
Abstract
The theory of sexual selection hypothesizes that, when females are selected to perform most parental investment, males compete for access to mates and polygyny evolves. Polygyny should result in high variance in male reproductive success, with a few males fathering a disproportionate number of pups. I tested this prediction by reviewing the literature on paternity, available for seven otariids. I identified a total of eight publications with data on paternity in seven species at 12 colonies. Overall, harem size produced an estimated mean of 20.7 pups per dominant male (assuming that the male fathered all pups born within its harem), but genetic analysis revealed that territorial males fathered 32.5% of pups born in colonies, resulting in an estimated two pups per territorial male, i.e. behavioural estimations of reproductive success was tenfold (20/2) higher than genetic ones; many males were unsuccessful in obtaining mates, even when following the dominant strategy; size of harem did correlate with paternity by territorial males but the slope was significantly lower than expected from behavioural estimations; differences in paternity between territorial and satellites males were not statistically significant in most studies. Generalizations of classical theories on the evolution of polygyny may need qualification in the light of evidence provided by molecular ecology.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Darwin’s (1871) theory of sexual selection hypothesised that physical and behavioral traits that provide advantages in male–male competition are, over time, selected for. Otariids (fur seals and sea lions) and elephant seals have been considered the most illustrative examples of this theory (Bonnes 1991; Cassini 1999, 2017). The theoretical approach is expressed in this sentence: ‘intense and bloody male aggression, extremely skewed male mating success, and a resulting extraordinary size dimorphism provide convincing evidence of the powers of sexual selection’ (Lindenfors et al. 2002, p. 188).
Trivers (1972) realized that differences in parental effort between the sexes are correlated with differences in sexual behavior and the reproductive system of a species. Polygyny is prevalent in species where males are freed from parental care duties. In other words, there is a phylogenetic potential for polygyny. In mammals, females are constrained by gestation to invest more parental effort, thus polygyny is expected to be the most common mating system in this taxon.
In another seminal paper written almost half a century ago, it was proposed that, in addition to phylogenetic factors, there are ecological features that determine the intensity of sexual selection in different species and populations (Emlen and Oring 1977). The environmental potential for polygyny depends on the degree to which multiple mates, or resources critical to gaining multiple mates, are economically defendable by males. Authors argued that the operational sex ratio predicts the intensity of sexual selection, defined as the average ratio of fertilizable females to sexually active males at any given time.
The development of molecular biology techniques during the last five decades has revolutionized most fields of biology, including the study of behaviour, ecology and evolution. Individual DNA profiling or fingerprinting became a standard tool after the development of the polymerase chain reaction (PCR) technique (Bartlett and Stirling 2003). Until the 1980s, it was assumed that genetic and behavioural levels of polygyny were correlated. At one extreme, in a monogamous system, a male should be the only father of the offspring born to his partner, while at the other, in a polygynous mating system, male mating success should be directly proportional to the number of females with which they are able to associate following successful competition with other males (Gonzalez-Suarez and Cassini 2014). The introduction of molecular markers provided opportunities to test these assumptions. Before analyzing the case of the Otariids, it is instructive to describe results from paternity estimates using genetic methods in birds.
Monogamy is the most common mating system in birds; traditionally over 90% of avian species were defined as monogamous (Lack 1968; Emlen and Oring 1977). Theory predicted that both parents are required for successful rearing of young, and that the costs of courting and mating with additional mates overcome benefits (Emlen and Oring 1977). Yet, molecular markers indicated that the incidence of extra-pair paternity was unexpectedly common. For example, in a review by Griffith (2007) of approximately 160 studies of avian parentage, extra-pair copulations occurred in over 130 bird species. Accordingly, in birds: (1) extra-pair paternity is recurrent, (2) variance in reproductive success (RS) among males is much higher than expected, and (3) less than 25% of avian species are actually genetically monogamous (Sheldon and Ellegren 1999; Whittingham and Dunn 2005).
