifferentiate, detach, and dive inward. They attach immediately, pushing the primitive endoderm (hypoblast) laterally. Subsequent cells play “leapfrog,” over their predecessors, migrate laterally and attach at the first available position. Recall the first wave of cells to ingress represents the chordamesoderm. These cells kick the hypoblast to either side and settle dead center, where they will ultimately form the notochord. The next wave of cells represents true endoderm. It pushes the hypoblast completely out from beneath epibast. The third wave of cells is the embryonic mesoderm.
Once the cells are in place, and the hypoblast has been pushed out of the embryo, the mesoderm closest to the midline, referred to as paraxial mesoderm (PAM), flanking the notochord. These cells acquire the homeotic code of the neural plate with which they are in register. This creates their anterior-to-posterior identity. They also acquire signals that will confer additional transverse spatial identities that become important during somitogenesis. Those that entered through lateral Hensen’s node become the medial halves of the somites, whereas those that entered 200 μm posteriorly through rostral streak become the lateral halves of the somites. This implies that the first wave of cells occupies the medial position; those that enter later are forced outside. Like students filing into an auditorium, latecomers must file past those already seated. When we later examine the development of the occipitocervical junction, we shall see that medial and lateral zones of the somites have different behaviors.
Intermediate mesoderm exists as a column of extending from neuromere c1 down through the lumbar-sacral neuromeres. IM is the source of the genitourinary system. It bears homeotic segmentation in register with paraxial mesoderm. This is manifested as a series of nephrotomes, one for each neuromeric level, and each with its segmental artery. At a metaneuromeric level, the embryonic kidney takes three iterations as it “migrates” caudally from the neck: pronephros, mesonephros, and metanephros. Aberrancies in the cervical spine have a correspondingly high incidence of genitourinary anomalies.
Lateral plate mesoderm does not display overt segmentation but its organization is homeotic. It has two layers: the somatic lamina (LPMS) is associated with ectoderm; the visceral lamina (LPMV) is associated with endoderm. Lateral plate mesoderm is organized neuromerically into two broad zones.
-
Cardiac LPM is a single mesenchymal mass in register with r0–r11. Its role is the formation of the heart, great vessels, and the blood supply to the head. Both LPMS and LPMV are involved in cardiac formation. LPMV forms aortic arches organized according to pairs of rhombomeres.
Posterior LPM begins at neuromere c1. Its role is the formation of the appendicular skeleton, the post-cranial vascular system, smooth muscle, and viscera. PLMP splits into two laminae: somatic lateral plate mesoderm LPMS and visceral lateral plate mesoderm. LPMS and LPMV cannot fuse together. The space between the layers is the intraembryonic coelom (IEC).
Lateral plate LPMV from c1 downward wraps around the GI tract to form its musculature and surrounds the larynx to envelop the respiratory diverticulum as the substance of the developing lungs. This mesenchyme drops down into the chest on either side of the heart, and descends from craniofacial LPM mesenchyme. For this reason, congenital malformations or aplasia of the lungs within the chest can co-exist in the presence of an intact heart.
Lateral plate LPMS from c1 to t1 relates to the upper extremity. LPMS from c1 to c6 forms pectoral girdle as the core of clavicle and 100% of scapula. LPMS from c5 to t1 forms the bones of the upper extremities. Its vascular core is surrounded by LPMV fascia.
From the above, the intraembryonic coelom begins at level c1 but manifests itself at level c4. How? The upper extremity develops as a diverticulum from levels c4 to t1. The skin envelope has PAM dermis. Just inside the envelope are PAM myoblasts. LPMS bones are at the center, accompanied by LPMV blood vessels. PAM muscles and their fascia attach to LPMS bones but they never fuse with LPMV fascia. For this reason, vascular pedicles always remain distinct from the muscle units. Furthermore, LPNV vascular pedicles never fuse with the LPMS periosteum. Based on these facts we conclude the following:
-
The intraembryonic coelom begins at c4 and extends downward into the chest.
-
The IEC is not seen in the neck because is expropriated by the upper limb.
-
The upper boundary of IEC is c4 and is subcutaneous in the neck behind the clavicle as the cupula which receives the apex of the lung.
The innervation of LPM gives us a clue as to its neuromeric anatomy. All lateral plate derivatives are hypaxial and are innervated by ventral roots. Furthermore, these are always organized as plexuses: cervical, brachial, celiac, superior mesenteric, inferior mesenteric, and lumbo-sacral. What that means is that LPM populations from individual neuromeres migrate in complex 3-D ways. They are not organized in a simple linear order, as are those from the somites. The clinical implication of this is that an event at a single neuromeric level might manifest itself within lateral plate mesoderm at several different levels.
A partial exception to this pattern is the innervation to LPM in the neck. Here, dorsal motor nucleus of vagus supplies esophagus but, as yet, individual segments have not been traced back to a specific rhombomere. Sympathetics from c1 to c3 through the superior cervical ganglion, from c4 to c5 through the middle cervical ganglion, and from c6 to c8 through the inferior cervical ganglion are distributed to the myenteric plexuses in their respective regions of the esophagus.
Neural Crest in the Neck
The role of neural crest cells in the neck is much more limited. In the upper neck they contribute to the cartilage structures of the fourth and fifth arches. The fasciae and connective tissues of branchiomeric muscles, including sternocleidomastoid and trapezius, are all neural crest whereas in all remaining cervical muscles these tissues are PAM. Neural crest is fundamental for the sympathetic nervous system and peripheral nerves throughout the body. Cervical dermis is PAM but contains neural crest melanocytes. The axial skeleton is exclusively PAM but the pectoral girdle contains neural crest elements: the cortex of clavicle and the spine of the scapula.
Osteology: Axial Bones
Why begin with the bones? Let us take a deep breath and a step backward. The spinal column represents the very core of the vertebrate body. It is also the world’s best place to observe the segmentation process at work. As we begin our story of vertebral assembly, we are going to start at the beginning, at its very first step: the segmentation of paraxial mesoderm. The next step is somitogenesis. Here, we shall see how vertebral bodies are formed from opposing halves of adjacent somite, a process termed parasegmentation. We shall then consider how each somite subdivides into functionally distinct units (sclerotome, myotome, and dermatome). Once these concepts are reviewed, we’ll look at the actual assembly process of “prototypical” vertebrae. This will be extended to the anomalous atlas and axis. Finally, we will delve into how the mammalian craniofacial junction evolved.
Certain evolutionary questions loom large here. Mammals have eight cervical somites but only seven vertebrae? Why do cervical nerves emerge above their respective vertebral arch, whereas spinal nerves at all other levels must pass underneath? Why on earth should strap muscles supplied by the cervical plexus insert on the sternum and clavicle? Why are the motor targets of spinal accessory nerve so far removed from the head and neck? I will beg the reader’s indulgence for any redundancy. The twists and turns of evolution are quite fascinating and terribly relevant. You won’t be bored (Figs. 10.1 and 10.2).

All mammals, regardless of size, have seven vertebrae. Compare the okapi versus giraffe. (a) brontosaurus, (b) velociraptor, (c) giraffe, (d) okapi, (e) human. Left: [Reprinted from Vidal D, Mocho P, Páramo A, Sanz JL, Ortega F (2020) Ontogenetic similarities between giraffe and sauropod neck osteological mobility. PLoS ONE 15(1): e0227537. https://doi.org/10.1371/journal.pone.0227537. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by-sa/4.0/deed.en.] Right: [Reprinted from Wikimedia. Retrieved from https://commons.wikimedia.org/wiki/File:Okapi_Giraffe_Neck.png. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by-sa/4.0/deed.en]

Camarasaurus. Sauropods had 11–19 cervical vertebrae. Backward-pointing cervical ribs are seen here. In mammals, these are bicipetal, fused to the vertebrae as transverse processes through which pass the vertebral artery, a longitudinal fusion of previous segmental cervical arteries. Note: amniotes divide into diapsids (reptiles and birds) and synapsids (mammals). In synapsids the number of cervical vertebrae stabilizes at the level of cynodonts to 7 whereas in diapsids, along the avian line, 8 or more are possible. [Reprinted from M Taylor MP, Wedel MJ, and Cifell RL. A new sauropod dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. Acta Palaeontologica Polonica 2011;56(1): 75–98. doi: https://doi.org/10.4202/app.2010.007. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by/4.0/]
The fundamental story of the mammalian cervical spine is the production of 8 vertebrae, Proatlas, the first cervical vertebra, has a complex evolutionary history in which it fragments into component parts to create the craniovertebral joint, that is, the atlas–axis complex. The fourth through the eighth cervical vertebrae repeat a basic developmental plan which is continued all the way to the sacrum. We shall discuss the osteology of the axial spine in terms of its building blocks (paraxial mesoderm and the somites), how these are assembled to form vertebrae, ossification centers of the vertebra as clues to developmental components, the developmental anatomy of the craniovertebral junction, and finally the individual bones beginning with the cranial base.
Somitogenesis
Immediately after gastrulation, paraxial mesoderm is a loose mesenchyme without cellular polarities. This presomitic mesoderm (PSM) quickly organizes itself due to the elaboration of cell adhesion molecules, such as fibronective and N-cadherin, a process known as compaction. The cells condense into somitomeres consisting of a basal lamina surrounding a single layer of epithelial cells with polarized organelles oriented radially around a central cavity (somitocoele) that contains mesenchymal cells. Craniofacial PSM has two zones with two distinct fates. The anterior zone extends from midbrain back to the otic vesicle and is registered with mesomeres m1–m2 and rhombomeres r0–r7 and is sometimes referred to as head mesoderm. It remains in the somitomeric state. The first seven somitomeres produce craniofacial muscles of the first three arches, the post-hypophyseal cranial base, and the primary blood vessel systems of the head and face. The posterior zone extends from the otic vesicle to the craniocervical junction, is in register with r8–r11, and undergoes transformation to somites (Figs. 10.3, 10.4, 10.5, 10.6).

Somitomeres and somites. Somitomeres and somitogenetic genes. The morphological pattern of somitomeres in the chick embryo is shown on the right, as a simplified illustration. Hypothetical somitomeres are numbered. On the left is shown the oscillating expression of a somitogenetic gene, chairy, in the early chick embryo.. Each oscillation is numbered together with the mesodermal part generated after that oscillation. Note that there are only two oscillations in the head mesoderm, one for the premandibular mesoderm, and the other for the rest of the cephalic mesoderm. Birds have 5 occipital somites and mammals have 4. Both birds and mammals have 7 somitomeres. These model, are physically confluent, have no internal compartments, and produce striated muscles only Somitomeres 1–3 and 5 are exclusive to the orbit. Somitomeres 4, 6, and 7 provide pharyngeal arches 1, 2, and 3 with mesoderm. is The fourth and fifth pharyngeal arches, not shown here receive banchiomeric mesoderm from somitomeres 8–11 prior to their reformatting into occipital somites with defined myotomes. Sm1–Sm4 produce the hypobranchial muscles of the tongue. [Reprinted from Kuratani S. Craniofacial development and the evolution of the vertebrates: the old problems on a new background. Zoological Sciences 2005; 22: 1–19. With permission from The Zoological Society of Japan]

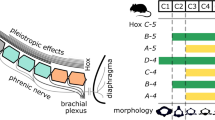
Somite distribution is controlled by Hox genes. Chick versus mouse pattern. Like all mammals, mice have 7 cervical vertebrae. Birds have longer necks. The ostrich Cervcal-thoracic boundary is Hox5 versus Hox6. [Reprinted from Gilbert SF, Barressi MJF. Developmental Biology, 11th ed. Sunderland, MA: Sinauer; 2016. Copyright © 2016. Oxford Publishing Limited. Reproduced with permission of the Licensor through PLSclear]

Organization of cervical mesoderm. Paraxial mesoderm is organized as somites. Intermediate mesoderm begins at level c1. It rounds up as the archicnephric duct. Individual nephrotomes and ductules are neuromeric. Lateral plate mesoderm posterior to cardiac LPM also begins at level c1. Its somatic and visceral layers are separate, creating the coelom. Pleuroperitoneal folds arise from the nephric ridge. Therefore conduction from the neck into the thorax is spatially organized [Courtesy of William E. Bemis]

Conversion of presomite mesoderm. Somitomeres after Sm8 are transient structure and immediately transition to somites. [Reprinted from Gilbert SF, Barressi MJF. Developmental Biology, 11th ed. Sunderland, MA: Sinauer; 2016. Copyright © 2016. Oxford Publishing Limited. Reproduced with permission of the Licensor through PLSclear]
Mammalian somitogenesis begins at Sm8 but, in terms of timing, this transformation occurs upon the development of the 19th somitomere. This is interesting because, in humans, these twelve levels correspond to the first 12 somite levels (4 occipital and 8 cervical). The first somite forms directly caudal to the otic vesicle. As the subsequent somitomere develops caudally (at position Sm20) Sm9 becomes the second somite. Thus, new pairs of somites are added on in rostrocaudal order until the number fixed for the species is reached. In humans, segmentation takes place from day 16 to day 28. It produces a total of 7 somitomeres and 42–44 somites at the rate of 4 per day. Recall that of the 8–10 coccygeal somites only 2–3 persist.
In sum, depending upon what stage the gastrulation process is observed, the expanding column of body mesoderm consists of (1) a rostral zone of mature somites that are differentiating into their subcomponents (e.g., sclerotome and dermomyotome); (2) an intermediate zone of pre-differentiated somites, and (3) a caudal zone of somitomeres just rostral to the retreating primitive streak (Fig. 10.7).

Somitogenesis. Cranial to caudal progression consists of 6 steps. Somite lower right is more mature and therefore are further along in the process. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
The “Clock and Wavefront” Model
The process of metameric transformation is regulated by the “clock and wavefront” model proposed first by Cooke and Zeeman in 1976 and subsequently elaborated by Pourquié in 2003. Presomitic mesoderm cells oscillate between a permissive and a non-permissive state for somite transformation. These changes of state are controlled by an autonomous segmentation clock, present in all vertebrates. It consists of a wavefront of maturation that sweeps caudally along the axis of the embryo, causing physiologic changes wherever it is located. The clock has an intrinsic periodicity (4 hours in humans) and is responsible for generating the segmentation of the somites [1] (Fig. 10.8).

Clock and wavefront. The body axis of vertebrates is composed of a serial repetition of similar anatomical modules that are called segments or metameres. This particular mode of organization is especially conspicuous at the level of the periodic arrangement of vertebrae in the spine. The segmental pattern is established during embryogenesis when the somites—the embryonic segments of vertebrates—are rhythmically produced from the paraxial mesoderm. This process involves the segmentation clock, which is a travelling oscillator that interacts with a maturation wave called the wavefront to produce the periodic series of somites. Segmental patterning of the vertebrate embryonic axis (PDF Download Available). Available from: https://www.researchgate.net/publication/5439063_Segmental_patterning_of_the_vertebrate_embryonic_axis [accessed Dec 15 2017].
The nuts and bolts are as follows. Notch is the putative stimulus for somitomere-somite transformation. It is a transmembrane receptor gene that gets periodically turned on by a number of time-based cycling genes known as the c-hairy1 family. These cycling genes cause coordinated pulses of notch mRNA to appear when a somite is being formed. In somitomeres, factor FGF8 keeps the cells in the mesenchymal state…and therefore prevents transformation into a somite. In any somitomere, the FGF8 gradient decreases toward the rostral axis. Thus, the rostral zone, where somitogenesis is taking place, has very low FGF8. This permits the cells to respond to Notch, to differentiate and acquire an epithelial boundary. In the caudal zone, high FGF8 prevents epithelialization of PSM cells. Thus, the wavefront of somitogenesis corresponds to the boundary between two domains of fgf8 expression: high/non-permissive (in the caudal PSM) and low/permissive (in the rostral PSM).
The body axis is growing and elongating caudally with constant addition of new somitomeres. Thus, as new PSM cells with strong fgf8 expression are added to the rear, the overall FGF8 gradient is displaced caudally as well. This guarantees that the metameric boundaries on both sides of each somite are separated by a cellular distance corresponding to the caudal displacement of the determination wavefront during a single oscillation cycle of the segmentation clock. The velocity of somite production is thus linked to the periodicity of the gene clock which is species-specific: 90 minutes for the chick, 2 hours for the mouse, and 4 hours in humans.
Differentiation of Somites
Somite boundaries are defined by epithelialization. This occurs when a mesenchymal > epithelial transformation takes place along the periphery of the somite. The somite is now physically separated from its more cranial neighbor by an epithelial segmental barrier. Within hours, further differentiation takes place along the dorsal–ventral axis. Ventromedial cells lose their epithelial characteristics. They are no longer bound to one another. They migrate medially to surround the notochord to form sclerotomes. In the meantime, dorsolateral cells retain some of their epithelial behaviors to form dermomyotomes. These immediately break apart. The superficial layer of mesenchyme interacts directly with the epithelium to form the dermatome giving rise to dermis and subdermal smooth muscle. The deep layer remains densely packed to form the myotome, the future source of axial striated muscle. Note that these changes take place due to external signals from the notochord; they are not intrinsic to the cells of the somite.
Sclerotomal cells differentiate in medial-lateral fashion according to their distance from the signaling centers of the notochord. Axial sclerotome compacts around the notochord to form the future vertebral body. Lateral sclerotome appears triangular and will form the remaining vertebral elements, and also displays a densely packed caudal zone and a loosely packed cranial zone; the zones are separated by the fissure of von Ebner. The cranial loose zone never chondrifies. It produces signals that stimulate the neuraxis to send out nerve tissue and neural crest into the space. In contrast, the caudal dense zone of the lateral sclerotome takes the form of a triangle. The base abuts up with the perichordal sclerotome (future vertebral body) to form the pedicle. The dorsolateral side produces the neural arch while the ventrolateral gives rise to the costal process (Fig. 10.9).

Differentiation of somite. (a) Ventromedial cells (VM) form the sclerotome. (b) Ventromedial sclerotome cells (Scl) de-epithelize from the somite and migrate towards the ventral notochord (NC). (c) Sclerotomal cells further subdivide into an axial cluster (Scl-A) surrounding the notochord, and paired lateral clusters (Scl-L) flanking the perichordal axial sclerotome. Dorsolateral somite retains its epithelial pattern to become the dermomyotome (DM). (d) The lateral sclerotome (Scl-L) forms a triangle next to the axial sclerotome. The three sides of the triangle become Anlagen for the pedicle (P), neural arch (N), and the costal process (C), respectively. The dermomyotome also subdivides into the dermatome (D) and the lateral migrating myotome (M).[Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]
The final result is a somite divided into distinct mesenchymal zones. The dermomyotome, responsible for dermis and muscle, requires some clarification. Until very recently, it was believed that somites were responsible for all populations of dermis. In reality, their role is limited to the skin overlying the epaxial muscles. In these locations, dermis is formed by a de-epithelialization of the underlying dermomyotome. Blood vessels to this dermis are also derived from dorsomedial dermomyotome. Hypaxial dermis is arranged in dermatomes in a common neuromeric register with its epaxial counterpart but the source material is somatopleuric lateral plate mesoderm in register with the overlying ectomere (Fig. 10.10).

Somite: component parts. Note internal cells of sclerotome become arthrotome to form joints. Dorsomedial sclerotome also forms dura. [Reprinted from Gilbert SF, Barressi MJF. Developmental Biology, 11th ed. Sunderland, MA: Sinauer; 2016. Copyright © 2016. Oxford Publishing Limited. Reproduced with permission of the Licensor through PLSclear]
The function of the myotome needs clarification. Myotomal cells subjacent to the dermatome behave exactly as expected to produce the epaxial muscles that correspond to their level.
The ventrolateral edge of somites produces hypaxial cells that behave in an altogether different manner, depending upon the neuromeric location of the somite. These populations do not aggregate around the spine. Instead, they migrate outward to cover the entire ventrolateral body wall and the extremities. In the case of somites located at the neuromeric levels of the limbs (c5–t1 and l3–s2) the migration deposits them into the limb bud. Similarly, somites located along the interlimb neuromeres (t2–l2) produce muscles that migrate outward along the body wall as intercostals and abdominal wall muscles.
Resegmentation of Somites
Remak in 1855 is credited with recognizing a shift in the metameric boundaries of somites at the level of the sclerotome such that the vertebral body develops from two adjacent somites whereas the neural arch arises from a single somite. This he termed neugleiderung, or resegmentation. Let’s first outline the process and then detail the mechanism [2] (Fig. 10.11).

Resegmentation of somites to form sclerotomes and changes of sclerotomal primordia to mature vertebral parts. The somitic and primordial origins of vertebral parts and phenotypic parts are colour-matched, and the locations of the somites, resegmented sclerotomes, and vertebrae along the embryonic axis are approximately counter-registered. During resegmentation, the sclerotome is formed from the caudal and rostral halves of two adjacent somites, such that the middle of the resegmented sclerotome lines up with the intersomitic cleft (IC). Both the axial sclerotome (Scl-A) and lateral sclerotome (Scl-L) develop dense and loose zones. The dense zone of the lateral sclerotome (Ld) becomes the neural arch (NA). Thus the neural archand is not resegmented. Pedicle (P), which is attached to the rostral part of the vertebral body (VB) chondrifies from the recombination of the loose zone of axial sclerotome (Al) and part of the dense zone of axial sclerotome (Ad). The rostral layer of the dense zone of the axial sclerotome soon forms the intervertebral boundary zone (IBZ) containing intervertebral boundary mesenchyme (IBM), which ultimately forms the annulus (A) and, together with notochord remnants (NC), the nucleus pulposus (NP) of the intervertebral disc (ID). The loose zone of the lateral sclerotome (Ll) does not form bone but promotes emergence of the nerve roots (NR). Thus, the neural arch is derived from a single somite but the vertebral body receives contributions from two adjacent somites. IV intersomitic vessel. Arrows indicate developmental fates of the sclerotomes. [Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]
After the lateral sclerotome develops its pedicular, arcual, and costal components, the mesenchyme of the axial sclerotome begins to show signs of compartmentalization as well. Its caudal zone receives mesenchyme from the dense zone of more distal somite. In the process, the caudal axial sclerotome becomes dense. The result is the unification of the vertebral body with the outlying pedicle. Note that the most medial cells of the axial sclerotome remain loose throughout development as the nucleus pulposus. The most cranial part of the axial dense zone is aligned with von Ebner’s fissure in the lateral sclerotome. It condenses dramatically to produce the intervertebral boundary zone (IBZ). The IBZ encircles the central core of nucleus pulposus and goes on to make the annulus fibrosus of the intervertebral disc. When fusion of the two pedicles to the body and the neural arches to one another is completed the resultant vertebral ring provides circumferential protection for the spinal cord. Note that the costal component gives rise to the transverse process and, potentially, to rib. Costal formation was secondarily eliminated in vertebrate evolution in the cervical and lumber regions as it gave greater mobility for the neck and trunk, although under certain congenital conditions cervical ribs can occur.
Since the two dense zones of axial and lateral sclerotome are lined up with the IBZ it makes sense that the mature pedicles will be attached to the cranial half of the vertebral body, and not to its caudal half. The position of the spinal nerve and intersegmental artery also makes sense because they pass through the loose cranial zone of lateral sclerotome and therefore must cross above the neural arch.
Examine Fig. 10.10 with care. Note that the sclerotome has two zones with two types of behavior. Both the axial sclerotome (Scl-A) and lateral sclerotome (Scl-L) develop dense and loose zones but the former re-segments, whereas the latter does not. Please bookmark this important point; we shall return to it later when discussing the O’Rahilly Müller model of occipitocervical development. These zones are likely genetic remnants of the original arculalia (Fig. 10.12).

Left Arculalia, embryonic (a) versus adult (b). Arculalia are seen in fishes as 4 segmented blocks of cartilage per segment, the result of an obvious built-in genetic program. Right These are not present in tetrapods because tetrapod centra result from perichordal mesenchyme. Nonethless, the spatial programming that determines the piscine system may be operative to determine zones of the sclerotome. Basodorsal (tan) = neural arch, Basoventral (blue) = intercentrum, Interdorsal (pink) = interneural arch, Intervertebral (green) = pleurocentrum. Source: Kardong 8.10 Scl-A—axial sclerotome, Ad = axial dense, Al = axial loose, Scl-L = lateral sclerotome, Ld = lateral dense zone, Ll = loose zone, IBZ = intervertebral boundary zone. [Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]
During re-segmentation, the sclerotome is formed from the caudal and rostral halves of two adjacent somites, such that the middle of the resegmented sclerotome lines up with the intersomitic cleft (IC). The dense zone of the lateral sclerotome (Ld) becomes the neural arch (NA). Thus, the neural arch is not resegmented. Pedicle (P), which is attached to the rostral part of the vertebral body (VB) chondrifies from the recombination of the loose zone of axial sclerotome (Al) and part of the dense zone of axial sclerotome (Ad). The rostral layer of the dense zone of the axial sclerotome soon forms the intervertebral boundary zone (IBZ) containing intervertebral boundary mesenchyme (IBM), which ultimately forms the annulus (A) and, together with notochord remnants (NC), the nucleus pulposus (NP) of the intervertebral disc (ID). The loose zone of the lateral sclerotome (Ll) does not form bone but promotes emergence of the nerve roots (NR). Thus, the neural arch is derived from a single somite but the vertebral body receives contributions from two adjacent somites. IV intersomitic vessels travel through the loose zone.
Arrows indicate the developmental fates of the sclerotomes. Intervertebral disc arises from rostral-most layer of the axial sclerotome dense zone. Neural arch develops from a single-level dense zone of the lateral sclerotome; it is not resegmented. Centrum results from resegmentation of two adjacent axial sclerotomes.
Sneak Preview: Resegmentation in the Ocicipito-cervical Junction
Prior to resegmentation, the eight cervical somites produce eight cervical vertebrae but the first one, proatlas, also called C0, is split up between the basioccipital bone and the atlas. In the process, its lateral derivatives (pedicle, neural arch) are lost. Thus, the C1 root that should have passed below proatlas now passes above the neural arch of atlas. The C8 nerve root passes below C7 neural arch but above the T1 neural arch because the T1 neural arch develops from the caudal half of the C8 somite. Note that resegmentation persists in the proximal ribs but the sclerotomes become mixed distally (Fig. 10.13).

Results of resegmentation. Proatlas the original first cervical vertebra (S4 + S5) loses the neural arch, donates its ventral body to foramen magnum, and its pedicles to the occipital condyles. The first cervical segmental nerve runs correctly beneath proatlas but upon the incorporation of proatlas into the cranial base, nerve C1 is now positioned atop atlas. All cervical nerve follow this pattern which “normalizes” numerically at neuromeric level t1, the first thoracic vertebra. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Assembly of Vertebra
As we have seen, sclerotomes arise in ventral zone of the somites. Each one quickly divides into a rostral half and a caudal half. This transition is marked by a boundary zone of the extracellular matrix called Von Ebner’s fissure. The cell population of the rostral half is loosely packed, while that of the caudal zone is densely packed. We need to make use of the following notation: rostral loose zone of somite N = SNR and the caudal dense zone of the same somite = SNC. Thus, for the first somite, S1 = S1R + S1C. When a somite subdivides, its outer layer, the epithelial plate remains unaffected. It spans both hemi-somites, as does its derivative, the dermomyotome. The perinotochordal sheath imparts a chondrogenic fate to those mesenchymal cells with which it comes into contact. Control of segmentation is dual. The notochord also controls ventral segmentation. When excised, ventral plate spinal ganglia control dorsal segmentation (Fig. 10.14).

Von Ebner fissure. Resegmentation of the sclerotome and its contribution to vertebrae. Schematic temporal sequence of sclerotome resegmentation (side view). Sclerotome rostral and caudal compartments are separated by the von Ebner fissure. The rostral compartment of one somite/sclerotome (yellow) fuses to the caudal compartment of the consecutive somite/sclerotome (blue) to form one vertebra. Thus, the somites and the vertebrae are out of register by one-half of a segment. The dorsal ectoderm (brown) and dermomyotome that do not resegment (purple) have been removed to visualize the underlying sclerotome. Fate of the rostral and caudal sclerotome compartments projected onto adult human vertebrae. Respective contribution of the somite caudal and rostral compartments is shown. The orientation of the embryonic axes is indicated in black bold in the circle and the corresponding medical terminology is shown in gray italics. (R) Rostral somite sclerotome compartments. (c) Caudal somite sclerotome compartments, (Nt) neural tube, (Nc) notochord, (Sc) spinal cord. [Reprinted from Chalupa’s J, Pourquié O. Patterning and Differentiation of the Vertebrate Spine. In: Pourquié O (ed) The Skeletal System. Cold Spring Harbor Laboratory Press, 2009, pp. 41–116. Cold Spring Harbor Laboratory Press]
Sclerotomes recombine to form vertebrae; this is termed re-segmentation. Each vertebra develops as the combination of the caudal (dense) sclerotome combined with the rostral sclerotome of the next-most-caudal somite. For example, the eighth thoracic vertebra develops from T7C + T8R. Using our numbering system (starting from the first occipital somite, S1), vertebra T8 = S19C + S20R. The fusion of sclerotomal tissue around the notochord creates the centrum of the vertebra. By Carnegie stage 15 human embryos demonstrate condensation around the notochord; neural arches, or processes, now enclose the spinal cord. They extend downward and make contact with the dorsolateral centrum. The centrum, located ventral to the spinal cord, encases the notochord. Neural arches have two sets of projections: dorsolateral laminae and ventrolateral pedicles. Note that fishes have entirely separate ventrally-located hemal arches, the function of which is to protect the aorta from compression during swimming. The hemal processes project upward to join the pedicles (Fig. 10.15).

Component parts of vertebra. Hemal arch found in fish for protection of aorta in mobile tail, in most reptiles, some birds and in mammals with long tails. Hypocentrum (intercentrum) is absorbed into the more proximal vertebra body N-1 where it receive the capitulum of rib from vertebra N. Left: [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.] Right: [Courtesy of Michael Carstens, MD]
To this point, two critical joint structures are missing: vertebra-to-vertebra and vertebra-to-rib. A solution to this problem occurs at the junction between the laminae and pedicles. Superior and inferior articular processes (zygapophyses) will create facet joints. The transverse process will form the costovertebral joint. What about the ribs? These grow out from the ventral junction of pedicle with centrum. They have their own ossification center. The coastal processes have a primary articulation here but they develop a secondary projection that expands backward and upward to make contact with the transverse process. Remember that the proximal ribs have a dual somitic origin (the same as that of its vertebra) whereas the distal rib comes from a single somite. Thus proximal fifth rib is constructed from caudal t4 and cranial t5. The distal fifth rib structure is unitary and is exclusively t5 (Figs. 10.16 and 10.17).

Development of vertebra Note chondrification centers are always paired but later coalesce; they become ossification centers. In early childhood, the centers of each vertebral arch fuse and are joined to the vertebral body by a cartilagsinous neurocentral junction. This junction allows growth to accommodate the enlarging spinal cord. Fusion of the neurocentral junction usually occurs between the third and sixth years. Anterior notching of the vertebrae is sometimes seen in the infant’s or child’s vertebrae and shows the site of somite fusion. Secondary ossification centers develop at the ends of the transverse and spinous processes and around the vertebral end plates at puberty. These fuse by age 25 years. Congenital defects are common in the axial system. Variations in the lumbar spine occur in about one-third of individuals. Spina bifida occulta is common. Hemivertebrae result from a failure of formation or segmentation. Such lesions are frequently associated with genitourinary abnormalities and less frequently with cardiac, anal, and limb defects; tracheoesophageal fistula; and conductive hearing defects if the cervical spine is involved. [Reprinted with permission from From Pansky B. Review of Medical Embryology. New York: Macmillan, 1982. https://discovery.lifemapsc.com/library/review-of-medical-embryology/chapter-66-development-of-the-axial-skeleton. © 1982]

Resegmentation of the rib includes the pedicle and the proximal rib, in which the contributions from cranial and caudal somites halves do not mix. The distal rib demonstrates more mixing. For this reason, sensory blocks for any give rib n must be also be placed into intercostal nerves n−1 and n + 1. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Like a suspension bridge, the vertebrate bodies are slung between two sets of vertical pylons (the extremitites). Ribs are struts essential for support. They relate to the vertebrae either by articulation or by fusion. Embryologically, ribs develop from cartilage within myoseptae, sheets of connective tissue in the coronal plane that partition off body segments. In fishes, there are two sets of ribs associated with each vertebral segment, a dorsal set, and a ventral set. Dorsal ribs form at the intersection of each myoseptum with a horizontal sheet of connective tissue, the horizontal septum, that runs longitudinally down the entire length of the fish body. It intersects with the ventral aspect of the vertebral column. Ventral ribs form at the intersection between the myoseptae and the coelom (visceral cavity). Ventral ribs of the fish body are serially homologous with the hemal arches of the fish tail. Ventral ribs are lost in tetrapods, but will occasionally make a guest appearance as a vertebral anomaly.
Ribs are named based on their type of distal articulation. True ribs articulate with the sternum, false ribs articulate with each other, and floating ribs have no articulation. True ribs have two proximal articulations: they are bicipital. This is best understood from the primitive condition. The tuberculum articulates with neural arch via the diapophysis. The capitulum articulates with the intercentrum via the paraphysis (Figs. 10.18 and 10.19).

Evolution of ribs Fishes have dorsal and ventral ribs with myoseptum dividing epaxial versus hypaxial muscles. In caudal vertebrae, the ventral ribs close together like pincers to form a hemal arch to protect the aorta from torsion while swimming. The hemal arch is analogous with the dorsal arch which protects the spinal cord. Amniote ribs gain stability by the addition of interlocking zygoapophyses in continuity with the pleurocentrum. Note insertions of each rib with tuberculum articulating with pleurocentrum of the same level and with capitulum articulating with intercentrum (which belongs to the preceeding vertebra). Note that false ribs have both insertions into the same level. [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]

Articulations of the rib. The seventh rib is shown. Head of rib articulates with the tubercle of the zygaphophysis of vertebra of its same number (pleurocentrum) and with the vertebral body preceeding it (a genetic remnant of the intercentrum). Re-segmentation process affects ribs. Upper part of fifth rib comes from T4C and lower half comes from T5R. This parasegmentation exists in the proximal have of the rib but the mesenchyme of the distal half is more uniformly from the same level. [Reprinted from Drake R, Vogel AW, Mitchell AWM. Gray’s Anatomy for Students, third edition. Philadelphia, PA: Churchill-Livingstone. 2015. With permission from Elsevier]
Intervertebral Discs: The Legacy of the Notochord
The notochord is a very ancient structure. Derived from the very first cell population to undergo gastrulation, it spans the length of the neuraxis, from Hensen’s node (r0) to the tail. As such, the notochord is a structural “fossil” that defines the anatomic site where gastrulation was initiated. In mammals, this is conveniently seen as the pituitary fossa. During early embryonic assembly, genes expressed by the notochord play a vital role in organizing the CNS into neuromeric compartments. The notochord runs all the way down the axial cranial base from the pituitary fossa, through the basisphenoid, and down the length of the basioccipital to emerge as the occipito-dental ligament (apical ligament of the dens). It subsequently travels through the centra along the entire length of the vertebral column.
The fate of the notochord depends upon whether it is encased within a vertebral body or whether it spans the interspace between the vertebrae. Recall that when the sclerotome has four subcomponents. Ventral sclerotome surrounds the notochord as the perinotochordal sheath. At 6 months of age, notochord cells within vertebral bodies begin to degenerate; by the second decade, they are all gone. Sometimes persistent notochord remnant can degenerate into a tumor known as a chordoma. Between the vertebrae, we have a very different story. Here the notochord and its sheath expand to form the nucleus pulposus, better known in its adult form as the intervertebral disc.
The original metameres of the body are co-extensive with the sclerotomes. Thus, every fissure of Von Ebner and every disc lie opposite the mid-point of a body segment. Recall that every somite has at its core, a transient hollow center, the somitocoele, a remnant of its original somitomeric form. Von Ebner’s fissure transects the somitocoele. When two somites recombine, the more cranial somite donates its caudal dense zone to the newly forming vertebra. Somitocoele cells that are dragged backward with this caudal half remain at the interface between the vertebra and its cranial predecessor. The intervertebral disc develops as an interaction between these somitocoele cells and the perinotochordal sheath.
Running through the center of the disc is notochord. Intervertebral discs first appear between the axis and the third cervical vertebra. Recall that the third cervical vertebra, C3 is the product of the caudal dense zone of S7 and the cranial loose zone of S8. Thus, “spacer” genes responsible for discs are not expressed between somites S1 and S7. What we have is a solid basioccipital bone, an occipito-atlantal joint, an atlantoaxial joint, and (finally) a run-of-the-mill intervertebral joint. Obviously, gene expression necessary for interspace anatomy does not take place cranial to axis-C3. But although there are no discs cranial to C3, don’t think for a moment that the notochord is not represented. After running downward through the center of the basioccipital bone notochord is continued by the occipitodental, or apical, ligament (a remnant of the loose zone of ancient proatlas), thence onward into the dens; and then finally into the body of axis.
Disc formation starts at stage 23 and thereafter. Centra of the vertebrae expand and compress down the notochord until it is eliminated. Abnormal persistence can lead to a chordoma. But between adjacent vertebrae the notochord persists as the central zone of the developing disc. Surrounding chondroblasts create a fibrous shell around the nucleus pulposus (Fig. 10.20).

Cross-section of cervical vertebra with disc. Intervertebral discs develop as a fibrous ring around nucleus pulposus, remnant of the notochord. Cervical disc herniation occurs most commonly in the fifth, sixth, and seventh vertebrae. These represent the “newer” vertebrae acquired in amniote evolution. Degeneration leads to Stress over time concentrates at the border between the embryonic pedicle and body, an anatomic weak spot, leading to degeneration and compression. This leads to a defect in the annulus allowing gelatinous nucleus pulposus to protrude outward. Left: [Reprinted from Wikimedia. Retrieved from: https://commons.wikimedia.org/wiki/File:Cervical_vertebra_english.png. With permission from Creative Commons License 3.0: https://creativecommons.org/licenses/by-sa/3.0/deed.en.] Right: [Reprinted from Wikimedia. Retrieved from: https://commons.wikimedia.org/wiki/File:Lagehernia.png]
Ossification of Vertebrae
Ossification centers might seem, at first glance, yet another example of anatomic minutia…fit only for small print. In reality, they represent centers of genetic activity from which bone formation spreads outwards. As such, we are really looking at the “maturity sequence” of a given bone. As such, we can expect that muscle attachment centers will develop in exactly the same sequence.
Once the sclerotomes surround the neural tube, they begin to express collagen II: conversion into bone follows. Stage 17 marks the onset of chondrification. It proceeds in a fixed sequence. Vertebrae typically ossify from four primary centers: one in each centrum and one in each neural arch. The centra have ossification centers located dorsal to the notochord. These are incorrectly described as being singular. Clear-cut bilateral ossifications can occur. Surpression of one centrum can produce a hemivertebra. During the initial postnatal years each centrum shares with the ipsilateral neural arch a synchondrosis, the neurocentral joint (Fig. 10.21).

Ossification of vertebrae. Left: Vertebra C1 (atlas) lacks a centrum. Vertebra C2 (axis) has two centra, one of which forms most of the dens. The development of “typical” cervical vertebrae is shown, including the primary ossification centers within the hyaline cartilage, The development of a thoracic vertebrae is shown, including the three primary ossification centers (observe the joints present at this stage), followed by secondary ossification centers at puberty. The development of the lumbar vertebrae is shown, including the primary and secondary ossification centers. Note the anular epiphyses separated from the body, and the anular epiphyses in place. The development of the sacrum is not shown. Note that the ossification and fusion of sacral vertebrae may not be complete until age 35. Right the primary and secondary ossification centers of the sternum (with ribs developed from costal elements. Left: [Reprinted from Pansky B. Review of Medical Embryology Macmillan, 1982 Chapter 66 Axial Skeleton, fig. 31 ISBN: 978-0023906206 https://discovery.lifemapsc.com/library/review-of-medical-embryology/chapter-66-development-of-the-axial-skeleton. With permission from LifeMap Sciences, Inc.] [Reprinted from Pansky B. Review of Medical Embryology Macmillan, 1982 Chapter 67 Appendicular Skeleton fig. 32 ISBN: 978-0023906206 https://discovery.lifemapsc.com/library/review-of-medical-embryology/chapter-67-appendicular-skeleton-and-skull-development. With permission from LifeMap Sciences, Inc.]
Ossification of the neural arches starts at the base (the root of the transverse process) and spreads in two directions: backward into the laminae and forward into the pedicles > further forward into the body > lateral into transverse processes > downward into the articular processes. At stage 23, the vertebral column looks like spina bifida: almost all vertebrae are present (32–33) but there are no spinous processes, the “tops” are open. Fusion of the spinous processes takes place much later, during the fourth postnatal month. Vertebral arch centers appear in craniocaudal order (some studies dispute this). Unification of the arches follows a distal–proximal sequence: lumbar > thoracic > cervical. Fusion of the centra with the arches follows a proximal–distal sequence: cervical > thoracic > lumbar.
Certain zones remain cartilaginous for many years. During puberty, six secondary ossification centers make their debut. The upper/lower borders of vertebral bodies form ring-like annular apophyses. From them will project costal articular facets. The apices of processes (spinous and transverse) each have a center.
Five Reasons Why Should We Care About Ossification Centers
Reason 1. Somites have a spatial “fate map” which is determined by the site at which epiblast cells ingress during gastrulation. Cells passing through Hensen’s node will be assigned to the medial somite. Those passing through the rostral streak will be positioned laterally.
Reason 2. Sclerotomes come from medial somites and these subdivide into four functional parts: central sclerotome = ventral neural arch, pedicles, and proximal ribs; ventral sclerotome = perinotochordal sheath; dorsal sclerotome = dorsal neural arch; and lateral sclerotome = distal ribs.
Reason 3. Ossification centers represent genetically distinct developmental fields. Thus, the pattern of vertebral development follows the order in which ossification centers appear. The temporal–spatial sequence is ventral > dorsal and medial > lateral.
Reason 4. Collagen II produced by ossification centers attracts myoblasts. Muscle insertions into the vertebra occur in the same temporal–spatial sequence as the ossification centers. This is particularly well demonstrated by the back muscles.
Pathologies or evolutionary rearrangements can selectively involve one vertebral field versus the rest. Inappropriate fusion of the lower cervical vertebrae (C3–C7) is seen in Klippel-Feil syndrome. A cuneiform vertebra caused by a missing centrum can lead to scoliosis. Differences in segmentation between the centra and the neural processes of somites 4–6 are responsible for the occipitocervical junction, a revolutionary development in tetrapods (see below).
Phylogeny of the Centrum
The four zones of the sclerotome are genetically distinct developmental fields. Each ossification center represents a single developmental field. An apparently singular ossification center may represent paired centers at the boundary zone of neighboring fields. Under certain conditions component parts may fail to fuse or display abnormal fusion patterns. These pathologies reflect the expression of otherwise suppressed developmental fields (Fig. 10.22).

Evolution of vertebrate centrae. (a) Basal tetrapod Ichtyostega had rhachitomatous vertebrae with U-shaped Intercentrum having a small aperture for a persistent but reduced notochord. NOTE Ancestrally, the neural arches simply protected the spinal cord. Once the the vertebral column began to support the animal’s weight, special articulations between the neural arches—the zygapophyses evolved. The first interation was between neural arch and centrum. This was transformed in amniotes to an articulation between the neural arches themselves. Most vertebrae had ribs. Sternum has not been identified at this level. The fusion of holospondylous vertebrae prevents identifying either intercentra or pleurocentra. Neck vertebra can barely be distinguished. (b) Amphibian neck consists of a single atlas with very limited mobility. (c) †Anthrocosaursn experimented with emolomerous centrae in which intercentrum and pleurocentrum were co-dominant. (d) In Reptilomorphs have schizomerous condition: the pleurocentrum is exerting dominance. The reptilomorph OCJ consists of a single occipital condyle that articulates with centrum of the atlas. The ancient condition is seen in rib insertion, in which tuberculum of rib N articulates with the pleurocentrum of vertebra N, while capitulum of rib N articulates with the intercentrum of vertebra N−1. E In synapsids, beginning with therapsids, the condyle divides and shifts dorsally. Mammals lose (1) proatlas entirely, and (2) neural spine of the atlas. Atlas and axis lose their zygapophyses. [Courtesy of William E. Bemis]
The number of centra per vertebral body is variable, ranging from none (aspondyly) to six (polyspondyly), seen in Holocephali (octopuses) and Dipnoi (lungfishes). In its primitive form, the tetrapod vertebra is diplospondylous; it consists of two paired parts, cranial pleurocentra, and caudal intercentra plus a neural arch. Such vertebrae, in which all three arch elements are discrete, are called aspidospondylous. Rhipidistians (lungfishes) had a particular type of rachitomous (Gr. rhachis = spine + tomos = cut) vertebra consisting of a large intercentrum, a small pleurocentrum, and a neural arch. The intercentrum was an incomplete U-shaped ring with its open segment directed dorsally whereas the pleurocentrum was U-shaped, but with its open segment facing ventrally. In the development of the craniovertebral joint, the intercentrum fragments along these lines.
The earliest tetrapods, such as Ichthyostega, had rachitomous vertebrae but invented a new lateral stabilizing element, the zygapophysis. An immediate split then occurred between the anamniote line leading to amphibians and the amniote line leading to anapsid reptilomorphs. In the former, the intercentrum became the main element of the vertebra body and is associated with the neural arch whereas, in our ancestral line, the pleurocentrum became dominant. The early reptilomorph condition explains the anatomy of modern-day rib articulations. With the appearance of definitive amniotes, the vertebrae become holospondylous, that is, characterized by the fusion of all component parts. The intercentrum, being absorbed into the posterior aspect of the preceding vertebral body, bears the insertion of the capitulum [3].
Vertebrae of the mammalian line are holospondylous, they form the fused centra with the pleurocentra dominant and the intercentra absorbed posteriorly. When intercentra are present, they remain as unossified intervertebral cartilages.
What is the significance of the above? (1) Developmental fields in the lateral zones of vertebrae can behave differently from those in the center. (2) Fields do not necessarily have to fuse with one another. Thus, ventral sclerotome, associated with the notochord, may remain distinct from the other parts of the sclerotome. (3) The diplospondylous condition is consistent with two genetic parts of the sclerotome: a cranial loose zone and a caudal dense zone. (4) The existence of rachitomous vertebrae proves that neural arches are assembled in a similar manner to the centra. (5) These facts put together will explain the various vertebral forms during evolution. We will sketch these out briefly in this chapter but will deal with them more completely in our discussion of neck evolution.
Rearrangement of Component Parts of the Centrum = A New Joint
Two fundamental events in the adaptation to land were the invention of the neck which granted mobility for predation and the liberation of the pectoral girdle from the skull. As we shall see later, in our discussion of the cucullaris muscle, the development of a moveable joint between the head and trunk was not a new idea. Placoderms had a pivot joint with a flexor/extensor system, the cucullaris, and levator capitis connecting the skull with rigid body armor. In subsequent fish evolution, these ideas were forgotten. When head trunk separation next appeared, a new innovation took place, a joint between the skull base and internal axial vertebral column. The cucullaris remained attached to the pectoral girdle, now displaced to the level of the fifth and sixth cervical vertebrae. The creation of this new joint required the recombination of somites at the cranial–vertebral junction, we shall now examine this process in detail (Figs. 10.23, 10.24, 10.25, 10.26, 10.27).

Formation of the human craniovertebral junction. Sclerotomal primordia and their vertebral phenotypes are colour-matched. Proatlas The proatlas resegmented sclerotome (PA) comes from the fourth and fifth somites. Derived from the proatlas are: the axial zones (Ad and Al) which become the basion (B) of the basioccipital or clivus (CL) and the apical segment of the dens (AD); the lateral dense zone (Ld) becomes the exoccipital comprising the occipital condyle (OC), and lateral rim and opisthion (OT) of the foramen magnum; the hypochordal bow of proatlas (HBp) forms the ventral clival tubercle (CT). C1 The C1 resegmented sclerotome (C1) comes from the fifth and sixth somites. Derived from the C1 sclerotome: the axial zones form the basal segment of the dens (BD); the lateral zone forms the posterior atlantal arch (C1P); the hypochordal bow (HBp) forms the anterior atlantal arch (C1A). C2 The C2 resegmented sclerotome (C2) comes from the sixth and seventh somites. Derived from the C2 sclerotome: the axial zone forms the C2 vertebral body (AB); the lateral zone forms the neural arch of C2 vertebra. IBZ (internvertebral boundary zone) between the proatlas and C1 sclerotome forms the upper dental synchondrosis (US) and the IBZ between the C1 and C2 sclerotomes forms the lower dental synchondrosis (LS). [Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]

Severance line traces the final separation of the skull from the cervical spine. It runs through the original resegmentation fronts of S4C and S5R. These are: (1) the junction between the basion and apical segment of the dens in the axial proatlas, and (2) the junction between the exoccipital, or future occipital condyle, and the lateral mass of C1 (derived from the lateral portion of the C1 resegmented sclerotome). [Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]

O’Rahilly/Müller model Centra, neural arches, spinal nerves and ganglia, comparative relationships. On left are the somites, prior to resegmentation, beginning with S1. Note that S1 has no dense zone, it is strictly a loose zone. S2R is loose so the combination of S1C and S2R does not convert caudal S1 into a dense zone. S4C + S5R becomes the proatlas. On the right shows the lateral masses all as dense zones from a single somite level (no resegmentation). The centra are resegmented. X, Y, and Z from proatlas, atlas, and axis form the tip of dens, body of dens and centrum of the axis. Tip of dens is S5C + S6R and the latter forms the upper synchondrosis of the dens. Body of dens is S6C + S7R with the latter loose zone persisting as the lower synchondrosis of the dens. Body of the axis is S7C + S8R. Note that the lateral masses are not resegmented. Body of the atlas comes strictly from S2. [Reprinted from Standring S. The Back. In: Gray’s Anatomy, 40th edition. Philadelphia, PA: Churchill-Livingstone; 2008: 763–774. With permission from Elsevier]

Staged development of occipital somites. (a) Stage 12 four somites (S1 = D) with hypoglosal nerves (yellow circles) and potential myoblasts arising from S2 to S4. Note that the occipital somites increase in size. Only S4 has a true neural arch, which is uses to make posterior foramen magnum (anterior formen magnum is basioccipital). Neural crest gangia (green) indicated. (b) Stage 14 S1–S4 now have sclerotomes and are positioned ventrally along the notochord. Loose zones (light green) and dense zones (dark green) are indicated. Nerves travel through loose zones. Occipital neural crest diminished and cervical neural crest has formed ganglia. (c) Perinotochordal sheath has reached tip of notochord (r1). [Reprinted from Standring S. The Back. In: Gray’s Anatomy, 40th edition. Philadelphia, PA: Churchill-Livingstone; 2008: 763–774. With permission from Elsevier]

Construction of the posterior fossa from occipital somites in the avian model. Contributions of individual somites to the occipital bone complex have been mapped. Paraxial mesoderm from each somite flows backward in concentric “swatches” which laminate together like Russian dolls. Recall that protoatlas was composed of S4C + S5R. The fifth somite is split between the foramen magnum and atlas. [Reprinted from Huang R, Quia Z, Patel K, Wilting J, Christ B. Contributions of single somites to the skeleton and muscles of the occipital and cervical regions in avian embryos. Anat Embryol 2000; 202(5):375–383. With permission from Springer Nature]
Frameshift of the Skull Base and First Three Spinal Vertebrae = A New Joint
-
Note on nomenclature: SomiteNumberCAUDAL/ROSTRAL.
-
Example: fourth somite, caudal zone = S4C; seventh somite, rostral zone = S7R.
Axial Sclerotomes
The fusion planes of S1–S3 have grooves denoting the previous existence of embryonic segmental arteries. The fourth somite is a normal, healthy somite with both loose and dense zones; hence the fusion between the S4R and its predecessors is marked by hypoglossal canal for the hypoglossal artery, the last remaining intersegmental vessel from the ancient longitudinal neural arteries. S4C recombines with S5R to form the sclerotome of the original primary cervical vertebra, the proatlas. The cranial region of proatlas (S4C) becomes incorporated as a ligament into the basion at the foramen magnum while S5R becomes the apical segment of the dens. The notochord runs downward through basioccipital, through this zone, and down into body of axis proper. The articulation of apical denta centrum with the basion creates the pivot point of the new craniovertebral junction.
The intervertebral boundary zone of the CVJ between somites 4 and 5 has unique properties. Recall that the IBZ is a tight compaction of cells located in the dense zone of the axial sclerotome in line with von Ebner’s fissure of the lateral sclerotome. Under normal conditions, IBZ forms the intervertebral discs. In proatlas IBZ, a physical disruption of cells, mediated by cleavage genes, separates the bony union between the dens and the basiocciput. The dentooccipital ligament and notochord remain behind in the midline. In this manner, the skull gains its independence from the vertebral column.
Dens is strapped into place by ligaments derived from dense proatlas cells ventral to the notochord, the hypochordal bow. These give rise to the anterior clival tubercle located on the ventral basioccipital. The lateral sclerotome of proatlas has important derivatives. Its dense regions form paired exoccipital bones. These fuse to the anterolateral border of the foramen magnum to form the two occipital condyles. The additional flexion of this system was an instant evolutionary advantage. The loose zones of the lateral sclerotome stimulate the development of the C1 nerve root.
Proatlas sclerotome, as the product of S4C and cranial S5R, is quite properly the first cervical vertebra, but it breaks up. Unfortunately, current nomenclature does not recognize the proatlas. Therefore, the product of S5C and S6R is falsely termed “the first cervical vertebra.” Rather than quibble with anatomic dogma, let’s just refer to proatlas as cervical vertebra 0.
Unlike all remaining sclerotomes caudal to S7, in which IBZ morphs into the disc, the IBZs of somites 6 and 7 disappear. Instead of having discs, the dens now has two synchondroses which ultimately ossify. In this way, the proatlas apex from S5R is cemented to the basal dens from S5C/S6R atlas, and the basal dens is united with the S6C/S7R body of the axis.
The upshot of resegmentation is to produce three midline components: the apical dental segment from caudal proatlas (cervical sclerotome 0), the basal dental segment from the first cervical sclerotome, and the body of axis from the second cervical sclerotome. Ossification of the midline is a lengthy process that takes place caudal to rostra. The lower synchondrosis between the axis body and the basal segment occurs at birth but the apical and basal dental segments are not united until adolescence. The apical ligament is derived from proatlas dens. The transverse atlantal and alar ligaments securing the CVJ arise from the S6 first cervical sclerotome and the basal dental segments—recall that the axial structures originally came from paired sclerotomes.
Lateral Sclerotomes
Lateral dense zone of first cervical sclerotome makes the posterior arch of atlas. Lateral dense zone of the second cervical sclerotome makes the posterior arch of axis. Loose zones promote second and third cervical nerves. Hypochordal bow of first cervical sclerotome forms anterior S4 and the anterior arch of atlas. No further equivalent hypochondral bows were produced.
To visualize this process, review Figs. 10.23, 10.24, 10.25. The first model is that of O’Rahilly and Müller (Fig. 10.23) and is based on direct observations of human embryos. It is schematic but tells the story. The second model, from Pang and Thompson (Figs. 10.24, 10.25) is more three-dimensional and presents the final anatomy in a color-coded lateral view of the occiput.
The osteology of the posterior cranial base is an important topic with little clarity in the literature (Figs. 10.26, 10.27, 10.28). We shall discuss it anon. Suffice it to say that the mammalian posterior fossa is made up of four occipital somites. The original three (alpha) fuse together and the fourth one (beta) is added on. Hypoglossal artery and nerve communicate through the interface between alpha and beta. This corresponds to the rostral (loose) zone of S4. During its early development S4 (unlike S1–S3) has proto-neural arches. Its dense zone produces an axial contribution to the terminal part of the basioccipital. The lateral elements of S4, acting as neural arches form the dorsal rim of the foramen magnum and part of the occipital condyles. Rostral S5 (proatlas) donates the rest of the condyles.

