
Abstract
Evapotranspiration (ET) is a key component in global water and energy cycles. This chapter presents and discusses recent research advances about ET over northern regions and watersheds. ET in northern regions tends to increase with the decrease of latitude. The largest ET typically appears in the forest ecosystem, while the grasslands and shrublands have small ET. While the seasonal variations in ET are usually high, the interannual variability in annual ET is usually low over the Arctic regions. Sublimation from snow cover accounts for about 15–25% of winter precipitation. Many factors, such as soil moisture, vegetation type and productivity, and ecosystem features affect ET over northern regions. In addition, precipitation plays a key role in impacting ET. ET is more sensitive to precipitation in the early growing season than in the late growing season. Furthermore, changes in freeze–thaw processes due to warming also affect land surface conditions and the ET processes. During 1983–2005, ET increased significantly in the Arctic region with a rate of 3.8 mm decade-1 because of regional warming and vegetation greening. Such an increase in ET may exert significant impacts on the regional hydrology and water resources. Advanced models can simulate past ET change over the large northern watersheds. Remote sensing has provided new ET data and information that support climate and hydrology research and applications. There is a key question: Will Arctic landscapes become wetter or drier as climate changes? According to global models and data analyses, annual ET has increased over the northern regions. In the future, summer PE is projected to decrease much of Canada, increase over Alaska, decrease over the western and northern Eurasian subarctic, and increase over parts of northeastern Russia. Over most of these areas, the sign of the projected change is not robust across the models at the 95% confidence level. Many factors contribute to the uncertainty in the projected changes in Arctic surface wetness. There is certainly a need to better quantify and narrow the uncertainties in global models in the northern regions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

1 Introduction
Evapotranspiration (ET) refers to the amount of water vapor evaporated from the unit area of the land surface during a unit of time and consists of evaporation from canopy-intercepted water, evaporation from soil (E), and transpiration from plants (T). ET is a key component in global water and energy cycles. On average, more than 60% of global land precipitation is returned to the atmosphere through terrestrial ET (Oki and Kanae 2006). ET as latent heat flux consumes roughly 50% of the solar radiation absorbed by the Earth’s surface (Trenberth et al. 2009). ET affects climate through a wide range of feedbacks to air temperature, humidity, and precipitation (Shen et al. 2015; Shukla and Mintz 1982; Zeng et al. 2017). Accurate estimation of ET is therefore fundamental to not only elucidating how the hydrological cycle responds to climate change but also to regional drought monitoring and water resources management (Fisher et al. 2017; Ma et al. 2019). However, investigating ET is challenging because of its complex interactions across the soil–vegetation–atmosphere interface (Katul et al. 2012; Matheny et al. 2014; Zhang et al. 2014). Historically, ET was related (via linear or nonlinear scaling with soil moisture) to direct measurements of its conceived maximum value, i.e., pan evaporation (Brutsaert 2013). However, neither water-limited nor energy-limited land surface ET acts as pan evaporation (Brutsaert 1982). With recent advances in instrumentations and data storage, a better understanding of ET from various land covers has been reported through the use of in situ flux observations (e.g., Baldocchi et al. 2004; Fischer et al. 2013; Ma et al. 2014; Mackay et al. 2007; Wilson and Baldocchi 2000). These studies have significantly contributed to our knowledge of regional hydrological regimes and their complex feedback mechanisms between land and atmosphere (Baldocchi 2014).
