
Abstract
North America has accumulated more naturalized species, or “alien species that form self-sustaining populations in new regions,” than any other continent (5958; 3513 of which come from outside the continent). Over the last 35 years, North America has seen a rapid increase in the number of plant invasions; species have arrived through the horticulture and aquarium trades, as agricultural contaminants, and via other accidental and intentional pathways. Introduced populations have persisted and expanded on the continent with the help of extensive land use change and growing transportation networks. The main driving forces of naturalization in North America have been found to be habitat legacy (i.e., the habit affinities of a plant species in their native range), propagule pressure, and residence time, which are modulated by specific biological traits (e.g., flowering periods, vigorous clonal growth, and tall stature) interacting differently with these drivers. Within North America, more invasive plant species are found in cold temperate and Mediterranean climate zones than in arid, temperate arid, subtropical, and tropical climates. Economic activity (measured as per capita gross domestic product; GDP) also heavily influences the distribution of non-native species; areas with higher per capita GDP (~17,000 USD) have over twice the number of non-native plant species compared to regions with lower GDP. Currently, in the United States alone, over $100 billion per year is spent on losses, damages, and the control of invasive species, with the bulk of those funds going toward weed control for crops, pastures, and forests. In addition to being a drain on the economy, invasive non-native plants disrupt ecosystems and can often have negative effects on ecosystem services, including altered hydrological and fire regimes, impacts on native species, and changes in soil properties and nutrient cycling. National legislation and international agreements have been implemented in attempts to reduce threats from non-native species to biodiversity, the economy, and human well-being.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Alien species
- Climate zone
- Global trade
- Invasive species
- Life history
- Non-native species
- Plant introductions
8.1 Introduction
This chapter focuses on plant invasions in North America and includes research from the United States (excluding territories), Mexico, and Canada. The aims for this chapter are to (1) describe the recorded history of biological invasions; (2) discuss the introduction pathways for non-native species into North America, including the roles of trade, transport, agriculture, horticulture, and disturbance on these introductions; (3) explore the plant traits and life histories that increase invasion success of non-native plants in North America; (4) describe the climate zones that are most heavily invaded by non-native plants; (5) examine the impacts that invasive plants have on ecosystem services, including impacts on hydrological and fire regimes, native species, soil properties, and nutrient cycling; and (6) discuss some of the national and international policy and legislation that has been enacted to control the spread of invasive plants in North America
8.1.1 A History of Biological Invasions in North America
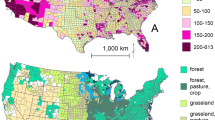
North America’s history of biological invasions can be divided into three periods. The first phase of modern biological invasions began around the year 1500 with the advent of global exploration, specifically the European rediscovery of the Americas, the birth of colonialism, and changes to agricultural trade and industry (Hewitt et al. 2009; Hulme 2009). By 1800 AD, the industrial revolution had begun, and along with it the second major phase of biological invasions in North America (Mack 2003; Hulme 2009). This period was defined by an exponential increase in global trade as well as a steady increase in the annual rate of plant introductions (Mack 2003; Hulme 2009). During this period, there were increases in international trade through new transportation routes and technologies, including canals, highways, railways, and steamships, and increases in emigration; over 50 million Europeans arrived in the United States between 1820 and 1930. This increase in international activity provided avenues for the introduction of non-native plants (McNeely 2006; Findlay and O’Rourke 2007; Hulme 2009). Over the last 35 years, North America has seen a rapid increase in the number of plant invasions, suggesting a “step change” in biological invasions, bringing us into the third and current phase, the Era of Globalization (Hulme 2009). Recently, Seebens et al. (2017) showed that this increase in the number of non-native species shows no sign of saturation at the global scale. North America has accumulated more naturalized species than other areas of the world (van Kleunen et al. 2015; Pyšek et al. 2017; Seebens et al. 2021); these species are distributed unevenly across countries and their respective states or provinces (Fig. 8.1).

Number of naturalized species per state or province for each North American country. Data was obtained from Global Naturalized Alien Flora (GloNAF) database (van Kleunen et al. 2019; GloNAF; https://glonaf.org/). (a) United States of America; (b) Canada; (c) Mexico
8.1.2 The Introduction of Non-native Species into North America
Non-native species have made their way into North America through a variety of pathways, including as stowaways on trade and transportation routes, and intentionally or unintentionally through agriculture, horticulture, and the aquarium trade. Land use change and disturbance have altered the receptivity of the landscape to invasions. Interestingly, we now know through advances in genetic technology and the tracing of genealogies of invasive plants that many species became invasive only after multiple introductions (Oduor et al. 2015). For example, chloroplast DNA sequences showed that black mustard (Brassica nigra) arrived in North America from multiple sources in its native range (Oduor et al. 2015).
8.1.2.1 Trade and Transport
Global trade and travel are primary drivers of the spread of invasive plants, and the step increase in invasive species recorded in recent decades could be linked to faster transportation routes, e.g., semi-trucks, trains, etc. (Meyerson and Mooney 2007; Hulme 2009; Seebens et al. 2015, 2017). International trade and travel are the primary sources of non-native plant stowaways or seed contaminants in goods and packaging material arriving by means of passenger planes and marine shipping ports (Hulme 2009; Tatem 2009; Levine and D’Antonio 2015; Bellard et al. 2016; Early et al. 2016). Generally, regions that have higher seaport and airport capacity and greater total imports are considered to be at greater risk of being invaded by a non-native species (Fig. 8.2; Early et al. 2016).

(a) Invasion threat to North America for the twenty-first century. Colors indicate the level of invasion threat, from very high to low. The scale was determined by ranking the threat values of each of the map grid cells. The cells were then binned into percentiles: 100–90%, very high (VH); 90–80%, high (H); 80–50%, medium (M); 50–20%, low (L); and 20–0%, very low (VL). (b) The impact of the combination of airport capacity and total imports on invasion threat. (c) Seaport capacity, an indicator of invasion threat from shipping. Colors are the same as indicated in Fig. 8.2a. All panels excerpted from Early et al. (2016)
International air travel is playing an increasingly significant role in driving increases in the rates of biological introductions globally (Tatem 2009). Global airline traffic connects regions of similar climates. Climatic similarity with the native region is one of the requirements for establishment of non-native species (Bellard et al. 2016), along with propagule pressure and suitability of other abiotic factors (Hulme 2009). Thus, increases in travel and local climatic changes could further increase the risk of the movement of non-native species and establishment in new areas (Ismail et al. 1999; Tatem 2009; Bellard et al. 2016). As illustrated in Fig. 8.2b, spatial patterns of air travel and importation of goods could affect the areas under threat from invasive species in the future (Early et al. 2016). In North America, the area that appears to have the greatest implied threat of invasive species arrival is the eastern portion of the United States.
Seaports have been a classic symbol for trade and economic development for centuries, but they are also gateways for introducing non-native species (Bellard et al. 2016). One means of introduction of non-native species in marine environments caused by shipping movements is the discharge of ballast material (Seebens et al. 2015; Bellard et al. 2016). In order for ships to remain balanced as they load and unload goods, the bottom of the ship is filled with ballast, any solid or liquid that is brought on the vessel to maintain stability. In the past, solid materials such as sand, rocks, and soils were commonly used and were often left behind at different destinations. This dry ballast was sometimes (if not frequently) contaminated with non-native plant seeds, resulting in new introductions (Mack 2003; Parks et al. 2005). One of the first studies exploring the possibility of alien plant species within dry ballast material found that of the 213 species identified, 93 species (~43%) were considered alien to the area investigated in Oregon, USA (Nelson 1917). As a result of these large number of alien species, this practice becomes obsolete, but now ships use seawater for ballast, transferring a significant amount of ballast water between different continents and oceans (Government of Canada 2010; Werschkun et al. 2014).