This discoveries produced an impact that was partly responsible for what was defined as the “the first major triumph of modern sexual selection research” (Jones and Ratterman 2009, p. 10002): extra-pair paternity exerts great sexual selection pressure on males even in assumed monogamous species; for females, it is an opportunity for active mate choice. The aim of sexual selection theory became to discover the mechanisms that could explain the evolution of these female preferences (Jones and Ratterman 2009). There was a proliferation of models of mate choice and quantitative approaches to measure selection differentials (e.g., Mead and Arnold 2004; Kotiaho and Puurtinen 2007; Kokko et al. 2007).
In birds, the use of molecular markers for determining paternity become a cornerstone of sexual selection research, changing our understanding of mating patterns in bird natural populations (Jones and Ratterman 2009). This then raises the question of how this may apply to mammals. Here I will review the impact of the use of molecular markers on our understanding of the role of sexual selection in the evolution of mating systems in the Otariids (sea lions and fur seals), one of the most polygynous taxa among mammals.
Size sexual dimorphism and high levels of behavioural polygyny are traits shared by all 14 species of Otariids (Berta 2018). Males are on average three times heavier, but can become up to five or more times heavier, than females (Fig. 6.1). An example of a study in northern fur seal (Callorhinus ursinus ) illustrates the extreme polygyny of this taxon: a single male was observed to mate with 161 females while hundreds of other males were excluded from mating (Mesnik and Ralls 2018). Most male Otariids typically defend territories containing resources needed by females, while lekking has been proposed in at least four species: Zalophus californianus , Arctocephalus australis , Phocarctos hookeri (Mesnik and Ralls 2018 and references therein), and Otaria flavescens (Soto and Trites 2011). The mating system of O. flavescens has been also been defined as direct defence of females and of territories, often present in the same colony (Campagna and Le Boeuf 1988; Franco-Trecu et al. 2014).

Image showing sexual dimorphism in southern sea lions
In his pioneering work, Bartholomew (1970) provided an explanation of the evolution of mating systems in Otariids. Even today, his model remains the core paradigm to explain Otariid polygyny (Miller 2018). Otariids are the only mammalian family that lives part of the time at sea but copulate and give birth on land. Otariid females aggregate and synchronize reproduction, providing to males high environmental potential for polygyny. Dominant males, capable of occupying a territory or defending a position among female aggregates, have a great chance of multiple copulations; subordinate males, conversely, are excluded from mating. Males can gain reproductive advantages during pre-copulatory contest competitions if large and belligerent, which may have selected for sexual size dimorphism (Bartholomew 1970). Subordinate males may resort to alternative strategies, which would yield lower reproductive success than dominant congeners.
In recent years, several studies have been published that estimated paternity based on molecular markers. Using their findings, I test the main assumptions of sexual selection theory applied to the evolution of polygyny in Otariids: that males show a large variance in reproductive success, and that the most successful mating strategy is performed by dominant males that aggressively defend a space or position within an aggregation of females.
2 Methods
A literature search in the Web of Knowledge database was conducted in August 2018 to find behavioural and genetic studies of RS and paternity in Otariids.
Behavioural Measures
Two reviews summarized results on behavioural measures of reproductive success in 14 species (Bonnes 1991; Lindenfors et al. 2002; Table 6.1). Bonnes (1991) discussed as indicators the operational sex ratio, the intensity of sexual selection (Is), and the maximum number of observed copulations by a single male (Table 6.1). Is is the variance in fitness scaled in relation to the square of the average fitness, which requires a measure of fitness for each adult male (Wade and Arnold 1980). Wade and Arnold (1980) applied this concept to data from a population of birds, using offspring/adult male and mates/adult male to calculate fitness. Bonnes (1991) reviews its use in Otariids and concludes that Is is the least reliable measure, because its failure to obtain a representative sample can produce extremely misleading values. Is is also very sensitive to the count of males that fail to reproduce. Lindenfors et al. (2002) provided data on harem size for most species of Otariids (Table 6.1).