Because transpiration from plants (T) links the water and carbon cycles, it is used to calculate carbon assimilation by terrestrial vegetation, so estimating T fluxes is a major focus in climate and ecology studies (Evaristo et al. 2015; Jasechko et al. 2013; Kool et al. 2014; Wei et al. 2017). As T process directly correlates with plant growth and the carbon cycle (Scott et al. 2006), quantitative estimation of T in the total evapotranspiration (T/ET) has long been acknowledged to play a crucial role in water resource management, yield estimation, water cycle, and climate change, from plot scale to global scale (Schlesinger and Jasechko 2014; Scott and Biederman 2017; Xiao et al. 2018). There is, however, a considerable discrepancy among global T/ET estimations by different methods. For example, the results of a combination of wide ranging, remotely sensed observations showed that approximately 80% of the annual land ET is attributed to T (Miralles et al. 2011). Wei et al. (2017) quantified the global T/ET with a leaf area index (LAI)-based ET partitioning algorithm and concluded that T accounts for 57% of ET. The distinct isotope effects of T and E based on the isotopic analysis of a global dataset of large lakes and rivers showed that T represents 80–90% of terrestrial ET (Jasechko et al. 2013), although this estimate was challenged by Coendersgerrits et al. (2014). The results of isotope mass budget-based simulations suggested that the transpired fraction of ET accounts for approximately 60% of the annual land ET (Good et al. 2015). There are also great challenges in the state-of-the-art land surface models and remote sensing models in representing the ratio of T to ET. Maxwell and Condon (2016) argued that partitioning ET is connected to water table depth and they found that including lateral groundwater flow in the model increases transpiration partitioning from 47 ± 13 to 62 ± 12%. This aspect was recently also acknowledged in Chang et al. (2018), in which terrain-driven lateral water flows spread out soil moisture to a wider range along hill slopes with an optimum subrange from the middle to upper slopes, where soil evaporation was more suppressed by the drier surface than T due to plant uptake of deep soil water, thereby enhancing T/ET. In terms of remote sensing models, large errors in representing the components of ET also exist in Moderate Resolution Imaging Spectroradiometer (PM-MODIS), the Priestley–Taylor Jet Propulsion Laboratory model (PT-JPL), and the Global Land Evaporation Amsterdam Model (GLEAM), which shows root-mean-square-error of 90–114% for soil evaporation and 54–114% for transpiration (Talsma et al. 2018).
This chapter reviews recent research and results of ET from various land surfaces (including different vegetation types) and also across large regions and watersheds. Specifically, it discusses ET process, pattern, and variabilities over space and time, such as regional/basin ET, its change, and impact to water balance. It also demonstrates model estimates/simulation of large-scale ET over the arctic domain and selected watersheds, highlights remote sensing development in ET estimation, and global model analysis of the net moisture flux (P-E) and its change across the northern regions.
2 ET Distribution and Variability
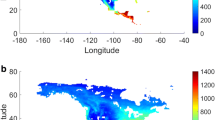
Atmospheric conditions, such as wind speed and saturation deficit, are dominant factors in determining ET variability. Zhang et al. (2003) summarized that daily mean evaporation over the Tibetan Plateau varied within the range of 0.3–3.5 mm on the permafrost surface, and regional differences in evaporation were strongly related to surface soil moisture. Locally, topography and its influence on surface soil moisture was found to control evaporation systematically. The seasonality of evaporation in permafrost regions is dominated by freeze–thaw cycles at the surface; evaporation from the melting permafrost surface is up to 4–7 times greater than that from the frozen ground. In forested terrain, the interception of precipitation can reduce daily evaporation by 60–70%. Sublimation from the snow surface observed at Tianshan Mountains and eastern Tibetan Plateau was in the range of 0.2–1.0 mm d−1. Mu et al. (2009) investigated the spatial pattern of evapotranspiration for the pan-Arctic domain and found large annual ET variability during 2000–2006 among the regional biomes. The largest annual ET rates occur over forests, while the lowest rates occur over grasslands and scrublands; annual ET rates for savanna and cropland areas are generally intermediate. Ecosystem processes in high-latitude boreal and tundra biomes are strongly constrained by low solar irradiance and freezing temperatures for much of the year, so that seasonal patterns in plant photosynthesis (GPP) and ET correspond closely and are generally confined to a relatively narrow growing season. The interannual variability in estimated annual ET is relatively low although the seasonal variation is high over the Arctic, which reflects the dominance of cold temperature constraints on boreal Arctic ecosystem processes (Fig. 4.1).