8.1.2.2 Agriculture, Horticulture, and the Aquarium Trade
Many non-native species have been introduced to North America as seed contaminants in agriculture, through the cut flower trade, through the aquarium trade, or through forestry and horticulture. Many non-native agricultural weeds have been accidentally introduced as contaminants in crop seeds, despite the Federal Seed Act (FSA) in the United States (Ismail et al. 1999). FSA regulates the interstate and foreign transport of agricultural and vegetable seeds, requiring labeling and purity standards in commerce (US FSA 7th Cong. 1551–1611 (1988)). Cut flowers can also have seed contaminants within the flowers, which can be transported to new regions when airplane passengers carry bunches of flowers from one country to another (Ismail et al. 1999). The majority of plants used in agriculture, forestry, and horticulture in North America are non-native (Reichard and White 2001), and while most of these plants do not become invasive, a small proportion of species has escaped cultivation, established self-sustaining natural populations without human intervention, and become invasive (Reichard and White 2001; van Kleunen et al. 2018). For instance, St. John’s wort (Hypericum perforatum) was introduced in the late 1700s as an ornamental for medicinal purposes in the eastern United States and was later found in the western United States in the mid-1800s (Campbell and Delfosse 1984; Reichard and White 2001; Mack 2003). H. perforatum is considered a noxious weed in both its native and non-native ranges and has harmful effects on livestock and both natural and agricultural lands (Mack 2003; Popay 2015). The species is now distributed throughout most states in the United States and in eastern provinces and British Columbia in Canada (Zouhar 2004); it is considered invasive in western North America (Popay 2015). Since its introduction, the abundance of H. perforatum has been effectively restrained in much of its new range through biological control (Mack 2003; Popay 2015).
The aquarium trade also presents a major risk for the introduction of aquatic non-native plants (Padilla and Williams 2004; Ricciardi et al. 2017; Della Venezia et al. 2018). In the United States, a major problem is that aquatic plants listed as federal or noxious weeds can nonetheless be purchased online, and most of the global trade likely takes place via this avenue. This creates difficulties for governments to enforce regulations, although US governmental agencies have been making headway in dealing with Internet trade (Padilla and Williams 2004). Water hyacinth (Eichhornia crassipes) is believed to have been introduced into Floridian waterways following the World’s Industrial and Cotton Centennial Exposition in New Orleans in 1884, when it was brought home as a souvenir and released. E. crassipes has been nicknamed “the world’s most troublesome weed” as a result of its ability to double in population size and create dense floating mats in about a week (Gopal and Sharma 1981; Padilla and Williams 2004). Since its introduction, Florida has appropriated millions of US dollars of state and federal funding to achieve limited control of E. crassipes (Padilla and Williams 2004). It is now found in several states, the province of Ontario in Canada, and in parts of Mexico.
While non-native plants are primarily introduced deliberately through the ornamental plant trade (Reichard and White 2001; Mack and Erneberg 2002; Lehan et al. 2013), accidental introductions through seed contamination are of increasing concern as a major pathway for invasive plants (Lehan et al. 2013; Fig. 8.3). In the eastern United States, the majority of non-native angiosperms (64% of surveyed species) were introduced deliberately, whereas only 2% arrived accidentally as seed contaminants and the remainder of species had unknown origins (Mack and Erneberg 2002; Lehan et al. 2013). In the western United States, a high proportion of invasive plants – primarily grasses and forbs – arrived accidentally as seed contaminants, whereas in eastern states non-native plants primarily arrived through deliberate introduction as ornamentals (Lehan et al. 2013). In fact, although all types of invasive plants are more likely to be introduced deliberately, introduction pathways vary considerably by plant growth habit; a significantly larger percentage of forbs and grasses were accidentally introduced compared to vines, shrubs and trees. In the United States, accidental introductions account for at least 12% of all invasive plants and 21% of noxious weeds; these are likely underestimates because many species have unknown introduction pathways (Lehan et al. 2013). Seed contaminants account for the majority of accidental introductions in the United States (Lehan et al. 2013).

Pathways for invasive plant introductions in the continental United States. (a) Deliberate, accidental, and unknown introduction pathways for invasive plants. Specific introduction pathways for accidentally (b) and deliberately (c) introduced invasive plants. Data from Mack and Erneberg (2002) and Lehan et al. (2013)
8.1.2.3 Land Use Change and Disturbance
As globalization increases, so does the amount of land use change and disturbance (Houghton and Nassikas 2017). In North America, the landscape is evolving to keep up with the demands of a rising population and global markets, as land is converted to support transportation routes, agricultural production, and energy infrastructure, among other things (Bradley et al. 2010; Houghton and Nassikas 2017). Disturbances to the land harm native plant populations and can provide opportunities for non-native species to benefit, for instance, by providing rapid large-scale increases in resource availability (Bradley et al. 2010). An example of this is deforestation, which increases light and belowground resource availability in a very short time frame. A strong positive relationship between physical disturbance and plant invasion has been found in several studies (as reviewed by Bradley et al. 2010). Disturbance types include roads (Larson 2002; Gelbard and Belnap 2003; Bradley and Mustard 2006), deforestation (Yates et al. 2004; Fan et al. 2013), urban areas (Turner et al. 2005; Pennington et al. 2010), energy development (Barney 2014; Villarreal et al. 2019), and agriculture (Pimentel 2009). The novelty, magnitude, and rate of disturbances often disadvantage native plants, causing the system to become less resistant to invasion (Bradley et al. 2010). Changes in disturbance regimes can allow for a different set of species, potentially invasive species, to persist and are therefore important for understanding invasive species establishment.
The intensity and global patterns of invasive plant establishment and disturbances are changing at a faster rate than any other period in recorded human history (Seebens et al. 2015; Early et al. 2016), indicating that the geographical patterns of future invasions will likely be very different from today as land use change continues to expand (Foley et al. 2005; Theoharides and Dukes 2007; Early et al. 2016). In North America, many non-native species take advantage of such disturbances to become widely established and can subsequently suppress native species’ populations (Bradley et al. 2010). In a study exploring the patterns of invasive plant diversity in Northwest mountain ecoregions, Parks et al. (2005) found that disturbed riparian systems and forests are especially vulnerable to plant invasion, whereas alpine and wilderness areas remain relatively unaffected by invasive plants. One such species that has been found to establish and thrive following a disturbance is pale swallow-wort (Vincetoxicum rossicum), a perennial, herbaceous vine native to southern Europe, which is of particular concern in the northeastern United States and southeastern Canada (Parks et al. 2005). It is expected that intense land use will likely continue to enhance invasive plant introductions into these low elevation mountain ecosystems (Parks et al. 2005).
8.2 Life History and Origin of Non-native Plants
Researchers often look to non-native plants’ traits or life histories, including properties such as growth rate, nutrient use efficiency, stress tolerance, herbivore resistance, and reproduction strategy, to better understand why certain species become more invasive than others (Theoharides and Dukes 2007; Pyšek and Richardson 2008; Ricklefs et al. 2008; Pyšek et al. 2017). For example, Pyšek et al. (2015) used a model to identify plant traits that promote naturalization of central European species in North America and confirmed that traits such as longer flowering periods, vigorous clonal growth, and tall stature promoted invasion success in North America. Ultimately, the main driving force of naturalization has to do with a myriad of life history traits, and Pyšek (2015) demonstrated that habitat legacy, propagule pressure, and residence time are important driving forces in North America.
While Pyšek et al. (2015) specifically explored the naturalization of European species in North America, invasive plants from other regions tend to possess these same traits. For example, the Asian species Japanese knotweed (Reynoutria japonica) is a highly aggressive, invasive perennial in Europe and North America. The clonal species was introduced into the United Kingdom as a garden ornamental around 1830 and was reported as naturalized in the United States by 1894 (Merhoff et al. 2003; Grimsby et al. 2007). Clonal spread by rhizomes and rapid growth have been suggested as the major characteristics that make R. japonica difficult to control and a problematic invader (Grimsby et al. 2007). Interestingly, Grimsby et al. (2007) found evidence that F. japonica implements different reproductive strategies in Europe and North America. In the United Kingdom, R. japonica is a single female clone reproducing exclusively through vegetative growth or hybridization with other Reynoutria spp. (Bailey 1994; Hollingsworth and Bailey 2000; Grimsby et al. 2007). However, in the United States, Grimsby et al. (2007) found evidence of both asexual and sexual reproduction, resulting in populations of knotweed that are genetically diverse, as these populations are not purely clonal. These complex reproductive dynamics of Reynoutria spp. and their widespread distribution indicate that the invasive potential of these species is far-reaching and is not fully realized (Grimsby et al. 2007). Further, this suggests that the invasion biology of Reynoutria spp. in the United States is different from that of knotweeds in Europe, demonstrating a need for regional studies of this likely rapidly evolving taxon in North America (Grimsby et al. 2007).
