
Abstract
North Patagonian Andean deep lakes (>100 m depth) are ultraoligotrophic, and thus are high light-low nutrient environments. In this chapter, we present examples and discuss the effect of particle input due to climate change (changes in glacial clay due to glacial recession) and volcanic eruption (volcanic ashes) in the water column of these lakes. The best descriptor of water column functioning was the vertical position of the deep chlorophyll maximum that was very sensitive to changes in underwater light and may be used as a simple variable to track changes in the dynamics of lakes. Mixotrophy is particularly important in the plankton of these lakes, and here we show how glacier recession affects, through the glacier clay, the bacterivory of these protists. Finally, we present results on how volcanic ashes triggered the disappearance of cladoceran populations, and changes in glacier clay affect zooplankton filter feeders, modifying the food quality (C:P ratio) that drives the coexistence between cladocerans and copepods.
‘Qué tranquilo y bello el cuadro en las cercanías del Leman argentino ¡más grandioso que el Suizo!.’ (How calm and beautiful scene in the surroundings of the Argentinian Leman, more magnificent than the Swiss one!)
F.P. Moreno – January 22, 1876.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
1 Introduction
The largest lake basin in Argentina is in the Andean-Patagonian region; these lakes are the heart and essence of the landscape of the region. Considering their volume, these deep and large lakes (area >5 km2, depth ≥100 m) are important water reserves that represent 77% of the water contained in lakes of Argentina (Quirós 1988).
The first data on these Patagonian lakes were obtained by geographers in the last decades of the nineteenth century, who referred to the similarity with lakes in the Alps (Biedma 1987). In the early twentieth century, salmonids were successfully introduced in most of the deep lakes (Quirós and Drago 1999). However, it was until the 1950s when biological samples were studied for the first time by the Swedish expedition of Kuno Thomasson (Thomasson 1959; Thomasson 1963). Thus, there are no scientific samples from before this introduction occurred.
The area has been affected by glacial processes that modeled the landscape including the lakes. In addition, the region has been affected by the activity of several volcanos producing volcanic ashes. These events (including glacial recession and catastrophic events such as moraine breaks and volcanic eruptions) cause changes in the input of inorganic suspended particles into lake ecosystems.
2 The Deep Andean-Patagonian Lakes of Argentina
Deep Andean-Patagonian lakes of Argentina (maximum depth >100 m) correspond to the Glacial lake district of the Southern Andes (Iriondo 1989) and the Wet Andes according to glacio-climatological regions (Lliboutry et al. 1998). This lake district in Argentina extends from 37° S (Lake Aluminé) to 55° S (Lake Fagnano), covering almost 2000 km (Fig. 3.1). During the late Pleistocene, the area was glaciated extensively and repeatedly. Thus, the landscape is dominated by glacial processes (moraines and glacial-fluvial plains) and also by volcanic events (Pereyra and Bouza 2019). Climate is cold temperate, with the prevalence of west-winds (westerlies) coming from the Pacific Ocean (Paruelo et al. 1998; Masiokas et al. 2008). Elevation of most mountain peaks and massifs does not exceed 4,000 m a.s.l. and the intense influence of the westerly circulation from the Pacific results in high precipitations (Kitzberger and Veblen 2003). Thus, the region is characterized by strong precipitation gradients, with clear contrasts between the western (~3000 mm y−1) (Viale et al. 2019) and the eastern slopes (2000–1500 mm y−1) (Paruelo et al. 1998; Viale et al. 2018). However, diverse macro-climatic phenomena such as SAM (South Annular Module), El Niño Southern Oscillation (ENSO), and the Pacific Decadal Oscillation (PDO) have affected interannual and month-to-month variations in the intensity of rains (Trauth et al. 2000; Rivera et al. 2018; Viale et al. 2019; Poveda et al. 2020). The region contains a wide variety of glaciers including permanent snowfields, mountain glaciers, valley glaciers, outlet glaciers, piedmont glaciers, icecaps, and extensive icefields (Masiokas et al. 2020; Chap. 4). The presence of a profuse hydrographic system including large deep lakes and rivers is characteristic of the Wet Andes, wherein 4800 km2 of the former extent of large glaciers are now occupied by more than 4,000 lakes (Wilson et al. 2018). However, the high precipitation amounts cause that most river flow is dependent on rainfall and snowmelt patterns (Masiokas et al. 2019). The rivers fed from Andean waters cross the Andes, flowing toward the Pacific Ocean or run through the Patagonian plateau steppe and outflow to the Atlantic Ocean (Chap. 9).

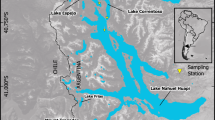
Maps of the Andean Patagonian lake area. (a) Location of the Andean lakes. (b) Andean lakes and the Liquiñe-Ofqui fault in the west side of the Andes. (c–e) Details of the most studied lakes. In (d), the sampling stations (numbered) are indicated along the Tronador arm of Lake Mascardi
The vegetation of the area is mainly composed of evergreen and deciduous trees dominated by species of the genus Nothofagus. In particular, the deciduous species of the southern beech, locally named “lenga,” N. pumilio (Poepp. et al.) Krasser, occurs from 35° S down to 55° S (Hildebrand-Vogel et al. 1990). Within the broad distribution area of more than 2000 km in length, the altitudinal distribution varies, constituting high mountain forests up to the timberline (Krummholz: stunted wind-blown trees) to pure stands at the sea level in the Southern Tierra del Fuego (Mathiasen and Premoli 2010). N. pumilio constitutes an important source of organic matter for both lakes and rivers (Albariño and Balseiro 2001; Bastidas Navarro et al. 2014; Bastidas Navarro et al. 2019).
The lakes exhibit a warm, monomictic thermal regime, with stable thermal stratification during late spring and summer (Baigún and Marinone 1995). Thermocline depth ranges between 15 and 40 m (or more), and this is related to lake morphometry, fetch, and interannual wind variations (Baigún and Marinone 1995; Pérez et al. 2007; Modenutti et al. 2008). Transparency is extremely high with very low vertical extinction coefficients of the different wavelengths (Kd PAR = 0.10–0.16 m−1) (Morris et al. 1995). However, some lakes are under the influence of clay discharged by glaciers such as Lake Argentino, Viedma, and Mascardi, and in these lakes, transparency decreases sharply (Modenutti et al. 2000; Richter et al. 2016). Earlier baseline studies indicated that nutrient concentrations are very low corresponding to the oligotrophic and undisturbed lake status (Calcagno et al. 1995; Markert et al. 1997; Quirós and Drago 1999).
Extensive studies of the whole lake district are scarce and most of them were carried out in a single summer campaign (Quirós 1988; Drago and Quiros 1995; Quirós 1997; Quirós and Drago 1999). However, the North Patagonian Andean lakes (around 39° to 42°S and 71°W, 400–750 m a.s.l.) were more intensively studied (Modenutti and Balseiro 2018). In this chapter, we will focus on this lake district located at mid-latitudes (Fig. 3.1). We aimed to summarize the events that occurred in the water column of deep oligotrophic lakes as a consequence of glacial recession due to climate change and natural catastrophic events (volcanic eruptions and sudden moraine breaks). These events produced noticeable changes in the light distribution of the water column because of the increase of suspended solids (volcanic ashes or glacial clay), affecting the plankton communities’ interactions, and thus pelagic ecosystem functioning.
3 Climate Change
In North Patagonian Andes, contemporary changes in the atmosphere have caused precipitation and mean temperatures to change at values previously experienced in geologic time; however, the velocity of change appears to be faster than that of similar periods (Marcott et al. 2013; Neukom et al. 2019). In particular, the increase in temperature (up to 1 °C since 1950) has been higher than in the rest of Argentina (Villalba et al. 2003; Barros et al. 2014), while precipitation significantly decreased (around 5% per decade) (Castañeda and González 2008; Masiokas et al. 2008). Thus, climate models predict around 10–30% less precipitation over northern Patagonia by the end of the century (Marengo et al. 2011; Barros et al. 2014; Pessacg et al. 2020), while in terms of temperature, simulations project an increase from 1.5 to 3 °C for the far future scenario (2071–2100) (Pessacg et al. 2020).
Glaciers are sensitive to climate change (Roe et al. 2017) and in the North Patagonian Andes (40.5°–44.5° S) glaciers decreased ~25% of their area between 1985 and 2011 (Paul and Mölg 2014), and this negative balance was confirmed by remote sensing data (Dussaillant et al. 2019; Chap. 4). As a consequence, the number of proglacial lakes and lake volume increased in response to climate change and glacier retreat (Paul and Mölg 2014; Shugar et al. 2020). In particular, the upper slopes of Monte Tronador host one of the largest contiguous ice covers (∼57 km2 in 2012) in the northern Patagonian Andes (Ruiz et al. 2015). Glaciers in Monte Tronador show a clear retreating and thinning pattern with a long-term frontal recession (Bown and Rivera 2007; Masiokas et al. 2010; Paul and Mölg 2014; Ruiz et al. 2017). As glacial lakes drain, hydrologic and geomorphic changes can occur (Kershaw et al. 2005; Harrison et al. 2018; Shugar et al. 2020) including catastrophic and hazardous events as glacial lake outburst floods (GLOFs) due to the sudden release of a glacier or moraine-dammed lakes (Masiokas et al. 2020; see Chap. 4). These GLOF events produce strong impacts in downstream valleys and the runoff of the outlet rivers (Dussaillant et al. 2012). In Monte Tronador in 2009, a GLOF event occurred due to heavy precipitation and ice blockage of the outlet, causing a break of the end moraine that impounded the proglacial Lake Ventisquero Negro (Worni et al. 2012). The event modified the downstream valley, and then the volume of the proglacial Lake Ventisquero Negro increased (Ruiz et al. 2017; Modenutti et al. 2018a).