Spatial pattern of mean annual and seasonal ET during 2000–2006 for the pan-Arctic domain (Mu et al. 2009)
Variation in evapotranspiration is influenced by all ecosystem parameters and processes, such as soil moisture content, vegetation productivity, and ecosystem nutrient and water budgets. The partitioning of available energy into evapotranspiration (latent heat flux) and sensible heat flux at a vegetation surface also affects aspects of weather and climate. As ET consists of soil evaporation, and transpiration from plants, the soil moisture, and biological feature are key factors to determine ET variation. Zhang et al (2005) demonstrated that E is more sensitive to surface soil moisture than T. The former takes water from a very thin soil layer and moves it to the atmosphere; the latter transfers water from the soil through root–stalk–leaf mechanisms and may take water from a thicker soil layer. Therefore, T is anticipated to be more independent of soil moisture variations than E. Figure 4.2 shows the relationship between E/Ep (Ep is potential evaporation) and the surface soil moisture. E/Ep increased almost linearly with ground surface moisture when the volumetric water content was less than 30% but varied little when moisture content beyond that level. This suggests that soil evaporation was constrained by the deficiency of available water when soil moisture was less than the critical value of 30%, illustrating the primary controls of soil water status on the soil evaporation.

Relationships between (left) E/Ep and surface moisture and (right) leaf conductance (Etrans/D * Pa, mms-2) and air temperature. D, leaf-to-air specific deficit; Pa, air density (After Zhang et al. (2005) for sparse grassland in northeastern Mongolia)
Transpiration rates are determined by two parameters: leaf conductance and leaf-to-air specific deficit (D). While it is impossible to present leaf conductance because of no CO2 concentration data were collected, the right panel of Fig. 4.2 shows the ratio of Etrans/D * Pa (D is the leaf-to-air specific deficit and Pa is air density), which implies a leaf conductance value, plotted against air temperature. The plots show that transpiration increased as temperatures increased. When temperatures became warm, higher leaf conductance allowed water uptake if soil moisture was low but higher than wilting point (2.8–4.1% in the root zone). Lower temperatures, lower D, and higher humidity accompany precipitation events, so plant transpiration decreased as soil evaporation approached to potential evapotranspiration (Fig. 4.2).
Projected increases in air temperature and precipitation due to climate change in Arctic wetlands could dramatically affect ecosystem function. As a consequence, it is important to define controls on evapotranspiration, i.e., the major pathway of water loss from the system. Liljedahl et al. (2011) quantified the multi-year controls on midday Arctic coastal wetland evapotranspiration, measured with the eddy covariance method at two vegetated, drained thaw lake basins near Barrow, Alaska. Variations in near-surface soil moisture and atmospheric vapor pressure deficits were found to have nonlinear effects on midday evapotranspiration rates. Data collections near Barrow over 3 years showed that vapor pressure deficits (VPD) near 0.3 kPa appeared to be an important hydrological threshold, allowing latent heat flux to persistently exceed sensible heat flux (Fig. 4.3). Dry (compared to wet) soils increased bulk surface resistance (water-limited). Wet soils favored ground heat flux and therefore limited the energy available to sensible and latent heat flux (energy-limited). Thus, midday evapotranspiration was suppressed from both dry and wet soils but through different mechanisms. They also found that wet soils (ponding excluded) combined with large VPD, resulted in an increased bulk surface resistance and therefore suppressing evapotranspiration below its potential rate (Priestley-Taylor coefficient < 1.26). This was likely caused by the limited ability of mosses to transfer moisture during large atmospheric demands. Ultimately, in addition to net radiation, the various controlling factors on midday evapotranspiration (i.e., near-surface soil moisture, atmospheric vapor pressure, and the limited ability of saturated mosses to transfer water during high VPD) resulted in an average evapotranspiration rate of up to 75% of the potential evapotranspiration rate. These multiple limitations on midday evapotranspiration rates have the potential to moderate interannual variation of total evapotranspiration and reduce excessive water loss in a warmer climate. Combined with the prevailing maritime winds and projected increases in precipitation, these curbing mechanisms will likely prevent extensive future soil drying and hence maintain the presence of coastal wetlands.