In addition to high genetic diversity, another trait that has been suggested to promote naturalization is a small genome size (Lavergne et al. 2010b; Pyšek et al. 2017, 2018). It has been theorized that small genomes allow species to have a shorter generation time and reproduce more quickly, leading to higher propagule pressure (Rejmanek and Richardson 1996; Pyšek et al. 2015). Comparative studies have shown that species with smaller genomes are more likely to become invasive than their relatives (Kubešová et al. 2010; Lavergne et al. 2010a, b; Pandit et al. 2014). Reed canary grass (Phalaris arundinacea), an invasive species in North America that is considered a major threat to native wetland vegetation and bird breeding habitat (Lavergne and Molofsky 2006; Lavergne et al. 2010b), has been used as a model to investigate whether reduced genome size could result in rapid phenotypic evolution (Lavergne et al. 2010b; Pyšek et al. 2018). It is suggested that the invasiveness of this species in North America was a result of multiple introductions and subsequent recombination of multiple European strains, allowing for rapid selection of novel genotypes with higher potential for vegetative reproduction (Lavergne and Molofsky 2004, 2007; Lavergne et al. 2010b). These novel genotypes of P. arundinacea in its invasive range have a smaller genome than European genotypes (Lavergne et al. 2010b), and this smaller genome size was associated with plant traits favoring invasiveness (long rhizomes, early emerging abundant shoots, resistance to aphid attack, and low C:N ratio; Pyšek et al. 2018).
8.3 Plant Invasions by Climate Zone
In North America, certain types of climate are more heavily invaded than others. In this section, we explore the number of invasive plant species that invaded each of North America’s five most common climate zones: tropical (equatorial and savanna), subtropical (arid), Mediterranean, temperate (warm, arid, and cold/boreal), and Arctic (excerpted from the zonobiomes outlined in Pyšek et al. (2017) and by Walter and Breckle (1991); Fig. 8.4a).

(a) Naturalization in North America according to zonobiome. The red dots indicate naturalization hotspots based on the percentage of naturalized species within regional floras. Excerpted from Pyšek et al. 2017. (b) Geographic patterns of forest plant invasion in the contiguous 48 states of the United States for (top) invasion richness and (bottom) invasion prevalence. Excerpted from Iannone et al. (2015)
In North America, the colder temperate and Mediterranean climate zones harbor approximately twice the number of non-native plant species as regions with arid, temperate arid, subtropical and tropical climates (Pyšek et al. 2017). For example, forests in the eastern United States have been found to harbor more invasive species than those in the western regions of the country (Iannone et al. 2015; Oswalt et al. 2015; Fig. 8.4b). Per capita GDP heavily influences the distribution of non-native species in temperate arid, subtropical, and tropical regions. Areas with higher per capita GDP (~17,000 USD per capita) have over twice the percentage of non-native plant species in their floras (16%) compared to regions with lower GDP (6%). Table 8.1 contains a list of the most widely distributed naturalized species in North America according to climate zone (Pyšek et al. 2017). Note that these widely distributed species are not necessarily those thought to have the greatest ecological or economic impacts, or those of the greatest management concern.
8.4 Impacts on Ecosystem Processes and Services
Invasive non-native plants threaten ecosystems, habitats, and native species and are key drivers of human environmental change. Ecosystems provide a number of benefits, both ecological and economic, known as ecosystem services, e.g., provision of habitat, storage of carbon, and pollination (Fisher et al. 2009; IPBES 2018). According to the most recent estimates, invasive species inflict over $100 billion per year in associated losses, damages, and management expenses in the United States alone (Pimentel et al. 2005). In addition to being a drain on the economy, invasive non-native plants disrupt ecosystems (Dukes and Mooney 2004), altering ecosystem processes, and often have negative effects on ecosystem services (Charles and Dukes 2007). Invasive plants are abundant in all major habitats in North America, but their impacts on biodiversity, cultural values, and economics differ among subregions (IPBES 2018). Here, we discuss the impacts that invasive plants have on ecosystem services in North America.
8.4.1 Hydrological and Fire Regimes
Terrestrial ecosystems provide hydrological services that benefit people by providing freshwater supplies (Brauman et al. 2007) and flood control. Invasive plants can alter the hydrology of ecosystems by changing the rate and/or timing of evapotranspiration (ET) or runoff, as a consequence of differences in transpiration rates, phenology, growth, or rooting depth of non-native versus native species (Levine et al. 2003). Some invasive plant species, particularly trees, use more water than native species (Calder and Dye 2001). Hydrological studies investigating the effects of removing invasive woody species have shown reductions in ET (Cleverly et al. 2006), rises in the water table (Asbjornsen et al. 2007), and increased water yield (Dye and Jarmain 2004). For example, a removal experiment in Hawaii exploring the effects of three invasive tree species, Cecropia obtusifolia, Macaranga mappa, and Melastoma septemnervium, on stands of the native tree species Metrosideros polymorpha found that stand-level water use within removal plots was half that of the invaded plots, despite a significant increase in compensatory water use by the native tree (Cavaleri et al. 2014). However, other experiments have shown the potential for increased stand ET after invasive woody plant removal as a result of compensatory water use by the remaining native species (Moore and Owens 2012) and little to no changes in water yield (Doody et al. 2011; Moore and Owens 2012). Alterations to the hydrology of the ecosystem as a result of invasion depend largely on the species involved and site-specific conditions (Dye and Jarmain 2004; Doody et al. 2011; Cavaleri et al. 2014). Changes in hydrology are not only caused by invasion of woody plant species; some herbaceous invasive species have been shown to alter water cycling. For example, in annual grasslands in western North America, yellow starthistle (Centaurea solstitialis) has increased summer water use by 105–120 mm year−1 (Gerlach 2001; Levine et al. 2003). Invasive annual grasses with shallow root systems, such as cheatgrass (Bromus tectorum), can change hydrology by competitively excluding deeper-rooted native perennials (Dyer and Rice 1999; Bradley et al. 2018; Fusco et al. 2019).
While fires are natural events in many ecosystems, invasive species can alter fire regimes, and there are many examples of invaders increasing fire frequency. This is particularly true of invasions of grasses into otherwise woody species-dominant systems; some non-native grasses create a more continuous fuel bed that was not previously found in the invaded system (Levine et al. 2003; Fusco et al. 2019). B. tectorum in particular has been found to dramatically accelerate fire regimes in western North America (Brooks et al. 2006; Balch et al. 2013; Bradley et al. 2018). In a study exploring fire frequency in the Great Basin, United States, from 1980 to 2009, where B. tectorum dominates at least 6% of landscape, B. tectorum was found to burn nearly four times more frequently than any other type of native vegetation (Balch et al. 2013). Fires were more likely to ignite in cheatgrass than in other types of vegetation, and cheatgrass was associated with increased fire frequency, size, and duration, suggesting even small amounts of cheatgrass in an ecosystem can increase fire risk and alter fire regimes (Balch et al. 2013; Bradley et al. 2018). Most of the native shrub species cannot persist in locations where B. tectorum increases fire frequency (Whisenant 1990; Brooks et al. 2006). In addition to B. tectorum, Fusco et al. (2019) found evidence for significant alteration to regional fire regimes for seven additional invasive grass species in different ecoregions of the United States: Taeniatherum caput-medusae in the Great Basin, Pennisetum ciliare and Schismus barbatus in the desert southwest, Microstegium vimineum and Miscanthus sinensis in eastern temperate deciduous forests, and Imperata cylindrica and Neyraudia reynaudiana in southern pine savannah and pine rockland communities. These eight invasive grass species are associated with increases in rates of fire occurrence of 27–230% in the United States (Fusco et al. 2019).