Bedrock abrasion by glaciers from Monte Tronador produce fine rock and mineral fragments which constitute the glacial flour (glacial clay) that is carried by meltwaters through streams (Chillrud et al. 1994) to proglacial lakes (Ariztegui et al. 2007). Glacial clay from the Argentinian side of the Monte Tronador is carried into three main proglacial lakes: Ventisquero Negro, Frias, and Mascardi affecting the light vertical distribution (Bonetto et al. 1971; Morris et al. 1995; Modenutti et al. 2000). Variations in the transport of glacial clay to the basin can be directly linked to changes in climate (Ariztegui et al. 2007). The marked retreating of Ventisquero Negro glacier (Ruiz et al. 2015) and the 2009 GLOF event (Worni et al. 2012) caused a concomitant change in sediment carried downstream by Upper Río Manso that produced substantial variation in the light distribution in Lake Mascardi Tronador Arm (Bastidas Navarro et al. 2018).
4 Volcanic Eruptions
In the southern Andes, from 40 to 46 °S, there is an extended fault zone called Liquiñe-Ofqui, which extends for about 1000 km NS with more than 40 active volcanos (Cembrano et al. 1996). This fault coincides with the Quaternary volcanic chain that contains recent active stratovolcanos as Chaiten and Puyehue-Cordón Caulle. In May 2008, the Chaiten erupted about 1 km3 of ashes (Carn et al. 2009), and in June 2011, the Puyehue-Cordón Caulle had a mega eruption with a spread of more than 100 million tons of pyroclastic material mostly carried by wind to the east side of the Andes, affecting a wide area of Argentina (Elissondo et al. 2016) including deep lakes (Elser et al. 2015). The eruption carried pumice of several sizes (4 mm to >10 cm in diameter) to the lake surface. Due to the low density of the sponge-like rocks, this pumice remained floating on the water surface from weeks to many months, creating new conditions for aquatic life (Elser et al. 2015; Modenutti et al. 2016). In addition to the pumice, ashes from the volcano affected lakes, as they drastically changed their optical climate, turning highly transparent waters to grey, cloudy ones (Modenutti et al. 2013b). At least six historical eruptions of the Puyehue-Cordón Caulle have been recorded (1759, 1893, 1921, 1960, 1990, 2011) (Elissondo et al. 2016). Thus, lakes in the area receive this volcanic input periodically.
5 Lake Water Column: Light, Temperature, and Nutrients
Light and nutrient supply dynamics interacting with thermal water column structure and internal food web interactions are decisive for pelagic ecosystem functioning (Sterner et al. 1997; Falkowski and Raven 2007). Geologically, the region of North Patagonian Andean deep lakes is dominated by a mixture of crystalline igneous, volcanic, and plutonic rocks, while sedimentary rocks are proportionally scarce (Flint and Fidalgo 1964). Thus, waters in lakes and rivers in the region are extremely dilute solutions in which major ion concentrations are below world averages (Pedrozo et al. 1993). Nutrient concentrations are also very low (total phosphorus <6 μg L−1 and total nitrogen <100 μg L−1) and no significant differences throughout the water column were noticed (Corno et al. 2009). Within the lake water column, light may have a complex pattern of spatial and temporal variability (Litchman 2003; Stomp et al. 2007a, b), and fluctuations in irradiance may affect photosynthesis and respiration (Quéguiner and Legendre 1986; Ferris and Christian 1991; Falkowski and Raven 2007), as well as primary producers’ growth rates (Litchman 2000, 2003). Light is a distinctive feature in deep North Patagonian Andean lakes since early studies have reported their high transparency to different wavelengths including ultraviolet bands, due to the low dissolved organic carbon (DOC) (Morris et al. 1995). The concentration of DOC shows values below 0.6 mg L−1 (Corno et al. 2009) and remains without changes along precipitation gradients (Zagarese et al. 2017; Queimaliños et al. 2019). Considering light and nutrients, these deep lakes can be described as high-light:low-nutrient environments (Balseiro et al. 2007), and this condition drives most processes that occur in the water column.
Comparing the different wavelengths, in most of the deep transparent lakes, the photosynthetically active radiation (PAR, 400–700 nm) can reach up to 40–50 m depth (euphotic zone) while hazardous UV-B (305 nm) and UV-A (320 nm) affect up to 12–20 m depth, respectively (Fig. 3.2a). However, this condition changes when lakes receive the input of glacial clay producing a reduction of the euphotic zone (e.g., in Fig. 3.2b see lakes Frías and Mascardi 3).

(a) Light vertical profiles in a North Patagonian Andean lake (Lake Nahuel Huapi), showing the transparency to several wavelengths, including ultraviolet radiation (UV-B, 305 nm and UV-A, 320, 340, and 380 nm) and photosynthetically active radiation (PAR, 400–700 nm). (b) Light vertical profiles of PAR of several lakes with different transparency. Less transparent lakes (Mascardi and Frías) are lakes with the input of inorganic particles (glacier clay). Mascardi 3 refers to Lake Mascardi sampling station number 3 (E3)
Lake thermal structure is also related to the light availability in the different layers of the water column. North Patagonian Andean lakes are warm monomictic, and summer stratification is characterized by a wind-mixed surface layer that is isolated from colder deep waters by a marked thermal gradient at the metalimnetic level (Pérez et al. 2002). Interestingly, extended euphotic zones include the epilimnion, the metalimnion, and, in certain cases, the upper part of the hypolimnion (Fig. 3.3). Vertical mixing can lead to a shortage of light if planktonic organisms are frequently dragged down to the deep dark layers, whereas stratification enhances light supply by decreasing mixing depth (Diehl 2002). This condition will imply that variation of the mixing depth affects the available mean light in the upper layer (Fig. 3.3). The available light in the mixing layer is defined as mean intensity in the mixolimnion (Im) (Helbling et al. 1994; Kirk 1994) and changes in Im affected the dominance of different planktonic species (Modenutti et al. 2008).

Light (blue) and temperature (red) vertical profiles of three different Andean Patagonian lakes. (a) and (b) have similar transparency (~Kd), but very different temperature profiles, (b) and (c) have similar temperature profiles, but (c) is less transparent. The yellow-shaded area represents the section averaged for the estimation of the irradiance of the mixed layer (Im). Note how Im changes with the variation in the temperature profile
As mentioned, glacial lakes frequently receive inputs of finely grounded rock particles of glacial origin; thus, upper glacial lakes often have a grey or whitish appearance. In contrast, the lower lakes in a series of glacial lakes can be blue because all particles have settled out of the water column. Early studies in Lake Mascardi (Bonetto et al. 1971) have shown that light was affected by glacial clay input, and Modenutti et al. (2000) showed a significant statistical relationship between the light extinction coefficient of PAR and total suspended solid concentrations. In particular, light extinction coefficients decreased steadily from the clay source (Upper Manso River) and along the Tronador Arm indicating an increase in transparency (Fig. 3.4). The GLOF event in May 2009 in Ventisquero Negro caused a significant decrease in turbidity over the years following the event (Fig. 3.4, black vs. red lines). Analysis of satellite images with SoPI (SRL 2018) revealed an acceleration in the increase of the lake area from 2.5 ha year−1 (before GLOF, 1998–2009) to 3.5 ha year−1 after the event (2010–2016). The increase in lake size increased the glacial clay sinking, and thus decreased the amount of suspended solids that were transported by the Upper Manso River downstream and into lake Mascardi Tronador Arm (Bastidas Navarro et al. 2018) (Fig. 3.5).

Light vertical profiles of photosynthetically active radiation (PAR) along the gradient of the Tronador arm of Lake Mascardi. Black lines correspond to data before the Glacier Lake Outburst Flood (GLOF) occurred in 2009 and red lines to data after the event. Note that the lake becomes more transparent from E1 to E6, before and after the GLOF

Satellite photographs of Lake Mascardi. RGB composites of the surface reflectance (ρs, Level-2) in all available wavelengths based on the atmospheric correction procedure using ACOLITE Python (https://github.com/acolite/acolite). Sensors used: Landsat 5 (L5/TM), Landsat 7 (L7/ETM), and Landsat 8 (L8/OLI). Date format on each map: year-month-day (hh:ss UTC). The sequence shows the change in reflectance (due to glacier clay) A-E: before (years 1986, 1998, 2000, 2005, and 2008), and F to I: after (2014, 2015, 2018, and 2021) the GLOF event
The eruption of the Puyehue-Cordón Caulle in 2011 discharged massive amounts of ash and pumice into the surrounding landscape in Argentina, producing an increase in total suspended solids (1.5 to 8-fold) in the surrounding lakes (Modenutti et al. 2013b). However, no differential specific absorption of the different light wavelengths occurred due to ash input, thus effects of volcanic particle inputs on the water column were comparable to those of glacial clay, decreasing light by increasing scattering, and thus light extinction coefficient. The result was a reduction of the euphotic zone during extended periods after the eruption occurred. Interestingly, glacial clay or volcanic ashes did not affect DOC concentration, however, volcanic ashes increased phosphorus in the water column.