Occipital somites S1–S4 produce only muscles dedicated to the head, either as branchiomeric muscles related to arches 3–5 or as the somatic hypobranchial muscles of the tongue. All muscles attached to the occipital bone are by secondary insertions (blue). Superior nuchal line bears secondary insertions of sternocleidomastod and trapezius, the latter is incorrectly labeled in red. Its fibers descend from C2 to C6 (somites S6–S10) to the scapula and then have return to the axial skeleton. Membranous occipital bone has four interparietal fields (the postparietals and tabulars) indicated in pink. Superior nuchal line bears the secondary insertion of occipitalis, incorrectly labeled in red. Note the inferior rim of foramen magunum and medial part of occipital condyles in mammals represent contributions from protoatlas. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
The cranial vertebral boundary in fishes lies between S3 and S4 (they have no neck). In diapsid amniotes (dinosaurs, birds, and true reptiles) it is between S4 and S5. Mammals incorporate the proatlas and have a complete reorganization of the atlas/axis complex. The mammalian cranial-vertebral boundary is between somites S5 and S6. Hypoglossal nerve and first cervical nerve in mammals are separated by the incorporated proatlas. The original first cervical vertebrae (proatlas) develop from the caudal half of S4 and the cranial half of S5. The joint that we use to flex our neck lies between proatlas (the occipital condyles) and atlas.
Development of the Occipital Somites
Nota bene Work of O’Rahilly and Müller constitutes the best resource for understanding the occipital region. The drawings are superb (Figs. 10.23, 10.26).
Stage 9 (1.5–2.5 mm, 26 days, 1–3 somite pairs)
First appearance of somites. The first somite relates to the posterior otic capsule and corresponds to rhombomere 8.
Stage 10 (2–3.5 mm, 29 days, 4–12 somites) pharyngeal arch 1
First somite is now the same size as somites 2–4. Somitocoeles still present. They disappear as differentiation takes place. Sclerotomes appear at ventromedial aspect of each somite.
Stage 11 (2.5 to 4.5 mm 30 days, 13–30 somites) pharyngeal arch 2
Rostral neuropore closed. Somitocoeles gone. Somite 1 is now smaller relative to the rest. It has no contact with surface ectoderm. Its sclerotomic material is confined to the caudal half. This will ultimately be carried forward as the hypoglossal cord It lies caudal to vagal and accessory nerve neural crest. No evidence of a hypoglossal nerve. Somite 2 in contact with surface ectoderm. Its sclerotome is interposed between the compact zone and the neural tube. Somite 3 has a more “regular” shape resembling somite 4. Its sclerotome closely follows that of the neural tube. Intersegmental arteries from the LNA (future basilar-vertebral system) are seen between S2–S3 and S3–S4. This is because intersegment S1–S2 is supplied by the fourth aortic arch.
Stage 12 (3–5 mm 31 days, 21–30 somites) pharyngeal arch 3
Dermatomyotomes 1–4 are still visible. For the first time, occipital somites are anatomically distinguished from cervical somites by two criteria: (1) primordial of hypoglossal nerve; and (2) the appearance of the cervical neural crest. The hypoglossal nerve presents as cellular strands that emerge from basal lamina of hindbrain and terminate in the myotomes of S2–S4. Cervical neural crest flows into somite 5 and further below. This will produce the posthyoid hypopharyngeal cord (strap muscles) c1–c3. These bind to the c1–c3 zones of clavicle and sweep forward under arches 5, 4, and 3.
None of the occipital somites has a somitocoele any longer. Somite 1 continues to have a dermatome but is being transformed into a loose mesenchyme. Although S2–S4 contain dermatome, myotome, and sclerotome, the ventral 2/3 of dermatome is becoming disorganized. The second intersegmental artery is being transformed into a hypoglossal artery between S3 and S4.
Pharyngeal arches 1–3 present with neural crest cells present. Epipharyngeal disc activity is seen (particularly above PA1). Disc cells have joined up with ganglia of V, VII, and X. PA3 contains neural crest from IX and X. Arch 4 contains neural crest destined for the larynx and tongue.
Stage 13 (4–6 mm 30+ somite pairs) pharyngeal arch 4
Dermomyotomes thinned out but are recognizable. Myotomes present in S1–S4 are separated by neural crest septae; they descend as the hypoglossal cord followed by XII nerve roots. This marks the early formation of the tongue. The first appearance of dense and loose zones is seen in S4 and in S5–S12.
Stage 14 (5–7 mm) pharyngeal arch 5, end of pharyngeal arch period
Rostral sclerotomes of S1–S3 are fused with perinotochord. Hypoglossal nerve root and hypoglossal artery separate them from the S4 sclerotome.
Stage 15–22 incorporation of S4 into the occiput
At stage 15 perinotochord of S4 has a dense zone that extends to the midline by stage 17. During stages 16–17, the occipital somites form a single unit extending forward toward the pituitary. This marks the consolidation of the basioccipital. In stage 18, cartilaginous exoccipital appears. Central segments X, Y, and Z represent S5–S7. In stage 19 the dense tissue below (ventral) to notochord condenses to form the hypochordal bow at the level of S5. This is the precursor of anterior arch of atlas. Basioccipital is mainly S3–S4.
At stage 20 the regular pattern of dense and loose zones seen in the central cervical region changes. The dense zones have two derivatives: (1) the central part surrounds the notochord, and; (2) the peripheral part becomes annulus fibrosus. Note that between X and Y there is no annulus fibrosus while between Y and Z it is present, but reduced in size. Well into the fetal period central column of axis continues to display these component parts. Thus, body of axis = S5 + S6 + S7. Neural arch of axis = S6 + S7 while anterior arch of atlas originates from S5.
In stage 21, occipital condyles come from two sources: central segment 4 and exoccipital. The central column of the axis originates from central segments 5–7. Neural arch of the axis comes from S6 to S7; accordingly, it is extra-large. Note that the position of the most superior ganglion is above (cranial) to neural arch. This is further evidence of the pre-existence of an original first cervical segment, the proatlas, constructed from S4 and S5.
In sum: Segmentation in human embryos is a sudden event at stage 9 marked by the appearance of primary rhombomeres [4], the first aortic arch, the otic disc, and three pairs of somites. These latter developed out of the more primitive somitomeres Sm8–Sm10. Neural crest is already present at this stage. Stage 10 is characterized by the appearance of the first pair of pharyngeal arches and the appearance of occipital sclerotomes. Stages 11–12 present with hypoglossal nerve, and spinal myotomes + neural crest. At stage 14 hypoglossal neural crest and the hypoglossal cord populate PA3 and PA4.
Humans have 37 pairs of somites (up to 44 during development). These are distributed as follows: 4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 8–10 coccygeal. In most of us, the coccygeal somites involute, leaving us with 1–3 remnants, but on occasion, persistence of coccygeal somites presents as a stubby tail. Contributions of occipital somites to the occiput have been mapped out by Huang (Fig. 10.27).
Descriptive Anatomy
Bookmark these fundamental evolutionary facts:
-
All somites from S4 backward express Hox3B; S1–S3 do not.
-
Fishes have three occipital somites; tetrapods have four.
-
The original first spinal vertebra in basal tetrapods is proatlas, S4C + S5R.
-
Fragmentation of proatlas contributes to cranial base and is responsible for the formation of the atlas–axis complex.
As previously stated, the composition of occipital bone has changed through the course of evolution. In its original piscine version, it was constructed from three occipital somites. In its current mammalian version, four occipital somites posterior fossa contain important contributions from proatlas, a bone not present in modern tetrapods. The anatomy of the proatlas is absolutely key to the understanding of the modern occipital bone and OCJ. Proatlas is synthesized from caudal S4 and rostral S5. We will use the following type of shorthand: S4C and S5R.
We will start with some generalities about the chondral occipital bone, emphasizing its three components: basioccipital, exoccipital, and supraoccipital. For a more in-depth description, including the multiple components of the membranous occipital bone, the reader is referred to Chaps. 8 (bones) and 12 (dura) (Fig. 10.28).
Sclerotomal fusion around the notochord is the earliest manifestation of the vertebral body. The standard pattern involves the unification of the upper half of one sclerotome (caudal) with the lower half of its neighbor (cranial). This pattern begins with S4–S5. We have previously emphasized that the four sclerotomes behave very differently: their central elements fuse completely, while their lateral elements maintain a separation.
Conceptually, it makes sense that the first priorities of the developing brain are to protect itself. It does so by means of blood supply and an underlying cartilaginous cranial base. At stage 9 (day 20) the first three somites are visible; the appearance of S4 is the definition of stage 10 (day 22). Unification of S1–S4 results in a single midline basioccipital bone, two lateral exoccipital bones, and the posterior rim of the foramen magnum. The anterior rim originates from elsewhere (the proatlas). Vascularization of the occipital bone from the basilar-vertebral system occurs at the same time. The caudal occipital segment is demarcated by the hypoglossal artery and first cervical artery.
Changes in the nervous system are taking place rapidly at this time. Stage 10 is also characterized by the formation of the neural tube, a process involving the fusion of the neural folds. Neurulation is initiated at the fourth somite; it proceeds forward from the occipitocervical junction. During weeks 5 and 6, the CNS differentiates into its component parts. The fourth ventricle roof thins out, giving rise in the midline to the foramen of Magendie and, laterally, to the foramina of Luschka. The fourth ventricle is now in communication with the subarachnoid space and CSF circulation begins.
Since the primordial function of the skull is the protection of the brain, chondrocranium is constructed first, during stages 8–13. Recall that all cranial base cartilages are neural crests, with exception of the otic capsule and the occipital bone complex. This process is described in Chap. 8.
After creating a supporting floor of cartilage, the brain seeks to protect its sidewalls with membranous bones. The process of membranous ossification occurs quickly because it does not require a cartilaginous intermediate. Membranous bone formation is completed by stages 16–17 (38–40 days). Occipital sclerotomes are in register with the segmental components of hypoglossal nerve. The nerves pass through the lateral loose zone of the sclerotomes as follows: the first root emerges between S1 and S2, the second root emerges between S2 and S3, the third root emerges between S3 and S4, and the last root defines emerges below the occiput.
The membranous and chondral components of the occiput have been erroneously referred to as a single bone, supraoccipital. As discussed previously, the upper membranous zone represents the four interparietal bone fields while the lower chondral zone of true supraoccipital also has four zones. IP and SO are divided by several landmarks. External occipital protuberance is readily palpable; it lies midway between the foramen magnum and the apex of the lambdoidal sutures. From this central point, two curvilinear lines arch outward (they are only visible on the dry skull). Uppermost is the highest nuchal line and, below it, the superior nuchal line. The importance of these lines has to do with the muscle attachments associated with them. Highest nuchal line bears along its lateral margin, the caudal attachment of the epicranial aponeurosis. This marks the most posterior insertion of the second arch superficial investing fascia and the caudal attachment of occipitalis. Superior nuchal line bears the proximal attachment of two muscles connecting the skull with the shoulder girdle. Along its medial half is the trapezius while the lateral half bears sternocleidomastoid. These muscles belong to the lateral motor column and develop later in time in comparison with muscles of the suboccipital triangle. They therefore must assume attachments at the distal margin of the chondral supraoccipital bone. Just below the sternocleidomastoid lies the distal attachment of splenius capitis. Running vertically downward from external occipital protuberance is the median external occipital crest. At the midpoint of this vertical line one encounters another transverse landmark, the inferior nuchal line. Two shallow fossae lie between superior and inferior nuchal lines. These are filled up with the broad distal attachments of semispinalis capitis and obliquus superior. Between inferior nuchal line and foramen magnum lies another set of fossae. These contain medially, rectus capitus posterior minor and more laterally, rectus capitus posterior major.
What we can glean from this anatomy? Three functional groups of muscles, arising from deep to superficial, achieve distal insertion into the supraoccipital/interparietal bone fields at progressively higher (and more superficial) levels. The four suboccipital muscles all have proximal attachments from the atlas or axis. All are innervated by dorsal ramus of C1. Three of the four fan upward and outward, seeking fame and fortune along the “occipital Riviera” below inferior nuchal line in the following sequence: rectus capitus posterior minor > rectus capitus posterior major > obliquus capitis superior. The Lone Ranger of this group is obliquus capitis inferior; its spans from the spinous process of the axis to the transverse process of the atlas. (2) Semispinalis capitis is supplied by descending branch of greater occipital nerve, dorsal C2, and dorsal C3. It is forced to attach above inferior nuchal line. A large muscle, it monopolizes the entire fossa all the way up to superior nuchal line. (3) Splenius capitus belongs to the most superficial quartile of the erector spinae muscle and is innervated by dorsal C2–C3. As such, it is forced to attach to the rough surface of occipital bone just below the sternocleidomastoid and, more laterally, to the mastoid process. The attachment of epicranius lies outside the chondral supraoccipital altogether. As a purely neural crest fascia, the SIF naturally relates to the purely neural crest membranous supraoccipital bone.
Exoccipital bones flank foramen magnum. They likely develop from serial contributions from S2 to S3. These quadrilateral flat bones are known as the jugular tubercles because they make up the posterior margin of the jugular foramen. From the ventral neural arches (S4C) of proatlas, the exoccipital bones acquire occipital condyles. These articulate with the superior articular facets of the transverse processes of the atlas. Anatomic confirmation of this is seen in the alar ligaments that unite the medial tubercle of each occipital condyle to the tip of the dens. The roughened area of the jugular process gives a distal insertion to the rectus capitis lateralis (supplied by ventral C1–C2) that spans upward from transverse process of atlas.
The basilar part of occipital bone shares a growth center with basisphenoid, which ossifies at age 25. The inferior margin of the foramen magnum comes from proatlas; it bears a tubercle that provides the suspension for the fibrous pharyngeal raphe. Just anterior to the occipital condyle is rectus capitis anterior (ventral C1–C2). Further forward and lateral to the tubercle is the proximal attachment of longus capitis (ventral C1–C3).
Mechanism of Development
Posterior fossa is constructed in mammals from four occipital somites plus the contribution from the rostral half of the first cervical somite, S5R. Occipital somites have the same component parts as their truncal counterparts but differ in behavior in several ways. Their topology arrangement is different. Instead of being separated by intervertebral discs, they are stacked together in succession, like Russian dolls to surround the spinal cord, as demonstrated by mapping work of Patel and Huang. Their dermatomes, initially present, involute. Their myotomes are strictly hypaxial, producing hypobranchial muscles dedicated to the tongue. Although they have rostral loose zones and caudal dense zones the axial components fuse into a solid block of bone whereas the lateral zones contain foramina that are limited to loose zones, through which cranial nerves pass to the periphery. Jugular foramen contains two cranial nerves, IX and X, and a spinal nerve, representing roots c1–c6, that is mislabeled as cranial nerve XI.
For these reasons, a standard model of parasegmentation does not apply well to the occipitovertebral region. Gray’s Anatomy states the following:
-
Occipital sclerotomes 3 and 4 are most distinct by stage 14, by which time the first three sclerotomes have fused. Vertebrae are formed from the fifth somite caudally: the first cervical vertebra is formed by the caudal half of occipital somite 4 and the cranial half of cervical somite 1. This shift in somite number accounts for the production of seven cervical vertebrae from eight cervical somites.
Seems plausible, doesn’t it? Certainly, it resolves the age-old student’s dilemma, why are eight cervical nerves but only seven cervical vertebrae? Unfortunately, this neat picture is flawed.
O’Rahilly and Müller to the Rescue
Occipitocervical development is best explained by the embryologic model of O’Rahilly and Müller, which takes into account differences in behavior between the centra and vertebral arches. Their conceptual breakthrough resulted from an analysis of the early development of the occipitocervical region not in terms of sclerotomes alone, but by taking into account differences in genetic expression between the centra and the vertebral arches. These translate into differences in behavior. All somites from S4 caudal express Hox3B; S1–S3 do not.
Models have to be predictive. The parasegmentation model fails to account for several anatomic facts. First, the basioccipital bone has two un-named foramina through which two of the three neurovascular pedicles of hypoglossal nerve make their exit. The third pedicle emerges through a separate hypoglossal canal between basioccipital and exoccipital bones. Second, atlas and axis are obviously different from all other vertebrae. Why is the atlas only a ring? Are the dens and the odontoid process one and the same? (3) What is the significance of the occipito-dental ligament (apical ligament of the dens)? Third, the explanation seems to fit mammals fairly well but how can we use it to explain the anatomy of the CVJ in other life forms, that is, fishes and early tetrapods?
O’Rahilly and Muller separated out occipital and cervical somites into their medial and lateral components. From a dorsal perspective, they created a map of the CVJ in which the centra are placed in the middle and the neural processes are placed laterally (cf Fig. 10.13).
Recall that:
-
ventral sclerotome = perinotochordal sheath, that is, the heart of the centrum;
-
central sclerotome = ventral neural arch, pedicle, and proximal rib;
-
dorsal sclerotome (late developing) = dorsal neural arch;
-
lateral sclerotome = distal rib.
When seen from this new perspective, lateral elements display a normal process of parasegmentation. Central elements, being genetically distinct, are free to behave differently (and do so). For this reason, the axial elements of S1–S3 fuse together. Parasegmentation at the axial zone first begins with S4. When S4 is incorporated into the tetrapod skull, its anterior loose zone combines with S3 which is strictly a loose zone. As a result, a mesenchymal gap demarcates the S3–S4 boundary.
The centra of the rostral 3 cervical somites (S5–S7) were designated by O’Rahilly and Muller as X, Y, and Z. Although equal in size to those of somites 8–12, X–Z display unusual behaviors, most (but not all) are explained by this model. At stage 17, X and Y are fused while an intervertebral disc exists (transiently) between Y and Z. By stage 21, Y and Z are fused. Thus, the dens consist of three centra, all of which belong to the axis. Using our terminology, the vertebral body of axis = c1 + c2 + c3.
In standard parasegmentation theory, the neural arch is composed in just the same way as the body. The cranial part is dense; it comes from the caudal zone of the previous somite. The caudal part is loose; it arises from the cranial zone of the same sclerotome.
Atlas is explained as follows. It has “given up” its body (the pleurocentrum) to the axis. The anterior arch remains unexplained. The posterior arch of atlas is quite narrow. It develops from the dense area of sclerotome 5; there is no contribution from the loose area of sclerotome 6.
Axis develops as follows. Its body consists of cranially projecting dens (X + Y) plus Z, X = pleurocentrum of atlas, Y = intercentrum of axis, and Z, the true centrum of axis = the dense zone of sclerotome 6 + the loose zone of sclerotome 7.
Nota bene these caveats to the FMROR model: (1) Terminology referring to the centra as X, Y, and Z is paleontologically fuzzy. Recall that the “centrum” previously consisted of a rostral intercentrum and a caudal pleurocentrum. It is easy to see how the “body” of the ancient proatlas could be split apart. (2) Soft tissue structures such as ligaments are not a part of the FMROR analysis. Paired alar ligaments connect dens to the medial aspect of occipital condyles. These are 11 mm long, thick, and stout. They serve a “check rein” function to prevent excessive rotation. The apical ligament of the dens starts from the anterior margin of the foramen magnum, its fibers coalesce to attach at the tip of the dens. It is flanked on either side by the alar ligaments. This ligament represents the continuation of the notochord from the skull base into the vertebral bodies. It is a direct product of the ventral sclerotome of the first cervical somite.
Now, for a moment, consider two fundamental differences between fishes and tetrapods. The fish skull is connected to the pectoral girdle…the head and trunk are welded together. Axial stability is provided by the notochord. It runs continuously from the skull all the way down the body. We’ll explore these ideas later on; suffice it to say that the earliest tetrapods, such as Acanthostega and Ichthyostega, demonstrate two revolutionary ideas, both of which are crucial for the creation of a neck: (1) disconnection of the head from the body, leading to greater mobility; and (2) a redesign of the first vertebra into a cervical vertebra permitting, for the first time, head rotation (Figs. 10.29, 10.30).

Mid-level sarcopteryigian became tetrapodomorphs. Tristichopterid fish, the oteolepiform Eustenopteron has dermal bones of pectoral girdle (white) attached to the skull at post-temporal bone. These arch backward from spinal 1 to spinal 6. Mesodermal pectoral girdle. The primitive chondral scapulocoracoid (pink) connects to the pectoral fine (yellow). The crown level tetrapodomorph, the elpistostegalid, Panderichthys, is very close to the earlies tetrapod, Tiktaalik. It has a humerus and the earliest evidence of bone marrow. In Acanthostega the pectoral girdle is separated from the skull, its dermal bones are simplified, and the chondral bones are enlarged. [Reprinted from Benton MJ (ed). Vertebrate Palaeontology, fourth Edition. Oxford, UK: Wiley Blackwell; 2014. With permission from John Wiley & Sons]

Separation of the pectoral girdle Eusthenopteron has opercular series (medium green) attached to cheek series (light green) and to skull roof (dark green). Dorsal view shows, just behind the parietals (in dark green) the post-temporals (medial) and tabulars (lateral). Posterior and internal to the operculars are medial and lateral extrascapulars (medium green). Panderichthys: Pectoral series (purple) remains attached to operular series. Acanthostega: disconnection of pectoral series from posttemporal. Opercular series is gone. [Courtesy of Dennis C. Murphy]
Recent labeling experiments tracking neural crest behavior indicate that the head–trunk interface in mammals does not lie at the occipitocervical boundary, but rather lies at the somite 3/4 level. This reflects the more ancient posterior occipital boundary. In fishes, only the first three somites contribute to the occipital bone. Furthermore, somites 1–3 develop normally. They all have cranial loose zones and caudal dense zones. They all have the expected proportions of sclerotome and dermomyotome compartments. Neural crest cells arising from rhombomere opposite somites 1–3 contribute to the posterior pharyngeal arches. Despite the fact that S1–S3 all possess posterior somitic “barrier” zones, crest cells seem to ignore them and proceed blithely forward. Neural crest cells arising opposite somites 4–5 do not contribute to the head. Instead, they behave in a “truncal” fashion, migrating ventrally, in a selective fashion, only through the “permissible” anterior loose zones.
Motor nerves, when they encounter occipital somites, behave differently. They proceed through loose zones only which correspond to developmental field boundary zones. Hypoglossal behaves in a “truncal” fashion, just as if it were a spinal nerve. Thus, while hypoglossal neurons distinguish between the anterior and posterior zones of S1–S3, neural crest cells do not.
Why should neural crest behavior change so radically at the somite S2–S3 boundary? All somites from the fourth somite onward, express Hox3B. Thus, differences in migratory behavior of anterior occipital and posterior occipital neural crest are not intrinsic to the crest cells but are dictated by the genetic environment of the individual somite.
In sum: the head-trunk interface lies at the somites 3–4 border. This is aligned with the head-trunk boundary of lower vertebrates. In both teleost fishes and amphibians, only the first three occipital somites make up the occipital bone. Evolution of amniotes involves the recruitment of an additional somite to make up the occipital bone but this incorporation was not accompanied by an expansion of the occipital NC domain.
Evidence of Intervertebral Discs in the Cranial Base
Recall those occipital somites have cranial loose zones and caudal dense zones; the boundary between them is Von Ebner’s fissure (AKA…you guessed it…the somitocoele). Thus, when somites recombine to make vertebrae, they “split apart” along this fissure. The intervening space becomes the disc.
At first glance, occipital somites have all the equipment to make discs but fail to do so. Basioccipital bone appears as a monotonous solid block, with tell-tale lateral hypoglossal foramina. The first attempt to make a disc is between Y and Z of the axis. This disc is unstable; its brief lifetime lasts from stage 17 to stage 21. The first permanent intervertebral disc appears between the axis and the third cervical vertebrae. What’s going on in the skull base?
Let’s take the Muller–O’Rahilly model and apply it, first to fishes and later to mammals. FMOR established differences in biological behavior between the central elements and lateral elements of sclerotomes. This idea is crucial: it allows us to reconstruct the anatomy of the basioccipital bone and occipitocervical joint in a way that accounts for all these elements. Recall that the cranial loose zone of the first occipital somite is fused with somitomere 7.
The central elements of S1–S3 do not have genes permissive for segmentation. Thus, the bodies and neural arches of S1 + S2 + S3R are fused. The lateral elements of S1–S3 do possess genes permissive for segmentation. Although the occipital sclerotomes do not have facet joints, they interact laterally with spaces that permit passage of hypoglossal neurovascular pedicles. Thus, genes permissive for segmentation are fully operative in the dorsal and lateral sclerotomes from S1 onward. Consequently, the pedicles of S1, S2, and cranial S3 are separate. They act like facet joints. The resulting spaces provide exit for the first two branches of the hypoglossal…the remaining two branches are truncal.
Phylogeny of the Occipital Bone
Fishes use the first three somites to construct the braincase. Basioccipital = S1 + S2 + S3R. First truncal vertebra = dense S3C + loose S4R Note that the S1–S3 are Hox3B negative, do not have parasegmentation, and are fused. Escape routes for hypoglossal nerve are occult boundary markers between these somites.
Basal tetrapods incorporate the rostral loose zone of the fourth somite into the braincase. This occurs probably at the level of Tiktaalik and is associated with the separation of the pectoral girdle (Fig. 10.31). Note that pectoral separation does not take place Basioccipital = S1 + S2 + S3 + S4R. First truncal vertebra (proatlas) = dense S4C + S5R. No axis is present. Transposition of the fourth somite from a trunk to the braincase resulted in (1) a more capacious posterior fossa; and (2) the incorporation of two spinal nerves, accessory and hypoglossal, into the braincase.

Phylogeny of the pectoral girdle: relation to vision? Note between Panderichthys and Tiktaalik loss of extrascapular series (blue) on the skull side and on the pectoral girdle, loss of post-temporal, supracleithrum and postcleithrum (anoclaiethrum). [Reprinted from Ahlberg PE, Clack J Paleontology: A firm stem from water to land. Nature 2006; 440(7085): 747–749. With permission from Springer Nature]
Advanced tetrapods (both lines, the temnospondyls and the anthracosaurs) incorporate proatlas partially, probably as ventral border of foramen magnum. Atlas does not have a ring. Axis is created but has no dens. The condyle is single and ventral.
Amniote synapsids (therapids) have more extensive incorporation of proatlas elements developing a double condyle from loose S5 (proatlas) lateral to the foramen magnum.
Mammals have fragmentation and remodeling of the primitive axis. Final incorporation of proatlas is occiput is the ligament of dens. The central elements of S1–6 are thus characterized by direct bone-to-bone continuity. Fusions. Dense S1–loose S5 = basioccipital bone. Dense S5 = rim of foramen magnum, medial condyles, and apical ligament of the dens. Loose S6–loose S7 = dens/body of axis. Dense S7–loose S8 = third cervical vertebra:
Proatlas is first vertebral body in early tetrapods during the Devonian period. No axis exists; all subsequent cervical vertebrae are alike. Proatlas was incorporated into the skull in subsequent anthracosaurs, such as Gephyrostegus, by which time the second cervical vertebra is transformed into an identifiable axis (although very different in form from its mammalian descendent). The ancient proatlas has been seen in mammals in two forms: (1) Hox gene manipulation in mice causing a “frameshift” mutation; and (2) congenital abnormalities. Proatlas has a “body” consisting of 6 elements: two anterior intercentra, two posterior pleurocentra, and paired dorsal neural arches. The intercentra develop from caudal S4 while the pleurocentra come from rostral S5. The dorsal neural arches of proatlas developed in the same manner. Projecting from the base of the neural arch were lateral masses containing posteriorly-directed articulatory surfaces; these formed a “standard” joint with the subsequent vertebra.
Proatlast remnants exist in a variety of forms [5]. The original form of proatlas without axis is reproduced in the Truang–Goehmann malformation [6] (Fig. 10.32).

Persistent proatlas. Dens is unfused with the vertebral rings of proatlas (*) and atlas (*). Os dontoideum is normal, presented as triangula nubbin. Lateral lateral view shows two separate bodies (*) [Reprinted with permission from Spittank H, Goehmann U, Hage H, Sacher R. Persistent proatlas with additional segmentation of the cranio-vertebral junction: the Tsuang-Goehmann malformation. Radiol Case 2016; 10(10):15–23. https://www.radiologycases.com/index.php/radiologycases/article/view/2890]
In the anapsid reptilomorphs. a fragment of proatlas neural arch is left behind, positioned dorsal to the atlas. This situation restricted movement. This remnant of proatlas persisted all the way into synapsids and disappear with mammals. The remaining fragment of proatlas was lost in mammals with a resultant gain in mobility for the neck.
Here is the final disposition of proatlas: it breaks up in three ways. (1) The centra split off from the body, leaving behind a ring-like neural arch. (2) Intercentra and pleurocentra separate along the coronal plane. The intercentra become the anterior tubercle of the clivus. The pleurocentra enclose the notochord as the apical ligament of the dens (occipito-dental ligament); they also form the tip of dens. The surviving centrum persists as the hypochordal bow. (3) The ventral and dorsal halves of the neural arch and “pedicles” also separate along the coronal plane. The rostral components (S4) are reassigned forward and ventral while the caudal components (S5) go backward and dorsal. Rostral neural arch forms the U-shaped anterior rim of foramen magnum. The rostral lateral masses fuse with the exoccipital bones to form the true occipital condyles. Alar ligaments connect these structures with the dens. Caudal neural arch forms the posterior arch of atlas (C1) and the alar and crucial ligaments. The dorsal lateral masses become the lateral masses of the atlas.
The secession of the proatlas centra from its body produces the S5 tip of dens. This will fuse with central elements of atlas and with body of axis to produce the odontoid process. The new “slimmed down” version of the proatlas body is known as the hypochordal bow. In general, it disappears along with the rest of proatlas. But at times the hypochordal bow of proatlas will persist. In such cases, it fuses with the anterior arch of the atlas. This produces an abnormal articulation between clivus, anterior arch of atlas, and the odontoid apex (Figs. 10.33, 10.34).

Apical and alar ligaments represent loose zone of proatlas (S5R). Exposure of the cruciform ligament after removal of posterior arches of atlas and axia removed andupward reflection of the tentorial membrane These span from dense zone of proatlas (S5C) to the anterior rim of foramen magnum which may represent S5R contribution or dense zone of fourth occipital somite (S4R). The centra of proatlas split up in the coronal plane. Intercentrum becomes anterior tubercle of clivus. Pleurocentra become the apical ligment of the dens and the dens apex. The neural arch of proatlas separates from the centra and breaks into to U-shaped segments. Rostral neural arch goes ventral and contributes to anterior rim of foramen magnum. Caudal neural arch goes dorsal to form posterior arch of the atlas. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

S1–S3 Mammalian atlas contains element of proatlas. The neural arch of proatlas splits in the coronal plane. Centrum breaks into three segments. Rostral intercentrum becomes anterior tubercle of clivus. Caudal pleurocentrum forms dens and the apical liament. The remaining fragment of centrum is the hypochordal bow. Neural arch of proatlas splits as well. Rostral neural arch shifts ventral to form anterior rim of foramen magnum (not seen here). Caudal neural arch shifts dorsal to form posterior arch of atlas. Left: [Reprinted from Keith A. Human Anatomy and Morphology, fourth ed. New York: Longmans, Green & Co; London: Edward Arnold, 1921.] Right: [Reprinted from Drake R, Vogel AW, Mitchell AWM. Gray’s Anatomy for Students, third edition. Philadelphia, PA: Churchill-Livingstone. 2015. With permission from Elsevier]
Atlas is, in reality, the second vertebral bone of the neck. Designed to permit flexion/extension of the head, atlas is a very “odd duck.” Embryologically complex, atlas is synthesized in a regular manner from caudal S5 and rostral S6 but also with contributions from proatlas. The atlas has the forma of an ovoid ring. It consists of two lateral masses connected by anterior and posterior arches. The lateral masses are bulkier anteriorly; they bulge into what would otherwise be a smoothly oval vertebral canal. This makes the anterior arch appear smaller. The inner “shoulders” of the lateral masses are connected by a transverse ligament. This ligament divides the vertebral canal into two compartments. The anterior third is occupied by the dens while the posterior two-thirds contain the spinal cord and its coverings. Both dens and cord are of similar diameter (Fig. 10.35).

Ossification of atlas and axis. By the same logic Axis has 8 centers with tips of dens and body coming from two sources each. The presence of these centers in the axis [Reprinted from Standring S. In: Gray’s Anatomy, 40th edition. Philadelphia, PA: Churchill-Livingstone; 2008. With permission from Elsevier]
Atlas lacks a true body. This odd situation seems more rational if we consider that its genetic components behave similarly to those of somites 1–5. That is, central elements maintain allegiance to one another and remain fused, while the lateral elements remain separate. The centra of the atlas behave in exactly the same manner as those of the proatlas: they also secede from the body of C1. Thus S5C–S6R body of the atlas fuses above, with S5R body of proatlas, and below, with the S6C–S7R body of the axis. The lateral masses of the atlas are proatlas remnants; these come from S5R neural arch. They bear two sets of articulations directed above, toward the occipital condyles, and below, toward the axis. The transverse processes of the atlas are long, almost equal to those of C7. They serve as leverage points for muscles controlling head position. The neural arch of the atlas is derived in a very standard way from S5C and S6R. In conclusion, we can see that both proatlas and atlas share very similar anatomy.
Ossification centers of the atlas are four. Those of the lateral masses appear at the seventh week. It spreads outward and backward along the neural arches; these unite at 3–4 years. Ossification of the anterior arch follows a slower course with paired midline centers (mistaken for a single center) appearing at about a year. At times ossification spreads forward from the lateral masses, another manifestation of the bilateral nature of the anterior arch.
Muscle attachments follow from key structural landmarks of the atlas. As expected, they follow the ossification sequence. The anterior tubercle on the anterior arch gives proximal attachment for longus colli. Anterior surface of lateral mass is the primary attachment for the upward-directed rectus capitis anterior. Flanking posterior tubercle are rectus capitis posterior minor. Transverse process bears rectus capitis lateralis superiorly. Behind that is obliquus capitis superior. The very apex of tranverse process bears a muscle going up, obliquus caitis inferior, and three muscles heading downward: levator scapulae, splenius cervicis, and scalenus medius.
Caveat
Recall that in mammalian evolution, the pleurocentra become the dominant component of the vertebral body. Intercentra are relegated to a low-status knob (parapophysis) wcapitulum) of the rib. We have to be careful about our terminology here. Previously, we used the anatomy of primitive tetrapod vertebrae to make a point. These creatures had bodies in which intercentra and pleurocentra were co-equal in size. We took advantage of this to point out the relationship to parasegmentation, in which pleurocentra were derived from the loose zone of the somite in question whereas intercentra originated from the dense zone of the next more cranial somite. In modern mammals, although the body is just pleurocentra that do not mean it arises from a single somite…the separate genetic centers behind parasegmentation are alive and well in twenty-first-century mammals:
Axis is designed to permit rotation of the head. It develops exclusively from sclerotomal elements of caudal S6 and cranial S7 but includes additions from proatlas and atlas. The body of the axis is the sum of the central sclerotomes (from which project the ventral neural arches) and the ventral sclerotomes (which enclose the notochord). Sitting atop the body is the odontoid process (dens), the formula for which is S4C + S5R + S5C. These are completely fused. At birth, a cartilaginous process representing an abortive attempt to create an intervertebral disc is seen between the odontoid process and axis body, that is, between S5C and S6R. This synchondrosis is present in most children up to age for but disappears by age 8 (Fig. 10.36).

Development of the axis. The axis is a composite structure: S5C (odontoid process of dens) + S6R (dens) + S6C (synchondrosis) + S7R. The three developmental phases of the axis (C2) and the three waves of ossification. The primordia for the dens components are assembled during the membranous phase. Upper and lower dental synchondroses are shown as dense lines. First wave of ossification at fourth foetal month consists of bilateral centres for the neural arches and a single centre for the centrum. Second wave at sixth foetal month consists of bilateral ossification centres for the basal dental segment. At birth, the basal dental centres should have integrated in the midline and begun to be fused to the centrum. Third wave of C2 ossification occurs from 3 to 5 years post-natal life at the apical dental segment, which does not become fused to the basal dens till the 6–ninth year, and fully formed during adolescence. [Reprinted from Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv System 2011; 27:523–564. With permission from Springer Nature]
The axis has primary and secondary ossification centers. Like any cervical vertebra, each vertebra arch has two centers; these appear in the seventh week. In the fourth–fifth month, ossification of the central is observed. This starts at the midline from paired centers (erroneously described as being single). The odontoid components from the axis show up at six months as paired centers. These generally fuse before birth. The odontoid components from proatlas demonstrate ossification between 5 and 8 years with fusion to the remainder of the dens by 12 years. Abnormalities of ossification can involve the apex with dens. If the odontoid process fails to fuse with the body the result is os odontoideum.
Anterior body of axis provides attachment for the longus colli. The tips of the transverse process are quite busy, containing proximal attachments for ventral scalenus medius and dorsal splenius cervicis and levator scapulae. Spanning between the tips are intertransversus muscles. The projection of the spinous process gives attachment first, to obliquus capitis inferior and further posteriorly, to rectus posterior major.
-
CVJ Derivatives: A Summary (Fig. 10.37)

Assembly of the atlas and axis Anapsid reptilomorph Dimetrodon (just before split into diapsid and synapsid lines) had two cervical vertebrae but no atlas and no axis. Centrum 1, intercentrum 1, and neural arch are separate and proatlas persists. Mammals shows fusion between intercentrum 1 and neural arch 1. Centrum 1 intercentrum 1 separate from atlas and fuse. They join with centrum 2 the body of the axis to become the tripartite dens. [Courtesy of William E. Bemis]
Somites 1–3 (alpha unit)
-
basioccipital bone,
Somites 2–3
-
exoccipital, including jugular tubercles.
Somite 4 (beta unit)
-
Centrum (S4C) is incomplete = posterior basioccipital, anterior tubercle of clivus.
Somite 5
-
Intercentrum (S5R) = apical, transverse ligaments of dens;
-
Pleurocentrum (S5C) = dens (apex), apical ligament.
-
Hypocentrum (what remains) = hypochordal bow.
-
Neural arch rostral (S5R) shifts ventral = anterior U-shape to foramen magnum, occipital condyles, alar, and cruciate ligaments.
-
Neural arch caudal (S5C) shifts dorsal = posterior arch of atlas (rostral), lateral atlantal masses.
Somite 6
-
Hypocentrum = anterior arch of the axis.
-
Centrum = dens (body).
-
Neural arch = posterior atlas arch (caudal).
Somite 7
-
Hypocentrum = disappears.
-
Centrum = (S7R) = body of axis.
-
Neural arch = posterior arch of the axis (rostral).
Cervical Vertebrae C3–C7: garden variety
We don’t have much to report here. It is as if, from c4 downward, nature simply repeats herself. The benefit for mammals is an increase in mobility. It’s not fair to compare mammals with birds because avian cervical vertebrae are amphicoelous, that is, they are double, opposing saddle joints… not unlike the human thumb. In humans, the transverse process is well out of the way of the foramen intertransversarium…let’s give preference to the vertebral arteries. These vertebrae do possess a coastal process but, under normal circumstances, it aborts. The distal parts of the cervical processes do not develop. But we are all aware of the so-called “cervical rib.” This is especially common at the level of c8 (the seventh cervical rib). Anomalies of lower cervical ribs occur via inappropriate cervical fusion: Klippel-Feil syndrome. The phenotype includes a low hairline, foreshortening of the neck, and reduced movement (Fig. 10.38).

Klippel-Feil Syndrome is defined by congenital fusion of two cervical vertebrae (at any level) with consequent limitation of motion. A short neck and low hairline are associated finidngs along with associated scoliosis and spna bifida. Left: [Reprinted from Ulmer J. Klippel-Feil Syndrome: Ct and MR of acquired and congenital abnormalities of the cervical spine and cord. J Comp Assist Tomo 1993; 17(2): 215–24. With permission from Wolters Kluwer Health, Inc.] Right: [Reprinted from McGaughran JM, Kuna P, Das V. Audiological abnormalties in Klippel-Feil Syndrome. Arch Dis Childhood 1998; 79(4): 352–355. With permission from BMJ Publishing Ltd.]
Cervical vertebra C3–C7 possess proximal muscle attachment sites that are analogous to those found in thoracic and lumbar vertebrae. The reader will note that these sites, whether dorsal or ventral, follow a similar pattern: midline-to-lateral, and from deep-to-superficial. The dorsal pattern exactly mimics the spread of ossification from the base of the neural arch (1) outward laterally along the lamina to the transverse process; and (2) outward dorsally along the spinous process.
Distal muscle insertions follow the same spatial pattern. Dorsal musculature is arranged in three layers. These mostly-forgotten muscles are important not for their specifics but for the biological pattern of their attachments. They are described in detail in the myology section.
Intrinsic layer muscles are monosomitic (with exception of; from deep to superficial and from medial to lateral: multifidus, semispinalis, interspinale, and intertransversari.
Erector spinae are polysomitic muscles arranged in three columns, from medial to lateral: spinalis cervicis, longissimus cervicis, and iliocostalis cervicis.
The splenii connect the occiput and the OCJ with the trunk: splenius capitus and splenius cervicis.
C7 does present a few peculiarities. Because of its long spinous process, this bone is often termed the vertebra prominens. Foramina transversaria transmits vertebral veins, but not the vertebral arteries. It has two very active insertion points. Anterior tubercle of the transverse process receives scalenus medius and levator costarum. Posteriorly, the tubercle of the spinous process receives the insertions of interspinales, multifidus, semispinalis thoracis, spinalis capitus, and, finally, trapezius.
Phylogeny of the Cervical Vertebrae
We are now going to use what we know from the fossil record to reconstruct the evolution of the cervical spine with emphasis on three specific issues.
-
What adaptations took place in tetrapod vertebrae for life on land?
-
How did the cervical spine expand in length?
-
What specific innovations took place in mammals to enhance control of the upper extremity?
Once upon a time, there was no such thing as a neck…just ask any fish! The terrestrial triumph of tetrapods involved multiple evolutionary advances including gills into lungs, fins into limbs, a novel joint interposed between the head and body, and a flexible neck designed for positioning of the head. As we shall see, the cutaneous envelope of neck and its musculoskeletal infrastructure developed as modifications of the piscine trunk, specifically involving somites 5–12. How the neck came into being will be discussed at the end of this chapter. For our purposes here, we are going to focus on the following topics: (1) the number of cervical vertebrae, (2) the parasegmentation of the occipitocervical junction, a structure that literally revolutionized life on land, and (3) the broader topic of vertebral evolution to the mammalian line (Fig. 10.39).

Phylogeny of vertebrae Neural and hemal arches do not become important until placoderms. Vertebral centra appear late in evolution of both chondrichthyan and osteichthyan fishes. In tetrapods, the centra replace the notochord except for nucleus pulposus. [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]
The Vertebral Axis in Fishes: Adaption for Swimming
Life in water in the water has its advantages. The axial skeleton of fishes faces few demands. Due to buoyancy, gravity has little influence. Body design must resist “telescoping,” contraction while swimming due to muscle contractions. The bauplan of the fish body is seen in the cephalochordate Branchiostoma, better known as amphioxus (Gr. amphi = both + oxy = sharp), so named for its two pointed ends. Amphioxus is supported by a notochord that runs the length of its body and can be stiffened by contraction. Its body wall is constructed from somites and it uses a series of V-shaped myomeres for swimming (Fig. 10.40).

Amphioxus Note the eight cartilaginous branchial baskets for filter feeding. The nerve cord has partial covering with a neural arch or spine. The perioral head has a neural crest annular cartilage. [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]
Although axial systems of extinct jawless fishes are as yet an evolutionary black box. Perhaps their body armor precluded the development of a spine. A few examples, such as osteostracans, have random skeletal elements surrounding the notochord and spinal cords. In living jawless descendents, hagfishes have no vertebral elements but lamprey has cartilaginous neural arches, arculalia, which partially protect the spinal cord. Jawless fishes do not have ribs.
A true axial skeleton evolved in gnathostomes. Chondrichthyans protect the spinal cord with a continuous jacket of segmented cartilage plates and poorly developed ribs. The vertebral column of bony fishes is divided into a trunk region and a tail region with modifications in the latter for swimming. Although the notochord continues straight into the skull the first trunk vertebra has articulation but essentially no mobility, due to the stiff notochord. The vertebrae in basal choanate fishes, such as the osteolepiform Eusthenopteron were rachitomatous with paired elements fused in the midline (referred to hereafter in the singular): small dorsal pleurocentra, dominant U-shaped intercentra, and neural arches. In the tail, mirror-image hemal arches remained to protect the aorta during swimming. Due to the low-stress conditions of water, fish vertebrae are connected with ligaments; there is no need for interlocking bony joints or zygapophyes (Figs. 10.41, 10.42).

Shark spine [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]

Osteolepiform Eusthenopteron vertebrae. The basal condition is intercentrum dominance. [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]
The Vertebral Axis in Tetrapods: Adaption for Weight-Bearing
Aquatic life placed little demand on the axial skeleton. Not unlike a weightless astronaut, you spend your time swimming around with a buoyance system. But the transition to land meant suspension of the body between limbs. Axial skeletal adaptions to the new mechanical demands resulted in vertebra with fused elements.
Basal tetrapods, the labyrinthodonts evolved directly from rhipidistians. They were ponderous creatures. We do not know if they had a sternum but most of their vertebrae were rib-bearing. Abdominal ribs were useful to present the collapse of their lungs and viscera as their massive bodies lay on the ground. It appears that they had primitive cervical vertebrae, the first of which was the atlas but not axis. The number of cervical vertebrae is uncertain (Fig. 10.43).

Cervical vertebrae in Ichthyostega. Ribs present at all levels. Number of cervical vertebrae is uncertain…based on cervical ribs the original tetrapod neck had 3 cervical vertebrae. Frogs and salamanders have a single cervical vertebra, the atlas. The frog brachial plexus consists of sp2 and sp3 with sp3 belonging to the first truncal vertebra. Recall that sp1 belongs to the proatlas. [Reprinted from Ahlberg PE, Clack JA, Blom H. The axial skeleton of the Devonian tetrapod †Ichthyostega. Nature; 2005; 437:137–141. With permission from Springer Nature]
The vertebrae of Acanthostega and Ichthyostega, were apsidospondylous and intercentrum dominant. This loose conglomeration of bony elements was ill-suited for life on land. They remained notochord dependent. Its elastic sheath provided form mobility which allowed them to incorporate the primitive neural connections used in side-to-side swimming motion of fishes into undulations used to propel one foot in front of another. Such lateral body swings are preserved today in salamanders. But their ponderous side-to-side pattern of locomotion placed undue torque on the spine; it åthis with two innovations. First, their cervical ribs were drastically reduced and eventually incorporated into the vertebra as zygapophyses. The processes reach across intervertebral joints as gliding articulations. They permit bending of the spine but resist twisting. The second innovation was the sacral region. By providing an attachment of the pelvic girdle to the vertebral column hindlimb, propulsive forces could be directly transferred to the spine.
Labyrinthodonts gave rise to two different lineages characterized by differences in the relative prominence of the centrum elements. Both directions reflect a reduced role for the notochord, with bony elements strengthened for load-bearing. Temnospondyls retained dominant intercentra. They are most closely associated with ribs and axial muscles used in swimming. Pleurocentra were not eliminated; they were eventually incorporated into the intercentrum as a single mass, the holospondylous condition, seen today in modern amphibians. The evolutionary consequence of holospondyly is possibly reflected in the foreshortened amphibian neck. Only a single cervical vertebra, the atlas, is present. Elsewhere in evolution, reshuffling the developmental subunits of the vertebra is responsible for the creation of a proatlas–atlas–axis complex. This is possible when these elements are fused (Figs. 10.44, 10.45, 10.46).

Vertebral body evolution from Rhipidistians forward can be classified by composition of vertebral centrum. Temnospondyls are intercentrum dominant and lead to all living land-dwelling anamniotes. Anthracosaurs are pleurocentrum dominante and lead to amniotes. [Reprinted from Kardong KV (ed). Vertebrates: comparative anatomy, function, evolution. McGraw-Hill Education; 2015. With permission from McGraw-Hill.]

Proatlas partial incorporation Atlas–axis complex appears in the bifurcation of basal tetrapods into temnosondyls and anthracosaura. Note proatlas fragment dorsal to neural arch of atlas in two temnospondyls Greererpeton burkemorani (left) and Eryops. Note the reduction of pleurocentra in the latter. Proatlas (C0) incorporation must be a concomitant event with the differentiation of C1 atlas and C2 axis. [Reprinted from Shishkin MA. Evolution of the cervical vertebrae in temnospondyl amphibians and differentiation of the early Tetrapods. Paleontological Record 2000; 34(5):534–546. With permission from Springer Nature]

Spine salamander. Note single cervical vertebra, pectoral girdle associated with second spinal vertebra. Note in amphibians, nerve sp1 is autonomous. The salamader brachial plexus is sp2–sp4. The frog brachial plexus is reduced to sp2 (flexors) and sp3 (extensors). This reflects the drastic reduction in its upper extremity. [Courtesy of William E. Bemis]
The other lineage of Labyrinthodonts was the Anthracosaurs (so-called because their skeletal remains were found in coal beds). The vertebrae of these creatures were characterized by expansion of the pleurocentrum at the expense of the intercentrum. In one direction, embolomeres, the two elements remained unfused but equal in size. In the other direction, intercentrum was reduced; this gave rise to amniotes.
Crown anamniotes are represented by the reptilomorph anthracosaur lineage. These creatures were embolomeres, meaning that their intercentra and pleurocentra were equally represented. The anthracosaurs ultimately gave rise to amniotes. Loss of cervical and abdominal ribs led to greater mobility.
In amniotes pleurocentra enlarged at the expense of the intercentra, ultimately eliminating them altogether. Pleurocentra function to support the neural arches. They undergo successive interlocking by virtue of their zygapophyses. This reduces axial flexibility but strengthens the vertebral column for load-bearing. The limbs became less ponderous leading to more speed and agility.
Tetrapod Vertebral Column: Regional Variation and Size
The tetrapod axial skeleton has more regional variation than in fishes due to multiple functions. For example, the stresses and mobility required for the upper extremities are different from the lower extremities. Living anamniotes went in a different direction. As amphibians reinvaded a watery habitat, swimming became important for the pre-adult phase. Subsequently, specializations for leaping were required. These include reduced numbers of vertebrae (they have but 1 cervical vertebra), enlarged centra, and extensive ossification of the vertebrae with the fusion of components: the holospondylous condition. These small creatures have a sternum but ribs for body support are not important. The sum of these changes strengthens the central core in preparation for propulsion by leaping.
Basal amniotes, the reptilomorphs, developed many innovations. They had anapsid skulls, without a temporal fenestration for specialized chewing muscles. The number of cervical vertebrae in anaspids is 5, including the invention of an axis, thereby allowing for enhanced neck mobility. Neck length of reptilomorphs immediately expands in subsequent lines. Diapsids increase their cervical vertebrae by expropriation from the trunk. Modern reptiles have 6–10 while birds have 11–25. This shifts the cervical–thoracic junction backward while maintaining a brachial plexus of 4 roots (Fig. 10.47).

Expansion of cervical vertebrae during amniote evolution through two crown amniote clades. Sauropsida are almost all diapsids. The extant (living) reptiles are Lepidosaurs and Archosaurs—the line leading to birds. Synapsids started out at the level of pelycosaurs with 5 vertebrae which increased to 6 with the therapids Dimetrodon† and stabilized at 7 with the cynodonts (Thrinaxodon† and mammals). [Reprinted from Hirasawa T, Kuratani S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
Reptilomorph needed a stronger skeleton to assist their invasion of the terrestrial environment so they reduced the intercentrum to a small piece and pushed in ventrally between the definitive centra (recall that all these pieces are fusions of bilateral embryonic components). Ribs were embedded in the body wall as important sources of lightweight support, extending all the way from atlas to the caudal (post-pelvic) vertebrae. In the anterior trunk, they developed a sternum, possibly as a fusion of anterior ventral cartilaginous rib elements, paraphernalia. Interclavicle appears for the first time and unites, together with coracoid, to the sternum, thereby locking pectoral girdle into a ventral position. Intercentrum, used by reptilomorphs as the attachment for capitulum of the rib, is incorporated into the pleurocentrum of the previous vertebral body so rib attachments now span two vertebrae. Accordingly, the rib from first vertebra of the trunk attaches to the upper sternum.
Throughout our discussion of diapsids, we have ignored the avian line because it does not lead to mammals. Some comments regarding avian vertebrae are appropriate for understanding a different form of neck. Despite the usefulness of wings, birds cannot use them for food acquisition. They compensate for this using an elaborate system for head positioning and control. Birds are sauropods, direct descendants of dinosaurs; accordingly, they have both an atlas–axis complex and cervical ribs. But the mobility they require for head positioning is achieved by increased neck length.
In synapsids, cervical vertebrae expand, becoming six in therapsids and seven in cynodonts. They accomplish this by duplicating levels C4–C5 using the same hox code.
and pushing additional codes backward. The resulting 6-root mammalian brachial plexus supplies greater complexity of upper extremity muscles (Table 10.1).
Phylogeny of the Occipital-cervical/Craniovertebral Articulation
We now turn our attention to the evolution of the vertebral column in general to that of the craniovertebral junction. First, we review the changing definition of the head-trunk interface over time. We then discuss how the anatomy of the joint evolved.
Basioccipital bone in primitive fishes develops from the fusion of three occipital vertebrae; its termination is the loose zone of S3. The first truncal vertebra is dense S3 + loose S4. The original head–trunk interface for all vertebrates is located at S3–S4. Fishes have direct contact between the head and pectoral girdle using a series of opercular bones. As they are swimming in a watery, low-impact environment, weight bearing is not an issue. “Shock absorber” discs are not needed.
The fish-tetrapod transition is characterized by three well-known organisms: Tiktaalik, Acanthostega, and Ichthyostega. Tiktaalik is the oldest and most primitive tetrapod. It demonstrates the loss of the opercular bone, the subopercular bone, and the extrascapular series (the two remaining opercular bones are lost later in evolution). The pectoral girdle is liberated from the skull and a neck joint exists. It had synovial joints in the forelimb. The hindlimb was incomplete. Although it had a pubis and ilium with an acetabular joint, ilium was not attached to the spine; no ischium was present. Acanthostega demonstrates the five basic elements of the pectoral girdle: anocleithrum, cleithrum, scapulocoracoid, clavicle, and interclavicle. Ischium and the sacrum appear for the first time.
When the upper limb was freed from the skull, the basic cervical plan was one of five vertebrae but fossils do not show us intact original proatlas. Fragments of it do persist up to and including the pre-amniote pelycosaurs. But today we do know what it looks like because of unusual congenital conditions in which this bone reappears (see Fig. 10.32). Genetic manipulation of the homeotic code can also cause it to be produced. The proatlas seen in fossils today is probably the neural arch of the original first cervical vertebra (call it C0) which was produced by neuromeres r11 and c1 in the first tetrapods (without an axis) and subsequently incorporated into the skull. Fossil evidence for an atlas–axis complex dates back to the anthracosaur line but where the incorporation of proatlas occurred is unclear.
At the dawn of our story, we shall start with basal anapsid amniotes. The reptilomorph neck seen in Paleothyris consists of five vertebrae. The craniovertebral joint consists of a single occipital condyle lying ventral to foramen magnum which articulates with a depression in the atlantal centrum. Just dorsal to this lies a fragment of proatlas that limits motion. This is followed by axis and by three standard cervical vertebrae. This allows for the existence of the primitive brachial plexus consisting of four roots, C3–C5 + T1. This plexus, exemplified in chameleons, has just two terminal branches: radial nerve and brachial nerve (Figs. 10.48, 10.49).

Evolution of proximal cervical vertebrae. Cynodonts are the last step before mammals. The transofrmation to mammalian atlas–axis complex, is shown in fig. 36 [Reprinted from Kardong K. Vertebrates: Comparative Anatomy, Function, Evolution, seventh ed. New York, NY: McGraw-Hill; 2015. With permission from McGraw-Hill Education]

Basic amniote pattern of the brachial plexus. br1–br4 (first through fourth spinal nerves) constitute the basic formula for the brachial plexus; brl, N. brachialis longus; rad, N. radialis; spc, N supracorcoideus. Note the amniote has the basic number of 5 cervical vertebrae. Radial and branchial nerves have two roots each and are connected. [Reprinted from Hirasawa T, Kuratani S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
The craniovertebral joint became more sophisticated in the synapsid transition. Our therapsid ancestor, Dimetrodon, showed a split of the occipital condyle, probably back into its two original embryonic parts. These are now relocated dorsally and are situated on either side of the foramen magnum. Concomitant with this was the formation of bilateral facets on the neural arch of atlas to receive the double condyles. Therapsids added a sixth cervical vertebra. All three parts of atlas (neural arch, intercentrum, and pleurocentrum) remained separate. The atlas–axis complex of cynodonts, such as Thrinaxodon still retained proatlas. The axis remains a full complement of seven cervical vertebrae that were now present. The penultimate iteration of the cervical spine.
In mammals, the proatlas is lost. The atlas undergoes a partitioning in which intercentrum joins with neural arch to make a ring while pleurocentrum is displaced backward to fuse with intercentrum of axis to form the dens which in turn fuses with pleurocentrum of axis, thus joining dens with the body. To facilitate rotation, the zygapophyses between the neural arches of atlas and axis are eliminated.
Osteology: Appendicular Bones
Organization of Lateral Plate Mesoderm (LPM)
Without exception, all bones of the head and neck are formed from either neural crest or paraxial mesoderm. Furthermore, all remaining bones of the axial skeleton, the vertebral column, and ribs, are PAM as well. Lateral plate mesoderm is the source of bone for the appendicular skeleton. The pectoral girdle, for evolutionary reasons we shall later explore, is an exception to this rule. The clavicle is a neural crest bone with a core of LPM marrow. Scapula is largely LPM, but specific sites along its periphery and the scapular spine bear neural crest cells.
Lateral plate mesoderm is produced by the same mechanism of gastrulation as its paraxial partner but, unlike its paraxial confrèr, it does not have an intermediate form of organization analogous to somites. First off, recall that the primary role of LPM is the production of the cardiovascular system, smooth muscle (as in the gut) the viscera, and the appendicular skeleton. Forward from neuromere c1 the role of LPM is strictly cardiovascular. Posterior to c1 the “portfolio” of LPM expands to form the bones of the appendicular skeleton and the smooth muscles of the gut and genitourinary system.
LPM has two regions: cardiac mesoderm (CM) and posterior lateral plate mesoderm (PLMP). PLMP is collinear directly behind CM and begins at neuromeric level c1. It is organized along the anterior–posterior axis of the embryo in the same way as somites, using Hox genes to establish positional specification. The separation of PLPM into somatic (LPMS) and splanchnic (LPMV) layers does not take place in the agnathic state. It is present in all gnathostomes [7] (Fig. 10.50).