In contrast, the invasion of stem-succulent plants can increase the moisture content of live fuels, making it more difficult for fires to ignite and spread (Brooks et al. 2006). The reduction of fires in some regions of North America reduces the recruitment and growth of native shrub species (D’Antonio et al. 1993; Brooks et al. 2006). Invasion of the non-native succulent, hottentot-fig (Carpobrotus edulis), has become a common event after a fire in the maritime chaparral in California, USA (Zedler and Scheid 1988). In this case, the invasion of C. edulis reduces fire frequency and can eventually lead to the conversion of maritime chaparral to a mix of succulent- and shrub-dominated vegetation (Brooks et al. 2006).
8.4.2 Consequences for the Displacement of Native Species by Non-native Plants
Invasive plant species displace and suppress populations of native plant species, which can have significant economic as well as ecological impacts on plant communities. For example, yellow starthistle (Centaurea solstitialis), which infests rangelands, is unpalatable to cows and toxic to horses and costs California $7.65 million annually in livestock forage loss and ranchers an additional $9.45 million in out-of-pocket expenditures (Eagle et al. 2007; Pejchar and Mooney 2009).
The impact of invasive species on agriculture is not just felt in terrestrial agriculture, but also in aquatic food production (Pejchar and Mooney 2009). Eurasian watermilfoil (Myriophyllum spicatum) is one of the most widely distributed non-native aquatic plants in North America; the weed is established in 48 states (expect Hawaii and Wyoming) and in the Canadian Provinces of British Columbia, Ontario, and Quebec (U.S. Fish & Wildlife Service 2018). Myriophyllum spicatum has high environmental and socio-economic impacts as well as moderate beneficial impacts in the Great Lakes. The environmental impacts of M. spicatum included a potential reduction in the abundance and diversity of non-native insects and other benthic macroinvertebrates compared to native communities (Keast 1984), a reduction in the growth and vigor of warm-water fishery, and a reduction in nutritional value for waterfowl compared to native plants it replaces (Aiken et al. 1979). The species can alter the hydrology of waterbodies and create stagnant water conditions for parasites that cause swimmer’s itch and promote mosquitoes (Jacobs and Margold 2009). Further, dense mats of M. spicatum can reduce water flow or clog agricultural, residential, or commercial water intakes, which can be expensive to unclog (Jacobs and Margold 2009; U.S. Fish and Wildlife Service 2018).
In addition to altering agroecosystems, invasive plants also have consequences for the biodiversity of native plant communities. Researchers often suggest strong competitive effects of invasive species on the growth, reproduction, and resource allocation of native residents as a mechanism for plant invasion (Levine et al. 2003). Dyer and Rice (1999) found that non-native annual vegetation changed seasonal patterns of resource availability in California’s inland grasslands. Here, increased competition for light in the spring, when rapid growth of annuals tends to occur, suppresses the growth of native perennial bunchgrasses and reduces access to belowground resources through competitive inference. These changes in resource availability eventually result in the loss of perennial grasses, general dominance of non-native annual species, and a relative underutilization of deep soil resources. This example shows how phenology of non-native species and the seasonal shifts in resource availability as a result of their dominance can shift the primary limiting resource from soil moisture to light and alter the seasonal timing of resource limitation (Dyer and Rice 1999).
In addition to affecting biodiversity through competition and by changing disturbance regimes, non-native plant species have been found to affect native plant species by hybridizing with them. Hybridization can have evolutionary consequences; native species can be reduced or lost as native genes are diluted by invasive genes. Hybridization can play a role in the successful spread of invasive plants, hybrids, and their genes (Ellstrand and Schierenbeck 2000; Blair and Hufbauer 2010). Hybridizations can be interspecific or intraspecific, so it is possible that a hybridization event among well-differentiated populations within the same species may act in the same way as hybridization among species and serve as a stimulus for the evolution of invasiveness (Ellstrand and Schierenbeck 2000). One intraspecific hybridization event(s) that increased invasiveness in North America took place in B. tectorum. Bromus tectorum in North America was found to have greater within-population genetic variation than populations in its native range, likely the result of hybridization of populations from multiple introductions (Novak and Mack 1993; Ellstrand and Schierenbeck 2000).
Invasive species with the ability to hybridize may benefit from evolutionary novelty and/or increased genetic variation. These properties may provide genetic material for rapid adaptation to abiotic and biotic conditions (Blair and Hufbauer 2010). Typha × glauca (Typhaceae) is a hybrid between T. latifolia (native) and T. angustifolia and is aggressive in disturbed wetlands in the eastern United States, such in the Great Lakes wetlands, especially when watersheds are urbanized (Zedler and Kercher 2004; Frieswyk and Zedler 2007). This species is sterile, but offspring spread aggressively through rhizomes that crowd out native species (Zedler and Kercher 2004). As mentioned previously, knotweed species (Fallopia spp.) hybridize in the United States but do not appear to have hybridized in their invasive range in Europe (Grimsby et al. 2007).
Hybridization among knapweed species (Centaurea spp.) makes these plants some of the most economically and ecologically detrimental introduced plants in western North America (Watson and Renney 1974; Blair and Hufbauer 2010). These species are capable of hybridizing with other knapweeds: Centaurea × psammogena is a hybrid between two non-native plants, spotted knapweed (C. stoebe) and diffuse knapweed (C. diffusa). Debate around Centaurea × psammogena has suggested that the plants are diffuse knapweed, expressing variable genotypes as a result of loose gene control (Watson and Renney 1974; Blair and Hufbauer 2010). More sophisticated genetic techniques have now made it possible to explore whether this hybridization in fact occurred. Blair and Hufbauer (2010) explored the hybridization of these two knapweeds at the molecular level and found hybridization had occurred in some individuals, but plants with intermediate morphology did not show evidence of mixed ancestry more often than plants with typical morphology of diffuse knapweed. Thus, in North America, sites that have both diffuse and spotted knapweed will likely include hybrid swarms (Blair and Hufbauer 2010).
8.4.3 Impacts on Soil Properties and Nutrient Cycling
Plants also modify the physical and chemical nature of the soil, altering biogeochemical processes and soil structure (Weidenhamer and Callaway 2010; Fei et al. 2014). Invasive plants primarily impact geomorphic properties of an ecosystem through bioprotection or bioconstruction (Fei et al. 2014). Bioprotection effects are properties that reduce or inhibit erosion or weathering, whereas bioconstruction refers to sedimentary accretion caused or facilitated by invasive species or through formation of organically dominated surface layers (e.g., leaf litter; Fei et al. 2014). Many invasive plants were originally introduced to novel ecosystems because they have geoprotective properties. A classic example in North America is the introduction of a vine native to Asia, kudzu (Pueraria montana), to the southeastern United States for erosion control. Pueraria montana forms a dense protective cover trapping sediments and infilling eroded gullies; however, since its introduction kudzu has overwhelmed the landscape in some areas, engulfing fields, trees, poles, and abandoned dwellings (Winberry and Jones 1973; Fei et al. 2014).
Similarly, European beachgrass (Ammophila arenaria) was introduced into San Francisco’s Golden Gate Park from northern Europe around 1869 with the intention of stabilizing sand dunes (Lamb 1898; Schroeder et al. 1977; Dukes and Mooney 2004). Ammophila arenaria has since colonized dunes along much of the US Pacific coast (Dukes and Mooney 2004). This species is thought to have become invasive for several reasons: (1) multiple introductions and introduction sites, via widespread planting of A. arenaria for 100 years after introduction (Wiedemann and Pickart 1996); (2) rapid within-site spread, through lateral growth of rhizomes; and (3) effective dispersal, as living rhizome fragments can wash down shore and colonize new locations (Wallén 1980). Ammophila arenaria collects sand more effectively than the previously dominant native dune grass Leymus mollis (Dukes and Mooney 2004; Barbour et al. 2007). This results in the development of steep, continuous foredunes, as high as 10 m, along the coast (Wiedemann and Pickart 1996). These large foredunes may inhibit active inland dune systems from collecting sand, resulting in more static systems that do not allow for establishment of native plants (Dukes and Mooney 2004).