Genetic Measures
Eight studies provided data on paternity in seven species of Otariids (Arctocephalus australis , Franco-Trecu et al. 2014; A. forsteri, Caudron et al. 2010; A. gasella, Gemmell et al. 2001; Hoffman et al. 2003; Callorhinus ursinus , Kiyota et al. 2008; Neophoca cinerea , Ahonen 2013; Zalophus californianus , Flatz et al. 2012; Z. wollebaeki , Porschmann et al. 2010; Table 6.2). Two studies were conducted at the same study site (Gemmell et al. 2001; Hoffman et al. 2003) and shared some results. Three studies (Caudron et al. 2010; Ahonen 2013; Flatz et al. 2012) provided information on more than one study site; these were analysed separately, bringing the total sample size to 12. When data covered several years, the information was pooled. All studies used microsatellites as molecular markers. Table 6.2A provides information on the number of highly polymorphic microsatellite loci, and the minor value of confidence level in CERVUS (some studies used two levels), which reflect the accuracy of the methods used. Total pups/year, total males/year and total territories/year provided an estimation of the population size at the study site (Table 6.2B). Sampled effort per study site was estimated from the percentage of total males sampled, study years, and number and percentage of genotyped pups (Table 6.2C). Total pup paternity was the total number of offspring identified with fathers in the colony . Few studies discriminated between territorial and satellite fathers, others sampled only territorial males (Table 6.2D). Mean pups/male was calculated as the ratio of total pups with paternity per total candidate males. There are several measures that provide an estimation of variation in paternity within the samples (Table 6.2F): mean number of pups/male, number of fathers with more than two pups, and the maximum number of pups assigned to one father (together with the number of fathers with this level of paternity ). The final estimation of RS of males was obtained by extrapolating paternity of sample males to all males of the colony . This was obtained as: total pups in the colony per year * proportion of pups sired by males/total males in the colony per year. Some studies measured paternity only for territorial males. Variance in paternity was obtained from Gonzalez-Suarez and Cassini (2014), with the exception of A. australis , which was provided by Franco-Trecu et al. (2014).
3 Summary of Results from Paternity Studies
In A. australis , Franco-Trecu et al. (2014) found that the overall average number of pups per putative father was 3.0 (SD = 2.9, range = 0–13). They also showed that the mean number did not significantly differ (Welch t = −1.9, p = 0.06) between territorial (mean ± SD = 3.4 ± 3.1) and satellite (1.9 ± 1.5) males.
Three breeding areas were studied in A. forsteri (Caudron et al. 2010): 13, 4, and 7 pups were assigned a known father in the Ohau Point, North and South study areas, respectively. Sampled males included territorial (n = 3) and non-territorial (n = 12) individuals at Ohau, and 5 territorial males at the South area. The 8 territorial males fathered a total of 9 (4-2-2-1-0-0-0-0) pups born in their territories, while 8 non-territorial males of Ohau fathered a total of 4 pups (1-1-1-1-0-0-0-0). Five additional pups sired by 3 non-territorial males were assigned to South or North study sites (2-2-1), while 2 of the 3 territorial males of Ohau fathered 4 pups from these two colonies (2-2). For the purpose of the present analysis, the first 5 pups were considered as extra-harem copulations, while the last 4 pups were excluded from the analysis (although they could be considered extra-harem paternities because they are not related to stable females of the harem ; they were probably related to females visiting male territories or to ex-territorial males acting as satellites in other colonies). In summary, intra-harem and extra-harem paternities coincided in 1.12 (9/8) pups per territorial/satellite male. Total territorial males in the main study site was seven and all were sampled. None of territorial males genotyped in the South colony fathered any of the offspring sampled; the estimate of reproductive success of the territorial males of that colony was zero.
