
Abstract
Chemical variation among streams is primarily governed by the type and composition of rocks in the drainage, and by the amount and chemical composition of precipitation. In addition, human activities can significantly influence the chemical composition of rivers and streams, indirectly by changing land use and the chemical composition of precipitation, and directly by the input of agricultural, industrial, and domestic waste. This chapter focuses on the dissolved major ions (Ca2+, Na+, Mg2+, K+, HCO3-, SO42-, Cl-) and gases (O2 and CO2). Exchange with the atmosphere maintains both gases at close to their equilibrium concentrations in solution, although photosynthesis and respiration have noticeable effects in highly productive systems, and pollution in the form of excessive organic waste can deplete O2 levels. Important causes of pollution of fresh waters includes salinization due to salts used to melt ice from roads and faulty irrigation practices, increased acidity from mine drainage and fossil fuel use, legacy contaminants such as PCBs and DDT, and emerging chemicals of concern such as pharmaceuticals and personal care products, and plastic. Where pollutants occur at sufficient concentrations, their presence is detectable in organism tissue and may significantly limit populations.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

We all have an intuitive appreciation that river water contains a variety of dissolved and suspended constituents. Mountain streams appear pure, farm creeks often are muddy with sediments, and drainages in limestone-rich regions are fertile while those containing only granitic rocks usually are less so. Rainwater and rivers throughout the world are polluted by organic and inorganic materials generated through human activities that impact the structure and function of freshwater systems.
Many factors influence the chemical composition of river water, producing variation from place to place. Rain is one source of chemical inputs to rivers, and a stream flowing through a region of relatively insoluble rocks can be chemically similar to rainwater in its composition. However, most streams and rivers contain much more suspended and dissolved material than is typically found in rain. Ultimately, all of the constituents of river water originate from dissolution of the earth’s rocks. Variation in water chemistry is driven by the world’s heterogeneous geology and by the magnitude of chemical inputs from other pathways including, but not limited to, precipitation, volcanic activity, and anthropogenic activities. Chemical composition of stream water also is altered by physical processes, such as evaporation and by chemical and biological interactions within the stream.
The materials transported in river water can be organized into dissolved versus suspended and organic vs. inorganic constituents. They include water, suspended inorganic matter, dissolved major ions (e.g., Ca2+, Na+, Mg2+, K+, HCO3−, SO42−, Cl−), dissolved nutrients (e.g., N, P, Si), suspended and dissolved organic matter, gases (e.g., N2, CO2, O2), and trace metals, both dissolved and suspended (e.g., Pb, Hg, Zn, Cu) (Berner and Berner 2012).
In this chapter, we focus on the major dissolved constituents and the gases in freshwaters. We will also discuss some of the interactions between water chemistry and the aquatic biota, and briefly review some of the chemical contaminants of freshwater systems. Readers wishing more detailed discussions of aquatic chemistry and geochemistry should consult Christensen and Li (2014), Weiner (2012), or other specialized volumes.
4.1 Dissolved Gases
Oxygen, carbon dioxide, and nitrogen occur in significant amounts as dissolved gases in river water. Although nitrogen gas (N2) can be incorporated into nitrogen cycling within stream ecosystems through specialized bacteria, the concentration of dissolved N2 in water is typically of little biological importance.
Both oxygen and carbon dioxide gas occur in the atmosphere and dissolve into water according to partial pressure and temperature (Table 4.1). The solubility of oxygen in freshwater is reduced at high elevations due to lower atmospheric partial pressure. Oxygen solubility also declines with increasing salinity. Air is nearly 21% O2 and just 0.03% CO2 by volume, but the latter is much more soluble in water. Hence, although saturated freshwater has higher concentrations of O2 than CO2, the difference is not as great in water as it is found in air. Groundwater frequently is very low in dissolved O2 and enriched in CO2 due to the microbial processing of organic matter as water passes through soil. Thus, streams receiving substantial groundwater inputs may have localized regions of high CO2 concentrations in the water column, but equilibration with the atmosphere usually occurs once hyporheic water enters the stream.
Lotic systems can experience spatial and temporal shifts in dissolved gas concentrations due to weather and natural changes in photosynthesis and respiration. In highly productive waters, whole-system photosynthesis results in elevated concentrations of oxygen during the day, while whole-system respiration causes oxygen to decline at night. These diel (24 h) changes in oxygen concentration provide a means of estimating photosynthesis and respiration of the total ecosystem, further discussed in Chap. 14. First applied to productive, slow-moving rivers and lentic waters where diffusion is relatively low and more easily estimated, recent improvements in measuring diffusion rates and detecting small changes in oxygen concentrations are extending this approach even to small woodland streams (Young and Huryn 1998). However, especially in small, turbulent streams receiving limited amounts of pollution, diffusion maintains O2 and CO2 near saturation. Should biological or chemical processes create a demand for or generate an excess of either gas within the water column, exchange with the atmosphere usually maintains concentrations very near to equilibrium. Gas concentrations often fluctuate seasonally, and longitudinally as water moves downstream. For example, water leaving Lake Constance—the headwaters of the Rhine River—is depleted in CO2 in summer months relative to atmospheric partial pressure due to the productivity of lake phytoplankton. However, inputs of organic pollution increase downstream, stimulating algal and microbial growth. High summer temperatures yield high respiration rates, producing downstream average pCO2 values about twenty times greater than atmospheric pCO2 (Kempe et al. 1991).
Supersaturation of CO2 in rivers is not uncommon. For example, of the ~6700 streams evaluated by Raymond et al. (2013), the average pCO2 was found to be almost eight times greater than atmospheric concentrations. Some of this carbon is lost from rivers and streams via evasion, or degassing. Recent modelling efforts suggest that the amount of carbon lost in CO2 evasion from streams and rivers is comparable to the amount of carbon exported from rivers into the ocean (Lauerwald et al. 2015; Magin et al. 2017). However, the source of the CO2 degassed from flowing waters seems to be related to the size of the system in question. Hotchkiss et al. (2015) suggest that CO2 emissions from smaller streams are dominated by terrestrially-derived CO2 entering streams from the surrounding landscape, whereas much of CO2 evasion from larger systems is primarily derived from internally generated CO2 produced through aquatic respiration. These, and other studies, indicate flowing waters may play an important but often underestimated role in global carbon cycling.
Though pCO2 in freshwaters can be exceptionally high and variable, changes in natural patterns of pCO2 in inland waters are expected to coincide with increasing concentrations of atmospheric CO2. Nonetheless, it is still largely unclear how increases in anthropogenically-derived CO2 emissions will influence the aquatic chemistry and ecology of freshwater systems (Hasler et al. 2015). Research suggests that the quality and quantity of both allochthonous (e.g., leaf litter) and autochthonous (e.g., algae) food resources will be impacted by increasing atmospheric CO2. For example, the coupled effects of increasing temperature and CO2 concentrations are expected to preferentially facilitate the growth of cyanobacteria over eukaryotic algae (Visser et al. 2016). Changes in the behavior and development of aquatic organisms are also expected if they experience stress derived from exposure to increasing pCO2 in inland waters and/or increases in acidity associated with increasing pCO2 (Hasler et al. 2015).
Animals and plants can be responsible for ecologically important shifts in dissolved gas concentrations in rivers. A fascinating example from the Kenyan portion of the Mara River demonstrates at least one way in which animals can influence oxygen dynamics in rivers (Dutton et al. 2018). Hippopotami are nocturnally-feeding animals that congregate in the Mara during daylight hours. During the dry season, excreting and egesting hippopotami become more densely packed in smaller and smaller volumes of water, producing a dense layer of organic-rich feces in the benthic regions of pools. In some cases, hippopotamus activity in pools can disturb this layer, aerating and mixing the water column. However, without enough aeration, anoxia can develop through the decomposition of organic matter (Fig. 4.1). When the rainy season begins, river discharge increases and flushes hippo pools; fish kills are not uncommon as volumes of anoxic water move downstream (Dutton et al. 2018).

(Reproduced from Dutton et al. 2018)
Observations of hypoxic events in hippo pools in the Mara River in Kenya. (a) Robotic boat surveying a hippo pool (image credit: Amanda Subalusky). (b) Dissolved oxygen (grey) and (c) discharge (grey) for a 3-month subset of data between November 2014 through January 2015. Black indicates discharge that was >2 times the calculated baseflow. (d) 3-D interpolation of the conductivity in pools containing or lacking high densities of hippopotami
Eutrophication occurs when a body of water is overly enriched with nutrients, such as nitrogen or phosphorus, which stimulate algal and plant growth and influence primary productivity and respiration. Though it is not as commonly studied as it is in lakes and reservoirs, eutrophication of rivers and streams is of global conservation concern (Dodds and Smith 2016; Smith 2016). When instream conditions facilitate the accumulation of algal biomass, harmful algal blooms (HABs) can occur (Downing 2014). Populations of eukaryotic algae or cyanobacteria can create HABs and may produce large shifts in dissolved gasses, via primary production and community respiration, which can influence other aquatic organisms and biogeochemical processes. Human-induced environmental change, including increasing atmospheric temperatures and CO2 concentrations, are expected to support increasing frequency of HABs (Reid et al. 2018).
Organic pollutants have a long and notorious history of altering oxygen and carbon dioxide concentrations in rivers and streams. As early as the 1600s, the River Thames rose to malodorous infamy, as human and animal waste discharged into the river were linked to “foulness” (Gameson and Wheeler 1977). In fact, 1858 was known as the “Year of the Great Stink” due to the ill-effects of organic pollution in the Thames. Wastewater treatment and more stringent environmental regulations have reduced sources of organic pollution in the Thames and in rivers found in richer economies throughout the globe. Data from the Delaware River in the eastern US demonstrate how enhancing wastewater treatment can support the recovery of heavily impacted systems. In 1950, ~280 municipal wastewater systems serving ~3.4 million people were discharging waste into the Delaware, and only half of the discharge was treated. Low instream dissolved oxygen (DO) concentrations reflected the negative impact that large volumes of organic waste were having in the system. By 1960, all wastewater was being treated prior to discharge in the river, and subsequent improvements in infrastructure in the 1970s and 80s were correlated with significant improvements in instream DO concentrations (Fig. 4.2, Marino et al. 1991). It is important to mention that organic pollution from human waste and other sources still remains an immense conservation challenge for resource managers throughout the world (Wen et al. 2017); this is especially true in poorer nations (Capps et al. 2016). As of 2017, the UN estimated more than 80% of global human wastewater is discharged into surface water without any treatment (Connor et al. 2017).