In addition to bioprotective properties, non-native plants also impact geomorphology through bioconstruction by altering sediment deposition or litter accumulation rates (Dukes and Mooney 2004; Fei et al. 2014). In riparian ecosystems, salt cedar (Tamarix spp.) encroachment upon formerly unoccupied sandbanks along riverbanks has resulted in stabilization of sediments and slowed water velocity (Blackburn et al. 1982). As water movement is reduced, sediment deposition increases further and river channels narrow, increasing the flooding frequency of rivers (Blackburn et al. 1982). Like Tamarix, other freshwater invasive plants have been shown to increase sedimentation. Floating plants such as water hyacinth (Eichhornia crassipes) increase sedimentation through their complex root structure (Gopal 1987), and emergent plants such as papa grass (Urochloa mutica) and submerged plants such as water thyme (Hydrilla verticillata) increase sediment accumulation rates by reducing flow velocity and litter accumulation (Langeland 1996; Bunn et al. 1998). Other invasive plants can accelerate erosion (Dukes and Mooney 2004; Fei et al. 2014). For example, spotted knapweed (Centaurea maculosa) displaced native bunchgrasses throughout many rangelands of western North America (Tyser and Key 1988; Lindquist et al. 1996). The presence of C. maculosa results in greater losses of sediment and greater runoff from areas dominated by this species than bunchgrass communities (Lacey et al. 1989).
Through these types of alterations to geomorphic properties, invasive plants can alter soil chemical and physical properties, influencing nutrient availability. Invasive species frequently have higher specific leaf area, growth rate, and leaf nutrient concentration compared to their native counterparts; these traits have the potential to accelerate decomposition and nutrient cycling (Allison and Vitousek 2004; Liao et al. 2008; Weidenhamer and Callaway 2010). For example, in deciduous forests in North America, areas invaded by garlic mustard (Alliaria petiolata), a biennial forb from Europe, have soils with significantly higher nutrient availability and soil pH (Reinhart and Callaway 2006; Rodgers et al. 2008). This may be the result of the significantly higher rates of leaf decomposition of this species compared to native trees (Reinhart and Callaway 2006; Rodgers et al. 2008). Higher nutrient contents in soils have also been found in western North America with the invasion of cheatgrass (B. tectorum; (Blank 2008) and the succulent, hottentot-fig (Carpobrotus edulis Renz and Blank 2004; Reinhart and Callaway 2006).
The examples above demonstrate the interwoven relationships of plants and soil biota known as plant-soil feedbacks (Reinhart and Callaway 2006). These feedbacks are considered positive when plant species increase beneficial soil biota, including mycorrhizal fungi, nitrogen-fixing bacteria, and other soil organisms that increase plant growth. Plant-soil feedbacks are considered negative when plants boost the abundance of soil-borne pathogens, herbivores, and parasites (Reinhart and Callaway 2006). In some invaded ecosystems, plant invaders promote soil biota that may even promote the establishment and invasion of non-native species, making plant-soil feedback processes an important component of invasion (Callaway et al. 2004). Rare species in North America consistently demonstrate negative feedbacks with soil microbes that promote biological diversity; conversely, abundant non-native and native species tend to produce positive feedbacks that can reduce biodiversity. Positive feedbacks occur when a plant species accumulates microbes near their roots that have beneficial effects on the plants that harbor them, such as mycorrhizal fungi or bacterial nitrogen fixers, which can lead to a reduction in biodiversity. Negative feedbacks occur when plant species accumulate pathogenic microbes in their rhizospheres that create increasingly hostile conditions to the plants that cultivate these pathogens, resulting in increased species turnover rates and, therefore, greater diversity. For example, soil microbes obtained from European soils in the native range of spotted knapweed (C. maculosa) were found to have stronger inhibitory effects on that plant’s growth than soil microbes in its non-native range within North America. In soils from North America, C. maculosa cultivated soil biota that had increasingly positive feedbacks on its growth, which may contribute to the success of this species on this continent (Callaway et al. 2004).
8.5 Policy and Legislation for Invasive Plants
North America has accumulated more invasive plant and animal species than any other continent (Fig. 8.5), and non-native species numbers are projected to continue to increase over the coming decades (Hulme 2009; Seebens et al. 2017, 2021). National legislation and international agreements have been developed in the last 100 years in attempts to reduce threats from non-native species to biodiversity, the economy, and human well-being (McGeoch et al. 2010; Seebens et al. 2017). Without legislation and international agreements, the number and impact of invasive species would probably be much greater (Seebens et al. 2017). Countries in North America are addressing non-native species in a variety of ways (Table 8.2), and there is a patchwork of additional legislation at state and local levels (Pyke et al. 2008; McGeoch et al. 2010). Much of this legislation focuses on prevention measures, as these are more cost-efficient and effective than combatting invasive species after their establishment and bearing the environmental and economic costs of invasion. Prevention measures are diverse, ranging from prohibitions on import, possession, or release of certain species to education to prevent accidental introduction of a species by the public (Pyke et al. 2008). Rapid response to initial sightings of a non-native plant species can be important for preventing subsequent invasion. For example, Caulerpa taxifolia, an invasive marine algae, was discovered in California in June 2000. Because C. taxifolia had a well-known 15-year history of spread in the Mediterranean Sea, this species was already on the US Federal Noxious Weed List in 1999. This awareness facilitated mobilization of a large number of state, federal, and local agencies, as well as private groups and non-governmental organizations (Anderson 2005), and the two populations in California were considered eradicated in June 2006 (NOAA 2019).

Predicted developments of alien, vascular plant species numbers on different continents until 2050. The dots represent means of up to 100 model runs. The shaded area represents the full range of predicted trajectories. Excerpted from Seebens et al. (2021)
In this era of globalization, it is projected that naturalization of non-native species will continue in North America. At the same time, as greenhouse gas emission rates continue to rise, climate change is also increasingly affecting North America’s ecosystems and the economy. Invasive plants and climate change are often treated as important, but independent, environmental issues. To date, there has been limited investment into forecasting how global climate change could shape future invasions, or how invasive species may affect the magnitude, rate, and impact of climate change (Pyke et al. 2008; Ricciardi et al. 2017). Thus, as new policies are developed in North America for invasive species, it will be beneficial to consider the implications of climate change and to develop policies that consider the interactions between invasive species and climate change that may combine to increase invasion risk to native ecosystems (Pyke et al. 2008; Bradley et al. 2009; Ricciardi et al. 2017). A changing climate creates risk as the climatic range for a given invasive species may expand into some new areas, but may also create opportunities for ecosystem restoration on invaded lands that have become climatically unsuitable for the invasive species (Bradley et al. 2009). Bradley et al. (2009) found that climate change can result in the potential for both range expansion and contraction of invasive plants in the western United States using habitat suitability modeling. Their analysis suggested Centaurea solstitialis and Tamarix spp. ranges are likely to expand with climate change, whereas Bromus tectorum and Centaurea stoebe are likely to shift in range, leading to both expansion and contraction (Bradley et al. 2009).
8.6 Conclusions
North America has more recorded naturalized plants than any other continent, and this number is expected to rise in the future as a result of human-related activities like transportation, agriculture, and climate change (van Kleunen et al. 2015; Pyšek et al. 2017; Ricciardi et al. 2017). The naturalization of these non-native species is often attributed to plant life history traits, and in North America, habitat legacy, propagule pressure, and residence time have been found to be important driving forces for non-native species to become invasive (Pyšek et al. 2015). These species are often distributed disproportionately across the continent, with the majority of non-native species being found in the colder temperate and Mediterranean climate zones (Pyšek et al. 2017). Invasive non-native plants threaten ecosystems, habitats, and native species and are key drivers of human environmental change in North America. Challenges associated with plant invasions are likely to grow, as international trade continues and environmental changes influence the rate and consequences of future invasions (Pyke et al. 2008; Ricciardi et al. 2017).