Lateral plate mesoderm evolution, plate 3. [Reprinted from Onimaru K, Shoguchi E, Kratani S, Tanaka M. Development and evolution of the lateral plate mesoderm: Compariative analysis of amphiosus, and lamprey with implircations for the acquisitions of acquired fins. Dev Biol 2011; 359(1):124–136. With permission from Elsevier]
In any case, LPM cell populations remain in register with the neuromeric level of the primitive streak from which they were produced and from which they are innervated. LPM of the head and neck receives its coding from the neural crest to penetrates it and surrounds it to make the fascia. Thus, scapula arises from LPM at levels c2–c6. The middle third esophagus has a mixture of PAM striated muscle and LPM smooth muscle, thus retaining somatotopic nocioception, whereas the muscle of distal third is purely LPM.
Phylogeny of Lateral Plate Mesoderm
Paraxial mesoderm with its subdivisions into somites has been present since the dawn of vertebrate evolution. Lateral plate mesoderm, in contrast, has undergone several iterations. The primitive state of LPM is seen in the cephalochordate amphioxus (the lancet), Branchiostoma. While its PAM is organized into myotomes. Its lateral plate mesoderm is not regionalized. The Forkhead gene FoxF is expressed throughout. Kuratani, working with lamprey, a living agnathan, established that its LPM is specified by Hox genes. Furthermore, it is regionalized into an anterior cardiac zone (CLMP) and a posterior truncal zone (PLMP). Lamprey CLPM has separate somatic and visceral layers but PLMP remains unseparated. No genes associate with limb production in gnathostomes are present in lamprey CLMP.
At the stem of gnathostomes, two changes take place in PLMP that are key to the production of fins/limbs. Its gene profile changes, such that it has a somatic layer, a requirement for the appendicular skeleton. Furthermore, it now expresses novel hox genes Tbx4 and Tbx5. TBX4/5 induces Fgf10 in mesoderm and Fgf8 in overlying ectoderm, both steps being requisite for limb formation. Nested expression of these genes specifies the “fin field.” As we shall see in our discussion of cervical neurology, the expression of Hox genes is involved in a shift from the ancestral pattern of neurons controlling the muscles of the pectoral fin to a new position coinciding with disconnection of the pectoral girdle from the skull.
Neuromeric Analysis: How to “Code” Lateral Plate Bones
Where does mesenchyme of clavicle and scapula originate? Neuromeric analysis of muscles having insertions/attachments to these bones allows us to “map” the bones into developmental zones. We can also reconstruct the order in which the bone fields are assembled. Here’s an example. Let’s recall (once again) that the origin of a muscle is given by the neuromeric location of its motor nerve. Levator scapulae is supplied by the dorsal scapular nerve (C3–C5), thus its myoblasts arise from the seventh, eighth, and ninth somites (S7–S9). The insertion/attachment of a muscle will be to a bone field that is registered with C3–C5. Coding is not determined by the secondary (distal) insertion) of the muscle. Levator scapulae develop from paraxial mesoderm. Levator scapulae make its primary insertion into the medial border of the scapula; its attachment is a triangular zone just medial to the spine. Its secondary insertions are tendinous slips to the transverse processes of atlas, axis, and the third and fourth cervical vertebrae. Thus, the mesenchyme making up this region of scapula originally came from lateral plate mesoderm in register with neuromeric levels C3–C5.
Warning
Traditional anatomic texts as misleading. Gray’s Anatomy uses the term “origin” in a purely descriptive way that has nothing to do with where the myoblasts of a particular muscle arise. In many instance (levator scapulae is a good example), the attachments of a muscle are exactly the reverse of its development. This is not to bash Gray. He simply described muscles arbitrarily based upon what he considered to be their fixed point (origin) and movement point (insertion). Moore’s text uses the terms “proximal attachment” and “distal attachment,” but again, uses them in a non-developmental way, leading often to a reversal (levator scapulae again). Such archaic terminology prevents us from seeing the unitary relationships that exist between bones and muscles in their earliest state.
Muscles attach to bone fields in a specified order. This can be determined by the spatial position of their motor nerves. At first glance, the process required to map out the bone fields of the appendicular skeleton (pectoral girdle and upper extremity) appears a bit more complex when compared to the mapping of the axial skeleton (skull and cervical vertebrae). Here’s why. Cervical muscles supplied by the medial motor column (spinal nerve) are midline or paramedian. Those specifically from C1 to C3 form the post-hyoid hypobranchial column which extends forward from the pectoral girdle, runs beneath the pharyngeal arches, and attaches via hyoid to the mandible. These muscles act as depressors of the lower jaw. Cervical muscles supplied by the lateral motor column relate to outboard structures. These are all hypaxial/ventral, even though, for evolutionary reasons, muscles associated with the scapula appear to occupy a “dorsal” position.
Embryonic muscles seeking attachment to the shoulder girdle and limb have complicated migration patterns. That is because their bony targets develop in an inboard–outboard, proximal–distal direction. Recall the evolutionary sequence: stylopodium, zeugopodium, and autopodium. Furthermore, the postaxial bone units subdivide, whereas the axial units simply add on linearly. For this reason, the segmental innervation of upper limb joints follows a strictly linear sequence with shoulder abductors and lateral rotators supplied by C5 while the intrinsic muscles of the hand are innervated by C8, T1. Finally, as the upper limb develops, it undergoes a series of movements: internal rotation of the humerus > elbow flexion (stage 20, 51 days) > pronation. These movements cause different zones of the developing bones to be “presented” to the developing muscle masses for attachment. This is particularly true for the humerus.
Because upper extremity myoblasts have complex migration patterns, their motor nerves often coalesce and then separate. This results in a plexus. The game plan of the cervical plexus is the most simplistic. Its superficial branches innervate the skin while its deep branches are motor. Muscles supplied are: (1) the anterior and lateral capitus group; (2) the superficial hypobranchial muscles connecting the ventral midline with hyoid and mandible; (3) the diaphragm; (4) the most medial muscles connecting head and cervical spine to should girdle, and; (5) a muscle (scalenus medius) connecting cervical spine to the first rib (c8–t1).
Brachial plexus is a far more complicated switchyard. Its organization is proximal–distal and also postaxial–preaxial: roots > trunks > divisions > cords > branches. Muscles are spatially organized in the same order. As one goes farther out the chain, more muscles are supplied.
Roots: dorsal scapular = levator scapulae + rhomboid; long thoracic = serratus.
Trunks: nerve to subclavius; suprascapular = supraspinatus + infraspinatus.
Divisions: no branches.
Cords: pectoral nerves; subscapular = teres major; thoracodorsal = teres minor + latissumus; and axillary = detoid. Note that the axillary nerve (C5–C6) is the last motor nerve of the posterior cord. It marks the endpoint for all muscles with primary (proximal) insertions to the shoulder girdle.
In our next section, we shall describe the embryology of the clavicle and scapula. Detailed information regarding the neuromeric coding of each muscle is presented. Illustrations depicting the developmental sequence are labeled using neuromeric terminology.
Hox Gene Mapping of the Neck Bones: New Rules of Engagement
The shoulder girdle represents an interface zone between the head, neck, and trunk. Our assumptions up until this point regarding what bone comes from neural crest and what bone comes from mesoderm have been based upon a classical ossification model in which bones formed by membranous ossification are presumed to arise from neural crest whereas those formed by chondral ossification are assumed to be of mesodermal origin. In the neck, these rules become flexible and give rise to the muscle scaffold model in which the mesenchymal origin of a region of bone is determined by the composition of the fascia accompanying its various muscle attachments. All muscle fibers are composed of mesoderm but their connective tissue elements and their fascial envelope can be neural crest or mesoderm. Muscles will attach.
As we shall see, clavicle and scapula are composite structures with certain zones developing from neural crest and others from mesoderm depending upon the muscle inserted into that zone. Furthermore, we find that post-otic neural crest (PONC) from r8 to r11 has multiple fates. (1) It synthesizes the cartilaginous structures of the fourth and fifth arches, that is the thyroid, cricoid, and arytenoids cartilage. (2) It contributes to the walls of the superior thyroid artery and its branches supplying the fourth and fifth arches. (3) It provides connective tissue and fascia for the muscles of the tongue as well as for the sternocleidomastoid and trapezius muscles.
Experimental work fundamental for our understanding of how the clavicle and scapula develop was reported by [8] using Wnt-1 and Sox-10-Cre recombinase-mediated fat mapping to answer three fundamental questions. First, can PONC form the chondral bone? The answer is yes. (2) Are the dermal bones behind the otic capsule strictly neural crest? The answer is no; in this zone, mesoderm can form membranous bone as well. (3) Does the distribution of neural crest and mesoderm into the shoulder girdle correlate with ossification type or muscle insertion sites? The answer is that insertion sites determine bone mesenchymal identity.
Note that, for the purposes of neuromeric coding, only the primary insertion of a muscle is important (Figs. 10.51, 10.52).

Mesenchymal analysis of plectoral girdle show diversity of ossification. Highly conserved neck muscle scaffolds (red) attach (hatched areas) on a shoulder skeleton (boxes 1–4) that displays variable dermal (light grey) and endochondral (dark grey) ossification type. Attachment regions (hatching) of the gnathostome trapezius muscle (T) are endochondral in sharks, salamanders5and all amniotes but are dermal in fish and frogs. A = limb muscles, Co1 and Co2 (coraco-branchialis and coraco-hyoideus); L = limb skeleton; R = trunk muscles; S = sterno-cleido-mastoid. In the shoulder skeleton, box 1 is the dorsal cleithrum (dermal) in bony fish (Polyodon or Amia) and frog (Rana) and the scapular region (endochondral) in salamander, mouse and living amniotes; box 2 is the acromio-coracoid (endochondral); box 3 is the clavicular region (dermal/dermal+endochondral), although in sharks, bone is absent and its space is taken by part of the scapulo-coracoid (stippled); and box 4 is the sternal region, comprising the sternum (endochondral) or connective tissue (bony fish). [Reprinted from Matsuoka T, Ahlberg P, Kesaris N, Iannerelli P, Dennehy U, Richardson WD, McMahon AP, Koentges G. Neural crest origins of the neck and shoulder. Nature 2005; 46:437–355. With permission from Springer Nature]

Genetic lineage labelling of PONC and somitic mesoderm. Otic PONC (green) and mesodermal (blue) populations are indicated. Note that the spine of scapula, the former cleithrum, is PONC. [Reprinted from Matsuoka T, Ahlberg P, Kesaris N, Iannerelli P, Dennehy U, Richardson WD, McMahon AP, Koentges G. Neural crest origins of the neck and shoulder. Nature 2005; 46:437–355. With permission from Springer Nature]
The sternocleidomastoid is innervated by the spinal accessory nerve from neuromeric levels r8–r11. It has an additional sensory supply from c1 to c4. Thus, although the muscle arises from four somites (occipital 1–4), it receives neural crest from neuromeric levels r8–r11 and c1–c4 which forms the connective tissue and fascia of SCM. These relationships play a role in the primary insertion of SCM to the crest of the occipital bone. Trapezius has a similar formulation.
Numerous pathologies obtain for defects in PONC. Pathological flexibility of PONC differentiation. Connective tissues can be over-expressed entirely leading to “pseudo” macroglossia (in patients with trisomies). PONC fate to become bone may change to soft tissue mesenchyme, as in Arnold-Chiari syndrome. PONC fated to become connective tissue may be for ectopic bone, as in Klippel-Feil syndrome (Fig. 10.53).

Pathologies of PONC differentiation. (1) Changes in PONC connective tissue per se. Trisomy pseudo-macroglossia (a). All tongue-muscle connective tissues are entirely derived from neural crest (blue areas) explaining enlarged tongues as neurocristopathic. (2) Changes in PONC fate from bone/periosteum (b) into connective tissue can explain localized cervical defects in cleidocranial dysplasia (c) where neural spines fail to close and Arnold-Chiari syndrome I + II (h). In the latter, the PONC-derived clivus (blue in g) of the otherwise mesodermal (mes) cranial base, which serves as the attachment point (attp) for pharynx constrictor muscles (constr. Phar. N.IX/X.) in front of the medulla (med), fails to form and is replaced by fragile connective tissue. (3) Changes in PONC fate from connective tissues into bone. CT of pharynx constrictor muscles (const.phar. N.IX/X in g that are connected to cervical vertebrae (e) can ectopically become a vertical bridge of bone (f), leading to neck immobility either in Klippel-Feil syndrome (f) of as ectopic, ‘omovertebral’ bones inside trapezius territory (stippled line in d) inpatients with Sprengel’s deformity, a frequent finding in Klippel-Feil syndrome 19, 20. Note also the concomitant loss of the PONC-derived (but not mesodermal) scapular spine in patients with Sprengel’s deformity (d). [Reprinted from Matsuoka T, Ahlberg P, Kesaris N, Iannerelli P, Dennehy U, Richardson WD, McMahon AP, Koentges G. Neural crest origins of the neck and shoulder. Nature 2005; 46:437–355. With permission from Springer Nature]
The Clavicle
Clavicle is a curvilinear bone derived and synthesized from postotic neural crest and lateral plate mesoderm from neuromeric levels c1–c4. It articulates between the manubrium sterni and the acromion of the scapula. Functionally it serves as a fulcrum enabling muscles to extend the upper extremity laterally. Animals using anterior prehension make active use of the clavicle but, in many carnivores, it is simply suspended in the muscle sling. For many reasons, the clavicle is unusual. Despite its position, draped over across the root of the neck, it belongs to the piscine head-trunk interface and is associated with the cucullaris muscle. Recall that cucullaris spans between the primitive pectoral girdle to the dorsal tips of the respiratory branchial arches, as far forward as r6–r7 (Figs. 10.54, 10.55, 10.56).

Development of the clavicle. Clavicle consists of a neural crest outer coating and a mesodermal core. Coding is c1– to c4–c5 from medial to lateral, Lateral articulaltion with acromion which is c4–c5. [Reprinted from Fawcett J. The Development and Ossification of the Human Clavicle. Am J Physiol 1913; 47(pt2):225–234]

Clavicle, superior. Each muscle has a primary insertion. Trapezius should be colored red. As part of the pectoral girdle, all muscles connecting to head follow an antegrade–retrograde have a primary insertion pattern. All muscles connecting clavicle to humerus are antegrade–antegrade. Trapezius is unusual: upper fibers are antegrade–retrograde and lower fibers are antegrade–antegrade. All insertions on clavicle are primary and are colored red. Trapezius has a primary insertion into clavicle and should be colored red. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Clavicle, inferior. As part of the pectoral girdle, all muscles connecting to head follow an antegrade–retrograde have a primary insertion pattern. All muscles connecting clavicle to humerus are antegrade–antegrade. Trapezius is unusual: upper fibers are antegrade–retrograde and lower fibers are antegrade-antegrade. All insertions on clavicle are primary and are colored red. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
But the limb migrates in mammals down to the level of c5 with muscles attaching to the scapula from this position. Clavicle follows along with the chondral component, scapulocoracoid, of the pectoral girdle. Thus, the phylogeny of the human clavicle reflects the caudal migration of the limb. Clavicle develops in two ways. Its external surfaces, both anterior and posterior, form in membrane. Clavicle is the first bone of the body to ossify, reflecting its relationship to the first post-cranial neuromeres. Recall that the original insertion of the pectoral chain was to the post-temporal bone of the occiput, likely at position r10–r11. Therefore, the most proximal zone of clavicle should code from c1. Recall that ossification of the cranium starts at the occipital somites and sweeps forward.
Clavicle ossifies from three constant centers, two primary and one secondary. At stage 17 (14 mm, 41–42 days), the clavicle is a mesenchymal condensation of mixed neural crest derivation slung between two mesodermal bones: the first rib and the acromion of the scapula. These latter two bones develop from paraxial mesoderm and lateral plate mesoderm, respectively. Within the band, medial and lateral zones of cartilaginous transformation (precartilage) appear. This “core of the clavicle” represents the ability of PONC to form endochondral bone. Next, two primary dermal ossification centers appear, one ventral and one dorsal. The ventral ossification center is of PONC derivation while the dorsal ossification center is from the lateral plate mesoderm. Mesoderm thus forms membranous bone on the back side of the clavicle. The two membranous centers quickly fuse around the chondral core by day 45. Next, the sternal and acromial zones become true cartilage from which endochondral ossification extends outward from the shaft. Expansion of diameter takes place via subperiosteal deposition [9, 10].
The medial center is responsible for 2/3 of the total length. Two secondary centers appear around age 20. The sternal center is constant and represents the remnant of the procoracoid while the acromial center is variable. The nutrient artery of the clavicle is derived from the suprascapular artery. It enters via a foramen at the lateral end of the subclavius groove.
As discussed previously, the early tetrapod pectoral girdle had a separate bone, the interclavicle, positioned medial to the clavicle. In the synapsid line, the interclavicle persists in therapsids and monotremes but in mammals, it is described as absent. We can postulate that the interclavicle may continue to exist as the secondary ossification site at the sternal border. The manubrium is physically different; it is a derivative of the ancient anterior coracoid process. The manubrium is a mammalian invention. We shall see that its original form fuses with interclavicle to create a larger structure with articulations for both clavicle and firest rib (see the development of manubrium).
Three hypaxial muscles having insertion on the clavicle define the neuromeric origins of the bone and divide the bone into two developmental fields. The connective tissue and insertions of all three muscles are derivatives of paraxial mesoderm The medial field is defined by sternohyoid (C2–C4), which attaches along the upper and anterior surfaces. The lateral field is defined inferiorly by subclavius (C5–C6) and by deltoid (C5–C6) superiorly. Note that the subclavius is innervated from the lateral trunk of the brachial plexus. Deltoid being situated more laterally has a most distal innervation from the posterior cord of the brachial plexus. This indicates that the subclavius gains access to the clavicle first, while the deltoid arrives later and thus is positioned more laterally. All remaining muscles attached to clavicle are hypaxial, with primary insertions from either head or trunk. The upper extremity has no secondary insertions into clavicle.
Thus, clavicle arises from postotic neural crest r8–r11 and lateral plate mesoderm in register with neuromeric levels c2–c6. Its two developmental fields each have a primary ossification center. Clavicular LPM from neuromeric levels c2–c6 associates itself medially with PAM from levels c8–t1 that forms the sternum and first rib and, peripherally, with lateral plate mesoderm assigned to the upper extremity.
Clinical Correlation: Cleidocranial Dysostosis
This neurocristopathy affects membranous bones, in particular causing hypoplasia or absence of the clavicle and an enlarged patent anterior fontanelle. Frontal and parietal bossing are prominent. Cranial suture may display small Wormian bones. Nasal bones may be missing. Hypoplasia of the zygoma is accompanied by faulty maxillary sinus development with chronic sinusitis. The sygomatic arch is thin or even discontinuous at the zygomaticotemporal suture. Mandible has a slime ramus and pencil-like coronoid. The mandibular symphysis is patent. Enamel issues and dental maldevelopment, including delays in eruption, are present. Excessive development of the dental lamina leads to hyperdontia and crowding (Figs. 10.57, 10.58).

Cleidocranial dysostosis. Absent collar bones, fforehead is prominent. Maxillary hypoplasia, both vetical and horizontal with mandibular prominence. X-ray shows open fontanelles, underdeveloped sinuses. The atlantoaxial complex is impacted up against the skull base. [Reprinted from Garg RK, Agrawal P. Clinical spectrum of cleidocranial dysplasia: a case report. Cases Journal BMC 2008; 1: 377–381. With permission from Creative Commons License 2.0: http://creativecommons.org/licenses/by/2.0]

Cleidocranial dysostosis. Multiple ectopic teeth, flattened nasal bridge, hyperteolorism is seen. [Reprinted from Garg RK, Agrawal P. Clinical spectrum of cleidocranial dysplasia: a case report. Cases Journal BMC 2008; 1: 377–381. With permission from Creative Commons License 2.0: http://creativecommons.org/licenses/by/2.0]
Mapping the Clavicle
Manubrium sterni is derived from anterior coracoid process. It codes to C1–C2.
It gives primary attachment to sternothyroid and sternohyoid.
Neural crest contributions to clavicle
Cartilagenous core is postotic neural crest r8–r11.
Stenocleidomastoid brings neural crest from its distal fibers to the medial 1/3 of anterior.
Medial 2/3 of Clavicle: C2–C4
Sternocleidomastoid arises from S5 to S6. Its motor supply is from the C1 to C2 roots of spinal accessory nerve. Its primary attachment to clavicle is at the anterior (neural crest) upper border. This muscle is branchiomeric with neural crest connective tissue likely originating from r8 to r11 leading it to connect it secondarily to the skull.
Pectoralis major arises from S9 to S13. It is supplied by C5, C6, C7, C8, and T1. It has two primary insertions: a clavicular head along the anterior lower border and a sternocostal head. Its secondary attachment is to the humerus.
Sternohyoid (lateral fibers) from C2 to C4 has a primary attachment situated at posterior (mesodermal) medial border, adjacent to manubrium. It ascends as part of the straps to the third arch border of the hyoid bone.
Lateral 1/3 of Clavicle: C5–C6
Subclavius This obscure muscle arises from S9 to S10. It is supplied by the subclavian nerve; that is, the trunks of C5–C6. It has its primary insertion within a groove on the inferior border of clavicle. Its secondary insertion is to first rib (T1 = S12–S13). Subclavius precedes deltoid.
Deltoid This large muscle arises from S9 to S10 as well. It is innervated by the axillary nerve, that is, by C5–C6 from the posterior cord. It has three primary insertion sites. The anterior fibers of this muscle have their primary insertion from anterior and superior lateral border of clavicle. Middle fibers are proximally attached at the acromion of the scapula whereas posterior fibers are located along the posterior border of the scapular spine.
Trapezius (posterior) has a primary insertion into posterior border of clavicle.
The Scapula
The scapula is a flat bone of triangular shape that it lies like a pancake over the dorsal surface of ribs 2–7. It is personified by 3’s. Three borders face superiorly toward the neck, medially toward the spinal column and laterally toward the armpit. Three angles make the triangle, superior sharp surface for the scapular elevator muscles, an inferior dagger for teres to pull the scapula toward the humerus and for serratus to retract it toward the chest wall, and a lateral labium to receive with humerus. Three processes projecting outward from the home plate in three different directions anchor the scapula and determine its movements. The spine projects dorsally and create two fossae for muscle attachments. Projecting laterally from the spine is an outward-looking peninsula, the acromion, designed for articulation with clavicle. The coracoid process projects ventrally and gives attachment to the three muscles, pectoralis minor is medial; lateral is biceps brachii and coracobrachialis (Figs. 10.59, 10.60, 10.61).

Scapula: neuromeric coding. The scapular plate is created from lateral plate mesoderm via chondral osteogenesis; scapular spine is LPM dermal bone. The bones are in register with neuromeres c3–t1. Connective tissues of pre-scapular muscles down to c3 have neural crest connective tissue. Post scapular muscles have mesodermal connective tissue. Muscles having primary insertion into scapula are in three classes. (1) Intrinsic muscles insert into the surface of the scapula and control internal/external rotation of shoulder joint and humeral abduction (subscapularis, teres minor and the supra- and infra-spinatus muscles). (2) Extrinspc muscles insert into projections of the scapula, that is, coracoid process, supra- and infra-glenoid tubercle of scapula, and scapular spine (biceps, triceps, and deltoid). (3) Stabilizers and rotators of the scapula muscles (trapezius, serratus, levator scapulae, rhomboids). [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Vascular supply of the scapula. Upper (rostral) half of scapula is supplied from the subclavian artery. Although subclavian has three parts, only thyrocervical trunk is important. First part of subclavian gives rise to thyrocervical trunk with three branches. After inferior thyroid goes into the neck suprascapular artery supplies structures along the clavicle as far as acromion; it then passes over to supply dorsal scapular muscles (supra- and infra-spinatus). Transverse cervical artery supplies the elevators (levator scapulae, rhomboids) and deep plane muscles of scapula to the second part of subclavian produces costocervical trunk. Second part of subclavian lies beneath scalenus anterior and gives rise to costocervical trunk with two branches, neither of which supplies scapula. Supreme intercostal goes down to posterior ribs 1–2 and pleura. Deep cervical goes up the neck in parallel to inferior thryoid. Third part of subclavian in 50% of cadavers gives rise to descending scapular artery which substitutes for deep branch of transverse cervical. The lower (caudal) half of scapular is supplied by axillary artery. Axillary artery also has three parts, scapula being supplied from the second and third. First part of axillary artery produces one artery, supreme thoracic (supremely unimportant) to pectorals and anterior chest wall. second part of axillary artery gives rise to two arteries: thoracoacromial trunk and lateral thoracic both perfuse the scapula. (1) Thoracoacromial has 4 branches. Pectoral branches go to the muscles attached to clavicle and coracoid process. Clavicular branch supplies subclavius. Acromial branch goes all the way to distal acromion and the humeral head. Deltoid branch supplies the muscle as its insertion into acromion. (2) Lateral thoracic artery descends along the chest wall to supply serratus. Third part of axillary artery has three arteries. (1) Subscapular artery supplies scapula via two branches. Circumflex scapular is larger and goes through triangular space to infraspinous fossa. It supplies the teres twins and subscapular. Thoracodorsal is smaller and is the continuation of subscapular to latissimus. (2 and 3) Anterior and posterior humoral circumflex are exclusive to the neck of humerus. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Ossification centers of the scapula. [Reprinted from Anwar I, Amiras D, Khanna M, Walker M. Physes around the shoulder girdle: normaldevelopment and injury pattern. Clin Radiol 2016; 71(7):702–209. With permission from Elsevier]
In conceptual terms, the scapula is a floating platform for the upper extremity that, via multiple muscles attached to the head, spine, and thoracic cage and via its articulation with the clavicle, permits placement of the hand in space. The mesenchymal origins of scapule demonstrate it to be mostly LPM.
Scapula appears shortly after stage 23, that is, during fetal life. It is synthesized from lateral plate mesoderm (LPM) in register with neuromeres c2 through t1. It also bears contributions from neural crest [11,12,13]. The overall flow of mesenchyme is from dorsal-to-ventral. This is evidenced by the identical innervation of the supra/infraspinatus twins (C5–C6) from the upper trunk which lie dorsal to the bone plate versus subscapularis (C5–C6) from the more distal posterior cord which occupies the ventral position. Scapular spine represents the ancient coracoid and bears the attachment of the trapezius from neuromeres c2 to c6. Bone formation has an epicenter at the suprascapular notch (zone c1–c2) from which it radiates both medially and laterally, and then downward. As the coracoid process represents an add-on, it is not surprising to find that this zone represented LPM from c5 down to t1.
Scapula ossifies from eight distinct centers: body (1), coracoid process (2), acromion (2), medial border (1), inferior angle (1), and lower glenoid rim (1). Ossification of the body of scapula takes after stage 23. It is initiated at the scapular neck, a collar-like ring of bone holding outward the glenoid fossa like a chalice. From there a flat quadrilateral plate of bone appears. In the third month, the blade of the scapula extends downward and the scapular spine, a derivative of the original anterior coracoid process, projects dorsally. This reflects the rich blood supply around the humerus and thence surrounding the borders of scapula. Failure of ossification at the acromion can create a painful non-union, os acromiale (Figs. 10.61, 10.62).

Os acromiale. [Reprinted from Barbier O, Block D, Dezaly C, Sirveaux F, Mole D. Os acromiale, a cause of shoulder pain, not to be overlooked. Orthopaedics & Traumatology: Surgery & Research 2013; 99(4): 465–472. With permission from Elsevier]
At birth, the scapula is largely osseous but all parts having to do with articulation with the clavicle and humerus remain cartilaginous: the glenoid, coracoid process, and acromion. Those parts having articulation with the thoracic cage, vertebral border, and inferior angle, also remain cartilaginous. The remaining flat surfaces develop via membranous ossification. Final ossification takes place during adolescence in this order: coracoid process, proximal acromion (base), inferior angle and lower vertebral border, distal acromion (apex), and upper vertebral border. These sites reflect vascular growth patterns. Superolateral border represents the axillary artery spreading outward, first ventrally toward coracoid and then dorsally to acromion. Inferomedial border is the scapular arcade: inferior half is subscapular and is followed by superior half, the descending branch of the transverse cervical.
Mapping the scapula: the insertion sequence of appendicular muscles.
Terminology note: At the primary insertion site a muscle can be said to arise without implying anything about its origin. Muscles always originate from the somite(s) of their motor nerve(s).
In all instances, appendicular muscles associated with the pectoral girdle have their primary insertions in the LPM bones with which they are in register. Bones mature in a fixed biological sequence: their binding sites become progressively available in a fixed spatiotemporal order: deep-to-superficial, distal-to-proximal, and ventral–dorsal. We can make the following generalization: for any given neuromeric level, muscles compete for binding sites according to the “landing site sequence.” As myoblasts enter an extremity they migrate first to the distal limit and then proceed to add up, like water filling up a glass.
Once a primary attachment is accomplished, muscles seek out their secondary insertion site(s). Those supplied by the cervical plexus swerve back to secondary targets at more proximal neuromeres along the midline. Strap muscles (C1–C3) from somites S5–S7, for example, seek out primary insertions into the manubrium and clavicle and thence rebound backward toward the body following the ventral margins of the branchial arches and terminating in the midline geniohyoids. Pectoral girdle muscles supplied by brachial plexus proceed forward to secondary targets at equal or more distal neuromeres along the body wall or upper extremity. Pectoralis minor (C8–T1) attaches first to coracoid process and inserts into third–fifth ribs (T3–T5).
What can insertion patterns reveal about the biological zones of the scapula? One boundary zone seems to occur along the scapular blade (both dorsal and ventral aspects), and along its margins. Levator scapulae (C3–C4) are inserted into the upper zone whereas the rhomboids (C5–C6) from brachial plexus insert below. The order of muscle attachments into specific layers is also useful. As scapula matures, it attracts insertions in the same sequence. Supraspinatus (C5) inserts above the spine; the larger infraspinatus (C5–C6) have a more distal root and inserts caudally. Thus scapular development proceeds, as predicted, from cranial to caudal. This gives us a picture of “mesenchymal flow” over scapula. Subscapularis and supra/infraspinatus attach first. The former grabs onto the chest wall and the latter two claim the highest position along humerus. Lateral border muscles such as teres minor and major come in next. They must attach to humerus more distally. Coracoid process muscles would be laid down in a late sequence. A great “cover-up” then takes place with the antegrade retrograde muscles attaching. On the ventral side and lateral sides, pectoralis major and deltoid are subcutaneous. Trapezius fulfills the same function over dorsal scapula. It covers over its competitors, supra- and infraspinatus (Fig. 10.63).

Muscle layering over the scapula. Muscles connecting pectoral girdle to the axial skeleton at or above neuromere t1 represent reassignment of previous muscles and continue to innervated by medial motor column, either directly or as cervical plexus. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Superior Border: LPM from c1 to c4
Omohyoid (inferior belly) arises from S6 (and sometimes S7). It is supplied from cervical plexus C2 (C3). The primary insertion is immediately medial to the suprascapular notch. It joins with an anterior belly that develops from S5 (S6). Together, the muscle sling makes a secondary insertion into hyoid just lateral to sternohyoid. Recall that we are coding manubrium as a c1 PAM bone. Spine of scapula—a c4 derivative—divides muscles connecting with the head from those extending medially toward the body. Blood supply to the upper omohyoid is lingual (second and third arches) and to lower muscle from superior thyroid (fourth and fifth arches). Thus, the vascular supply betrays the function of this muscle as it connects the pectoral girdle to the pharyngeal arches.
Acromion: LPM from c5 to c6
Deltoid (middle fibers) arises from S9 to S10 and is supplied by axillary nerve (C5–C6, posterior cord). Because it spans from pectoral girdle to humerus its blood supply reflects distinct zones of axillary artery. The proximal muscle receives the thoracoacromial from second part of axillary artery while the distal muscle is supplied by the anterior and posterior humeral circumflex branches from third part of axillary. At the tip os of.
Coracoid Process: LPM from c5 to t1
This process projects forward from the upper border at almost a right angle. It lies below the lateral one-fourth of the clavicle. It has three muscles associated with it. Recall that the coracoid process represents the incorporation of the ancient posterior coracoid into scapula. It develops from medial to lateral, that is, short head of biceps > coracobrachialis > pectoralis minor. Because the latter two muscles have a common motor nerve, coracoid has two ossification centers: an inferior center represents the musculocutaneous components while the superior center represents the medial pectoral nerve-supplied pectoralis minor.
Biceps (short head) arises from S9 to S10 and is supplied by musculocutaneous nerve (C5–C6). The primary insertion is at the lateral apex of the coracoid process (in common with coracobrachialis). It terminates at the tuberosity of the radius (posterior side).
Coracobrachialis arises from S10 to S11 and is supplied by the musculocutanous nerve (C6–C7), the terminal branch of lateral cord. The primary insertion is at the medial apex of the coracoid process. It has its secondary insertion on the humerus between the primary insertions of triceps, proximally, and brachialis. Blood supply comes from the third part of axillary as anterior humeral circumflex.
Pectoralis minor arises from S12 to S13 and is supplied by the medial pectoral nerve (C8, T1) from brachial plexus and medial cord. The primary insertion is the superior aspect of the process. It fans out to insert into the chest wall at ribs 3–5. Pectoralis minor defines the three parts of axillary and is supplied by thoracoacromial axis from its second part. Its medial border also receives the superior thoracic artery, the sole branch from first part of axillary artery.
Medial Border, Posterior: LPM from c3 to c5
Levator scapulae arises from S7 to S9 and is supplied by spinal nerves of C3 and C4 plus C5 via dorsal scapular nerve. Its primary attachment is to superior medial border of scapula above the spine. It ascends to make secondary attachments to the transverse processes of atlas and axis, C3, and C4. It is supplied by two vessels. Ascending cervical is from inferior thyroid branch of thyrocervical trunk whereas transverse cervical arises directly from thyrocervical trunk, from first part of subclavian immediately lateral to the vertebral axis.
Rhomboid minor and major both originate from S8 to S9 and are supplied by a branch of dorsal scapular nerve, C4, C5. Its primary insertion is to inferior medial border of scapula below the spine. Blood supply is from transverse cervical artery from thyrocervical trunk. This latter, being proximal to suprascapular, descends under the rhomboids to reach the tip of scapula.
Medial Border, Anterior: LPM from c5 to c7
Serratus anterior arises from S9 to S11 and is supplied by long thoracic nerves C5, C6, and C7. It has its primary insertion to medial border of scapula. Secondary insertions are sought out to ribs 1–8 (and sometimes to ribs 9 and 10). It is supplied by the lateral thoracic artery from second part of the axillary artery.
Lateral Border, Posterior: LPM from c5 to c7
Triceps brachii (long head) arises from S9 and is likely supplied by the upper components (C5–C5) of radial nerve (C5–T1) from the posterior cord. The secondary insertion is at the infraglenoid tuberosity: exactly the location of an ossification center! The primary supply of triceps is distal, away from the muscles associated with scapula, specifically from profunda brachii from superior ulnar collateral. However, the proximal zone of the long head is perfused from posterior humeral circumflex.
Teres minor arises from S9 to S10. It is supplied by axillary nerve, C5, and C6. Its insertion is upper 2/3 of posterior lateral border and the lateral 1/3 of the fossa. It has a dual blood supply, the proximal scapular zone perfused by circumflex scapular, from subscapular artery, the largest branch of the third part of axillary artery. The distal zone is supplied by posterior humeral circumflex.
Teres major arises from S9 to S11. It is supplied by lower subscapular nerves C5, C6, and C7. Its primary insertion is from the lower 1/3 of the posterior lateral border and the lateral 1/3 of the fossa.
Pectoralis minor arises from S12–S13 and is supplied by medial pectoral nerve (C8 and T1) from brachial plexus, medial cord. The primary insertion is superior aspect of the process. It fans out to insert into the chest wall at ribs 3–5. Like teres minor, it also has a dual blood supply with proximal zone from thoracodorsal closer to the chest wall leading to posterior humeral circumflex at the arm.
Dorsal Scapula: LPM from c5 to c6
Supraspinatus arises from S9 to S10; it is supplied by suprascapular nerve (C5 and C6) from the upper trunk. It has an insertion from the medial 2/3 of the fossa. Two arteries perfuse it. Suprascapular artery is a branch of thryocervical trunk, dorsal scapular artery is the deep branch from transverse cervical, and also a branch of thyrocervical trunk.
Infraspinatus has the same characteristics as supraspinatus. Its blood supply comes from suprascapular and circumflex scapular, a descending branch of subscapular.
Ventral Scapula: LPM from c5 to c6
Subscapularis arises from S9 to S10; it is innervated by subscapular nerve (C5, C6), the first branch from the posterior cord. The branch sequence is subscapular > axillary > radial. This large muscle is perfused by branches from three sources: axillary to the upper lateral half, and subscapular to the lower lateral half, with a contribution of suprscapular.
Scapular Spine: LPM from c3 to c6
Trapezius has primary insertion along the entire spine from lateral to medial and is encoded probably c3–c4 to c6. It has three distinct vascular zones. The upper zone is supplied by the transverse muscular branch of occipital artery, with neural crest related to the second arch. This may relate to the embryologic part of the muscle that originally was associated with the hindbrain. The middle zone is supplied by the transverse cervical artery from thyrocervical trunk from the first part of subclavian. The lower zone is supplied by dorsal (descending) scapular artery which arises from the second or third part of subclavian.
Clinical correlations of pectoral girdle coding: the strap muscles demystified.
From neuromeric levels, r8–r11 and c1–c3 somites produce two groups of hypobranchial muscles that follow a curvilinear pathway downward and forward to their targets. They bear the sobriquet hypobranchial because they must migrate around the boundaries of the arches. The first group from S1 to S4 provides the extrinsic muscles of the tongue and has primary attachments from the second arch (styloglossus and hyoglossus) and first arch (genioglossus) into the tongue matrix. S5 produces genioglossus which is mistakenly included with the tongue muscles…really is the anterior extension of the strap muscles.
The second group of hypobranchial muscles comes from S5 to S7 and is known as the strap muscles. These descend downward first to the sternum and clavicle and thence upward beneath the fourth and third arches to attach to the hyoid bone. Their primary insertions are into the sternum, manubrium, and clavicle in medial-to-lateral sequence. Because none of the strap muscles are innervated by c4, the insertion of the omohyoid stops short of distal clavicle.
Note that second arch platysma muscle fibers drape over the clavicle but are separated from it by neural crest SIF fascia whereas the fasciae enclosing muscles with true attachments to the clavicle is DIF and is mesodermal.
Phylogeny of the Pectoral Girdle
Life on land would be unthinkable without locomotion. The transition from fins into limb is a fascinating story, well worth an entire chapter (or even a book). Because the appendicular skeleton is well represented in the fossil record, we can directly observe the structural details of animals long extinct; we can also follow the phylogenetic changes leading to the modern human limb. For our purposes, we shall focus on the relationship of the pectoral girdle (and perforce the upper extremity) to the head and neck. In our previous discussion of the neuromuscular organization of the neck, we noted that the spinal accessory nerve and the eight cervical nerves innervate several functional classes of muscles: (1) cranium to pectoral girdle, (2) cervical vertebrae to cranium, (3) cervical vertebrae to each other, (4) cervical vertebrae to pectoral girdle, and (5) cervical vertebrae to trunk. Obviously, the development of the clavicle and scapula is inseparable from that of occipital bone, mastoid, and the cervical column…but how? [14] (Figs. 10.64, 10.65, 10.66, 10.67, 10.68, 10.69).

The pectoral fins in actinopterygian fishes, seen here as Polypterus, is suspended from a pectoral girdle with two components. (1) 5 neural crest dermal bones (red) and mesodermal chondral bones (blue). The dermal series hugs the posterior margin of the opercular series covering the gills. Post-temporal articulates with the skull and clavicle extends to the gulars. Chondral scapulmesodermal articulates with 3 fused basal pterygiophore. These elements are discrete in chondrichthyans. Multiple radial pterygiophores extend outward but are united by distal arc of bone. Lepidotrichia (lepido—too many to count) complete the fin. The pectoral girdle in sarocopterygian Eustheopteron retains the dermal bone series but shows the etrapod form of chondral bones: stylopodium, zeugopodium, autopodium. [Courtesy of William E. Bemis]

Eogyrinus Attheney represents the basal tetrapod stance for terrestrial life. Pectoral girdle overlaps initial 3 cervical vertebrae. [Reprinted from Wikimedia. Retrieved from: https://commons.wikimedia.org/wiki/File:Eogyrinus_Attheney._Wellcome_M0006666.jpg. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by/4.0/deed.en]

Reduction of dermal bones. [Reprinted from: http://bio.sunyorange.edu/updated2/comparative_anatomy/anat_3/a_shoulder.htm. With permission from Dr. Walter Jahn]

Tetrapod limb design show shift of the limb axis to the ulna. Stylopodium (purple) = purple. Zeugopodium (orange) = ulna/radius. Autopodum (yellow) = hand/foot [Courtesy of William E. Bemis]

Comparison of pectoral girdle in amniotes from †Dimetrodon to primitive mammal, Didelphusi, the opossum. Cleithrum is relocated to dorsal aspect of scapula as the scapular spine, terminating in acromion and glenoid fossas. Anterior coracoid becomes manubrium; it maintains its relationship to calvicle. Posterior coracoid becomes the true coracoid process. [Courtesy of William E. Bemis]

Evolution of pectoral girdle. Tetrapods replace dermal bones of fishes with LPM. Early tetrapods lose posttemporal and gain interclavicle. Note: cucullaris in fishes inserts into anocleithrum. Loss of this bone at petoral dissociation forces cucullaris to relocate back to scapula. Note: the function of clavicles and coracoids is to serve as a brace against the sternum. Coracoid develops from embryonic coracoid plate. Anterior ossification centers give rise to procoracoids; posterior ossification centers give rise to coracoids. Clavicle absent in most non-avian reptiles; present in most mammals. [Reprinted from McGonnel IM. The evolution of the pectoral girdle J Anat 2001; 199:189–194. With permission from John Wiley & Sons]
Let’s get a quick bird’s-eye view of the subject. The appendicular system consists of pectoral and pelvic girdles that support the limbs. The girdles differ radically in composition. The pectoral girdle appears first in evolution with Tiktaalik; it is composed of both dermal and endochondral elements. The pelvic girdle occurs later in time with tetrapods Acanthostega/Ichthyostega; it is strictly endochondral. The existence of a dual design suggests differing evolutionary mechanisms.
Endochondral bones arose from the basal supporting structures of the forefin. During evolution, these lateral plate structures expanded outward into the surrounding skin envelope, where dermal bones derived from the original encasing bones were encountered. Thus, coracoid articulates posterior to the neural crest bone series that is suspended between the skull and the gular bones and follow the posterior borders of the opercular series.
Primitive fishes carry a variety of projecting spines, knobs, and lobes. Fins are different: they are thin and membranous; they also are internally supported by fin rays. Like scales, fin rays develop at the epidermal/dermal interface, but subsequently sink into the dermis, where they are known as dermal fin rays. These take various names. Ceratotrichia are keratinzed rods in elasmobranchs. Bony fishes have fin rays and lepidotrichia; these are often ossified. The proximal fin near the body is reinforced by pterygiophores. The proximal fin contains basals while slender radials extend from the basals into the substance of the fin. Fishes have two types of fins. Paired fins are pectoral and pelvic; these articulate with girdles inside the body wall; the rest are singular. The upper limb arises from the pectoral fins.
Paired fins were an early innovation in piscine evolution, bringing added mobility and stability to fishes swimming within a watery environment in constant motion. They first appeared in the ventrolateral fin folds of early agnathans. Ostracoderms had only pectoral fins but later ancient fishes (placoderms, acanthodians, and the chondrichthyans) had both pectoral and pelvic girdles. Rhipidistians, such as Eusthenopteron, made use of fleshy fins that acted as pivots.
The tetrapod limb, formally termed the chiridium, is, by definition, digit-bearing. It always consists of three regions. The stylopodium is closest to the body; it is always singular (humerus and femur are analogous). The middle limb, zeugopodium, always has two elements: ulna and radius (tibia and fibula) in the forelimb. The distal upper limb, termed autopodium, contain the carpus and manus (tarsus and pes).
The pectoral fin was originally attached to the skull via a series of dermal bones: postemporal, supracleithrum, postcleithrum (anocleithrum), cleithrum, clavicle, and (later) inteclavicle. The evolution of the pectoral girdle in advanced teleosts and tetrapods involved the progressive loss of dermal elements. In the transition to tetrapods, the endochondral element of fishes, scapulocoracoid, became dominant with two separate ossification centers: scapula and procoracoid.
Early amniotes added a brand-new endochondral element, the coracoid (=posterior coracoid). Modern tetrapods retain both “coracoids.” Procoracoid becomes the dominant ventral shoulder bone in amphibians, reptiles, and birds, whereas therian mammals (that includes us) retain only the coracoid. Anterior coracoid is the likely source of the upper manubrium, where it fuses with lateral plate mesoderm to form a single structure. The cleithrum persists as the spine of the scapula.
Origin of Paired Fins: The Fin-Fold Theory and Homeotic Genes
In the mid-nineteenth century, E.M. Balfour and J.K. Thacher put forth a remarkably prescient theory regarding limb position that turned out to have a molecular rationale. Fishes were considered to have a continuous set of ventrolateral fin folds. These were reinforced by two kinds of endoskeletal struts called pterygiophores. These had two components: basal bones were proximal and extended inward; distal to the basal were radial bones. Over the course of time, the basal elements fused across the midline to form the shoulder and pelvic girdles. Dermal bones, originally part of the skin of armored fishes, became “add-ons” to the pectoral girdle (but not to the pelvis) (Fig. 10.64a).
Several lines of evidence support the fin-fold theory; these range from indirect paleontologic findings to molecular mapping. Prehistoric agnathic fishes such as Myllokunmingia and Haikouichthyes had fin folds. Presumably, pectoral and pelvic fins arose at the same time. Shark embryos have continuous lateral wall ectodermal ridges.
Fins arise at specific neuromeric levels. Jarvik made two important contributions. First, he hypothesized that the mesenchymal core of the fin folds was capable of forming endoskeletal basals and radials. These, in turn, would serve as attachment sites for muscles from nearby segmental myotomes, providing control of the fin. Overlying scales became supportive dermal fin rays, an event documented in the embryogenesis of many living fishes. Dermal bone consolidation offered convenient sites of attachment along the anterior surface of the bones. At the same time, the dermal girdle would form the posterior wall of the pharynx, could protect the heart, and would provide attachment sites for pharyngeal arch muscles.
Jarvik emphasized the importance of the pectoral girdle at the transition point between axial musculature and branchial (pharyngeal) arch slits (Kardong, pp. 246–247). If we assign one mesodermal element (somitomeres or somite) per arch, then the primitive fish formula is 3 somitomeres +12 somites. The formula for sharks is 3 + 4; that for mammals is 3 + 2. Chondrichthyan fins arise at the fourth somite. Recall that the maximum number of branchial arches documented in the fossil record is 15. Thus, the primitive fish fin can arise as far back as the 12th somite. The pectoral fin zone extends from S4 to S12. In mammals, the S12 level marks the transition between the seventh cervical vertebra and the first thoracic. By extension, the mammalian pectoral zone extends from the OCJ down to T1.
Homeotic genes provide the underlying basis for the fin-fold hypothesis. Expression of Engrailed-1 defines the ventral compartment of the embryonic body. Within the ventral zone, pre-vertebrates such as amphioxus express a single T-box gene, AmphiTbx4/5. This is the ancestral gene of the fin fold. In sharks, further genetic takes place, with the anterior fin defined by Tbx5 and the posterior limb by Tbx4. The tetrapod limb makes use of an entirely new gene, Sonic hedgehog, to accomplish three revolutionary changes: (1) Shh promotes outgrowth of the limb from the body wall; (2) it liberates the limb from being in parallel with the body, and; (3) it sets up a proximal-to-distal limb axis.
The Pectoral Girdle in Fishes
Ostracoderms Although agnathic fishes dating back to the Cambrian had ventrolateral fin folds, paired pectoral fins made a rudimentary appearance in these armored fishes. They were definitely not buoyant. Like acanthodians, chondrichthyans, and placoderms, the ostracoderms lacked a swim bladder. These bottom-dwellers made use of their folds to undulate over the ocean floor.
Placoderms appeared in the Silurian and radiated widely, being active swimmers with strong jaws. They had a fused endoskeletal pelvis and a pectoral girdle consisting of several dermal bones attached proximally to the postemporal bone. These served as armor plate and reinforced a singular endoskeletal bone, the scapulocoracoid.
Chondrichthyans used their fins as stabilizers. In more derived sharks the paired basal components extended along the midline of the body and fused into a U-shaped scapulocoracoid bar. More caudally they formed a similar puboischiac bar as well. Chondrichthyans do not form membranous bones. From the get-go, they lost all dermal contributions to the shoulder girdle. In their modern iteration, sharks have three large pterygiophores extending off the pectoral girdle.
Acanthodians had fins with large spines along their leading edges. The sole manifestation of a pectoral girdle was a connection between a pectoral spine and the scapulocoracoid.
Actinopterygians are modern bony fishes. They have a pectoral girdle that is mostly dermal. Because they have a swim bladder, these fishes are buoyant, so they use the pectoral fins for maneuvering. The shoulder girdle is U-shaped and draped behind the gill chambers. The dermal girdle attaches to the skull by means of the posttemporal bone. This is followed by a supracleithrum to which are attached, in some fishes, additional dermal bones. Specifically, postcleithrum (also called anocleithrum) is found in this position. The largest dermal element, cleithrum, articulates with endoskeletal scapulocoracoid. Finally, a mixed membranous-chondral bone, clavicle, extends across the midline beneath the gill chambers to fuse with its opposite member as a symphysis. It does so via the union of paired intervening bone field, the interclavicles. This is not the source for manubrium (it originates from endochondral procoracoid).
Dermal elements of the pectoral girdle are found in bony fishes, sarcopterygians, and early tetrapods before fading from the scene. Following Kardong, we shall refer to these collectively as the “postcleithrum,” even though this breaks with traditional nomenclature. Comparative anatomists apply the term anocleithrum in sarcopterygians and tetrapods to the postcleithrum of actinoptyerigians. This redundancy makes the literature confusing.
Sarcopterygians have dermal fins supported by muscles and internal supportive elements. These are represented today by three kinds of lungfishes and the deep-dwelling coelacanth, Latimeria. These have reduced shoulder elements. Fortunately, fossil rhipidistians left behind a detailed record of their appendages. These points the way toward tetrapod evolution. Eusthenopteron in the late Devonian had dermal fins supported by bones homologous to tetrapods. Pectoral fin articulates first with scapulocoracoid and a series of supporting dermal bones. A new element, interclavicle, is first seen here. It will be retained in some later tetrapods, but not in mammals (Fig. 10.64b).
The Pectoral Girdle in Tetrapods
Early tetrapods spent the early part of their life cycles in water. Skulls of Europs show the continued presence of an aquatic sensory system, the lateral line canals. Between the Carboniferous and Permian periods, tetrapod girdles and limbs increased in strength as an adaptation to weight bearing. The primitive state is shown by Eogyrinus, a Carboniferous tetrapod. Despite a length of nearly 6 feet, it had relatively small and cartilaginous limbs. Movement was likely similar to that of salamanders, in which the limbs merely serve as pivot points around which the body swings and sways. The limbs of Europs were stout and densely ossified, consistent with a primary function of ambulation (Fig. 10.65).
The pectoral girdle changed significantly between rhipidistian fishes and tetrapods.
Concomitant, or shortly after, arrival on land, tetrapods lost the posttemporal bone. This freed up the shoulder from the skull and eliminated the constant jarring of the skull from ambulation. The dorsal series of dermal bones, such as supracleithrum and postcleithrum, are lost. The remaining elements were ventral: endochondral cleithrum, dermal clavicle, and paired interclavicles that met in the midline in front of the neck. In early amniotes cleithrum “disappears.” In its place are two distinct endochondral entities: a dorsal scapula and a ventral coracoid (Fig. 10.66).
Innovations in the Tetrapod Limb
Tetrapod limbs are characterized by a common developmental plan. Although all limbs possess an axis of symmetry, the postaxial elements predominate. The most proximal element is axial and singular: the stylopodium (humerus and femur). A second-order zeugopodium consists of two elements: the postaxial ulna and preaxial radius order elements that develop from the postaxial primordial. The subsequent development of tertiary elements is very different. All branching units of the manus are postaxial in origin. The preaxial element can produce linear third and fourth-order structures but cannot branch. This genetic pattern is very ancient. Chondrichthyans, primitive actinptyerygeians, and rhipidistian fossils all show a metapterygian stem branching into postaxial elements. Comparative analysis of Hox gene expression in vertebrate limbs shows that tetrapods deform the straight metapterygial axis of fishes. The axis becomes bent anteriorly to support a distal limb such that all the digits arose from the posterior elements (Fig. 10.67).
In primitive amniotes (reptilomorphs) a brand-new endochondral element appears, confusingly named the new coracoid. It is located just behind the original coracoid. We can avoid getting fouled up in terminology and also track the evolution of these components by giving them separate names. The older bone element, homologous to the original tetrapod coracoid, is the anterior coracoid or procoracoid. The newer bone element is the posterior coracoid, or metacoracoid.
The three-part scapula evolved in two directions. Sauropsids (birds and reptiles) maintain the three-part version. Procoracoid is the dominant ventral shoulder bone. Synapsids, preserve the dermal elements to varying degrees. Clavicle is retained across the board in all taxa. Monotremes continue to possess interclavicles but marsupials and mammals lose the interclavicles, thus permitting direct articulation between clavicles and sternum. In mammals, the two coracoids split up, allowing the glenoid fossa to face ventrally instead of posteriorly. This positions the upper limb beneath the thorax and is useful for enhanced mobility. Posterior coracoid remains behind and fuses to the scapula as the coracoid process. It is positioned ventral to clavicle. Anterior coracoid shifts medially to form the manubrium. Mammalian evolution found dorsal repositioning of the otherwise hypaxial pectoral girdle to be a selective advantage as it gave greater leverage for limb control. Scapula shifted dorsally to provide more efficient insertions. Cucullaris, a branchial arch dilator (and therefore hypaxial) split apart into sternocleidomastoid and trapezius. The former remained in register with clavicle while the latter chose the dorsal margin of scapula—and an opportunistic series of insertions along the “siderails” of the thoracic spine (for details, vide infra) (Figs. 10.68 and 10.69).
The Sternum: Phylogeny and Development
The sternum is classified as part of the appendicular skeleton. It develops in humans by chondrification of lateral plate mesoderm in the distal interneuromeric spaces. The presternal cartilages are distal to the ribs. Because they come from LPM they do not have parasegmentation. Therefore they readily undergo longitudinal fusion (much like internal thoracic/mammary artery) to create a sternal band. These subsequently fuse with one another in the midline. Fusion failures result in a cleft sternum or even ectopia cordis. Longitudinal failures can occur as well (Figs. 10.70, 10.71).

Development of the sternum BMC. Sternum develops from ossified lateral plate mesoderm cartilages in the distal interneuromeric space. These are distal to, and separate from, the corresponding ribs. Because they are LPM (and do not come from somites) these distal components do not have parasegmentation. In the parasternal position the segments unite longitudinally. Failure of fusion of both sides is seen in sternal fissures. Left: [Reprinted from Carlson BM. Human Embryology and Developmental Biology, sixth edition. St. Louis, MO: Elsevier; 2019. With permission from Elsevier.] Right: [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Failure of fusion of the sternal bands can be either cranil-caudal or caudal-cranial. A superior vertical defect is seen, In severe cases this may lead to ectopia cordis. Left: [Reprinted from Acastello E, Majluf R, Barbosa LM, Peredo A. Sternal cleft: a surgical opportunity. J Ped Surg 2003; 38(2): 178–183. With permission from Elsevier.] Right: [Reprinted from Engum S. Embryology, sternal clefts, ectopia cordis, and Cantrell’s pentad. Semin Ped Surg 2008; 17:154–160. With permission from Elsevier]
Stenum is lacking in fishes. It is not seen in stem fossil tetrapods like Acanthostega, perhaps because its origins were in cartilage, but it is present in amphibians so we assume it appeared somewhere in the tetrapod transition. Amphibian ribs do not fuse with the sternum. Amniotes achieve a fused condition.
In synapsids, interclavicle is a presumed candidate as the source of the sternal bands. Interclavicles quickly become interposed between the clavicles, forcing them to retreat laterally. Shortly after the initial syapsid state, paired manubria appear caudal to the interclavicles; they fuse together into manubrium. From its very inception manubrium has an articulation. Basal theriodonts show shortening of interclavicles such that they equal the manubrium in size. In the crown state, theriodonts show complete incorporation of interclavicles into the manubrium to become the first sternal segment. Upper part of manubrium (interclavicle) therefore articulates with clavicle and lower part of manubrium articulates with the first rib (Fig. 10.72).

Phylogeny of the pectoral girdle of basal mammals and their relatives. Note the presence of an interclavicle (red), clavicles (green) and a new bone, the manubrium (deep blue), which develops where the sternum develops in other tetrapods. Note zone of clavicle-interclavicle contact (dark green) in immediate pre-mammals (4) Interclavicle is interposed between manubrium and clavicle. Evolution of interclavicle and anterior sternal structure in the cynodont–mammal transition. (Placement of Pseudotribos in mammal phylogeny is shown in Supplementary Information.) a, Cynodont Probelesodon. b, Mammaliamorph Bienotheroides. c, Mammaliaform Sinoconodon. d, Pseudotribos. e, Monotreme Ornithorhynchus. f, Tachyglossus. g, Eutriconodont Jeholodens. h, Marsupial Didelphis. Apomorphies of (1) mammaliaforms: widening anterior end of interclavicle; posterior club-foot of interclavicle to the same width of manubium for extensive attachment of pectoralis muscles. (2) Yinotheria: ‘T-shaped’ manubrium. (3) Monotremata: gracile and elongate lateral process of interclavicle overlapping two-thirds the length of clavicle; enlargement of (meta)coracoid to articulate directly with the interclavicle; presence of procoracoid for strengthening the girdle. (4) Theriimorpha: shortening of interclavicle to equal manubrium; reduction of the lateral process of interclavicle; mobile claviculo–interclavicle articulation. (5) Crown Theria: incorporation of embryonic interclavicle into the manubrium, the first sternal segment in articulation of clavicle and thoracic rib 1. cl, clavicle (green); cl–ic contact zone between clavicle and interclavicle, articulation or overlap of clavicle and interclavicle (yellow); ic, interclavicle (red); mb, manubrium sternebra 1 (blue); mc, metacoracoid; pc, procoracoid; sc, scapula (grey); stb2, second sternebra; tr1 and tr2, thoracic rib costal cartilage 1 and 2, respectively. [Reprinted from Luo Z-X, Ji Q, Yuan C-X. Convergent dental adaptations in pseudo-tribosphenic and tribosphenic mammals. Nature 2007; 450:93–97. With permission from Springer Nature]
Neurology of the Neck
Introduction
Go on, jump in, the water is fine! People say ridiculous things like this when you are standing at the edge of a swimming pool…and you know the water is not warm. So why should we have to immerse ourselves in things such as the spinal cord, the cervical roots, and the cervical plexus? And what should we gain from our efforts?
Our goals are simple: we are out to discover the what, the how, and the why of the neck. Like any building, the structure of the neck depends upon a blueprint. In previous chapters, we have sought out relationships between the nervous system and peripheral tissues. We’ll follow the same methodology here. We shall see that the dermal innervation patterns of the neck and its various muscles reflect a spatial blueprint within the spinal cord. When we compare the motor columns of the brain with those of the spinal cord, a remarkable and very functional system emerges. Muscle structures clustered around the vertebral axis are the most “primitive.” They are supplied from the midline. As fishes became sophisticated enough to develop fins, a new system of peripheral muscles evolved to furnish control. The vertebrate body in some way anticipates the head. Thus, the anatomic “idea” of spatial separation of neurons seen in the spinal cord is “copied” in the design of the head muscles and their motor nuclei. We shall look at how these patterns are reiterated.
As cervical nerves leave the cord they undergo a series of simple and predictable changes that will convey them to their respective targets. There must be a way to separate out the nerve supply for axial versus peripheral structures. Muscles have to be very careful about how they find their target insertion sites, sometimes their pathways are circuitous. After all, a lot of things are going on in the neck. It’s hard to find your way around! But the cervical plexus was not designed by Salvador Dali. Its goals are simple: The plexus will organize and spatially distribute sensory nerves to broad swatches of skin. It does so in a very logical way. The plexus will also separate out motor nerves for several groups of muscles: (1) muscles connecting the axial skeleton to itself, controlling movements of the head and spine, (2) the peripheral control muscle of respiration, the diaphragm, (3) muscles connecting the axial skeleton to the appendicular skeleton, that is, the shoulder girdle.
We have left out one final group of muscles, the non-striated ensheathment of the esophagus. Because it is derived from lateral plate mesoderm its motor control is from the vagus nerve. We will consider the esophagus separately.
We are left with the larger question: how and why did the neck evolve? This will be the subject of a separate later section in this chapter. It requires a multi-system approach including the development of fins and the transition to limbs, structural requirements of air-breathing, and the whole issue of where the cervical neuromeres came from in the first place. One thing is sure…the history of the neck itself is embedded in its neuromuscular design. The muscles of the neck are arranged in functional classes according to the grand evolutionary design of the region. Positioning of the head for feeding and survival is the responsibility of the axial muscles. Life on land demands an air pump. Respiratory and digestive passages must be separated. Limbs provide locomotion. So, understanding of blueprint of the cervical nerves can lead to a rock-solid appreciation for how function follows form.
A Cook’s Tour of the Spinal Cord: Grey Matter
Heads versus tails…the best place to understand the vertebrate blueprint is, of course, the fish trunk. Here, developmental segments are laid out in a repetitive sequence along the spinal cord. A series of septae divide the myotomes into epaxial (dorsal) and hypaxial (ventral) components. Motor nerves follow the same terminology. Hypaxial muscles are supplied by ventral roots. Epaxial muscles receive dorsal roots.
How are these neurons laid out in the spinal cord? We all are familiar with cross sections of the cord in which grey matter is shaped like the letter “H,” or like a butterfly. In the center of the H is a central canal with spinal fluid. The horizontal limb of the H straddles the canal as the dorsal and ventral commissures. At any given level, the ventral horn contains afferent somatic motor neurons whose axons will leave the cord via ventral roots. Localized collections of afferent SANS motor neurons form the lateral horn. The dorsal horn marks the termination of efferent primary sensory fibers entering the cord via the dorsal roots to create sensory columns Remember: spinal cord = neural tube = a flat neural plate rolled up on itself like a handlebar mustache or a cigar. Thus, as you “follow the curve” of spinal cord grey matter, you are merely progressing from medial to lateral. When you unroll it and flatten it out, voila! You wind up recreating the original neural plate (Figs. 10.73, 10.74, 10.75, 10.76, 10.77).