(Reproduced from Marino et al. 1991)
Historic dissolved oxygen profiles for the Delaware Estuary during summer months
4.2 Major Dissolved Constituents of River Water
Salinity is a term that refers to the sum of the concentrations of all of the dissolved ions in water. However, four major cations, sodium (Na+), potassium (K+), calcium (Ca2+), magnesium (Mg2+), and four major anions, bicarbonate (HCO3−), carbonate (CO32−), sulfate (SO42−), and chloride (Cl−), are the dominant ions in fresh waters. Other ions, including those of nitrogen (N), phosphorus (P), and iron (Fe), are biologically important but contribute relatively little to total ionic concentrations. A similar metric used to describe the concentration of dissolved material is total dissolved solids (TDS), or sum of the concentrations of the aforementioned major ions plus organic matter. In relatively unpolluted waters, TDS is often very similar to salinity. However, in systems influenced by wastewater and other types of organic pollution, TDS can far exceed salinity measurements (Thompson et al. 2006).
The world average of TDS in rivers is about 100 mg L−1 (Table 4.2). Variation in the concentrations of individual constituents and of TDS among river systems results from regional and local variation in natural and anthropogenic inputs of ions within a watershed. However, the vast majority of the world’s rivers are dominated by HCO3− (>50% of the TDS), Cl−, and SO42− (Cl− + SO42− ~10–30% of the TDS), reflecting the important contribution of the weathering of sedimentary rocks to riverine chemistry. This pattern is unsurprising, as three-quarters of the earth’s land surface is covered by sedimentary rocks, which are rich in carbonate minerals and contribute the majority of total dissolved solids to rivers (Berner and Berner 2012).
Precipitation is the ultimate source of water flowing through rivers and streams; yet, river water typically has greater ionic concentrations than rainwater (Berner and Berner 2012). The ionic concentration of rainwater (Table 4.3) is typically low, with average concentrations of a few milligrams per liter. However, these values are highly variable, and can be strongly influenced by both natural and anthropogenic sources. The ions Na+, K+, Ca2+, Mg2+, and Cl− are derived primarily from particles in the air, whereas SO42−, ammonium (NH4+), and nitrate (NO3−) are derived mainly from atmospheric gases. Marine salts (NaCl) are especially important near the oceans, and a transition to calcium sulfate (CaSO4)- or calcium bicarbonate (Ca(HCO3)2)-dominated rain occurs as one proceeds inland (Berner and Berner 2012). Roughly 10–15% of the Na+, Ca2+, and Cl− in US river water comes from rain, compared to one-fourth of the K+, and almost half of the SO42−. In contrast, almost none of the SiO2 or HCO3− comes from rain. The relative importance of chemical inputs to rivers from precipitation can vary seasonally and over short distances, as Sutcliffe and Carrick (1983) document for streams of the English Lake District. These data emphasize the need to examine the origins of each of these major cations and anions in order to understand what influences their concentrations in rivers and streams.
Calcium is the most abundant cation in the world’s rivers. It originates almost entirely from the weathering of sedimentary carbonate rocks, although pollution and atmospheric inputs constitute small sources of the element. Along with magnesium, the concentration of calcium is used to characterize “soft” versus “hard” waters, which are discussed more fully below. Magnesium also originates almost entirely from the weathering of rocks, particularly from magnesium-silicate minerals and dolomite. Atmospheric inputs are minimal, and pollution contributes only slightly to the concentration of magnesium in fresh water.
Sodium is generally found in association with chloride, reflecting their common origin. Weathering of NaCl-containing rocks accounts for most of the sodium and chloride found in river water. However, rainwater inputs from sea salts can contribute significantly to total sodium and chloride concentrations, especially near coasts. Berner and Berner (2012) estimate that worldwide, approximately 28% of the sodium and 30% of the chloride in rivers is derived from pollution. Though chloride is chemically and biologically unreactive, and often is used as a conservative tracer in experiments examining nutrient dynamics in streams, the impact of anthropogenically-derived sodium on aquatic communities can be profound. We will discuss some of the sources of anthropogenically-derived sodium and its impact on aquatic communities later in the chapter.
Potassium (K+) is the least abundant of the major cations in river water, and its concentration is the least variable among systems. Roughly 90% of riverine potassium originates from the weathering of silicate materials, especially potassium feldspar and mica. Concentrations of potassium in river water vary with underlying geology, and tend to increase substantially when moving from polar latitudes toward the tropics. This pattern is apparently due to more complete chemical weathering at higher temperatures. Biological activity can also influence patterns in silica concentrations. Silica (also called silicon dioxide; SiO2) is derived from the weathering of silica-rich minerals that are dominant parts of the earth’s crust. Silica is used by diatoms in the formation of their external cell wall and can on occasion limit algal productivity.
Riverine bicarbonate (HCO3−) is derived almost entirely from the weathering of carbonate minerals. However, the immediate source of the majority of bicarbonate in river water is CO2 produced by bacterial decomposition of organic matter that is dissolved in soil and groundwater. Bicarbonate is a biologically important anion. High concentrations of bicarbonate are reflected in measures of alkalinity and are indicative of fertile waters. The carbonate buffer system, alkalinity, and hardness are interrelated and will be discussed more fully below. Anthropogenic increases in acidity, caused by acid precipitation or mining, reduce bicarbonate levels through the formation of carbonic acid (H2CO3).
Sulphate has many sources, including the weathering of sedimentary rocks and pollution from fertilizers, organic waste, mining activities, and the burning of fossil fuels. Biogenically-derived sulphate in rain and volcanic activity are additional inputs. In areas of sulphuric acid rain, sulphate concentrations are high relative to overall ionic concentrations. In most river water, sulphate and bicarbonate concentrations are inversely correlated, especially in low alkalinity areas.
The concentration of hydrogen ions (H+) is very important both chemically and biologically, because it determines the acidity of water. This is expressed as pH, on a logarithmic scale in which a tenfold change in H+ activity corresponds to a change of 1 pH unit. A pH of 7 is neutral, higher values are alkaline, and lower values are acidic. CO2 concentration and pH are interdependent. CO2 readily dissolves in water to form carbonic acid (H2CO3). Carbonic acid dissociates first to a bicarbonate (HCO3−) ion and H+, and then to a carbonate (CO32−) ion and H+; hence, changes in CO2 concentrations can produce significant shifts in the acidity of stream water.
We will address two key nutrients, nitrogen and phosphorus, more fully in Chap. 13, including their forms (e.g., nitrate, ammonium, phosphate, etc.) and their influence on ecosystem processes. Dissolved inorganic phosphorus and nitrogen are primary nutrients that limit plant and microbial production, and cycle rapidly between their inorganic forms and their incorporation into the food web.
The concentration of major ions can be measured in many ways. For example, TDS is measured by evaporating an aliquot of filtered stream water and weighing the remaining residue. Additionally, colorimetric methods (specific ions react with specific chemicals to form colored compounds), ion chromatography, and ion-specific probes can be used to measure ionic concentrations. Dissolved constituents are reported as units of mass, mg L−1 (equal to parts per million, ppm), or as chemical equivalents. In the latter case, milliequivalents per liter are calculated from mg L−1, by dividing the concentration by the equivalent weight of the ion (its ionic weight divided by its ionic charge).
Conductivity is a measure of electrical conductance of water, and is an approximate measure of total dissolved ions. Distilled water has a very high resistance to electron flow, and the presence of ions in water reduces that resistance. Differences in conductivity result mainly from the concentration of the charged ions in solution, and to a lesser degree from ionic composition and temperature. Conductivity measurements by probes can compensate for temperature (20 or 25 °C) and values are reported as μS ∙ cm−1 (microSiemens per centimeter) or in the older literature as μmho ∙ cm−1 (the reciprocal of ohms). Total dissolved solids can also be estimated from specific conductance (SC). The relationship between TDS and SC is typically linear (TDS = k * SC) with a value of k between 0.55 and 0.75. However, the value of the constant varies with location and must be established empirically (Walling 1984). In fresh water, salinity is usually estimated from conductivity and temperature by using salinity sensors and values are reported as ppt (parts per thousand).
4.2.1 Variability in Ionic Concentrations
The chemistry of fresh waters is quite variable, rivers usually more so than lakes. Natural spatial variation is determined mainly by the type of rocks available for weathering, the climate, and by the chemical composition of rain, which in turn is influenced by proximity to the sea and by anthropogenic activities. The ionic concentration of rivers draining igneous and metamorphic terrains is roughly half that of rivers draining sedimentary terrain, because of the differential resistance of rocks to weathering. All of these factors provide the opportunity for substantial local variation in river chemistry. As a consequence, the concentration of total dissolved ions can vary considerably amongst the headwater branches of a large drainage. However, these heterogeneities tend to average out and ionic concentration tends to increase as one proceeds downstream (Livingstone 1963).
Although small streams in the same region often are chemically similar, they can also differ markedly. In one of the first such studies, Walling and Webb (1975) reported a concentration range of total ions from 25 to 650 mg L−1 among a series of small streams in southwest England, resulting from small-scale shifts between igneous and sedimentary geology and variation in land use. Research from around the globe has continued to document similar chemical variation at relatively small spatial scales. Small streams draining a volcanic landscape in central Costa Rica exhibited pronounced differences in solute concentrations depending on geology, soil types, and elevation. Concentrations of phosphorus, several major cations and anions, and trace elements were higher in headwater streams draining younger volcanic landscapes and were much lower in streams draining older lava flows (Pringle and Triska 1991). Another intriguing example of small-scale spatial variation in stream chemistry comes from the Taylor Valley in Antarctica, where streams fed by glacial meltwater that are strongly influenced by chemical weathering exhibit a range of chemical compositions and TDS that are similar to ranges found in larger temperate and tropical systems (Welch et al. 2010).
Drastic differences in ion concentrations can also be seen at the confluence of rivers with disparate water chemistry. The blackwater Río Negro and whitewater Solimões that converge to form the Amazon River dramatically illustrate the chemical differences between tributaries draining distinctive landscapes. The Río Negro drains well-weathered, crystalline rock and is much lower in ions and much higher in organic acids, whereas the Amazon mainstem drains the comparatively young Andes and has a much higher dissolved load. At their confluence near Manaus, Brazil, known as “the meeting of waters”, the tea-colored waters of the Rio Negro run parallel to the sediment-laden café-au-lait waters of the Solimões without mixing for about 6 km, because of differences in temperature and water density. The unique chemical signatures of both mighty rivers can be detected as much as 100 km below their confluence.
Within-site, temporal variation in water chemistry is also common in stream and rivers due to the influence of fluctuating discharge, precipitation, and biological activity. Discharge has especially strong effects on ionic concentrations in river water. Rivers are fed by a combination of groundwater and surface water, resulting in variation due to local geology and precipitation. Because of its longer association with rocks, the chemistry of groundwater is often more concentrated but less variable than surface waters. Increases in flow due to rain events typically dilute stream water, although it is not a simple relationship to predict (Livingstone 1963). Golterman (1975) argued that there are two broadly-defined patterns relating discharge with water chemistry. The TDS may be inversely related to discharge, which is the expected dilution effect when the input of materials is relatively constant and chemicals are homogenously mixed in the river. Alternatively, ion concentrations may remain relatively constant in response to fluctuations in discharge. This pattern can be explained by two mechanisms. First, the water may be reaching equilibrium with soils; therefore, ion concentration would remain constant regardless of flow volume. Second, chemical concentrations may remain relatively constant if they are approaching saturation values for the system. The response to rising discharge in the Orinoco River in northern South America differs depending on the source of ions. This very large river system is characterized by a very high runoff rate and very low concentrations of geologically-derived nutrients because a large fraction of its catchment is underlain by a shield rock that is resistant to weathering and covered with undisturbed forest. Seasonal increases in discharge result in a dilution response of geologically-derived major ionic solids, including soluble silica and phosphorus. In contrast, biologically-derived substances, including organic carbon and nitrogen, exhibit a purging response in which their ion concentrations increase with increasing discharge (Lewis and Saunders 1989).
Studies of stream water draining the Hubbard Brook Experimental Forest in the northeastern US illustrate how ionic concentrations can change in response to seasonal variation in precipitation inputs, discharge, and the cycle of growth of the terrestrial vegetation (Likens and Bormann 1995). The relative constancy in stream chemistry evident in these data is notable and is probably typical of intact, undisturbed ecosystems (Fig. 4.3). Most dissolved substances vary within a narrow range (less than twofold), whereas stream flow can vary as much as four orders of magnitude over an annual cycle. In the Hubbard Brook Experimental Forest, virtually all drainage water must pass through its mature and highly permeable podzolic soils. This affords considerable buffering capacity, and accounts for the relatively constant chemical composition of stream water at these sites (Likens et al. 1970).