Gemmell et al. (2001) studied paternity in A. gazella territorial males in a colony with an average of 264 males and 700 pups per season. They sampled 243 territorial males and 184 pups, and they assigned a total of 65 paternities (35%) to the males (190 males with no paternities, 45 with 1, 4 with 2, and 4 with 3 paternities/male), given a RS of 0.34 pups/territorial male. Hoffman et al. (2003) studied the same species in the same colony and reported: (i) a mean number of pups assigned per male of 0.93, (ii) 58.8% of pups with assigned paternity (with the least conservative method), and (iii) 2.49 pups/sampled father.
Callorhinus ursinus was studied by Kiyota et al. (2008). They collected genetic samples from 37 males and 85 pups in 5 years in a colony with an average of 9 territories and 100 pups per year. Territorial males were classified into the following categories, according to their timing and duration of territory tenure: primary males (males that held territories before July 1 before the arrival of most females); secondary males (males that held territories after June 30); and temporary males (males that stayed in the breeding area for <1 day). They differentiated between associate and non-associate (primary or secondary) males, based on whether pups were identified to born or not in their territories. 98.2% of paternity corresponded to territorial males associated (45 pups) or not associated (10 pups) to females, and average reproductive success of territorial males was 10.9 pups/male (100 pups * 0.98/9 territorial males). However there were 29 pups without assigned males, 27 of which had a candidate father but the authors did not state whether they were territorial or temporary males. Had the 29 pups been sired by satellite males, paternity of territorial males would drop to 64.7%, with a reproductive success of 7.2 pups/territorial male. This result is significantly higher than those in other species, probably due to two characteristics of the study: (i) the study area was an isolated cove where satellite males would have low chances to interact with females, and (ii) the study focused more on older females who came ashore in the beginning of the reproductive season, and were more likely to copulate with territorial males than with satellite males (M. Kiyota, pers. commun.).
Ahonen (2013) studied three colonies of Neophoca cinerea during two seasons. Assigned paternities were 39.1%, 31.4%, and 19.3%, while total pups-total males were 196-112, 66-40, and 104-59 for Olive, Lilliput and Blefuscu sites, respectively. Estimated reproductive success of territorial males was: 0.59 (77/112), 0.52 (21/40), and 0.34 (20/59) pups/territorial male, respectively.
Flatz et al. (2012) studied two colonies of Zalophus californianus . Overall, they found considerably lower variation in male RS than expected in a species that exhibits behavior associated with strongly polygynous mating. With one exception, no male was identified as the father of more than two pups, and mean RS for San Jorge and Los Islotes colonies was 0.71 and 0.21 respectively (Table 6.2).
Zalophus wollebaeki was studied by Porschmann et al. (2010) in the Galapagos islands. They obtained 53.7% of pup paternity assignments. They sampled all males and pups in the colony , thus RS estimation was straightforward: 0.41 (154/374) pups/male.
Overall, harem size produced an estimation of 20.1 pups per dominant male (Table 6.1), while territorial males fathered only 32.5% of pups born in colonies, giving an estimation of 2.0 pups per territorial male (Table 6.2). These results indicated that behavioural estimations of reproductive success in Otariids are more than tenfold higher than genetic ones.
4 Does Theory Meet Facts?
The most obvious and characteristic feature of Otariids is the remarkable sexual dimorphism in size. The scene of two territorial males fighting in a colony saturated with females is spectacular and fascinates anyone interested in animal behaviour. It seems evident that the size and aggressiveness of these males serve for intra-sexual competition for access to reproductive partners. Added to these conspicuous features, Otariids fit in the theory, which explains the evolution of mating systems by sexual selection: environmental potential for polygyny is high because most species display female spatial clustering during a synchronous birthing season, giving some males the opportunity to gather numerous matings in a short time.
However, this review indicates that, in the face of paternity data, the main assumptions of sexual selection theory applied to explain the evolution of mating systems of Otariids are not met for several studies and species. Variance in the reproductive success of males is substantially lower than what was believed when the data to measure it was behavioural (operational sex ratio, copulations, harem size). A territorial male can be surrounded by females and even copulate with a large number of them, but that does not mean that he will sire many offspring. Also, there are numerous offspring whose paternities were not identified, which are probably the result of alternative strategies used by satellite males.