6 Deep Chlorophyll Maximum (DCM) as a Sensitive Variable
The high transparency (both to PAR and UVR, Fig. 3.2a) strongly reduces primary production and phytoplankton growth rates in the upper layers of the water column (Callieri et al. 2007). The net effect on the phytoplankton community is strong photoinhibition with DNA damage (Villafañe et al. 2004). These upper layers are inhabited by pigmented phytoplanktonic species that have high carotenoid content (Pérez et al. 2007). In addition, in many lakes, these upper layers are dominated by mixotrophic dark-pigmented ciliate species such as Stentor araucanus (Modenutti et al. 2005). The success of this species in highly illuminated layers has been associated to the pigment stentorin and the high concentration of mycosporine-like aminoacids (MAAs) that contribute to the high performance of these ciliates under UVR (Modenutti et al. 1998; Tartarotti et al. 2004). In particular, this species profits from high irradiances and is particularly abundant when the epilimnion mixing layer is reduced by calm wind weather. In this case, differences in Im with an increase in mean light in the upper levels favored the prevalence of the dark ciliate (Modenutti et al. 2008).
Because of the avoidance of high irradiances in the epilimnetic mixing layer, the distribution of phototrophic organisms throughout the water column results in the development of a meta-hypolimnetic deep chlorophyll maximum (DCM) located approximately at 1% of surface PAR (Pérez et al. 2002; Modenutti et al. 2004; Modenutti et al. 2013a). The colonization of these deep levels by phototrophic organisms represents a trade-off between higher survival (decrease of UVR effect) and lower cell-specific primary production (low irradiance) (Modenutti et al. 2004). Static primary production experiments, both in situ and in experimental system incubations, showed that bright light is a major factor reducing primary production (Callieri et al. 2007) and that is not compensated by the addition of nutrients (N and P) (Modenutti et al. 2013b). Although in lake experiments with light intensities below 10% of surface PAR (<200 μmol photon m−2 s−1), production increases significantly (Modenutti et al. 2004), this condition is achieved within the mixing layer where cells are dragged to higher damaging irradiances (Diehl 2002). Thus, in deep stratified lakes, primary production at these mixed levels remains low and increases only below the thermal gradient of the metalimnion. In fact, the development of DCM in North Patagonian Andean lakes, as in oceans and other deep lakes, has been associated with the existence of stable dim-illuminated metalimnetic layers (Kirk 1994; Sharples et al. 2001), and the variables that explained the DCM depth were the light extinction coefficients of PAR and 320 nm (Kd 320 and Kd PAR) (Modenutti et al. 2013a).
The low DOC concentration observed in these lakes causes that the underwater wavelength-specific absorption depends mainly on the water itself, and thus the light at the DCM is dominated by green and blue wavelengths of the visible light spectrum (Pérez et al. 2002). For phototrophic organisms, this condition corresponds to the blue spectral niche where phycoerythrin and chlorophyll b are the accessory pigments better adapted (Stomp et al. 2007a, b; Holtrop et al. 2021). Indeed, at this level, the contribution of photosynthetic accessory pigments, such as fucoxanthin, phycoerythrin, and chlorophyll b increased (Pérez et al. 2007), and correspond to the observed increase in dinoflagellates, picocyanobacteria, and the Chlorella-bearing mixotrophic ciliate Ophrydium naumanni (Modenutti and Balseiro 2002; Modenutti et al. 2004; Callieri et al. 2007; Bastidas Navarro et al. 2018). Thus, at the DCM, different competition and predator-prey relationships co-occur. At these levels, autotrophs (both prokaryotes and eukaryotes) compete for light and nutrients, but at the same time, mixotrophs (i.e., nanoflagellates and ciliates) prey actively upon picocyanobacteria, other Eubacteria and Archaea (Modenutti et al. 2008). Furthermore, light is a decisive factor for clearance rate in ciliates and nanoflagellates. Thus, the vertical distribution of both predator and prey (picocyanobacteria) show a high coincidence (Modenutti and Balseiro 2002; Schenone et al. 2020).
The depth of the DCM appeared to be very sensitive to changes in light distribution. As mentioned for Lake Mascardi, the input of glacial clay produces light extinction coefficients to steadily decrease from the clay source (Upper Manso River) and along the Tronador Arm (Fig. 3.4). This light gradient causes, in turn, that DCM becomes deeper along the Tronador Arm (Hylander et al. 2011). As a consequence of the GLOF event in 2009 (Worni et al. 2012), the volume of the Lake Ventisquero Negro increased and the amount of suspended solids that were transported downstream into Lake Mascardi decreased. This condition resulted in a decrease in the light extinction coefficient (Fig. 3.4), and this higher transparency provoked the deepening of the DCM in the years after GLOF (Fig. 3.7) without changes in autotrophic species composition (Bastidas Navarro et al. 2018).
On the other hand, geological events such as volcanic eruptions can also increase the amount of suspended solids and decrease water transparency. During the Puyehue-Cordón Caulle eruption, the turbidity caused by ash fall triggered an upper position of DCM in the affected lakes. In Lake Espejo, the DCM moved upward from 40 m to 15 m depth (Fig. 3.6). However, ashes mainly composed of silica carried other elements as P, Fe, etc. (Caneiro et al. 2011) that changed the phytoplankton composition favoring diatoms such as Tabellaria flocculosa and Aulacoseira granulata.

(a) Lake Espejo before and (b) 5 months after the eruption of Puyehue-Cordón Caulle volcanic complex. Note the change in the color of the lake water and the floating pumice near the lakeshore in (b). Lower panel: vertical light and chlorophyll a profiles: (c) before eruption and (d) 5 months after the eruption of Puyehue-Cordón Caulle volcanic complex

Changes in the depth of the deep chlorophyll maximum (DCM) along the transparency gradient of Lake Mascardi: black dots and line: before the Glacier Lake Outburst Flood (GLOF) event; red dots and lines: after GLOF event. Note the deepening of the DCM after the GLOF with the increase in transparency (see Fig. 3.4)
Summarizing, both suspended particles, glacial clay, and volcanic ashes have shown a similar effect decreasing water transparency, and consequently, provoking an upper location of the DCM. Again, the analysis of environments with glacial clay inputs (before and after GLOF event) and volcanic ash influence indicated that PAR and UV-A (320 nm) wavelengths have major importance in determining DCM depth (Modenutti et al. 2013a). These two catastrophic events provided unique opportunities to directly test the effect of changes in inorganic particle input and to relate different variables with previous lake conditions. These two events can be seen as natural experiments in which DCM was the most sensitive biological variable that quickly and accurately reacted to changes in light extinction.
7 Mixotrophic Nanoflagellates: Light and Feeding
Mixotrophy is a widespread strategy among phytoplankton in highly transparent environments, wherein mixotrophic nanoflagellates (MNF) account for up to 80–90% of total phytoplanktonic cells. MNF have a combination of different feeding strategies: while phagotrophy primarily provides nutrients and carbon for biosynthesis, photosynthesis provides carbon for both biosynthesis and respiration (Jones 2000; Berge et al. 2017; Hansson et al. 2019). In transparent lakes with high light and low nutrient concentration, small picoplanktonic cells (i.e., heterotrophic bacteria and picocyanobacteria) are favored since they are better competitors for P than large osmotrophic phytoplankton because of their higher surface/volume ratio (Gurung et al. 1999; Danger et al. 2007a, b). In this scenario, MNF have an advantage over strict osmotrophic algae by feeding on these P-rich picoplanktonic cells (i.e., bacterivory). Thus, MNF can obtain P by preying on competitors. MNF bacterivory is now recognized as a key pathway of energy and matter transfer through the planktonic food web (Mitra et al. 2014). Recent research has pointed out that mixotrophy is particularly successful in oligotrophic environments with high light-low nutrient conditions, whereas more productive, less illuminated systems would favor strict heterotrophic and phototrophic organisms (Fischer et al. 2017; Waibel et al. 2019). Phagotrophy by heterotrophic nanoflagellates (HNF) is negatively affected by suspended solids (Sommaruga and Kandolf 2014) while that of MNF has a more complex response because of their dependence on light (Schenone et al. 2020).
North Patagonian Andean deep lakes are very transparent, displaying different levels of turbidity due to glacial influence and volcanic activity, and both events produce an increase in suspended solids by the input of fine minerogenic solid particles affecting bacterivory. On the one hand, suspended solids reduce light penetration for phototrophic organisms for photosynthesis (Kirk 1994). Light has a non-monotonic effect on phytoplankton’s phototrophy, where too low or too high light levels will reduce carbon uptake (Jassby and Platt 1976; Litchman 2003). On the other hand, bacterivory studies have suggested a simple and monotonic light dependence on phagotrophy of MNF (Jones 1997; Flynn and Mitra 2009), however, these studies evaluated a narrow range of light intensities. Interestingly, under a wider light range, Schenone et al. (2020) also found a non-monotonic response of phagotrophy to light in MNF. Based on this evidence and combining bacterivory experiments in a gradient of suspended clay with modelling, it was observed that MNF bacterivory is affected by turbidity and particle interference (Fig. 3.8) (Schenone et al. 2020).