Motor and sensory tracts of the spinal cord. Grey matter has three horns with three kinds of nuclei: ventral horn, c1 to s4 = somatic motor, (GSE) lateral horn, t1 to l3 = SANS motor, (GVE); dorsal horn (posterior columns), s5 to c1: non-nocioceptive sensory (GSA). White matter conveys 3 motor tracts (but no nuclei) from forebrain (corticospinal), midbrain (rubrospinal), and hindbrain (vestibulospinal) and and 1 nocioceptive sensory tract. Motor tracts of the spinal cord Grey matter has three horns with three kinds of nuclei: ventral horn, c1 to s4 = somatic motor, (GSE) lateral horn, t1 to l3 = SANS motor, (GVE); dorsal horn (posterior columns), s5 to c1: non-nocioceptive sensory (GSA) White matter conveys 3 motor tracts (but no nuclei) from forebrain (corticospinal), midbrain (rubrospinal), and hindbrain (vestibulospinal) and and 1 nocioceptive sensory tract. Sensory tracts of the spinal cord Touch and pressure, kinesthesia pathways: dorsal columns of the extremities (1) gracilis (blue) = leg, (2) cuneatus (green) = arm, and (2) Nocieptive pathways (red): = spinthalamic tracts (anterior and lateral) Propioceptive pathways (yellow) spinocerebellar tracts convey position information from the body to the brain: anterior (Gower’s tract) and posterior (Flechsig’s tract) [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones].

Cranial nerve columns of the brain stem Motor = red Sensory = blue. Somatic motor column (red) supplies midline muscles unrelated to the pharyngeal arches. It is analogous to medial motor column (MMC) in the spinal cord. Dorsal vagus (blue) supplies PANS motor to glands. Nucleus ambiguus column, r6–r11 (purple) supplies pharyngeal arch muscles (palate, pharynx, larynx), continuous with central motor column (CMC) in the cervical spinal cord, but not analogous to lateral motor column (LMC) in the spinal cord. Somatic sensation (orange) is represented by a singe nucleus of spinal tract of trigeminal. Spinal V is somatotopic. Taste (green) is in same neuromeric zone as facial and nucleus ambiguus (facial nucleus not depicted). [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Motor neuron columns and Hox proteins in the mammalian spinal cord. Motor neuron columns are generated at specific positions along the rostrocaudal axis of the spinal cord detrmined by the homeotic code. Hox6 specifies c1–c8. Hox9 specifies t1–t12. Hox10 specified ls3. Lateral motor columns (LMCs) are generated at brachial and lumbar limb levels and innervate limb muscles. At thoracic levels, the preganglionic column (PGC) innervates the sympathetic ganglia (scg), whereas the hypaxial motor column (HMC) innervates the body wall muscles. Median motor columns (MMCs) are present at all levels of the spinal cord and innervate the dorsal and ventral axial musculature (note: ventral motor neurons to spine not shown here). Hox6, Hox9, and Hox10 are expressed in specific regions of the spinal cord along the rostrocaudal axis and direct the identity of motor neurons and their connectivity to peripheral targets. Hox6 controls brachial LMC identity, Hox9 controls PGC and HMC identity, and Hox10 controls lumbar LMC identity. Note [Reprinted from Murakami Y, Tanaka M. Evolution of motor neurons to vertebrate fins and limbs. Dev Biol 2014; 355(1):164–172. With permission from Elsevier]

Neuromeric organization of spinal motor columns. Schematic summarizing the segmental distribution of spinal motor columns (adapted from Dasen and Jessell, 2009). While the medial motor column (MMC, brown) is present all along the rostro-caudal axis, the spinal accessory column (SAC, purple) is restricted to the five first cervical segments (C1–C5). The phrenic motor column (PMC, red) is confined between C3 and C5. The preganglionic column (PGC, orange) extends through the thoracic segments until the second lumbar segments (L2) as well as well as between sacral segments 2 and 4 (S2–S4). The hypaxial motor column (HMC, light blue) is exclusive of the thoracic segment where as the lateral motor column (LMC, dark and light green) is located at limb levels: brachial (C5–T1) and lumbar segments (L1–L5). Note conspicuous absence of representation for the cervical plexus muscles, which belongs to MMC. Note phrenic motor nerve (unique in mammals) may have belonged originally to medial group of brachial plexus. Note cervical level is defined by Hox5. [Reprinted from Stifani N. Motor neurons and the generation of spinal motor neuron diversity. Fronters in Cellular Neuroscience 2014; 8: article 293. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by/4.0/]

Motor columns in the cervical spine Note the position of phrenic nerve and spinal accessory nerve intermediate between medial motor column (c1–c8) to axial muscles and lateral motor column to brachial plexus Cervical plexus strap muscles are not represented but belong to MMC, as they represent original axial muscle that pre-date the evolution of the upper extremity. LMC neurons, c5–t1, have a new set of genetic markers and occupy a new position in the cord. MMC, medial motor column; PMC phrenic motor column; SAC, spinal accessory column; LMC, lateral motor column; HMC, hypaxial motor column; PGC, preganglionic column (SANS motor) [Reprinted from Stifani N. Motor neurons and the generation of spinal motor neuron diversity. Fronters in Cellular Neuroscience 2014; 8: article 293. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by/4.0/]
Ventral Horn
If we look at the spinal cord as a whole, ventral horn neurons run in three vertical columns, often extending through many segments. The horns are organized into medial, central, and lateral cell columns. But remember, our focus is segmental, so the best way to appreciate functional relationships is on cross-section. Medial cell groups supply axial musculature. Lateral cell groups supply the limbs: (1) trunk-to-limb, (2) intrinsic limb, and (3) hand/foot. Don’t get fooled. Some texts describe limb and foot as being dorsal to trunk. In reality, they are just “following the outer curve” of ventral horn.
Remember: the ventral horn is like a handlebar mustache. When you unroll it, you come up with the original neural plate.
Medial motor column extends the entire length of the spinal cord. The homolog of MMC in medulla is r8–r11 hypoglossal nucleus.
-
MMC medial (MMCM) supplies the epaxial erector spinae group. These muscles extend the head, neck, and trunk. They are innervated via dorsal primary rami.
-
MMC lateral (MMCL) innervates the following hypaxial groups: the prevertebral muscles of the neck, the intercostals, and anterior wall muscles of the trunk. These muscles flex the neck and trunk. They are innervated by ventral primary rami.
-
MMC cervical plexus involves neuromeres c1–c4. It supplies the strap muscles of the neck. It provides sensation to the hypaxial neck down to clavicle.
Central motor column has two discrete parts. These are found exclusively in the cervical spinal cord and consist of neuromeres c1–c6. Its functional importance is disproportionate to its size.
-
Spinal motor column (SMC) supplies sternocleidomastoid and trapezius. It is located at the ventral periphery of ventral horn, extending from c1 to c6. In the myology section, we shall see how this rationalizes the insertions of these two muscles. The cervical roots become confluent as the spinal root of the cranial nerve XI, the neuromeric definition of which makes it truly accessory, if not downright irrelevant. The homolog of SMC in the medulla is r6–r11 nucleus ambiguus.
-
Phrenic motor column (PMC) supplies the diaphragm. It is interposed between SMC and MMC and runs between c3 and c5. The primary root of phrenic nerve is from C4 but it gets contributions from C3 and C5.
Lateral motor column is discontinuous. It supplies muscles that are evolutionarily new. The neurons are always ventral and they pass through a new-fangled structure…every student’s nightmare…a plexus. LMC has no homolog in the medulla.
-
Brachial plexus (neuromeres c5–t1): all roots are both motor and sensory to the upper extremity.
-
Lumbosacral plexus (neuromeres t12–s4): roots l2–s3 are motor to the lower extremity; other roots supply muscles of the pelvis.
-
Coccygeal (pudendal) plexus (neuromeres s5–cx1): strictly sensory.
Lateral Horn
Lateral horn contains the cell bodies of pre-ganglionic sympathetic autonomic nervous system (SANS) neurons. These project laterally from the “H” from the first thoracic to the second lumbar segments. This column (GVE) is lateral to LMC in the neural plate and therefore lies dorsal to it in the spinal cord. Because these neurons are motor, they must exit via the ventral root. But the SANS neurons, like escaping criminals, must return to do their time on the vertical “chain ganglia.” Because the criminals have surrendered (waving a white flag) they leave the ventral root and return to the ganglia via white rami communicantes. The ganglia are vertically connected. In this way, SANS neurons ascend to populate three cervical ganglia, the largest one with the greatest distribution being the superior cervical ganglion. Craniofacial SANS fibers run with the branches of carotid system and therefore are in the company of the neural crest fibers that envelope these arteries. Homeotic coding of this neural crest at various junctures along the arterial system may serve as a signal for the formation of individual branches and these carry SANS fibers with them to their respective targets.
Nuclei of pre-ganglionic paraympathetic nervous system neurons (PNAS) are located in the nuclei of cranial nerves, III, VII, IX, and X and S2–S4 in the spinal cord. In the hindbrain, r6–r7 inferior salivatory nucleus and r8–r11 dorsal motor column of the vagus form a single, albeit discontinuous column which, just like SANS, is lateral to LMC in the neural plate and dorsal to it in the neural tube. In spinal cord, S2–S4 have the same location but the neurons are small and do not occasion an outward projection for the “H.”
Dorsal Horn
The spinal cord conveys two types of sensory information back to the brain:
-
Proprioception (fine touch, pressure, and position sense) are carried in grey matter.
-
Nocioception (pain and temperature) is carried in white matter.
Grey matter contains proprioceptive nuclei and fibers in the dorsal horns. These are conveyed upward to cerebellum and higher centers for processing. Beginning caudally, the neurons add up from medial to lateral creating, in the thoracolumbosacral cord a single nucleus gracilis, representing the lower extremity and trunk. In the cervical cord, complex information from the upper extremity and neck necessitates an additional and physically separate nucleus cuneatus. The posterior columns are thus somatotopic.
A Cook’s Tour of the Spinal Cord: White Matter
Three motor tracts (no nuclei) convey impulses from the tri-partiate embryonic brain.
-
Hindbrain is ventral (medial) and carries the vestibulospinal tract.
-
Midbrain is intermediate and contains rubrospinal tract.
-
Forebrain is dorsal and has the corticospinal tract.
These descending tracts add up from medial to lateral. Thus, primitive fibers from hindbrain occupy the midline (ventral) position while those from the cortex are the most lateral and are dorsal to the other two tracts.
White matter contains nociceptive nuclei and fibers in the ventral periphery and conveys them to the thalamus for processing. Like proprioception, these add up from medial to lateral. The anterior spinothalamic tract is medial–ventral representing the trunk and lower extremity while the posterior spinothalamic tract is lateral/dorsal and carries additional information from more rostral levels.
Spinal Cord Tracts Are Somatotopic, That Is, Neuromeric
Although the pathways conveying sensory information from the body up through the spinal cord to the brain are also very different from motor pathways, both systems make use of somatotopic organization. The peripheral nervous system is of neural crest construction (the Schwann sheath) each nerve connects a neuromeric level with target tissues produced at that level. The main center for somatic information from the head and face is trigeminal nucleus. From the neck on down, we see the pathways separate out. Somatic information is conveyed in the spinothalamic tracts. These are spatially organized by function. We’ll use the neural plate model, so from medial-to-lateral, the nociceptive sequence is pressure > touch > pain > temperature.
The spinothalamic tracts are also laid out by neuromeric segments in a ventral-to-dorsal sequence: s5 > c4. The initial sacral fibers are ventral. Thus, as one proceeds upward, the fibers add up and the spinothalamic tract gets bigger. Recall that the nociceptive tract for the head and upper neck is r1–c4. The so-called trigeminal nucleus has a spinal extension down to c4. Why? Once again, we observe the developmental importance of the c4–c5 junction. It represents the primary (head-related) neck versus the secondary (trunk-related) neck.
Proprioception, exteroception, and vibration are conveyed in the posterior columns (dorsal funiculi). The spatial layout of these tracts is diametrically opposite that of their spinothalamic siblings. Functional modalities have a dorsal-to-ventral sequence: touch > pressure > movement > vibration > pressure. The segmental sequence is medial-to-lateral: c4 > s5. Morphologically the columns are described as fasculi. The more medial fasciculus cuneatus contains mid-thoracic to sacral fibers. The more lateral fasciculus gracilis contains mid-thoracic to sacral fibers. Thus, as one proceeds upward along the cord the posterior columns get wider.
The Unique Role of C4 as a Neuromeric “Faultline”
Pain referral to the trigeminal nucleus continues downward to C4, thereafter being referred to a separate structure for the spinothalamic tract. Is there something about the fourth cervical neuromere that separates a “head-associated neck” from a “trunk-associated neck?” Actinopterygian fishes have three occipital somites, S1–S3, each having a motor nerve to the pectoral fin. The actinopterygian head-trunk junction is S3/S4. The addition by tetrapods of a fourth occipital somite shifted the head/trunk junction backward from one neuromere to S4/S5. Liberation of the pectoral girdle from the head was accompanied by a backward transposition of the pectoral girdle to level C4. The original blueprint for brachial plexus in all tetrapods, except mammals, consists of four roots. These brachial plexus nerves, br1–br4, always bracket the head/trunk interface, the trunk being represented by br4. Thus, in basal amniotes (reptilomorphs) with six cervical vertebrae, the brachial plexus consists of C4–C5–C6–T1. Muscles interposed between the head and pectoral girdle continued to retain branchiomeric neural crest connective tissues and retained neurologic connections to these tissues as well. Thus, the existence spinal nucleus of V down to c4 reflects the persistence of these relationships. Somatic sensory events distal to c4 are referred to the trunk-associated spinothalamic tract.
Spinal Nerves: Anatomic Components
What could be more boring that reviewing spinal nerves? Let’s spiff up this necessary section by a radical statement. There are 35 (not 31) pairs of spinal nerves: 4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 1 coccygeal. [Recall that there were originally 5 coccygeal somites but these normally regress.] There is good evolutionary evidence to indicate that spinal accessory nerve is a strictly peripheral nerve that becomes incorporated into the jugular foramen during the addition of the fourth occipital somite. After all, both its target muscles, SCM and trapezius, are part of the pectoral girdle (we will consider this fascinating story later).
In any case, all spinal nerves except XI and C1 make their exit through intervertebral foramina. The first cervical nerve is designed to exit from beneath the proatlas (the vertebral product of somites 5–6). Because proatlas is absorbed into the skull, C1 exits about atlas. C2, obediently, travels below atlas. Recall from a developmental standpoint how the components of the foramina flow around the pre-existent nerves. XI is, of course, an exception. Indeed, as we shall see, it does not have a true cranial root and it is not a cranial nerve. The spinal roots C1–C4 of this transitional nerve no longer are individually inserted into the cucullaris. Instead, they link up with one another longitudinally. The first root, C1, becomes entrapped by the incorporation of the fourth somite and winds up exiting the jugular foramen in the company of vagus. The roots of the transitional nerve keep adding up caudally and the nerve finally exits via foramen magnum.
-
Ventral (anterior) roots are centrifugal; they contain motor neurons from anterior and lateral grey column of the cord.
-
Dorsal (posterior) roots convey centripetal fibers carrying information back from the periphery to the brain. Each rootlet pierces the dura but remains covered by pia mater and thence by arachnoid. In the neck, dorsal roots are three times bigger than the ventral roots.
-
Spinal ganglia are situated in the intervertebral foramina. Just behind them lies the vertebral artery.
-
Spinal nerves refer to the zone just beyond the ganglia. Here the dorsal and ventral roots temporarily fuse.
-
Meningeal branches are given off at this point.
-
Note that the cervical nerves get progressively larger from C1 to C8.
-
-
Spinal roots come next, that is, the dorsal and ventral spinal rami.
-
Dorsal rami travel upward to supply epaxial muscles in medial > lateral order.
-
Ventral rami immediately receive sympathetic fibers because just lateral to each ramus is a vertically oriented chain of sympathetic ganglia. The SANS fibers travel from ganglion to ventral ramus via a grey ramus communicans.
-
In summation, hypoglossal nucleus (r8–r11) in brainstem innervates hypaxial somite-derived muscles. It continues into spinal cord (c1–s4) as medial motor column all the way down the trunk innervating epaxial muscles continuously and hypaxial muscles discontinuously (not c5–t1 and not t12–l5). Nucleus ambiguus (r6–r11) supplies the hypaxial branchiomeric muscles of arches 3–5 (somitomeres 7–11) to palate, pharynx, and larynx in somatotopic fashion. It continues into spinal cord (c1–c6) as central motor column. Lateral motor column appears in the neck for the first time at levels c5–t1.
Clinical Application
Trauma to cervical spinal roots can be caused by herniation of the disc. Nerve roots live immediately next to the disc. This takes place in the root sleeve, a section of the nerve where it is still encased by dura. This condition is called lateral recess syndrome. Just outside the dura in the foramen is the dorsal root ganglion. Nerve roots can also be afflicted by neurofibromas. The symptoms of root compression are dermatome = numbness + paresthesia; myotome = pain.
Here let’s confront (once again) terminology. Nerve roots always exit below their respective vertebra. The reason for this is that the caudal part of the sclerotome is made up of the loose zone. T5 is made from T4C + T5R and the fifth thoracic nerve travels through the permissive territory of T5R. However, cervical vertebrae are numbered differently due to lack of appreciation for the proatlas, Nerve roots are considered to exit above the corresponding cervical vertebrae. For this reason, disc prolapse between vertebrae T4 and T5 will compress spinal root T4 but the same process between C4 and C5 will compress spinal root C5.
Spinal Nerves: Functional Classification
Somatic
-
Somatic efferent = motor control.
-
traditional terminology divides nerves to striated muscles into GSE, general somatic efferent, for garden-variety muscles and SVE, special visceral efferent (sic) for branchiomeric so-called “visceral” muscles.
-
-
Somatic afferent = sensory information from skin, fascia, muscles, and joints.
Visceral
-
Visceral efferent SANS preganglionic sympathetic neurons have their cell bodies in the lateral grey column of the spinal cord from T1 to L3. When fibers exit, they are myelinated and travel into the sympathetic trunk via white rami communicantes. Upon synapsing the ganglia, post-ganglionic neurons travel out to non-striated muscles or glands.
-
Visceral efferent PANS preganglionic neurons have their cell bodies in the lateral grey column of S2 to S4.
-
Visceral afferent neurons have their cell bodies in the spinal ganglia. Their axons run through the white rami communicantes into SANS ganglia (where they do not synapse) and then out to their targets in the viscera.
Meningeal
Branches to meninges are found at each vertebral level. The numbers 2–4 on each side. They all receive SANS input from grey ramus communicans. Within the spinal canal, they track along blood vessels. These mixed/SANS fibers innervate dura, blood vessels, periosteum, and ligaments. Ascending branches from C1 to C3 are distributed to dura mater of posterior cranial fossa. This anatomy if important for understanding occipital headache.
Cervical Nerves and Plexuses
In this section, we shall discuss the general characteristics of the cervical nerves, individual cervical nerves C1, C2, C3, the cervical plexus, phrenic nerve, and the spinal accessory nerve.
Different Rami, Different Roles
Dorsal rami lead to a very monotonous existence. Beginning with C2, they all divide into medial and lateral branches. All of them supply muscles. Skin is innervated by medial branches of the second to the fifth dorsal rami. Dorsal rami are motor for epaxial muscles of the spine supplied from the medial motor column.
Ventral rami are more complex. Not only must they supply the needs of the body axis but they are called upon, at varying anatomic levels, to innervate the appendages. At all levels of the spinal cord, ventral rami are larger than dorsal rami. There are three reasons for this. First, the volume of hypaxial muscle controlling the axial skeleton of the neck and trunk is considerably larger. These muscles have more complex tasks. Not only do they flex and rotate the spine but they control motion (and function) of the entire thoracic cage and abdomen. Second, tetrapods an entirely distinct set of hypaxial muscles are assigned to the appendicular skeleton. Third, mammalin evolution converts a muscle associated with the pre-mammalian brachial plexus into an entirely novel structure, the diaphragm, which they internalize into the chest cavity. It also results in a subdivision of the cucullaris muscle into sternocleidomastoid and trapezius with extensive attachments to a new version of the pectoral girdle.
It is not surprising then that such phylogenetically new muscles should be innervated from separate zones of the spinal cord, the lateral motor column. Thu, s ventral rami carry neurons from 3 distinct motor columns: (1) hypaxial muscles of the axial skeleton from the medial motor column, (2) the muscles of the upper extremity via the lateral motor column, and (3) the diaphragm and cucullaris derivatives via the central motor column. This section deals with only those muscles with primary attachment to either the cervical spine or the pectoral girdle.
With exception of the first ramus, all cervical ventral rami make their debut between the guywires suspending transverse processes of the neck vertebrae, anterior intertransverse, and the posterior intertransverse muscles. Because hypaxial muscles are attached internally to the spine (and skull) and externally to the shoulder girdle, their respective motor nerves must make complex spatial decisions in order to get to their correct destinations. This results in the formation of two plexuses: the upper four cervical nerves are organized into the cervical plexus and the lower four cervical nerves are organized into the brachial plexus. All plexuses are strictly hypaxial; dorsal rami are not involved.
The contribution of the sympathetic autonomic system to the cervical nerves is uniform. They all receive SANS neurons via one or more grey rami communicantes. There are three sources for SANS: superior cervical ganglion > C1–C4; middle cervical ganglion > C5–C6; and inferior (cervico-thoracic) ganglion > C7–C8. Thereafter the ventral rami divide; this will become the basis for understanding the cervical plexus.
Cervical Nerves C4–C8
Dorsal rami
After negotiating their way around the vertebral pillars, these form medial and lateral branches directed to the epaxial muscle of the spine. Medial branches supply (from deep to superficial): interspinous > multifidus > semispinalis cervicis, + semispinalis capitis. Lateral branches (from medial to lateral) longissimus capitis > longissimus cervicis > iliocostalis cervicis. These muscles develop right away and occupy virtually all the dorsal binding sites of the cervical spine. Later on, when hypaxial muscles such as levator scapulae or the rhomboids seek a secondary attachment, they are forced to occupy more peripheral sites. Recall that scapula, a true hypaxial structure, shifts dorsally. Muscles dedicated to the pectoral girdle follow suit. Thus, although the dorsal scapular nerve is ventral, its muscles attach to the dorsal aspect of the vertebral column.
C4–C8 ventral rami all emerge reliably between scalenus anterior and scalenus medius. Ventral neurons arising from medial motor column continue the same pattern to supply the intervertebral muscles along the ventral aspect of the spine. Central motor column (C1–C6) produces phrenic nerve (C3–C5), a unique adaptation in mammals of a muscle formerly belonging to the brachial plexus, subcoracoideus, that migrates into the chest to form the diaphragm. It also produces and spinal accessory nerve. Those arising from the lateral motor column combine to form the brachial plexus (C5–T1).
Medial Motor Column: C1–C8
First Cervical Nerve
C1 is unique in that has a minimal sensory representation. Somite 5 has no dermatome but does provide PAM periosteum to ventral foramen magnum, the so-called and mis-named “meningeal branch” of C1. It has anterior and posterior rami and targets can be organized as follows:
1. Strap muscles (rectus cervicis)
-
Geniohyoid via anterior loop, then via branch to hypoglossal. This indicates that the four-somite model of tongue muscles exists from S2 to S5.
Thyrohyoid via anterior loop, then via branch to hypoglossal
-
Sternohyoid via Ansa
-
Omohyoid via Ansa
2. Muscles from atlas to skull, monosomitic
-
Rectus capitis anterior
-
Rectus capitis lateralis
-
Rectus capitis posterior
3. Muscles from atlas/axis to skull, polysomitic
-
Longus capitis
-
Splenius cervicis
4. Muscle from atlas to scapula, polysomitic
-
Levator scapulae
C1 dorsal ramus (suboccipital nerve) is much larger than its ventral fellow. It makes sense, the axial head extensors are bulky, whereas axial head flexors are ribbon-like. Remember that the ancient proatlas is formed by r11–c1, that is, by somites 4 and 6. C1 should rightfully lie below proatlas, but, alas, proatlas has been absorbed into the skull. Atlas is next-in-line. It is formed from c1 to c2, that is, by somites 6 and 7. Thus dorsal C1 comes out above the arch of atlas. It enters suboccipital triangle to supply five muscles: rectus capitus posterior major and minor, obliquus capitis superior and inferior, and semispinalis capitis.
C1 ventral ramus (suboccipital nerve, SON) emerges above the ring of the atlas. It dives forward, skirting the lateral mass of the atlas, staying medial to vertebral artery. It innervates rectus capitis lateralis and then emerges medial to it. It descends to join with the ascending branch of the second cervical ventral ramus.
Second Cervical Nerve (Greater Occipital Nerve GON)
Arises between atlas and axis. Just the opposite of C1 it has limited motor innervation, (semispinalis capitis) but extensive sensory representation to the back of the scalp, ear, and over parotid gland.
C2 dorsal ramus (greater occipital nerve, GON) is the largest of all the cervical dorsal rami…thus its name. It exits between the posterior arch of atlas and the posterior lamina of axis. This puts it directly below inferior oblique muscle, which it proceeds to innervate and then divides. Medial branch is strongly sensory. It travels through the attachments of semispinalis capitis and trapezius, where it is joined by dorsal C3 and by occipital artery. Next, it sends connecting branches to lesser occipital nerve. It proceeds upward to innervate scalp all the way to the vertex. On occasion, it provides sensation to the back of the ear.
Considerable developmental information is embedded in the dermatomal relationship between the greater occipital nerve versus its sidekick, the lesser occipital nerve. Both supply dermis from the second cervical dermatome. They indicate a distinction between epaxial versus hypaxial scalp skin (Fig. 10.84). Lesser occipital shares a common border with V3 while the greater occipital is co-extent with V1. Although the terms epaxial and hypaxial do not apply to the head, the boundary between these two zones replicates the territory of frontonasal neural crest.
Medial branch is motor to the most rostral of the transversospinal group: semispinalis capitis. Lateral branch is motor to most rostral muscles of erector spinae: splenius cervicis > longissiums capitus > longissiums cervicis.
GON neuralgia can result from (1) entrapment; (2) arthritis of the upper apophysis next to the second cervical root; and herpes indwelling in the geniculate ganglion. This is transmitted to C2 in several ways: (1) via facial nerve directly via the auricular branch of posterior auricular nerve; and (2) posterior auricular nerve makes a second connection to lesser occipital nerve (LON) the superficial branch of ventral C2.
C2 ventral ramus projects outward between atlas and axis. After emerging between longus capitis and levator scapulae, it divides. An ascending branch connects with the descending branch of the first cervical ventral ramus. A descending branch joins third cervical nerve.
Third Cervical Nerve
C3 dorsal ramus exits medial to posterior intertransverse muscle and divides. Medial branch ends up in lower occipital skin. Lateral branch joins with C2 dorsal. Sometimes the dorsal rami of C1 to C3 form loops known as the occipital plexus.
C3 ventral ramus exits between longus capitis and scalenus medius. It sends a medial branch to contribute to the phrenic nerve.
Fourth to Eighth Cervical Nerves
C4–C8 dorsal rami After negotiating their way around the vertebral pillars, these form medial and lateral branches directed to the epaxial muscles of the spine.
-
Medial branches supply (from deep to superficial): interspinous > multifidus, semispinalis cervicis, semispinalis capitis.
-
Lateral branches supply (from medial to lateral); longissimus capitis, longissimus cervicis > iliocostalis cervicis.
All these muscles develop right away and occupy virtually all the available binding sites along the cervical spine. Later, when hypaxial muscles such as levator scapulae or the rhomboids seek a secondary attachment, they are forced to use more peripheral binding sites. Recall that scapula in mammals shifts dorsally, bringing all of its muscles along with it.
C4–C8 ventral rami all emerge reliably between the scalenous anterior and scalenus medius. Ventral neurons arising from medial motor column take two different routes. Some continue the same axial pattern as in C1–C3 to supply intervertebral muscles along the ventral aspect of the spine. Others participate in the cervical plexus to supply the strap muscles and elevators of the scapula. Ventral neurons from central motor column also have two distinct patterns. Phrenic nerve arises from C4 via cervical plexus and dives into the chest. Spinal accessory nerve forms from separate branches of C1–C6 that do not participate with cervical plexus. Ventral neurons from the lateral motor column (C5–T1) form the brachial plexus.
The Cervical Plexus, C1–C4
Let’s begin with a misleading statement. Traditional texts describe the ventral rami of C1–C4 as joining together to make up the cervical plexus. From a functional standpoint the motor components of cervical plexus are really C1–C3. All these muscles come from MMC. C4 is different. Yes, C4 is sensory to the skin of the neck. But in its original design, C4 is also motor to subcoracoideus, a muscle mass connecting pectoral girdle with the upper extremity. In birds, this elevates the wing. In mammals, subcoracoideus becomes the future diaphragm. Thus, from a sensory standpoint, we can talk about cervical plexus as being C1–C4 but from a motor standpoint C4 in mammals is transitioning to the brachial plexus (Fig. 10.78).

Cervical plexus. The plexus is made up of ventral roots from spinal nerves C1–C4. These are medial motor column as they supply muscles to the axial spine. The strap muscles are derived from original rectus cervicis. C1 is strictly motor and supplies geniohyoid, the most distal strap muscle. Phrenic nerve mostly C4 reflects original tetrapod position of muscle from the neck to the pectoral girdle (see discussion on phylogeny of diaphragm). C4 is unique. It supplies the following muscle groups: (1) head-pectoral girdle (spinal accessory), (2) neck-pectoral girdle, and (3) pectoral girdle-humerus. It does not contribute to pectoral girdle-pharyngeal arch control. Spinal accessory nerve, cranial nerve XI innervates two hypaxial branchiomeric muscles sternocleidomastoid and trapezius. It has four–six nuclei in the central motor column, a continuation of nucleus ambiguus. Transition between cervical plexus and brachial plexus takes place at level C4. [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones]
But, we get ahead of ourselves…so let’s proceed with a standard description. The cervical plexus is made up of ventral roots that coalesce to take the shape of an elongated ellipse. It is located at the same level as the first four cervical vertebrae; its surface markings are defined by a line running from the auricle to thyroid cartilage. The plexus is deep to sternocleidomastoid and just anterior to scalenus medius and levator scapulae.
Branches from the cervical plexus have both sensory and motor functions. Sensory branches innervate skin territories of the head, neck, and chest. Motor branches supply: (1) muscles connecting the axial skeleton to the skull, (2) muscles connecting the axial skeleton to itself, (3) muscles connecting pectoral girdle to the axial skeleton, (4) the strap muscles connecting pectoral girdle to pharyngeal arches. The diaphragm, traditionally considered part of the cervical plexus, arises from a separate motor column and merely “goes along for the ride.”
Conceptually, the cervical plexus is quite elegant. As we stated earlier, all the ventral rami divide into ascending and descending branches. The ascending branch of C1 wanders off by itself (it travels forward with hypoglossal to supply geniohyoid) but the remaining roots combine to form a series of loops. The first loop, made from C2 to C3, provides superficial branches in the form of named nerves to the head and face. These are lesser occipital, great auricular, and transverse cutaneous nerves. The second loop, made from C3 to C4, provides sensory supply to the neck and chest as the supraclavicular nerves. Thus, superficial branches are sensory to skin, while deep branches supply muscle. A single medially-directed branch, C4, off the second loop, is motor to the diaphragm. The diaphragm exists only in mammals. From an evolutionary standpoint, it represents the displacement of the subcoracoid muscle originally connecting pectoral girdle to the upper extremitiy. Thus, C4 in pre-mammals is part of brachial plexus but in mammals becomes distinct: phrenic nerve is really the upper-most component of the brachial plexus.
Sensory Branches: Superficial Ascending
Lesser occipital nerve (C2). It curls around the spinal accessory nerve and ascends along the posterior border of the sternocleidomastoid. Here it penetrates upward through the deep investing fascia of SCM and divides. Auricular branch supplies the upper posterior pinna. It connects with great auricular nerve, posterior branch. Occipital branch of LON supplies hypaxial posterior scalp. It interfaces with greater occipital nerve, which supplies epaxial posterior scalp.
Great auricular nerve (C2–C3) This is the largest sensory branch. It wraps around posterior border of SCM and then crosses the muscle deep to platysma. It then proceeds to parotid gland, where it divides. Anterior branch supplies the skin over the parotid gland. Within the substance of parotid, it connects with facial nerve. Posterior branch goes backward over the mastoid process. It supplies the lower posterior pinna to the occipital. It pierces the concha to supply the anterior (external) ear skin of the lobule and concha. It also connects with lesser occipital to the scalp and posterior auricular VII.
Transverse cutaneous nerves (C2–C3) These follow around the posterior border of SCM at its halfway point. Ascending branches innervate submandibular skin and upper neck. Descending branches traverse platysma to supply neck anterolateral neck skin down to sternum.
Sensory Branches: Superficial Descending
Supraclavicular nerves (C3–C4) These nerves all arise from a single trunk constructed from C3 to C4, that is, from the inferior loop. The trunk comes out from behind sternocleidomastoid, these tracks down beneath platysma, and then tricurcates. Medial branches extend to the midline and down to the second rib. They innervate sternoclavicular joint. This is important because we can neuromerically code the joint and the adjacent manubrium as potential derivatives of the c2 lateral plate mesoderm. Intermediate branches cross over the clavicle down to second rib. The dermatome of C3–C4 abuts against that of T2, all intermediate dermatomes (C5–T1) being expropriated away from the trunk by upper extremity. The sensory boundary is very sharp; there is virtually no overlap. Lateral branches flow over trapezius and acromion process. Thus upper shoulder skin has C4 innervation.
Sensory Branches, Deep
Propioceptive branches from C1 to C4 connect with spinal accessory nerve within the substance of sternocleidomastoid and trapezius. Recall that SAN being motor general somatic efferent is exclusively motor. Thus, these communicating branches provide much-needed coordination. They have complex central connections for head-turning, especially the startle response.
Motor Branches to Muscles of the Axial Skeleton
The cervical plexus conveys motor branches from the medial motor column to muscles connecting the axial skeleton with itself. Four axial muscles are supplied by roots that reflect the spatiotemporal order of their development. All have primary insertions from the vertebra. Those that are directed upward are rectus capitis lateralis (C1) and rectus capitus anterior (C1–C2). Those directed downward are longus capitis (C1–C2–C3) > longus colli (C2–C3–C4–C5–C6). Although these muscles are hypaxial they are not related to the upper extremity and have their nuclei in the medial motor column. Motor nerves from C1 to C3 to these muscles are frequently depicted as part of the cervical plexus but do not fit the functional pictures. The cervical plexus is also a switch yard for muscles connecting the trunk to the pharyngeal arches. These are all medial motor column muscles that represent the ancient coracomandibularis in sharks.
Motor Branches to the Hypobranchial Muscles
Hypobranchial muscles come in two flavors. The prehyoid column originates from the occipital somites. These muscles connect the tongue with mandible and hyoid, that is, with the first, second, and third pharyngeal arches. They are innervated by r8–r11 hypoglossal nerve. The posthyoid column originates from the first three cervical somites. These muscles connect the appendicular skeleton (the pectoral girdle) with ventral pharyngeal arches, PA1–PA4. They are supplied by the c1–c3 cervical plexus. The motor components of cervical plexus are as follows.
Communicating branches pass from the C1 to C2 loop. The hypoglossal branch follows along XII and then splits off separately to supply thyrohyoid and geniohyoid. Together these two muscles constitute the superior strap muscles. These unite the ventral midline of pharyngeal arches 1–4. A vagal branch and sympathetic branches are described.
Ansa cervicalis is an inferiorly-directed loop from cervical plexus that supplies the inferior strap muscles, including omohyoid. The purpose of these muscles is to connect the midline pectoral girdle, the manubrium (ancient coracoid), and clavicle with the lower jaw via TH and GH. Ansa has two roots: inferior root (posterior) and superior root (anterior). Inferior root of ansa cervicalis comes from the medial series from a branch of C2 and a branch from C3. It links up with superior root just in front of common carotid. Note the absence of C4 in motor branches. Note: C4 motor fibers are reserved for phrenic nerve and represent the transition to brachial plexus.
Beware the masquerader! Spinal accessory nerve is comprised of strictly segmental roots from C1 to C5 (or C6). They are not supplied by cervical plexus. Sternocleidomastoid and trapezius are developmentally unrelated to the strap muscles and have their motor nuclei in a separate functional column.
Central Motor Column: C1–C5
The rationale for a central motor column in the rostral cervical spinal cord is poorly understood. It supplies two sets of muscles lateral to those of the axial spine, and therefore distinct from MMC. The derivatives of cucullaris (sternocleidomastoid and trapezius) and diaphragm are wildly different in position and function. Each has its own distinct motor column. Spinal accessory column (SAC) spans from c1 to c5 (some consider it to extend as caudal as c7). SAC is positioned at the lateral margin of ventral the horn. Phrenic motor column (PMG) belongs to neuromeres c3–5. It is intermediate in position between MMC and SAC. Yet all have a common denominator, a neuromeric relationship with the pectoral girdle.
These two motor columns obviously overlap. Why is SAC lateral to PMC? This might seem trivial. After all, diaphragm is certainly internal to the scapula. The reason lies deeper (no pun intended). If we look at the organization of the LMC to the upper limb (cf Fig. 10.75) nuclei for the ventral muscles of the limb are in the medial LMC whereas those for dorsal muscles are lateral. We shall see that subcoracoracoideus inserts into the ventral aspect of the pectoral girdle. Trapezius is inserted on the opposite side of scapula. Thus, SMC is lateral to PMC.
We shall take now consider the innervation of the muscles of the central motor column. The nerve to the cucullaris complex, spinal accessory, comes first, as it is the more primitive of the two. Cucullaris is present in extinct placoderm fishes. Subcoracoideus does not appear in evolution until tetrapods. Its final iteration as the true diaphragm exists only in mammals.
Spinal Accessory Nerve (C1–C5)
Nucleus Ambiguus: Neuromeric Organization
Spinal accessory nerve in the spinal cord represents the physical continuation of nucleus ambiguus of the brainstem. This subject is of critical embryological and evolutionary interest because the appearance in mammals of sternocleidomastoid and trapezius, as derivatives of the ancient cucullaris muscle, marks a critical turning point in neck anatomy. Examine the neuroanatomy with care (Figs. 10.79, 10.80, 10.81, 10.82).

Cranial nerve IX. Note single motor neuron from upper nucleus ambiguus directed to Sm7 stylopharyngeus. It has primary insertion into r6–r7 pharynx and secondary (retrograde) insertion into r4–r5 styloid process. Sensory distribution of IX is extensive, covering the entire third arch from the back of the fauces down to thyroid cartilage. Remainder of Sm7 muscles supplied from rostral nucleus ambiguus routing via vagus nerve. [Reprinted from Hanes DE. Fundamental Neuroscience for Basic and Clinical Applications. Philadelphia, PA: Saunders; 2006. With permission from Elsevier]

Cranial nerves X, XI. Nucleus ambiguus is motor to the palate-pharyngo-laryngeal nerve. Vagus nerve from midsection of nucleus ambiguus supplies palatal and pharyngeal muscles, Caudal nucleus ambiguus supplies laryngeal muscles via the vagus. These rootlets have been erroneously termed the cranial root of XI. Spinal accessory nucleus does not join with vagus and does participate in pharyngeal arch innervation. Cranial nerve XI does not exist. CN XI supplies upper third of sternocleidomastoid and descends to the trapezius, indicating the phylogenetic relationships of these muscles to the first five neuromere of the spinal cord (c1–c5). So-called “cranial part” of XI given off by r10–r11 and destined for the pharynx via vaguas: palato-pahaygo-laryngeal nerve. Left: [Reprinted from Hanes DE. Fundamental Neuroscience for Basic and Clinical Applications. Philadelphia, PA: Saunders; 2006. With permission from Elsevier.] Right: [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones]

Spinal accessory nerve in situ dissection. Left Rootlets from nucleus ambiguus indicated by * and # crossing the posterior fossa to join with vagal bundle (VB) to form common palate-pharyngo-laryngeal motor nerve. Brainstem rotated, showing olivary eminence. SAN is clearly distinct. Cranial nerve VIII, IX, and XI labeled. VB = vagal bundle. Right: inferior cerebellar peduncle sectioned, jugular foramen opened, showing SAN heading caudally toward sternocleidomastoid and trapezius. SAN remains distinct from X without contributions from caudal medullary rootlets. Hypoglossal nerve XII (unlabeled) seen ventral to the rootlets, showing four roots converging to enter hypoglossal foramen (unlabeled). [Reprinted from Wiles CCR, Wrigley B, Greene JRT. Re-examination of the medullary rootlets of the accessory and vagus nerves. Clini Anat 2007; 20(1):19–22. With permission from John Wiley & Sons]

Spinal accessory nerve showing rootlets from caudal nucleus ambiguus apparently joining with spinal accessory nerve (but not sharing fascia) and eventually joining with vagus. Recall that the pleuro-peritoneal folds are associated with the pronephros sector of nephric ridge which begins at level c1 in the neck. Note that anterior scalene intervene between right phrenic n. and second part of subclavian whereas left phrenic n. descends more medially, directly over first part of subclavian. Phrenic nerve supplies diagphragm via three branches: sternal, anterolateral and posterolateral. These corresponds to congenital defects of Morgagni, eventration, and Bochdalek. Note: paired pericardiophrenic nerves penetrate to inferior surface of diaphragm; they receive pain signals from the underlying visera: right pericardiophrenic (liver), left pericardiophrenic (left lobe of liver, stomach, spleen and left adrenal). Phrenic nerve is NOT a part of cervical plexus and NOT a part of brachial plexus, as currently defined but phylogenetically it belongs to the brachial plexus. Its insertion pattern is antegrade-antegrade [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Nucleus ambiguus is a collection of motor neurons located in r6–r11 of the hindbrain that supply so-called “branchiomotor” muscles of the third, fourth, and fifth pharyngeal arches. The traditional name for this functional column is special visceral efferent (SVE), a term that is completely wrong on two out of three counts. There is nothing special at all about muscles that originate from somitomeres 7–11. They are striated, voluntary, with myoblasts of paraxial mesoderm origin and connective tissue stroma made from neural crest. Furthermore, it perpetuates the long-debunked proposition that, since pharyngeal arches are in continuity with the GI tract, their musculature should be “visceral” or smooth but somehow isn’t. Nucleus ambiguus is lateral in the brainstem. Medial to it lies dorsal nucleus of vagus r6–r11which PANS motor to the entire presacral GI tract and represents the so-called general visceral efferent (GVE) column supplying the smooth muscle and glands of the gut. Most medial is hypoglossal nucleus r8–r11 which is generally somatic efferent to the striated tongue muscles.
The organization of NA is somatotopic. It has three neuromeric sections, but these are represented by only two cranial nerves. Rhombomeres r6–r7 are motor for the muscles of somitomere 7: stylopharyngeus, soft palate muscles, and superior constrictor and middle. These neurons can travel via one of two routes to their targets. The nuclei of r6–r7 supply two types of motor nerves. A single branch to the stylopharyngeus passes out via glossopharyngeal nerve, as it travels directly to the posterior pharynx.
All remaining muscles from Sm7 are innervated by nerves from r6 to r7 that join with those of r8–r11 to form via vagus nerve. Rhombomeres 8–11 are motor for middle and inferior pharyngeal constrictors via pharyngeal plexus, for the fourth arch muscles of the larynx via r8–r9 superior laryngeal nerve, and for the muscles of the fifth arch via r10–r11 inferior laryngeal nerve. We can rightly term this motor nerve of X as the palato-pharyngo-laryngeal nerve (PPL).
Herein let’s debunk an anatomic anachronism. The most caudal two rhombomeres r10–r11 have been traditionally (and mistakenly) termed a separate structure, the cranial root of spinal accessory nerve. In reality, these two roots simply complete the PPL. Understanding the motor supply to arches 3–5 is much simpler and neuroanatomically accurate if we use the neuromeric model.
Cranial Nerve XI, Does It Exist?
Spinal accessory has traditionally been described as having two anatomically separate components. This concept has been disproven [15, 16].
Cranial part (ramus internus) has 4–5 rootlets arising in series from nucleus ambiguus, representing levels r10–r11. These are slightly displaced away from those of r6 to r9. As these rootlets approach and enter the jugular foramen, they join together just superior to the inferior (nodose) ganglion to form common vagus, that is, palato-pharyngo-laryngeal nerve (PPLN). The rootlets of PPLN share a combined sheath of arachnoid. PPLN in jugular foramen remains completely separate from cranial nerve IX. Glossopharyngeal nerve comes exclusively from levels r6–r7 and is sensory for nocioception and taste and PANS motor to salivary glands; it has but a single somatic motor branch to stylopharyngeus. Within jugular foramen PPLN and glossopharyngeal nerve IX remain separate: IX has its own arachoid sheath. Spinal accessory nerve also has its own sheath of arachnoid. The periosteum of the posterior fossa is falsely labeled dura—see Chap. 12. Intracranial periosteum is continuous with extracranial periosteum so nerves IX, X (its caudal roots being the imposter XI) are all covered with “dura.”
Upon exit from jugular foramen IX and PPLN immediately diverge. So too does SAN. It takes off posteriorly and laterally in the general vicinity of internal jugular vein. After crossing transverse process of C1, it is crossed by occipital artery. Spinal XI follows the superior sternocleidomastoid branch of occipital artery and enters the muscle. Within SCM, spinal root has various options. It can anastomose with the ansa of Maubrac, that is, C2–C3 fibers from ansa cervicalis. It can also join with the McKenzie branch, that is, the anterior root of C1. XI usually emerges from SCM at its midpoint, called Erb’s point. This generally occurs above (or within 2 cm) of the exit point of greater auricular nerve (C2–C3).
Spinal part (ramus externus) consists of 6 roots in series that coalesce to form a single peripheral nerve. Because the first root in evolution belongs to first truncal somite and because the latter is absorbed into the skull, SAN becomes entrapped by the jugular foramen. Its gross anatomy was difficult to discern in the nineteenth century and it was considered a part of what was thought to be a separate nerve from the rootlets of the posterior medulla: thus the legend of the XI was born and has persisted by default until recent neurosurgical dissections and dye studies have clarified the situation.
In sum: sternocleidomastoid and trapezius are the inheritors of cucullaris. Their myology, long misrepresented for lack of fossil evidence, has come to light revealing these muscles as likely branchiomeric but with a unique history connected with the evolution of the tetrapod neck which we shall examine in greater detail, vide infra.
Phrenic Nerve (C3–C4–C5)
Diaphram is unique to mammals and, from an evolutionary perspective, is a derivative of the brachial plexus, vide infra. Its sole source of supply is phrenic nerve in which C4 is the predominant root. C4 constitute an evolutionary anatomic break-point. Recall our previous discussion of the caudal translocation of the brachial plexus that takes place in mammals. It is found along the lateral border of scalenus anterior (the most midline of the scalenii). Of course, we know phrenic nerve must follow an internal course. In order to do this, phrenic nerve must cut across planes. It penetrates the prevertebral fascia surrounding scalenus anterior and then proceeds to track across the muscle underneath its fascia. Muscle and its fascia are coded by c3–c6 so it makes sense that c4 nerve to diaphragm follows along this plane. Its downward course takes it beneath omohyoid and between subclavian artery (behind) and vein (in front). While traveling through the neck, phrenic nerve receives SANS fibers from the cervical sympathetic ganglia. As it enters the thorax it crosses in front of internal thoracic artery (Figs. 10.83, 10.84).

Phrenic nerve C3–C5 Its anatomic course recapitulates entry of subscapular anlage into thorax above first rib, that is, at interface of c8–t1. Recall thath the pleuro-peritoneal folds are associated with the pronephros sector of nephric ridge which begins at level c1 in the neck. Left: [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones] Right: [Reprinted from Standring S. In: Gray’s Anatomy, 40th edition. Philadelphia, PA: Churchill-Livingstone; 2008. With permission from Elsevier]

Left: Cervical plexus with motor nerves (green) and sensory nerves (orange) demonstrates phrenic nerve arising directly from roots C3 and C5 with a contribution from C5. Phrenic nerve is NOT a part of cervical plexus and NOT a part of brachial plexus, as currently defined but phylogenetically it belongs to the brachial plexus. Its insertion pattern is antegrade-antegrade. Right: Brachial plexus. Standard description of the mammalian brachial plexus fails to recognize the diaphragm as essentially a brachial plexus muscle, with roots C4 and C5 innervating supracoradoideus, the precursor of diaphragm. Note the evolution of more complex motor control of the upper extremitiy results in an additional third motor branch, the median nerve. Thus median remain in the original position of radial while radial shifts backward two neuromeres to receive roots C6–C7 with a cross-branch to the union of C8–T1. Note the critical transitional nature of C4. Left: [Courtesy of Michael Carstens, MD] Right: [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
Differences exist between the two phrenic nerves regarding their course within the thorax but these are not germane for our purposes. Suffice it to say that the phrenic nerves provide sensory input to the mediastinal pleura, fibrous pericardium, and the parietal (external) pericardium. Clinical correlation: The communality of C4 from the heart constitutes an important source of referred pain to the neck…a tightening of the throat.
At the central tendon of diaphragm, phrenic nerve sends branches to parietal pleura above and parietal peritoneum below. It then trifurcates. Anterior branch runs forward to the sternum. Anterolateral branch supplies the lateral leaf of the central tendon. Posterior branch supplies the crura. Below the diaphragm, branches of the phrenic nerve make clinically important connections with the celiac plexus. Sources of referring fibers are the liver, gall bladder, and the adrenals. Shoulder pain may thus reflect pathology in the thorax, hepatobiliary system, or kidneys.
Trauma to the phrenic nerve can occur in the neck at scalenus, beneath the clavicle, and from injuries to the upper brachial plexus.
Lateral Motor Column: C4–T1
Although we have foresworn venturing too deeply into the mysteries of the brachial plexus, it is very useful to dissect it apart from a functional and neuroanatomic standpoint. Although all of its muscles come from cervical somites (we must include somite 13, T1 for the sake of completeness, it is useful to consider those muscles as having a relationship with the pectoral girdle. If we do so, we can gain useful insights into the possible rationale behind the layout of brachial plexus. So be patient. We shall get into the myology again in the next section (Fig. 10.85).