In summary, although only seven species have been surveyed, a preliminary conclusion is that research on genetic paternity challenges traditional theory on the evolution of mating systems in Otariids, as it did in studies of birds. Unexpected high levels of extra-harem paternities were found, similarly to the extra-pair paternities discovered in birds. However the effect in terms of male reproductive variance of these extra-paternities is exactly the opposite: while in birds this variance is higher than expected by behaviour, in Otariids it was significantly lower. If variance is low, and there are no large differences in the breeding success of dominant and satellite males, then the intensity of sexual selection would be considerably less than it was assumed in traditional models.
Larger males appear to be the most successful in maintaining a position within the colony (Lidgard et al. 2005; Lindenfors et al. 2002; Meise et al. 2014). This pattern has been interpreted as the result of sexual selection operating on large males that provides success in agonistic interactions, dominance and efficient territorial defence (Lindenfors et al. 2002). Porschmann et al. (2010) postulated an alternative hypothesis. They proposed that fasting ability selects for male size rather than just fighting ability and dominant status. Larger males have greater fat reserves, allowing them to extend tenure without the need to go to the sea for food. Large attendance alone would be enough to access to more estrus females.
Cassini (2017, Chap. 6) proposed that the size of Otariid males is determined primarily by natural selection, due to the advantages it could bring to foraging in cold seas. This hypothesis complements that proposed by Porschmann et al. (2010) because the largest males can access the best foraging areas (or any other advantage that body size could provide during foraging), they may build more fat reserves, so they would be better prepared to follow a strategy based on a long attendance at a site where females aggregate. On the other hand, small males cannot stay long in the colony , as they are forced to go foraging.
This natural selection hypothesis produces the same prediction that sexual selection hypothesis: large males will use the territorial strategy, and the small males will use alternative strategies. The most important difference between the two hypotheses is that natural selection does not need to assume that ‘dominant’ males will have greater fitness per reproductive season than satellite males. This hypothesis interprets territorial defence as an opportunistic strategy of males with large reserves of fat, and also as a secondary evolutionary product that originates from the advantages that large body size provides for survival at sea.
Krüger et al. (2014) found support to the hypothesis that male body size may have evolved by natural selection pressures. They analyzed 11 life-history traits in 35 pinniped species to determine their co-evolutionary dynamics and inferred their most likely evolutionary trajectories in the evolution of this taxon. They found that sexual size dimorphism had evolved prior to changes in the mating system. Only subsequently did polygyny evolve, leading to further coevolution as the strength of sexual selection intensified. They called for the inclusion of ecological variables when studying sexual selection and argue for caution when assuming causality between coevolving traits. They proposed that differences in body sizes between sexes could be the consequence of niche partitioning during aquatic foraging, or in combination with sexual selection on males to enforce copulations on females. Assuming that natural selection pressures are a determinant of body size, a question remains as to why females are smaller than males. There are at least four hypotheses that explain the origin of sexual size dimorphism by processes different to sexual selection (for an explanation, see Chap. 7): (1) non-adaptive processes (Leutenegger and Cheverud, 1982, 1985), (2) inter-sexual competition, (3) bimodal niches, and (4) dimorphic niches (Slatkin 1984; Cassini 2017).
Several alternative strategies have been described in Otariids. Some of them were not observed but inferred after discovering the significant occurrence of extra-harem paternities. In A. australis , satellite males stayed intermittently in a fraction of the study area farthest from the coastline that was partially used by females and pups whenever the tide level rose (Franco-Trecu et al. 2014). In O. flavescens , non-territorial males group raid territories and breeding aggregations of females, or they try stop females entering the colony from the sea (Campagna et al. 1988, Chap. 9). In A. gazella, Gemmell et al. (2001) suggested the existence of aquatic or nocturnal mating by satellite males. In Z. wollebaeki , a fraction of males seems to be successful by paying only short visits to the colony (Porschmann et al. 2010). In Z. californianus , Flatz et al. (2012) suggested that copulations may be occurring outside the boundaries of the colonies, such that some males intercept females traveling to and from the rookery on foraging trips. In Z. wallobaeki, Meise et al. (2014) propose that intermediate-sized males can hide among females when the territorial male is close by, thus avoiding being spotted and chased away.