(Reproduced from Likens et al. 1967)
The relationship between the concentration of major ions and stream discharge in a small forested catchment in the Hubbard Brook Experimental Forest, New Hampshire, between 1963 and 1965
The relationship between discharge and ionic concentration may also vary among ions within a given site. For example, associations between cation concentrations and discharge were ion-specific over a two-year period in a single watershed in Hubbard Brook (Fig. 4.3). When all data were pooled in the analyses, there were no significant relationships between potassium, calcium, or magnesium and discharge. However, sodium concentrations were negatively related to flow. This pattern is most likely due to a dilution effect resulting from limited sodium availability in the system (Likens et al. 1967). In contrast, similar work conducted in mountain streams in California documented negative relationships between calcium and magnesium concentrations and discharge (Johnson and Needham 1966), highlighting how regional differences in the physicochemical environment can influence stream water chemistry.
Although solute concentrations may exhibit only modest variation in response to discharge fluctuations in an intact ecosystem, human activities in a watershed, such as timber harvest and road construction, are significant disturbances that influence solute export. Following deforestation and suppression of regrowth by herbicides in a catchment of Hubbard Brook, the concentrations of most major ions increased in stream water, and total output of ions increased six-fold. Only ammonium and carbonate remained low and constant, and sulphate declined because of reductions in sulphate generation by sources internal to the ecosystem. The average concentrations of calcium and magnesium increased by over 400%, sodium by 177%, and potassium concentrations increased over 18-fold. Altered ion concentrations were attributed to increased discharge, changes in the nitrogen cycle within the ecosystem, and higher temperatures (Likens and Bormann 1995). Use of best management practices (BMPs) can moderate the response of stream water chemistry to the disturbances associated with timber harvest. A long-term study of a well-managed clear-cut of a forested catchment found that almost all solutes and nutrients showed elevated concentrations and exports after harvest. However, overall losses were relatively small and judged not detrimental to forest productivity because the forest was harvested using several BMPs including cable logging, which minimized the need for roads and dragging of felled trees (Swank et al. 2001).
Climate also exerts considerable influence over regional variation in the chemical composition of rivers. Across a gradient from arid to humid conditions, a general inverse relationship is seen between annual precipitation and total solute concentration. High concentrations of total dissolved ions are found in rivers draining arid areas due to the small volumes of precipitation and runoff, salt accumulation in the soil, and evaporation (Walling 1984). Changes in the timing and intensity of precipitation associated with anthropogenic climate change has the potential to influence the chemical composition of rivers and streams. For example, work in boreal streams illustrates the impact of increasing discharge and winter climate conditions on inter-annual variation in dissolved organic carbon concentrations in streams during snowmelt (Ågren et al. 2010).
Research from long-term (~20 year) studies demonstrates that concentrations of solutes in stream water seem to have seasonal patterns that are driven by geology, terrestrial vegetation, and temporal patterns in climate variables, including insolation (a measure of incident solar radiation per unit area and time), temperature, and precipitation (Lucas et al. 2013; Lutz et al. 2012; Navrátil et al. 2010). Long-term studies also provide insight into how environmental change may alter relationships between ion concentrations, especially Ca2+, Mg2+, and SO42−, and other watershed attributes. In a 20-year study of the Walker Branch Watershed in Tennessee in the southeastern US, researchers documented a ~1.0 °C increase in mean annual temperature, a ~30% increase in the rates of forest evapotranspiration, and a ~20% decline in the amount of precipitation entering the watershed, which were linked to a ~34% decline in river runoff (Lutz et al. 2012). Concentrations of solutes changed significantly with discharge, showing dilution for Ca2+ and Mg2+, and increasing concentrations for SO42− (Fig. 4.4). This difference is because calcium and magnesium are controlled by bedrock weathering, whereas the main source for sulphate in this system is wet and dry deposition from nearby coal-fired power plants. In addition, over the 20-year record, concentrations of Ca2+ and Mg2+ increased, and this is attributable to reduced precipitation. As a result of climate-related declines in runoff, groundwater, rich in Ca2+ and Mg2+, has become an increasing proportion of stream flow.

(Reproduced from Lutz et al. 2012)
Relationships between the concentration of geochemical solutes and discharge. Original concentration units are mg L−1 and discharge units are in L s−1. Values are ln-transformed. Regression summaries are listed above each panel and all relationships are significant
In response to the implementation of environmental regulations, such the 1970 Clean Air Act in the US, many, but not all watersheds, have experienced declines in SO42− deposition (Lutz et al. 2012; Navrátil et al. 2010). Coal-fired power plants are still active in many regions throughout the globe, and nearby watersheds may still receive relatively large inputs of SO42− through deposition (e.g., Fig. 4.4) that can influence concentrations of other ions. Once deposited in the watershed, the impact of SO42− on stream water chemistry is variable and depends largely upon the volume of surface water runoff, soil chemistry, and underlying geology. Analyzing a 20-year dataset from 60 Swedish streams, Lucas and others (2013) documented declines in Ca2+, Mg2+, K+, and Na+ since the early 1990s that were related to declines in SO42− in streams in southern Sweden. In contrast, cation concentrations in streams in northern Sweden—a region that was not affected as strongly by anthropogenically derived SO42− deposition—were characterized by seasonal variability linked to climate variables, such as temperature and precipitation (Lucas et al. 2013).
Rising ion concentrations due to other anthropogenic activities, such as agricultural development, wastewater discharge, and natural resource extraction, also threaten the chemical integrity of freshwaters throughout the globe (Griffith 2014). Increases in some ions, particularly bicarbonate salts, chlorides, and sulphates, can alter stream community structure and function by decreasing the number of species able to live in a system and increasing the mortality of stream organisms (Johnson et al. 2015; Tyree et al. 2016).
4.2.2 The Dissolved Load
The dissolved load of a river is the product of concentration and discharge, and usually is expressed as kg day−1 or metric tons yr−1. In comparing catchments or river basins it is helpful to present this as a yield per unit area, by dividing by the drainage area. Streamflow is more variable than ionic concentration, and so between-year variation in the export of ions can depend strongly on inter-annual variation in discharge (Fig. 4.5). Because discharge and ionic concentration often are inversely related, the range of the dissolved load of ions transported by the world’s major rivers varies over only two orders of magnitude, from around 3 to as high as 500 t · km−2 yr−1, with highest values observed in small alpine rivers (Meybeck 1977). The dissolved load typically experiences less temporal variation than does the amount of suspended solids carried by rivers. Greater discharge of rivers in wetter regions of the globe more than compensates for lower ionic concentrations; therefore, the dissolved load is usually smaller in arid regions when compared to areas with more surface runoff.

(Reproduced from Likens and Bormann 1995)
Between-year variation in the gross output of calcium, sodium, magnesium, and potassium (kg ha−1) primarily depends on between-year variation in discharge for undisturbed catchments of the Hubbard Brook Experimental Forest. Data were collected between 1963–1974
4.2.3 Chemical Classification of River Water
If one relates the relative proportions of principle anions and cations in the world’s surface waters to the concentration of total dissolved solids, a curve with two arms emerges (Fig. 4.6). Gibbs (1970) interpreted this as evidence that three major mechanisms control surface water chemistry. At the left side of the “boomerang” lie systems where the rocks and soils of river basins are the predominant source of their dissolved materials. Relief, climate, and age and hardness of rocks determine the positions of rivers within this grouping. Proceeding along the lower arm to the right of the figure we encounter waters that are lower in ions and whose chemical composition most closely resembles the rain. These are mainly the tropical rivers of Africa and South America, with their sources in highly leached areas of low relief. In tropical humid regions, precipitation with a composition similar to seawater is an important source of ions. The average ionic concentrations of river water in these regions are typically less than 30 mg L−1. Proceeding along the upper arm to the right, we encounter systems with high concentrations of dissolved ions and again a relative predominance of Na+ and Cl−. These are rivers of hot, arid regions, where the combined influence of evaporation and precipitation of CaCO3 from solution accounts for their higher ionic concentrations. Total dissolved salts exceed 1000 mg L−1 in streams in these regions, and can be as high as several g L−1, as seen in some rivers of central Asia (Crosa et al. 2006). Thus, three major mechanisms—atmospheric precipitation, dissolution of rocks, and the evaporation-crystallization process—are considered to account for the principal trends of dissolved ions in the world’s surface waters. Other factors, including topography, vegetation, and the composition of rocks and soils then can be invoked to explain differences in stream chemistry within these major groupings.

(Modified from Gibbs 1970)
A classification of surface waters of the world based on ratios of sodium to calcium and chloride to bicarbonate, in relation to total dissolved salts. As one proceeds from left to right along the lower arm, inputs shift from a dominance of rock dissolution to a dominance of precipitation. The majority of large tropical rivers are found to the lower right. As one proceeds from left to right along the upper arm, sodium and chloride increase. These high salinity rivers lie in arid regions where evaporation is great. Note the vertical axis also reflects a gradient from high precipitation and runoff at the base to arid regions at the top
Justifiably, critics of this classification scheme question the interpretation of control at the ends of the boomerang. For example, the Río Negro’s chemistry is equally a consequence of its long history of intense weathering, and its basin, which is predominantly comprised of silicious rocks (Stallard and Edmond 1983). Similarly, saline rivers can be strongly influenced by near-surface halite deposits (Kilham 1990). These examples downplay the roles of precipitation and evaporation, and suggest that local geology can be of primary importance in determining river chemistry over all extremes. Regardless, most of the world’s rivers are closer to the middle than the ends of this diagram, are low in Na+/(Na+ + Ca2+), and are dominated by Ca2+ and HCO3− from carbonate dissolution. This is in accord with the long-held view that the weathering of sedimentary rocks provides most of the dissolved ions in most of the world’s major rivers (Berner and Berner 2012).
4.3 The Bicarbonate Buffer System
Dissolved CO2 reacts with H2O to form carbonic acid (H2CO3), a weak inorganic acid that occurs at low concentrations relative to unhydrated CO2 at pH values <8 (Wetzel 2001). H2CO3 further dissociates to form hydrogen (H+), bicarbonate (HCO3−), and carbonate (CO32−) ions. The latter in turn react with water to form hydroxyl ions (OH−). When the natural content of carbonate rocks is high, as is common in the sedimentary rocks that make up much of the earth’s surface, these reactions result in sufficient hydroxyl ions to produce alkaline waters. This is known as the CO2–HCO3−–CO32− buffering system, because it resists change in pH. Although addition of hydrogen ions will neutralize hydroxyl ions, more hydroxyl ions are formed immediately through the reaction of water with carbonate, and so pH resists change. However, once the supply of carbonate is exhausted, which occurs most rapidly in igneous drainages, pH can drop quickly.
The relative proportions of CO2, HCO3−, and CO32− are pH-dependent (Fig. 4.7). At a pH below 4.5 only CO2 and H2CO3 are present, and almost no bicarbonate or carbonate can be found. Indeed, bicarbonate concentration commonly is measured by titration with a strong acid until reaching a pH of about 4.3. At higher pH values dissociation of carbonic acid occurs, bicarbonate and carbonate are present, and CO2 and H2CO3 are no longer detectable. At intermediate pH values, HCO3− predominates. These dissociation dynamics are influenced by both temperature and ionic concentrations, and the relationships shown in Fig. 4.7 may not be valid for water of very high ionic concentration. The inorganic carbon required for photosynthesis is most easily obtained from CO2, although many photosynthetic organisms can utilize bicarbonate. By influencing the forms of inorganic carbon available, shifts in pH can influence the efficiency of different primary producers in the ecosystem.