Results of the modeling of mixotrophic nanoflagellate bacterivory under a light and glacial clay gradient. The different curves represent the decay in bacterivory as glacier clay increases. For details in the model equations, see Schenone et al. (2020)
In addition to the effect on light penetration, volcanic particles release P to the environment. This would imply less competition for P among osmotrophic cells and trigger the dominance of autotrophy over mixotrophy (Fischer et al. 2017). After the 2011 Puyehue-Cordón Caulle volcanic eruption, community-driven changes were observed in the dominance of MNF species in lakes with high suspended solids due to volcanic particles, from highly bacterivorous species (Chrysochromulina parva) (Gerea et al. 2019) to more phototrophic ones (Plagioselmis lacustris) (Modenutti et al. 2013b).
8 Changes in C:P and Crustacean Zooplankton Distribution
The crustacean zooplankton communities of the North Patagonian Andean lakes are dominated by calanoid copepods of the Family Centropagidae, in particular by Boeckella gracilipes and B. michaelseni, and among cladocerans, Bosmina chilensis and B. longirostris and daphnids, like Ceriodaphnia dubia and Daphnia commutata (Menu Marque and Marinone 1986; Modenutti et al. 2003; Balseiro et al. 2007; Balseiro et al. 2008). In several lakes at very deep layers, the presence of predaceous copepods, such as the calanoid Parabroteas sarsi and the cyclopoid Mesocyclops araucanus, was observed (Reissig et al. 2004; Modenutti et al. 2018b).
Crustacean zooplankton performance is commonly known to be affected by numerous environmental factors, such as fish predation pressure (Brooks and Dodson 1965), food particle size (Sommer 1989), and food quantity (Lampert 1977; Smith and Cooper 1982). Light penetration has a critical role in many features of oligotrophic aquatic ecosystems including zooplankton vertical distribution. In this sense, zooplankton evaded the higher level of the water column due to UVR and PAR+UVR (Alonso et al. 2004; Modenutti et al. 2018b). In addition, light can shape the composition of zooplanktonic communities through food quality. As stated by the “light:nutrient hypothesis” (LNH), the elemental carbon:phosphorus ratio (C:P) of primary producers is a result of the relation between light penetration in the water column and inorganic P availability (Sterner et al. 1997). Under high light intensities (PAR) and low levels of inorganic P, the nutrient limitation for phytoplankton becomes more severe, resulting in a disproportionate accumulation of C relative to P in algal tissue. Food quality has a strong influence on the fitness of herbivorous zooplankton (Sterner and Elser 2002). Hence, the variable C:P ratio in the phytoplankton, as a result of varying light intensities in the water column, represents a nutritional challenge for zooplankton. In turn, the elemental composition of zooplankton varies significantly among grazer species and taxonomic groups. For example, copepods have and require relatively low amounts of P, while cladocerans, in particular Daphnia, have higher P content and requirements (Andersen and Hessen 1991; Hall et al. 2004). As a consequence, the impact of different light penetration (Fig. 3.2b) in P limited systems varies for different zooplankton species according to their somatic P requirements, via the indirect effect of light as a modulator of food quality.
In glacier-influenced lakes inputs of finely ground rock particles cause a turbidity-induced attenuation of photosynthetically active radiation (PAR) and, as stated by the LNH, affect the nutrient composition of phytoplankton. Such shifts in the effects of glacial clay on the light environment can be observed among lakes (i.e., Lake Frías compared with other lakes, Fig. 3.2b), and within a single lake as glacial clay settles, moving away from the source input (i.e., Lake Mascardi). The northernmost end of the Lake Mascardi receives the Upper Manso River with a high load of glacial clay, generating a longitudinal gradient in light (Fig. 3.9). As predicted by the LNH, as light penetration increases with increasing distance from the clay input, also sestonic C:P increases, with a consequent decrease in food quality for grazers (Laspoumaderes et al. 2013; Laspoumaderes et al. 2017). Thus, there is a switch from the dominance of P-rich Daphnia in the turbid, low C:P area of the lake to the dominance of P-poor copepods as the distance from the clay input increases, turbidity decreases and sestonic C:P is higher (Fig. 3.9).

Schematic representation of the light gradient of the Tronador Arm of Lake Mascardi and zooplankton distribution. Note the change in the C:P ratio of phytoplankton and the concomitant change in the relative abundances of cladocerans and copepods along the gradient. Relative size of the C and P represent changes in C:P ratio
In addition to the better food quality in the turbid area of the lake, also ultraviolet radiation (UVR) is attenuated. Stoichiometric constraints (high food C:P ratio) affect Daphnia’s capacity to respond to the oxidative stress caused by UVR (Balseiro et al. 2008). This means that in the more transparent end of the gradient with higher P limitation, the fitness of Daphnia is reduced by its inability to cope with the effect of UVR. Although the copepods could suffer a similar effect of food quality on cellular defense (Souza et al. 2010), due to their lower P requirements, they experience a less severe stoichiometric imbalance than Daphnia.
The ingestion of particles, such as suspended clay, in concentrations over 50 mg L−1, is known to reduce fecundity and survival in daphnids (Kirk and Gilbert 1990), but the highest total suspended solids reported in Lake Mascardi were 5 mg L−1. In this lake, Daphnia coexists with clay particles that have a net positive effect on their fitness (UVR protection, lower C:P food) over the negative effect that can pose a feeding interference. In an experimental clay gradient from 0 to 5 mg L−1, Laspoumaderes et al. (2017) found that glacial clay alone is indeed a source of stress for Daphnia as it affects its enzymatic activities, which represents a feeding interference, and affects respiration rates. However, in the lake, this negative effect is overcome as clay decreases underwater light intensity (both PAR and UVR), which results in a better food quality and a decrease in UVR damage and visual fish predation.
Turbidity affects the foraging of visual planktivorous fishes (Vinyard and O’Brien 1976; Gregory and Northcote 1993; Utne 1997). In particular, members of the Galaxiidae family (Galaxias maculatus and Aplochiton zebra) are important zooplankton feeders in North Patagonian Andean deep lakes (Barriga et al. 2002; Lattuca et al. 2007). While G. maculatus is not affected by turbidity (Stuart-Smith et al. 2007), A. zebra is strongly affected (Jönsson et al. 2011). In addition, eye-size growth trajectories in A. zebra differ between turbid and clear water environments (McDowall and Pankhurst 2005; Lattuca et al. 2007). In Lake Mascardi, A. zebra foraging success decreases as an effect of impaired vision and increasing difficulty in finding prey when the water gets more turbid (Jönsson et al. 2011). Thus, changes in suspended solid may also affect other trophic links between zooplankton and fishes.
9 Volcanic Eruption and Cladoceran Disappearance
The eruption of the Puyehue-Cordón Caulle volcanic complex in 2011 discharged massive amounts of ash and pumice into the surrounding landscape in Argentina, producing an increase in total suspended solids in the surrounding lakes. Particle inputs from either glacial clay or volcanic ashes produce comparable effects on the water column, by generating a decrease in the underwater light penetration and in the depth of the DCM, and a consequent decrease in the C:P ratio of primary producers. Remarkably, the two natural phenomena that seem similar have had contrasting effects on the zooplankton communities.
The deposition of volcanic ash in the lakes affected by the eruption caused a decrease in light and a decrease in sestonic C:P, the same as observed with glacial clay in Lake Mascardi. Before the volcanic eruption, zooplankton communities in lakes Espejo, Correntoso, and Nahuel Huapi (affected by the eruption) were dominated by the copepod Boeckella gracilipes and Ceriodaphnia dubia as the dominant cladoceran, followed by Bosmina chilensis (Balseiro et al. 2007). However, in the summer following the eruption, Wolinski et al. (2013) observed an opposite shift in the zooplankton ensemble to the one observed in Lake Mascardi. Although ash inputs generated a decrease in light penetration and a consequent decrease in the phytoplankton C:P ratio (such as glacial clay in Lake Mascardi), cladocerans disappeared and copepod populations decreased, but the latter only in the lake with the highest ash concentration.
Volcanic particles are very similar in size to glacial clay (Modenutti et al. 2013a), but they have a crystalline structure, mainly composed of silica, which grants abrasive features (Caneiro et al. 2011). Therefore, filter feeders ingest ash as they do with clay, but the damaging effect of ash is much stronger and occurs at lower concentrations than that of clay. Ash was observed to be captured by D. commutata during the feeding process, resulting in a gut completely filled with ashes, a strong decrease in food uptake, and physical gut damage (Fig. 3.10) (Wolinski et al. 2013).