Research has focused on territorial males, while the study of satellite males has been notably less intense. There are very few studies measuring the reproductive success of alternative strategies, probably due to two reasons: more theoretical interest in the dominant strategy, and more difficulty in studying satellite males. It has been proposed that satellite males would incur lower energetic costs from male–male interactions, thermoregulation, and site tenure than territorial males, thus being potentially able to invest in post-copulatory traits by augmenting both the quantity and quality of their sperm and seminal fluids (Simmons and Fitzpatrick 2012; Franco-Trecu et al. 2014). A possible aspect of low variance in male reproductive success that could be studied in the future relates to the effectiveness of fertilization. Although the frequency of copulations in dominant males seems to be high, fertilizations are lower than expected. Could the cause be in the low production of sperm by the large males? It maybe that smaller males ultimately may be more successful in postcopulatory competition.
5 A Mixed Model of the Evolution of Sexual Size Dimorphism and Mating Systems in Mammals
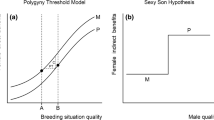
Recent studies that estimates intensity of sexual selection using molecular measures of male reproductive success have challenged the hypothesis that sexual selection is the only selective pressure involved in the evolution of mammalian mating system. Taxa that show higher levels of polygyny and dimorphism (and that have been used as biological models of the theory of sexual selection) have shown lower variances in male fitness than expected by behavior or dimorphism (in pinnipeds this chapter and Gonzalez-Suarez and Cassini (2014), in primates Cassini (2020a), in artiodactyls, Cassini (2020b)). Cassini (2020c) developed a theoretical model to explain these differences between what was obtained by ethological studies and what was obtained with molecular techniques (Fig. 6.2). The mixed model of the evolution of sexual size dimorphism and mating systems in mammals proposes an initial phase where natural selection operates on body size, followed by increases in sexual dimorphism (Fig. 6.3). Ecological factors and male disturbance promote female grouping. Males compete for access to these groups according to two possible evolutionary scenarios comprising directional or equilibrium sexual selection, where both produce a similar type of behavioral polygyny, but they differ in terms of the intensity of intra-male pre-copulatory sexual selection.

Relationship between genetic estimation (average paternity of territorial males) and behavioural estimation (harem size) of RS

(a) Sequence of evolutionary events postulated by the classical sexual selection model of the evolution of polygyny and sexual size dimorphism in mammals. (b) Mixed model of the evolution of sexual size dimorphism and mating systems in mammals (described in text)
6 Final Remarks
The theory of sexual selection applied to mating systems has been previously challenged by new discoveries originating in molecular techniques. The first major challenge occurred with monogamous species, when it was discovered that many animals that form long-term pair bonds produce extra-pair offspring as a norm. Studies employing molecular markers have demonstrated that in many cases levels of extra-pair parentage cannot accurately be estimated from behavioural observations alone (Hughes 1998). These results stimulated research in the field of mate selection by females, which became one of the most fruitful fields within behavioural ecology research during the late twentieth century.
At present, we are facing a similar phenomenon regarding sexual selection in polygynous mating systems. In this study, I reviewed published studies on genetic paternity in Otariids and concluded that classical theories on the evolution of polygyny in mammals should be revised in light of the new results provided by molecular ecology, specifically the high level of extra-harem paternity . I proposed a new approach that relativizes the role of sexual selection in favour of a more holistic approach, in which the influence of different forms of selection is evaluated. Efforts should be made to increase the number of studies on genetic paternity , and on the behaviour of adults of both sexes who move between areas of a colony and between colonies.