The relationship between mean hourly air vapor pressure deficit (VPD) and a Bowen ratio (β) or b Priestley–Taylor α during differing soil moisture conditions at the BE site, 2006–2008. Dry soils represent a soil water potential <−0.13 MPa at 10 cm depth. The vertical dashed lines represent the identified critical value of VPD. VPDs above this threshold resulted in a <1 and a Priestley–Taylor near or above 1. The identified VPD thresholds were 0.25 (2006), 0.31 (2007), and 0.28 kPa (2008) for wet soils and 1.19 kPa for dry soils (2007). (© Liljedahl et al. 2011, distributed under the Creative Commons Attribution 3.0 License. use with permission from the authors)
Evapotranspiration dominates hydrological processes on the Arctic Coastal Plain for a couple of months after snowmelt until soil moisture declines (Kane et al. 2000, 2008). The majority of studies on ET in the Arctic tundra focus on whole-ecosystem fluxes, with rates of approximately 1–3 mm day−1 (Liljedahl et al. 2011; Mendez et al. 1998). However, a whole-ecosystem approach to determining ET rates does not allow for quantifying the variability in fluxes associated with the heterogeneous landscape (Oren et al. 2006), particularly on the Arctic Coastal Plain (Oechel et al. 1998). Further, a whole-ecosystem approach does not allow for partitioning ET into its components of evaporation and transpiration. Spatial heterogeneity in soil moisture, soil temperature, and plant composition likely affect how ET is partitioned into evaporation and transpiration in the Arctic Coastal Plain (Oberbauer and Dawson 1992). It is critical to understand the partitioning of evapotranspiration because environmental processes control evaporation and transpiration differently (Jasechko et al. 2013). While both respond to surface energy, atmospheric demand, and soil water availability (Betts et al. 1999; Calder 1998), evaporation is a physical process but transpiration is a plant physiological process controlled by stomata (Wullschleger et al. 1998).
On the tundra surface near Barrow, Alaska, Raz-Yaseef et al. (2017) found that variation in environmental conditions and plant community composition, driven by micro-topographical features, has a significant influence on ET. ET had high variability across the field site, i.e., the fluxes were highest over mosses and open water, lower from grasses and sedges (65% of those from mosses and open water), and lowest over bare ground and lichens (50% of those from mosses and open water) (Fig. 4.4).

Two years ET measured with the portable chamber. Values are averages for each plant type and for open water; bars denote standard deviation (After Raz-Yaseef et al. 2017)
Among plant types, ET from moss and inundated areas was more than twice that from other plant types. ET from troughs and low polygonal centers was significantly higher than that from high polygonal centers. ET also varied seasonally, with peak fluxes of 0.14 mm h−1 in July. Diurnal fluctuations in incoming solar radiation and plant processes produced a diurnal cycle in ET. Observed patterns with projections for the impact of permafrost degradation on polygonal structure suggest that micro-topographic changes associated with permafrost thaw have the potential to alter the tundra ecosystem ET (Young-Robertson et al. 2018). Yuan et al. (2010) examined the impacts of precipitation seasonality and ecosystem types on ET quantified by eddy covariance towers from 2002 to 2004 in three ecosystems (grassland, deciduous broadleaf forest, and evergreen needle leaf forest) in the Yukon River Basin, Alaska. The annual precipitation changed greatly in both magnitude and seasonal distribution through the three investigated years. Observations and model results showed that ET was more sensitive to precipitation scarcity in the early growing season than in the late growing season, which was the direct result of different responses of ET components to precipitation in different seasons. The results demonstrated the importance of seasonal variations of precipitation in regulating annual ET and overshadowing the function of annual precipitation. Comparison of ET among ecosystems over the growing season indicated that ET was largest in deciduous broadleaf, intermediate in evergreen needle leaf, and lowest in the grassland ecosystem. These ecosystem differences in ET were related to differences in successional stages and physiological responses.