Brachial plexus and phrenic nerve C5 sends branch to C4 phrenic nerve prior to participating with any other roots, that is, prior to entry into the plexus. All motor branches of brachial plexus supply cervical somite muscles antegrade–antegrade insertion sequence into pectoral girdle and distal neuromeres of the thorax and humerus. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Why Is a New Motor Column Needed in the Lowevr Neck
The neck is a mixture of the old and the new. It represents the forward continuation of purely axial trunk musculature (both epaxial and hypaxial) represented by the medial motor columns. On the other hand, the cervical region gives rise to an evolutionary innovation, the limb system. We see this transition for the first time in sarcopterygian fishes as muscles to control the fins migrate out from the body into the fin itself thus causing a bulkiness from which their name is derived. Muscles controlling the limb are, of course, ventral. But, by virtue of being in a new anatomic environment, one associated with the lateral plate mesoderm of the limb itself, the neuromuscular system makes a binary adjustment. (1) Muscles that continue to connect the pectoral girdle back to the head body at neuromeric levels c4 or more proximal than their insertion represent a reassignment of previous piscine muscles. These retain an innervation pattern based on the original motor columns, medial motor column, or as a caudal extension of nucleus ambiguus, central motor column. (2) Muscles that attach the pectoral girdle to the body wall distal to c4 or to upper extremity represent an innovation. These muscles cannot be supplied by the original primary ventral roots; their motor nuclei become spatially distinct within the spinal cord as lateral motor column (cf Fig. 10.73).
In tetrapods, the situation becomes more complex. New genes are involved. During development, the limbs, to which these muscles attach, undergo predictable patterns of motion. In order to find their proper insertions myoblast groups must traverse complex pathways. Remember that the bones of the shoulder girdle and upper extremity possess many binding sites for muscles. These will fill up in an orderly manner, using three gradients: proximal–distal, posterior–anterior, and ulnar–radial. A new muscle seeking an available binding site will migrate in such a way as to respect the positional priority of their predecessors. This means that motor neurons will follow along previous pathways through the plexus until arriving at an appropriate exit point. Plexuses thus take on the appearance of busy switchyards. But the apparent anatomic complexity of a plexus belies the fact that it is simply a compilation of all migration pathways. The developmental sequence of limb muscles is faithfully reproduced by its motor plexus.
Ventral rami of C5–C8 join up with the ventral ramus of T1 to form brachial plexus. It is well described in Gray’s anatomy so we shall be very selective here. Our emphasis is the neck and its connections, we will draw the line in terms of neuromuscular commentary with the pectoral girdle and its connections. This makes developmental sense. The clavicle arises from LPM of c1–c6 and scapula from c3 to c6, and it functions as a critical waystation between the trunk and the upper limb. It is a moveable platform containing muscles connecting two types of muscles: those connecting scapula with the axial skeleton and those connecting it with the humerus.
Secrets of Insertion
Pectoral girdle muscles with an antegrade/retrograde insertion sequence seek a secondary attachment at neuromeric sites proximal to the primary insertion. The myoblasts first gather around the pectoral anlage at the base of the limb bud, attach to their assigned sites, and then migrate backward to the midline to form secondary insertions. The secondary insertion sites follow a cranial to caudal and medial to lateral progression as the muscles seek out a(n) available binding site(s). All the most desirable parking spaces have been previously taken by the axial muscles. This pattern applies to the muscles supplied by the cervical plexus. These have secondary insertions into the skull and cervical spine.
Pectoral girdle muscles with an antegrade/antegrade insertion sequence have secondary attachments at neuromeric sites equal to, or distal to, the primary insertion sites. Fortunately, the upper limb follows the same pattern, with secondary binding sites on the humerus opening up in the same sequence, so that subsequent muscles insert further down on the shaft.
Functional Dissection of the Brachial Plexus to the Pectoral Girdle
Branches of the brachial plexus supply two classes of muscles, based on the insertion patterns previously described. We shall further group the targets of these nerves by insertion site. Muscles inserting into primary insertion into clavicle are indicated in red; those inserting into scapula are indicated in blue. A single muscle, deltoid, inserts into both clavicle and scapula.
-
(1)
Antegrade/Retrograde Insertion
-
Muscles that attach the clavicle (including manubrium) to the pharyngeal arches.
-
sternothyroid, sternohyoid, and omohyoid (C1–C3) = cervical plexus.
-
-
Muscles that attach clavicle and scapula to the skull/vertebral column.
-
Sternocleidomastoid and trapezius (C1–C6) = spinal accessory nerve.
-
levator scapulae (C3–C4) = cervical plexus (plus C5 branch).
-
-
-
(2)
Antegrade/Antegrade Insertions
-
Muscles that attach the clavicle and scapula to the chest wall.
-
subclavius (C5–C6),
-
rhomboid major and rhomboid minor (C5),
-
serratus anterior (C5–C7),
-
pectoralis major (C5–T1) and pectoralis minor (C8–T1).
-
-
Muscles that attach scapula to humerus, acting on the shoulder.
-
deltoid (C5–C6),
-
subscapularis (C5–C6),
-
supraspinatus (C5) and infraspinatus (C5–C6),
-
teres major (C5–C6) and teres minor (C5).
-
-
Muscles that attach scapula to humerus acting on the arm:
-
coracobrachialis (C6–C7),
-
short head of biceps (C5–C6) and long head of biceps (C5–C6),
-
long head of triceps (C7–C8).
-
-
Motor Nerves of Brachial Plexus to Pectoral Girdle
Roots
-
Communicating to phrenic (C5).
-
Nerve to scaleni (C5–C8) has three parts. Scalenus anterior (C4–C6) is synthesized first. Scalenus medius (C3–C8) picks up three additional somites. Its mesenchyme is co-extensive and caudal to that of sternocleidomastoid (C1–C2). Scalenus posterior (C7–C8) picks up mesenchyme from the lower two roots. Its secondary insertion is into second rib because there is no further additional room. It lies deep to serratus, which forms later.
-
Nerve to longus colli (C5–C8) also has three parts, each with a different primary insertion. Superior oblique LC (C2) is the smallest and is located at the atlas (a combination of S5C and S6R). Middle (vertical) LC (C2–C4) is the largest. Inferior oblique LC (C5–C6) completes the sequence.
-
Dorsal scapular nerve (C3–C5) has a very misleading name. Its primary contributor is the ventral root of C5 (with contributions from C3 and C4) and arises at the intervertebral foramen. It then penetrates scalenus medius to achieve a dorsal position. It then runs along the deep surface of levator scapulae which it supplies until reaching the dorsal border of scapula. From here it innervates rhomboid major and rhomboid minor. Because all three muscles are specific to the pectoral girdle, their nuclei belong to lateral motor column.
-
Long thoracic nerve (C5–C7) This counterpart to dorsal scapular nerve supplies serratus anterior. It arises at the root level of the brachial plexus. Its nuclei are somatotopic and in line with dorsal scapular nerve. Interesting parallels exist between long thoracic and dorsal scapular nerve. Their nuclei with lateral motor column are continuous. DSN is parallel with phrenic nerve of the central motor column. All four muscles belonged at one point in evolution to a common blastema, the ancient subscapularis or subcoracoscapular. Its fragmentation created in mammals the two functional sets of muscles: those connecting pectoral girdle to the spine (levator scapulae and rhomboids) and those connecting it to the chest wall (subclavius, serratus anterior, pectoralis major and minor) plus the muscular diaphragm.
Trunks
-
Dorsal scapular nerve (C3–C5) This nerve has a very misleading name. Its primary derivation is C5 with some contributions from C3 and C4. It arises at the intervertebral foramen. It then penetrates scalenus medius to achieve a dorsal position. DSN subsequently runs along the deep surface of levator scapulae (which it supplies) until reaching dorsal border of scapula. From here, it innervates rhomboid major and rhomboid minor. As it emanates from C3 to C5 nerve roots, dorsal scapular is sometimes not considered part picks of brachial plexus because of a mistaken model that the plexus has five roots. This mistake is corrected when we consider the phylogeny of the brachial plexus. In evolution, motor control of the upper extremity can begin as far forward as the second spinal nerve in anurans (frogs) but this is a backward step in evolution. The basal amniote plexus begins with C4 and has four roots in the context of 5 cervical vertebrae with a formula C4–C5–C6–T1. Mammals duplicate two neuromeres C4–C5 with a formula of C4–C5–C6–C7–C8–T1 to create a brachial plexus of six roots and seven cervical vertebrae. Dorsal scapular is most certainly a full-fledged member of the plexus.
-
Nerve to subclavius (C5) and accessory phrenic nerve (C5) Both these nerves are mononeuromeric. The latter can originate from subclavius n. or as a distinct nerve. Despite the difference in their names, both relate to clavicle. Recall that diaphragm is a derivative of the supracoracoideus which connects sternum to the ancient anterior coracoid in premammals. We distinguish this from the posterior coracoid which appears for the first time in the synapsid line leading to mammals. The former becomes manubrium and the latter the final coracoid process of the scapula. This neurology supports the origin of diaphragm as a re-directed brachial plexus muscle with C4 arising at the level of the cervical nerve and the C5 fibers traveling outward more laterally, being admixed with those supplying subclavius.
-
Suprascapular nerve (C5–C6) is more lateral on the trunk.
Cords
-
Lateral pectoral nerve (C5–C7) and medial pectoral nerve (C8–T1) arise from the lateral cords and medial cords, respectively. Pectoralis major receives both nerves while pectoralis minor is supplied by just the medial nerve.
-
Upper subscapular nerve (C5–C6) is proximal, as it supplies upper subscapularis.
-
Middle subscapular nerve, or thoracodorsal nerve (C6–C8) follow subscapular artery, the first branch of the third zone of axillary artery which is immediately proximal to the humerus, and descends with it for latissumus.
-
Lower subscapular nerve (C5–C6) is distal, as it supplies lower subscapularis and teres major.
In Summation
Cervical plexus innervates muscles attaching pectoral girdle to the skull or cervical spine at proximal neuromeres: these are all antegrade-retrograde.
Brachial plexus innervates muscles attaching pectoral girdle to thoracic spins, thorax, or humerus at distal neuromeres: these are all antegrade–antegrade:
-
only the upper trunk contributes to the pectoral girdle, and,
-
no posterior division of any of the three trunks supplies the pectoral girdle.
Phyologeny of Motor Neuron to the Vertebrate Pectoral Girdle: A Preview
In the history of evolution, the fins-to-limb transition had neurologic implications. A formerly simplistic system of flexor/extensor muscles inside the body proceeding past a fixed pectoral girdle welded to the head cavity and connected to the fin undergoes a series of drastic changes. The pectoral girdle disconnects and becomes mobile, requiring motor control. New muscles extend outward into the extremity requiring new muscles supplied from an entirely different motor column. In this process, motor neurons actually migrate caudally out of the hindbrain and into the spinal cord due to a frameshift of Hox expression along the rostrocaudal axis. This neuromeric shift is crucial to the formation of the neck. We shall look at this process in greater detail at the conclusion of the myology section, once we have discussed the developmental anatomy of cucullaris muscle and its derivatives (sternocleidomastoid and trapezius) as well as that of the uniquely mammalian innovation, the diaphragm.
Myology of the Neck
How Cervical Muscles Originate and Insert
Cervical muscles originate from paraxial mesoderm in the myotomes of somites. The sole exception to this rule is the possibility of lateral plate mesoderm contributing to the muscles of cucllaris, sternocleidomastoid and trapezius. This difference is academic as the connective tissue stroma within these muscles arises from the neural crest of cervical neuromeres c1–c6. Using our neuroanatomic model, we can organize the muscles of the neck as follows.
-
Medial motor column supplies vertebral muscles.
-
The anterior flexors are uniformly polyneuromeric and are innervated by ventral branches of segmental nerves C1–C6.
-
The posterior extensors MMC are both mononeuromeric and polyneuromeric. They are innervated by dorsal branches of segmental nerves C1–C8.
-
Cervical plexus (C1–C3) supplies hypobranchial muscles (cervical somites 1–3). These represent the ancient coracomandibularis which connects sternum with pharyngeal arches innervated by cervical plexus.
-
-
Lateral motor column supplies type three types of hypaxial muscles.
-
Segmental nerves (C1–C8)—not via a plexus—supply lateral vertebral muscles arise from cervical somites 3–8, attach to ribs 1–2 and receive motor innervations from. All muscles control the upper limb girdle.
-
Cervical plexus (C1–C3) supplies hypobranchial muscles (cervical somites 1–3 and represent the ancient coracomandibularis which connects sternum with pharyngeal arches innervated by cervical plexus.
-
Brachial plexus (C4–T1) supplies the muscles of pectoral girdle and upper extremity.
-
-
Central motor column muscles are unique.
-
Sternocleidomastoid (C1–C2), and trapezius (C2–C6) represent an evolutionary transition between the skull and pectoral girdle. They are supplied by a polyneuromeric segmental nerve, spinal accessory.
-
Diaphragm cervical C3–C5), found only in mammals, represents a pectoral girdle muscle internalized into the chest. It marks the most cranial boundary of brachial plexus.
-
The piscine forefin is positioned at the junction of head and trunk. Like all limbs, it is strictly hypaxial. It consists of a dermal pectoral girdle (including clavicle) and a cartilage-replacement fin (the proximal bone of which is scapula). Over the course of evolution scapula assumes an epaxial position but, because it is inseparable from upper extremity, its neuromuscular apparatus retains the original hypaxial identity. Leaving aside the XI-innervated sternocleidomastoid and trapezius, all other scapular muscles behave like limb muscles. They are innervated by ventral motor nerves acting through a plexus. Cervical plexus and brachial plexus are co-extent with C4 being the transition point.
As we proceed forward with our analysis of muscle groups, a great deal of attention will be placed on where each muscle attaches and in what order these attachments take place. You will note that the words “origin” and “insertion” are deliberately not used in these contexts. All striated muscles from level r8 and more caudal originate from the myotomes of somites. The neuromeric level(s) of origin for any muscle is identical to that of its motor nerve. Pectoralis minor is innervated by medial pectoral nerve (C8–T1). Its myoblasts therefore arise from S12–13, that is, the eighth cervical and first thoracic somites. Muscle attachments proceed along three gradients: cranial-caudal, deep-superficial, midline-lateral. Mononeuromeric muscles have priority over polyneuromeric muscles.
Let’s see how this process works for the muscles of the anterior midline of the neck. Rectus capitis anterior minor (c1) has midline priority with primary insertion at mass of atlas and the medial transverse process. It gains a privileged secondary insertion site into posterior basioccipital just in front of foramen magnum. Due to its c1 predecessor, rectus capitis lateralis (c1–c2) is forced to take a primary insertion on lateral transverse process of atlas and must also seek out a more lateral secondary insertion site on jugular process of exoccipital bone. Longus capitis (c1–c3), otherwise known as rectus capitis anterior major, must take a more anterior primary insertion into basioccipital just behind basisphenoid.
Cervical Muscle Migration: Axial Muscles Versus Appendicular Muscles
Cervical muscles interconnecting the axial skeleton have a simple (somitic) embryonic origin. They can be dorsal or ventral and are always supplied by individual roots, not by a plexus. Their primary insertions are always local and follow the neuromeric and spatial constraints as previously described. Secondary insertions can be proximal or distal depending upon site availability. Muscles from the atlas are bidirectional, spanning upward to the skull and downward as erector spinae.
Cervical muscles associated with the pectoral girdle develop by two mechanisms described by Huang. Myoblasts move into their corresponding neuromeric zone around the lateral plate mesoderm of the clavicle and scapula. After developing their primary insertions, they follow two options: (1) The muscles translate backward to available sites on axial skeleton (skull, ligamentum nuchae, vertebral spines). (2) The muscles translate forward from pectoral girdle to humerus. The former process is called antegrade–retrograde migration while the latter process is referred to as antegrade–antegrade migration. Once again, primary insertion always takes place within the neuromeric territory of origin; secondary insertion is opportunistic.
Consider these examples. Rhomboid minor (C4–C5) is a somitic muscle innervated by dorsal scapular nerve. It first attaches to the c4–c5 zone of scapula just at medial border of the spine. Subsequently, it attaches to the spinous processes of the seventh cervical and first thoracic vertebrae. Rhomboid major (C4–C5) must attach to medial scapula distal to rhomboid minor; thus, it inserts from below the medial spine all the way down to the inferior angle of scapula. Its secondary attachments are also in sequence with the spinous processes of second–fifth thoracic vertebrae. Sternocleidomastoid myoblasts descend from S1 to S2 to posterior margin of manubrium and medial clavicle, make their attachments and return to the r6–r7 mastoid process and r8–r11 superior nuchal line.
Principles of Motor Column Analysis
With these principles in mind, let’s examine the muscle groups following the same developmental schema. The epaxial muscle group is more simplistic. The suboccipital muscles and the deep back muscles are anatomically similar and form a single functional unit supplied by dorsal spinal nerves. We will then turn our attention to the hypaxial muscle group. Anterior vertebral and lateral vertebral muscles interconnect head, neck, and trunk by means of ventral spinal nerves. The more superficial infrahyoid (sic) muscles connect the pharyngeal arch system with the trunk by means of the cervical plexus. Next, we shall examine muscles of occipitocervical origin that connect upper limb to vertebral column and muscles of cervical origin that connect upper limb to thoracic wall. Finally, we conclude our survey of the cervical musculature with a discussion of muscles of having extraordinary evolutionary significance: (1) a forgotten member of the brachaial plexus, the diaphragm; and (2) the cucullaris muscle and its mammalian derivatives, sternocleidomastoid, and trapezius, including implications for the evolution of the neck.
Cervical somites differ from occipital somites in two fundamental ways. First off, they have a full complement of subunits. Recall those occipital somites have sclerotomes; their myotomes are hypaxial only, and their dermatomes are involute. Somites mature in a cranial-caudal progression. Somite 5 (C1) has an epaxial myotome, but no dermatome. Somite 6 has a fully functional dermatome supplying both hypaxial neck skin and epaxial scalp.
With the exception of esophagus, all neck muscles arise from PAM and are somitic. Esophageal muscle originates in lateral plate mesoderm. Motor columns differ by dorsal/ventral and by function: the role of pharyngeal arch muscles as “the outsider” is assumed by muscles assigned to the pectoral girdle and upper extremity.
-
MMC rostral, c1–c4: occipital cervical and upper paraspinous muscles (S5–S8).
-
MMC caudal, c5–c8: lower paraspinous muscles (S9–S12).
-
LMC rostral, c1–c4: cervical plexus muscles (S5–S7) > pectoral girdle.
-
LMC caudal, c5–t1: brachial plexus muscles. (S8–S13) > upper extremity. We should include c4/S8 in this category as well for its contribution to diaphragm, a surprising original member of the “brachial plexus breakfast club.”
-
CMC, c1–c6: diaphragm (S9) from phrenic motor column (PMC) and sternocleidomastoid/trapezius (S5–S9) from spinal accessory motor column (SAC).
Phylogeny of the Pectoral Girdle: A Quick Review
In the ensuing section, we shall be discussing the individual muscles of the neck from a neuromeric, neuroanatomic, and functional consideration. Because many of these muscles insert into the pectoral girdle, its evolutionary components need to be kept in mind. Although this material was presented under the section of osteology, a summary is worthwhile.
The earliest gnathostomes were armored fishes. The most basal forms, such as the osteostracans, had no jaws but did have paired pectoral fins with a shoulder girdle fused to the skull at the level of the heart. Jaws appear in the more advanced placoderms, so-called for the ring of armor surrounding the anterior trunk. These latter split into the defunct †acanthodians and the ray-finned actinopterygians, an offshoot from which became our distant ancestors, the lobe-finned sarcopterygians.
In one group of placoderms, the †arthrodires (ancestral to tetrapods), separation of the head and body is first seen using a pivot joint that permitted elevation of the head under muscle control involving the prototype of the scapula. The prototype of the pectoral girdle is desmontrated in the arthrodrthrodire †Dunkleosteus. It consisted of five dermal bones suspended from the skull by post-temporal bone and followed in succession by supracleithrum, postcleithrum, cleithrum, and the most ventral, clavicle. Immediately behind this bony suspension bridge was a mesodermal scapulocoracoid cartilage articulated between cleithrum and the forefin. Control of the arthrodire pivot joint involved two anatogonistic muscles: the dorsal levator capitis, attached to the thoracic armor and the ventrolateral cucullaris, attached to scapula. This initial genetic experiment (a homeotic switch) was abandoned in the subsequent evolution of fishes into the chondrichthyan line and osteichthyans.
In basal actinopterygians s split occurred in which scapulocoracoid split into two separate bones, scapula and coracoid. These were dorsal to clavicle. Although pretetrapod sarcoptyergians such as the osteolepiform †Eusthenopteron retained the scapulocoracoid complex, the basics of a limb are now present.
The subsequent development of the pectoral girdle in tetrapods is one in which its the dermal elements are reduced or lost and mesoderm elements become dominant. Loss of post-temporal was the fundamental step required in separation of the pectoral girdle from the head. In the early tetrapod †Europs. only cleithrum and clavicle remain (the interclavicle will be lost in mammals). A hollowing-out of scapulocoracoid creates a glenoid fossa, fit for receiving a weight-bearing extremity.
Amniotes are divided into the diapsids and synapsids; in the latter scapulocoracoid once again fragments, now into three pieces: scapula, the (original) anterior coracoid, and a new element, the posterior coracoid. Mammals position the limbs beneath the body with elbows directed posteriorly and knees anteriorly, producing a longer stride and more efficient gait. Motor needs drove change. Massive ventral muscles needed to pull the body off the ground were not needed. The shoulders being mobile versus the pelvis required expansion of dorsal muscles for stability. In some important cases, ventral muscles shifted dorsally, without a change in their neural control. This explains the innervation of trapezius, levator scapulae, and the rhomboids. Consequently, in therapsids the coracoids got smaller. Both are retained monotremes (the all-purpose iconoclast, the platypus) but the anterior coracoid is lost (but not really) in therians, and subsequently in their descendants, the mammals. “Loss” of anterior coracoid permitted the glenoid fossa to redirect ventrally atop the humerus. Posterior coracoid remains adjacent to glenoid fossa as the coracoid process. Cleithrum becomes repositioned over scapula as the scapular spine. Its lateral margin becomes acromion.
Interclavicle persists into synapsids as far as the monotremes but is lost in therians (mammals). Some running mammals lose the clavicle but it is retained in most as a shoulder brace. Recent evidence suggests that anterior coracoid is not really lost, but rather is reincarnated as manubrium.
Epaxial Muscles Interconnecting the Skull and Cervical Spine (MMC)
Suboccipital Muscles
These muscles connect the upper three cervical vertebrae to the skull. All four muscles in this group arise from the first cervical somite (somite 5). Depending upon their relative biologic maturity, they choose different primary and secondary sites of attachment as these become available. Taken together, they comprise an equilateral suboccipital triangle. Its base is transverse, in parallel with the posterior arch of the axis and is defined by obliquus capitis inferior. The apex of the triangle points upward toward greater occipital nerve. Its medial limb is rectus capitis posterior major. The medial border of its lateral limb, obliquus capitis superior, overlaps RCP major, and leads directly to greater occipital nerve. This implies that REC major is laid down first, followed by OCS. Note that three out of four of these muscles form a secondary attachment to the spinous process of the axis. Let’s discuss them in their developmental sequence: rectus capitus minor > rectus capitis major > obliquus capitis superior > obliquus capitis inferior (Fig. 10.86).

Dorsal rami of C1–C3 C1 is strictly motor. C2 has extensive sensory distribution up the lateral aspect of the occiput while C3 gives sensory innervation to the medial zone. C3, C4, and to a lesser extent C5 supply diaphragm. [Reprinted from Standring S. In: Gray’s Anatomy, 40th edition. Philadelphia, PA: Churchill-Livingstone; 2008. With permission from Elsevier]
The suboccipital muscles have distal attachments to occipital bone at the r11–c1 zone and higher up to r8 (superior nuchal line). Their proximal attachments are to the c1–c2 neural arch of the atlas; and to the c2–c3 zone, that is, the axis.
-
Rectus capitis posterior minor This muscle is more midline than RCPmaj so it migrates earlier. Its primary attachment is the tubercle of posterior atlas arch; its secondary attachment is medial inferior nuchal line.
-
Rectus capitis posterior major This muscle develops later than its minor colleague RCPmin so it assumes a more lateral course. Its primary attachment is the spinous process of axis and it secondarily attaches to lateral inferior nuchal line of chondral supraoccipital bone. It is crossed superficially by two nerves, first, the posterior division of the first cervical nerve and, superficial to that, the greater occipital nerve.
-
Obliquus capitis superior OCS can be seen overlapping RCPmaj so we know it is laid down last. Its primary attachment is lateral mass of atlas. Its secondary attachment is occipital bone; it lies lateral to RCPmaj to fill out the lateral end of the fossa between superior and inferior nuchal lines.
-
Obliquus capitis inferior Larger than its colleague, the primary attachment of OCI is apex of spinous process of axis. Its secondary attachment is to the dorsal part of transverse process of atlas.
Epaxial Muscles Interconnecting the Cervical Spine and Thorax (MMC)
These muscles exist in three layers. These are, from deep-to-superficial: intrinsics, erector spinae, and splenii. The extent of muscle insertions increases in the same sequence. The deep layer consists of small muscles. The splenials extend all the way down to the ribs (Fig. 10.87).

Medial motor column supplies anterior neck muscles either directly by ventral roots or via cervical plexus. Note key role of Scalenus muscles. Longus colli (C1–C6) innervated via the cervical plexus, C1–C4, and as isolated branches from C5 to C6. Starts at S6–S9, sparing the dens (S5). It is in continuity with scalenus medius. Longus capitis connects r8 basioccipital to anterior neck and is in continuity with scalenus anterior. Both anterior and medial scalen are innervated by C4 via cervical plexus. Scaleus posterior is not supplied by C4. C4 innervates supraclavicular c4 zone. C4 from central motor column supplies diaphragm. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Intrinsics
Three of these four muscles are monosomitic; from deep to superficial and from medial to lateral, they are:
-
Multifidus muscles have a rotatory function. They span from the caudal surface of the laminae outward to the transverse process over levels C2–C5.
-
Semispinalis spans from the spinous processes of C2–C5 to a more lateral position on posterior surface of transverse process 6 segments below.
-
Interspinales These muscles are short and stubby. They connect the apices of spinous processes spanning from one vertebra segmentally to the next. Thus, they exist as pairs straddled on either side of the interspinous ligament. There are three groups: cervicis, thoracis, and lumborum. Of interest to us is interspinales cervicis. It has six pairs. The first interspinal is between axis and C3; the last one is between C7 and T1. We don’t find analogous muscles between atlas and axis, nor do they exist between skull base and atlas. Evolutionary relationships between proatlas, atlas and axis, and atlas explain this curious absence.
-
Posterior intertransversari These worm-like muscles connect the transverse processes of neighboring vertebrae. Although they are found along the entire length of the spine, they are best developed in the cervical region, where they exist as seven paired slips, directed posteriorly and anteriorly. ITC has a medial slip that connects to the posterior tubercle of the next caudal transverse process. It belongs to the intrinsic back muscles, so its nerve is posterior primary ramus. ITC also has a lateral slip that connects to the anterior tubercle of the next caudal transverse process. It is therefore analogous to intercostal muscle. It is therefore served by anterior primary ramus. Nested in between these muscles at every level is the ventral primary division of the respective cervical nerve. The biologic significance of these small muscles is as a watershed between ventral primary rami and dorsal primary rami.
Erector Spinae
The erector spinae group are polysomitic muscles. They are large and fleshy, extending from skull to sacrum. They are arranged in three vertical columns lying on either side of the vertebral column extend from skull to sacrum. These are spinalis (lateral column), longissiums (intermediate column), and iliocostalis (lateral column). Four regional groups are determined by the superior point of attachment: capitis, cervicis, thoracis, and ililumborum. Innervation is by dorsal primary division of spinal nerves.
-
Spinalis cervicis is poorly represented. When present, it comes from higher up on the spinous processes of axis, C3, and C4 with secondary insertion into the midline ligamentum nuchae and spinous processes of C7 and T1(T2). On occasion, contained within spinalis cervicis is an unusual muscle, spinalis capitis.
-
Longissimus capitis This muscle arises from articular processes of cervical vertebrae 5–8. It insets into mastoid process deep to splenius capitis (and to sternocleidomastoid). This implies it develops prior to splenius. Its nerve supply is c5–c8 (lower cervicals + middle).
-
Longissimus cervicis spans from the posterior tubercle of transverse process from C2 to C6. The muscles slips extend into the thoracic transverse processes of T1–T5.
-
Iliocostalis cervicis spans from posterior turbercles of transverse process of C4–C6 into the angle of ribs 3–6.
Splenii
-
Splenius capitis Innervation is from posterior rami of C4–C5. Muscle originates from the first cervical somite (S5) from occipital bone immediately inferior to the lateral 1/3 of superior nuchal line and from mastoid process. It is covered by sternocleidomastoid. It swoops downward, bypassing the neck altogether. It touches down with secondary attachments to the spinous process of the seventh cervical vertebra and to thoracic vertebrae 1–3. Innervation.
-
Splenius cervicis The primary attachment of this muscle is to the posterior tubercles of transverse processes of cervical vertebrae 2–3. The secondary attachment is to spinous processes of thoracic vertebrae 3–6.
Hypaxial Muscles Interconnecting the Skull and Cervical Spine (MMC)
The muscles under consideration are: rectus capitis anterior, and rectus capitis lateralis, longus capitis, and longus colli. These four muscles can be considered as a single functional unit. RCA and RCL stabilize the atlanto-occipital joint. RCA and LCp antagonize cervical extension and rotate the head. The most significant point is the manner in which three of four muscles “map out” the incorporation of proatlas into posterior cranium. The attachment patterns of these four muscles, based on their developmental “maturity,” illustrate perfectly the rules described in the introduction. Rectus capitis (c1) has priority for attachment over rectus capitis lateralis (c1–c2). Both recti have priority over longus capitis (c1–c3) and longus colli (c2–c6).
-
Rectus capitis anterior This muscle develops from the first cervical somite (S5). S6 may contribute. It is hypaxial, a flexor, and innervated by anterior ramus of C1 (plus, sometimes, C2). The muscle is situated directly behind longus capitis. Its primary attachment is to the posterior lip of foramen magnum, that is, r11. It passes downward and laterally to a secondary attachment to lateral mass of atlas. Together with longus capitis, RCA antagonizes the extensors of the head. As yet another example of r11–c1 mesenchymal continuity, RCA is considered a cranial continuation of the anterior intertransverse muscles of cervical vertebrae. In point of fact, RCA unites proatlas with the true second cervical vertebra.
-
Rectus capitis lateralis This muscle develops from first and second cervical somites (S5–6). RCL is hypaxial, a flexor, and innervated by a branch coming off the loop between ventral C1 and C2. RCL lies lateral to RCA. Its primary attachment is to jugular foramen (lateral r10–r11). The secondary attachment is to transverse process of atlas. Its action is to laterally flex the head and, with RCA, to antagonize the extensor muscles.
-
Longus capitis This muscle is supplied by C1–C3 therefore it originates from the first-third cervical somites (S5–7). Its primary attachment is to r10–r11 basioccipital bone lateral to pharyngeal tubercle, that is, anterior and neuromerically cranial to anterior arch of atlas. Its most cranial somite of origin is cranial to that of longus colli. Therefore, longus capitis develops and migrates prior to longus colli. Its secondary insertion is to anterior tubercles of transverse processes of cervical vertebrae 3–6. This attachment pattern makes sense for two reasons. Fourth occipital somite and first cervical somite are directly adjacent. Basioccipital binding sites mature prior to those of cervical vertebrae. The action of longus capitis is to flex the head and upper cervical vertebrae.
-
Longus colli This muscle develops from the second-sixth cervical somites (S6–10). It is hypaxial and therefore a flexor; it is therefore innervated by the ventral rami of C2–C6.
-
The muscle has three parts. (1) Vertical portion arises from anterior surface of bodies of cervical vertebrae 2–4 and inserts into anterolateral bodies of cervical vertebra 5–7 and thoracic vertebrae 1–3. (2) Superior oblique portion arises from transverse process anterior tubercles of cervical vertebrae 3–5. It ascends to the atlas, inserting into a tubercle on the anterior arch. The tubercle also bears anterior atlanto-occipital membrane ligament, representing r11–c1 proatlas. (3) inferior oblique portion arises from transverse process anterior tubercles of cervical vertebrae 5–6; it inserts into the anterior bodies of thoracic vertebrae 1–3.
-
Development of longus colli involves a temporal-spatial gradient. The first phase occurs as mesoderm of the vertical portion flows from somites 6 to 8, “spilling” downward and centric. The second phase involves “overflow” from somites 7 to 9 in the opposite direction. This sequence is identified because somite 2 does not contribute and because flow is “mirror image,” that is, upward and centric from the most cranial (most mature) population. The final phase involves contributions from Inferior oblique portion flows from somites 9–10 but, because midline binding sites are occupied by the previous vertical portion that has commandeered the midline, the myoblasts are forced to make use of more lateral insertion sites. The actions of this muscle are nothing to write home about. It weakly flexes, rotates, and laterally bends the neck. Taken together, longus capitis and longus colli replicate the neuromeric distinction between neck and thorax. The cranial attachment of the former duplicates the proatlas. The caudal attachments of the latter occupy the anterior vertebral column from atlas to the third thoracic vertebra.
-
Hypaxial Muscles Connecting Neck and Trunk (MMC)
The three scalenes have shared functions: (1) fixation of the rib cage on forced inspiration, (2) ipsilateral bending of the neck, and, due to obliquity, rotates it. All scalenes are hypaxial; they are supplied by the ventral division of the respective cervical nerve. The scalenes have a different embryologic basis and develop prior to sternocleidomastoid. Absence of the latter has been reported in isolation from the scalenes.
-
Scalenus anterior This muscle develops from somites 8–10. It is supplied by C4–C6. Scalenus anterior lies under cover of sternocleidomastoid. Its primary insertion is as the confluence of slips from the tubercles of transverse processes of cervical vertebrae 3–5. The secondary attachment is to scalene tubercle of upper inner surface of first rib at the scalene tubercle.
-
Scalenus medius This muscle develops from somites 7–12. It is supplied by C3–C8. The most extensive of the scalenes, its primary attachment is as confluent slips from the posterior tubercles of cervical vertebrae 3–8. The secondary attachment is to cranial first rib. It lies directly anterior to the sternocleidomastoid.
-
Scalenus posterior This muscle develops from somites 10–12. It is supplied by C6–C8.
Muscles Connecting Pectoral Girdle with Pharyngeal Arches (MMC)
Posterior Hypobranchial Muscles
In contrast to the anterior hypobranchial muscle series from occipital somite and dedicated to the tongue, posterior hypobranchial muscles arise from mesenchyme from the first–third cervical somites (S5–S7). These muscles are derived from the coracomandibularis in fishes which serves as a depressor of the mandible and is supplied by occipital nerves. In tetrapods, posterior hypobranchial muscles function as stabilizers of mobile structures (hyoid and thyrohyoid) during the act of swallowing. They are innervated by cervical plexus.
The migration pattern of these muscles can be deduced from the anatomy of the ansa cervicalis, a component of the cervical plexus. The motor nuclei dedicated to these muscles reside in the lateral motor column of c1–c3. (Previous opinion had erroneously held these fibers to be hypoglossal…they aren’t!) Infrahyoid myoblasts follow the “slime trail” laid down by pioneering tongue myoblasts from the occipital somites. Thus, they slavishly follow the hypoglossal nerve. At the base of the tongue, they encounter hyoid, which offers them four very attractive binding sites. These are arranged along the hyoid body by order of maturity, from posterior-to-anterior: thyrohyoid. Omohyoid, sternohyoid, thyrohyoid, and geniohyoid. Thus, they insert successively into fourth, third, second, and first arches. Like jet fighters, the myoblasts (along with their attendant motor nerves) will peel off in succession as they seek attachment to the hyoid binding sites. The consequent neuroanatomy is a faithful replica of this biologic sequence.
Despite some old-fashioned and confusing terminology, ansa cervicalis is as simple as “1–2–3.” (1) Superior root of ansa is made predominantly from one nerve, C1. (2) Inferior root of ansa contains two nerves, C2 and C3. Together these three nerves innervate six muscle bellies (1 + 2 + 3 = 6). The union of the two roots forms a loop lateral and ventral to common carotid. Nota bene: because C1 has been long confused with XII, the superior root is sometimes called descendens hypoglossi whereas inferior root has been named descendens cervicalis. My advice is to ignore these terms…just know where they came from.
Fibers from C1 and C2 (via a secondary loop with C1) “hitch a ride” with hypoglossal. At hyoid bone, some of them hop off, providing motor nerves to geniohyoid, thyrohyoid, and omohyoid. Other C1–C2 fibers continue downward to form the superior root of ansa. The departure of the first three motor nerves is somatotopic. Geniohyoid travels the farthest along XII. Next, thyrohyoid parts company with XII at the posterior border of hyglossus. Finally, omohyoid starts out with the superior root of ansa but jumps ship soon thereafter. to innervate the muscle. Fibers from C2 and C3 form the inferior root of ansa. Right where the two roots join together, two more nerves jump off to innervate first sternohyoid, then sternothyroid. Finally, at the very bottom of the loop, the nerve to the inferior belly of the omohyoid is given off. Note that C4 is sometimes attributed to this plexus but, in reality, it remains the supply for the diaphragm. Recall that C4 provided a motor supply for the ancient subcoracoideus which in the reptilomorphs, becomes internalized inside the chest and invades the lateral thoracic fold that grows inward as the anlagen of the diaphragm.
Migration sequence of the hypobranchial muscles: cranial-to-caudal.
The hypobranchial (sic) muscles originate from eight consecutive somites. Occipital somites S1–S4 give rise to the muscles of the tongue and to a single prehyoid strap muscle, geniohyoid (Chap. 9). The first four cervical somites, C1–C4, give rise to the post-hyoid strap muscles. Their function is to connect the clavicle and manubrium (the ancestral coracoid) of the pectoral girdle with the ventral aspects of the pharyngeal arches all the way forward to the mandible. As such, they act as depressors of the lower jaw.
-
Geniohyoid The first “target” of C1, this muscle “hitches a ride” with hypoglossal nerve, thus betraying its origin from S5. For this reason, it is not considered a true part of ansa cervicalis. Geniohyoid is the forward continuation of the coracomandibularis; for this reason, it is referred to as the prehyoid strap muscle. Geniohyoid has its primary attachment on the superior surface of hyoid and greater cornu (second arch). It is deep to the V3 mylohyoid. Having a narrow belly, GH runs forward to insert on the most “mature” sector of the r3 mandible, the genial tubercle. Tightening of GH “fixes” the floor of mouth during food ingestion. It originates from a single myotome (somite 5) and is supplied by C1.
-
Thyrohyoid The first target of ansa is also small muscle from the fifth somite. It has a primary insertion from the oblique line of thyroid cartilage (fourth arch) and spans over to the inferior surface of the hyoid greater cornu (third arch). Its action is a bit indecisive: it can either depress hyoid or elevate thyroid. True to its size, TH merits a single nerve, C1.
-
Superior belly of omohyoid The third target of ansa is a long-distance muscle (OHs) originating from lower border of hyoid posterior to thyrohyoid and lateral to sternohyoid. It forms a central tendon which is tethered at the level of cricoid cartilage by thickening of deep investing fascia. It acts with its companion inferior belly to stabilize hyoid on the clavicle (more on this anon). As a larger three-myotome muscle, OHs are supplied by C1–C3.
-
Sternothryoid The first target of ansa, this muscle (ST) is misnamed. Although described as “inserting” on the oblique line of thyroid cartilage, its fibers of secondary inerstion are confluent with the most posterior binding site on hyoid bone, deep to sternohyoid. It therefore seeks out fourth arch and third arch. ST migrates earlier than SH. Accordingly, its primary insertion is the lowest on sternum, at the manubrium and on the cartilage of first rib, spilling over to the second rib.
-
Sternohyoid The second target of ansa, this muscle (SH) has its primary insertion into the posteromedial clavicle, that is, the c1 pole, and sternum’s “backside.” Its secondary insertion is into lower body of the r6 hyoid bone, that is, third arch. Swallowing activates suprahyoid muscles. These elevate hyoid and larynx. Sternohyoid provides an opposing, stabilizing force. At times, it is reinforced by an additional cleidohyoid muscle, forming sternocleidohyoid. These muscles develop in the first, second, and third cervical myotomes. They are supplied by C1–C3.
-
Inferior belly of omohyoid The final target of ansa, second belly of the longest strap muscle (OHi) sweeps over SH and ST, passing deep to r8–r11/S1–S4 sternocleidomastoid in search of a rather elusive binding site. This is not an easy proposition. All the “parking spots” around the “sternal square” are taken. The clavicle is also off limits because its entire superior surface is taken up by a pair of “occipital occupiers,” sternocleidomastoid and trapezius. So, in desperation, omohyoid gloms onto the posterior aspect of superior border of scapula. Taken together, the action of OHsup and OHinf is not only a depressor of hyoid but also a dorsal retractor. Cervical fascia is thereby tensed.
Muscles Connecting Pectoral Girdle with Vertebral Spine: MMC, LMC, and SAC
We shall discuss these muscles in the spatiotemporal order in which they are produced. This involves analysis of the neuromeric order of muscle origin, followed by that of its primary and secondary attachments. For example, both rhomboid minor and major originate from the fifth and sixth cervical somites, S9–10. The minor is C4 and the major, being larger is C4–C5. Primary attachments for both are to medial border of scapula, a chondral bone originating from lateral plate in register with levels c4–c8. The rhomboids choose their binding sites based on the relative biologic maturity of medial scapula following a cranial-caudal gradient. Thus, minor binds more cranial than major. The area of insertion of major, being a two-myotome muscle, more extensive. Secondary insertion sites follow the same gradient as well. Thus, rhomboid minor from upper medial scapula attaches to the seventh cervical and first thoracic vertebrae while rhomboid major from lower medial scapula attaches to the second-seventh thoracic vertebrae.
These muscles then are arranged in two layers. The ventral layer consists of levator scapulae, followed by rhomboid minor and major. The dorsal layer contains trapezius (a product of cervical somites 1–5) and latissimus (a product of cervical somites 6–8). The primary attachment site of trapezius, scapular spine, is a neomorph. It was not part of the bones making up the original scapular anlage. Synthesis of this muscle group is ventral-dorsal. Hence, the medial border muscles are covered over by trapezius.
-
Levator scapulae (MMC) This muscle develops from the third and fourth cervical somites (somites 7–8) and is innervated by C3–C4 from cervical plexus, a long lever arm. Its primary attachment is to transverse processes of atlas, axis, cervical vertebrae C3–4. The secondary attachment is to superior medial border of scapula.
-
Rhomboid major and minor (LMC) Both of these muscles arise from the fifth cervical somite (somite 9) and are innervated by C5 via dorsal scapular nerve of brachial plexus. Major has its primary insertion into medial border of scapula between the root of the spine above and the inferior angle below. Its secondary attachment is from spinous processes of thoracic vertebrae 2–7 (these comprise neuromeres t1–t8). The primary insertion of rhomboid minor into scapula lies above that of major, into the triangle-shaped medial surface of spine. Its secondary attachment to the spinous process of seventh cervical and first thoracic vertebrae. The actions of the rhomboids are to adduct scapula, i.e. they stabilize it when it bears weight. Inward rotation of angle of scapula by the lower muscle helps the adduction of the arm.
-
Trapezius (SAC) Discussed separately under muscles connecting pectoral girdle to the skull.
Muscles Connecting Pectoral Girdle with Body Wall (LMC)
Ventral Body Wall
-
Subclavius A cylindrical muscle, subclavius arises from fifth to sixth cervical somites and is supplied by their respective segmental nerves (C5–C6), its primary attachment is the groove inferior surface of middle 1/3 clavicle. Its secondary attachment is to first rib and the first costochondral junction. It stabilizes the bone during movement.
-
Pectoralis minor Smaller than its confrere, this muscle arises from the eighth cervical and first thoracic somite. It is innervated by medial pectoral nerve (C8 and T1). Its primary attachment is to medial-superior surface of coracoid process. Its secondary attachment is to the third–fifth ribs. Note that two other muscles have primary attachments to coracoid process: c5–c7 coracobrachialis and c5–c6 short head of biceps. The implication is that the coracoid process has three developmental zones, arranged from lateral to medial. Biceps (short head) binds first, coracobrachialis follows, and pectoralis minor is last.
Dorsal Body Wall
-
Serratus anterior This thin muscle arises from fifth to seventh cervical somites; it is supplied by long thoracic nerve of brachial plexus (C5–C7). Its primary attachment is to the ventral surface of medial scapula. In this regard, it lies opposite the c5 rhomboids, which are attached to dorsal surface of medial scapula. The secondary attachments are to the superior surfaces of ribs 1–9. Each digitation (except the first) is intercostal, that is, it is attached to two ribs.
-
Latissimus dorsi Not technically part of the pectoral girdle, but it has a physical relationship to the dorsal surface of scapula, being superficial to it. This large muscle is of enormous utility for reconstructive surgeons, It develops from cervical somites 6–8. It is supplied by C6–C8 thoracodorsal nerve. Its primary attachment is via 7-cm quadrangular tendon which passes anterior to teres minor and is anchored to intertubercular groove of humerus (zone c6–c8). Its secondary attachment is very extensive: to the spinous processes of T7–T12, via thoracolumbar fascia all the way down to sacral vertebrae; laterally it attaches to iliac crest. It is covered over from T7 to T12 by trapezius, as the latter muscle forms later.
Muscles Connecting Pectoral Girdle to the Upper Extremity (LMC)
Nine muscles extend from scapula to proximal half of humerus to provide control of the arm. Apart from supraspinatus of them are polyneuromeric. All derive at least part of their mesenchyme from the fifth cervical somite. Secondary attachment sites vary according to the relative biologic maturity of the muscle and of its osseous attachment site along the humerus.
-
Supraspinatus Arising from the fifth cervical somite (S9), this muscle fills the supraspinous fossa. Secondary attachment to humerus at the most proximal fossa of greater tubercle. It is supplied by suprascapular nerve (C5), the upper division of upper trunk. Recall that the lower division contributes to lateral cord. A rotator cuff muscle stabilizes the shoulder by tractioning the humerus inward to glenoid fossa. It rotates humerus laterally. Supraspinatus also may initiate abduction, one of the reasons that pain with rotator cuff is initiated with abduction.
-
Infraspinatus This muscle is much larger, arising from fifth and sixth cervical somites (S9–S10). It is supplied by suprascapular nerve (C5–C6). Its fibers are confluent laterally with teres minor and inferiorly with teres major. Secondary attachment to humerus is into fossa #2 (intermediate) of greater tubercle. Also rotator cuff muscle, its actions are similar but it is not involved in abduction.
-
Subscapularis This muscle from fourth and fifth cervical somites; supplied by upper and lower subscapular nerves. Its primary insertion covers virtually the entire ventral surface of the scapular body. Secondary attachment to lesser tubercle and anterior capsule of shoulder joint. Supplied by C4–C5 upper and lower subscapular nerves from posterior cord. These are proximal to the terminal branches, axillary nerve, and radial nerves.
-
Teres minor develops from the fifth and sixth cervical somites (S9–10) and has a broad primary insertion along upper medial (axillary) border of scapula. It is supplied by C5–C6 via the posterior cord and axillary nerve. Teres minor wraps around the border of scapula in two directions. Posteriorly, its fibers are confluent with infraspinatus which is innervated by C5–C6 suprascapular nerve. Anteriorly it borders with subscapularis which is supplied by C5–C6 upper and lower subscapular nerves. These muscles plus supraspinatus, all originating from S9 to S10, constitute a functional unit known collectively as the rotator cuff. Its secondary insertion into the humerus consists of two tendons: upper fibers attach to fossa #3 (the most distal) of the greater tubercle. Lower fibers go distal.
-
Teres major is larger. It develops from cervical somites 5–7 (S9–S11). Its primary insertion is to lower the medial border of scapulal; it covers the angle. It inserts secondarily into the humerus at the medial lip of intertubercular sulcus. Teres major is innervated by a branch of lower subscapular nerve, a branch of posterior cord. It shares a common nerve supply with lower subscapularis. These two muscles plus supra/infraspinatus constitute the rotator cuff. They have a common root.
-
Regarding binding sites on humerus, both of the teres muscles demonstrate spatial precedence over latissimus. Minor binds to C4 greater tubercle, major binds to medial lip of intertubercular sulcus, whereas latissumus binds to the bottom of intertubercular sulcus (the C6–C8 zone of humerus). From here, latissiumus makes very extensive secondary attachments. The myoblasts migrate around lateral border of teres majors and spill out over the surface of underlying serratus to gain attachment to thoracolumbar fascia.
-
-
Coracobrachialis This is the second muscle to have its primary insertion into coracoid process. It inserts secondarily into the medial shaft of humerus deep to and proximal to biceps, between the origin of triceps and brachialis. It is innervated by usculocutaneous nerve from upper trunk (C5, C6) and lower trunk (C7).
-
Short head of biceps brachii This muscle is the third and most lateral of the “coracoid chorus.” It joins long head of biceps to gain secondary insertion into radial tuberosity. The insertion site of short head is into tuberosity of the radius, which is more distal than that of long head of biceps. The origin is C5–C6 via musculocutaneous nerve.
-
Long head of biceps brachii Originating from C5 to C6 (S9–S10), long head has its primary attachment to the supraglenoid tuberosity at the superior margin of glenoid labrum.
-
Pectoralis major This muscle arises from the fifth–eighth cervical somites and the first thoracic somite; it is innervated by two nerves: lateral pectoral nerve (C5–8 and T1) and by medial pectoral nerve (C8–T1). This muscle has two heads: (1) clavicular head inserts into medial ½ of clavicle; (2) sternocostal head inserts into the junction of ribs 1–7 with sternum. The secondary insertion of pectoralis major is into the humerus as a flat tendon 5 cm in width attaching to the terminus of lateral lip of intertubercular sulcus (bicipital groove). The primary action of pectoralis major is adduction of the arm and (secondarily) as medial rotator of humerus. Clavicular fibers flex and adduct the humerus. Sternocostal fibers extend, against resistance, the flexed humerus against the side of the chest…as in giving a backwards elbow to someone standing behind.
-
Deltoid Arising from fifth and sixth cervical somites; deltoid is supplied by axillary nerve (C5–6). Deltoid tuberosity ½ way down shaft, lateral side. Deltoid is a “late-comer” which covers over other muscles. Furthermore, it is further out on the brachial plexus, being innervated by one of the two terminal branches of posterior cord via C5–C6 axillary nerve.
Note that the deltoid and pectoralis “layer over” the anterior scapular muscles. They accomplish this because they migrate beneath the skin, giving passage for migration of deeper muscles that achieve more direct attachment to the pectoral girdle.
Muscles Connecting the Pectoral Girdle to the Skull (SAC = CMC Lateral)
Sternocleidomastoid
Sternocleidomastoid is a hypaxial polyneuromeric somitic muscle. It develops from the myotomes of the first two to three cervical neuromeres, that is, at c1, c2, and c3. It receives motor innervation from C1 to C3 via spinal accessory nerve. The central motor column in which SAN is located is a continuation of branchiomeric nucleus ambiguus in the medulla but there is no physical connection between it and the putative intracranial division of cranial nerve XI (Fig. 10.88).

Sternocleidomastoid. SCM has primary insertion into neuromeric sites c1–c3 along clavicle. Spinal accessory nerve CN XI develops in the lateral motor column of r8–r11 and the embryonic basal plate c1–c6. SCM is likely supplied by cranial + c1–c2 whereas trapezius receives motor contributions down to c6. Left: [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918.] fig. 386. Right: [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones]
Neuromeric Coding of Clavicle and Manubrium
To understand sternocleidomastoid, we must bear in mind the neuromeric coding of its primary insertion sites, clavicle and manubrium, and of its secondary insertion sites, the mastoid and occiput. Clavicle is like a sausage-like rectangular box made of two types of mesenchyme. Its peripheral “skin” is neural crest and its core is mesoderm. The medial articulation with manubrium is c2 and the lateral articulation with scapula is c6. The dorsal and superior sides are a gradient from c2 to c6. The ventral and inferior sides are exclusively c5–c6. OK, so what about the coding for c1? Why don’t we see it in clavicle? The answer lies in the mysterious manubrium.
Manubrium is considered part of sternum but appearances can be deceiving. Recall that the body of the sternum is formed by the vertical fusion of five parallel bars of lateral plate mesoderm. It forms separate articulations with manubrium and xiphoid. These seven segments articulate with seven ribs. Manubrium is a fused Y-shaped structure consisting of a central rectangle of l1 lateral plate mesoderm from which project two sets of shoulder-like processes. The inferior processes are located at the ¾ mark of the bone. These articulate with first rib, a combination of C8C and T1R paraxial mesoderm. They also bear on their anterior surface the upper limit of insertion for pectoralis major. The superior processes project from the upper ¼ of manubrium; they articulate with clavicle and bear the insertion of the C1 sternomastoid. It is likely that the upper ¼ of manubrium is phylogenetically distinct. It represents the fusion of two c1 anterior coracoid processes. For this reason, the posterior aspect of manubrium bears in its upper 1/4 the insertions of two strap muscles. Sternohyoid attaches to its upper posterior margin and to posterior clavicle. Below SH is sternothyroid which also attaches to the cartilage of first rib. Evidence for the separate nature of upper manubrium exists in running mammals that lack clavicles but present a functional sternum bearing a normal set of first ribs.
Ossification centers provide additional clues about the distinct nature of manubrium. In humans, the ossification center for manubrium is considerably larger than those of the sternal body. This perhaps indicates the coalescence of four centers. Clavicle manifests two primary centers in its shaft which ossify at weeks 5–6 of life. A separate secondary center at the sternal end of clavicle appears at ages 18–20 and unites with the rest of the bone at age 25.
Mastoid process, as previously described is derived from post-otic paraxial mesoderm corresponding to r6–r7. It develops after birth and its expansion is ascribed to traction placed on it by the neural crest fascia of sternocleidomastoid. Mastoid is confluent with parietal, interparietal and supraoccipital. The boundary between interparietal and supraoccipital is superior nuchal line. The latter bone develops from occipital somites, S1–S4 so superior nuchal line is likely encoded r8–r11. The order of insertion into mastoid is, from medial to lateral: sternomastoid, cleidomastoid, and cleido-occipitalis.
Knowing these facts, we can better understand the insertions of SCM and how they relate to those of neighboring strap and scalene muscles.
Motor Control and Insertions of Sternocleidomastoid
As previously described, in the rat model the motor nucleus of SCM within the central motor column exists in two discrete locations. Sternomastoid and cleidomastoid are supplied from the dorsomedial edge of the ventral horn between C1 and C3. Their colleague, cleido-occipital is supplied from the dorsolateral edge of ventral horn from C2 to C3. It shares this position with trapezius muscle C2–C6. For this reason, the secondary insertion of the three parts of SCM into the skull follows a ventral to dorsal pattern with cleido-occipital forming last and taking a higher primary insertion site. Also, SCM develops prior to its partner, trapezius, and inserts deep to it [17, 18].
Sternocleidomastoid follows an antegrade–retrograde insertion sequence. The muscle has two functional components and three primary insertions, into the ventral components of the pectoral girdle.
The sternal head is attached to the C1 manubrium whereas the clavicular head is attached to C2–C3 clavicle. Recall that manubrium represents the ancient mesodermal anterior coracoid. The sternal component of SCM is rounded and is attached to the upper border of the manubrium. The clavicular component is broad and flat. It is attached along the anterior surface of the medial 1/3 of clavicle. SCM may be somatotopic throughout its length. Sternomastoid and cleidomastoid are medial to occipito-mastoid with an insertion sequence of c1–c2–c3. Recall that c1–c2 manubrium is derived from the original anterior coracoid process.
As we shall see, the posterior surface of the SCM interacts with the hypobranchial strap muscles sternohyoid and sternothyroid, both of which arise from S5 to S7 and are innervated by C1–C3 via cervical plexus. These muscles develop prior to SCM are positioned posterior to it. Sternothryoid is most medial. It binds to lower posterior manubrium and opportunistically to adjacent first rib. Thus, ST lies below SH. Sternohyoid inserts into posterior manubrium above ST but below and behind sternal head of SCM. It also inserts into posterior surface of medial clavicle. This suggests that clavicular development proceeds from posterior (dorsal) to anterior (ventral).
The secondary insertion of SCM into mastoid process is perfectly logical. Recall that first cervical somite, S5, participates in the formation of the craniovertebral joint. The insertion sequence of muscles follows a strict spatiotemporal sequence. Medial motor column muscle development preceeds that of central motor column so MMC muscles get “first choice” for binding sites. [For this reason, congenital absence of SCM has been reported as a rare clinical deformity but the developmental failure of the suboccipital muscles would be catastrophic, incompatible with life.] All S5 muscles supplied by dorsal branch C1 from the medial motor column span upward from the suboccipital triangle and attach to binding sites on the chondral occipital bone complex. SCM, being thus excluded from the “occipital parking lot,” has as its first available site the mastoid process of temporal bone. It inserts into the mastoid process, all the way from its superior border to the apex. In addition, cleido-occipital inserts via a thinner aponeurosis higher up on the skull, leaping over C1 epaxial muscles to find a vacant parking spot along the superior nuchal line, that is, at the border between neural crest membranous bone and PAM chondral bone. Both these insertions make neuromeric sense. Mastoid, being r6–r7, borders directly on the r8 occipital parking lot. Superior nuchal line is also a boundary between the supraoccipital complex and the interparietal complex.
The neurovascular supply to SCM is strongly segmental and represents four distinct sources. Proximal third is supplied ECA branches representing AA2 and AA3. second arch is represented by occipital artery and third arch is represented by ascending pharyngeal artery. APA has an interesting course. It has two divisions, pharyngeal and neuromeningeal. From the latter, an odontoid arch branch supplies the craniovertebral junction. The latter gives off a musculospinal branch that connects at level C3 with ascending cervical artery from the thryocervical trunk. Middle third receives fourth arch superior thyroid artery, and third arch ECA is too far internal to lend a hand. Distal third is supplied by variable branches from thryocervical trunk: inferior thyroid, suprascapular, ascending cervical or transverse cervical. These latter represent the substitute for the defunct AA5 to the fifth arch.
Is there a hidden significance to this vascular pattern? SCM straddles from skull to pectoral girdle. Deep to its proximal 2/3 lie the structures of pharyngeal arches PA2–PA5. It makes sense that these neuromeric zones, these angiosomes include their muscle cover. Distal 1/3 of SCM covers the zone which is caudal to the fifth pharyngeal arch and includes trachea and esophagus. All these structures lie distal to the aortic arch system; accordingly, they are supplied by derivatives from second axis off the subclavians, that is, the vertebrals.
Using the Mathes/Nahai classification, sternocleidomastoid is a type V muscle. It does not have an axial vessel. Instead, it picks up arteries from surrounding structures along its course (in this case, the skin). Gracilis is another type V muscle. Because these muscles do not have a dominant pedicle they have a short arc of rotation and their usefulness for reconstructive surgery is limited [9, 19, 20].
We conclude with comments regarding congenital absence of sternocleidomastoid, an extremely rare clinical entity, with four cases reported. A review by Vajramani documents a case with MRI showing involvement of both SCM and trapezius with no other anomalies noted, including deficits higher up involving palato-pharyngo-laryngeal nerve. This supports the clinical independence of spinal accessory nerve. SCM absence has been found in isolation with a normal trapezius indicating potential autonomy of the roots. Strap muscles and scalenes are intact. Reports of congenital absence of trapezius are much more numerous. This raises the question as to why SCM is more “protected.” Its synthesis could be more primordial that that of trapezius. As we shall see, mammalian division of cucullaris into these two muscles involves a concomitant process of change at the level of neuromeres c4–c5 which results in additional lengthening of the neck and expansion of the brachial plexus. Since trapezius is a c2–C6 and the only neuromere it shares in common with SCM is c2. Perhaps the pathology occurs in a posterior-to-anterior manner, with level c2 being hit last. We shall look at this topic further when we assess the spectrum of variations seen in trapezius [17, 18] (Fig. 10.89).

Absent right sternocleidomastoid No other findings. Ipsilateral trapezius intact with intact spinal accessory nerve. [Reprinted from Singh HP, Kumar S, Agarwal SP. Congenital asymptomatic absence of unilateral sternocleidomastoid muscle. BMJ Case Rep 2014. DOI: 10.1136/bcr-2013-202,786 PMID: 24445852. With permission from BMJ Publishing Group Ltd.]
Trapezius
Trapezius is a hypaxial polyneuromeric muscle found only in mammalian. it develops from somites S6–S10, levels C2–C6. Recall that sternocleidomastoid arises one somite higher than trapezius, from somites S5–S6. The muscles share motor innervation is from spinal accessory nerve with serial nuclei nucleus in the cervical spinal cord from levels c1 to c6. The nucleus for trapezius is located at the posterolateral margin of anterior horn; it also supplies the cleido-occipital part of Scm (Fig. 10.90).

Spino-Trapezius extends from r8 to r11, c1–c8, and T1–T12. Despite its dorsal position, trapezius is a hypaxial muscle—its motor nerves are ventral. Upper fibers (r8–r11 to c1–c7) are supplied by CNCXI and ventral branches of c2–c4; with sensory and proprioceptor innervations from c1 to c4. Middle fibers (c7–t3); lower fibers (t4–t12). The implication for motor control is that spian accessory nerve is extensive. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Trapezius and the Neuromeric Coding of Scapula
Scapula develops from lateral plate mesoderm in register with neuromeres c2–c6. Trapezius inserts along the scapular spine which is likely encoded from medial to lateral. Straddling on either side of it are the c5–c6.
Trapezius has many characteristics in common with sternocleidomastoid. (1) Both muscles are innervated from the central motor column of the cervical spinal cord, which represents the caudal continuation of nucleus ambiguus. (2) The muscles receive their blood supply, in part, from the external carotid artery and in part from subclavian. (3) Trapezius and sternocleidmastoid are descendents of the ancient cucullaris muscle, a lineage extending back 500 million years ago to the placoderms of the Devonian period. (4) Their ontology reflects the changes wrought by evolution in constructing the neck, changes that are inseparable from the transition to land.
Trapezius in mammals terminates at somite level S10…this is significant. First off, in basal amniotes, that is, the anapsids, have six cervical somites and the number of cervical vertebrae stabilizes at five. The neck-trunk interface in anapsids lies between somites S10 (sp6 or C6) and S11 (sp7 or T1). The brachial plexus in basal amniotes (and in all non-mammals) has four spinal roots (C4–C5–C6–T1). As a consequence, br1 is located at somite level S8, precisely at C4. S8 represents a fork in the road for evolution. Be it in the direction of diapsids (birds and reptiles) or synapsids (mammals), necks got longer. In contrast, brachial plexus remains invariable in diapsids. It continues to have four roots regardless of its location.
Mammals are innovators. By duplicating levels C4–C5, they create a brachial plexus of six spinal roots (C4–C5–C6–C7–C8–T1). Nonetheless, even in mammals, C4 remains a key transition point. It marks the first neuromere of the brachial plexus. C4 is also the site of origin for diaphragm. This puts the final mammalian somite count to 8 (and the number of cervical vertebrae to 7).
Evolution has embedded something else of deep significance at level C4. The primitive branchial arch system of armored fishes stabilizes in chondrichthyes at 7 gill arches. Given the principle of two neuromeres per arch and with the first branchial arch in register with r2–r3, the seventh branchial arch becomes located at spinal neuromere level sp3–sp4. Thus, the location of the upper extremity, at the interface between neck and trunk, becomes located in early tetrapods, at the end of the branchial arch system.
Trapezius myoblasts originate from neuromeric levels c2–c6 (S6–S10). Like SCM, trapezius is an appendicular muscle: it therefore develops later in time compared with axial counterparts from the same c1–c6 levels. Hence, trapezius must be a master opportunist, glomming on to whatever bindings sites have been left vacant by its predecessors.
Trapezius, like SCM, follows a two-stage model of antegrade and retrograde insertions. In the first stage, myoblasts and accompanying neural crest fascia from c2 to c6 migrate directly outward underneath the skin where they come into contact with their target bone, the scapula, made from c1 to c6 lateral plate mesoderm. Thus, the initial migration of myoblasts takes place within their neuromeres of origin. After making primary attachments, the muscles migrate retrograde to the body axis to seek out available binding sites, sweeping from lateral-medial from r8 to r11 and downward in the midline from c1 to the S5.
Trapezius has three zones of primary attachment to upper posterior scapula from which its fibers reobound medially, fanning out to insert secondarily into the fibro-osseous structures of the vertebral midline. The extent of its attachments are surprisingly meagre. At the time of trapezius development, the bulk of dorsal scapular binding sites are already “taken” by other muscles which previously migrated to the site. These are (above the spine) levator scapulae and supraspinatus and (below the spine) rhomboids, infraspinatus, teres minor, and teres major. Poor trapezius has to settle for the “leftovers.”
The primary insertions of trapezius are as follows:
-
(1)
Superior insertion is relegated to the lateral 1/3 of posterior clavicle. Why such a limited zone? Recall that the anterior clavicle is completely covered over by platysma. The medial 2/3 of posterior clavicle harbors SCM and the strap muscles. So trapezius takes what remains. From there, its fibers span upward and medially to insert secondarily into the skull and the spine. Its occipital insertion is into superior nuchal line along its medial 1/3 because SCM fibers occupy the lateral 2/3. From external occipital protruberance it attaches in a vertical line downward along ligamentum nuchae from C1 to C7.
-
(2)
Middle insertion is restricted to the medial aspect of acromion and lateral scapular spine. Despite the large surface area of the scapular bade per se, no other sites are available because it is completely covered by pre-existing muscles…but the scapular spine sticks upward and is available. From here, Tpz fibers span medially to the neural spines of C8 to T3.
-
(3)
Inferior insertion is located along medial scapular spine (T4–T12). The fibers flow downward and medially; they pass over a bare area between the two fossae of supraspinatus and infraspinatus but do not attach there. They terminate medially on the neural spines of T4 to L1.
Textbook descriptions of trapezius are confusing because the muscle is described as if it had 19–20 somites of “origin.” No such muscle exists in the entre body. Why should such a massive muscle be controlled by a spinal accessory nerve from only neuromeres c2–c6? Furthermore, the midline attachments of the muscle are fibroaponeurotic, not muscular. These make sense only as secondary insertions. Recall that the extent of the insertions depends strictly on (1) the mesenchymal mass of the muscle, (2) the capacity of each binding site to accept a finite amount of insertion, and (3) the total number of sites available. Recall the “toothpaste” model: a finite amount trapezius is expressed, it flows outward in all directions seeking out all available binding sites.
Trapezius, like SCM, is a type V muscle. Its vasculature is defined by its cutaneous environment. The upper 1/3 is supplied by second arch occipital artery, transverse muscle branch. The remainder of the muscle is supplied by the costocervical trunk. The middle 1/3 receives the superficial branch of transverse cervical (or the superficial cervical). The lower 1/3 is supplied by dorsal scapular artery. This reflects the relationship of the first four somites to ECA territory, whereas the terminal eight somites are beyond the distribution of pharyngeal arch circulation.
Trapezius in all non-mammalian amniotes is made from four somites. Why should trapezius expand to six neuromeric levels in mammals? Tetrapod evolution achieves changes in neck length by expropriation of truncal neuromeres into a cervical fate. Mammals are different. They lengthen their necks by duplicating levels sp4–sp5. This interposes two new cervical vertebrae and creates a six-root branchial plexus. Because the mammalian neuromeres c6–c7 are a duplication of c4–c5, two new levels of mesoderm are dedicated to making the dorsal aspect of cucullaris. Sternocleidomastoid stays the same but trapezius has more mesenchymal mass.
Development of Trapezius
Description of trapezius and SCM development are based on embryo dissections by Keibel and Mall based on crown-rump length; we have correlated them with staging as per the Carnegie Foundation.
Stages 14–15 in the 7-mm embryo trapezius develop with sternocleidomastoid as part of the common anlage of cucullaris. During stages 16–17 (11 mm) the anlage divides with trapezius being positioned dorsal. Clavicle and scapula are now distinct. At stages 18–19 (16 mm) trapezius gains attachment to scapula and extends upward toward, but is not in contact with, ligamentum nuchae. The stage 20 (20 mm) embryo demonstrates full secondary attachments of trapezius to the midline.
Trapezius is an adductor. Because of its geometry, it can be considered to have two functional components: upper fibers elevate the scapula while the lower ones draw it downward. Absence of upper trapezius has been reported as have deficiencies of the lower muscle.
Congenital Anomalies of Trapezius
Developmental defects of the trapezius muscle are also uncommon. A 2006 review found eight cases in the literature, at times accompanied by aplasias in other muscles as well. In five cases bilateral aplasia has been reported. Rarer still are deficits involving both Tpz and Scm: only four such cases have been reported [18]. Trapezius deficits occur as a spectrum and these variations provide us with a number of clues as to its development. Unilateral 50% aplasia presented as a progressive dropout of lower fibers with absent insertion below T9. This indicates that the developmental pattern of the muscle is cranial-caudal. Like paint dripping downward on a wall, when the mesenchyme runs out it simply fails to reach the most distal binding site [21]. Unilateral agenesis of the lower trapezius muscle has been reported with absent fibers from T6 to t12. In a cadaver case, a lower defect was explained by the absence of the spinal accessory nerve and blood supply. However, in the same patient, sternocleidomastoid function remained, functioning on upper roots of the SAN. This finding supports the autonomy of the two muscles with SCM originating independently from C1 to C2 roots, somites S5–S6 [22]. A clear-cut distinction seems to exist between upper cervical fibers and those of the middle and lower thirds [23]. Bergin presented a case in which ¾ of the descending (lower) trapezius appeared separate from the remainder of the muscle. The isolated part inserted into clavicle as an independent tendon [24] (Fig. 10.91).