References
Ahonen H (2013) Population structure and mating system of the Australian sea lion (Neophoca cinerea). Neurons and Cognition [q-bio.NC]. Université Paris Sud - Paris XI
Bartholomew GA (1970) A model for the evolution of pinniped polygyny. Evolution 24:546–559
Bartlett JMS, Stirling D (2003) A short history of the polymerase chain reaction. Methods Mol Biol 226:3–6
Berta A (2018) Pinnipeds. In: Würsig B, Thewissen JGM, Kovacs K (eds) Encyclopedia of marine mammals. Academic, New York, pp 734–740
Bonnes DJ (1991) Determinants of mating systems in the Otariidae (Pinnipedia). In: Renouf D (ed) The behaviour of Pinnipeds. Chapman & Hall, New York, pp 1–44
Campagna C, Le Boeuf BJ (1988) Reproductive behaviour of southern sea lions. Behaviour 104:233–260
Campagna C, Le Boeuf BJ, Cappozzo HL (1988) Group raids: a mating strategy of male southern sea lions. Behaviour 105:224–249
Cassini MH (1999) The evolution of reproductive systems in pinnipeds. Behav Ecol 10(5):612–616
Cassini MH (2017) Role of fecundity selection on the evolution of sexual size dimorphism in mammals. Anim Behav 128:1–4
Cassini MH (2020a) A mixed model of the evolution of polygyny and sexual size dimorphism in mammals. Mammal Rev. https://doi.org/10.1111/mam.12171
Cassini MH (2020b) Sexual size dimorphism and sexual selection in primates. Mammal Rev. https://doi.org/10.1111/mam.12191
Cassini MH (2020c) Sexual size dimorphism and sexual selection in Artiodactyles. Behav Ecol. https://doi.org/10.1093/beheco/araa017
Caudron AK, Negro SS, Fowler M, Boren L, Poncin P, Robertson BC, Gemmell NJ (2010) Alternative mating tactics in the New Zealand fur seal (Arctocephalus forsteri): when non-territorial males are successful too. Aust J Zool 57(6):409–421
Darwin C (1871) The descent of man, and selection in relation to sex. Wiley, London
Emlen ST, Oring LW (1977) Ecology, sexual selection and the evolution of mating systems. Science 197:215–223
Flatz R, González-Suárez M, Young JK, Hernández-Camacho CJ, Immel AJ, Gerber LR (2012) Weak polygyny in California Sea lions and the potential for alternative mating tactics. PLoS One 7:e33654
Franco-Trecu V, Costa P, Schramm Y, Tassino B, Inchausti P (2014) Sex on the rocks: reproductive tactics and breeding success of South American fur seal males. Behav Ecol 25(6):1513–1523
Gemmell NJ, Burg TM, Boyd IL, Amos W (2001) Low RS in territorial male Antarctic fur seals (Arctocephalus gazella) suggests the existence of alternative mating strategies. Mol Ecol 10:451–460
Gonzalez-Suarez M, Cassini MH (2014) Variance in male RS and sexual size dimorphism in pinnipeds: testing an assumption of sexual selection theory. Mammal Rev 44:88–93
Griffith SC (2007) The evolution of infidelity in socially monogamous passerines: neglected components of direct and indirect selection. Am Nat 169(2):274–281
Hoffman JI, Boyd IL, Amos W (2003) Male reproductive strategy and the importance of maternal status in the Antarctic fur seal Arctocephalus gazella. Evolution 57:1917–1930
Hughes C (1998) Integrating molecular techniques with field methods in studies of social behavior: a revolution results. Ecology 79:383–399
Jones AG, Ratterman NL (2009) Mate choice and sexual selection: what have we learned since Darwin? Proc Natl Acad Sci 106(Suppl 1):10001–10008. https://doi.org/10.1073/pnas.0901129106
Kiyota M, Insley SJ, Lance S (2008) Effectiveness of territorial polygyny and alternative mating strategies in northern fur seals, Callorhinus ursinus. Behav Ecol Sociobiol 62:739–746