(Reproduced from Wetzel 2001)
Influence of pH on the relative proportions of inorganic carbon species, CO2 (+ H2CO3), HCO3−, and CO32− in solution
Fresh waters can vary widely in acidity and alkalinity due to both natural and anthropogenic factors. Very acid and very alkaline waters are harmful to most organisms, and so the buffering capacity of water is critical to the maintenance of life. The pH of water is a measure of the concentration of hydrogen ions, hence the strength and amount of acid present. Because the scale of pH is logarithmic to the base 10, a decline of one pH unit represents a tenfold increase in hydrogen ion concentration. In natural waters, pH is largely governed by H+ ions from the dissociation of H2CO3 and OH− ions from the hydrolysis of bicarbonate. Rainwater is usually acidic (typically near a pH of 5.7) because of its carbon dioxide content, and also due to naturally occurring sulphate. Normally these acids are neutralized as rainwater passes through the soil. However, in catchments primarily composed of hard rocks, little buffering capacity, and large surface water (as opposed to groundwater) inputs, stream water will be acidic even if pollution is absent. Organic acids also contribute to low pH values. Where decaying plant matter is abundant, especially in swamps, bogs, and peaty areas, humic acids result in “brown” or “black” waters, and a pH in the range of 4–5. In addition, volcanic fumes and local seepage from sulphurous or soda springs can produce natural extremes of pH.
Alkalinity refers to the quantity and kinds of compounds that collectively shift the pH into the alkaline range (>7.0 pH). It is a measure of the capacity of the solution to neutralize acid, and is determined by titration with a strong acid and expressed as milliequivalents per L or mg L−1. The bicarbonate buffering system is mainly responsible for alkalinity. Hence, alkalinity is often measured as mg L−1 of CaCO3, which measures the acid-neutralizing capacity due to carbonate and bicarbonate. Because alkalinity is estimated from a filtered sample, it measures the capacity of solutes to neutralize acid. Acid neutralizing capacity (ANC) is similar, but is determined for an unfiltered sample and thus measures the capacity of solutes plus particulates to neutralize acid.
Hardness is another commonly used term to describe water quality. It is calculated by measuring the concentration of cations that form insoluble compounds with soap; hence it is primarily a measure of the concentration of calcium and magnesium salts. The common co-occurrence of calcium and bicarbonate has led some to equate estimates of hardness with measures of alkalinity. However, it is possible to document very high alkalinity in water with very little calcium or magnesium; thus, it can be inappropriate to equate these terms.
4.4 Biological Implications of Varying Ionic Concentrations
The ionic concentration of water can affect species distributions and biological productivity in rivers and streams throughout the world. This is especially evident when examining stream communities that are adapted to extremely high or extremely low ionic concentrations. Thus, human activities promoting changes in water chemistry may consequently lead to changes in the abundance and distribution of aquatic organisms and produce shifts in ecosystem processes.
4.4.1 Variation in Ionic Concentration
If one wished to develop an entirely artificial medium for the culture of freshwater invertebrates, as has been done for algae and zooplankton, a long list of chemicals would be needed in the mixture to successfully support population growth (Kilham et al. 1998). Unquestionably, stream-dwelling organisms require water of some minimal ionic concentration to survive. The majority of the evidence linking the ionic content of water to the stream biota comes from surveys, and streams with very low ionic concentrations often support smaller numbers of species and fewer organisms. Hynes (1970) described a number of natural examples where the species of algae, mosses, and higher plants differ between soft versus hard waters.
Among the invertebrates, it appears that mollusks, crustaceans, and leeches are more sensitive to the range of ionic concentrations than are aquatic insects. Such a dependence on water chemistry is expected for shell-building organisms because CaCO3 is necessary for shell deposition and growth. For instance, the amphipod Gammarus can be commonly found in streams of the English Lake District that have at least 3 mg L−1 calcium, but are rare in streams of lower concentrations. Similarly, historical surveys of mollusks have described relationships between species diversity and hardness. According to Hunter et al. (1967), regional mussel diversity is dependent upon a large range of chemical conditions. Roughly 5% of the molluscan species of a region will occur in extremely soft waters (<3 mg L−1 Ca2+), ~40% of the species are dependent upon moderately soft water (<10 mg L−1), ~55% of species will be found in intermediate waters (10–25 mg L−1), and the remaining species will be associated with hard water (>25 mg L−1). Research examining the potential range expansion of invasive zebra (Dreissena polymorpha) and quagga (D. rostriformis bugensis) mussels in the Laurentian Great Lakes suggests that invasion success is strongly related to ambient calcium concentrations (Baldwin et al. 2012).
The role of ionic concentrations in governing the condition and distribution of freshwater species is context dependent. Lodge et al. (1987) argue for a limiting role of calcium concentrations in governing the distribution of freshwater snails when <5 mg L−1, but suggest that other abiotic and biotic factors become more important when calcium concentrations are greater. The chemical characteristics of a system may also interact with local biological factors to influence species interactions. For example, snails reared in water of differing calcium concentrations produced larger, heavier, and thicker shells at higher calcium concentrations and also when exposed to water-borne cues of a predatory crayfish, Procambarus, indicative of the importance of calcium availability to predator avoidance (Bukowski and Auld 2014).
Alkalinity is widely used as a surrogate for stream fertility, linking alkalinity to productivity of fish populations via their food supply. Compiling data on salmonid production and stream alkalinity from numerous studies in the US, Kwak and Waters (1997) demonstrated a statistically significant but nonetheless imperfect relationship between the two variables (Fig. 4.8). However, their own data from 13 southeastern Minnesota streams did not show any dependence of salmonid production on stream alkalinity, which the authors suggested may be due to the modest range of alkalinity values among sites as well as the influence of additional variables including temperature and biological interactions. Recent work examining links between alkalinity and freshwater organisms has often focused on the relationships between alkalinity and toxicity of heavy metals in rivers and streams.

(Reproduced from Kwak and Waters 1997)
Relationship between the annual production of salmonids and alkalinity for streams in southeastern Minnesota (black circles) and other locations throughout the US.
4.4.2 Salinization and Alkalinization of Freshwater Systems
The natural range of salinity and alkalinity in inland waters is considerable. The impacts of salinization, or the process of increasing concentrations of dissolved salts in fresh waters, has received much attention by researchers. As described in Sect. 4.2, salinity differs from TDS in one important way: its measurement does not include the concentrations of organic compounds in water. Therefore, in relatively unpolluted water, TDS is often very similar to salinity and the terms are often used interchangeably. Alkalinization refers to the process of increasing concentrations of dissolved alkaline substances (e.g., bicarbonate, carbonate, and hydroxide) in freshwater systems. In comparison with acidification, alkalinization is not as well studied. Yet, increases in stream alkalinity are also associated with the application of road salts, accelerated weathering due to the deposition of acidic compounds, and altered microbial activities. In some cases, the processes are linked. For instance, the application or dissolution of salts containing strong bases and carbonates can increase both salinity and alkalinity in receiving waters (Kaushal et al. 2018).
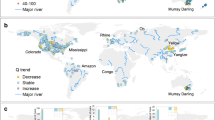
Globally, average salinity in rivers is approximately 120 mg L−1, a value thought to be elevated relative to natural conditions due to anthropogenically derived pollution (Berner and Berner 2012). Recent work has suggested that anthropogenic activities are largely responsible for increasing salinity and alkalinity in inland waters. This pattern, termed the freshwater salinization syndrome (Kaushal et al. 2018), can be seen over large spatial scales and across heterogeneous landscapes (Fig. 4.9). Specific drivers of the syndrome include salt pollution (irrigation runoff, road deicers, sewage), increased weathering of geological materials by strong acids (acid rain, acid mine drainage, fertilizers), and the abundance of easily weathered materials in agricultural (e.g., lime) and urban (e.g., concrete) environments (Kaushal et al. 2018).

(Reproduced from Kaushal et al. 2018)
A conceptual model of the freshwater salinization syndrome. At least three sets of processes contribute to freshwater salinization. They are listed in order from upstream to downstream including: (i) accelerated weathering throughout the network; (ii) human salt inputs; and (iii) enhanced biological alkalinization in larger systems in response to increased light and nutrient availability. Underlying geology results in different starting points in headwaters based on the potential for chemical weathering. Weathering processes introduce alkalinity, bicarbonate, and base cations along drainage networks. The relative influence of changes in weathering may respond to increasing disturbance along a river network, as salt and nutrient pollution typically change in response to hydrological modifications and stream size. The spatial and temporal extent of the freshwater salinization syndrome varies with land use, climate, and underlying geology
In colder regions of the globe, salts are used on roads to melt ice and reduce vehicle accidents (Fay and Shi 2012). Runoff carrying salts and other de-icing compounds can significantly elevate the salinity of receiving waters, and cause large fluctuations in salinity over relatively short time scales. The volume of deicing salts used on roadways has increased dramatically through time (Schuler and Relyea 2018a). Between 1950 and 2014, the volume of road salt applied to roads in the US increased from less than 1 million tons to approximately 20 million tons. Similar patterns of road salt application have been documented in other regions of the world (Schuler and Relyea 2018b).
Runoff from road salts can negatively affect aquatic populations, communities, and ecosystems (Schuler and Relyea 2018a). Laboratory experiments have repeatedly documented how road salts are toxic to individual species and have negative effects on animal populations. For example, work with rainbow trout (Oncorhynchus mykiss) demonstrated the distinct effects of three, commonly-used types of road salt on trout growth and development. When scaled to the population level, researchers suggested that the reduced growth caused by salts at critical early-life stages has the potential to alter population structure by reducing trout recruitment (Hintz and Relyea 2017). Field-based observational and experimental research indicates exposure to increasing salinity from road salt can shift community composition, initiate trophic cascades, and alter important ecosystem processes, such as denitrification. Notably, many negative effects of road salt have been observed at concentrations below the chronic and acute thresholds (230 mg Cl− L−1 and 860 mg Cl− L−1, respectively) that have been established by the US Environmental Protection Agency, suggesting that environmental regulations may not adequately protect the integrity of aquatic communities (Schuler and Relyea 2018b).
Results from a long-term ecological research project of urban areas as ecological systems, focused on the city of Baltimore in the eastern US, document the long-term trends and implications of increasing salinity in urban streams. Kaushal et al. (2005) reported chloride concentrations as high as 25% of seawater during winter, and this trend continues to increase (Fig. 4.10). Similarly, Bird et al. (2018) used 16 years of water chemistry data to investigate relationships between major ions and land cover in the Baltimore region through time. Though deicing salt and concrete were the principal nonpoint source contributions to ionic concentrations, higher concentrations of all major ions were recorded in regions with more urban land cover. Unexpectedly, the concentrations of most major ions were also increasing in urban streams through time, even with no major changes in land cover during the study. This pattern was not evident in forested and rural sites (Fig. 4.11). Collectively, these results provide additional evidence that road salt application has large impacts on freshwater communities, and that human activity can have long-term effects on salinity and alkalinity in streams.

(Reproduced from Kaushal et al. 2005)
The mean annual concentration of chloride increases with impervious surface area (an indicator of urbanization) for streams along a rural to urban gradient near Baltimore, Maryland. Dashed lines represent thresholds for damage to some land plants and chronic toxicity to sensitive aquatic organisms