Congenital absence of cervical trapezius. Congenital absence of thoracic trapezius. Left: [Reprinted from Mehra L, Tuli A, Raheja S. Dorscapularis triangularis: ebryologic and phylogentic characterization of a rare variation of trapezius. Anat & Cell Biol 2016 Sep;49(3):213–216. With permission from Creative Commons License 4.0: http://creativecommons.org/licenses/by-nc/4.0/.] Right: [Reprinted from Emsley JG, Davis M. Partial unilateral absence of the trapezius muscle in a human cadaver. Clin Anat 2001; 14(5): 383–386. With permission from John Wiley & Sons]
Upper defects of trapezius also exist. In one such case, the upper fibers were replaced by a fibroaponeurosis spanning from the occiput to C4 and a normal nerve, indicating survival of neural crest connective tissue but not myoblasts. Compensatory hypertrophy of rhomboids was noted but this could reflect a diversion of mesenchyme from one muscle to the other at the level of the myotome [25]. In another case with Tpz loss down to C4 the clavicular insertion was missing [26].
Together these studies suggest that knock-outs or dysfunction may exist in either the upper somites, the lower somites, or both. An intact SAN in the absence of upper fibers implies survival of neural crest mesenchyme with persistence of all roots of SAN. Loss of the muscle “target” would be expected to lead to loss of motor neurons. When SAN and Tpz are both absent we can see that the neural crest (critical for the development of the peripheral nerves) is also absent. Finally, 17 cases of muscular anomalies concomitant with Tpz aplasia are reviewed by Yiyit, who noted a commonality with Poland syndrome [27, 28]. This suggests two basic mechanisms. (1) Common mesenchymal deficit characterizes anomalies of Tpz with Scm. (2) Common neuroangiosome or neighboring neuroangiomes along the subclavian characterize deficits of Tpz with pectoralis major (thoracoacromial) or with serratus anterior and latissimus dorsi (both thoracodorsal) [29].
Phylogeny of Sternocleidomastoid and Trapezius: The Cucullaris Muscle
The anatomy of sternocleidomastoid and trapezius in mammals is the result of two evolutionary processes: (1) the break-up of a precursor muscle, the cucullaris; and (2) the transformation of a pectoral girdle of dermal bones attached to skull to an independent claviculo-scapular assembly, a subject reviewed previously in this chapter. Understanding the anatomy of the cucullaris and its innervation through evolution is critical. Until recently, the phylogeny of cucullaris in prehistoric fishes (when it appeared and what was its function) was uncertain, as only skeletal remains were available for study. Because sharks are considered a link between ancient gnathostomes and our ancestral bony lungfishes, assumptions about cucullaris have been based on its anatomy in extent chondrichthyans. The recent discovery of placoderm fossils with preserved musculature has provided a radically different model, one offers insights into the evolution of the neck. In addition, new studies using hox gene analysis combined with micro CT scanning demonstrate that cucullaris anatomy and function are dynamic over time, as is its motor innervation. Analysis of these changes provide a powerful window on the evolution of the neck.
The Importance of Placoderms
During the Silurian-Devonian period, eight groups of armored fishes appeared, all of which descended from a common (unknown) basal ancestor. Their emergence marked the great divide between cyclostomes (represented today by extent hagfishes and lampreys) and jawed gnathostomes (all other vertebrates). Cyclostomes (“round mouths”) have a circular oral aperture with no specialized perioral bones. Their branchial apparatus consists of 8 primitive cartilaginous branchial baskets. The “gnathostome revolution” involved the invention of branchial arches, the subsequent transformation of the first two arches into jaws, paired appendages (future limbs), a pectoral girdle, and eventually a neck region. Radical changes in muscles were required in order to control these new moving parts (Figs. 10.92, 10.93, 10.94, 10.95, 10.96).

Placoderms are in direct lineage to mammals as evidenced by four inventions: paired fins, a head–body joint, cucullaris muscle, and transverse abdominal muscles. The articulation and the transverse abdominal muscles temporarily disappear in the course of fish evolution but are reincarnated in tetrapods. [Reprinted from Trinajstic K, Ahlberg, E, et al. Fossil musculature of the most primitive jawed vertebrate. Science 2013; 341(6142):160–164. With permission from The American Association for the Advancement of Science]

Eastmanosteus skull showing the cranial-trunk joint. †Eastmanosteus calliaspis showing dermal neck joint (red ring). This was the original idea for the future craniovertebral joint. The placoderm skull plates have distinct terminology: ADL, anterior dorsolateral plate; AL anterior lateral plate; MD, medial dorsal plate; MV, medial ventral plate; Nu, nuchal plate; PNu, paranuchal plate, PVL, posterior ventrolateral plate. [Reprinted from Trinajstic K, Ahlberg, E, et al. Fossil musculature of the most primitive jawed vertebrate. Science 2013; 341(6142):160–164. With permission from The American Association for the Advancement of Science]

Placoderm musculature. Cucullaris is the precursor of trapezius. Its primary insertion is into ADL, anterior dorsolateral plate. Its secondary insertion is into Pnu, paranuchal plate. [Reprinted from Trinajstic K, Ahlberg, E, et al. Fossil musculature of the most primitive jawed vertebrate. Science 2013; 341(6142):160–164. With permission from The American Association for the Advancement of Science]

Neck and cucullaris evolution based on Trinajstic. Cyclostomes have no jaw and no neck. Agnathans represented by Cephalaspis. Placoderms are considered in the stem group. (a) Head shield of Cephalaspis contains pectoral girdle precursors. Existence of cucullaris unknown. (b) The placoderm pectoral girdle articulates with dermal cranium. (c) Ancestral crown gnathostome pretetrapod has cucullaris attached to the dermal chain of bones. With tetrapods, disconnection of the pectoral girdle from skull happens with loss of upper dermal bones. Cucullaris inserts into chondral scapulocoracoid. Cucullaris persists until mammals when it breaks up into sternocleidomastoid and trapezius. [Reprinted from Kuratani S. A muscular perspective on vetebrate evolution. Science 2013; 341(6142):139–140. With permission from The American Association for the Advancement of Science]

Sharks and sarcopterygians have 7 branchial arches. Cucullaris inserts into the fourth and fifth ceratobranchial cartilages. It is in series with the levatores arcus branciales. [Courtesy of William E. Bemis]
Gnathostomes (“jawed mouth”) are divided into a crown group and a stem group. All members of the crown group, be they living or extinct, have the full set of jawed vertebrate characteristics, although paired appendages are lost in snakes. All members of the stem group are extinct. The most primitive stem gnathostomes, such as ostracoderms, did not have jaws but these earliest fishes did possess pectoral fins and a pectoral girdle fused to a massive head shield. They resembled a giant vacuum cleaner, a sort of armor-plated lamprey well-suited for life on the ocean floor. Notably, ostracoderms did not have transverse abdominal muscles.
Separation of the head and pectoral girdle took place with the appearance of jaws and presumably cucullaris, to control the pectoral girdle, evolved at the same time. At the upper end of stem gnathostomes, the placoderms represent the earliest form of vertebrate anatomy with jaws. These creatures, such as Dunkleosteus had oral bones consisting of plates. In a more advanced iteration, Entelognathus, these plates became premaxilla, maxilla, and mandible. One member of this group, the †Arthrodires (Gr., arthrodes = well-jointed) had a joint between the head and the thoracic plates which allowed to the skull to be raised, increasing the gape of the jaws This vertical pivot function would require evolution of elevator and depressor muscles. When placoderms gave rise to both the chondrichthyans and the osteichthyans the original head-trunk pivot was lost and cucullaris was forced to relocate. Transverse abdominal muscles were lost as well, not to appear again until the advent of tetrapods.
In tetrapods, simultaneous appearance of limbs with transverse abdominal muscles is a non-trivial detail because they are involved in the control of respiration. These muscles have to travel a considerable distance to insert muscles that serve as evolutionary markers concomitant evolution of appendicular muscles and the relationship between the ancient placoderm pivot joint and the tetrapod neck. In both instances, an original muscle plan is “remembered” in evolution. This indicates the importance of studying the placoderm model because this vertebrate form is ancestral to all living tetapods (Fig. 10.92).
Primordial Attachments of Cucullaris Muscle
Ostracoderms, jawless stem gnathostems basal to placoderms had head shields with large plates and no cranio-thoracic pivot joint suggesting (1) that the shoulder girdle of placoderms results from a subdivision of the ostracoderm shield; and (2) cucullaris is not present in ostracoderms but appears for the first time in placoderms. Arthrodires have a regionalized body plan with rigid body armor and scales. The basal condition of cucullaris in placoderms is the control of a constrained pivot joint, implying the need for antagonistic elevator and depressor muscles. Preserved neck and trunk muscles in three species of placoderms were reported by Trinajstic [30] in 2013. In these specimens, elevation of the dermal neck joint was achieved by two muscles: levator capitis major and levator capitis minor. These were antagonized by cucullaris which was found lateral to the joint, connecting the inner surface of the dermal girdle, anterior dorsolateral plate (ADL), with a hollow in the inner surface of the skull roof, paranuchal plate (PNu). Cucullaris in its original form thus acted as a depressor of the skull (Figs. 10.93, 10.94, 10.95).
Although the cranial pivot joint is lost in derived fishes, cucullaris is stably conserved across gnathostomes as a link between head and neck. Cucullaris adapts to varying evolutionary scenarios by seeking out analogous neuromeric insertions. Neuromeric relationships are fundamental. Caecilians are limbless burrowing amphibians lacking a pectoral girdle and gills. In these creatures, cucullaris is found to span dorsal trunk fascia forward to the otic capsule and down to the fascia of ventral trunk muscles at the same level. With the loss of branchial arches in tetrapods the proximal insertion of cucullaris once again returns to skull but the scapula is now disconnected from the skull and mobile. Thus, cucullaris becomes capable of elevating it away from the ventral midline. The mammalian neck is marked by multiple extensors without a specific antagonist. We shall see that the subdivision of cucullaris that results in an independent sternocleidomastoid serve permits it a double function as both flexor and rotator.
Gross Anatomy and Developmental Origins of Cucullaris
Cucullaris is a thin flat muscle, easily disrupted by dissection and difficult to visualize in three dimensions. High-resolution micro-CT scanning offers across a spectrum of gnathostomes demonstrate remarkable uniformity of anatomy [31]. In bony fishes such as the actinopterygian bichir and the sarcopterygian lungfish, cucullaris is referred to as protractor pectoralis. In the lungfish Latimeria it does not reach the dorsal midline and being bounded by epaxial muscles. It spans from the anocleithrum of pectoral girdle forward and inserts into the fifth ceratobranchial cartilage (i.e., branchial arch 7). As such, it is in series with gill levators of the first 4 gill arches. Just anterior to cucullaris the levator of the fourth-gill arch is greatly enlarged, leading to speculation that this muscle should be included as part of cucullaris. This latter is a non-trivial finding. Recall from our model of two neuromeres per arch that the third-gill arch (fifth pharyngeal arch) receives its mesenchyme from neuromeres r10–r11. Therefore fourth and fifth-gill arches in fishes are located at truncal neuromeres sp1–sp4. In mammals, these correspond to levels c3–c4. As we shall see, in mammals the innervation of cucllaris mammals shifts out from hindbrain by two units, becoming completely cervical. Thus, the territory of cucullaris is redefined levels to c1–c6, exactly as we now see it! (Figs. 10.96, 10.97, 10.98).

Cucullaris is the dogfish shark, Squalus. [Courtesy of William E. Bemis]

Cranial muscle anatomy in Coelacanth. Lateral view shows: (1) pectoral girdle (white); (2) branchial arches 3–7: first–fourth ceratobranchial cartilages (yellow) and fifth ceratobranchial cartilage (pink), musculature of levatores archs branchiales (green) and cucullaris (blue). In ventral view, all ceratobranchiales are colored white. Cucullaris is clearly confluent with the gill levators at BA4–BA5. Thus, cucullaris is in register with the fifth and sixth gill arches, spinal neuromeres sp1–sp4. [Reprinted from Sefton EM, Bhullar B-A, Mohades Z, Hanken J. Evolution of the head trunk interface in tetrapod vertebrates. eLife 2016;5:e09972. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by/4.0/]
Where is the source for the mesenchyme of cucullaris? Fate mapping data shows it to arise from unsegmented cranial mesoderm. It appears lateral to the occipital somites and extends backward as far as the fifth, sixth, or seventh arches (depending on the species). Recall that basal fishes have only three occipital somites. The fourth is added at the sarcopterygian–tetrapod transition.
This has given rise to a point of confusion. Based on work in chickens, unsegmented mesoderm fated for cucullaris has been found outside the confines of the somites; thereby some to describe cucullaris as originating within lateral plate mesoderm. This flies in the face of what we know about mesoderm for three reasons: physical, neuromeric, and neuroanatomic.
First: as we know, LPM is the source of smooth muscle, the heart, and cardiovascular system but not conventional striated muscle. Scanning electron microscopy demonstrates that the PAM that contributes to pharyngeal arches is continuous, like a strand of pearls, whether it is in the form of somitomeres or somites.
Second: differences in plasticity exist between branchiomeric and somitic mesoderm. Mesoderm giving rise to branchiomeric muscle, including cucullaris, when transplanted into a more posterior position cannot generate muscle whereas truncal somite muscle transplanted into the head forms pharyngeal arch muscle. This implies two types of genetic programs, one for the head and one for the trunk. The connective tissues of branchiomeric muscles are always neural crest and relate back to the original 7 branchial arch system while connective tissues in somitic muscles are PAM. It has been shown that the CTs of sternocleidomastoid and trapezius are neural crests.
Finally, the motor nuclei supplying cucullaris in anamniotes reside in the nucleus ambiguus; the muscle is innervated by a branch of vagus and by spinal branches as well. In amniotes, motor nuclei for cucullaris reside within the central motor column, continuation of nucleus ambiguus into the cervical spinal cord. In both situations, neural control for cucullaris remains in the same genetic environment regardless of neuromeric level.
We can conclude (and will demonstrate) the following about cucullaris in both its ancestral and derived states:
-
Cucullaris is a branchiomeric muscle throughout evolution.
-
As such, the mesenchyme of cucullaris is unsegmented PAM.
-
The PAM of ancestral cucullaris is lateral to occipital somites S1–S4 and is neuromeric levels r8–r11.
-
The motor supply of cucullaris is located in nucleus ambiguus and/or central motor column.
-
The ancestral nerve for cucullaris joins with caudal vagus nerve but shifts abruptly in birds and mammals out of medulla to spinal cord as spinal accessory nerve.
-
The above change was the result of a frameshift mutation causing a posterior translocation of hox genes coding for cucullaris from rhombomeres into the spinal myelomeres.
-
The mesodermal source for cucullaris moved backwards as well, becoming in register with somites S5–S10, corresponding in mammals to cervical neuromeres c1–c6.
-
In mammals, the cervical spine lengthens two units by a duplication of neuromeres c4–c4. Thus, trapezius gets more mesenchyme. Instead of being a four-somite muscle, the mammalian cucullaris is a six-somite muscle.
-
These changes are integral to the creation of the cranio-vertebral joint.
Phylogeny of Spinal Accessory Nerve
The pectoral fin in stem gnathostomes was welded into the dermal bones of the braincase and was relatively immobile, serving as a stabilizer. Its mesenchyme was synthesized. The advent of a pectoral girdle distinct from the skull in placoderms was accompanied by a change in its neuromeric definition. It was now synthesized from truncal mesenchyme, both as the dermal bone chain and as mesodermal scapulocoracoid. But the muscle that evolved with it, cucullaris, remained branchiomeric; its connective tissue was neural crest from r8 to r11.
The closest we can come to the neuroananatomy of ancestral cucullaris is to study its innervation in living chondrichthyan and sarcopterygian fishes. In selachians (sharks) cucullaris is large and superficial. It is attached to the fascia of epaxial muscles connected to the skull and thence to the most posterior (7th) branchial arch and to the pectoral girdle which it serves to elevate and protract. It is innervated by a branch of vagus, ramus accessories, which is functional, being a special visceral efferent.
Studies in the clear nose skate, Raja eglantera, provide further definition of neuromeric anatomy of cucullaris in the basal state. Barry described a motor column ventrolateral to the dorsal motor nucleus of vagus (PANS) which he termed the ventral motor nucleus of vagus. This column extended into the spinal cord down to level Sp5 giving off motor rootlets. We now identify these structures as homologs of nucleus ambiguus and the accessory nerve. Barry also made an erroneous but prescient speculation: that the ventral nucleus of vagus represents a delamination from the dorsal nucleus…more on the signficance of this a bit later [32].
Skates are closely related to sharks but have a fusion of the proximal vertebrae, the synarcium, which in turn is fused to the pectoral girdle but nevertheless has a cucullaris. These fishes have three dorsal muscles (medial, intermediate, and lateral) which, together, are precursors of what will become trapezius. We can term these the cucullaris complex. This complex has a rostral attachment is to dorsal synarcual process. Its dorsal boundary is the midline epaxial muscle column. Laterally it is confluent with the levatores arcuum branchiales. Posteriorly it is inserted into the rostral suprascapula [33]. The lateral muscle receives an accessory branch from the caudal hindbrain that runs in the substance of the 4th (intestinal branch) of the vagus nerve. Innervation of the intermediate and medial muscles is from spinal roots (Figs. 10.99, 10.100, 10.101).

Cucullaris complex consists of three muscles. The lateral-most one gets direct innervation from caudal ventral motor nucleus of vagus, neuromeres r10–r11. Intermediate and medial muscles are supplied from spinal nerves. Extrapolating from sharks and skates, cucullaris in the basal state of placoderms was a branchiomeric muscle spanning the head-neck joint and supplied by hindbrain. Whether neuromeric territory extended further caudal into the spine is unknown. [Reprinted from Sperry DG, Boord RL. Central location of the motoneurons that supply the cucullaris (trapezius) of the clearnose skate, Raja eglanteria. Brain Res 1991; 582:312–319. With permission from Elsevier]

Insertions of cucllaris into the pectoral girdle in skates. Comparative studies of primitive chondrichtyans give us a clue as to the original anatomy of cucullaris in placoderms. The synarcium in these creatures is a fusion is a fusion of vertebral column to the otic capsule. Scapulocoradoid is fused to synarcium, making it immobile. Lateral muscle (supplied by vagus) and intermediate muscle (supplied by ventral spinal nerves) both insert into scapulocoracoid. Medial muscle inserts solely into suprascapula, a transverse process fused to synarcium and unrelated to the pectoral girdle. From a functional standpoint cucullaris has two components, hindbrain and spinal. In mammals, trapezius arises strictly from cervical mesoderm, eliminating the need for hindbrain innervation. For this reason, there is a disconnect between the root of r10–r11 that join vagus 4 en route to laryngeal muscles and the spinal accessory nerve, sp1–sp6. [Reprinted from Sperry DG, Boord RL. Central location of the motoneurons that supply the cucullaris (trapezius) of the clearnose skate, Raja eglanteria. Brain Res 1991; 582:312–319. With permission from Elsevier]

The spinal accessory nerve is ancestral to all vertebrates. In elasomobranchs. It occupies 6 neuromeres: r10–r11, sp1–sp4. [Reprinted from Sperry DG, Boord RL. Central location of the motoneurons that supply the cucullaris (trapezius) of the clearnose skate, Raja eglanteria. Brain Res 1991; 582:312–319. With permission from Elsevier]
Using horseradish peroxidase (HRP) staining Sperry and Boord showed that motor neurons for cucullaris in Raja originate from caudal nucleus ambiguus, probably from levels r10–r11. None were found more rostrally. Vagus has four major roots. The accessory neurons were traced through the fourth root of X. This root is directed to the fifth arch (fifth gill arch). Accessory neurons are admixed with other motor neurons; these are unlabeled by HRP; in mammals, they are directed toward the muscles of the larynx. The Sperry study did not document contributions to cucullaris from sp1–sp4 and considers cucullaris the lateral muscle only. The broader definition by Marian includes all three muscles of the cucullaris complex, it makes sense that the intermediate and medial muscles represent cucullaris arising from the proximal cervical mesoderm, and innervated by the backward extension of nucleus ambiguus into levels sp1–sp4 [33,34,35].
Thus, the potential innervation of the cucullaris complex from mucleus ambiguus spans a total of six neuromeres across the hindbrain-spinal boundary. This basal gnathostome pattern is universal in anamniota (fishes, amphibians) reptiles and persists in amniote reptiles. This neuroanatomy changes in birds and mammals. Birds have a true accessory nerve that is physically separate from vagus but retains its hindbrain innervation. In mammals, the roots of accessory nerve are completely divorced from the medulla. Due to the hox gene translocation, the definition of what neuromeric levels produce cucullaris muscle shifts backwards from r10–c4 to levels c1–c6. Cucullaris now arsies strictly from cervical mesoderm. This redefines its innervation—the need for hindbrain nuclei is eliminated; accessory nerve becomes strictly spinal.
The break-up of cucullaris into a sternocleidomastoid from c1 to c2 versus trapezius from c4 to c6 is likely related to original genetic differences between mesenchyme originally from r10 to r11 versus that from c1 and beyond. Perhaps sternocleidomastoid represents the lateral muscle mass of the cucullaris complex whereas trapezius, being spinal, is homologous to the intermediate and mediall muscles.
In summation, the cucullaris complex in mammals undergoes a homeotic shift backward two neuromeres to occupy somite levels S5–S10 (c1–c6). The muscle breaks into two units. The split between sternocleidomastoid S5–S6 (c1–c2) and trapezius S7–S10 (c3–c6) reflects the original homeotic “breakpoint” between head and neck, that is, between neuromeres r11 and sp1. It should be noted that the homeotic changes responsible for retrodisplacement of cucullaris in mammals occur much later in evolution than those required for redefinition of cervical muscles for the pectoral girdle and upper limb.
Phylogeny of the Strap Muscles and Prepectoral Muscles
In mammals, the ventral roots of C1–C4 emerge from the medial motor column to face a binary choice. They can take a direct route to innervate muscles associated with the ventral cervical spine such as rectus capitis, longus capitis, or longus colli. They can join up together to form the cervical plexus. In the latter case, they can either supply the hypobranchial strap muscles, the scalenes, or the levator scapulae. The first group of muscles is defined by secondary insertions within the axial spine (or skull). The second group of muscles has secondary insertions outside the axial spine: to pharyngeal arches, to the rib cage, or to the pectoral girdle; they are all outboard to the axial spine.
Hypobranchial muscles as so-named because of their migration below the branchial/pharyngeal arches. In fishes, these muscles originate from the myotomes of the four occipital somites (S1–S4) whereas in tetrapods they are produced from the first four cervical somites as well (S5–S8). Hypobranchial muscles in fishes are a rostral extension of hypaxial muscles supplying the pectoral girdle; they seek out insertions into the floor of the pharynx. Although not branchiomeric, they connect coracoid process with the ventral gill arches and mandible. They act as depressors of the lower jaw.
In chondrichthyans, the hypobranchial muscles are organized into two layers. Posthyoid muscles are referred to as rectus cervicis. These deep consist of (1) coracoarcualis and coracobranchialis that connect to the gill apparatus; and (2) coracohyoid that connects the pectoral girdle with the second/third arch hyoid bone. Prehyoid coracomandibularis is the most superficial layer and runs all the way forward to the lower jaw. Advanced osteichthyans show two innovations. Rectus cervicis gives rise to a rostral extension, sternohyoideus. This makes sense because cartilaginous fishes do not have a sternum. Coracomandibularis subdivides, giving a distinct geniohyoideus (Fig. 10.102).

Hypobranchial muscles in shark, Squalus and mammal, Felix. Hypobranchial muscles in fishes are in three layers. Subdivision into a tongue take place with tetrapods. [Courtesy of William E. Bemis]
Let’s now consider the pectoral muscles controlling the fin. The pectoral girdle in fishes is fixed, these muscles extend outward from the scapulocoracoid, traversing a single joint to insert into the dorsal and ventral aspects of the fin. With exception of cucullaris, fishes lack the prepectoral muscles used by tetrapods to control the scapula (trapezius and levator scapulae). Recall that cucullaris in placoderms crosses the head-trunk pivot joint from paranuchal plate to anterior dorsolateral plate. Recall as well that cucullaris is subverted in crown fishes to become a branchial arch dilator. Its role reappears in tetrapods.
The origin of pectoral muscles in fishes involves four myotomes, M2–M5. M2 and M3 belong to the posterior two occipital somites and are innervated by occipital nerves Oc1 and Oc2. M4 and M5 belong to the first somites of the trunk and are supplied by spinal nerves Sp1 and Sp2. Additional myotomes contribute to the fish fin but can run as far back as sp11. The basal condition of the shoulder muscles becomes more complex in the sarcopterygians, as exemplified by Coelacanth Latimeria. The scapula is not mobile. Thus, the fin is controlled by a deltoid extending from cleithrum to humerus, by an adductor running from cleithrum distally, and by 11 pairs of supinator and pronator muscles (Fig. 10.103).

Dorsal view of shoulder in Latimeria Cucullaris is proximal to cleithrum. Deltoid (Dl) joins cleithrum to humerus. Control of the fin consists of an antagonistic adductor and abductors and a 11 pairs of supinator and pronator muscles. These extend past one joint only, the glenohumeral joint. Cervical plexus muscle mass such as rectus cervicis appears in tetrapods. [Reprinted from Miyake T, et al. Pectoral fin muscles of the Coelacanth Latimeria chalumnae: functional and evolutionary implications for the fin-to-limb transition and subsequent evolution of tetrapods. Anat Rec 2016; 299(9):1203–1223. With permission from John Wiley & Sons]
When pectoral girdle becomes independent the fin rays consolidate and subdivide to make stylopodium, zeugopodium, and autopodium, interconnected with additional joints. The original muscles attached to scapula subdivide. The adductor on the ventral side of the scapula persists as serratus. On the dorsal side of scapula abductor mass splits. The lower part becomes the rhomboids. The upper part becomes the levator scapulae and assumes a new function, elevation of the scapula. Like a telescope, the autopodium expands outward and the new muscles arise by subdivision. They continue to be supplied by anterior (ventral) divisions of cervical nerves.
Fishes do not have tongues. Their hypobranchial muscles are innervated by the four branches of occipital nerves. There is no hypoglossal nerve. The motor neurons for the occipital nerves arise from midline nucleus in r8–r11 that is analogous to the hypoglossal nucleus. It has been shown that primitive pectoral innervation in all fishes originated from both the hindbrain and the spinal cord. Furthermore, Ma et al. found the pattern conserved in sarcopterygian lungfishes in the tetrapod lineage. They hypothesized that a backward shift in Hox expression affecting both neuroepithelium and mesoderm caused muscles to relocate along the rostrocaudal axis along with their neurons. This led to a decoupling of the motor neurons from the hindbrain. Initially, the disconnection was osseous, as in Tiktaalik. with a neuroanatomical translocation as a secondary event [36, 37]. As we shall see, the “tetrapod shift” forced changes in the spinal cord leading to a cervical plexus from MMC and a brachial plexus from a new entity, the LMC.
Anamniote tetrapods, as represented by amphibians, undergo further subdivision of their hypobranchials into two functional groups. These recognize the relocation of the pectoral girdle and the appearance of a new structure, the tongue (although a formal hypoglossal nerve does not appear in evolution until amniota). A prehyoid group arises from sessile myotomes. Coracomendibularis splits into a deep genioglossus and a superficial geniohyoideus that connects the second arch hyoid to first arch mandible. The posthyoid group originates from translocated myotomes. Omoarcuals continiue to connect the coracoid process of pectoral girdle with the gill arches. They are the forerunner of omohyoid. Rectus cervicis spans between the ventral midline of the pectoral girdle and third arch hyoid bone. Cullaris inserts into the dorsal pectoral girdle (cleithrum). Pectoriscapularis runs obliquely downward from the gill region. It is flanked ventrally by rectus cervicis and dorsally by cucullaris. It has two insertions: fascia dorsalis of the epaxial dorsalis trunci muscles and ventral pectoral girdle (clavicle). Pectoriscapularis is the anterior-most muscle of the pectoral girdle and inserts into clavicle, pulling the scapula upwards. As such, it is neuromerically caudal to its confrères and may be the sole muscle corresponding to level c4. Amniote evolution involves increasing complexity of the tongue supplied by a coalescence of the occipital nerves into a single hypoglossal nerve. In mammals the intrinsic lingualis muscles of the tongue result from a delamination genioglossus, thereby explaining their location in the center of the tongue. Geniohyoid produces hyoglossus and styloglosssus.
Between pre-mammal anapsids such as the pelycosaures †Dimitrodon and mammals, the two remaining dermal bones rostral to scapula undergo translocation. Dorsal cleithrum migrates to the back of scapula, taking trapezius with it. Ventral clavicle dissociates from cleithrum and moves to the extreme medial/ventral corner of scapula; causing a similar dislocation of sternocleidomastoid. Sternohyoid and sternothyroid follow the dissociation of anterior coracoid process in the development of manubrium (Figs. 10.104, 10.105).

Phylogeny of hypobranchial muscles. Basal proximal insertion of muscles into pectoral girdle is retained. Relation to branchial arches reduced in amphibians (mudpuppy). Sternum appears in amniotes with manubrium from the anterior coracoid process. [Courtesy of William E. Bemis]

Dermal bones of the pectoral girdle bear insertions of hypobranchial and shoulder muscles. Into clavicle are inserted coraohyoid (green) and pectoriscapularis (tan). Cleithrum has cucullaris (the future trapezius). When cleithrum translocates to the back of the scapula it brings trapezius along with it. [Courtesy of William E. Bemis]
What relationship exists between the strap muscles and the muscles of the shoulder? Shoulder muscles can be considered in three groups. Axioscapular muscles (trapezius, levator scapulae, rhomboids, and serratus) position the scapula, and therefore the glenohumeral joint, correctly in space. Scapulohumeral muscles (deltoid, supra/infraspinatus, and teres major) control the position of humerus within the glenohumeral joint. Axiohumeral muscles (pectoralis major, subscapularis, and latissimus) position humerus independently from the scapular. Our focus here is solely on the first two axioscapular muscles.
The boundary between cervical plexus and brachial plexus in amniotes lies at neuromere c4. The levator scapulae (C3–C4) insert into medial border of scapula above (rostral to) to the spine and are supplied by cervical plexus. The rhomboids insert into medial border of scapula below (caudal to) the dorsal side (C4–C5) belong to brachial plexus. Note that scapular spine defines the breakpoint. It is the sole component of scapula derived from dermal bone. With the exception of the medial border rostral to the spine and the scapular spine itself, the entire muscle mass of scapula is supplied by c5 and below, that is, from brachial plexus. The independence of acromion is seen as a separate chondral ossification center and even as a separate oa acromiale. For this reason, it constitutes a breakpoint between the primary insertions of trapezius directed to the skull and deltoid, directed to humerus (Fig. 10.106).

Evolution of pectoral muscles. In fishes the muscles extend past the joint to insert into the distal appendage. Tetrapods have secondary insertions into the proximal stylopodium (humerus) with a second tier of muscles extending to the zeugopodium (ulna/ventral and radius/dorsal). This activates. Dorsal muscles (dark green); ventral muscles (light green). [Courtesy of William E. Bemis]
In summation, axioscapularis muscles do not change in antero-posterior position with evolution. They control the fins in fishes and the scapula in tetrapods. For this reason, their motor nerves, although described as part of either cervical plexus or brachial plexus, in reality are just garden-variety ventral roots. Strap muscles, on the other hand, result from a homeotic shift backwards causing the cervical somites to express additional programs hypobranchial muscles “stolen” from the occipital somites. These “posteriorized” hypobranchial muscles are innervated by the medial motor column of the cervical spinal cord. The extent of the shift varies with distribution of Pax6. Recall that in fishes Pax6 spans from r8 to sp5. In the homeotic shift the definition of Pax6 is pushed back from sp1 to sp9. In mammals, this corresponds to c1–t1. Thus, the prepectoral girdle musculature of the straps extends from c1 to c3, levator scapulae are supplied from c3 to c4, the diaphragm is at c4 and brachial plexus spans from c4 to t1. The appearance of the phrenic motor column, spinal accessory motor column, and lateral motor column in the cervical spinal cord reflects the evolutionary overlap of these target muscles.
Phylogeny of the Cervical Plexus
The basal form of the peripheral nervous system is seen in cyclostomes (lamprey) in which the homologs of ventral and dorsal nerve roots exist as distinct spinal nerves. Ventral nerves are segmental and somatic motor. They directly enter the myomeres. Dorsal roots are intersegmental; they contain both motor and visceral sensory neurons. They pass between myomeres and extend outward to the skin and inward toward the gut. Note: visceral motor neurons gain access to the viscera in lampreys via cranial nerves, especially the vagus [38] (Fig. 10.107).

Phylogeny of motor neurons in ventral horn (VH) of spinal cord. In lampreys, MMC-like motor neurons (Mn) are in the ventral horn but no distinction is made between dorsal and ventral. Fishes do not have motor columns in teleosts such as zebrafish and the sea robin Prionotus carolinus, which possess movable fin rays, are segregated into discrete pools for dorsal and ventral muscles. In the lineage of tetrapods, including amphibians, reptiles, and mammals, motor neurons located in the lateral portion of the ventral horn segregate into the LMC and innervate limbs. Motor neurons in the python snake form a single continuous MMCcolumn and lack the LMC. ASL, accessory spinal lobes; DH, dorsal horn; DMn, dorsal motor neuron; MMC, medial motor column; LMC, lateral motor column; VH, ventral horn; VMn, ventral motor neuron [Reprinted from Murakami Y, Tanaka M. Evolution of motor innervation to vertebrate fins and limbs. Dev Biol 2011; 355(1):164–172. With permission from Elsevier]
This system changes in hagfishes forward throughout vertebrate evolution as ventral and dorsal nerve roots attached to the spinal cord at a given neuromere. Furthermore, somatic motor fibers and visceral sensory fibers (both somatic and visceral) now separate out into the ventral and dorsal roots. Evolution of motor neurons is slower. Anamniotes use both roots while amniotes concentrate all visceral motor fibers into the ventral nerve root. The territory of nerves serving the pectoral girdle and upper extremity is Hox related. In mammals, Hox5 defines 8 cervical neuromeres and Hox6 extends from first thoracic neuromere to the first lumbar neuromere.
In lampreys, motor neurons for the upper extremity are located in the ventral horn but without spatial orientation. Actinopterygian (ray-finned) fishes have a forefin controlled by a very simple system of dorsal extensors and ventral flexors attached to an immobile scapulocoracoid. Although they do not form motor columns they demonstrate segregation of the dorsal and ventral neurons. Their pectoral muscles have proximal attachment within the body Sarcoptyergian (fleshy finned) fishes are so-named because their muscles extend outward from the body into the “limb.”
Fish fins are rather simplistic. Using a tail for propulsion the fins serve as stabilizers. These provide maneuverability and braking. In combination with the lateral line sensory system (previously discussed), the fins enable sudden, darting motions ideal for prey capture or escape. The musculature is very simple. Greater complexity is achieved by subdivision of the dorsal/ventral muscle masses.
In contrast to the simple paddle-like structure of the fin articulating from a fixed pectoral girdle, tetrapods have a mobile pectoral girdle and a three-tiered limb (stylopodium, zeugopodium, and chiropodium). The system thus advances from one joint to four joints with a consequent increase in mobility and greater complexity of muscle control. Note that proximal control of the scapula involves the subdivision of existing muscles connecting dorsal and ventral scapula to the body. Control from scapula to limb requires more elaborate set of divisions and reassignments. Innervation of this greater muscle mass is accomplished by creation of a separate lateral motor column. In mammals this extends from neuromeric levels c4 to t1.
Innervation of pectoral muscles in fish fins: relationship to the evolution of the neck.
Fishes have three somites. The first somite does not produce muscle; therefore hypobranchial muscles arise one somite backward, from S2 to S3, and are innervated by occipital nerves Oc1 and Oc2. The basal plan for muscles controlling the pectoral fin in all bony fishes involves 4 myotomes from the occipital somites S2–S3 and the first two somites of the trunk S4–S5. Let’s refer to these myotomes as M1–M4. Recall that in the tetrapod transition a fourth somite is added to the braincase [37] (Fig. 10.108).

Location of motor neurons to pectoral fin muscles in crown bony (actinopterygian) fishes. Occipital motor neurons (gray) innervate muscles of the second and third myotome producing the hypobranchal anlage (e.g., coracomandibularis). Pectoral motoneurons (red) form a single column extending from hlndbrain into at least the first two neuromeres of the spinal cord. Because the neurons are admixed in the spinal cord with standard hyaxial motor neuron to the muscles of the ventral they are functionally like MMC. [Reprinted from Ma L-H, Gilland E, Bass AH, Baker R. Ancestry of motor innervation to pectoral fin and forelimb. Nature Communications 2010; 1(49) doi: 10.10.38/ncomms1045. With permission from Springer Nature]
Pectoral motorneurons of all fishes arise from both the hindbrain and spinal cord, rather than solely from spinal cord. Mapping experiments demonstrate that this dual pattern in conserved all the way from basal forms such as Polydon spathula (paddlefish) through advanced teleosts, the midshipman Porichthys notatus. In the basal piscine pattern two occipital nerves (Oc1 and Oc2) emerging than an occipital foramen (OcF) just anterior to the craniao-vertebral junction to supply hypobranchial muscles. They also join with two spinal nerves sp1 and sp2 to supply the pectoral fin.
Lungfishes add a fourth occipital somite. They therefore have three occipital motor nerves. Oc1 and Oc 2 from somites S2 and S3 form a distinct hypobranchial nerve to the coracomandibularis-in-transition. Oc3 immediately joins Sp1–Sp3 to form the pectoral nerve. With additional complexity, fishes can have up to 11–13 spinal nerves contributing to the forefin. For our purposes, what is important is the basic tetrad (Figs. 10.109, 10.110, 10.111).

Pectoral fin innervation in crown actinopterygian Midshipman fish. Left: hindbrain/spinal cord boundary (hatched yellow line) OcF-occipital foramen. Right: Motor control from two occipital nerves, Oc1–Oc2 and three spinal nerves, Sp1–Sp3. [Reprinted from Ma L-H, Gilland E, Bass AH, Baker R. Ancestry of motor innervation to pectoral fin and forelimb. Nature Communications 2010; 1(49) doi: 10.10.38/ncomms1045. With permission from Springer Nature]

Pectoral innervation in Dipnoi (lungfish). We can translate the same analysis from actinoptyergian to sarcopterygian fishes. Sarcopterygians have a fourth occipital somite and consequently 3 occipital nerves. Hindbarin/spinal cord boundary is between Oc3 and Sp1. Note appearance of a distinct hypobranchial nerve Oc1–Oc2 representing somites S2–S3. Oc3 joins with Sp1–Sp3 to supply the pectoral muscles. In tetrapods, when Oc3 is displaced backwards from S4 into S5, the cervical spinal cord will have in the medial motor column four motor roots corresponding to the upper extremity. [Reprinted from Ma L-H, Gilland E, Bass AH, Baker R. Ancestry of motor innervation to pectoral fin and forelimb. Nature Communications 2010; 1(49) doi: 10.10.38/ncomms1045. With permission from Springer Nature]

Diaphragm component parts. [Reprinted from Gilbert SF, Barressi MJF. Developmental Biology, 11th ed. Sunderland, MA: Sinauer; 2016. Copyright © 2016. Oxford Publishing Limited. Reproduced with permission of the Licensor through PLSclear]
Let’s introduce a definition. Prepectoral muscles are those that connect the pectoral girdle to the body. Their motor nuclei (neuromeres sp to sp4) arise from two motor columns. MMC via cervical plexus supplies the strap muscles and levator scapulae. SAC via spinal accessory nerve supplies trapezius. Postpectoral muscle are all those connecting either scapula or body wall with the upper extremity. They are supplied by a brachial plexus which, in non-mammalian amniotes, consists of four roots. The more complex mammalian brachial plexus has six roots (vide infra). The brachial plexus is physically separate from cervical plexus. It is neurologically requiring a new, autonomous motor column, the LMC [39].
Distinct motor columns do not appear in evolution until tetrapods. This caudal shift in the location of pectoral plexus and thus its motor nuclei (or vice versa) took place at the sarcopterygian–tetrapod transition, probably with Tiktaalik. With the creation of the neck, all four roots of the original pectoral nerve are now outside the skull. In this transition the expression of the original occipital muscles, they are split into the prehyoid lingual group which continue to be supplied from medulla and a posthyoid cervical group.
The function of occipital nerves changed as well. There were now four occipital somites and four-root hypoglossal nerve dedicated solely to the tongue. Spinal nerves sp1 and sp2 assumed a new role. Not only were they responsible for the axial musculature but they became the supply of hypobranchial muscles formerly innervated by Oc1 and Oc2. MMC from sp1 to sp4 is thus an admixture: the usual motor neurons for axial muscles plus retrodisplaced hypobranchial motor neurons—the future cervical plexus.
It is tempting to speculate what the consequences of this neuromeric shift were for the cervical neuromeres. Recall that in fishes neuromeres caudal to sp2 are involved in the fin field. With the “loading up” of c1–c3 with hypobranchial muscles did this push the program for a brachial plexus backward to c4–c5–c6–t1? Did this necessitate the creation of an additional lateral motor column?
Our knowledge of the cervical region in the basal tetrapods is very limited. We can surmise the situation by looking at extent amphibians. Amphibians have no atlas–axis. Frogs have only three cervical vertebrae. Their diminutive forelimbs are supplied by a rudimentary brachial plexus consisting of two spinal nerves, sp2 and sp3. Salamanders have four cervical vertebrae with a brachial plexus from root sp3 and sp4. We do know that in early tetrapods the position of the pectoral girdle was immediately behind the head. Presumably, the rectus cervis muscles are supplied by at least one spinal nerve.
Amniotes are more sophisticated. Buccal pumping was abandoned. The circulatory system was remodeled. The neck expanded to five cervical vertebrae in chameleons and subsequently to six in pre-reptiles. As we shall see, the amniote pre-reptilian brachial plexus has four roots with the final root always positioned between the last cervical vertebra and the first thoracic vertebra, thereby indicating the location of the pectoral girdle. When the rostral limit of the brachial plexus reaches neuromeric level c4 we see the first signs of a true three-root cervical plexus.
Pectoral Girdle Muscle Displaced into Thorax: (PMC = CMC Medial)
Diaphragm
We conclude our exploration of cervical myology with the diaphragm, the third muscle of the central motor column, and one unique to mammals. Its evolution is intimately involved with the expansion of the mammalian neck and involves the transposition of the brachial plexus [40] (Fig. 10.111).
Mammals have a high metabolic rate permitting them to function at a wide range of temperatures. This required highly efficient multi-lobulated lungs with a large surface area but with the drawback of low compliance. To ventilate the lung, mammals invented a unique muscle, the diaphragm, capable of overcoming the compliance issue. The diaphragm stems from muscles “originating” from vertebrae and the sternum at the boundary of the thorax and abdomen. Why should these muscle fibers be innervated by the C3–C5? What could explain myoblast migration into this site? To appreciate these questions, let’s look at the structure of the diaphragm, blood supply and innervation, and development [41].
Structure
The diaphragm is a complex musculotendinous structure constructed from four components, each from a different tissue source. Significance.
-
Septum transversum is the most significant as it makes central tendon. It lies ventral.
-
Pleuroperitoneal folds (PPFs) fuse with two structures: dorsal mesentery of esophagus and dorsal part of septum transversum. Together, these form the primitive diaphragm. The membranes are dorso-lateral and constitute the portal of entry of cervical myoblasts.
-
Dorsal mesentery of the esophagus (mesoesphagus) is an extension of lateral plate mesoderm hanging down into the abdominal cavity. It fuses with both septum transversum and pleuroperitoneal membranes. It forms the midline part of diaphragm. LPM smooth muscle surrounding the esophagus invades the dorsal mesentery to produce right and left crura. These are dorsal.
-
Body wall gives off an internally-directed shelf of mesenchyme which forms a semi-circumferential arcade peripheral to pleuroperitoneal membranes.
Nota bene: The classical embryologic model above is under challenge. Animal models of congenital diaphragmatic hernia induced by the teratogen nitrofen demonstrate that the PPFs constitute the major (and perhaps the sole) contributor of muscle to diaphragm [42].
Blood supply consists that two major and two minor sources.
-
Phrenic arise paired or singular directly from the aorta or a common branch from the coeliac trunk. They are embryologically associated with the foregut. The phrenic arteries supply the central diaphragm. At the posterior border of central tendon they divide. Medial branch runs anteriorly to joint with pericardiophrenic and the musculophrenic from the periphery. Lateral branch travels outward to anastomose with the intercostal arteries.
-
Posterior intercostal/subcostal arteries from two sources. Those for the first and second interspaces come from costocervical trunk. Those arteries supplying interspaces 3–12 arise directly from the aorta. Posterior intercostals associated with the lower 6 ribs supply the body wall component of the diaphragm.
-
Pericardiophrenic arteries are small vessels that arise from the internal thoracic arteries. As such, they arise from longitudinal anastomoses of transverse segmental vessels and accompany the phrenic nerves. The neurovascular pedicles pass between the pleura and pericardium, supplying the latter.
-
Musculophrenic artery is one of two terminal branches of internal thoracic, the other being superior epigastric. Both branches are anterior. Prior to its division, internal thoracic gives off anterior intercostal arteries to the upper 6 intercostal spaces. It then divides, with musculophrenic running behind the costal cartilages to supply the lower 6 intercostal spaces.
Phrenic nerves descend into the chest from C4 with minor contributions from C3 and C5. The right phrenic is forced to deviate by the presence of anterior scalene which excludes it from contact with the second segment of subclavian. Left phrenic runs a direct course and passes anteriorly to first segment of subclavian. In the thoracic cavity the phrenic nerves, accompanied by pericardiophrenic arteries, pass anterior to hilum sandwiched between fibrous pericardium and the pleura of mediastinum. Here they pick up blood supply from internal thoracic artery as its pericardiophrenic branches. Once the phrenic nerves gain access to the superior surface of diaphragm they divide into four branches. Sternal branch runs anteriorly. Anterolateral branch runs in front of the lateral lamina or leaf of central tendon. Posterolateral branch runs behind the lateral lamina. Crural branch runs posteriorly to the crura.
Neuroangiosome failure at boundary zones between neuroangiosomes correlates with diaphragmatic hernias. Hernia at the foramen of Morgagni is anterior, anterolateral defects can be large, permitting massive herniation into the chest. The Bochdalek hernia is posterolateral (Fig. 10.112).

Diaphragm: normal anatomy and sites of congenital defects. Sites correspond to watershed zones of neuroangiosomes. Pleuroperitoneal folds bear the bulk of diaphragmatic myoblasts. CDH sites are located at the border of the folds with the septum transversum and the LPM musculature surrounding the esophagus. Eventration is not a true hernia but represents upward displacement by the viscera due to hypoplasia of the scaffold. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Pain sensation is relayed in three ways. Direct transmission from the diaphragm travels via the phrenics and refers to the dorsolateral shoulder. Inflammation at the periphery of the diaphragm can be pick up by the lower six posterior intercostals and perceived as pleuritic pain. Finally, small penetrating branches from the phrenics can detect subdiaphragmatic inflammation, on the right side from liver, and on the left side, from left lobe of liver, stomach, left adrenal, and spleen. Thus, splenic injury can refer to the left shoulder.
Congenital Defects
Diaphragmatic hernias are a well-described problem in pediatric surgery with numerous reviews available. There are three forms. Bochdalek hernia is most common (95%). It is posterolateral with left-side predominance (85%). This preference has been ascribed to a protective effect of the liver on the right side. Complete absence of the diaphragm has been reported [43]. The Morgagni hernia represents only 2% of reported cases. It is retrosternal, just behind xiphoid process. Both these defects are located along the border of septum transversum. Eventration of the diaphragm Is not a true hernia. It is caused by hypoplasia of the scaffold leading to an upward displacement of the diaphragm by the abdominal contents. Eventration has been reported in association with Poland syndrome [44,45,46,47] (Figs. 10.113, 10.114, 10.115, 10.116).

Diaphragmatic hernia. Morgani is retrosternal; it is much less common, CDH, when large, is termed eventration of the diaphragm. It involves territory anterior to the foramen of Bochdalek. Left: [Reprinted from Wikimedia. Retrieved from: https://en.wikipedia.org/wiki/Congenital_diaphragmatic_hernia#/media/File:Morgagni_Hernia.PNG. With permission from Creative Commons License 4.0: https://creativecommons.org/licenses/by-sa/4.0/deed.en.] Right: [Reprinted from Fisher JC, Bodenstein L Computer simulation analysis of normal and abnormal development of the mammalian diaphragm. Theor Biol Med Model: 2006;3:9. With permission from Creative Commons License 2.0: http://creativecommons.org/licenses/by/2.0]

Bochdalek hernia, senso strictu, refers to small posterolateral defects on the left side. A Plain x-ray showing bowel loops in the left chest. B Loops of small bowl entering the chest through the defect. C Defect after reduction of the bowel. D Autopsy specimen showing hypoplastia of the lungs, left > right [Reprinted from Tovar JA Congenital diaphragmatic hernia. Orphanet J Rare Dis: 2012, 7;1. With permission from Creative Commons License 2.0: http://creativecommons.org/licenses/by/2.0]

Eventration of the diaphragm represents thinning out due to absence of myobplasts. It can be isolated or can occur as part of Poland syndrome. Left: [Reprinted from Ravisagar P, Abhinav S, Matthur RM, Anula S. Eventration of the diaphragm presenting as recurrent respiratory tract infetions—a case report. Egyptian Journal of Chest Diseases and Tuberculosis 2015; 64:291–293. With permission from Elsevier.] Right: [Reprinted from Kulkarni MI, Sneharoopa B, Vani HN, Nawaz S, Kannan B, Kulkarni PM. Eventration of the diaphragm and associations. Indian J Ped 2007; 74(2):202–205. With permission from Springer Nature]

Septum transversum (gray) is a semicircular shelf that grows from ventral body walls to ventral foregut (yellow), dividing the heart from the liver. It partially cuts apart the primary coelom, but paired pleuropericardial canals (tan) persist dosal to septum transversum. The lungs, as foregut diverticula, will invade the canals. [Reprinted from Carlson BM. Human Embryology and Developmental Biology, sixth edition. St. Louis, MO: Elsevier; 2019. With permission from Elsevier]
Development of the Diaphragm
The conceptual structure of the diaphragm consists of the creation of a platform or scaffold which subsequently is populated by myoblasts and migrates into position. Despite its position at neuromeric level t10, the diaphragm is not derived from the thorax. It is not commonly appreciated that the scaffold arises at the level of the occipital somites. During its descent, it acquires myoblasts from the myotomes of C3–C5.
Topologically the components of the diaphragm fit together l(dorsa-ventral) like the letter “T” with septum transversum as a vertical post in the midline and the pleuroperitonesal folds like the crossbar. Its three-dimensional relations with the heart, foregut, and pleuroperitoneal canals are depicted in Fig. 10.116. Lateral to the passage way between chest and lung, nephric folds run longitudinally down the embryo. These are thought to contribute to the pleuroperitoneal folds.
Cervical somite myoblasts from c3 to t1 form a common anlage. Pre-diaphragmatic myoblasts innervated from the PMC segregate out and inter the pleuroperitoneal folds and thence septum transversum, bringing their innervation from c4 to c5 as phrenic nerve.
In mammals, the intracoelomic scaffold becomes populated with non-cervical somite local myoblasts throughout. This thoracic mesenchyme gives the diaphragm some additional substance but whether or not they pick up innervation from phrenic nerve is unclear. The origin of this somatic lateral plate mesenchyme is from the lateral thoracic wall. It forms a semicircular arcade wrapped completely around the pleuroperitoneal folds and partially around the septum transversum. The somatic innervation of this zone is from intercostal nerves; whether it receives phrenic nerve innervation is unclear.
Muscles that migrate great distances such as those of the tongue and extremities can be traced by means of Pax3 expression. Extrapolations from rats using this experimental method shows give us the following developmental sequence in Carnegie stages [48, 49] (Fig. 10.117).
-
Stage 8 At 22–23 days the heart has folded beneath the pharyngeal arches.
-
Stage 10 Extrapolating from rat model, the muscle precursors of the diaphragm delaminate from somites in Carnegie stage 10, at which time S1–S10 are fully functional, that is, down to the sixth cervical somite (C2). Mesenchyme from the caudal part of the pericardium extends downward and spans between the ventrolateral body wall and the foregut.
-
Stage 11 Motor axons enter the cervical anlage.
-
Stage 13 upper limb buds appear. Septum transversum has descended from C2 to C4. It Is penetrated at its superior aspect by cervical myoblasts from C3 to C4–C5. Pleuroperitoneal folds (PPFs) project into coelom and migratory cells track along the brachial plexus but are still within the body wall. They have not yet entered PPFs or the limb bud.
-
Stage 14 Myoblasts enter the PPFs and limb bud. Liver tissue invades the inferior aspect of septum transversum.
-
Stage 15 Phrenic nerve axons enter the PPFs and innervate the myoblasts. Diaphragm begins to translocate caudally.
-
Stage 16 The lung bud is growing downward surrounded by body wall and with the liver beneath. Because septum transversum is primarily a ventral structure, dorsal to it are the pleuropericardial canals, mesonephric ridges of intermediate mesoderm, and future suprarenal glands. Expanding pleural cavities will burrow into this mesenchyme dorsally. In so doing they will strip LPMS mesenchyme away from the dorsal body wall. This will be the future source for the peripheral diaphragm.
-
Stage 17 Muscle forms in the diaphragm. Phrenic nerve trifurcates. Pleuroperitoneal folds fuse to septum transversum and diaphragm closes.
-
Stages 21–23 are marked by closure of the pleuroperitoneal canals under pressure from surrounding tissues (liver and adrenals). Diaphragm reaches its final position at stage 23.

Embryonic development of the diaphragm. (a) Sequence of development: E11.5 = Carnegie stage 11; E12.5 = Carnegie stage 13; E13.5 = Carnegie stage 15; E14.5 = Carnegie stage 17; E15 = Carnegie stage 18. (b) Sagittal section of a mouse embryo at an early stage of phrenic axon elongation, (c), at a later stage of phrenic axon elongation, and (d) at an early stage of phrenic nerve branching. Dotted area indicates a cell population of migratory muscle precursors (MMPs) migrating into PPF. ant ca ve, anterior cardinal vein; du cu, ductus cuvieri; eso, esophagus; liv, liver; lu b, lung bud; perc cav, pericardiac cavity; phr, phrenic nerve; po ca ve, posterior cardinal vein; s tr, transverse septum. [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
Phylogeny of the Diaphragm: An Unrecognized Brachial Plexus Muscle
The mammalian diaphragm originates from a subpopulation of cells destined in pre-mammal synapsids to become supracoracoideus, connecting pectoral girdle to the humerus. The migration of this muscle is arrested. It remains within the body wall but retains its motor supply from neuromere c4. The evolution of this muscle can be understood on the basis of its motor innervation (Fig. 10.118).

Evolution of the pectoral girdle—leading to the diaphragm. Diaphragm originates from a subpopulation of cells that migrate toward brachial plexus yet remain within the body wall. In basal tetrapods, a common muscle mass, supplied from the first two spinal nerves br1–br2, of the brachial plexus, connects pectoral girdle to humerus. In amniotes br1 and br2 are located at C4–C5. The muscle anatomy diverges. In basal amniotes it splits into supracoracoscapularis/supracoracoideus and subscapularis. In diapsids (birds) supracoracoideus persists as the antagonist to pectoralis to elevate the wing or stabilize the upper extremity. Birds do not make a diaphragm. In synapsids (mammals) the original coracoid process is lost and is reincarnated as spine of scapula. The Pectoralis major/minor remains attached to the chest, but C4–C5 subscapularis fragments (1) part remains in situ, (2) a c5 part relocates dorsally on either side of the scapular spine as supaspinatus and infraspinatus, and (3) a C4 part attached to chest wall becomes internalized, losing its connection with pectoral girdle to become the diaphragm. Left: [Reprinted from https://commons.wikimedia.org/wiki/File:Wing_Muscles,_color.svg. With permission from Creative Commons License 2.5: https://creativecommons.org/licenses/by-sa/2.5/deed.en.] Right: [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones]
Recall from our previous discussion of the cervical spine that the common ancestor of all amniotes had five cervical vertebrae and that the number of cervical vertebrae increases in each lineage. On the diapsid side, †dinosaurs had 11 to 19 (or more), birds go from 8 to 20, and crown reptiles have 8. On the synapsid line, non-therapsids had five, therapsids had six, with the final number settling at seven with cynodonts and eventually mammals (Fig. 10.119).