Kokko H, Jennions MD, Houde A (2007) Evolution of frequency-dependent mate choice: keeping up with fashion trends. Proc R Soc B Biol Sci 274(1615):1317–1324
Kotiaho JS, Puurtinen M (2007) Mate choice for indirect genetic benefits: scrutiny of the current paradigm. Funct Ecol 21(4):638–644
Krüger O, Wolf JBW, Jonker RM, Hoffmann JI, Trillmich F (2014) Disentangling the contribution of sexual selection and ecology to the evolution of size dimorphism in pinnipeds. Evolution. https://doi.org/10.1111/evo.12370
Lack D (1968) Ecological adaptations for breeding in Birds. Methuen, London
Leutenegger W, Cheverud J (1982) Correlates of sexual dimorphism in primates: ecological and size variables. Int J Primatol 3:387–402
Leutenegger W, Cheverud J (1985) Sexual dimorphism in Primates. The effects of size. In: Jungers WL (ed) Size and scaling in primate biology. Plenum, New York, pp 33–60
Lidgard DC, Boness OJ, Bowen WD, McMillan JI (2005) Statedependent male mating tactics in the grey seal: the importance of body size. Behav Ecol 16:541–549
Lindenfors P, Tullberg BS, Biuw M (2002) Phylogenetic analyses of sexual selection and sexual size dimorphism in pinnipeds. Behav Ecol Sociobiol 52:188–193
Mead LS, Arnold SJ (2004) Quantitative genetic models of sexual selection. Trends Ecol Evol 19(5):264–271
Meise K, Piedrahita P, Kruger O, Trillmich F (2014) Being on time: size-dependent attendance patterns affect male RS. Anim Behav 93:77–86. https://doi.org/10.1016/j.anbehav.2014.04.019
Mesnik SL, Ralls K (2018) Mating systems. In: Würsig B, Thewissen JGM, Kovacs K (eds) Encyclopedia of marine mammals. Academic, New York, pp 712–719
Miller EH (2018) Territorial behavior. In: Würsig B, Thewissen JGM, Kovacs K (eds) Encyclopedia of marine mammals. Academic, New York, pp 1156–1166
Porschmann U, Trillmich F, Müller B, Wolf JBW (2010) Male RS and its behavioural correlates in a polygynous mammal, the Galapagos Sea lion (Zalophus wollebaeki). Mol Ecol 19:2574–2586
Sheldon BC, Ellegren H (1999) Sexual selection resulting from extrapair paternity in collared flycatchers. Anim Behav 57(2):285–298
Simmons LW, Fitzpatrick JL (2012) Sperm wars and the evolution of male fertility. Reproduction 144(5):519
Slatkin M (1984) Ecological causes of sexual dimorphism. Evolution 38:622–630
Soto KH, Trites AW (2011) South American sea lions in Peru have a lek-like mating system. Mar Mamm Sci 27:306–333
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual selection and the descent of man, 1871–1971. Aldine-Atherton, Chicago, pp 136–179
Wade MJ, Arnold SJ (1980) The intensity of sexual selection in relation to male sexual behavior, female choice, and sperm precedence. Anim Behav 28:446–461
Whittingham LA, Dunn PO (2005) Effects of extra-pair and within-pair reproductive success on the opportunity for selection in birds. Behav Ecol 16(1):138–144
Acknowledgements
I thank Claudio Campagna for the invitation that this work originated from. MHC is a researcher of the Argentinean Research Council CONICET.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Cassini, M.H. (2021). Polygyny in the Era of Molecular Biology: Revisiting Bartholomew’s Model. In: Campagna, C., Harcourt, R. (eds) Ethology and Behavioral Ecology of Otariids and the Odobenid. Ethology and Behavioral Ecology of Marine Mammals. Springer, Cham. https://doi.org/10.1007/978-3-030-59184-7_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-59184-7_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-59183-0
Online ISBN: 978-3-030-59184-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)