References
Aiken SG, Newroth PR, Wile I (1979) The biology of Canadian weeds. 34. Myriophyllum spicatum L. Can J Plant Sci 59:201–215
Allison SD, Vitousek PM (2004) Rapid nutrient cycling in leaf litter from invasive plants in Hawai’i. Oecologia 141:612–619. https://doi.org/10.1007/s00442-004-1679-z
Anderson LWJ (2005) California’s reaction to Caulerpa taxifolia: a model for invasive species rapid response. Biol Invasions 7:1003–1016. https://doi.org/10.1007/s10530-004-3123-z
Asbjornsen H, Mora G, Helmers MJ (2007) Variation in water uptake dynamics among contrasting agricultural and native plant communities in the Midwestern U.S. Agric Ecosyst Environ 121:343–356. https://doi.org/10.1016/j.agee.2006.11.009
Bailey JP (1994) Reproductive biology and fertility of Fallopia japonica (Japanese knotweed) and its hybrids in the British Isles. In: DeWaal LC, Child LE, Wade MP, Brock JH (eds) Ecology and management of invasive riverside plants. Wiley, Chichester, pp 141–158
Balch JK, Bradley BA, D’Antonio CM, Gómez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980-2009). Glob Chang Biol 19:173–183. https://doi.org/10.1111/gcb.12046
Barbour M, Keeler-Wolf T, Schoenherr AA (eds) (2007) Terrestrial vegetation of California. University of California Press, Berkeley
Barney JN (2014) Bioenergy and invasive plants: quantifying and mitigating future risks. Invasive Plant Sci Manag 7:199–209. https://doi.org/10.1614/ipsm-d-13-00060.1
Bellard C, Leroy B, Thuiller W et al (2016) Major drivers of invasion risks throughout the world. Ecosphere 7:e01241. https://doi.org/10.1002/ecs2.1241
Blackburn WH, Knight RW, Schuster JL (1982) Saltcedar influence on sedimentation in the Brazos River. J Soil Water Conserv 37:298–301
Blair AC, Hufbauer RA (2010) Hybridization and invasion: one of North America’s most devastating invasive plants shows evidence for a history of interspecific hybridization. Evol Appl 3:40–51. https://doi.org/10.1111/j.1752-4571.2009.00097.x
Blank RR (2008) Biogeochemistry of plant invasion: a case study with downy brome (Bromus tectorum). Invasive Plant Sci Manag 1:226–238. https://doi.org/10.1614/ipsm-07-026.1
Bradley BA, Mustard JF (2006) Characterizing the landscape dynamics of an invasive plant and risk of invasion using remote sensing. Ecol Appl 16:1132–1147. https://doi.org/10.1890/1051-0761(2006)016[1132:CTLDOA]2.0.CO;2
Bradley BA, Oppenheimer M, Wilcove DS (2009) Climate change and plant invasions: restoration opportunities ahead? Glob Chang Biol 15:1511–1521. https://doi.org/10.1111/j.1365-2486.2008.01824.x
Bradley BA, Blumenthal DM, Wilcove DS, Ziska LH (2010) Predicting plant invasions in an era of global change. Trends Ecol Evol 25:310–318. https://doi.org/10.1016/j.tree.2009.12.003
Bradley BA, Curtis CA, Fusco EJ et al (2018) Cheatgrass (Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions. Biol Invasions 20:1493–1506. https://doi.org/10.1007/s10530-017-1641-8
Brauman KA, Daily GC, Duarte TK et al (2007) The nature and value of ecosystem services: an overview highlighting hydrologic services. Annu Rev Environ Resour 32:67–98. https://doi.org/10.1146/annurev.energy.32.031306.102758
Brooks M, D’Antonelo C, Richardson DM et al (2006) Effects of invasive alien plants on fire regimes. BioScience 54:677. https://doi.org/10.1641/0006-3568(2004)054[0677:eoiapo]2.0.co;2
Bunn SE, Davies PM, Kellaway DM et al (1998) Influence of invasive macrophytes on channel morphology and hydrology in an open tropical lowland stream, and potential control by riparian shading. Freshw Biol 39:171–178. https://doi.org/10.1046/j.1365-2427.1998.00264.x
Calder I, Dye P (2001) Hydrological impacts of invasive alien plants. Land Use Water Resour Res 7:1–12
Callaway R, Thelen G, Rodriguez A et al (2004) Soil biota and exotic plant invasion. Nature 427:731
Campbell MH, Delfosse ES (1984) Biology of Australian weeds. 13. Hypericum perforatum L. J Aust Inst Agric Sci 50:63–73
Cavaleri MA, Ostertag R, Cordell S, Sack L (2014) Native trees show conservative water use relative to invasive trees: results from a removal experiment in a Hawaiian wet forest. Conserv Physiol 2:1–14. https://doi.org/10.1093/conphys/cou016
Charles H, Dukes JS (2007) Impacts of invasive species on ecosystem services. In: Nentwig W (ed) Biological invasions. Springer, Berlin/Heidelberg, pp 217–237
Cleverly JR, Dahm CN, Thibault JR et al (2006) Riparian ecohydrology: regulation of water flux from the ground to the atmosphere in the Middle Rio Grande, New Mexico James. Hydrol Process 30:3207–3225. https://doi.org/10.1002/hyp
D’Antonio CM, Odion DC, Tyler CM (1993) Invasion of maritime chaparral by the introduced succulent Carpobrotus edulis – the roles of fire and herbivory. Oecologia 95:14–21. https://doi.org/10.1007/BF00649501
Della Venezia L, Samson J, Leung B (2018) The rich get richer: invasion risk across North America from the aquarium pathway under climate change. Divers Distrib 24:285–296. https://doi.org/10.1111/ddi.12681
Doody TM, Nagler PL, Glenn EP et al (2011) Potential for water salvage by removal of non-native woody vegetation from dryland river systems. Hydrol Process 25:4117–4131. https://doi.org/10.1002/hyp.8395
Dukes J, Mooney H (2004) Disruption of ecosystem processes in western North America by invasive species. Rev Chil Hist Nat 77:411–437
Dye P, Jarmain C (2004) Water use by black wattle (Acacia mearnsii): Implications for the link between removal of invading trees and catchment streamflow response. S Afr J Sci 100:40–44
Dyer AR, Rice KJ (1999) Effects of competition on resource availability and growth of a California bunchgrass. Ecology 80:2697–2710. https://doi.org/10.1890/0012-9658(1999)080[2697:EOCORA]2.0.CO;2
Eagle AJ, Eiswerth ME, Johnson WS et al (2007) Costs and losses imposed on California ranchers by yellow starthistle. Rangel Ecol Manag 60:369–377
Early R, Bradley BA, Dukes JS et al (2016) Global threats from invasive alien species in the twenty-first century and national response capacities. Nat Commun 7:1–9. https://doi.org/10.1038/ncomms12485
Ellstrand NC, Schierenbeck KA (2000) Hybridization as a stimulus for the evolution of invasiveness in plants? PNAS 97:7043–7050
Fan Z, Moser WK, Hansen MH, Nelson MD (2013) Regional patterns of major nonnative invasive plants and associated factors in upper midwest forests. For Sci 59:38–49. https://doi.org/10.5849/forsci.10-100
Fei S, Phillips J, Shouse M (2014) Biogeomorphic impacts of invasive species. Annu Rev Ecol Evol Syst 45:69–87. https://doi.org/10.1146/annurev-ecolsys-120213-091928
Findlay R, O’Rourke K (2007) Power and plenty: Trade, war and the world economy in the second millennium. Inst Int Integr Stud:1–16
Fisher B, Turner RK, Morling P (2009) Defining and classifying ecosystem services for decision making. Ecol Econ 68:643–653. https://doi.org/10.1016/j.ecolecon.2008.09.014
Foley JA, DeFries R, Asner GP et al (2005) Global consequences of land use. Science 309:570–574
Frieswyk CB, Zedler JB (2007) Vegetation change in Great Lakes coastal wetlands: deviation from the historical cycle. J Great Lakes Res 33:366–380. https://doi.org/10.3394/0380-1330(2007)33[366:vciglc]2.0.co;2
Fusco EJ, Finn JT, Balch JK et al (2019) Invasive grasses increase fire occurrence and frequency across US ecoregions. Proc Natl Acad Sci USA 116:23594–23599. https://doi.org/10.1073/pnas.1908253116
Gelbard JL, Belnap J (2003) Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv Biol 17:420–432. https://doi.org/10.1046/j.1523-1739.2003.01408.x