Evolution of cervical vertebrae in amniotes. The two crown clades are diapsida and synapsida. Diapid taxa are the lepidosaurs (modern reptiles) and archosaurs (dinosaurs and birds). Synapsids are mammals. Extant (living) taxa are in bold. [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
In all basal tetrapods, control of the upper extremity is accomplished by common muscle mass, divided into flexors and extensors and usually supplied by four spinal nerves br1–br4, the first three are always cervical and the last one thoracic. Thus, the non-mammals tetrapod formula is CX–CY–CZ–T1. Where brachial plexus begins, that is, the location of br1, varies with evolution. In lizards, br1 can be found at C3, in basal amniotes br1 is at C4. Advanced amniotes have longer necks. In crown reptiles, br1 is at C7 and in the ostrich, the plexus begins at C18…excellent for burying one’s head in the sand!
Let’s compare the situation in the chicken that that of mammals. In Gallus the 4-root brachial plexus extends from sp13–sp16 to the cervical–thoracic junction between vertebrae C15 and T1. Roots br1 and br2 give rise to nerves to the supracoracoid and subscapularis muscles. The same pattern is seen in the basal amniote plexus (Fig. 10.120a).

Caudal transition of the brachial plexus. (a) Basic pattern of the brachial plexus of basal amniotes, based on the inference from extant reptiles and birds. (b) Basic pattern of the brachial plexus of mammals. br1–br4, first through fourth spinal nerves for the basic pattern of amniote brachial plexus. KEY: br1′ and br2′, remnants of the first and second, respectively, spinal nerves of the ancestral brachial plexus; brl, N. brachialis longus; C4–C8, T1 cervical spinal nerves 4–8 and thoracic spinal nerve 1; med, median nerve; phr, phrenic nerve; rad, radial nerve; sbcs, subcoraco-scapularis nerve; sbs, subscapularis nerve; spc, supracoracoideus nerve; sps, suprascapularis nerve; uln, ulnar nerve; X, communication between br2′ and br2; Y, communication between br2′ and br3–br2. Lateral nerves marked by circles; medial nerves marked by boxes. Keep in mind these homologies between amniotes and mammals: subcorascapularis = subscapularis and supracoracoidus – suprascapularis. [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
The mammalian plexus is more complex, with the original four roots shifted backward to the sixth–ninth spinal nerves, that is, C6–T1. On the other hand, suprascapular does not change position; it remains behind C5. This implies that mammals achieve the configuration of eight cervical neuromeres and seven cervical vertebrae by virtue of a partial duplication in the fourth and fifth spinal nerves. Mammals are thus unique in having six spinal nerves by virtue of duplicating br1–br2. They also have an additional nerve ord, the median nerve. This more sophisticated system enables enhanced motor control of the upper extremity with the formula CX–CY–CX–CY–CZ–T1. In sum, the true mammalian brachial plexus is C4–C5–C6–C7–C8–T1 (Fig. 10.120b).
The phrenic nerve can be understood as a remnant of the ancestral brachial plexus. The suprascapular nerve in mammals, by remaining a C5, retains its original amniote identity. In contrast, subscapular nerve moves backwards two neuromeres. Recall that subscapular or subcoracoscapular arise from br1 to br2 in the original amniote pattern at level C5. We now find them in mammals at level C7. The myoblasts of subscapularis arise from the cranial part of the muscle mass dedicated to the forelimb. In pelycosaurs, as in mammals, it attaches scapula to chest wall. In this regard, the myoblasts of diaphragm, abandoned back at levels C4–C5, pursue a similar course but penetrate the chest to enter the pleuroperitoneal folds. In the process, they lose their proximal attachment to the pectoral girdle but their innervation is retained! (Fig. 10.121).

†Dimetrodon and the basal diaphragm. In synapsid pelycosaur, Dimetrodon† show five cervical vertebra. Heart is just medial to pectoral girdle. Subscapular muscle (sbsc) becomes inserted into thoracic wall directly opposite the potential pleuroperitoneal folds. Proximalhoulder muscle derivatives enter coelomic cavity at arrow, populate the folds and bring cervical innervation with them. Left: [Reprinted from Wikimedia. Retrieved from: https://commons.wikimedia.org/wiki/File:Dimetrodon_incisivum_01.jpg. With permission from Creative Commons License 3.0: https://creativecommons.org/licenses/by-sa/3.0/deed.en] Right: [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons.]
Phylogeny of the Diaphragm
The muscle mass that will give rise to the diaphragm originates from the first two cervical somites of the plexus (C4–C5). Its spinal nerves br1–br2 connect the pectoral girdle to humerus.
In basal amniotes, the anapsids, br1 and br2 shift backward from the original tetrapod position becoming located at C4–C5. The muscle anatomy diverges into an extensor of the humerus, supracoracoscapularis/supracoracoideus, and a flexor, the pectoral mass called subscapularis. In diapsids, supracoracoideus persists as the antagonist to pectoralis to elevate the wing in birds and stabilize the upper extremity in reptiles. Note that birds do not make a diaphragm. They suffocate if the chest is compressed.
In synapsids (pre-mammals) the original coracoid process is lost and is reincarnated as spine of scapula. The muscle complex fragments. C5–T1 remains attached to the chest as pectoralis major and minor. C4–C5 relocates to the scapula, becoming worthy of its name. In mammals, the original coracoid process becomes the scapular spine. Subscapular splits. Half of the muscle remains in situ on the ventral aspect of scapula. The other half shifts dorsally to form, on either side of the spine, supaspinatus, and infraspinatus. Finally, a C4 portion migrates into the pleuroperitoneal folds to become a diaphragm, bringing its cervical nerves with it.
Let’s describe how this happens. In the basal synapsid pelycosaurs and pre-mammals subscapular muscle spans across the internal aspect of scapula (Fig. 10.121). It is located in the body wall just behind the heart, facing the coelomic cavity just opposite the pleuroperitoneal fold. This muscle mass is termed the proto-diaphragm. In the Hirasawa and Kuratani model, the driver for diaphragm development is the caudal shift of the forelimb bud. Seven-vertebra status was completed with the cynodonts in the Triassic, as in Thrinaxodon†. Sealing up the diaphragm closed down the aperture for the esophagus (a potential problem for large chunks of food). This did not present a problem as Thrinaxodon was a heterodont (having specialized teeth) and had a secondary hard palate indicating advanced food processing. The pre-cynodont pelycosaur myoblast population positioned beneath scapula (including primitive diaphragm) in cynodonts now formally subdivides into subscapularis and diaphragm. Thus, the HK model demonstrates the origin of diaphragm from the forelimb muscle mass assigned to the brachial plexus and supports the idea of C4 as the basic take-off point for the plexus (Fig. 10.122).

Kuratani Hirasawa hypothesis regarding the migration of the diaphragm. Transposition of the diaphragm comes about via caudal transposition of the forelimb. Muscle mass of subscapularis will subdivide with caudal component remaining in relation to the limb and the dorsal diaphragmatic component inserting itself into the pleuroperitoneal fold. (a) Pelycosaur-grade synapsid pectoral region (ventral view), showing the positions of the subscapular muscle (m sbsc), the respiratory and circulatory systems (left), and the skeleton (right). The drawing of the skeleton is based on that in Romer & Price (1940). (b) Embryonic development of the hypothetical proto-diaphragm (ventral view). (c) Embryonic development of the diaphragm in extant mammals (ventral view). atr, atrium; aor, aorta; aor ar, aortic arch; ant ca ve, anterior cardinal vein; ant cor, anterior coracoid; br ve, brachial vein; cl, clavicle; glen, glenoid; du cu, ductuscuvieri; f cor, coracoid foramen; icl, interclavicle; jug ve, jugular vein; lu b, lung bud; pm dph, premuscle mass for the diaphragm; pm sbsc, pre-muscle mass for the subscapular muscle; po ca ve, posterior cardinal vein; po cor, posterior coracoid; scap, scapula; sin ven, sinus venosus; s tr, transverse septum; vent, ventricle. [Reprinted from Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat 2013; 222:504–517. With permission from John Wiley & Sons]
Congenital diaphragmatic hernias have received a great deal of attention in the pediatric surgery literature. The posterolateral Bochdalek form accounts for 95% of cases, the remainder divided between the hernia of Morgagni just behind the xiphoid and eventration, a condition in which a floppy diaphragm billows upward into the chest. The c-met null-mutant mouse has an amuscular diaphragm and has been used to show that the Bochdalek defect is primarily due to mesenchymal defect in the pleuroperitoneal folds such that they do not provide a sufficient scaffold for myoblasts to populate. We can surmise that the position of the Foramen of Bochdalek indicates the perhaps the final site of pleuroperitoneal fold development, one which is last to be completed and first to demonstrate tissue insufficiency.
Phylogeny of the Phrenic Nerve and Brachial Plexus
The brachial plexus in all tetrapods (except mammals) has four spinal nerves, br1–br4. These are paired, respectively, as radial nerve (br1–br2) and long brachial nerve (br3–br4) to produce two y-shaped nerves. These subsequently marry up transversely to form a “double-Y” (cf Fig. 10.119).
Of critical importance is that the position of the four-nerve plexus along the neuromeric axis depends on the number of cervical vertebrae. Also, the neck trunk interface is always between br3 and br4 In chameleons, with five cervical vertebrae, the plexus begins at spinal nerve 3. Amniotes in general have six cervical vertebrae with the rostral limit of pectoral forelimb innervation being at spinal nerve 4 and the trunk beginning at sp7 Ostriches have 20 cervical vertebrae so the plexus begins at C18.
This variation in the number of cervical vertebrae is reflected in the paleontological record. The amniote bauplan begins with an anapsid skull and five vertebrae. It then bifurcates into the saurapsida leading to reptiles and birds and the synapsida leading to mammals. Saurapsids split into the dead-end eurapsids with five vertebrae and the diapsids with 6. These latter subsequently expand to seven–eight in extent reptiles and eight or more in birds. Synapsid are more sedate. The therapsid line goes from five to six. The cynodonts (immediately prior to mammals) stabilize at seven cervical vertebrae (cf Fig. 10.119).
Mammalian brachial plexus reflects the anatomic consequences of cervical expansion. The original four roots are pushed backward to levels C6–T1 maintaining their anatomy all the while. What remains behind at levels C4–C5 are two roots with exactly the same Y configuration as before. Thus, the mammalian brachial plexus consists of three Y-shaped nerves, in antero-posterior order: median, radial, and ulner. All three of the Ys are cross-linked.
Key motor nerves are affected by these anatomic shifts. In basal amniotes, the brachial plexus is at the fourth–seventh spinal nerve. Supracoroideus (spc) and subscapularis (sbcs) are supplied from the common stem of br1 and br2, that is, from the radial nerve (cf Fig. 10.120a).
To the amniote pattern, mammals add two additional cervical vertebrae, causing the position of the brachial plexus to shift backward to the sixth–ninth spinal nerves. This forces a partial duplication of the original plexus in which the original pattern of C4–C5 is repeated at levels C6–C7. Suprascapular nerve (sps) remains connected with neuromeres c5–c6; it is now innervated by a new nerve, the median. Recall the median is a mere repetition of radial and that radial and ulnar are cross-connected. Because the median repeats the ancestral condition, it too sends a cross branch backwards to connect with ulnar, bypassing the radial. It remains attached to radial nerve and is dragged backward with it to level c6–c7. Meanwhile (back at the ranch) a new nerve, phrenic, appears at the br1–br2 level; it is supplied by c4–C5 (cf Fig. 10.120b).
Implications of the Brachial Plexus Shift for Trapezius
Conflictual data exists regarding the motor supply to mammalian trapezius: does it terminate at C4 or extend down to the sixth spinal nerve? In the creation of the mammalian brachial plexus. Based on our discussion of the amniote brachial plexus neuromuscular structures at levels c4–c5 are duplicated or altered to create an 8-nerve plexus. Subscapularis therefore moves backward from sp5 to sp7. For this reason, we propose that cucullaris simply expanded its available sources of mesenchyme and split into Scm and Tpz.
Finally, we ask the question: why are all these hypaxial muscles inserting into the skull and spine? Cucullaris in placoderms is clearly paired with an extensor antagonist. It was clearly supplied by a ventral nerve. The limbs in primitive tetrapods, later amphibians and reptile required powerful ventral muscles to control splayed-out limbs. Changes in the shoulder and hip joints allowed for mammals to positioned their limbs beneath the body. Dorsal muscles became much more important to retract the humerus. More simplistic muscle masses in reptilomorphs broke into subunits which became more sophisticated for complex and agile movements. For this reason, it is no surprise that in mammals, motor control of the dorsal scapula involves the transposition of muscles to the dorsal midline.
Myology of the Neck: Final Thoughts
We have now completed our survey of the muscles of the neck. As always, the assignment of myoblasts to a particular neuromeric level is based on the neuromeric principle of a one-to-one correspondence between motor nerve and somite. The origin of a muscle is determined by the root(s) of its motor nerve. Migration of myoblasts for each muscle follows a strict spatial–temporal order, an order that determines the final outcome of the system. Migration is a very physical, real-time process. Successive groups of myoblasts find a CAM “slime trail” and venture out, pushing and shoving their way along established tissue planes until they encounter an available binding site.
Important sidebar: The neuromuscular relationship is reciprocal. At each neuromeric level, a level-specific homeotic code is shared with all tissue derivatives, including somites, intermediate and lateral plate mesoderm, endoderm, neural crest, and neural tube. This resolves the issue of whether the spinal cord has individual myelomeres. Although its neuroanatomy consists of ascending and descending tracts, synaptic connections between spinal nuclei and ganglions are level-specific. The homeotic coding, seen peripherally, imposes itself on the CNS.
Muscles possess intrinsic properties; these determine four things: (1) when they migrate, (2) how they interact with their neighbors, (3) where they form a primary attachment (insertion), and (4) where they make a secondary attachment (insertion). Myoblast development is a tightly choreographed cranio-caudal dance in which the final anatomy of a muscle respects the territory of previously established muscle units. For example, myoblasts departing from somite Sn at time t1 are followed by those departing at t2. Maturation of myoblasts within somite Sn + 1 occurs slightly later, therefore Sn + 1 myoblasts will always respect the pathways of their predecessors. Muscles “flow” like amoebae along spaces defined by their surroundings. Consummate opportunists, embryonic muscles always “choose” the nearest available binding site. Recognition of the binding site is the responsibility of the fascia surrounding the myoblasts. The primary attachment site is usually a bone originating from the same neuromeric level, but there are exceptions. If, after all, muscles from Sn have taken up their primary attachments, an unfilled binding site persists within the territory of neuromere, then the first available muscle from Sn + 1 will seize the opportunity and attach there. Secondary attachments develop according to the same rules. Finally, muscles insertions follow two patterns: (1) antegrade–antegrade and (2) antegrade–retrograde, as in the pectoral girdle.
Time and again we have seen how these principles are reflected in the attachment sequence of muscle units. Consider the intrinsic muscles of the larynx (somite 1) versus cricothyroid (somite 2). These are supplied by superior and inferior laryngeal nerves respectively. The intrinsics take up binding sites within the larynx while cricothryoid is forced outside, binding to r8–r9 interface between inferior border of r8 thyroid and superior border of r9 cricoid. Migration patterns of tongue and infrahyoid are very orderly. Omohyoid follows the same rules as its inferior belly seeks out a primary C2–C3 binding site on scapulae; it then doubles back to insert on hyoid. Longus capitis originates from cervical somites 1–3 whilst longus colli originates from somites 2–6; origins/insertions of longus capitis take precedence of those of longus colli. The 3-stage developmental sequence of longus colli illustrates what happens when one part of a muscle “blocks out” it neighbor. The vertical portion (S6–8) “spills” downward and centric. “Overflow” of myoblasts from somites 7–9 tracks upward in the opposite direction to attach at the skull base. Finally, the seemingly-absurd anatomy of the multi-layered epaxial muscles falls into a familiar (and understandable) neuromeric pattern. These muscles begin with deep-lying mononeuromeric units (intertransverse-interspinalis) which take up available attachment sites along the midline. Additional poly-neuromeric muscles develop in progressively more superficial and lateral layers. The attachments of these muscles are determined by the principle of “first come, first serve.” The origins and insertions of these later muscles utilize those sites “left over” after the synthesis of the predecessor muscles.
As one considers the organizational plan of these muscles, the most striking observation is the overall simplicity of the system. Muscles come from somitic PAM. These migrate depending upon their biologic maturity within the myotome. This is a function of their spatial location within the myotome, itself determined by homeotic genes that provide system of “coordinates” within the myotome, a sort of “street map,” as it were. Axial bones come from somitic sclerotomes, whereas appendicular bones arise from lateral plate mesoderm. Each bone has a developmental sequence which is manifested as a series of collagen-II muscle binding sites laid out in spatial-temporal order. Musculoskeletal development, that is, the origin-insertion system, is a simple but elegant process in which spatiotemporal units of muscle are “matched up” with spatial–temporal system of bone. In summation, mesodermal units of muscle and bone in genetic register with one another constitute a unique and elegant system of self-assembly.
Angiology of the Neck
Introduction: Arteries of the Neck Don’t Seem Segmental…But They Are
In this, our fourth and final section, we will take on the arterial system of the neck. At first glance, this anatomy seems very different from that of the head. Instead of individual arteries assigned to an organized series of pharyngeal arches what we observe is a series of long vessels extending upward from the subclavian artery in the chest. These vessels give off branches to multiple neuromeric levels. Our goal is to understand how and why the seemingly random arrangement of these arteries develops according to a Cartesian plane.
Let’s begin with a simple model of neck artery development. We place a transparent graph over a segment of human embryo from neuromeres c1 to c8. Our embryo is oriented in the vertical position. The y-axis is midline of the embryo, that is, the CNS. The x-axis is remainder of the embryo away from the midline, that is, all non-CNS tissues. We start out with a simple blood supply consisting of paired longitudinal tubes (dorsal aortae) connected to a heart. From these tubes, a series of primary horizontal segmental arteries project medially, one for each neuromere. The resulting system looks like a ladder with the neuromeres supplied by individual rungs. Now, let’s make our embryo more complex. We will add on additional tissues on either side of the neuraxis. Supporting arteries for these tissues extend outward from the rungs of our previous “ladder.” These secondary horizontal arteries supply tissues that are genetically in register with the same neuromere. Now, let’s link up our secondary segmental arteries using a series of vertical interconnecting arteries. The original ladder now looks like a lattice.
Let’s now focus on the lowest arterial “rung” in the mammalian neck: the seventh intersegmental artery. This vessel runs between the seventh cervical vertebra and the first thoracic vertebra. It is destined to become the subclavian artery. Because it is positioned at the cervicothoracic junction, this artery will ultimately supply the upper limb. Prior to entry into the limb bud, the individual segments of the vertical interconnecting arteries are now fused into longitudinal axial arteries arising from the subclavian. Some ascend to supply the neck (vertebral, thyrocervical, and costocervical. Others descend to supply the sternum and chest (internal thoracic) or the muscles controlling the pectoral girdle (thoracoacromial and long thoracic). Finally, let’s fuse the dorsal aortae such that they no longer supply the hindbrain and neck. The brain and peripheral tissues are now dependent on the axial arteries, each of which maintains original longitudinal segmental branches to specified tissues.
So that’s our model of cervical blood supply. Now, let’s look at the evolution of this system in real-time, using staged human embryo dissected and drawn by Dorcas Padget. Obviously, we are reworking the material discussed in Chaps. 7 and 8 on the vascular system. But the neck is sufficiently different in design that it makes sense to consider her work anew. Embryonic events involving the aortic arch arteries and the pharyngeal arches are rehashed…but from the perspective of tissues from c1 and below.
The embryology of the cervical arteries will be a “thrice told tale.” First, we’re going to review in broad brush strokes the sequence of events by which craniofacial blood supply is set up. This will include the “inside story” about how the heart and dorsal aortae are originally connected. Knowing this, the true identity of the first aortic arch artery will be revealed …are you getting curious? We will reinforce our narrative using a timetable of events relevant to the neck organized by Carnegie stages. Our second task is to develop a visual appreciation of vascular development in staged human embryos as depicted by Padget. In particular, we will focus on the embryogenesis of the vertebral artery because this model applies to all other derivatives of the subclavian. Our third iteration of this story explores the embryologic rationale behind these events. The mammalian neck is both a conduit and a container. Structures related initially to the pharyngeal arches, such as the heart and thymus, pass through it into the chest. Contained within the neck are the transition zone between pharynx and esophagus, endocrine glands (thyroid and parathyroid), the immune system (thymus), and the take-off of the respiratory system. As these structures emerge, the grid-like vascular plan of the primitive state (based strictly on neuromeres) gives way to a very different arrangement. Understanding the how and why of this process is our final goal. This requires careful study of how the great vessels develop, and in particular, the embryologic relationships that exist between them, the aortic outflow tract, and the aortic arch arteries. Background preparation for this story will involve a brief sojourn through relevant aspects of cardiac development.
In sum, the initial vascular system of the neck is simplistic and Cartesian. During the mid to late embryonic period (stages 12–23) rapid growth and differentiation force changes in the vascular system that result in an adult arrangement very different in form, but ultimately segmental in function (Figs. 10.123, 10.124, 10.125, 10.126, 10.127, 10.128, 10.129, 10.130, 10.131, 10.132).

(DHP 14). Stage 13, early Embryo 588a (4 mm) depicts baseline vascular relationships. This embryo (the first of Padget’s series) has fully formed aortic arches 3 and 4. Aortic arch arteries 1 and 2 have involuted; the remnants of first and second pharyngeal arches are supplied by a tangle of vessels connected to the outflow tract. Brain circulation is as follows. The hindbrain is fed via the primitive trigeminal artery, located just above the first pharyngeal arch. This vessel marries up the dorsal aortae with the longitudinal neural artery. Note that the caudal end of the longitudinal neural artery is fed by the primitive hypoglossal and first cervical segmental arteries. The dorsal aortae cranial to the first aortic arch artery acquire a new name: the internal carotid arteries. These supply the midbrain and forebrain. This embryo demonstrates the earliest stages in the involution of the first two aortic arch arteries. AA1 and AA2 are relatively intact here. The tangle of vessels within the arches that will become the ventral pharyngeal artery is not yet present. Cervical somites are supplied by segmental arteries. During stage 13 the dorsal aortae fuse to form a median vessel, the descending aorta as seen in the adult. The unified dorsal aortae give rise to three groups of arteries. (1) Unpaired, ventral, visceral (splanchnic) branches supply visceral lateral plate mesoderm. The celiac, superior mesenteric, and inferior mesenteric arteries supply the gut. (2) Paired, lateral, visceral (splanchnic) branches supply intermediate mesoderm. These include the suprarenal, renal and gonadal arteries to retroperitoneal structures. (3) Dorsolateral, parietal (somatic) branches supply paraxial mesoderm and parietal lateral plate mesoderm.

(DHP 1a) Stage 13, late Embryo 588b (4 mm) is slightly later in development. Within pharyngeal arches 1 and 2 are vessels that will be the forerunners of the external carotid system. These connect up with the cardiac outflow tract at the junction of aortic arches 3 and 4. The former insertions of AA1 and AA2 into the dorsal aortae are marked by two remnants vessels hanging down from the dorsal aortae: the mandibular artery and the hyoid artery. The caudal end of each longitudinal neural artery is clearly supplied by the first cervical segmental artery from its respective, as-yet unfused, dorsal aorta. Because the first and second aortic arches are now interrupted, blood flow forward from the heart to the brain is entirely dependent on the third aortic arch artery. For this reason, the portion of the dorsal aortae extending forward from AA3 is now defined as the primitive internal carotid arteries. AA4 is plugged into the dorsal aorta just behind AA3. Blood flow backward into the remainder of the embryo takes place through AA4

(DHP 15) Stage 14, early Embryo 2841 (5.3 mm) demonstrates further changes in the arterial supply to pharyngeal arches. What were formerly a tangle of vessels within the arches have now consolidated into the ventral pharyngeal artery. Although the proximal take-off of the VPA is unclear, it extends as far forward as the downgrowth of V3. The hyoid artery (future stem of the stapedial system) is prominent. Backward anastomosis between the caudal division of internal carotid and the longitudinal neural arteries marks the completion of brain circulation. As a consequence, smaller interconnecting arteries (primitive trigeminal and primitive hypoglossal), formerly essential for supplying the longitudinal neural artery to the hindbrain, have now involuted.

(DHP 3a) Stage 14, late Embryo 3960 (5.5 mm) shows us more clearly how the stem of the external carotid does not take off from the third aortic arch per se. Instead, it extends forward from a segment of outflow tract common to both AA3 and A4. The sixth aortic arch artery, now briefly seen, has given rise to the primitive pulmonary artery. Cervical blood supply is still strictly a function of the segmental system. These arteries are clearly intersegmental. The first cervical intersegmental lies dorsal to the first cervical nerve. It supplies both the caudal termination of the longitudinal neural arteries (the anatomic boundary of the unpaired basilar arteries) and the first cervical somite. From the seventh cervical intersegmental artery, the subclavian stem of the subclavian artery is now seen. Aortic arch artery 6, often termed the pulmonary arch, arises from two sources. A ventral stem arises at stage 13 from the aortic sac while a dorsal stem arises at stage 14 from the dorsal aorta. When these unite they connect the parent structures; no intervening pharyngeal arch is present. There is no such thing as the sixth pharyngeal arch. At stage 14 three important structures arise from the structures encoded to r8–r9. Lung buds (endoderm) project into lateral plate mesoderm. The trachea and the esophagus differentiate with lateral plate mesoderm. A rich blood supply develops within LPM, from which arises the pulmonary vascular plexus. It explains why bronchial vessels are supplied separately from the pulmonary arteries per se. Once the 6th (pulmonary) arch forms, it sends out the true pulmonary artery to connect to the pulmonary vascular plexus

(DHP 16). Stage 15 Embryo 163 (9 mm) demonstrates characteristic cerebral hemispheres. Transverse anastomoses have formed uniting all seven cervical segmental arteries. This is the precursor state of the vertebral artery. At their caudal termination the longitudinal neural arteries are still unpaired. Although we know that the vertebrals will eventually anastomose with the LNAs, blood flow to the hindbrain in stage 15 is still predominantly from the internal carotid. The subclavian is now prominent. It clearly relates to the paired dorsal aorta

(DHP 17) Stage 16 Embryo 1771 (12.5 mm) marks the end of the pharyngeal arch phase. The stapedial system is developing from the hyoid artery. Intracranial stapedial provides circulation to the meninges and the orbit and medial nasopharynx. Extracranial stapedial becomes a distal “add-on” to external carotid, the internal maxillo-mandicular artery. Its branches supply the lateral nasopharynx, oropharynx, and face. The heart has not yet descended into the chest. The future vertebral artery is irregular but clearly seen, its ventral connections to the dorsal aortae have involuted, save at the level of the seventh intersegmental artery. Here, the take-off of the subclavian artery is seen. This stage is transitional in higher vertebrates from a cerebral circulation exclusively supplied by the carotid to the later iteration in which hindbrain is supplied by the vertebrals

(DHP 6) Stages 17–18 Embryo 940 (14 mm) demonstrates annexation of the distal ventral pharyngeal artery (external carotid) by the extracranial stapedial at the level of V3

(DHP 18) Stages 19–20 Embryo 1390 (18 mm) shows adult form for the external carotid artery and most cerebral arteries. The stapedial system now has orbital, maxillary, and mandibular divisions. Subclavian now arises opposite the pulmonary arch. It is the sole source of vertebral artery. Thryocervical trunk present as well.

(DHP 19) Stages 21–22 Embryo 632 (24 mm) has secondary anastomoses of the stapedial artery: StV1 orbital to the primitive ophthalmic to form definitive (hybrid) ophthalmic artery and extracranial StV2 internal maxillary to the intracranial StV2 to form middle meningeal artery. Subclavian has the full complement of branches: vertebral, thryocervical trunk, internal mammary, costocervical trunk is not shown but is developmentally present, and axillary (brachial) artery

(DHP 20) Stage 23+ Embryo 886 (43 mm) shows posterior communicating artery reversing its position from caudal-cranial 180 degrees to cranial-caudal. Stems of both inferior cerebellar arteries (anterior and posterior) are visible and dead-end in the choroid plexus of fourth ventricle. Choroid plexus of lateral ventricle supplied by anterior and posterior choroidal aa. Lacrimal branch (representing the StV1–StV2 supraorbital artery) connects with ophthalmic
Beginning of Embryonic Circulation
In its most primitive state, the unflexed embryo has four vascular systems that develop in situ. Vitelline vessels arising from the yolk sac are extraembryonic. Within the embryo, the embryonic CNS (future hindbrain) is supplied by paired primitive hindbrain channels (PHC) while the non-CNS embryo is nourished by paired dorsal aortae (DA) running its entire length. The dorsal aortae develop from lateral plate mesoderm; they are situated peripherally to the paraxial mesoderm of the somitomeres and somites. At their anterior terminus, each dorsal aorta is plugged into the footplates of a horseshoe-shaped zone of cardiogenic mesoderm.
All other arterial systems in the embryo develop by induction and budding. To keep ourselves oriented, let’s enumerate these important secondary systems in the order of their appearance.
-
Aortic arch arteries AA1 is merely a stretched-out continuation between the ventrally-flexed heart tubes and the dorsal aortae. The six aortic arches appear between stages 9 and 15.
-
Primitive internal carotid artery sprouting from the genu where the dorsal aortae bend downward as the first aortic arch arteries. Primitive ICA is defined as extending cranial from AA1.
-
Transverse interconnecting arteries between the outboard dorsal aortae and the primitive hindbrain channels: primitive trigeminal, primitive otic, primitive hypoglossal.
-
Reconfiguration and renaming of the PHCs into the longitudinal neural arteries.
-
Breakdown of aortic arch arteries within pharyngeal arches leading to the development of the ventral pharyngeal artery (VPA) or primitive external carotid.
-
Degeneration of DA between AA3 and AA4 leads to definitive internal carotid artery.
-
Hyoid artery > stapedial artery system programmed by cranial nerves:
-
intracranial stapedial programmed by cranial nerves > meningeal arteries,
-
extracranial stapedial programmed by chorda tympani > internal maxilla- mandibular artery and its branches.
-
-
Fusion of IMMA with VPA > definitive external carotid artery.
-
Degeneration of stapedial stem > anastomoses.
-
StV1 with internal carotid > ophthalmic artery;
-
StV2 with external carotid > middle meningeal a.; other meningeal aa “originate” from ECA.
-
Early development of the heart takes place in the visceral layer of lateral plate mesoderm. Three phases are described. During the plexiform phase (stage 9) LPMV gives rise to an endothelial plexus; this is quickly surrounded by a myocardial mantle. The primitive heart is non-contractile. It consists of paired heart tubes linked distally in the form of an inverted U-shaped loop. Proximally the loop is continuous with the dorsal aortae. The connection between the future aortic sac and the dorsal aortae is Y-shaped. Distally, the future sinus venosus is continuous with the vitelline vessels. In the straight tubular heart phase (stage 10) the endocardial plexus organizes into a single tube. It has a linear fate map, the segments of which are (from proximal to distal): aortic sac, truncus arteriosus, conus cordis, RV, LV, LA, RA, and sinus venosus. Three sets of feeding vessels plug into the sinus venosus: cardinal, omphalo-mesenteric, and umbilical vein. The looped heart phase (stages 10–11) takes place during flexion of the embryo. Anatomic fates are assigned to sectors of the heart. The distal loop assumes an atrial fate; it receives the future gut vessels. More proximal segments form the ventricles and the cardiac outflow tract.
During the pharyngeal arch phase, the embryonic heart lies below the pharynx. From the aortic outflow tract, a succession of six aortic arch arteries appears. The first five develop within the central core of the five pharyngeal arches; they connect the now-beating embryonic heart with the overlying dorsal aortae. The sixth aortic arches form the pulmonary circulation. Note that the development of the first aortic arch artery is unique. It does not “sprout” from the aortic sac. When the embryonic flexion carries the heart ventrally, the aortic sac pulls the dorsal aortae downward: a 90-degree bend is created at the fourth somitomeres, that is, at rhombomeres 2 and 3. From this site sprout: (1) the primitive internal carotid artery and (2) the primitive trigeminal artery. The first aortic arch artery = the vertical segment of dorsal aorta spanning from the Sm4 downward to the aortic sac.
Note that at no point in development are all aortic arch arteries present at the same time. They form and involute in strict sequence. Despite this instability, blood flow from heart to embryo is always maintained. Just when AA1 and AA2 are falling apart, AA3 and AA4 are plugged into the DAs. AA5 has a short half-life. It bequeaths its vascular territory to AA4. As the heart descends into the chest, ventral remnants of aortic arch arteries 1–4 reorganize into ventral pharyngeal artery. VPA forms the external carotid system up to the external maxilla-mandibular (facial) artery. The dorsal remnants of the aortic arch arteries also have important roles. For our purposes, the key player is hyoid artery of the second aortic arch. It gives rise to the stapedial arterial system, the intracranial branches of which supply the dura while the extracranial branches form the internal maxilla-mandibular artery. Thus, remodeling of aortic arches 1–4 supplies all derivatives of pharyngeal arches 1–5.
Blood supply to the remainder of the embryo is organized along very different lines. From the paired dorsal aortae segmental vessels are directed upward to supply each neuromeric zone of the embryo. The first pair of such vessels is the primitive hypoglossal arteries. These connect the DAs to the longitudinal neural arteries supplying r8–r11. Immediately caudal are the segmental (cervical) arteries. From this point backward, each somite is supplied by a segmental artery, originating from the ipsilateral dorsal aorta. Obviously, this situation is unstable. The heart will descend into the chest, the great vessels will arise and the dorsal aortae will fuse. In response to these events, the anatomic basis of blood supply to the neck will also change. We will follow this developmental sequence in staged human embryos as per Padget.
Timetable of Arterial Development by Stages
The aortic arch artery period lasts from stage 9 to stage 14. One arch develops per stage.
Stage 9 (first aortic arch artery): This stage is marked by the appearance of the first three occipital somites and the formation of heart tubes from lateral plate mesoderm (visceral lamina). First aortic arch artery is present at this stage.
Stage 10 (first pharyngeal arch, second aortic arch artery): A single tubular heart is present, functional embryonic circulation is established and the pharynx develops. The embryo is flexed, the heart is now ventral and the heartbeat is initiated. The second aortic arch artery is present. The heart is now connected to the remainder of the embryo via aortic arch arteries 1 and 2; and these arteries course upward through the cores of their respective pharyngeal arches to reach the dorsal aortae.
Stage 11 (second pharyngeal arch, third aortic arch artery): The oropharyngeal membrane, located at the boundary between pharyngeal arches 2 and 3, ruptures. The OPM also marks the boundary between the ectoderm of the oro-nasopharynx and the endoderm of the pharynx proper.
Stage 12 (third pharyngeal arch, fourth aortic arch artery) dorsal aorta begins to fuse with the process continued through stage.
Stage 13 (fourth pharyngeal arch, fifth aortic arch artery, involutes in tetrapods) ventral sprout of pulmonary artery from AA4, dorsal aorta begin to fuse distal to it.
Stage 14 (fifth pharyngeal arch, sixth aortic arch artery) dorsal sprout of pulmonary artery arises from dorsal aortae. Union of the two sprouts becomes AA6. It has nothing to do with a pharyngeal arch. Pulmonary artery appears to arise from AA6 but, in actual fact, is biologically correlated with AA4, that is, r8. Transverse branches from paired aortae supply the segment.
Stage 15 Longitudinal neural arteries fuse together from r1 to r8 to form the basilar. Interconnections form linking the segmental aortic branches. These will unite to form the vertebrals.
Stage 16 subclavian artery defined. Vertebrals now connect the subclavian to the cerebral circulation.
Stage 17 Brachiocephalic trunk and left common carotid, stapedial artery system, fusion of dorsal aortae is now complete.
Stage 18 and Stage 19.
Stage 20 Descent of the thymus from r8 into chest.
Stage 21 and Stage 22.
Stage 23 Vascularization of the thymus by internal thoracic artery.
During the aortic arch artery period the heart lies cephalad to the occipital somites, that is, cranial to r8. During stage 12, after the formation of AA4, it descends to the level of the somites r8–r11. By the eighth week of development, the heart lies at the mid-thorax.
Cervical Artery Development in Stages
The following illustrations from Padget’s work demonstrate the time course of vessel development. These are based on dissections from the original Carnegie Embryo Collection. The reader is referred to Figs. 10.123, 10.124, 10.125, 10.126, 10.127, 10.128, 10.129, 10.130, 10.131, 10.132 for a stage-specific presentation of the cranial arterial development sequence.
The Carnegie collection along with contributions from many other sources has been made digital under the direction of Dr. Mark Hill at University of New South Wales http://human-embryology.org
Readers can readily access the collections: https://human-embryology.org/wiki/Carnegie_Collection
Arteries of the Neck
Arteries supplying the structures of the neck originate from two sources. Fourth and fifth pharyngeal arch structures involving the larynx are supplied from the most caudal branch of the external carotid system, the superior thyroid artery. Recall that this vessel originates from the stem of the fourth aortic arch artery; it is the right and proper supply of the fourth pharyngeal arch. However, due to the untimely death of the fifth aortic arch artery, AA4 is forced to assume the duties of supplying the fifth pharyngeal arch. Given its embryologic assignment, superior thyroid terminates within the larynx. It does not relate to the musculature of the lower pharynx, specifically to middle constructor and below. These structures are supplied by the inferior thyroid artery.
Subclavian Artery
Subclavian artery develops from the seventh cervical intersegmental artery. It is located at the genetic take-off of the upper extremity. The four arterial stems of the subclavian are as follows: vertebral, thyrocervical trunk, internal thoracic, and costocervical trunk. These vessels likely develop in a proximal to distal sequence.
With regard to the neck and pectoral girdle, the branches of subclavian supply: (1) structures within the neck up to the territory of fourth arch (external carotid), (2) the pectoral girdle including the rostral, medial, and rostro-lateral scapula, and (3) structures connecting the neck with the pectoral girdle (Fig. 10.133).

Subclavian artery. Branches supply (1) structures within the neck up to the territory of fourth arch (external carotid), (2) the pectoral girdle including the rostral, medial and rostro-lateral scapula, and (3) structures connecting the neck with the pectoral girdle. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
Vertebral Artery
The first stem of the subclavian arises from the first part (medial to scalenus anterior). It avoids the transverse process of the seventh cervical vertebra but at level C6 and above it passes through foramina in the transverse processes. It has four parts: (1) From its stem to C6, (2) from C6 to C2, (3) arising medial to rectus capitus lateralis, it curves backward and medially around the lateral mass of C1.
-
Spinal branches enter via intervertebral foramina. They supply the spinal cord and its meninges. Paired lateral chains lie above the posterior surface of the vertebral bodies just at the attachment of the pedicles.
-
Muscular branches are assigned to the muscles of the suboccipital triangle. Muscles arranged along the ascent of the vertebral artery are supplied from other sources.
Internal Thoracic Artery
The second stem of the subclavian might seem very far afield of the neck but several of its branches have interesting developmental features. First off, internal thoracic exits from the first portion of subclavian directly opposite the thyrocervical trunk. As we discussed previously, internal thoracic is the mere continuation of a longitudinal union of a series of segmental vessels. The mesenchymal targets of these vessels include structures derived from both paraxial mesoderm and lateral plate mesoderm.
-
Pericardiophrenic artery is the most proximal branch of internal thoracic. It accompanies (and supplies) the phrenic nerve (C4) into the chest, coursing in between pleura and pericardium. The artery is slender; one should consider it as primarily a means to support the motor nerve.
-
Mediastinal branches are distributed anteriorly to supply the parietal (sternocostal) surface of pericardium and regional lymph nodes.
-
Thymic branches remind us of the embryologic location of this gland. Thymus arises from the endoderm of the caudal portion of the third pharyngeal pouch, that is, between the third and the fourth arches. That places it at level r8. Primordial thymus contains two different types of tissue. The rostral part of pouch 3 produces medullary duct epithelium while caudal pouch 3 may be the site of reticular epithelium. Neural crest cells are responsible for the connective tissue framework of the thymus. They may also be responsible for inducing the pharyngeal pouch epithelium to produce and maintain the differentiation of lymphoid stem cells. At stage 20, lateral plate mesoderm penetrates the thymus, bringing in arterial supply. The thymic parenchyma becomes divided into lobules. Circulating stem cells from the bone marrow take up residence in the newly vascularized thymus. These cells produce: (1) thymocytes committed to T-cell lineage, and (2) dendritic cells. During the third trimester functionally mature thymocytes migrate from the thymus to populate peripheral lymphoid tissues permits circulating systemic dispersal of stem cells into the rest of the body. Note that thymic development cannot proceed in the absence of neural crest cells from levels corresponding to the third pouch, that is, from r7 and r8.
-
Bronchial branches supply the lower trachea (lateral plate mesoderm) as well as neighboring bronchi.
-
Sternal branches are six in number, the uppermost one supplies manubrium. The intermediate four branches are distributed to the four segments of the sternal body. The sixth and last branch supplies the xiphoid. [Recall that manubrium also is also supplied by suprasternal branch of suprascapular artery and by the clavicular branch of thoracoacromial artery. This explains why manubrium has six centers of ossification; three per side].
-
Anterior intercostal arteries are also six in number. The remaining interspaces are supplied by the musculophrenic artery, the penultimate branch of internal thoracic. These arteries supply the intercostal muscles. At the midaxillary line, they anastomose with their counterpart posterior intercostal arteries from the aorta. Penetrating branches supply overlying pectoralis and breast tissue. The fourth intercostal is particularly important in that regard.
-
Perforating branches pass through five–six interspaces at the sternal border to gain the subcutaneous plane. Here they travel laterally, accompanied by the anterior cutaneous branches of intercostal nerves. They initially run with pectoralis fascia just under the muscle. More laterally they become more superficial to supply the muscle itself and skin. Branches of the second, third, and fourth interspaces support the breast.
-
Musculophrenic artery follows the posterior surfaces of ribs 8, 9, and 10. In like manner to the anerior intercostals it gives off branches to the seventh, eighth, and ninth interspaces. Terminal branches access the lower part of diaphragm.
-
Superior epigastric artery is the terminal branch of internal thoracic. Its anastomosis with inferior epigastric demonstrates that the segmental neurovascular organization is continued all along the anterior abdominal wall. Internal thoracic follows the same principles as vertebral with multiple intersegmental branches linking up together longitudinally and then, due to changes in flow and the losing connection with the individual intersegmental vessels.
Thyrocervical Trunk
The third stem of the subclavian, thyrocervical trunk arises from the first part of subclavian, just behind the medial edge of scalenus anterior. It provides branches to all neuromeric levels from c8 up to c1. It provides blood supply for structures lying outboard to those supplied by the vertebral.
Inferior Thyroid
Inferior thyroid is the first artery of the thyrocervical trunk. Inferior thryoid tracks upward in front of the vertebral artery, swerves internally to run dorsal to carotid sheath, and then terminates in the thyroid gland at levels r8–r11. It has the following important branches.
-
Muscular branches These supply infrahyoid, and three multi-segment muscles: longus colli (atlas to T3), scalenus anterior (the anterior tubercles of the third through sixth cervical vertebrae represent neuromeric levels c3–c7), and inferior constrictor (a Sm9 muscle attaching to the back of thyroid and cricoid cartilages).
-
Ascending cervical This artery can arise in two locations: behind (dorsal) to carotid sheath from the parent artery or directly from the trunk. In any case, it skirts along the anterior aspect of the cervical vertebrae, at the level of the transverse processes. This places it between scalenus anterior and longus capitus. From this location, it sends spinal branches into the intervertebral foramina and thence into the vertebral canal. These provide segmental supply to the vertebral bodies and spinal cord in exactly the same way as the spinal branches given off by the vertebral artery.
-
Esophageal branches These arteries provide segmental supply to the ventral aspect of the esophagus. They have anastomoses with segmental arteries to the dorsal esophagus arising from the aorta.
-
Tracheal branches are distributed segmentally along the trachea. These anastomose inferiorly with bronchial arteries and superiorly with tracheal branches of superior thyroid artery (the arterial axis of the fourth and fifth pharyngeal arches). Note that bronchial arteries arise from the ventral side of thoracic aorta and from the upper intercostal arteries.
-
Inferior laryngeal artery runs along the dorsal aspect of trachea to the dorsal larynx. In this position, it is covered by inferior constrictor. Recurrent laryngeal nerve travels along with it.
In sum: The developmental rationale of the inferior thyroid artery is to form a segmental system in parallel with the vertebral. It is responsible for structures within the core of the neck lying outboard to those supplied by the vertebral.
Suprascapular Artery
The second artery of the thryocervical trunk, subscapular is the first of three arterial systems dedicated to the pectoral girdle. The arterial axis pursues a downward and lateral course from the thyrocervical trunk crossing in front of scalenus anterior and phrenic nerve. Sternocleidomastoid lies on top of it. It then travels underneath clavicle, lying atop subclavian artery and the brachia plexus. It can be found beneath the inferior belly of omohyoid. When it reaches the superior border of the scapula it passes backward over the transverse scapular ligament beneath which lies suprascapular nerve. The artery now tracks directly on the bone of the supraspinous fossa. It continues laterally around the neck of the scapula. It now passes through the great scapular notch beneath the inferior transverse ligament to gain access to the infraspinous fossa. Here it makes two anastomoses: (1) with transverse cervical artery via its descending scapular branch; and (2) with subscapular artery via its circumflex scapular branch.
-
Muscle branches This artery supplies the inferior 1/3 of sternocleidomastoid, subclavius, supra- and infraspinatus. It represents the first of three angiosomes dedicated to the pectoral girdle, all of which arise from subclavian.
-
Suprasternal branch curves over the sternal margin of clavicle to supply chest skin.
-
Acromial branch supplies skin over the acromion, traversing trapezius in the process. Here it makes an anastomosis with the thoracoacromial artery via its own acromial branch. Recall that acromion has two ossification centers. It is reasonable to associate these with the two arterial systems. Suprascapular axis, arising more proximally from subclavian, would logically be assigned to the more medial ossification centers.
-
Articular branches supply the joint between distal clavicle and scapula (thoracromial joint).
-
Osseous nutrient branches supply the clavicle and scapula per se.
In sum: The developmental rationale of suprascapular artery is to supply structures related to the clavicle and rostral aspect of scapula. It supplies skin along the clavicle from manubrium as far lateral as acromion, sternocleidomastoid, omohyoid, and subclavius. It plays a supporting anastomotic role to supra and infraspinatus.
Transverse Cervical Artery
The third artery of the thryocervical trunk, the course of transverse cervical runs parallels that of suprascapular artery, running above and behind it. It too crosses in front of scalenus anterior and phrenic nerve (deep to sternocleidomastoid). More laterally, it crosses in front of the trunks of brachial plexus. Just above it lies platysma, which it supplies. As it approaches the anterior border of trapezius it divides into two distinct branches, deep and superficial. It is an artery of considerable variation. Sometimes it is absent. In such cases, the regions supplied by the superficial and deep branches of TCA are served by two completely different arteries. Superficial TCA is replaced by superficial cervical artery and deep TCA is replaced by descending scapular artery. For this reason, in the descriptions below we’ll have to refer to both situations.
Situation 1:
-
Superficial branch of TCA runs deep to hypaxial trapezius and then divides. An ascending branch tracks up along trapezius to anastomose with occipital artery via its descending branch. Descending branch follows the course of spinal accessory under the trapezius.
Deep branch of TCA pursues an epaxial course to supply levator scapulae and adjacent muscles. It reaches the superior angle of scapula and then runs downward along its medial (vertebral) border until reaching the inferior angle. It runs in company with dorsal scapular nerve (C5), which is motor to both levator and the rhomboids. It also supplies subscapularis and serratus posterior. Thus deep br. of TCA is the neuroangiosome of medial border of the scapula.
Situation 2:
-
Superficial cervical artery arises directly from thyrocervical trunk and pursues a purely superficial course. There is no deep branch. It basically supplies the trapezius.
-
Descending scapular artery is seen only when transverse cervical is absent. It arises independently from the third part of the subclavian. The structures it supplies are exactly the same as those supplied by the deep branch of TCA.
In sum: The developmental rationale of TCA is to supply the muscles of scapula from cranial to caudal and from medial to lateral: levator scapulae, rhomboids, on the dorsal side, supra and infraspinatus; on the ventral side, subscapularis and serratus posterior. These muscles connect scapula to chest wall and span from upper lateral scapula to the neck of humerus.
Costocervical trunk is the fourth stem of the subclavian. It arises from two different locations. On the left side comes off the first part of subclavian, just distal to thyrocervical trunk. On the right side, it comes from the second part of subclavian.
Supreme intercostal artery descends just in front of the neck of the first and second ribs. It gives off the first and second posterior intercostal arteries. This is compensation of an oddity of the aorta. The most superior intercostal branch of aorta supplies the third intercostal space. For developmental reasons, aorta is not capable of directly supporting the first two interspaces. Supreme intercostal to the rescue! It anastomoses with the more superior aortic branch.
Deep cervical artery is analogous to the dorsal branch of the aortic posterior intercostal artery. It ascends along the back of the neck between semispinalis capitis and semispinalis cervicis all the way up to the axis. Here are anastomoses with occipital artery (descending branch) and with branches of the vertebral.
Axillary Artery
The axillary artery represents the continuation of the subclavian into the upper extremity. It gives rise to five arterial stems: supreme thoracic, thoracoacromial, lateral thoracic, subscapular, and humeral circumflex (Fig. 10.134).

Axillary artery. With regard to the pectoral girdle, the branches of axillary have a very limited role, supplying the following structures: (1) clavicle, (2) distal projections of the rostral scapula (acromion and the coracord process), (3) caudo-lateral scapula, and (4) muscles connecting pectoral girdle to the humerus and to the anterolateral chest wall. [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]
With regard to the pectoral girdle, the branches of axillary have a very limited role, supplying the following structures: (1) clavicle, (2) distal projections of the rostral scapula (acromion and the coracord process), (3) caudolateral scapula, and (4) muscles connecting pectoral girdle to the humerus and to the anterolateral chest wall.
Supreme Thoracic Artery
Renowned anatomist Richard Snell at George Washington University described this structure rather pithily as “supremely unimportant.”
Thoracoacromial Artery
-
Pectoral branches descend between pec major and pec minor supplying them and the breast.
-
Acromial branch travels laterally under deltoid to access the acromial process where it makes a number of anastomoses to form the acromial rete.
-
Clavicular nutrient branch travels upward and medially all the way to the sternoclavicular joint. It may be the arterial rationale for the secondary sternal ossification center of the clavicle.
-
Deltoid branch supplies pectoralis major and deltoid. It ends up in the acromial rete.
Lateral thoracic artery comes off axillary artery only 30% of the time. In other cases, it comes off thoracromial or subscapular. It supplies serratus anterior and thus the anterior side of the medial (vertebral) border of scapula. For that reason, medial border has two ossification centers. The anterior one is supplied by lateral thoracic. The posterior one is supplied by the artery to the levator scapulae and the rhomboids, the deep branch of transverse cervical artery.
Subscapular Artery
This is the “big boy,” the most sizeable branch of the axillary artery. It arises just distal to the lateral border of subscapularis; it then quickly divides.
-
Circumflex scapular artery is the larger of the two branches. At the lateral border of scapula is passes through the triangular space, bounded above by subscapularis, below by terres major, and laterally by long head of triceps. It travels along the bone of the infraspinatus fossa, supplying it. This is the likely source for one of the ossification centers of the scapular body (the other being supplied by suprascapular artery).
-
Thoracodorsal artery.
Humeral Circumflex Arteries
-
Posterior humeral circumflex artery arises just beyond distal border of subscapularis. It is accompanied by axillary nerve through quadrangular space defined above by subscapularis and teres minor, below by teres major, medially by long head of triceps, and laterally by neck of the humerus. It serves the deltoid and the shoulder joint.
-
Anterior humeral circumflex artery arises just opposite its twin. It supplies the head of the humerus and shoulder joint.
Just like the vertebral and the thyrocervical trunk, internal thoracic was originally a series of segmental arteries coming from the dorsal aortae in the unfolded embryo. Recall that at this embryonic stage, the dorsal aortae run all the way forward and unite together into horseshoe-shape. The most distal parts of the inverted U will become paired heart tubes. The dorsal aortae give rise to three sets of arterial branches, arranged neuromerically, and assigned to specific embryonic structures. Dorsal intersegmental arteries give off dorsal branches supplying the spinal cord and the vertebrae. Lateral intersegmental branches from the dorsal intersegmental arteries supply the neuromeric levels of the neck and trunk. In the thorax, these are represented by the segmentals from the aorta.
Organ Systems of the Neck
Esophagus
The esophagus is a muscular tube extending from the r11 terminus of cricoid cartilage just opposite to vertebral level C6 25 cm downward to the stomach. It thus span from a c1–c8 and t1–t12, a total of 20 neuromeric units. It consists of four layers: the serosa, an external fibrous/fascial layer, the muscularis with two muscle planes, a submucosa comprised of lateral plate mesoderm admixed with sympathetic ganglia and containing blood vessels and an endodermal mucosa (Figs. 10.135, 10.136).

Neuromeric levels of the esophagus. Lateral plate mesenchyme, although it does not proceed through an intermediate somite, is neuromerically organized. Each segment of the esophagus is innervated by a branch from the sympathetic chain, faithfully recapitulating the individual somitic contributions to the rest of the thoracic wall. Cervical esophagus c1–c4 recapitulate the 7-arch branchial arch system. Levels BA6 and BA7 are transition zones where striated PAM muscle transitions to smooth LPM muscle. In levels c5–c8 the eosophagus becomes entirely smooth muscle. Recall that Thoracic esophagus has 10 neuromeric levels with 10 sympathetic ganglia and 10 segmental branches from the thoracic aorta. Note pathway of the left phrenic nerve coursing over the c1–c4 pericardium and its relationship to the ipsilateral vagus [Reprinted from Lewis, Warren H (ed). Gray’s Anatomy of the Human Body, 20th American Edition. Philadelphia, PA: Lea & Febiger, 1918]

Arterial supply to esophagus is via segmental aa. cervical = Inferior thyroid; upper thoracic = descending aorta via bronchial aa; lower thoracic; left gastric right lateral and splenic left lateral [Reprinted from TeachMeAnatomy, courtesy of Dr. Oliver Jones]
The lining of the esophagus is in continuity with the oropharynx. It is a non-keratinized stratified squamous epithelium which, due to its thickness, is protective in nature. It is populated by Langerhans cells which process and present antigens. Lamina propria contains lymphoid tissue and mucous glands with a strange distribution, being found only near the pharynx and at the gastroesophageal junction. This is followed by a muscularis muscosa layer (found only in esophagus and rectum) consists of longitudinal smooth muscle which is thin and whispy near the pharyx but thickens progressively. The function of this layer is to maintain the epithelial surface of the gut in motion so that material is expressed out of the crypts and that the surface lining is kept in contact with the luminal contents. The submucosa contains mucous and serous glands that communicate with the lumen. The muscular layer has a deep plane of circular muscle and a superficial layer of longitudinal muscle.
The mesenchymal composition, blood supply, and innervation of the esophagus differ from those of the midgut and hindgut. We can best appreciate its development in neuromeric terms. The entire GI tract is regionally specified through Homeotic genes. Its basic structural components of endoderm and lateral plate mesoderm, visceral layer (LPMV), are neuromerically coded. Neural crest populations are segmental as well. The muscular layer of the gut has two strata of smooth muscle: the inner layer circular and the outer layer is longitudinal. The muscle layer of esophagus is unique in that its cervical zone has striated muscle. Moreover, the circular muscle layer is in direct continuity with inferior constrictor and is therefore intrinsically branchiomeric.
Esophagus has three sectors based on the mesenchyme of its muscular layer. Cervical esophagus is the upper third and has striated muscle from PAM. In the thorax, middle third of esophagus muscular layer is a mixture of striated and smooth muscle while the distal third is exclusively smooth muscle. Lateral plate mesoderm, visceral layer (LPMV) is the source of the smooth muscle from LPM. Paraxial mesoderm provides cervical muscle. The origin of striated muscle in the thoracic esophagus is uncertain but most likely represents a transformation of local LPMV from a smooth to striated fate.
Vascular supply to cervical esophagus is segmental and arises from inferior thyroid artery. As previously discussed ITA represents the unification of transverse branches originally given off by the dorsal aortae prior to the great remodeling that takes place after the pharyngeal arch period. Each neuromeric segment of esophagus is supplied segmentally from the inferior thyroid axis. Motor innervation of this muscle is supplied by nucleus ambiguus lateral motor column acting through the vagus. Nucleus ambiguus extends downward into the cervical spine at four neuromeric levels. More distal smooth muscle sector is innervated by dorsal motor nucleus of vagus.
Nerve endings for pain and stretch route through segmental spinal nerves in the cervical esophagus whereas in the thoracic esophagus they connect to celiac plexus. Thus, upper esophageal pain can be localized (vaguely) to “find the proverbial fishbone.” Lower afferents convey more generalized sensation.
Note that neural crest cells migrate along the plane of the circular layer at week 6. They then penetrate internally to enter the submucosa where they form the myenteric plexus.
Neuromeric Model of the Cervical Esophagus
Paleontologic Footprint
Recall that in the original agnathic vertebrates, 12 arches have been documented. This implies that 24 neuromeric levels. Recall as well that fish arches represent communications with the pharynx. They were present well beyond the head into the body. The implication is that primitive vertebrate anatomy included branchiomeric muscles for control of the gill arches for 4 cranial and 20 postcranial segments. In humans that translates to c1–c8 + t1–t12 = 20. This represents the thoraco-abdominal boundary. Thus, the neuromeric definition of foregut versus midgut territories resides at the 20th postcranial neuromere?
The anatomy of the mammalian foregut (esophagus) is remarkably consistent with this ancient original fish model. As we have previously discussed, the pharyngeal constrictor muscles above the esophagus arise from PAM before it has organized into the somites. Thus, branchiomeric myoblasts from neuromeres r6–r11 are somitomeric. Striated muscle in the cervical esophagus arises in the same way…outside of the normal somite myotomes. PAM for striated muscle arrives to populate the cervical esophagus from somitomeres Sm9–Sm12 (neuromeric levels c1–c4) most intensely. These levels represent the ancient sixth and seventh branchial arches. Striated muscle is present to a lesser degree in the lower cervical esophagus. This can only be explained by contributions from Sm13–Sm16 (neuromeric levels c5–c8). And the only way to rationalize this is to invoke the embedded genetic capacity of c5–c8 for branchiomeric muscle, presumably a throw-back to the eighth and ninth arches.
This is not to imply that arches 6–9 exist in mammals, simply that the somitomeres Sm9–Sm16 retain the genetic capacity to send out a select population of PAM with segmental pain fibers that track superficial to LPMV. Together they populate the wall of esophagus.
A neuromeric model of the esophagus begins with an endodermal tube surrounded by lateral plate mesoderm, visceral layer (LPMV). The mesenchyme of muscularis mucosa and submucosa is in register with c1–c8. An external layer of striated muscle is contributed from PAM of Sm9–S12 as above. In the lower cervical esophagus LPMV smooth muscle begins to appear. Once inside the thorax no further PAM is found in the esophagus: it is surrounded by smooth muscle.
This behavior fits with observations about the unique contribution of lateral plate mesoderm to the muscular suspension of the pectoral girdle from the head: sternocleidomastoid and trapezius. These hypoaxial muscles may be possibly be derived from lateral plate mesoderm rather than PAM. As such they represent levels c1–c2 for sternocleidomastoid and c2–c6 for trapezius. Perhaps the limited availability of LPM at cervical levels for esophagus is due to “mesenchymal diversion” for the external neck.
Larynx, Trachea, and Thyroid
Key events by developmental stages have been correlated to Carnegie stages [50].
Stage 11
Ventral epithelium of the foregut thickens as respiratory primordium.
Stage 12
Diverticulum develops in the respiratory primordium. Primitive pharyngeal floor is isolated and will become epiglottis. Cephalic RD becomes the infraglottic region and gives off bronchopulmonary buds.
Stages 13–14
Bronchopulmonary buds migrate downward as the carina. Space between RD and carina becomes the trachea. Growing esophagus and trachea constitute a watershed zone vulnerable to vascular compromise leading to esophageal atresia, trachea-esophageal fistula, tracheal stenosis, or even agenesis.
Stage 15
Ventral primitive laryngopharynx is compressed bilaterally by the cartilages and muscles of the larynx.
Stage 16
Epithelial lamina obliterates primitive laryngopharynx ventral-dorsal leaving behind a narrow communication between the hypopharynx and infraglottis. A space appears, the laryngeal cecum, between the arytenoids and the epiglottis. The space deepens along the ventral aspect of the epithelial lamina. The primitive laryngopharynx has the shape of the letter “T.”
Stages 17–18
Laryngeal cecum pushes downward to reach to glottis, where it stops.
Stages 19–23
Recanalization of the epithelial lamina takes place from dorsal-cephalic to ventral-caudal. The process terminates at the glottis. Failures in this process can create webs or stenosis.
Fetal Period
When recanalization of the epithelial lamina is complete, communication is established between the supraglottis and the infraglottis. Laryngeal ventricles are outgrowths of the cecum. Innnervation is established with the myenteric plexus at 13 weeks. At 16 weeks the cartilaginous vocal cords, ventricle, and saccule are formed. Fetal swallowing begins. At 6 months the epiglottis has fibrocartilage and fetal breathing is established.
Phylogeny of the Larynx and Mechanisms of Ventilation
Lungfishes have a primitive epiglottis to separate air and food. Soft tissue airway anatomy for extinct tetrapods is not available to us so we have to content ourselves with amphibians. Frogs have short nasal chambers that open through choanae anterior to palate. The amphibian glottis is suspended by lateral laryngeal cartilages that are derived from the fifth arch (no sixth arch). Behind the glottis is a small triangular laryngotracheal chamber that leads directly into the lungs. Inside the chamber are vocal cords. Inspiration is controlled by buccal pumping with expiration a combination of elastic recoil of the lungs plus the contraction of transversus abdominis muscle, reincarnated from placoderms (Fig. 10.137).