Microphotographs of Daphnia commutata exposed to volcanic ashes in concentration from 0 to 8 mg L−1. (a) Without volcanic ashes, note the green color of the gut due to algae. (b, c) D. commutata exposed to increasing concentration of volcanic ashes ((b) 2 mg L−1 and (c) 5 mg L−1). Note the change in color of the gut in (b) that is filled with ashes and the intensive damage in (c) (dead Daphnia)
Regardless of the better food condition generated by the lower light penetration and the inorganic P provided by the ashes, cladocerans could not deal with the ingestion of volcanic ashes. However, copepods seemed to suffer less due to their ability to select motile cells (Balseiro et al. 2001). While 5 mg L−1 of glacial clay is still a beneficial concentration promoting population growth in Daphnia (as reported for Lake Mascardi), ash concentration between 2 and 8 mg L−1 was sufficiently harmful to affect the survival and reproduction of natural and experimental populations of cladocerans. Indeed, the experiments with 8 mg L−1 of ashes caused the complete disappearance of Daphnia populations within 5 days (Wolinski et al. 2013). As the sedimentation process occurred, ash concentrations decreased, favoring population recovery, as observed in the following summer (18 months after the eruption). When the lakes recovered their original transparency with low total suspended solids values, the cladoceran populations also returned to their historical abundances.
10 Concluding Remarks
Long-term studies monitoring North Patagonian Andean deep lakes allowed us to analyze how different events affect internal lake processes and eventually how resilient these lakes are to the input of inorganic particles and the consequent light changes. The most sensitive biological variable to these external factors is the location of the DCM in the water column. This is of particular interest as chlorophyll is a very easy-to-measure variable, both in the laboratory or directly in the field by a portable fluorometer. Thus, this variable represents an important tool for lake monitoring since the depth of the DCM reacted quickly and reliably to changes in light extinction. However, important changes can occur considering the composition of the DCM because species react differently to particle interference (as shown for nanoflagellates) (Schenone et al. 2020) and to the addition of Si and P (as shown by the diatom increase after the volcanic eruption) (Modenutti et al. 2013b). Future scenarios of climate change include differences in optical light climate both decreasing and increasing the diffuse extinction coefficient of lake water. Also, possible changes in land use with the increment of new roads and touristic center developments will affect the input of particles into the lakes with a possible loss of transparency. This will change the location of the DCM, and thus this parameter can be an invaluable tool for understanding lake dynamics and future production trends. Therefore, we encourage agencies to consider this parameter as a quick response of planktonic communities to changes in light conditions.
The relationship between light and nutrients is also determinant of the relative dominance of zooplankton taxa requiring different amounts of P in grazer assemblages and allows to predict shifts in grazer composition with changes in light and nutrient supplies (Elser et al. 2000; Hall et al. 2004; Laspoumaderes et al. 2013). How grazer assemblages react to changes in the light:nutrient environment does not only depend on their P requirements but also on the relationship between the type of particles that generate the light attenuation and their feeding strategy. Hence, the decrease in the performance of planktivorous fishes will affect the transference of secondary production to higher trophic levels, thus impacting the trophic web structure.
References
Albariño RJ, Balseiro EG (2001) Food quality, larval consumption, and growth of Klapopteryx kuscheli (Plecoptera: Austroperlidae) from a south Andes stream. J Freshw Ecol 16:517–526
Alonso C, Rocco V, Barriga JP, Battini MA, Zagarese H (2004) Surface avoidance by freshwater zooplankton: field evidence on the role of ultraviolet radiation. Limnol Oceanogr 49:225–232
Andersen T, Hessen DO (1991) Carbon, nitrogen, and phosphorus content of freshwater zooplankton. Limnol Oceanogr 36:807–814
Ariztegui D, Bösch P, Davaud E (2007) Dominant ENSO frequencies during the Little Ice Age in Northern Patagonia: the varved record of proglacial Lago Frías, Argentina. Quat Int 161:46–55. https://doi.org/10.1016/j.quaint.2006.10.022
Baigún C, Marinone MC (1995) Cold-temperate lakes of South America: do they fit Northern hemisphere models? Arch Hydrobiol 135:23–51
Balseiro E, Souza M, Modenutti B, Reissig M (2008) Living in transparent lakes: Low food P:C ratio decreases antioxidant response to ultraviolet radiation in Daphnia. Limnol Oceanogr 53:2383–2390
Balseiro EG, Modenutti BE, Queimaliños C, Reissig M (2007) Daphnia distribution in Andean Patagonian lakes: effect of low food quality and fish predation. Aquat Ecol 41:599–609
Balseiro EG, Modenutti BE, Queimaliños CP (2001) Feeding of Boeckella gracilipes (Copepoda, Calanoida) on ciliates and phytoflagellates in an ultraoligotrophic Andean lake. J Plankton Res 23:849–857
Barriga JP, Battini MA, Macchi PJ, Milano D, Cussac VE (2002) Spatial and tempòral distribution of landlocked Galaxias maculatus and Galaxias platei (Pisces, Galaxiidae) in a lake in the South Andes. N Z J Mar Freshwat Res 36:345–359
Barros V, Field C, Dokke D, Mastrandrea M, Mach K, Bilir T, Chatterjee M, Ebi K, Estrada Y, Genova R (2014) Climate change 2014: impacts, adaptation, and vulnerability. Part B: regional aspects. Contribution of Working Group II to the fifth assessment report of the Intergovernmental Panel on climate change. Fifth assessment report of the Intergovernmental Panel on climate change. Cambridge University Press, Cambridge
Bastidas Navarro M, Balseiro E, Modenutti B (2014) Bacterial community structure in patagonian Andean Lakes above and below timberline: from community composition to community function. Microb Ecol 68:528–541. https://doi.org/10.1007/s00248-014-0439-9
Bastidas Navarro M, Díaz Villanueva V, Modenutti B (2019) High phosphorus content in leachates of the austral beech Nothofagus pumilio stimulates bacterioplankton C-consumption. Freshw Sci 38:435–447. https://doi.org/10.1086/704752
Bastidas Navarro M, Martyniuk N, Balseiro E, Modenutti B (2018) Effect of glacial lake outburst floods on the light climate in an Andean Patagonian lake: implications for planktonic phototrophs. Hydrobiologia 816:39–48
Berge T, Chakraborty S, Hansen PJ, Andersen KH (2017) Modeling succession of key resource-harvesting traits of mixotrophic plankton. ISME J 11:212–223. https://doi.org/10.1038/ismej.2016.92
Biedma JM (1987) Crónica histórica del lago Nahuel Huapi (Historical chronicle of Nahuel Huapi Lake). Editorial Del Nuevo Extremo
Bonetto AA, Dioni W, Depetris P (1971) Informe preliminar sobre las investigaciones limnológicas de la cuenca del Río Manso y Lago Mascardi (Río Negro - Patagonia) (Preliminary report on the limnological survey of the Río Manso and Lago Mascardi basins). Fundación Bariloche 4:1–62
Bown F, Rivera A (2007) Climate changes and glacier responses during recent decades in the Chilean Lake District. Global Planet Change 59:79–86
Brooks JL, Dodson SL (1965) Predation, body size, and composition of plankton. Science 150:28–35
Calcagno A, Fioriti M, Pedrozo F, Vigliano P, López HL, Rey C, Razquin ME, Quirós R (1995) Catálogo de lagos y embalses de la Argentina (Catalog of the lakes and reservoirs of Argentina). ARM & Asociados
Callieri C, Modenutti BE, Queimaliños C, Bertoni R, Balseiro EG (2007) Production and biomass of picophytoplankton and larger autotrophs in Andean ultraoligotrophic lakes: differences in light harvesting efficiency in deep layers. Aquat Ecol 41:511–523
Caneiro A, Mogni L, Serquis A, Cotaro C, Wilberger D, Ayala C, Daga R, Poire D, Scerbo E (2011) Análisis de cenizas volcánicas del Cordón Caulle (complejo volcánico Puyehue-Cordón Caulle) (Analysis of the volcanic ashes of Cordón Caulle (Puyehue-Cordón Caulle volcanic complex)). Comisión Nacional de Energía Atómica
Carn SA, Pallister JS, Lara L, Ewert JW, Watt S, Prata AJ, Thomas RJ, Villarosa G (2009) The unexpected awakening of Chaitén volcano, Chile. Eos Trans Amer Geophys Union 90:205–206
Castañeda M, González M (2008) Statistical analysis of the precipitation trends in the Patagonia region in southern South America. Atmosfera 21:303–317
Cembrano J, Hervé F, Lavenu A (1996) The Liquiñe Ofqui fault zone: a long-lived intra-arc fault system in southern Chile. Tectonophysics 259:55–66
Chillrud SN, Pedrozo FL, Temporetti PF, Planas FH, Froelich PN (1994) Chemical weathering of phosphate and germanium in glacial meltwater streams: effects of subglacial pyrite oxidation. Limnol Oceanogr 39:1130–1140
Corno G, Modenutti BE, Callieri C, Balseiro EG, Bertoni R, Caravati E (2009) Bacterial diversity and morphology in deep ultraoligotrophic Andean lakes: Role of UVR on vertical distribution. Limnol Oceanogr 54:1098–1112
Danger M, Leflaive J, Oumarou C, Ten-Hage L, Lacroix G (2007a) Control of phytoplankton-bacteria interactions by stoichiometric constraints. Oikos 116:1079–1086
Danger M, Oumarou C, Benest D, Lacroix G (2007b) Bacteria can control stoichiometry and nutrient limitation of Phytoplankton. Funct Ecol 21:202–210
Diehl S (2002) Phytoplankton, light, and nutrients in a gradient of mixing depths: theory. Ecology 83:386–398
Drago E, Quiros R (1995) The hydrochemistry of the inland waters of Argentina: a review. Int J Salt Lake Res 4:315–325
Dussaillant A, Bastianon E, Bertoldi W (2012) Outburst floods and morphology of Colonia and Baker rivers, Patagonia: climate change, extreme flood impacts and sustainable hydropower.