Gerlach JDJ (2001) A model experimental system for predicting the invasion success and ecosystem impacts of nonindigenous summer-flowering annual plants in California’s Central Valley grasslands and oak woodlands. University of California, Davis
Gopal B (1987) Water hyacinth. Elsevier Science Publishers, New York
Gopal B, Sharma KP (1981) Water-hyacinth (Eichhornia crassipes) the most troublesome weed of the world. Hindasia, New Delhi
Government of Canada (2010) Ballast water defined. In: Gov. Canada. https://www.tc.gc.ca/eng/marinesafety/oep-environment-ballastwater-defined-249.htm
Grimsby JL, Tsirelson D, Gammon MA, Kesseli R (2007) Genetic diversity and clonal vs. sexual reproduction in Fallopia spp. (Polygonaceae). Am J Bot 94:957–964
Hewitt CL, Gollasch S, Minchin D (2009) The vessel as a vector – biofouling, ballast water and sediments. In: Rilov G, Crooks JA (eds) Biological invasions in marine ecosystems. Springer, Berlin/Heidelberg, pp 117–131
Hollingsworth ML, Bailey JP (2000) Evidence for massive clonal growth in the invasive weed Fallopia japonica (Japanese Knotweed). Bot J Linn Soc 133:463–472. https://doi.org/10.1006/bojl.2000.0359
Houghton RA, Nassikas AA (2017) Global and regional fluxes of carbon from land use and land cover change 1850–2015. Global Biogeochem Cycles 31:456–472. https://doi.org/10.1111/1462-2920.13280
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46:10–18
Iannone BV, Oswalt CM, Liebhold AM et al (2015) Region-specific patterns and drivers of macroscale forest plant invasions. Divers Distrib 21:1181–1192. https://doi.org/10.1111/ddi.12354
IPBES (2018) Summary for policymakers of the regional assessment report on biodiversity and ecosystem services for the Americas. IPES Secretariat, Bonn
Ismail AA, Baskin YC, Clout MN et al (1999) Invasive alien species how to address one of the greatest threats to biodiversity: a toolkit of best prevention and management practices. In: Wittenberg R, Cock MJW (eds) Global invasive species programme. Kuala Lumpur, Malaysia
Jacobs J, Margold J (2009) Plant guide for Eurasian watermilfoil (Myriophyllum spicatum L.). Bozeman, MT
Keast A (1984) The introduced aquatic macrophyte, Myriophyllum spicatum, as habitat for fish and their invertebrate prey. Can J Zool 67:1289–1303
Kubešová M, Moravcová L, Suda J et al (2010) Naturalized plants have smaller genomes than their non-invading relatives: a flow cytometric analysis of the Czech alien flora. Preslia 82:81–96
Lacey JR, Marlow CB, Laned JR (1989) Influence of spotted knapweed (Centaurea maculosa) on surface runoff and sediment yield. Weed Technol 3:627–631
Lamb F (1898) Sand dune reclamation on the Pacific coast. The Forester 4:141–142
Langeland KA (1996) Hydrilla verticillata ( L.F.) Royle (Hydrocharitaceae), “the perfect aquatic weed”. South Appalach Bot Soc 61:293–304
Larson DL (2002) Native weeds and exotic plants: relationships to disturbance in mixed-grass prairie. Plant Ecol 169:317–333. https://doi.org/10.1023/A:1026046810307
Lavergne S, Molofsky J (2004) Reed canary grass (Phalaris arundinacea) as a biological model in the study of plant invasions. CRC Crit Rev Plant Sci 23:415–429. https://doi.org/10.1080/07352680490505934
Lavergne S, Molofsky J (2006) Control strategies for the invasive reed canarygrass (Phalaris arundinacea L.) in North American wetlands: the need for an integrated management plan. Nat Areas J 26:208–214. https://doi.org/10.3375/0885-8608(2006)26[208:csftir]2.0.co;2
Lavergne S, Molofsky J (2007) Increased genetic variation and evolutionary potential drive the success of an invasive grass. Proc Natl Acad Sci USA 104:3883–3888. https://doi.org/10.1073/pnas.0607324104
Lavergne S, Mouquet N, Thuiller W, Ronce O (2010a) Biodiversity and climate change: integrating evolutionary and ecological responses of species and communities. Annu Rev Ecol Evol Syst 41:321–350. https://doi.org/10.1146/annurev-ecolsys-102209-144628
Lavergne S, Muenke NJ, Molofsky J (2010b) Genome size reduction can trigger rapid phenotypic evolution in invasive plants. Ann Bot 105:109–116. https://doi.org/10.1093/aob/mcp271
Lehan NE, Murphy JR, Thorburn LP, Bradley BA (2013) Accidental introductions are an important source of invasive plants in the continental United States. Am J Bot 100:1287–1293. https://doi.org/10.3732/ajb.1300061
Levine JM, D’Antonio CMD (2015) Forecasting biological invasions with increasing international trade. Conserv Biol 17:322–326
Levine JM, Vila M, Antonio CMD et al (2003) Mechanisms underlying the impacts of exotic plant invasions. Proc R Soc B Biol Sci 270:775–781. https://doi.org/10.1098/rspb.2003.2327
Liao C, Peng R, Luo Y et al (2008) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytol 177:706–714. https://doi.org/10.1111/j.1469-8137.2007.02290.x
Lindquist JL, Maxwell BD, Weaver T (1996) Potential for controlling the spread of Centaurea maculosa with grass competition. Gt Basin Nat 56:267–271
Mack RN (2003) Plant naturalizations and invasions in the eastern United States: 1634-1860. Ann Missouri Bot Gard 90:77–90
Mack RN, Erneberg M (2002) The United States naturalized flora: largely the product of deliberate introductions. Ann Missouri Bot Gard 89:176–189
McGeoch MA, Butchart SHM, Spear D et al (2010) Global indicators of biological invasion: species numbers, biodiversity impact and policy responses. Divers Distrib 16:95–108. https://doi.org/10.1111/j.1472-4642.2009.00633.x
McNeely JA (2006) As the world gets smaller, the chances of invasion grow. Euphytica 148:5–15. https://doi.org/10.1007/s10681-006-5937-5
Merhoff LJ, Silander JJA, Leicht SA et al (2003) IPANE: invasive plant atlas of New England. Department of Ecology and Evolutionary Biology University of Connecticut. http://www.ipane.org
Meyerson LA, Mooney HA (2007) Invasive alien species in an era of globalization. Front Ecol Environ 5:199–208
Moore GW, Owens MK (2012) Transpirational water loss in invaded and restored semiarid riparian forests. Restor Ecol 20:346–351. https://doi.org/10.1111/j.1526-100X.2011.00774.x
Nelson JC (1917) The introduction of foreign weeds in ballast as illustrated by ballast-plants at Linnton, Oregon. Torreya 17:151–160
NOAA (2019) Caulerpa taxifolia. NOAA Fisheries West Coast Region. https://www.westcoast.fisheries.noaa.gov/habitat/aquatic_invasives/caulerpa_taxifolia.html
Novak SJ, Mack RN (1993) Genetic variation in Bromus tectorum (Poaceae): comparison between native and introduced populations. Heredity (Edinb) 71:167–176. https://doi.org/10.1038/hdy.1993.121
Oduor AMO, Gómez JM, Herrador MB et al (2015) Invasion of Brassica nigra in North America: Distributions and origins of chloroplast DNA haplotypes suggest multiple introductions. Biol Invasions 17:2447–2459. https://doi.org/10.1007/s10530-015-0888-1
Oswalt CM, Fei S, Guo Q et al (2015) A subcontinental view of forest plant invasions. NeoBiota 24:49–54. https://doi.org/10.3897/neobiota.24.8378
Padilla DK, Williams SL (2004) Beyond ballast water: aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front Ecol Environ 2:131–138. https://doi.org/10.1890/1540-9295(2004)002[0131:BBWAAO]2.0.CO;2
Pandit MK, White SM, Pocock MJO (2014) The contrasting effects of genome size, chromosome number and ploidy level on plant invasiveness: a global analysis. New Phytol 203:697–703. https://doi.org/10.1111/nph.12799
Parks CG, Radosevich SR, Endress BA et al (2005) Natural and land-use history of the Northwest mountain ecoregions (USA) in relation to patterns of plant invasions. Perspect Plant Ecol Evol Syst 7:137–158. https://doi.org/10.1016/j.ppees.2005.09.007
Pejchar L, Mooney HA (2009) Invasive species, ecosystem services and human well-being. Trends Ecol Evol 24:497–504. https://doi.org/10.1016/j.tree.2009.03.016