Phylogeny of ventilation. Tetrapods invented the transverse abdominal muscles…homologous with those of placoderms. Enhancement of exhalation preceded that of inhalation. [Reprinted from Kardong K. Vertebrates: Comparative Anatomy, Function, Evolution, seventh ed. New York, NY: McGraw-Hill Education; 2015. With permission from McGraw-Hill]
Neck length increases in amniotes. In reptiles, the laryngotracheal chamber subdivides into larynx, with supporting arytenoid cartilages, a cricoid, and trachea (Gr. tracheia = rough artery). The developmental advantage of the cartilage rings is to maintain the patency of the lengthened airway. Reptiles make use of intercostal muscles to expand and contract the chest cavity. Crocodiles have unusually paired diaphragmatic muscle (unrelated to mammalian diaphragm) which stretch longitudinally from liver back to the pelvic girdle and open the pleural cavity.
A secondary palate for separation air from food appears in therapsids and continues into mammals. Enlarged nasal cavities with conchae warm, humidify, and cleanse the air. The mammalian larynx, characterized by a new thyroid cartilage, houses an epiglottis and vocal folds.
It is not generally appreciated that lungs are a basic feature of all bony fishes. Air breathing is present in primitive ray-finned fishes, such as Polypterus and Amia, as well as the lungfishes. In Amia, the proportion of O2 taken from lungs versus gills is 25% but as the water warms up this can rise to 75%. Actinopterygians use a 4-stroke buccal pumping mechanism and external oblique muscles to ventilate. Lungfishes have hearts and circulatory anatomy similar to amphibians and a primitive epiglottis to separate food and air. Ventilation is accomplished using a 2-stroke buccal pump with additional internal oblique muscles for forced inhalation.
The tetrapod transition took place in warm shallow waters where aqueous oxygen supply was poor so evolutionary pressure favored the use of the lungs. Early tetrapods continued to ventilate the lungs using buccal pumping instead of ribs. This is related to the loss of opercular-gular bone series permitting the mouth to gape open more widely, and fusion between the braincase and palate. Tetrapod ventilation was made more efficient by a third muscle layer, the transverse abdominals, muscles that existed long before in placoderms but which were expressed during fish evolution. The transverse muscles assist in forced exhalation. Costal ventilation appeared in the amniotes and was supported by inhalation and exhalation. The final innovation is that of the mammalian diaphragm.
Phylogeny of the Lungs
Throughout vertebrate evolutions, lungs arise as endodermal outgrowths from the gut. Specifically, the transition takes place below the fifth arch, that is, as the boundary between r11 and the first truncal neuromere. In mammals, this is the r11–c1 junction. In almost all cases, the lungs are paired, and ventral. In mammals, they connect with the pharynx via trachea (Fig. 10.138).

Phylogeny of the lungs. Lungs are a feature of all bony fishes. [Reprinted from Kardong K. Vertebrates: Comparative Anatomy, Function, Evolution, seventh ed. New York, NY: McGraw-Hill Education; 2015. With permission from McGraw-Hill]
The primal origin of lungs is the gas bladder which appears with ray-finned fishes and is connected via a pneumatic duct to the gut. When functions to maintaining buoyancy, that is, the position of the fish in the water, it is referred to as the swim bladder. When its internal walls are infolded and heavily vascularized, it is referred to as a respiratory bladder, or lung. Swim bladders are dorsal and generally singular, with venous blood returning into the systemic circulation. Lungs are ventral and paired; their blood return and enters the heart directly. Both bladders originate from the gut and have a common form of innervation. Fish evolution reveals repeated episodes of reversal between the two forms, depending upon the respiratory versus buoyancy requirements.
Neither form of bladder is present in agnathans, placoderms, or chondrichthyans (sharks). Lungs are actually the most basal form. They arose in the common ancestor interposed between placoderms and the two lines of osteichthyans, actinopterygians, and sarcopterygians. Lungs begin with sarcopterygians. The primitive form, seen in the Australian lungfish is single and dorsal (one side is atrophic) but in the more derived South American and African lungfishes a dorsal–ventral hox gene places paired lungs in the final, ventral position. The swim bladder in the ray-finned fishes may have evolved separately or as a derivative of the lung.
Neuromeric Rationale of the Respiratory System
The pulmonary diverticulum from the foregut at r11–spinal1 is universal among tetrapods. Recall further that the neuromeric transition point for the brachial plexus and upper limb is at spinal 4 (C4 in mammals) and that cucullaris spans from sp4 forward to occiput. Recall that the pharyngeal arches share a common mapping system of distal-less (dlx) genes. Thus, neuromere c1 may be located at a transition posterior to the termination of the dlx system. Bookmark this point: interesting and instructive parallels exist between the location of the tetrapod partitioning mechanism and the respiratory system of insects (Fig. 10.139).

Evolution of the Dlx code and serial homology of phyarngeal arch endoskeletal elements. The Dlx code arose along the gnathostome stem, and was primitively deployed in all pharyngeal arches. The evolutionary relationship between the Dlx code of gnathostomes and the nested expression of DlxA-D in the pharyngeal arches of lamprey remains unclear. Dorsal (Dlx1–2-expressing) and ventral (Dlx1–6-expressing) domains of the Dlx code would have primitively given rise to dorsal “epimandibular,” “epihyal,” and “epibranchial” elements and ventral “ceratomandibular,” “ceratohyal” and “ceratobranchial” elements (in the mandibular, hyoid and gill arches, respectively), while intermediate (Dlx1, 2, 5 and 6-expressing) domains would have given rise to the region of articulation between these elements. The primitive role for the Dlx code in patterning the mandibular, hyoid and gill arch endoskeletal segments has been conserved in elasmobranchs, and presumably in non-teleost actinopterygians (e.g., paddlefish), while post-hyoid arch expression of the Dlx code has been modified or obscured in amniotes (e.g., mouse), and possibly in teleosts. at, ala temporalis; cb, ceratobranchials; “ch,” hypothetical ceratohyal; ch, ceratohyal; “cm,” hypothetical ceratomandibula; eb, epibranchials; “eh,” hypothetical epihyal; “em,” hypothetical epimandibula; hm, hyomandibula; in, incus; mk, Meckel’s cartilage; pq, palatoquadrate; sp., styloid process; st, stapes. [Reprinted from Gillis AJ, Modrell MS, Baker CVH. Developmental evidence for serial homology of the vertebrate jaw and gill arch skeleton. Nat Commun 2013; 4:1436. With permission from Springer Nature]
First off, foregut partitioning begins at the node between weeks 3 and 4 of development.
The site of the respiratory diverticulum along the ventral foregut wall at level c1 is marked by the expression of Nkx 2.1 Just opposite, the dorsal foregut wall contains the expression of Sox-2. Wingless (wnt) signals from surrounding lateral plate mesenchyme cause Beta-catenin to accumulate in the ventral gut tube between r11 and c1. B-catenin is required for separation to occur. It acts like the Pied Piper to attract the diverticulum. If expressed ectopically, it can form extra lungs.
Meanwhile, interaction between the epithelium of the future esophagus and trachea with surrounding mesenchyme determines the type of epithelial lining. Mesenchyme surrounding the dorsal gut tube contains transcription factor Barx1 which insures the production of soluble Frizzle-related proteins (sFRPs). The sFRPs bind to Wnt so they cannot access the gut tube, thereby blocking their activity. In contradistinction, ventral mesenchyme does not produce sFRPs. Note how this mechanism could explain the dorsal versus ventral positioning of the gas bladder in evolution. The presence or absence of Wnt signaling directly affects the histology of the epithelium. When blocked, the lining of dorsal esophagus is squamous epithelium. When active, Wnt creates a ciliated respiratory epithelium that lines the trachea (Fig. 10.140).

Trachea appears immediately after lung buds at stage 13. Separation from esophagus is complete at stage 14. Tracheo-esophageal fistula can occur at this time. Descent of the trachea into the chest is complete at stage 23. D Roles of Wnt signaling and mesenchyme expressing Barx1 causes differentiation of epithelia. Presence of Barx1 blocks Wnt and causes region to become esophageal. [Reprinted from Gilbert SF, Barressi MJF. Developmental Biology, 11th ed. Sunderland, MA: Sinauer; 2016. Copyright © 2016. Oxford Publishing Limited. Reproduced with permission of the Licensor through PLSclear]
In the later fourth week the posterior aspect of the respiratory diverticula produces lung buds. The lung buds in turn induce from the LPM just dorsal to the paired mesodermal ridges. These approximate one another in the midline and fuse in a dorsal to vental manner. This makes sense because the LPM folds are posterior to the lung buds. The fusion of the folds creates a septum that separates trachea from esophagus.
Lung development is also homeotic. Combinatorial patterns of hox genes Hoxa3 to Hoxa5 and Hoxb3 to Hoxb6 are responsible for regional specification of the respiratory track such as bronchi to bronchioles and the lobes of the lung. Furthermore, coding exists in the mesoderm surrounding the tract. The more proximal zone inhibits branching and results in trachea whereas more distal mesenchyme supports branching.
A Sidebar on the Thyroid
It is significant that the NKx 2.1 zone in the c1 ventral foregut is associated with localization of the thyroid gland. Thyroid tissue likely contains r10–r11 markers from the fifth arch. Thyroid tracks toward the tongue along the hyopobranchial genetic tract, the same one used by the strap muscles running between the manubrium and the 1st/second arch interface in the tongue. Thryoid tissue probably migrates in company with the occipital myoblasts, squarely in the midline. When it returns to the neck it follows this selfsame pathway backward to manubrium and into the chest.
Lessons from Drosophila
Nomenclature of Drosophila genes: distal-less (dll), buttonhead (btd), trachealess (trh), wingless (wg), and ventral veinless (vvl).
The arthropod (insect) exoskeleton contains ectodermal tracheal placodes, numbering 10 in Drosophila. These become spiracles that penetrate the skeleton to reach a longitudinal tracheal system running the length of the body. Spiracles congregate around the three pairs of legs with the two thoracic spiracles associated with the first pair of legs. Respiratory spiracle/trachea cells and leg cells exist in a common pool and are related. Tracheal precursors develop where trh and vvl are expressed. Leg precursors appear where Dlli is expressed. It appears that the decision to make trachea versus legs depends on the presence of wingless expression. Wg blocks trh and therefore prevents trachea formation; it promotes Dll and causes legs to develop (Figs. 10.141, 10.142).

Indian moon moth (Actias selene) showing respiratory spiracles transmitting O2 through exoskeleton. These arise from placodes and lead into longitudinal internal trachea which subdivides to supply all parts of the body. It is the homolog of lungs. [Reprinted from Wikimedia. Retrieved from: https://commons.wikimedia.org/wiki/File:Actias_selene_5th_instar_spiracles_sjh.jpg. With permission from Creative Commons License 2.5: https://creativecommons.org/licenses/by-sa/2.5/deed.en]

To trachea or not to trachea? Insect exoskeleton tiny valves called spiracle which admit oxygen. These are lines with hairs to keep dust out. O2 and CO2 are transported by tracheae; these terminate in tracheoles where gas exchange occurs. Role of wingless (wg) in repressing tracheal fate. In Drosophila, 10 tracheal placodes give rise to spiracles that connect with the trachea. Note two thoracic placodes are associated with first pair of legs. Tracheal cells are specified by trachealess (tll) and ventral veinless (vvl) along the body axis. Tracheal cells and leg primordia cells exist in a common pool. Legs form where Dll is expressed. The decision trachea versus legs depends upon the activation state of the wingless (wg) signaling pathway. In aquatic arthropods such as crustaceans, respiratory gills and appendages are also associated. Role of wingless (wg) in repressing tracheal fate. Genes: distal-less (dll), buttonhead (btd), trachealess (tll), wingless (wg) and ventral veinless (vvl). Wingless represents a “switch” between trachea and appendages. When not expressed respiratory system develops. Note relationship between arthropod distal-less (dll) and vertebrate distal-less (dlx) that is expressed in the pharyngeal arches. Cells of the respiratory system in Drosophila arise in close association with cells that give rise to the extremities. In aquatic arthropods such as crustaceans, respiratory gills and appendages are also associated. Genes: distal-less (dll), buttonhead (btd), trachealess (tll), wingless (wg) and ventral veinless (vvl) [Courtesy of Science Photo Library].
Primitive aquatic arthropods such as crustaceans demonstrate a similar relationship between the respiratory structures and appendages [51]. Crustacean gills develop as a dorsal lobe of their appendages (epipods). Franch-Marro et al. [52] found homologs of Drosophila tracheal inducer genes expressed in crustacean epipods. This suggests that tracheal systems and gills in arthropods evolved from a common precursor.
The relationship between the forelimb and the anterior-most component of the respiratory system becomes relevant when we consider the role in humans of Wnt signals in (1) the partition of foregut into trachea and esophagus and (2) the creation of respiratory epithelium all the way down the trachea. This event takes place precisely at the neuromeric levels c–c4 (and beyond) associated with the forelimb. The primal events involved in the creation of the gas bladder likely involved similar mechanisms at the very same neuromeric level.
Coda: Thoughts on the Assembly of the Neck
In our discussion so far, we have considered the neck as a series of structural components: bones, nerves, muscles, arteries, and the endodermal-encoded tubes (the esophagus and respiratory tract). In this final section let’s consider how these structures are assembled, and how this assembly changed from primitive tetrapods to the amniote condition. Some of this story is by way of review (based upon what we know from the fossil record), some of the stories is best-guess speculation…the final picture awaits future research. The neck being a truly integrative structure, our story begins with concomitant events that changed both the vertebral axis and the pectoral girdle.
Fish vertebrae are seemingly monotonous, the only difference between trunk vertebrae and those of the tail is the presence of a hemal arch in the latter as a protective device for the aorta to prevent it from kinking during swimming. Yet, beginning with transitional form Tiktaalik, a drastic transformation takes place in vertebral column causing specialization. From the earliest true tetrapods such as Ichthyostega onwards, cervical, thoracic, lumbar sacral, and caudal regions can always be identified. In the future neck, the homeotic transformation that contributes to new cervical vertebrae is an anteriorization of identity of truncal somites, regardless of whether the total number of vertebrae changes or not.
The evolution of the neck results from two fundamental processes.
-
Homeotic transformation refers to the anatomic changes that results when neuromeric levels formerly responsible for the structures of the trunk are reassigned to produce new derivatives with new functions. Transformation implies just that: it is not an addition of a level but a change in the definition of that level from trunk to neck. Bear in mind that the transformation of a somite with a thoracic homeotic signature into one with a cervical hox code represents not just a change in osseous structure but a change in the entire portfolio of tissues at that level.
-
Homeotic duplication refers to the repetition of units within the same anatomic region resulting in new anatomic structures. It is unique to mammals where the neuroanatomic contents of levels C4–C5 are repeated to create and add the median nerve to the brachial plexus and more complex system of muscles. The result of duplication is to force the expression of anatomic structures into more posterior neuromeres.
Key Evolutionary Steps Determining the Size of the Mammalian Neck
Step 1: the first iteration of parasegmentation
In the pre-tetrapod evolution of late sarcopterygians, a fourth occipital somite appears, the result of the incorporation of the first truncal somite into the skull. Recall the first three occipital somites do not manifest parasegmentation. They are fused together as a unit. In sarcopterygians the fourth somite demonstrates a new characteristic: the rostral part of S4 centrum is loose. For the first time in evolution, it breaks apart and fuses with S1–S3. This creates the modern cranial base, S1–S2–S3–S4R. The caudal part of S4 centrum is now free to interact with rostral half of S5 centrum. The recombination of S4C with S5R creates a modified spinal vertebra, the protoatlas, S4C–S5R. Although lungfishes do not have neck joint…we are just waiting for Tiktaalik to come along…the protoatlas creates the conditions for a joint. There is nothing special about the appearance of protoatlas. It looks like just another spinal vertebra. But its subsequent dissolution caused a revolution.
Step 2: the second iteration of parasegmentation
Lightning strike twice. Tetrapods apply the same mechanism to protoatlas. It splits apart…asymmetrically. Instead of becoming totally intracranial…in effect becoming a fifth occipital somite…its rostral centrum contributes to foramen magnum. Voila! A joint is created. This process leaves behind a fragment of the neural arch of protoatlas C0, referred to as the proatlas, intercalated above the new vertebra in contact with the skull, Atlas, now has the formula S5C–S6R and a very different shape.
We have seen in the fossil record, in humans, an intact proatlas with a total of eight cervical vertebrae has been documented clinically (Fig. 10.32). Proatlas appears as part of a proatlas–atlas–axis complex followed by three additional cervical vertebrae, C3–C5. This model is seen in Ichthyostega and Acanthostega. Thus, the common anamniote ancestor of amniotes had 5 cervical vertebrae. The boundary between cervical and thoracic ribs is thus defined by the third spinal nerve of the brachial plexus, br3; this is spinal nerve C5. The boundary of the thorax is C5–T1. The evolution of extant amphibians such as frogs and salamanders proceeds in the opposite direction with reductions in the brachial plexus formula (vide infra).
Step 3 The amniote transition was accompanied by changes in the neck. In both the reptilian/avian (diapsid) line and the mammalian (synapsid) additional numbers of cervical vertebra were produced and a caudal shift of the thorax took place. Diapsids are more prolific. The ostrich has 20 cervical vertebrae with the first thoracic vertebra located at S25, In the mammalian line, non-therapsids have 5, therapsids have 6, and cynodonts achieve the final mammalian number of 7.
We can follow this evolutionary story by making use of two anatomic principles.
-
The neuromeric position of the pectoral fin or upper extremity is located at the head-thorax boundary in fishes and the neck-thorax boundary in tetrapods. Anatomic differences between cervical and thoracic ribs permit the identification of the boundary in all fossil taxa.
-
A universal pattern of the brachial plexus exists in all amniotes (mammals being an important exception) that allows one to work backwards (rostral) from the trunk to determine the identity of the neuromeres supplying the upper limb. Brachial plexus neuroatomy faithfully recapitulates the evolution of the neck.
Caudal Shift and Homeotic Duplication of the Brachial Plexus in Mammals
The brachial plexus of all non-mammal amniotes consists of four spinal nerves, three of which are of cervical origin and the last one from neuromeric level t1. These are referred to as br1–br4. Pairing of br1–br2 creates the radial nerve for extensors while pairing of br3–br4 creates the brachial nerve for flexors. Thoracic neuromere t1 is responsible for br4. Brachial and radial nerves are interconnected and supply the simple adductor and abductors muscles groups of the limb. This simple configuration goes back to the beginnings of vertebrate evolution.
Even though the mammalian brachial plexus is more complex, the same rules hold true. Mammals duplicate roots br1–br2 to create a new median nerve in a more rostral position. In the process, the rest of the plexus is pushed back to two neuromeric units, and a 6-nerve brachial plexus results. Nevertheless, the penultimate nerves c8 and t1 continue to define the neck–trunk boundary.
Let’s look at how this system arose. Recall that bony fishes, osteoichthyes, evolve in two lines: actinopterygian (ray-finned) fishes and sarcopterygian (fleshy-finned) fishes. In the basal state, both lines have only two muscles associated with the pectoral fin, one abductor and one adductor. These are innervated by a plexus of nerves, designated as either occipital (Oc) or spinal (Sp), depending upon the location of their roots in the medulla or spinal cord.
Using lipophilic dyes [37] mapped out the alignment of the fin bud, myototomes, and neuroepithelium in four species of actinopterygians, from the primitive basal paddlefish, Polydon spatula, to the highly derived teleost midshipman, Porichthys notatus. The pectoral motoneurons shared a conserved pattern with no migration during embryogenesis. Porichthys shows us a simple system of two occipital roots and spinal roots distributed across the hindbrain-spinal boundary. Although the number of roots could vary during development, all were derived from two distinct populations corresponding to the adductor and abductor muscles. Furthermore, these motoneurons formed a single column distinct from the posterior r10–r11 dorsal motor nucleus of vagus. Four myotomes were supplied. The anatomic parallels with tetrapods are obvious.
The pectoral fin in the sarcopterygian line is much more complex. It has recognizable bones (stylopodium, zeugopodium, and autopodium) connected with additional joints, and controlled by new muscles (arising by subdivisions for the pre-existing muscle mass). This condition exists today in the African lung fish, Dipnoi species Protopterus dolloi. Lungfish have three occipital neve roots that combine into a single pectoral nerve Oc3 which then joins spinal nerves Sp1–Sp3 to form a plexus. The presence of an additional third occipital nerve is of great significance. Although not confirmed experimentally, the origin of the fourth occipital somite probably takes place in late sarcopterygians just before the tetrapods.
These findings are consistent with the hypothesis that a dual hindbrain-spinal origin for pectoral motoneurons in jawed fishes was translocated backward into a spinal-only innervation of the tetrapod forelimb. The sequence Oc1–Oc2 and Sp1–Sp2 become br1–br4. We can imagine a scenario in which hox shifts cause the occiput of the fish skull to change from S3 to S5R. This introduction of new programming causes a disruption in levels r10–r11. The genetic program for the peripheral hypobranchial muscles of rectus cervicis in fishes is kicked back in tetrapods to c1–c4. At the same time, the neuromeres of the tetrapod hindbrain (r8–r11) assume a more modest role in supplying the hypobranchial muscles of the tongue. The question arises, after these initial events, what happened next?
Our ability to reconstruct the life history of Devonian tetrapods is limited due to inadequate fossil evidence regarding the cervical spine and neck joint. Nonetheless, Acanthostega represents a model for the initial iteration of the brachial plexus, as the entire length of its skeleton has been reconstructed. It had a well-defined atlas–axis complex. The arch of atlas is small and, while there are no pleurocentra, the intercentrum is robust. No proatlas has as yet been identified. Five cervical vertebrae are present. Nothing is known about the craniovertebral junction in Ichthyostega. http://pondside.uchicago.edu/oba/faculty/coates/Coates_1996.pdf
Our next clues come from Greererpeton a crown tetrapod of the mid-Carboniferous. Here a small proatlas was present and the atlas arch embraces that of the axis. This enlarged axis supported expanded muscle insertions for greater control of the head. In the transition to the amniotes atlas pleurocentrum and axis intercentrum became firmly fused as dens to support the ring of the atlas. The neural arch of axis took the decision to marry up its pleurocentrum, a condition persisting to this day. Proatlas continues to be represented in the stem amniote Paleothyris and continues into the synapsids as far as Dimetrodon.
The basic pattern of four spinal nerves and the position of the br3–br4 nerves at the cervicothoracic junction is shared by virtually all living amniotes. The only difference between taxa is the total number of cervical neuromeres: the last three will always contain br1–br3. We know that amniotes start out with five cervical vertebrae with an atlas/axis complex and a brachial plexus br1 (spinal nerve 4) is located at c3. This situation leaves three spinal nerves available for cervical plexus. Thus: sp1 is below proatlas; sp2 is below atlas; and sp3 is below axis. Exactly as in humans. The sleight of hand in mammalian evolution is a duplication of br1 and br2 to produce six roots and two additional cervical vertebrae.
Frogs have rudimentary neck and a deemphasized upper limb with a two-nerve brachial plexus supplying a simple flexor/extensor. Brachial plexus consists sp2 below the axis and sp3 below first thoracic vertebra, thus conserving the piscine model with two roots straddling the cervicothoracic junction. Nota bene: The first spinal nerve never innervates the limb. This is probably because the forward shift of proatlas takes that neuromere out of the genetic territory of Hox5, the territory of upper extremity. Formerly known as the “fin fold field.” With proatlas plastered up against the skull, sp1 emerges cranial to the atlas. Furthermore, the existence of sp1 above atlas demonstrates that urodeles incorporated the protoatlas, C0, with sp1 left behind. This makes sense because, in all taxa, first spinal nerve is simply not programmed for the extremity. Its purpose is to supply the axial muscles controlling the head.
-
In sum: the inference of amphibians is that the presence of an atlas is evidence of the prior forward shift of proatlas.
-
first spinal nerve in frogs contributes to hypoglossal.
-
The amphibian brachial plexus has two nerves; it begins with second spinal nerve: (br1 = sp2 and br2 = sp3).
Pre-mammals start their brachial plexus with br1 as sp4 just below C3. With evolution, they proceed to add duplicate b1–br2 creating the formula. This forces the transfer of the original 4-root plexus from spinal nerves Sp4–Sp7 backward to Sp6–Sp9. But even though the hox code for c4–c5 changes, the neuroanatomic contents remain much the same. Thus c4–c5 combine in the same way to produce a new structure, median nerve. Even the original cross-bridge between radial and brachial is duplicated, with median connected to brachial (now renamed the ulnar) and radial connected to brachial.
-
The formula for the mammalian brachial plexus is: br1′–br2′–br1–br2–br3–br4.
-
Supracoracoideus nerve (phrenic nerve) retains its ancestral state at c4 (br1′).
-
Subscapular nerve, remains attached to br1 so it shifts backward one neuromere to c4–c5.
-
Median nerve br1′–br2′ at roots c4–c5 is an exact carbon copy of radial nerve br1–br2 which has been shifted backward two neuromeres at roots c6–c7.
Mesenchymal consequences of neck development: making room for mesenchyme.
In anamniote evolution, the neck remained simplistic. Salamanders have a three-spinal nerve brachial plexus with little complexity of either extremity. This is reduced to 2-spinal nerve in frogs. Snakes lose the extremities altogether. Note that they lose the lateral motor column as well. In early tetrapods, the shoulder girdle is closely tucked into the head. These animals used buccal pumping as a means to drive air into the lungs. The musculature required for this was similar to that of the gill apparatus and these muscles were inserted into the pectoral girdle. The basic structures of the esophagus and the airway (the glottis) are present in amniotes. As the neck expands, additional neuromeric units of endoderm and lateral plate, like a candy cane, simply add more stripes to the structures transmitted through the neck and into the chest. In the mammalian line lengthening of the neck was accompanied by repositioning of the extremities beneath the body expansion of the brachial plexus for more greater complex control of the forelimbs, and the invention of the diaphragm with enhanced ventilation (buccal pumping was abandoned) supporting higher oxygen demand, as in a cheetah running at top speed.
Mechanism: Is There a Common Event?
Previously, we discussed the phylogeny of the scapula and clavicle. We turn now to the evolution of their overall positioning. How does this square with the expansion of in the number of cervical vertebrae? The story of fins-to-limbs is a sequence of events: (1) the addition of the fourth occipital somite, (2a) the creation of an articulation between skull and spine, (2b) loss of bony connections between the head and pectoral girdle, (4) addition of cervical units, (3) repositioning of the pectoral girdle and brachial plexus, and (5) development of a specialized pectoral girdle. Let’s look at this sequence in greater detail.
The first evidence of paired pectoral fins with attachment to the axial skeleton occurs in the agnathan armored fishes, the osteostracans. As we know, these creatures had a massive head shield of dermal bones. A single endochondral bone, the future scapulocoracoid, articulated with the posterolateral side of the head. In a later group of placoderms, the †arthrodires, two important changes are noted. First, dermal elements are incorporated into a cleithral group (supracleithrum, postcleithrum or anocleithrum, dorsal cleithrum, and ventral cleithrum) and a clavicle. The latter bone is thought to arise from a modified dermal scale. The dermal elements are much larger than scapulocoracoid. Second, the entire pectoral complex is anchored to a single cranial bone, the posttemporal.
In subsequent osteoichththyans, the dorsal and ventral cleithra unite into a single bone. The dermal bones become regularized and identifiable. Anterior to the pectoral girdle the opercular and postopercular bones, part of the 4-bone opercular complex covering the gills. Above the pectoral girdles an extrascapular series is interposed between them it and the postparietals.
Dissociation of the pectoral girdle takes place in the Devonian period with the transitional sarocopterygian, Tiktaalik. Because it had well-developed gills and no jaw features of tetrapods, this important creature stands between the true crown sarcoptyerygians, Panderichthys, and the tetrapod Acanthostega. This is where we shall place our attention. Gone are the intervening extrascapulars, opercular and subopercular, eliminating all bony connection between pectoral girdle and the skull. The tabulars and postparietals move into position as a replacement for the extrascapulars. These provide enhanced insertions for cervical muscles.
Many changes are noted in the skull. The postfrontal is elevated above the level of the orbital margin, forming a ridge consistent with dorsalization of the eyes for better vision on land (more on this concept later). The ethmosphenoid lengthens relative to the oto-occipital complex. This consistent with the development of a snout. The dermal intracranial joint is lost and the palate is less mobile. The effect of this crocodile-like head is to increase the biting surface. The middle ear of tetrapods is anticipated by a widening of fish spiracle as a potential communication into the pharynx and alterantions in the hyomandibula which is no longer bomerang-shaped but short, straight, and stout. It is the precursor for stapes in Acanthostega.
Bone changes also have respiratory implications. The skull is wider with a more voluminous buccal cavity. Expansion of gular plates and branchial elements support more aggressive buccal pumping of air. In obligate gill-breathers hyomandibular is involved in the forces associated with pumping of water through the gills. This function is unnecessary for terrestrial life so hyomandibula is reassigned to the ear. Decreased reliance on water pumping is associated with increased use of air breathing.
Tiktaalik has approximately 45 presacral vertebrae (Acanthostega has 30). Cervical vertebrae are not documented. The ribs are expanded and wider than those of Panderichthys. Plate-like flanges extend caudally. These changes presumably assisted in thoracolumbar support necessary for weight bearing.
Finally, there are immediate advantages of cranial mobility and a neck. Fishes can readily orient themselves in water to position the mouth for prey capture. On land, body position is fixed so the head position becomes independent. Increasing sophistication of control of extremities. Tiktaalik lived in the mud flats and was able to lift itself out of the water and snap at its target [53].
What mechanisms can be inferred from Tiktaalik? Disconnection of the pectoral girdle from the skull involves the loss of three series. The posttemporal is a dermal bone of the temporal series that directly connects the pectoral girdle with the r6–r7 opisthotic. Remaining behind in tetrapods from this series are intertemporal, supratemporal, and tabular bones. Medial and lateral extrascapular bones are gone as are the opercular bones. But the mesenchyme is likely not lost…it is merely reassigned. Experimental work tracing the homeotic signature of the posttemporal may reveal the final destiny of this mesenchyme. Could it be converted to fascia or muscle?
Addition of the fourth occipital somite and first cervical somite likely occurs as a two-step process, as suggested by the lungfish, protoatlas brought inside the braincase with the proatlas added in later. It could also have happened simultaneously. In any case, this brought additional homeotic complexity to the occiput. S4 joins with S3 to form the exoccipitals. By the time of Greererpeton the notochordal cotyle of the occiput disappears and the S4C moiety of the exoccipitals combines with S5R to make a single occipital condyle. Could the introduction of new paraxial mesoderm into the occiput be a contributing factor to the demise of the posttemporal?
Regarding lengthening of the neck, the selective advantages of a more cervical vertebrae giving a more mobile head for food gathering are obvious. Apart from the synapsids which fix the number at 7, diapsids are wildly diverse. The plesiosaur †Muraenosauraus had 76 cervical vertebrae! Snake lack them altogether. We have discussed the homeotic mechanisms previously.
The variations introduced by the reassignment of additional cervical neuromeres have little effect on the epaxial anatomy. Intersegmental muscles are simply added on. All the variations take place seen in hypaxial structures. The position of the pectoral girdle to the axial skeleton remains hypaxial, despite the apparent dorsal position of scapula. For this reason, all muscles connecting it to the head, neck, or trunk are innervated by accessory nerve r8–r11 (sternocleidomastoid, trapezius) or ventral roots of c1–c4 (the straps). Muscles from c5 to c8 (brachial plexus) that have primary insertion on scapula connect with the arm. Thus, pectoral girdle can be thought of as an island connecting two muscle groups: those of proximal cervical plexus with those of distal brachial plexus.
Dermatomal distribution is a particularly good example of hypaxial versus epaxial variation. As upper extremity is hypaxial, its entire skin envelope is hypaxial as well. Thus, ventral neck skin is mapped out into four quadrants representing C2–C4, whereas dorsal neck skin is a smooth continuum from C2 to C8 to T1–T12. Each dermatome is supplied by a segmental dorsal branch. All remaining ventral skin from C5 to T1 is expropriated into the arm. Therefore, a sharp boundary exists over the infraclavicular skin between the distribution of C4 and that of T2. The entire hypaxial region of C2–T1 is innervated by means of either the cervical or brachial plexus. Segmental ventral innervation does not recommence until level T2.
Finally, we must consider the simultaneous changes that took place within the upper limb itself. The homeotic regulation of appendicular development has been extensively documented. The homeotic code for each level consists of a combination of hox genes. These spatially distant regions may share homeotic genes common to both. What is certainly true of the tetrapod transition is that the axial patterning process controlled by homeotic genes affect patterning of multiple tissues in all three body axes during early organogenesis. Even minor disturbances can cause serious downstream effects. For this reason, the number of cervical vertebrae in mammals is highly conserved. At stake are many traits involving craniofacial structures, larynx/trachea/lungs, the kidneys, and limbs. This explains the large number of craniofacial syndromes reported.
The Evolutionary Impact of an Enhanced Visual System
Life for predators is a simple equation: you eat what you kill and you can’t kill what you can’t perceive. Water is a difficult medium for visual hunting. Low levels of light and the density of the medium pose extreme limitations on predation. MacIver and Schmitz applied a novel approach to understand the transition to terrestrial life: the evolution of visual acuity. Measurements of eye socket volume and the anatomic position of the spiracle in 59 tetrapodomorph taxa that span the water-land transition yield startling results. Eyes tripled in size with vision through air improved improving a one million-fold increase in the amount of space within which objects (prey) could be perceived. The orbits, though still lateral, moved upward on the skull. At the same time changes in breathing that evolved as a compensation for the reduced oxygen content of the Devonian favored crocodilian predatory behavior by looking for prey just above the water line [54] (Fig. 10.143).

Evolution of visual range. [Reprinted with permission from MacIver MA, Schmitz L, Mugan U, Murphy TD, Mobley CG. Massive increase in visual range preceded the origin of terrestrial vertebrates. Proc Natl Acad Sci U S A. 2017 Mar 21;114(12):E2375–E2384. With permission from Copyright © 2017 National Academy of Sciences]
It all started with the sarcopterygians (no surprise there). The changes are seen in three stages: finned tetrapods, transitional tetrapods, and digital tetrapods. From the osteolepiform Eusthenopteron to the panderichthid Panderichthys orbital volume went up 1.42×. The transitional Tiktaalik had a volume increase of 1.43×. This exponential expansion peaked with Acanthostega which has an orbital volume 1.52× baseline. Enhanced socket size correlates directly with larger eyes and larger pupils (Fig. 10.144).

Resizing and repositioning of the orbit and spiracle Having invaded shallow waters, with access to more, translocation of eye sockets moved to the top of the skull, providing upward vision, as shown here for Panderichthys. Low oxygen conditions favored, breathing through spiracles dorsalized to just behind the eyes in the elpistostegalians, as shown here for Tiktaalik. With continued surfacing and selection of changes to lens and cornea enabling a focused image of prey, during the 12-My transition from finned to digited tetrapods, enlarged eye sizes developed, enabling the full power of long-range vision. Simultaneously, with the selective advantages of limbs, animals like Acanthostega became better suited for longer forays onto land, culminating in more terrestrial forms, such as Pederpes, 30 My after Tiktaalik. Spiracles (green), the precursors of the Eustachian tube, were likely used for breathing at the water surface while using aerial vision. Eyes dorsalize between Eusthenopteron and Tiktaalik, and enlarge in tetrapods. S Total animal lengths are between 50 cm and 1.5 m (not drawn to scale). Age spans from 385 My for Eusthenopteron to 355 My for Pederpes. [Reprinted with permission from MacIver MA, Schmitz L, Mugan U, Murphy TD, Mobley CG. Massive increase in visual ragne preceded the origin of terrestrial vertebrates. Proc Natl Acad Sci USA. 2017 Mar 21;114(12):E2375–E2384. With permission from Copyright © 2017 National Academy of Sciences]
The depth of visual field for Eusthenopteron was roughly its body length, Although these evolutionary changes led to a tripling of eye size, they were not of great use underwater. However, when aerial vision became available, depth of field increased to 100 body lengths and the gain in visually monitored space under light conditions increased 1,000,000-fold.
The position of the orbits changed as well. Beginning with Panderichthys the eyes moved to the top of the head. The eyes of Tiktaalik are definitely crocodilian. Elevated bony prominences caused by the prefrontal and postfrontal bones appear at this time, the harbinger of supraorbital ridges. This proved to be a critical innovation because it permitted Tiktaalik’s eyes to be out of water while at the surface. Of note, the changes seen in the orbits of Panderichthys are part of a more radical restructuring of the face characterized by primarily by elongated jaws, an adaptation for more successful prey capture. The palate widened and the skull became flatter, presenting a lower along the surface of the water [55, 56].
A third factor contributing to enhanced predation was respiration. Water is a difficult medium for oxygen capture compared with air. Not only is it 800× more dense, but its O2 content is 1/30th that of air. Aqueous respiration requires a mass flux 24,000 time greater per unit of extracted oxygen. Devonian conditions were terrible due to a decline in the amount of available oxygen. Sarcopterygians responded by enlargement of the breathing passageways called spiracles. The spiracle was derived from the first branchial cleft between the first and second branchial arches in agnathic fishes. When jaws were invented and the first two arches merged together, the remnant of the cleft became the spiracle. Because it sits in front of the otic capsule (the future ear), it is also known as the otic notch. It tetrapods the spiracle is converted into the Eustachian tube. Repositioning of the spiracle is seen in elpistostegalians. Panderichthys had a flattened skull, resembling that of a crocodile; it could lurk at the water’s edge, acquire oxygen with less energy consumption, and scan the environment for prey. This provided an intermediate step between specializations for aquatic predation and innovations propitious for brief ventures onto land without a commitment to a full-time terrestrial existence.
In sum, enhanced visual acuity, a much larger depth of field, and improved respiration in the aerial environment were propitious for exploration and penetration of the terrestrial environment. Significant neuroanatomic and behavioral benefits accrued. Short-range vision (single body length) permits reactions to just-in-time stimuli. Long-range permits more complex forms of decision-making using different neurocircuitry. Multiple options for pursuit of prey or evasion of predators were developed by trial-and-error behaviors. These are dependent on the hippocampus for memory and learning. Hippocampus is present in the common amniote ancestor of birds and mammals which emerged during the late Carboniferous shortly after tetrapods hit the land. We can therefore postulate that the primitive neural elements of planning were created in part for the “visual explosion” immediately prior to emergence onto land.
The Buena Vista Hypothesis and the Neuromeric Model
MacIver proposed that greatly enhanced vision and improved access to oxygen led to energy-efficient predation and facilitated the transition to land where new forms of information-gathering could be exploited for expansion into multiple ecologic niches [57]. In the follow-up study using orbital volumes the explosion in vision occurred in sarcopterygians while under water, an environment in which this innovation was undoubtedly of value, albeit limited, for hunting. What is striking are the changes in the rest of the face, especially involving dermal bones from r2 and r3 neural crest. Virtually all the changes seen in the orbits reside in changes in size or position of the r2 maxillary bone complex and palate which expanded the space available for a larger globe. Mirror-image changes took place in the r3 mandible which lengthened concomitantly. Because the spiracle was positioned between the r2–r3 first arch and the r4–r5 second arch, it was passively brought forward. The sum of these innovations, plus a modest increase in aquatic vision meant that, under low oxygen conditions, surface predation brought sudden and unexpected benefits.
Restructuring of the sarcopterygian face occurred at the same time as fins became digit-bearing. It is possible that a common denominator exists between the homeotic driver of the pectoral apparatus and the altered hox code of the face. In any case, once these adaptations were present in Panderichtys, the stage was set for Tiktaalik to separate the pectoral girdle from the skull and convert the gas bladder to a lung-like organ. In the process, an articulation was created between the skull and pectoral girdle (Fig. 10.145). Areas for extensive cervical muscle insertion appeared along the dorsum of the skull, permitting head mobility [36, 58]. Further refinements in the craniovertebral joint took place in the digital tetrapods: the neck was born (Fig. 10.146).

Association of the clavicle with the first cervical vertebra. This establishes the original neuromeric coding of clavicle with c1 neural crest. Left; Dorsal view, note the presence of a cervical vertebra flanked laterally by cleithrum. The exact nature of the articulation is not well-defined. Right ventral view showing clavicle closely apposed to the skull beneath the first cervical vertebra. Abbreviations: an, anocleithrum; bb, basibranchial; co, coracoid; clav, clavicle; clth, cleithrum; cbr, ceratobranchial; ent, entopterygoid; hu, humerus; lep, lepidotrichia; mand, mandible; nar, naris; or, orbit; psp, parasphenoid; ra, radius; suc, supracleithrum; ul, ulna; uln, ulnare. Scale bar equals 5 cm [Reprinted from Daeschler EB, Shubin NH, Jenkins FA. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 2006; Vol 440:757–763. With permission from Springer Nature]

Birth of the neck. [Reprinted from Shubin NH, Daeschler EB, Jenkins FA Jr. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. NatureI 2006; 440:764–771. With permission from Springer Nature]
Clinical Impact of Homeotic Transformation
A radiologic survey of autopsy material involving 1389 fetuses/infants with detectable ossification centers was carried out to assess the effects of vertebral anomalies. Homeotic pathologies were extremely common with cervical ribs present in/3 of the cases. Only 20% had normal vertebral counts (7 cervical, 12 thoracic, and 5 lumbar). The severity of malformations was correlated with the severity of vertebra aberrancy. These included craniofacial clefts and mesenchymal deficiencies or excesses. Pathologies seemed to indicate local signaling problems rather than generalized defects of germ layer migration. For example, the development of cleft lip/anterior palate due to a defect of premaxilla likely has to do with downstream event involving the specific vascular axis (medial nasopalatine) rather than a problem upstream in the original neural crest population migrating from the second rhombomere [59].
Vertebral events with disturbances in A-P patterning were likely local with the majority being ipsilateral, involving genetic expressions on one side only, such as unequal ribs, and hemivertebra. These suggest strong coupling between the A–P patterning of paraxial mesoderm and the preservation of left-right symmetry. Craniofacial NC contributes strongly to the construction of multiple structures, including the heart, lungs, GI tract, and brain whereas trunk-derived neural crest has only minor, very conscripted roles. For this reason, craniofacial neural crest malformations had an extremely high degree of vertebral abnormality, more so than those involving trunk-derived neural crest. What all these points to are that early organogenesis is intensively interactive with the process of axis formation because gastrulation and segmentation lay down the neuromeric building blocks for the rest of the body.
Vertebral A–P pattern defects involving homeotic transformations are not randomly distributed over the vertebral column. Only certain regions are affected. Most changes involve frameshifts in one or at most two neuromeric levels. For example, the cervical region can vary between 6 to 8 vertebrae: anything different is lethal. The effects can be physical or biochemical. Shifts in vertebral boundaries usually do not coincide with the boundaries of outlying tissues. Compression of vascular or neural structures can result. Lumbo-sacral transitions are associated with intervertebral disc compression or degeneration and narrowing of the pelvic outlet in women affecting the birth canal. Abnormal boundaries can indicate a reduction in the mesenchyme that affect the expression of gene products critical for development either locally or peripherally. Thus, changes in a single neuromere can wreak havoc on an entire organ system.
Conclusion
In this chapter, we have examined the neck from the standpoint of both its anatomic components as well that of a multi-organ system communicating between head and trunk. In every instance, neuromeric coding was useful in understanding tissue origins and assembly. Mesenchyme from neural crest, paraxial mesoderm, lateral plate mesoderm, and epithelial tissues such as endoderm and ectoderm, all arise from specific sites of the neuromeric map with consequent restrictions placed upon the genetic expression in each neuromeric zone. We then proceeded to review the process by which the neck is constructed, taking care to respect the neuroanatomy of the brachial plexus. Although we are left with many more questions than answers the pathway forward to understanding the development of the neck will surely involve the neuromeric mapping of its parts and the tracing of the final derivatives back to their beginnings.
A somitogenesis model integrating the segmentation clock and determination front. The system of opposing gradients of FGF (fibroblast growth factor)–Wnt signaling (purple) and retinoic acid (green) signaling was proposed to position the determination front (black line) along the presomitic mesoderm (PSM). This particular level is characterized by a signaling threshold at which the cells become competent to respond to the segmentation clock signal and is conceptually similar to the wavefront of the original Cooke and Zeeman model 14. The clock signal is still poorly characterized but probably involves three signaling pathways experiencing periodic activity: FGF, Wnt, and Notch. The wave of cyclic gene expression controlled by the segmentation clock oscillator is shown in orange on the left side of the embryos. When competent cells that pass through the determination front receive the clock signal, they simultaneously activate Mesp2 (mesoderm posterior 2; shown in black), thereby defining the future segmental domain as shown on the right side of the embryos. In this model, the size of the segment (the future somite) is defined by the distance traveled by the wavefront during one oscillation of the segmentation clock. However, the role of retinoic acid in this model remains debated (see text). During the next cycle Mesp2 expression becomes restricted to the anterior compartment of S–I (grey). T, time in segmentation clock cycle unit. [Reprinted from Dequéant M-L, Pourquié O. Segmental patterning of the vertebrate embryonic axis. Nature Rev. Genet 2008; 9(5):370–382. with permission from Springer Nature]
References
Bénazéref B, Pourquié O. Formation and segmentation of the vertebrate body axis. Annu Rev Cell Dev Biol. 2013;29:1–26. https://doi.org/10.1146/annurev-cellbio-101011-155703. Epub 2013 Jun 26. Review PMID: 23808844.
Pang D, Thompson DNP. Embryology and bony malformations of the craniovertebral junction. Childs Nerv Syst. 2011;27:523–64.
Shishkin MA. Evolution of the cervical vertebrae in Temnospondyl amphibian and differentiation of the early tetrapods. Paleontol J. 2000;34(5):534–46.
O’Rahilly R, Müller F. Human embryology and teratology. 3rd ed. New York: Wiley-Liss; 2001.
Menezes AH, Fenoy KA. Remnants of occipital vertebrae proatlas segmentation abnormalities. Neurosurgery. 2009;64(5):945–54.
Spittank H, Goehmann U, Hage H, Sacher R. Persistent proatlas with additional segmentation of the cranio-vertebral junction: the Tsuang-Goehmann malformation. Radiol Case. 2016;10(10):15–23.
Onimaru K, Shoguchi E, Kratani S, Tanaka M. Development and evolution of the lateral plate mesoderm: compariative analysis of amphiosus, and lamprey with implircations for the acquisitions of acquired fins. Dev Biol. 2011;359:124–36.
Matsuoka T, Ahlberg PE, Kessaris N, Iannarelli P, Dennehey U, Richardson WD, McMahon AP, Koentges G. Neural crest origins of the neck and shoulder. Nature. 2005;436(7049):347–55.
Kierner AC, Zelenka AM, Reidel G, Burian M. Blood supply of SCM muscle and its clinical implications. Arch Surg. 1999;134(2):144–7.
Nagashima H, Sugahara F, Watanabe K, Shibata M, Chiba A, Sato N. Developmental origin of the clavicle and its implications for the evolution of the neck and the paired appendages in vertebrate. J Anat. 2016;229:536–48.
Ericsson R, Knight R, Johanson Z. Evolution and development of the vertebrate neck. J Anat. 2013;222:67–78.
Huang R, Quia Z, Patel K, Wilting J, Christ B. Contributions of single somites to the skeleton and muscles of the occipital and cervical regions in avian embryos. Anat Embryol. 2000a;202:375–83.
Huang R, Zhu Q, Patel K, Wilting J, Christ B. Dual origin and segmental organization of the avian scapula. Development. 2000b;127:3789–94.
McGonnell IM. The evolution of the pectoral girdle. J Anat. 2001;199:181–94.
Lachman N, Acland RD, Rosse C. Anatomical evidence for the absence of a morphologically distinct cranial root of the accessory nerve in man. Clin Anat. 2002;15:4–10.
Tada MN, Kuratani S. Evolutionary and developmental understanding of the spinal accessory nerve. Zool Lett. 2015;1:4. https://doi.org/10.1186/s40851-014-0006-8.
Singh PH, Kumar S, Agarwal SP. Congenital asymptomatic absence of unilateral sternocleidomastoid muscle. BMJ Case Rep. 2014;2014:bcr2013202786. https://doi.org/10.1136/bcr-2013-202786.
Vajramani A, Witham FM, Richards RH. Congenital unilateral absence of sternocleidomastoid and trapezius muscles: a case report and literature review. J Pediatr Orthop B. 2010;19(5):462–4. https://doi.org/10.1097/BPB.ObO13e32833ce404.
Hacein-Bey L, et al. The ascending pharyngeal artery: branches, anastomoses, and clinical significance. Am J Neuroradiol. 2002;23:1246–56.
Leclère FM, Vacher C, Benchaa T. Blood supply to the human sternocleidomastoid and its implications for mandible reconstruction. Laryngoscope. 2012;122(11):2402–6.
Emsley JG, Davis MD. Partial unilateral absence of the trapezius muscle in a human cadaver. Clin Anat. 2001;14:383–6.
Allouh M, Mohamed A, Mhanni A. Complete unilateral absence of trapezius muscle. McGill J Med. 2004;8:31–3.
Garbelotti SA Jr, de Sousa Rodrigues CF, Sgrott EA, Prates JC. Unilteral absence of thoracic part of the trapezius muscle. Surg Radiol Anat. 2001;23(2):131–3.
Bergin M, Elliott J, Jull G. The case of the missing lower trapezius muscle. J Orthop Sports Med. 2011;41(8):614–5.
Tigga SR, Goswami P, Khanna J. Congenital partial absence of trapezius with variant pattern of rectus sheath. Acta Med Iran. 2016;54(4):280–2.
Mehra L, Tuli A, Raheja S. Dorsoscapularis triangularis: embryological and phylogenetic characterization of a rare variation of trapezius. Anat Cell Biol. 2016;49:213–6.
Witbreuk MM, Lambert SM, Eastwood DM. Unilateral hypoplasia of the trapezius muscle in a 10-year old boy: a case report. J Pediatr Orthop B. 2007;16:229–32.
Yiyit N, Isitmangil T, Öztúrker C. The abnormalities of trapezius muscle might be a component of Poland’s syndrome. Med Hypotheses. 2014;83:533–6.
Nooij LS, Oostra RJ. Trapezius aplasia: indications for a dual developmental origin of the trapezius muscle. Clin Anat. 2006;19:547–9. https://doi.org/10.1002/ca.20325.
Trinajstic K, Sanchez S, Dupret V, Tafforeau P, Ahlgren PE. Fossil musculature of the most primitive jawed vertebrates. Science. 2013;341(6142):160–4. https://doi.org/10.1126/science.1237275. Epub 2013 Jun 13. PMID: 23765280
Sefton EM, Bhullar B-A, Mohades Z, Hanken J. Evolution of the head trunk interface in tetrapod vertebrates. elife. 2016;5:e09972. https://doi.org/10.7554/eLife.09972.
Barry MA. Central connections of the IXth and Xth cranial nerves in the clearnose skate, Raja eglantera. Brain Res. 1987;425:159–66.
Boord RL, Sperry DG. Topography and nerve supply of the cucullari (trapezius) of skates. J Morphol. 1991;207:165–72.
Marion GE. Mandibular and pharyngeal muscles of Acanthius and Raja. Am Nat. 1905;39:891–924.
Sperry DG, Boord RL. Central location of the motoneurons that supply the cucullaris (trapezius) of the clearnose skate, Raja eglanteria. Brain Res. 1992;582:312–9.
Daeschler EB, Shubin NH, Jenkins FA. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature. 2006;440:757–65.
Ma L-H, Gilland E, Bass AH, Baker RS. Ancestry of motor innervation to pectoral fin and forelimb. Nat Commun. 2010;1:49. https://doi.org/10.1038/ncomms1045.
Murakami Y, Tanaka M. Evolution of motor innervation to vertebrate fins and limbs. Dev Biol. 2011;355:164–72.
Stifani N. Motor neurons and the generation of spinal motor neuron diversity. Front Cell Neurosci. 2014;8:293. https://doi.org/10.3389/fncel.2014.00293.
Pickering M, Jones JFX. The diaphragm: two physiological muscles in one. J Anat. 2002;201:305–12.
Merrell AJ, Kardon G. Development of the diaphragm, a skeletal muscle essential for mammalian respiration. FEBS J. 2013;280(17):4026–35. https://doi.org/10.1111/febs,12274.
Fisher JC, Bodenstein L. Computer simulation analysis of normal and abnormal development of the mammalian diaphragm. Theor Biol Med Model. 2006;3:9. https://doi.org/10.1186/1742-4682-3-9.
Toran N, Eery J. Congenital bilateral absence of diaphragm. Thorax. 1891;36:157–8.
Clugston RD, Greer JJ. Diaphragm development and congenital diaphragmatic hernia. Semin Pediatr Surg. 2007;16:94–100.
Kulkarni ML, Sneharoopa B, Vani HN, Nawaz S, Kannan B, Kulkarnin PM. Eventration of the diaphragm and associations. Indian J Pediatr. 2007;74:202–5.
Mirza B, Bashir Z, Sheikh A. Congenital right hemidiaphragmatic agesis. Lung India. 2012;29(1):53–5.
Ravisagar P, Abhinav S, Mathur RM, Anua S. Eventration of diaphragm presenting a recurrent respiratory tract infections—a case report. Egypt J Chest Dis Tuberc. 2015;64:291–3.
Hirasawa T, Kuratanin S. A new scenario of the evolutionary derivation of the diaphragm from shoulder muscle. J Anat. 2013;222:504–17.
Hirasawa T, Fujimoto S, Kuratani S. Expansion of the neck reconstituted the shoulder-diaphragm in amniote evolution. Dev Growth Evol. 2016;58:143–53.
Kakodkar KA, Shroeder JW Jr, Holinger LD. Laryngeal development and anatomy. In: Hartnik CJ, Hansen MC, Gallaghen TQ, editors. Advances in oto-rhino-laryngology, pediatric airway surgery, vol. 73. Basel: S Karger, AG; 2012. p. 1–13.
Brusca RC, Brusca GJ. Invertebrates. Sunderland: Sinauer; 1990.
Franch-Marro X, Martin M, Averof M, Casanova J. Association of trachal placodes with leg primordia in Drosophila: implication for the origin of the insect tracheal system. Development. 2006;133:785–90.
Ahlberg PE, Clack JA. A firm step from water to land. Nature. 2006;440:747–9.
McIver MA, Schmitz L, Mugan U, Murphey TD, Mobley CD. Massive increase in visual range preceded the origin of terrestrial vertebrates. Proc Natl Acad Sci USA. 2017;114(12):E2375–84. https://doi.org/10.1073/pnas.1615563114. Epub 2017 Mar 7. www.pnas.org/cgi/doi/10.1073/pnas.1615563114.
Ahlberg PE, Clack JA, Luksevics E. Rapid braincase evolution between Panderichthys and the earliest tetrapods. Nature. 1996;381:61–4.
Schultze M, Arsenault M. The Panderichthid fish Elpigostega—a close relative of Tetrapods? Paléo. 1985;28(2):293–309.
McIver MA. Neurobiology: from morphological computation to planning. In: Robbins P, Aydede M, editors. The Cambridge handboook of situated cognition. New York: Cambridge Univ. Press; 2009. p. 480–504.
Shubin NH, Daeschler EB, Jenkins FA. The pectoral fin of Tiktaalik roseaa and the origin of the tetrapod limb. Nature. 2006;440:764–71.
ten Broek CMA, Bugiani M, Gallis E. Evo-devo of the human vertebral column: on homeotic transformation, pathologies, and prenatal selection. Evol Biol. 2012;39:456–71.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this chapter
Carstens, M.H. (2023). The Neck: Development and Evolution. In: Carstens, M.H. (eds) The Embryologic Basis of Craniofacial Structure. Springer, Cham. https://doi.org/10.1007/978-3-031-15636-6_10
Download citation
DOI: https://doi.org/10.1007/978-3-031-15636-6_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-15635-9
Online ISBN: 978-3-031-15636-6
eBook Packages: MedicineMedicine (R0)