Dussaillant I, Berthier E, Brun F, Masiokas M, Hugonnet R, Favier V, Rabatel A, Pitte P, Ruiz L (2019) Two decades of glacier mass loss along the Andes. Nat Geosci 12(10):802–808. https://doi.org/10.1038/s41561-019-0432-5
Elissondo M, Baumann V, Bonadonna C, Pistolesi M, Cioni R, Bertagnini A, Biass S, Herrero J-C, Gonzalez R (2016) Chronology and impact of the 2011 Cordón Caulle eruption, Chile. Nat Haz Earth Syst Sci 16(3):675–704. https://doi.org/10.5194/nhess-16-675-2016
Elser JJ, Bastidas M, Corman JR, Emick H, Kellom M, Laspoumaderes C, Lee ZM, Poret-Peterson A, Balseiro E, Modenutti B (2015) Community structure and biogeochemical impacts of microbial life on floating pumice. Appl Environ Microbiol 81(5):1542–1549. https://doi.org/10.1128/AEM.03160-14
Elser JJ, Fagan WF, Denno RF, Dobberfuhl DR, Folarin A, Huberty A, Interlandi S, Kilham SS, McCauley E, Schulz KL, Siemann EH, Sterner RW (2000) Nutritional constraints in terrestrial and freshwater food webs. Nature 408(6812):578–580
Falkowski PG, Raven JA (2007) Aquatic photosynthesis, 2nd edn. Blackwell Science, Malden
Ferris JM, Christian R (1991) Aquatic primary production in relation to microalgal responses to changing light. Aquat Sci 53:187–217
Fischer R, Giebel H-A, Hillebrand H, Ptacnik R (2017) Importance of mixotrophic bacterivory can be predicted by light and loss rates. Oikos 126(5):713–722. https://doi.org/10.1111/oik.03539
Flint RF, Fidalgo F (1964) Glacial geology of the East flank of the Argentine Andes between Latitude 39 10′ S. and Latitude 41 20′ S. Geol Soc Am Bull 75:335–352
Flynn KJ, Mitra A (2009) Building the "perfect beast": modeling mixotrophic plankton. J Plankton Res 31(9):965–992. https://doi.org/10.1093/plankt/fbp044
Gerea M, Queimaliños C, Unrein F (2019) Grazing impact and prey selectivity of picoplanktonic cells by mixotrophic flagellates in oligotrophic lakes. Hydrobiologia 831:5–21. https://doi.org/10.1007/s10750-018-3610-3
Gregory RS, Northcote TG (1993) Surface, planktonic, and benthic foraging by juvenile chinook salmon (Onchorhynchus tshawytscha) in turbid laboratory conditions. Can J Fish Aquat Sci 50:233–240
Gurung TB, Urabe J, Nakanishi M (1999) Regulation of the relationship between phytoplankton Scenedesmus acutus and heterotrophic bacteria by the balance of light and nutrients. Aquat Microb Ecol 17:27–35
Hall SR, Leibold MA, Lytle DA, Smith VH (2004) Stoichiometry and planktonic grazer composition over gradients of light, nutrients and predation risk. Ecology 85:2291–2301
Hansson TH, Grossart HP, del Giorgio PA, St-Gelais NF, Beisner BE (2019) Environmental drivers of mixotrophs in boreal lakes. Limnol Oceanogr 64:1688–1705. https://doi.org/10.1002/lno.11144
Harrison S, Kargel JS, Huggel C, Reynolds J, Shugar DH, Betts RA, Emmer A, Glasser N, Haritashya UK, Klimeš J (2018) Climate change and the global pattern of moraine-dammed glacial lake outburst floods. Cryosphere 12:1195–1209
Helbling EW, Villafañe V, Holm-Hansen O (1994) Effects of ultraviolet radiation on the Antarctic marine phytoplankton photosynthesis with particular attention to the influence of mixing. Antarct Res Series 62:207–227
Hildebrand-Vogel R, R G, A V (1990) Subantarctic-Andean Nothofagus pumilio forests. Distribution area and systematic overview, vegetation and soils as demonstrated by an example of a South Chilean stand. Vegetatio 89:55–68
Holtrop T, Huisman J, Stomp M, Biersteker L, Aerts J, Grebert T, Partensky F, Garczarek L, Woerd HJV (2021) Vibrational modes of water predict spectral niches for photosynthesis in lakes and oceans. Nat Ecol Evol 5:55–66. https://doi.org/10.1038/s41559-020-01330-x
Hylander S, Jephson T, Lebret K, Von Einem J, Fagerberg T, Balseiro EG, Modenutti BE, Souza MS, Laspoumaderes C, Jönsson M, Ljungberg P, Nicolle A, Nilsson PA, Ranåker L, Hansson L-A (2011) Climate-induced input of turbid glacial meltwater affects vertical distribution and community composition of phyto- and zooplankton. J Plankton Res 33:1239–1248. https://doi.org/10.1093/plankt/fbr025
Iriondo MH (1989) Quaternary lakes of Argentina. Palaeogeogr, Palaeoclimatol, Palaeoecol 70:81–88
Jassby AD, Platt T (1976) Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol Oceanogr 21:540–547
Jones HLJ (1997) A classification of mixotrophic protists based on their behaviour. Freshw Biol 37:35–43
Jones RI (2000) Mixotrophy in planktonic protists: an overview. Freshw Biol 45:219–226
Jönsson M, Ranåker L, Nicolle A, Ljungberg P, Fagerberg T, Hylander S, Jephson T, Lebret K, von Einem J, Hansson L-A, Nilsson P, Balseiro EG, Modenutti BE (2011) Glacial clay affects foraging performance in a Patagonian fish and cladoceran. Hydrobiologia 663:101–108. https://doi.org/10.1007/s10750-010-0557-4
Kershaw JA, Clague JJ, Evans SG (2005) Geomorphic and sedimentological signature of a two-phase outburst flood from moraine-dammed Queen Bess Lake, British Columbia, Canada. Earth Surface Processes Landforms: J Br Geomorphol Res Group 30(1):1–25
Kirk JTO (1994) Light and photosynthesis in aquatic ecosystems. Cambridge Univ. Press
Kirk KL, Gilbert JJ (1990) Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 71:1741–1755
Kitzberger T, Veblen TT (2003) Influences of climate on fire in northern Patagonia, Argentina. In: Fire and climatic change in temperate ecosystems of the western Americas. Springer, pp 296–321
Lampert W (1977) Studies on the carbon balance of Daphnia pulex de Geer as related to environmental conditions. IV. Determination of the “threshold” concentration as a factor controlling the abundance of zooplankton species. Arch Hydrobiol Suppl 48:361–368
Laspoumaderes C, Modenutti B, Souza MS, Bastidas Navarro M, Cuassolo F, Balseiro E (2013) Glacier melting and stoichiometric implications for lake community structure: zooplankton species distributions across a natural light gradient. Glob Chang Biol 19:316–326. https://doi.org/10.1111/gcb.12040
Laspoumaderes C, Souza MS, Modenutti BE, Balseiro E (2017) Glacier melting and response of Daphnia oxidative stress. J Plankton Res 39(4):675–686. https://doi.org/10.1093/plankt/fbx028
Lattuca M, Ortubay S, Battini M, Barriga J, Cussac V (2007) Presumptive environmental effects on body shape of Aplochiton zebra (Pisces, Galaxiidae) in northern Patagonian lakes. J Appl Ichthyol 23:25–33
Litchman E (2000) Growth rates of phytoplankton under fluctuating light. Freshw Biol 44:223–235
Litchman E (2003) Competition and coexistence of phytoplankton under fluctuating light: experiments with two cyanobacteria. Aquat Microb Ecol 31:241–248
Lliboutry L, Williams R, Ferrigno J (1998) Glaciers of Chile and Argentina. J Geophys Res 1386:1103
Marcott SA, Shakun JD, Clark PU, Mix AC (2013) A reconstruction of regional and global temperature for the past 11300 years. Science 339:1198–1201. https://doi.org/10.1126/science.1228026
Marengo JA, Pabón JD, Díaz A, Rosas G, Ávalos G, Montealegre ER, Villacis M, Solman S, Rojas M (2011) Climate change: evidence and future scenarios for the Andean Region. In: Climate change and biodiversity in the tropical Andes. IAI-SCOPE-UNESCO, Paris, pp 110–127
Markert B, Pedrozo F, Geller W, Friese K, Korhammer S, Baffico G, Diaz M, Wolfl S (1997) A contribution to the study of the heavy-metal and nutritional element status of some lakes in the southern Andes of Patagonia (Argentina). Sci Total Environ 206:1–15
Masiokas MH, Cara L, Villalba R, Pitte P, Luckman B, Toum E, Christie D, Le Quesne C, Mauget S (2019) Streamflow variations across the Andes (18–55 S) during the instrumental era. Sci Rep 9:1–13
Masiokas MH, Rabatel A, Rivera A, Ruiz L, Pitte P, Ceballos JL, Barcaza G, Soruco A, Bown F, Berthier E, Dussaillant I, MacDonell S (2020) A review of the current state and recent changes of the Andean cryosphere. Front Earth Sci 8. https://doi.org/10.3389/feart.2020.00099