Pennington DN, Hansel JR, Gorchov DL (2010) Urbanization and riparian forest woody communities: diversity, composition, and structure within a metropolitan landscape. Biol Conserv 143:182–194. https://doi.org/10.1016/j.biocon.2009.10.002
Pimentel D (2009) Invasive plants: their role in species extinctions and economic losses to agriculture in the USA. Springer, Dordrecht
Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52:273–288. https://doi.org/10.1016/j.ecolecon.2004.10.002
Popay I (2015) Hypericum perforatum (St John’s wort). CABI Invasive Species Compend. https://www.cabi.org/isc/datasheet/28268
Pyke CR, Thomas R, Porter RD et al (2008) Current practices and future opportunities for policy on climate change and invasive species. Conserv Biol 22:585–592. https://doi.org/10.1111/j.1523-1739.2008.00956.x
Pyšek P, Richardson DM (2008) Traits associated with invasiveness in alien plants: where so we stand? In: Nentwig W (ed) Biological invasions. Ecological Studies (analysis and synthesis), 193rd edn. Springer, Berlin/Heidelberg, pp 97–125. https://doi.org/10.1007/978-3-540-36920-2_7
Pyšek P, Manceur AM, Alba C et al (2015) Naturalization of central European plants in North America: species traits, habitats, propagule pressure, residence time. Ecology 96:762–774. https://doi.org/10.1890/14-1005.1
Pyšek P, Pergl J, Essl F et al (2017) Naturalized alien flora of the world: species diversity, taxonomic and phylogenetic patterns, geographic distribution and global hotspots of plant invasion. 89:203–Preslia, 274. https://doi.org/10.23855/preslia.2017.203
Pyšek P, Skálová H, Čuda J et al (2018) Small genome separates native and invasive populations in an ecologically important cosmopolitan grass. Ecology 99:79–90. https://doi.org/10.1002/ecy.2068
Reichard SH, White P (2001) Horticulture as a pathway of invasive plant introductions in the United States. Bioscience 51:103. https://doi.org/10.1641/0006-3568(2001)051[0103:haapoi]2.0.co;2
Reinhart KO, Callaway RM (2006) Soil biota and invasive plants. New Phytol Tansley Rev 170:445–457. https://doi.org/10.1111/j.1469-8137.2006.01715.x
Rejmanek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1661
Renz MJ, Blank RR (2004) Influence of perennial pepperweed (Lepidium latifolium) biology and plant-soil relationships on management and restoration. Weed Technol 18:1359–1363
Ricciardi A, Blackburn TM, Carlton JT et al (2017) Invasion science: a horizon scan of emerging challenges and opportunities. Trends Ecol Evol 32:464–474
Ricklefs RE, Guo Q, Qian H (2008) Growth form and distribution of introduced plants in their native and non-native ranges in Eastern Asia and North America. Divers Distrib 14:381–386. https://doi.org/10.1111/j.1472-4642.2007.00457.x
Rodgers VL, Stinson KA, Finzi AC (2008) Ready or not, garlic mustard is moving in: Alliaria petiolata as a member of eastern North American forests. Bioscience 58:426–436. https://doi.org/10.1641/B580510
Schroeder PM, Dolan R, Hayden BP (1977) Vegetation changes associated with barrier-dune construction on the outer banks of North Carolina. Environ Manage 1:105–114. https://doi.org/10.1007/BF01866101
Seebens H, Essl F, Dawson W et al (2015) Global trade will accelerate plant invasions in emerging economies under climate change. Glob Chang Biol 21:4128–4140. https://doi.org/10.1111/gcb.13021
Seebens H, Blackburn TM, Dyer EE et al (2017) No saturation in the accumulation of alien species worldwide. Nat Commun 8:1–9. https://doi.org/10.1038/ncomms14435
Seebens H, Bacher S, Blackburn TM et al (2021) Projecting the continental accumulation of alien species through to 2050. Glob Chang Biol 27:970–982. https://doi.org/10.1111/gcb.15333
Tatem AJ (2009) The worldwide airline network and the dispersal of exotic species: 2007-2010. Ecography (Cop) 32:94–102. https://doi.org/10.1111/j.1600-0587.2008.05588.x
Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stage of invasion. New Phytol 176:256–273. https://doi.org/10.1111/j.1469-8137.2007.02207.x/pdf
Turner K, Lefler L, Freedman B (2005) Plant communities of selected urbanized areas of Halifax, Nova Scotia, Canada. Landsc Urban Plan 71:191–206. https://doi.org/10.1016/j.landurbplan.2004.03.003
Tyser RW, Key CH (1988) Spotted knapweed in natural area fescue grasslands: an ecological assessment. Northwest Sci 62:151–160
U.S. Fish & Wildlife Service (2018) Eurasian watermilfoil (Myriophyllum spicatum) ecological risk screening summary
van Kleunen M, Dawson W, Essl F et al (2015) Global exchange and accumulation of non-native plants. Nature 525:100–103. https://doi.org/10.1038/nature14910
van Kleunen M, Essl F, Pergl J et al (2018) The changing role of ornamental horticulture in alien plant invasions. Biol Rev 93:1421–1437. https://doi.org/10.1111/brv.12402
van Kleunen M, Pyšek P, Dawson W et al (2019) The global naturalized alien flora (GloNAF) database. Ecology 100:1–2. https://doi.org/10.1002/ecy.2542
Villarreal ML, Soulard CE, Waller EK (2019) Landsat time series assessment of invasive annual grasses following energy development. Remote Sens 11:1–18
Wallén B (1980) Changes in structure and function of ammophila during primary succession. Oikos 34:227–238
Walter H, Breckle S (1991) Ökologie der Erde. Band 1. Grundlagen. Gustav Fischer, Stuttgart
Watson AK, Renney AJ (1974) The biology of Canadian weeds. Centaurea diffusa and C. maculosa. Can J Plant Sci 54:687–701
Weidenhamer JD, Callaway RM (2010) Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J Chem Ecol 36:59–69. https://doi.org/10.1007/s10886-009-9735-0
Werschkun B, Banerji S, Basurko OC et al (2014) Emerging risks from ballast water treatment: The run-up to the International Ballast Water Management Convention. Chemosphere 112:256–266. https://doi.org/10.1016/j.chemosphere.2014.03.135
Whisenant SG (1990) Changing fire frequencies on Idaho’s Snake River plains: ecological and management implications. In: Proceedings of the symposium cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management, pp 4–10. https://doi.org/10.1016/0006-3207(92)90659-b
Wiedemann AM, Pickart A (1996) The Ammophila problem on the Northwest coast of North America. Landsc Urban Plan 34(3–4):287–299
Winberry JJ, Jones DM (1973) Rise and decline of the “miracle vine”: Kudzu in the Southern Landscape. Southeast Geogr 13:61–70. https://doi.org/10.1353/sgo.1973.0004
Yates ED, Levia DF, Williams CL (2004) Recruitment of three non-native invasive plants into a fragmented forest in southern Illinois. For Ecol Manage 190:119–130. https://doi.org/10.1016/j.foreco.2003.11.008
Zedler JB, Kercher S (2004) Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. CRC Crit Rev Plant Sci 23:431–452. https://doi.org/10.1080/07352680490514673
Zedler PH, Scheid GA (1988) Invasion of Carpobrotus edulis and Salix lasiolepis after fire in a coastal chaparral site in Santa Barbara County, California. Madroño 35:196–201
Zouhar K (2004) Hypericum perforatum. Fire effects information system. https://www.fs.fed.us/database/feis/plants/forb/hypper/all.html
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Ploughe, L.W., Dukes, J.S. (2022). Plant Invasions in North America. In: Clements, D.R., Upadhyaya, M.K., Joshi, S., Shrestha, A. (eds) Global Plant Invasions. Springer, Cham. https://doi.org/10.1007/978-3-030-89684-3_8
Download citation
DOI: https://doi.org/10.1007/978-3-030-89684-3_8
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-89683-6
Online ISBN: 978-3-030-89684-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)