(Reproduced from Bird et al. 2018)
Temporal changes in calcium concentrations in streams along a gradient of urbanization. Major ion concentrations were greater in areas with more urban land cover. Concentrations increased through time in areas with more urban development even through there were no corresponding major changes in land cover
Agricultural development and resource extraction in a watershed also can influence freshwater salinity (Tyree et al. 2016). In the US, agricultural runoff is a major source of excess Na+ and Cl− to rivers and streams. Increasing K+ concentrations are associated with rivers in the Midwestern US and most likely are due to the application of agricultural fertilizer (potash) rich in K+. Agricultural liming to support greater crop production also may enhance stream alkalinity and the concentration of dissolved salts in rivers and streams (Kaushal et al. 2018). Secondary salinization, or the salinization of soil, surface water, or groundwater due to human activities, is a particular problem in arid and semi-arid areas due to high demands for both surface and groundwater for irrigation to support dryland agriculture. Irrigation in dryland regions concentrates salts in streams in two ways: by reducing total water volume entering streams due to increased rates of evapotranspiration, and by increasing the salts entering streams from soil leachate due to increased infiltration rates. In Australia, where salinization is widespread in semiarid agricultural areas, salinity in the lower South Australian Murray River averages ~0.5 g L−1 (Williams and Williams 1991). Salinization of the Pecos River typifies challenges faced in arid watersheds in the US and Mexico. Reduced flows, inputs of high salinity groundwater, and increased evapotranspiration have collectively increased its salinity (Hoagstrom 2009).
Mining activities in a watershed can also increase salinity and alkalinity in streams and rivers, as chemicals added to mine effluent to neutralize pH, precipitate metals, and oxidize sulfur compounds prior to discharge can introduce large volumes of water containing salts, lime, and other constituents (Kimmel and Argent 2010). For example, stream reaches downstream of discharges from a Pennsylvanian mine complex in the eastern US had TDS and specific conductance values more than an order of magnitude greater than reference sites. Fish community metrics, including species richness (the total number of species), species density, and species diversity (species richness and species evenness), were much lower in sites with the highest TDS and specific conductance values relative to the reference sites (Kimmel and Argent 2010).
The ecological response to anthropogenically-driven changes in salinity and alkalinity is varied. Many aquatic species, especially those adapted to life in arid regions, can tolerate large fluctuations in and high concentrations of ions. For example, in two river systems of Australia, researchers found no relationship between macroinvertebrate community assemblages and salinity levels that exceeded 2 g L−1. Similarly, several fish species in the Murray River, also in Australia, survived laboratory exposures to salinities up to 30 g L−1, possibly reflecting a relatively recent marine ancestry for these species (Williams and Williams 1991). However, salinity tolerance can also structure aquatic communities. In the Red River in Texas, US, where salinities range from ~200 to ~35,000 mg L−1 TDS, fish communities are grouped into low-, medium-, and high-salinity assemblages. Conspicuously, the species most sensitive to higher salinities have experienced the greatest decline (Higgins and Wilde 2005), indicating that even in streams where organisms are adapted to higher ionic concentrations, increasing salinity presents a conservation challenge (Hoagstrom 2009; Hoagstrom et al. 2010). Similar patterns have been documented in rivers in other regions of the world. For instance, secondary salinization is responsible for changing diversity and abundance of the freshwater fish community rivers in southwestern Australia (Beatty et al. 2011).
4.4.3 Effects of Acidity on Stream Ecosystems
Fresh waters may be naturally acidic due to the decay of organic matter, and anthropogenically acidified by atmospheric deposition of strong inorganic acids formed from sulphate and nitrous oxides released in the burning of fossil fuels, or from acids leached from mining deposits. Naturally acidic waters, tea-colored from the breakdown of organic matter occur in diverse settings including northern peatlands, tropical regions such as the aptly named Río Negro, and blackwater rivers draining swamp forests such as the Ogeechee River in the southeastern US.
Acid precipitation is a phenomenon due to industrialization, and has its greatest influence in streams and rivers in regions of poor buffering capacity, especially those in granitic catchments. The strong inorganic acids H2SO4 and HNO3, formed in the atmosphere from oxides of sulphur and nitrogen released in the burning of fossil fuels, have lowered surface water pH in many regions of Europe and North America. The deleterious effects of acidic stream water on freshwater organisms are well established, primarily in terms of reduced numbers of species and individuals, but there also is evidence of altered ecosystem processes in response to declining pH. The relative impact of increased acidity on aquatic biodiversity is influenced by the amount of deposition, the buffering capacity of the system, and the life history and physiology of aquatic organisms.
Direct physiological effects of acidity are implicated by both field and laboratory studies of aquatic organisms. Reductions of pH have been linked to increased mortality rates and issues with the development of animals (Willoughby and Mappin 1988). Most likely, the negative outcomes are due to an increasing inability to regulate ions as acidity increases, including declining sodium concentrations in body tissues, and increasing inability to obtain sufficient calcium from surrounding waters (Økland and Økland 1986). Field collections indicate that the effects of lower pH may be taxon-specific, as plecopterans and tricopterans seem to tolerate waters of lower pH more effectively than do ephemeropterans and some dipterans. Hall and Ide (1987) speculate that differences in life cycle and respiratory style of these groups account for their differential susceptibility to acid stress.
Evidence suggests that the timing and intensity of exposure to waters of varying pH can produce varied outcomes in the behavior of aquatic organisms and aquatic community structure. For example, the availability and spatial location of refuge streams of moderate to high alkalinity influenced the habitat use of brook trout in streams of the central Appalachian Mountains of West Virginia, US. Spawning and recruitment occurred primarily in small tributaries with alkalinity above 10 mg L−1, whereas large adults apparently dispersed throughout the catchment (Petty et al. 2005). Similarly, Lepori et al. (2003) were able to differentiate between macroinvertebrate assemblages in streams that became acidic during snowmelt (pH reduced to 5) and those in well-buffered streams (pH remained above 6.6) in alpine regions of Switzerland.
Ecosystem processes also are affected by anthropogenic changes in stream pH (Ferreira and Guérold 2017). Inputs of autumn-shed leaves are an important energy supply to woodland streams, and breakdown rates respond to a number of environmental variables. Breakdown rates of beech leaves Fagus sylvatica varied more than 20-fold between the most acidified and circumneutral sites in 25 woodland headwater streams along an acidification gradient in the Vosges Mountains, France (Dangles et al. 2004a). More acidic streams experienced lower rates of microbial respiration and a reduction of microbial species that were associated with decaying leaves. It is important to note that field data indicate that naturally acidic streams may not behave like streams experiencing human-driven increases in acidity. For example, even at a pH of 4, neither the number of microbial taxa nor leaf decomposition rates were strongly depressed relative to more alkaline systems in naturally acidic streams of northern Sweden (Dangles et al. 2004b). The streams in question contain a unique fauna, suggesting that communities found in naturally acidic systems were adapted to the physiological challenges of this environment.
Acidic precipitation promotes the leaching of metals from soils and increased concentrations of free metal ions in affected rivers and streams, and this can affect aquatic organisms and ecosystem processes. Increasing free metal concentrations can be very deleterious to aquatic organisms, as they can accumulate on the surface of gills and impair osmoregulatory processes. For example, aluminum commonly occurs at elevated concentrations in acidic waters (Fig. 4.12). Separate and combined additions of aluminum compounds and inorganic acids to stream channels have been used to distinguish the direct influence of hydrogen ion concentration from the effects of elevated aluminum. In a short-term (24 h) manipulation of a soft-water stream in upland Wales, two salmonid species exhibited far greater susceptibility to the combined effects of acid and aluminum versus sulphuric acid alone, apparently because of respiratory inhibition (Ormerod et al. 1987). Episodes of acidification and elevated aluminum concentrations restricted stream fishes from sites in the northeastern US that had suitable chemical conditions (pH > 6 and inorganic Al < 60 μg L−1) at low flow (Baker et al. 1996). Abundances of the brook trout (Salvelinus fontinalis) were reduced and the blacknose dace (Rhinichthys atratulus) and sculpin (Cottus bairdi and C. cognatus) were eliminated from streams that episodically experienced pH < 5 and inorganic Al > 100–200 μg L−1. Behavioral avoidance is one cause of decline, and offers the possibility of subsequent recolonization if alkaline refuge areas are available. Baker et al. suggested that lower mobility in sculpins relative to brook trout may explain why the former were eliminated from acid-pulsed streams. Ecosystem processes, such as leaf-litter decomposition, can also be affected by exposure to Al. In a decomposition study of leaf litter in eight, first- and second-order streams in northeastern France, Ferreira and Guérold (2017) documented reductions in leaf decomposition rates of three tree species with increasing acidity (i.e., decreasing pH) and with increasing Al concentrations (Fig. 4.13).

(Reproduced from Wigington et al. 1996)
Relationship between aluminum concentrations (measured as inorganic monomeric Aluminum, Alim) and pH for streams of northeastern US. Note the wide range in Alim at low pH values

(Reproduced from Ferreira and Guérold 2017)
Relationship between the decomposition rates of leaves of three tree species that were enclosed in fine (a, c) and coarse (b, d) mesh bags, and pH (a, b) and total Al concentrations (c, d)
In response to increasingly stringent air pollution regulations in Europe and North America, there have been sharp reductions in the emission of chemicals contributing to acid rain in these regions. Recovery from acid rain is evident in the chemical characteristics of many systems. Though the biological response to the reductions has been slower than changes in water chemistry, evidence suggests that populations of freshwater organisms are beginning to recover in response to increasing pH in the water column (Warren et al. 2017).
Though acid rain is on the decline in many regions of the world, acidification of surface waters can be driven by other activities including mining. In addition to decreasing pH associated with acid mine drainage, mining activity in a watershed is often associated with increased concentrations of mixtures of heavy metals (e.g., lead, zinc, copper, and aluminum). Ambient metal concentration has been linked to declines in macroinvertebrate species richness and abundance. Notably, the effects of contaminants may influence organisms differently depending on their ontogeny, or life stage. For example, Schmidt et al. (2013) documented the impacts of metal bioavailability on larval and adult insect populations along a gradient of metal concentrations in streams in the Colorado Mineral Belt, a region that has been mined for the last two centuries. There was a significant negative relationship between metal concentrations and larval densities along the gradient, but densities fell precipitously when metal concentrations exceeded aquatic life criteria (cumulative criterion accumulation ratio (CCAR) ≥ 1). A contrasting pattern was seen in the relationship between metal concentration and the emergence of adult insects. Sharp declines in emergence were seen at lower concentrations, while smaller changes were seen in emergence rates in reaches with concentrations above the CCAR (Fig. 4.14). This work indicates that the emergence of adults was a more sensitive indicator of metal contamination, pointing to the need to consider environmental impacts across all stages of an organism’s development in order to effectively protect freshwater communities.