Masiokas MH, Villalba R, Luckman BH, Lascano ME, Delgado S, Stepanek P (2008) 20th-century glacier recession and regional hydroclimatic changes in northwestern Patagonia. Global Planet Change 60:85–100
Masiokas MH, Villalba R, Luckman BH, Mauget S (2010) Intra-to multidecadal variations of snowpack and streamflow records in the Andes of Chile and Argentina between 30 and 37 S. J Hydrometeorol 11:822–831
Mathiasen P, Premoli AC (2010) Out in the cold: genetic variation of Nothofagus pumilio (Nothofagaceae) provides evidence for latitudinally distinct evolutionary histories in austral South America. Mol Ecol 19:371–385. https://doi.org/10.1111/j.1365-294X.2009.04456.x
McDowall R, Pankhurst N (2005) Loss of negative eye-size allometry in a population of Aplochiton zebra (Teleostei: Galaxiidae) from the Falkland Islands. N Z J Zool 32:17–22
Menu Marque SA, Marinone MC (1986) El zooplancton de seis lagos del Chubut (Argentina) y sus probables relaciones con la ictiofauna y algunos factores ambientales (The Zooplancton of six lakes from Chubut (Argentina) and their possible relationship with ichthyofauna and some environmental factors). In: Vila I, Fagetti E (eds) Trabajos presentados al taller internacional sobre ecología y manejo de peces en lagos y embalses. Santiago de Chile, 5-10 nov. 1984. FAO, Roma, pp 90–114
Mitra A, Flynn KJ, Burkholder JM, Berge T, Calbet A, Raven JA, Granéli E, Glibert PM, Hansen PJ, Stoecker DK, Thingstad F, Tillmann U, Våge S, Wilken S, Zubkov MV (2014) The role of mixotrophic protists in the biological carbon pump. Biogeosciences 11(4):995–1005. https://doi.org/10.5194/bg-11-995-2014
Modenutti B, Bastidas Navarro M, Martyniuk N, Balseiro E (2018a) Melting of clean and debris-rich ice differentially affect nutrients, dissolved organic matter and bacteria respiration in the early ontogeny of the newly formed proglacial Ventisquero Negro Lake (Patagonia Argentina). Freshw Biol 63:1341–1351. https://doi.org/10.1111/fwb.13161
Modenutti B, Wolinski L, Souza MS, Balseiro EG (2018b) When eating a prey is risky: implications for predator diel vertical migration. Limnol Oceanogr 63:939–950. https://doi.org/10.1002/lno.10681
Modenutti BE, Balseiro EG (2002) Mixotrophic ciliates in an Andean lake: dependence on light and prey of an Ophrydium naumanni population. Freshw Biol 47(1):121–128
Modenutti BE, Balseiro EG (2018) Preface: Andean Patagonian lakes as sensors of global change. Hydrobiologia 816(1):1–2. https://doi.org/10.1007/s10750-018-3622-z
Modenutti BE, Balseiro EG, Bastidas Navarro M, Laspoumaderes C, Souza MS, Cuassolo F (2013a) Environmental changes affecting light climate in oligotrophic mountain lakes: the deep chlorophyll maxima as a sensitive variable. Aquat Sci 75(3):361–371. https://doi.org/10.1007/s00027-012-0282-3
Modenutti BE, Balseiro EG, Bastidas Navarro MA, Lee ZM, Souza MS, Corman JR, Elser JJ (2016) Effects of Volcanic Pumice inputs on microbial community composition and dissolved C/P ratios in lake waters: an experimental approach. Microb Ecol 71(1):18–28. https://doi.org/10.1007/s00248-015-0707-3
Modenutti BE, Balseiro EG, Callieri C, Bertoni R (2008) Light versus food supply as factors modulating niche partitioning in two pelagic mixotrophic ciliates. Limnol Oceanogr 53(2):446–455
Modenutti BE, Balseiro EG, Callieri C, Bertoni R, Queimaliños CP (2005) Effect of UV-B and different PAR intensities on the primary production of the mixotrophic planktonic ciliate Stentor araucanus. Limnol Oceanogr 50(3):864–871
Modenutti BE, Balseiro EG, Callieri C, Queimaliños C, Bertoni R (2004) Increase in photosynthetic efficiency as a strategy of planktonic organisms exploiting deep lake layers. Freshw Biol 49(2):160–169
Modenutti BE, Balseiro EG, Elser JJ, Bastidas Navarro M, Cuassolo F, Laspoumaderes C, Souza MS, Dıaz Villanueva V (2013b) Effect of volcanic eruption on nutrients, light, and phytoplankton in oligotrophic lakes. Limnol Oceanogr 58(4):1165–1175. https://doi.org/10.4319/lo.2013.58.4.0000
Modenutti BE, Balseiro EG, Moeller R (1998) Vertical distribution and resistance to ultraviolet radiation of a planktonic ciliate Stentor araucanus. Verhandlungen Internationale Vereinigung Limnologie 26:1636–1640
Modenutti BE, Perez GL, Balseiro EG, Queimaliños CP (2000) Relationship between light availability, chlorophyll a and total suspended solid in a glacial lake of South Andes. Verh Int Verein Limnol 27(5):2648–2651
Modenutti BE, Queimaliños C, Balseiro EG, Reissig M (2003) Impact of different zooplankton structures on the microbial food web of a South Andean oligotrophic lake. Acta Oecol 24(Suppl 1):289–298
Morris DP, Zagarese H, Williamson CE, Balseiro EG, Hargreaves BR, Modenutti BE, Moeller R, Queimaliños C (1995) The attenuation of solar UV radiation in lakes and the role of dissolved organic carbon. Limnol Oceanogr 40(8):1381–1391
Neukom R, Barboza LA, Erb MP, Shi F, Emile-Geay J, Evans MN, Franke J, Kaufman DS, Lücke L, Rehfeld K (2019) Consistent multi-decadal variability in global temperature reconstructions and simulations over the Common Era. Nat Geosci 12(8):643
Paruelo JM, Beltran A, Jobbágy E, Sala O, Golluscio R (1998) The climate of Patagonia: general patterns and controls on biotic processes. Ecol Austral 8(2):85–101
Paul F, Mölg N (2014) Hasty retreat of glaciers in northern Patagonia from 1985 to 2011. J Glaciol 60(224):1033–1043. https://doi.org/10.3189/2014JoG14J104
Pedrozo F, Chillrud S, Temporetti P, Díaz M (1993) Chemical composition and nutrient limitation in rivers and lakes of northern Patagonian Andes (39.5°-42° S; 71° W) (Rep. Argentina). Verh Int Verein Limnol 25:205–214
Pereyra FX, Bouza P (2019) Soils from the Patagonian region. In: The soils of Argentina. World Soils Book Series, pp 101–121. https://doi.org/10.1007/978-3-319-76853-3_7
Pérez G, Queimaliños C, Balseiro EG, Modenutti BE (2007) Phytoplankton absorption spectra along the water column in deep North Patagonian Andean lakes (Argentina): Limnology of Temperate South America. Limnologica 37(1):3–16
Pérez GL, Queimaliños CP, Modenutti BE (2002) Light climate and plankton in the deep chlorophyll maxima in North Patagonian Andean lakes. J Plankton Res 24(6):591–599
Pessacg N, Flaherty S, Solman S, Pascual M (2020) Climate change in northern Patagonia: critical decrease in water resources. Theor Appl Climatol:1–16
Poveda G, Espinoza JC, Zuluaga MD, Solman SA, Garreaud Salazar R, van Oevelen PJ (2020) High impact weather events in the Andes. Front Earth Sci 8:162. https://doi.org/10.3389/feart.2020.00162
Quéguiner B, Legendre L (1986) Phytoplankton photosynthetic adaptation to high frequency light fluctuations simulating those induced by sea surface waves. Mar Biol 90(4):483–491
Queimaliños C, Reissig M, Perez GL, Soto Cardenas C, Gerea M, Garcia PE, Garcia D, Dieguez MC (2019) Linking landscape heterogeneity with lake dissolved organic matter properties assessed through absorbance and fluorescence spectroscopy: Spatial and seasonal patterns in temperate lakes of Southern Andes (Patagonia, Argentina). Sci Total Environ 686:223–235. https://doi.org/10.1016/j.scitotenv.2019.05.396
Quirós R (1988) Relationship between air temperature, depth, nutrient and chlorophyll in 103 Argentinian lakes. Verhandlungen Internationale Vereinigung Limnologie 23:647–658
Quirós R (1997) Classification and state of the environment of the Argentinean lakes. In: Study report for the lake environment conservation in developing countries: Argentina (Ed. ILEC Workshop on Better Management of the Lakes of Argentina), ILEC Workshop on Better Management of the Lakes of Argentina, pp 29–50
Quirós R, Drago E (1999) The environmental state of Argentinean lakes: an overview. Lakes Reserv Res Manag 4(1-2):55–64