(Reproduced from Schmidt et al. 2013)
Effects of metal bioavailability on adult and larval aquatic insects. The Cumulative Criterion Accumulation Ratio (CCAR) is derived from the US EPA Aquatic Life Criteria for metals, which is based on the concentration of metals that can be present in surface water before it is likely to harm plant and animal life
Synergistic interactions between heavy metal pollution and land use have the potential to mediate the impact of metal contamination on aquatic ecosystems. Metal toxicity likely is less important in naturally brown- and blackwater streams (Collier et al. 1990; Winterbourn and Collier 1987) or in watersheds with wetlands, owing to the chelating abilities of humic acids, which bind metal ions mobilized at low pH. However, although some evidence indicates algal communities may be resistant to heavy metal contamination, research does suggest that microbial communities can be strongly impacted by metals (Schuler and Relyea 2018a).
Addition of lime to neutralize acid conditions is widely practiced. The River Auda in Norway had lost its anadromous salmon and sensitive mayflies due to anthropogenic acidification when liming commenced in 1985. Within two years, sensitive mayflies had returned, and additional macroinvertebrates appeared over the following five-plus years. However, in some cases liming has not been sufficient to offset the effects of episodic acidification. In three acidified Welsh streams that were evaluated for 10 years following the liming of their catchments, pH increased to above 6 and the number of macroinvertebrate species increased, but relatively few acid-sensitive species recovered (Ormerod and Edwards 1987). The occasional appearance but limited persistence of acid-sensitive taxa in limed streams led the authors to suggest that episodes of low pH continued to affect acid-sensitive taxa even after liming. Whether it makes sense to add lime to naturally acidic streams presents a further complication. In Sweden, approximately US $25 million has been spent since 1991 to lime some 8000 lakes and 12,000 km of streams to restore their condition, and as Dangles et al. (2004b) point out, expending funds to lime naturally acidic systems may not be wise management.
4.5 Legacy and Emerging Chemical Contaminants
Globally, rivers and streams face pollution problems associated with both legacy and emerging contaminants. Legacy contaminants are chemicals often produced through industrial and agricultural activities that remain in streams and rivers long after they are first released into the environment (Macneale et al. 2010). Emerging freshwater contaminants can be defined as naturally occurring or manmade substances that present realized or potential risks to the structure and function of rivers and streams (Nilsen et al. 2019). For example, substances that have never previously been detected or substances that were previous found in much lower concentrations in flowing waters may be considered emerging contaminants because their impacts on human health and the environment are not well understood.
4.5.1 Legacy Contaminants in Rivers and Streams
Examples of legacy contaminants that threaten the structural and functional integrity of freshwaters include metals (e.g., mercury and lead) and pesticides (e.g., dichloro-diphenyl-trichloroethane (DDT)), and polychlorinated biphenyls (PCBs). Often, legacy contaminants collect in sediments that can be repeatedly resuspended in the water column to interact with other chemical constituents and aquatic biota. Some legacy contaminants such as DDT, PCBs, and mercury accumulate in animal tissues because they are differentially retained rather than excreted. Therefore, the longer animals are exposed to the contaminant, the higher the concentration of the contaminant in their tissues will be. Some legacy contaminants also biomagnify, meaning contaminant concentrations increase in tissues of animals feeding at higher trophic levels (i.e., a tertiary consumer would be expected to have higher tissue concentrations than a primary consumer in the same environment). Rachel Carson famously wrote about the environmental implications of the broad consequences of legacy contaminants in her book, Silent Spring (Carson 1962).
A notable example of a biomagnifying legacy contaminant is DDT, a synthetic insecticide that was widely and effectively used in the 1940s and 50s to control mosquito populations and agricultural pests. Three million tons of DDT had been produced and used by 1970 (Zimdahl 2015). After a time, mosquito populations evolved resistance to the pesticide, and coupled with increasing amounts of data highlighting the disastrous environmental effects of DDT application, the US Environmental Protection Agency delivered a cancellation order for DDT in the US in 1972. Like many pesticides, DDT can be toxic to non-target organisms (Macneale et al. 2010). It has been linked to egg-shell thinning and subsequent population declines in many North America bird species. This was especially true for birds of prey, including the osprey (Pandion haliaetus) and bald eagle (Haliaeetus leucocephalus), as they accumulated high concentrations of DDT through biomagnification.
Though use of DDT in the US declined precipitously after the cancellation order, application of the pesticide was still permitted with federal exemption. Additionally, until 1985 the US continued to produce and export the pesticide to countries where there were fewer restrictions on pesticide use (Turusov et al. 2002). As of 2003, only three countries continue to produce DDT—China, the Democratic People’s Republic of Korea, and India. However, the emergence and spread of mosquito-borne illnesses, such as Zika and Chikungunya, and the continued persistence of malaria and dengue, support the continued use of the pesticide. For example, DDT continues to be used in many countries for the control of malaria and leishmaniosis; yet, data suggest that disease vectors in affected regions are developing resistance to the pesticide, and that DDT application may eventually decline further (Van Den Berg et al. 2017).
4.5.2 Emerging Contaminants in Running Waters
In a review assessing the potential risk of contaminants of emerging concern (CEC) on aquatic food web ecology, Nilsen et al. (2019) asserted there are currently five primary challenges in evaluating these risks. First, we lack detailed information about complexity of mixtures of CECs that exist in the environment. Second, there is insufficient understanding of the sublethal impacts of CECs on aquatic organisms. Third, research is needed to elucidate some of the biological consequences of the frequency and duration of exposure to CECs and whether these effects are evident across generations (e.g., maternal effects). Fourth, we need a better understanding of how exposure to other environmental stressors (e.g., changes in temperature or flow) may exacerbate or reduce the impact of exposure to CECs. Last, research is needed to understand how CEC exposure can influence trophic relationships. The term “emerging” implies a potential for change—a substance defined as a CEC may later be defined as a legacy contaminant or may be recognized as a new, but relatively benign addition to freshwater systems. In light of this uncertainty, our discussion focuses on CECs that have been observationally or experimentally linked to changes in the structure and function of aquatic systems, while acknowledging this is not an exhaustive list. For those interested in learning about the newest CECs and the methods to detect them in aquatic systems, Dr. Susan Richardson leads a biennial review of developments in water analysis for CECs in Analytical Chemistry (e.g., Richardson and Kimura 2016; Richardson and Temes 2018).
In response to the well-known, negative environmental effects of many pesticides and herbicides, there has been a large investment in developing new chemicals and methods to combat agricultural pests and arthropod vectors of human disease. Currently, glyphosate‐based herbicides, marketed under the trade name Roundup®, are among the most widely used agricultural chemicals globally (Annett et al. 2014). There is a wide diversity of formulas and applications of glyphosate; hence, aquatic communities may be simultaneously exposed to a variety of formulas and concentrations of the herbicide, with poorly understood consequences. It is likely that glyphosate toxicity is species-specific, and may occur through multiple mechanisms, influenced by the timing, formulation, and quantity of the herbicide applied(Annett et al. 2014). Amphibians seem to be particularly sensitive to exposure to the herbicide due to aspects of their physiology and life history. In a mesocosm study investigating the impact of glyphosate on aquatic macroinvertebrates, initial evidence suggests that under experimental conditions, sedimentation, rather than pesticide addition, had a stronger impact on drift and adult emergence (Magbanua et al. 2016).
Fracking, the hydraulic fracturing of rock by injection of a pressurized liquid, has markedly increased the amount of natural gas used to produce energy in the US and elsewhere. Through this process, a mixture of water, sand, and other chemicals are injected at high pressure into a wellbore, or deep hole, to create cracks in deep rock formations to release natural gas and other materials. If not treated properly, the wastewater produced from fracking can contaminate surface and groundwater supplies. Reliance on coal may be reduced, improving air quality, but other environmental harm can occur due to high water consumption, contaminants in return water, and release of methane, a powerful greenhouse gas. In watersheds with newly drilled natural gas wells, streams and rivers are threatened with increases in sediment runoff, reduced streamflow, and contamination from introduced chemicals and the resulting wastewater (Entrekin et al. 2011). There is evidence linking fracking to increases in concentrations of ions in surface waters used as drinking water (Harkness et al. 2015). In a study of streams in the Marcellus Shale basin in Pennsylvania in the eastern US, researchers examined stream physiochemical properties, trophic relationships, and mercury levels to assess whether fracking activities were correlated with changes in water chemistry, macroinvertebrate diversity, and trophic ecology. Fracking activities were associated with elevated methyl mercury concentrations, decreased stream pH, elevated dissolved mercury in stream water, and reductions in environmentally-sensitive macroinvertebrate taxa (Grant et al. 2016). In an examination of the relationships between instream microbial diversity and well-pad density, Trexler et al. (2014) documented lower diversity in bacterial communities collected in sites with natural gas development relative to reference sites. Moreover, community structure was more similar among bacterial communities collected from sites with gas development than communities at reference sites. This may be because fracking results in a decline in pH, which in turn is responsible for influencing microbial diversity at the population and community level.
Pharmaceuticals and personal care products (PPCPs) occurring in the water column, sediments, and organisms found in freshwater systems are an important class of chemical of emerging concern (Ebele et al. 2017). The diversity and use of PPCPs is increasing in both human and veterinary medicine throughout the globe. Caffeine and other stimulants, antihistamines, amphetamines, antibiotics, and antidepressants are all PPCPs commonly found in streams and rivers (Richmond et al. 2017). Many PPCPs are persistent in surface water, and are not removed by conventional wastewater treatment; hence they can move from wastewater effluent into freshwater systems. In addition, PPCPs often enter the natural environment via veterinary pathways without treatment (Ebele et al. 2017). Therefore, even with enhanced treatment technologies in existing systems or the construction of new facilities, increasing concentrations of PPCPs in freshwater systems will most likely remain a challenge for decades to come. Though research on PPCPs is relatively limited, researchers have begun to assess the risk they pose to the environment by examining their persistence in the environment, whether or not they bioaccumulate in aquatic food webs, and if and in what concentrations they are toxic to aquatic organisms (Ebele et al. 2017; Nilsen et al. 2019).
Existing literature on the potential ecological impacts of PPCPs gives reason for concern. Evidence indicates that these compounds are being consumed by aquatic organisms, may be transferred across terrestrial-aquatic boundaries through trophic interactions, can alter the growth and behavior of aquatic organisms, and can influence ecosystem processes (Richmond et al. 2017). A study examining relationships between wastewater discharge and PPCP concentrations in consumers documented increased concentrations of PPCPs in both aquatic and riparian consumers at sites with increasing influence from wastewater inputs (Fig. 4.15a; Richmond et al. 2018). Comparable within-site concentrations of PPCPs in aquatic invertebrate larvae and riparian predators indicate there may be a direct trophic transfer from emerging insects to riparian predators. Additionally, vertebrates feeding on aquatic macroinvertebrates could be consuming a wide variety of PPCPs, and in the case of some drugs (i.e. antidepressants), they may be ingesting as much as one-half of a recommended therapeutic dose for humans (Fig. 4.15b).

(Modified from Richmond et al. 2018)
(a) Pharmaceutical concentrations in aquatic invertebrates and riparian spiders. Total pharmaceutical concentration (ng g−1 dry weight) in riparian web-building spiders (dark grey) and aquatic invertebrates (black), arranged in decreasing order of wastewater influence. Violin plots illustrate kernel probability density and horizontal lines within each plot reflect median concentrations. (b) Estimated dietary intake of pharmaceuticals by two invertebrate predators compared to recommended pharmaceutical doses for humans. Dietary intake rates are displayed as the percentage of recommended human doses by therapeutic class for platypus (black) and brown trout (grey) in Brushy Creek in southeastern Australia. (CCBs calcium channel blockers, GORD gastroesophageal reflux disease, NSAID non-steroidal anti-inflammatory drugs, RAS renin angiotensin system)
Exposure to PPCPs may influence ecosystem function, as Rosi et al. (2018) showed by exposing biofilm communities to drugs commonly found in wastewater. Biofilm respiration rates varied in response to exposure to caffeine, cimetidine (acid-reducer, commonly used to treat ulcers), ciprofloxacin (antibiotic), and diphenhydramine (antihistamine) among four streams along an urban gradient. Respiration rates were low in the least urban stream when exposed to the PPCPs, but respiration in the most urban stream was resistant to drug exposure (Fig. 4.16). This suggests that biofilm communities in more urban systems may be exposed to a suite of PPCPs that select for populations of bacteria that can maintain functional processes even when exposed to contaminants.

(Reproduced by Rosi et al. 2018)
Respiration rates of biofilm communities grown on organic substrates and exposed to pharmaceuticals in streams along an urbanization gradient (left to right) (a) caffeine, (b) ciprofloxacin, (c) cimetidine, and (d) diphenhydramine. Bars indicate response ratios of respiration rates for treatments with pharmaceuticals to no pharmaceutical controls. The dashed line is equal to a response ratio of 1, indicating no effect of exposure. Lowercase letters indicate significant differences between sites (Tukey’s honestly significant difference; P < 0.05). Error bars represent standard error
Engineered nanomaterials (ENMs) are small (1–100 nm) particles used in various manufacturing applications. They have high surface to volume ratios and can exhibit unique physical and chemical properties, making it difficult to predict their impact on stream ecosystems. Often, ENMs are engineered with surface coatings that can increase their bioavailability and bioreactivity by orders of magnitude (Reid et al. 2018). Sensitivity to ENM toxicity varies among species and between stages of development (Callaghan and MacCormack 2017). Pesticides, herbicides, and PPCPs can all be converted into ENMs; thus, the ecological risks posed by ENMs also include the aforementioned risks presented by emerging contaminants already discussed.
4.6 Plastic Pollution in Freshwater Systems
Microplastics fragments (<5 mm) resulting from the fragmentation of plastic debris or consumer and industrial products are a newly recognized contaminant of aquatic ecosystems worldwide (Hoellein et al. 2014, 2019). First recognized in marine systems, an estimated 80% of plastic in the sea originates from inland sources and is transported by rivers to the oceans (Mani et al. 2015). A survey of microplastics over 820 km of the Rhine River from Basel to Rotterdam, the first and last major industrial cities along this large European river, found microplastic particles everywhere, but most abundantly near large cities (Fig. 4.17). Sources of variability include proximity to wastewater treatment plants, hydrological effects of mixing and turbulence, and timing of sampling relative to timing of releases. Given the origin of many microplastics from the fragmentation of synthetic fabrics and abrasives in personal care products, it is not surprising that concentrations are highest below wastewater treatment plants, as McCormick et al. (2016) reported from surveys upstream and downstream of WWTP effluent sites at nine rivers in Illinois, US.

(Reproduced from Mani et al. 2015)
Microplastics profile of the Rhine River in Europe. Horizontal columns indicate microplastic abundance 1000 m−3 and the fraction of the kinds of microplastic. Letters indicate the position in the cross section of the river (L: left bank, M: mid-river, R: right bank, T: transect)
Although research into ecological effects are at an early stage, negative consequences for growth and survival of aquatic organisms are a concern. Chemical contaminants can adhere to particles that are small enough to be ingested by invertebrates, and passed up the food chain. Ingestion poses risks of gut irritation and blockage, and of nutritional value that may be low or absent (Scherer et al. 2018). Transfer of toxins adhered to plastic into organisms may pose an even greater risk, but that has yet to be ascertained.
Macroinvertebrates collected from several rivers in southern Wales, UK, identified microplastic particles in roughly half of the individuals sampled (baetid and heptageniid mayflies and hydropsychid caddisflies) and at all sites (Windsor et al. 2019). Particles were significantly reduced when invertebrate gut contents were evacuated prior to analysis, indicating that most particles were acquired by ingestion. Microplastics are indeed entering freshwater food webs, with the potential for trophic transfers, and they have been observed in the guts of predatory fish in UK waters (Horton and Dixon 2018). Microplastic also serves as a substrate for bacterial assemblages, but whether this benefits particle consumers is uncertain. RNA sequencing revealed that microplastics supported distinct bacterial assemblages that differed from those found on natural particles in having fewer taxa, apparently those best able to degrade plastic polymers (McCormick et al. 2016).
4.7 Summary
The constituents of river water include suspended inorganic matter, dissolved major ions, dissolved nutrients, suspended and dissolved organic matter, gases, and trace metals. The dissolved gases of importance are oxygen and carbon dioxide. Exchange with the atmosphere maintains the concentrations of both at close to the equilibrium determined by temperature and atmospheric partial pressure, especially in streams that are small and turbulent. Photosynthetic activity in highly productive settings can elevate oxygen to super-saturated levels during the day. Respiration has the opposite effect, reducing oxygen and elevating CO2. High levels of organic waste can reduce oxygen concentrations below life-sustaining levels, and CO2 can be elevated from groundwater inputs or biological activity.
Many factors influence the composition of river water and, as a consequence, it is highly variable in its chemical composition. The concentration of the dissolved major ions (Ca2+, Na+, Mg2+, K+, HCO3−, SO42−, Cl−) in river water is roughly 120 mg L−1 on a world average. However, the chemical makeup of river water is highly variable, ranging from a few mg L−1 where rainwater collects in catchments of very hard rocks to some thousands of mg L−1 in arid regions.
Chemical variation among streams is primarily governed by the type and composition of rocks and by the amount and chemical composition of precipitation in a given region. For instance, the concentration of total dissolved salts is roughly twice as great in rivers draining sedimentary terrain compared to rivers flowing through regions dominated by igneous and metamorphic rock, owing to differences in weathering and chemical composition of the parent material. Due to increased rates of dilution and decreased rates of evaporation, regions with large amounts of precipitation and surface water runoff typically have stream water with lower ionic concentrations relative to water flowing through arid climates. The influence of precipitation on water chemistry is typically minimal except in regions with high surface runoff flowing over recalcitrant rocks.
River chemistry changes temporally under the multiple influences of seasonal changes in discharge regime, precipitation inputs, and biological activity. Groundwater typically is both more concentrated and less variable in ionic concentration than surface waters, because of its longer association with the underlying geology. In undisturbed catchments, some ions are remarkably constant across discharge fluctuations spanning several orders of magnitude. However, because rainfall increases the surface water contribution, increases in flow often dilute ion concentration.
Natural waters contain a solution of carbon dioxide, carbonic acid and bicarbonate, and carbonate ions in an equilibrium that serves as the major determinant of the acidity-alkalinity balance of fresh waters. Rivers can vary widely in acidity and alkalinity. Extreme values of pH (much below 5 or above 9) are harmful to most organisms. The bicarbonate buffer system, consisting of the CO2–HCO3−–CO32− equilibrium, provides the buffering capacity of fresh water that is critical to the health of aquatic biota. The abundance and diversity of riverine organisms are often strongly connected to the chemical composition of water. Water of very low ionic concentration appears to support a reduced fauna, particularly of crustaceans and mollusks. The number of species commonly increases with hardness, and many taxa are distinctly “soft-water” or “hard-water” forms.
Anthropogenic activities can significantly impact the chemical composition of rivers and streams. Land use conversion from natural to agricultural, industrial, or urban landscapes can alter the volume and chemical composition of precipitation, surface water, and groundwater entering watersheds. Human-derived changes in pH, salinity, and the concentrations of heavy metals are linked to changes in the structure and function of aquatic communities. Research focused on emerging contaminants, such as PCPPs, plastics, and nanoparticles, has demonstrated the ubiquity of anthropogenically-derived chemical changes in the world’s aquatic systems. However, much more research is needed to elucidate the response of aquatic organisms and ecosystem processes to acute and chronic exposure to known and emerging contaminants, and to the potentially interactive effects among anthropogenic stressors in freshwater systems.
References
Ågren A, Haei M, Kohler S et al (2010) Regulation of stream water dissolved organic carbon (DOC) concentrations during snowmelt: the role of discharge, winter climate and memory effects. Biogeosciences 7:2901–2913
Annett R, Habibi HR, Hontela A (2014) Impact of glyphosate and glyphosate-based herbicides on the freshwater environment. J Appl Toxicol 34:458–479
Baker J, Van Sickle J, Gagen C et al (1996) Episodic acidification of small streams in the northeastern United States: effects on fish populations. Ecol Appl 6:422–437
Baldwin BS, Carpenter M, Rury K et al (2012) Low dissolved ions may limit secondary invasion of inland waters by exotic round gobies and dreissenid mussels in North America. Biol Invasions 14:1157–1175
Beatty SJ, Morgan DL, Rashnavadi M et al (2011) Salinity tolerances of endemic freshwater fishes of south-western Australia: implications for conservation in a biodiversity hotspot. Mar Freshw Res 62:91–100
Berner EK, Berner RA (2012) Global environment: water, air, and geochemical cycles. Princeton University Press, Princeton, NJ
Bird DL, Groffman PM, Salice CJ et al (2018) Steady-state land cover but non-steady-state major ion chemistry in urban streams. Environ Sci Technol 52:13015–13026
Bukowski SJ, Auld JR (2014) The effects of calcium in mediating the inducible morphological defenses of a freshwater snail, Physa acuta. Aquat Ecol 48:85–90
Callaghan NI, MacCormack TJ (2017) Ecophysiological perspectives on engineered nanomaterial toxicity in fish and crustaceans. Compar Biochem Physi Part C: Toxic Pharma 193:30–41
Capps KA, Bentsen CN, Ramirez A (2016) Poverty, urbanization, and environmental degradation: urban streams in the developing world. Freshw Sci 35:429–435. https://doi.org/10.1086/684945
Carson R (1962) Silent spring. Houghton Mifflin Harcourt, New York
Christensen ER, Li A (2014) Physical and chemical processes in the aquatic environment. Wiley, Hoboken, NJ
Collier KJ, Ball OJ, Graesser AK et al (1990) Do organic and anthropogenic acidity have similar effects on aquatic fauna? Oikos 33–38
Connor R, Renata A, Ortigara C et al (2017) The United Nations World Water Development Report 2017. Wastewater: The untapped resource. The United Nations World Water Development Report. United Nations Educational, Scientific and Cultural Organization, Paris
Crosa G, Froebrich J, Nikolayenko V et al (2006) Spatial and seasonal variations in the water quality of the Amu Darya River (Central Asia). Water Res 40:2237–2245
Dangles O, Gessner MO, Guerold F et al (2004a) Impacts of stream acidification on litter breakdown: implications for assessing ecosystem functioning. J Appl Ecol 41:365–378
Dangles O, Malmqvist B, Laudon H (2004b) Naturally acid freshwater ecosystems are diverse and functional: evidence from boreal streams. Oikos 104:149–155
Dodds WK, Smith VH (2016) Nitrogen, phosphorus, and eutrophication in streams. Inland Waters 6:155–164. https://doi.org/10.5268/IW-6.2.909
Downing JA (2014) Limnology and oceanography: two estranged twins reuniting by global change. Inland Waters 4:215–232
Dutton CL, Subalusky AL, Hamilton SK et al (2018) Organic matter loading by hippopotami causes subsidy overload resulting in downstream hypoxia and fish kills. Nat Commun 9:1951
Ebele AJ, Abdallah MA-E, Harrad S (2017) Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerging Contaminants 3:1–16
Entrekin S, Evans-White M, Johnson B et al (2011) Rapid expansion of natural gas development poses a threat to surface waters. Front Ecol Environ 9:503–511
Fay L, Shi X (2012) Environmental impacts of chemicals for snow and ice control: state of the knowledge. Water Air Soil Pollut 223:2751–2770
Ferreira V, Guérold F (2017) Leaf litter decomposition as a bioassessment tool of acidification effects in streams: evidence from a field study and meta-analysis. Ecol Indic 79:382–390
Gameson A, Wheeler A (1977) Restoration and recovery of the Thames estuary. In: Cairns J, Dickinson KL, Herricks EE (eds) Recovery and restoration of damaged ecosystems. University of Virginia Press, Charlottesville, VA, pp 72–101
Gibbs R (1970) Mechanisms controlling world water chemistry. Science 170:1088–1090
Golterman H (1975) Chemistry. In: Whitton B (ed) River ecology. Volume 2. Studies in ecology. University of California Press, Berkeley, pp 39–80
Grant CJ, Lutz AK, Kulig AD et al (2016) Fracked ecology: response of aquatic trophic structure and mercury biomagnification dynamics in the Marcellus Shale Formation. Ecotoxicology 25:1739–1750
Griffith MB (2014) Natural variation and current reference for specific conductivity and major ions in wadeable streams of the conterminous USA. Freshw Sci 33:1–17
Hall RJ, Ide FP (1987) Evidence of acidification effects on stream insect communities in central Ontario between 1937 and 1985. Can J Fish Aquat Sci 44:1652–1657
Harkness JS, Dwyer GS, Warner NR et al (2015) Iodide, bromide, and ammonium in hydraulic fracturing and oil and gas wastewaters: environmental implications. Environ Sci Technol 49:1955–1963. https://doi.org/10.1021/es504654n
Hasler C, Butman D, Jeffrey J et al (2015) Freshwater biota and rising pCO2? 19:98–108. https://doi.org/10.1111/ele.12549
Higgins CL, Wilde GR (2005) The role of salinity in structuring fish assemblages in a prairie stream system. Hydrobiologia 549:197–203
Hintz WD, Relyea RA (2017) Impacts of road deicing salts on the early-life growth and development of a stream salmonid: salt type matters. Environ Pollut 223:409–415
Hoagstrom CW (2009) Causes and impacts of salinization in the lower Pecos River. Great Plains Res 2009:27–44
Hoagstrom CW, Zymonas ND, Davenport SR et al (2010) Rapid species replacements between fishes of the North American plains: a case history from the Pecos River. Aquat Invasions 5:141–153
Hoellein T, Rojas M, Pink A et al (2014) Anthropogenic litter in urban freshwater ecosystems: distribution and microbial interactions. PLoS One 9:e98485