Reissig M, Modenutti BE, Balseiro EG, Queimaliños C (2004) The role of the Predaceous Copepod Parabroteas Sarsi in the Pelagic Food Web of a large deep Andean lake. Hydrobiologia 524(1):67–77
Richter A, Marderwald E, Hormaechea JL, Mendoza L, Perdomo R, Connon G, Scheinert M, Horwath M, Dietrich R (2016) Lake-level variations and tides in Lago Argentino, Patagonia: insights from pressure tide gauge records. J Limnol 75(1). https://doi.org/10.4081/jlimnol.2015.1189
Rivera JA, Araneo DC, Penalba OC, Villalba R (2018) Regional aspects of streamflow droughts in the Andean rivers of Patagonia, Argentina. Links with large-scale climatic oscillations. Hydro Res 49(1):134–149
Roe G, Baker M, Herla F (2017) Centennial glacier retreat as categorical evidence of regional climate change. Nat Geosci (10):95–99. https://doi.org/10.1038/ngeo2863
Ruiz L, Berthier E, Masiokas HM, Pitte P, Villalba R (2015) First surface velocity maps for glaciers of Monte Tronador, North Patagonian Andes, derived from sequential Pléiades satellite images. J Glaciol 61(229):908–922. https://doi.org/10.3189/2015JoG14J134
Ruiz L, Berthier E, Viale M, Pitte P, Masiokas MH (2017) Recent geodetic mass balance of Monte Tronador glaciers, northern Patagonian Andes. Cryosphere 11(1):619–634. https://doi.org/10.5194/tc-11-619-2017
Schenone L, Balseiro EG, Bastidas Navarro M, Modenutti BE (2020) Modelling the consequence of glacier retreat on mixotrophic nanoflagellate bacterivory: a Bayesian approach. Oikos 129(8):1216–1228. https://doi.org/10.1111/oik.07170
Sharples JC, Moore M, Rippeth TP, Holligan PM, Hydes DJ, Fisher NR, Simpson JH (2001) Phytoplankton distribution and survival in the thermocline. Limnol Oceanogr 46(3):486–496
Shugar DH, Burr A, Haritashya UK, Kargel JS, Watson CS, Kennedy MC, Bevington AR, Betts RA, Harrison S, Strattman K (2020) Rapid worldwide growth of glacial lakes since 1990. Nat Clim Change 10(10):939–945. https://doi.org/10.1038/s41558-020-0855-4
Smith DW, Cooper SD (1982) Competition among cladocera. Ecology 63(4):1004–1015
Sommaruga R, Kandolf G (2014) Negative consequences of glacial turbidity for the survival of freshwater planktonic heterotrophic flagellates. Sci Rep 4:4113. https://doi.org/10.1038/srep04113
Sommer U (1989) The role of competition for resources in phytoplankton succession. In: Sommer U (ed) Plankton ecology: succession in plankton communities. Springer-Verlag, Berlin, pp 57–106
Souza MS, Modenutti BE, Carrillo P, Villar-Argaiz M, Medina-Sánchez JM, Bullejos F, Balseiro EG (2010) Stoichiometric dietary constraints influence the response of copepods to ultraviolet radiation-induced oxidative stress. Limnol Oceanogr 55(3):1024–1032
SRL SET (2018) Software de Procesamiento de Imágenes (SoPI) (Software for Image Processing (SoPI)). 3.0 edn. Comisión Nacional de Actividades Espaciales (CONAE) de la República Argentina, https://www.argentina.gob.ar/ciencia/conae/unidad-educacion/software/sopi
Sterner RW, Elser JJ (2002) Ecological stoichiometry. The biology of elements from molecules to the biosphere. Princeton University Press, Princeton
Sterner RW, Elser JJ, Fee EJ, Guildford SJ, Chrzanowski TH (1997) The light:nutrient ratio in lakes: the balance of energy and materials affects ecosystem structure and process. Am Nat 150(6):663–684
Stomp M, Huisman J, Stal LJ, Matthijs HC (2007a) Colorful niches of phototrophic microorganisms shaped by vibrations of the water molecule. ISME J 1(4):271–282. https://doi.org/10.1038/ismej.2007.59
Stomp M, Huisman J, Voros L, Pick FR, Laamanen M, Haverkamp T, Stal LJ (2007b) Colorful coexistence of red and green picocyanobacteria in lakes and seas. Ecol Lett 10(4):290–298
Stuart-Smith RD, Stuart-Smith JF, White RWG, Barmuta LA (2007) The impact of an introduced predator on a threatened galaxiid fish is reduced by the availability of complex habitats. Freshw Biol 52(8):1555–1563
Tartarotti B, Baffico G, Temporetti P, Zagarese HE (2004) Mycosporine-like amino acids in planktonic organisms living under different UV exposure conditions in Patagonian lakes. J Plankton Res 26(7):753–762. https://doi.org/10.1093/plankt/fbh073
Thomasson K (1959) Nahuel Huapi: Plankton of some lakes in an Argentine national park, with notes on terrestrial vegetation. Acta Phytogeogr Suec 42:1–83
Thomasson K (1963) Araucanian lakes. Acta Phytogeogr Suec 47:1–139
Trauth MH, Alonso RA, Haselton KR, Hermanns RL, Strecker MR (2000) Climate change and mass movements in the NW Argentine Andes. Earth Planet Sci Lett 179(2):243–256
Utne ACW (1997) The effect of turbidity and illumination on the reaction distance and search time of the marine planktivore Gobiusculus flavescens. J Fish Biol 50:926–938
Viale M, Bianchi E, Cara L, Ruiz LE, Villalba R, Pitte P, Masiokas M, Rivera J, Zalazar L (2019) Contrasting climates at both sides of the Andes in Argentina and Chile. Front Environ Sci 7. https://doi.org/10.3389/fenvs.2019.00069
Viale M, Valenzuela R, Garreaud RD, Ralph FM (2018) Impacts of atmospheric rivers on precipitation in southern South America. J Hydrometeorol 19(10):1671–1687
Villafañe VE, Buma AGJ, Boelen P, Helbling EW (2004) Solar UVR-induced DNA damage and inhibition of photosynthesis in phytoplankton from Andean lakes of Argentina. Arch Hydrobiol 161(2):245–266
Villalba R, Lara A, Boninsegna JA, Masiokas M, Delgado S, Aravena JC, Roig FA, Schmelter A, Wolodarsky A, Ripalta A (2003) Large-scale temperature changes across the southern Andes: 20th-century variations in the context of the past 400 years. In: Climate variability and change in high elevation regions: past, present & future. Springer, pp 177–232
Vinyard GL, O’Brien WJ (1976) Effects of light and turbidity on the reactive distance of bluegill (Lepomis macrochirus). J Fish Board Canada 33(12):2845–2849
Waibel A, Peter H, Sommaruga R (2019) Importance of mixotrophic flagellates during the ice-free season in lakes located along an elevational gradient. Aquat Sci 81(3). https://doi.org/10.1007/s00027-019-0643-2
Wilson R, Glasser NF, Reynolds JM, Harrison S, Anacona PI, Schaefer M, Shannon S (2018) Glacial lakes of the Central and Patagonian Andes. Global Planet Change 162:275–291. https://doi.org/10.1016/j.gloplacha.2018.01.004
Wolinski L, Laspoumaderes C, Bastidas Navarro M, Modenutti BE, Balseiro EG (2013) The susceptibility of cladocerans in North Andean Patagonian lakes to volcanic ashes. Freshwat Biol 58(9):1878–1888. https://doi.org/10.1111/fwb.12176
Worni R, Stoffel M, Huggel C, Volz C, Casteller A, Luckman B (2012) Analysis and dynamic modeling of a moraine failure and glacier lake outburst flood at Ventisquero Negro, Patagonian Andes (Argentina). J Hydrol 444-445:134–145. https://doi.org/10.1016/j.jhydrol.2012.04.013
Zagarese HE, Ferraro M, Queimaliños C, Diéguez MC, Suárez DA, Llames ME (2017) Patterns of dissolved organic matter across the Patagonian landscape: a broad-scale survey of Chilean and Argentine lakes. Mar Freshw Res 68(12). https://doi.org/10.1071/mf17023
Acknowledgments
This work was supported by FONCyT PICT2017-1940, PICT2018-1563, PICT2019-0950, CONICET PUE2016-0008, and UNComahue B236.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Balseiro, E., Modenutti, B., Bastidas Navarro, M., Martyniuk, N., Schenone, L., Laspoumaderes, C. (2022). North Patagonian Andean Deep Lakes: Impact of Glacial Recession and Volcanic Eruption. In: Mataloni, G., Quintana, R.D. (eds) Freshwaters and Wetlands of Patagonia. Natural and Social Sciences of Patagonia. Springer, Cham. https://doi.org/10.1007/978-3-031-10027-7_3
Download citation
DOI: https://doi.org/10.1007/978-3-031-10027-7_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-10026-0
Online ISBN: 978-3-031-10027-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)