Hoellein TJ, Shogren AJ, Tank JL et al (2019) Microplastic deposition velocity in streams follows patterns for naturally occurring allochthonous particles. Sci Rep 9:1–11
Horton AA, Dixon SJ (2018) Microplastics: an introduction to environmental transport processes. Wiley Interdiscip Rev-Water 5:10. https://doi.org/10.1002/wat2.1268
Hotchkiss E, Hall R Jr, Sponseller R et al (2015) Sources of and processes controlling CO2 emissions change with the size of streams and rivers. Nat Geosci 8:696
Hunter WR, Apley ML, Burky AJ et al (1967) Interpopulation variations in calcium metabolism in the stream limpet, Ferrissia rivularis (Say). Science 155:338–340
Hynes HBN (1970) The ecology of stream insects. Ann Rev Ent 15:25–42
Johnson C, Needham P (1966) Ionic composition of Sagehen Creek, California, following an adjacent fire. Ecology 47:636–639
Johnson E, Austin BJ, Inlander E et al (2015) Stream macroinvertebrate communities across a gradient of natural gas development in the Fayetteville Shale. Sci Total Environ 530:323–332
Kaushal SS, Groffman PM, Likens GE et al (2005) Increased salinization of fresh water in the northeastern United States. Proc Natl Acad Sci 102:13517–13520
Kaushal SS, Likens GE, Pace ML et al (2018) Freshwater salinization syndrome on a continental scale. Proc Nat Acad Sci 115:E574–E583
Kempe S, Pettine M, Cauwet G (1991) Biogeochemistry of European rivers. In: Degens ET, Kempe S, Richey JE (eds) Biogeochemistry of major world rivers. SCOPE 42. Scientific Committee on Problems of the Environment (SCOPE). Wilely, New York, pp 169–212
Kilham P (1990) Mechanisms controlling the chemical composition of lakes and rivers: data from Africa. Limnol Oceanogr 35:80–83
Kilham SS, Kreeger DA, Lynn SG et al (1998) COMBO: a defined freshwater culture medium for algae and zooplankton. Hydrobiologia 377:147–159
Kimmel WG, Argent DG (2010) Stream fish community responses to a gradient of specific conductance. Water Air Soil Pollut 206:49–56
Kwak TJ, Waters TF (1997) Trout production dynamics and water quality in Minnesota streams. Trans Amer Fishs Soci 126:35–48
Lauerwald R, Laruelle GG, Hartmann J et al (2015) Spatial patterns in CO2 evasion from the global river network. Glob Biogeochem Cycle 29:534–554
Lepori F, Barbieri A, Ormerod SJ (2003) Effects of episodic acidification on macroinvertebrate assemblages in Swiss Alpine streams. Freshw Biol 48:1873–1885
Lewis WM, Saunders JF (1989) Concentration and transport of dissolved and suspended substances in the Orinoco River. Biogeochemistry 7:203–240
Likens GE, Bormann FH (1995) Biogeochemistry of a forested ecosystem. Springer, Berlin
Likens G, Bormann F, Johnson N et al (1967) The calcium, magnesium, potassium, and sodium budgets for a small forested ecosystem. Ecology 48:772–785
Likens GE, Bormann FH, Johnson NM et al (1970) Effects of forest cutting and herbicide treatmetn on nutrient budgets in a Hubbard Brook watershed-ecosystem. Ecol Monogr 40:23–47. https://doi.org/10.2307/1942440
Livingstone DA (1963) Chemical composition of rivers and lakes (vol. 440). US Government Printing Office, Washington, DC
Lodge DM, Brown KM, Klosiewski SP et al (1987) Distribution of freshwater snails: spatial scale and the relative importance of physicochemical and biotic factors. Am Malacol Bull 5:73–84
Lucas RW, Sponseller RA, Laudon H (2013) Controls over base cation concentrations in stream and river waters: a long-term analysis on the role of deposition and climate. Ecosystems 16:707–721
Lutz BD, Mulholland PJ, Bernhardt ES (2012) Long-term data reveal patterns and controls on stream water chemistry in a forested stream: Walker Branch, Tennessee. Ecol Monogr 82:367–387
Macneale KH, Kiffney PM, Scholz NL (2010) Pesticides, aquatic food webs, and the conservation of Pacific salmon. Front Ecol Environ 8:475–482
Magbanua FS, Townsend CR, Hageman KJ et al (2016) Individual and combined effects of fine sediment and glyphosate herbicide on invertebrate drift and insect emergence: a stream mesocosm experiment. Freshw Sci 35:139–151
Magin K, Somlai-Haase C, Schäfer RB et al (2017) Regional-scale lateral carbon transport and CO2 evasion in temperate stream catchments. Biogeosciences 14:5003–5014
Mani T, Hauk A, Walter U et al (2015) Microplastics profile along the Rhine River. Sci Rep 5:17988
Marino GR, DiLorenzo JL, Litwack HS, Najarian TO, Thatcher ML (1991) General water quality assessment and trends analysis of the delaware estuary, part one: status and trend analysis. Najarian Associates, Eatontown, NJ. http://www.najarian.com/
McCormick AR, Hoellein TJ, London MG et al (2016) Microplastic in surface waters of urban rivers: concentration, sources, and associated bacterial assemblages. Ecosphere 7:22. https://doi.org/10.1002/ecs2.1556
Meybeck M (1977) Dissolved and suspended matter carried by rivers: composition, time and space variations and world balance. Paper presented at the Interactions between sediments and freshwater, Amsterdam
Navrátil T, Norton SA, Fernandez IJ et al (2010) Twenty-year inter-annual trends and seasonal variations in precipitation and stream water chemistry at the Bear Brook Watershed in Maine, USA. Environ Monit Assess 171:23–45
Nilsen E, Smalling KL, Ahrens L et al (2019) Critical review: grand challenges in assessing the adverse effects of contaminants of emerging concern on aquatic food webs. Environ Toxicol Chem 38:46–60. https://doi.org/10.1002/etc.4290
Økland J, Økland K (1986) The effects of acid deposition on benthic animals in lakes and streams. Experientia 42:471–486
Ormerod S, Edwards R (1987) The ordination and classification of macroinvertebrate assemblages in the catchment of the River Wye in relation to environmental factors. Freshw Biol 17:533–546
Ormerod S, Boole P, McCahon C et al (1987) Short-term experimental acidification of a Welsh stream: comparing the biological effects of hydrogen ions and aluminium. Freshw Biol 17:341–356
Petty JT, Lamothe PJ, Mazik PM (2005) Spatial and seasonal dynamics of brook trout populations inhabiting a central Appalachian watershed. Trans Am Fish Soc 134:572–587
Pringle CM, Triska FJ (1991) Effects of geothermal groundwater on nutrient dynamics of a Lowland Costa Rican stream. Ecology 72:951–965. https://doi.org/10.2307/1940596
Raymond PA, Hartmann J, Lauerwald R et al (2013) Global carbon dioxide emissions from inland waters. Nature 503:355
Reid AJ, Carlson AK, Creed IF et al (2018) Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol Rev 94:849–873
Richardson SD, Kimura SY (2016) Water analysis: emerging contaminants and current issues. Anal Chem 88:546–582. https://doi.org/10.1021/acs.analchem.5b04493
Richardson SD, Temes TA (2018) Water analysis: emerging contaminants and current issues. Anal Chem 90:398–428. https://doi.org/10.1021/acs.analchem.7b04577
Richmond EK, Grace MR, Kelly JJ et al (2017) Pharmaceuticals and personal care products (PPCPs) are ecological disrupting compounds (EcoDC). Elementa-Sci Anthrop 5:8. https://doi.org/10.1525/elementa.252
Richmond EK, Rosi EJ, Walters DM et al (2018) A diverse suite of pharmaceuticals contaminates stream and riparian food webs. Nat Commun 9:9. https://doi.org/10.1038/s41467-018-06822-w
Rosi EJ, Bechtold HA, Snow D et al (2018) Urban stream microbial communities show resistance to pharmaceutical exposure. Ecosphere 9:16. https://doi.org/10.1002/ecs2.2041
Scherer C, Weber A, Lambert S et al (2018) Interactions of microplastics with freshwater biota. Freshw Microplast. Springer, Cham, pp 153–180
Schmidt TS, Kraus JM, Walters DM et al (2013) Emergence flux declines disproportionately to larval density along a stream metals gradient. Environ Sci Technol 47:8784–8792. https://doi.org/10.1021/es3051857
Schuler MS, Relyea RA (2018a) A review of the combined threats of road salts and heavy metals to freshwater systems. Bioscience 68:327–335
Schuler MS, Relyea RA (2018b) Road salt and organic additives affect mosquito growth and survival: an emerging problem in wetlands. Oikos 127:866–874
Smith VH (2016) Effects of eutrophication on maximum algal biomass in lake and river ecosystems. Inland Waters 6:147–154
Stallard R, Edmond J (1983) Geochemistry of the Amazon: 2. The influence of geology and weathering environment on the dissolved load. J Geophys Res: Oceans 88:9671–9688
Sutcliffe D, Carrick T (1983) Relationships between chloride and major cations in precipitation and streamwaters in the Windermere catchment (English Lake District). Freshw Biol 13:415–441
Swank WT, Vose J, Elliott K (2001) Long-term hydrologic and water quality responses following commercial clearcutting of mixed hardwoods on a southern Appalachian catchment. For Ecol Manage 143:163–178
Thompson K, Christofferson W, Robinette D et al (2006) Characterizing and managing salinity loadings in reclaimed water systems. American Water Works Association, Alexandria, VA
Trexler R, Solomon C, Brislawn CJ et al (2014) Assessing impacts of unconventional natural gas extraction on microbial communities in headwater stream ecosystems in Northwestern Pennsylvania. Front Microbiol 5:522
Turusov V, Rakitsky V, Tomatis L (2002) Dichlorodiphenyltrichloroethane (DDT): ubiquity, persistence, and risks. Environ Health Perspect 110:125–128
Tyree M, Clay N, Polaskey S et al (2016) Salt in our streams: even small sodium additions can have negative effects on detritivores. Hydrobiologia 775:109–122
Van Den Berg H, Manuweera G, Konradsen F (2017) Global trends in the production and use of DDT for control of malaria and other vector-borne diseases. Malaria J 16:401
Visser PM, Verspagen JM, Sandrini G et al (2016) How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 54:145–159
Walling D (1984) Dissolved loads and their measurement. Erosion and sediment yield: some methods of measurement and modelling. Geo Books, Regency House, Norwich, pp 111–177
Walling D, Webb B (1975) Spatial variation of river water quality: a survey of the River Exe. Trans Instit Brit Geogr 1975:155–171. https://doi.org/10.2307/621615
Warren DR, Kraft CE, Josephson DC et al (2017) Acid rain recovery may help to mitigate the impacts of climate change on thermally sensitive fish in lakes across eastern North America. Glob Change Biol 23:2149–2153
Weiner ER (2012) Applications of environmental aquatic chemistry: a practical guide, 3rd edn. CRC Press
Welch KA, Lyons WB, Whisner C et al (2010) Spatial variations in the geochemistry of glacial meltwater streams in the Taylor Valley, Antarctica. Antarct Sci 22:662–672
Wen Y, Schoups G, Van De Giesen N (2017) Organic pollution of rivers: combined threats of urbanization, livestock farming and global climate change. Sci Rep 7:43289
Wetzel RG (2001) Limnology: lake and river ecosystems. Academic Press, San Diego, CA
Wigington PJ Jr, DeWalle DR, Murdoch PS, et al (1996) Episodic acidification of small streams in the northeastern United States: ionic controls of episodes. Ecol Appl 6:389–407
Williams M, Williams W (1991) Salinity tolerances of four species of fish from the Murray-Darling river system. Hydrobiologia 210:145–150
Willoughby L, Mappin R (1988) The distribution of Ephemerella ignita (Ephemeroptera) in streams: the role of pH and food resources. Freshw Biol 19:145–155
Windsor FM, Tilley RM, Tyler CR et al (2019) Microplastic ingestion by riverine macroinvertebrates. Sci Total Environ 646:68–74
Winterbourn M, Collier K (1987) Distribution of benthic invertebrates in acid, brown water streams in the South Island of New Zealand. Hydrobiologia 153:277–286
Young RG, Huryn AD (1998) Comment: improvements to the diurnal upstream-downstream dissolved oxygen change technique for determining whole-stream metabolism in small streams. Can J Fish Aquat Sci 55:1784–1785
Zimdahl RL (2015) Six chemicals that changed agriculture. Academic Press, London
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Allan, J.D., Castillo, M.M., Capps, K.A. (2021). Streamwater Chemistry. In: Stream Ecology . Springer, Cham. https://doi.org/10.1007/978-3-030-61286-3_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-61286-3_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-61285-6
Online ISBN: 978-3-030-61286-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)