Sublimation from snow surface has been identified as an important hydrological process at high altitudes and in high-latitude regions, involving complex mass and energy exchanges. In the Colorado Frontal Ranges, measurements from snow evaporation pans indicated that net sublimation for the 5-month winter period from December to April was 135 mm (Meiman and Grant 1974). Berg (1986) estimated sublimation losses from snow cover to be 30–51% of precipitation over a 2-year period from 1973 to 1975. Kattelmann and Elder (1991) estimated sublimation from snow to be 18% of total precipitation over 2 years in the Sierra Nevada. In the subarctic region, several studies have shown that sublimation from snow cover is a non-negligible hydrological component that also affects river discharge and regional water resources. In western Canada, sublimation from snow during the winter season consumed from 15 to 40% of seasonal snowfall (Woo et al. 2000) and 12 to 33% of annual snowfall (Pomeroy and Li 1997). Suzuki et al. (2002) estimated that sublimation from snow cover in eastern Siberia was significant at 25.6% of precipitation from October to April. However, these estimations were derived from modeling and lack of observational verification. Subarctic ground surfaces are dominantly covered by subalpine and boreal forests. The impact of forests on snow cover has been extensively investigated via accumulation and melting processes. An increase of 30–45% in seasonal snow accumulation was measured after the removal of the evergreen forest (Pomeroy and Gray 1995; Pomeroy and Li 1997). Pomeroy et al. (1998) also found that snow water equivalent (SWE) generally increases with evergreen canopy density in boreal forests. Simulation models for snowmelt under a forest canopy have been developed to examine the relationship between snowmelt and forest density (Barry et al. 1990; Yamazaki and Kondo 1992; Wigmosta et al. 1994). Canopy density is important in controlling snow ablation timing and rates because tree height and canopy properties control the transmission of solar radiation (Davis et al. 1997; Ni et al. 1997).
Leydecker and Melack (1999) discussed the sensitivity of sublimation over a short time scale using snow surface roughness, instrument height, and wind speed and demonstrated that wind speed was the critical variable for determining sublimation and that doubling wind speed can triple the sublimation. Zhang et al. (2008) demonstrated that when the wind speed is less than 2. 0 m s−1, the saturation deficiency is predominant in determining sublimation, and sublimation increases significantly when the wind speed is above 2.0 m s−1. The variation in vapor transfer coefficient versus wind speed can be deduced (Fig. 4.5). This result shows that the vapor transfer coefficient for the snow surface is larger under low wind and decreases sharply as wind speed increases. However, the vapor transfer coefficient is nearly constant when the monthly mean wind speed is above 2.0 m s−1. The impact of the coefficient on the sublimation rate is not so clear. Daily sublimation shows a tendency toward correlation with the vapor transfer coefficient (Fig. 4.5). The values of 20.3–21.6% of total snowfall lost to sublimation compares favorably with snow cover lost due to sublimation of 25.6% in eastern Siberia (Suzuki et al. 2002), or 12–33% of annual snowfall in Canada (Pomeroy and Li 1997).

Vapor transfer coefficient versus wind speed (left) and daily sublimation versus the vapor transfer coefficient (right) (Zhang et al. 2008)
Snow cover models were used to quantify sublimation. Sexstone et al. (2018) demonstrated that snow sublimation rates corresponding to climate-warming simulations remained unchanged or slightly increased but total sublimation losses decreased by up to 6% because of a reduction in snow-covered area and duration. Seasonally snow-covered forests in western North America have experienced substantial disturbance from mountain pine beetle (Dendroctonus ponderosae) and spruce beetle (Dendroctonus rufipennis) outbreaks (Potter and Conkling 2016), which have resulted in widespread tree mortality and thus changes to forest structure that are particularly relevant to canopy and surface sublimation processes. Field studies that have focused on measuring SWE in both unimpacted and disturbed forests have inferred decreasing (e.g., Boon 2012; Pugh and Small 2012) as well as steady or increasing (e.g., Biederman et al. 2014; Harpold et al. 2014) net sublimation fluxes in the presence of disturbance. In contrast, distributed watershed modeling studies considering beetle-induced forest mortality (Penn et al. 2016) have generally reported decreased evapotranspiration but have not specifically focused on the sublimation component of evapotranspiration. Although changes to snow accumulation and melt processes from climate warming have been studied widely (Musselman et al. 2017a, b), the response of sublimation to climate change has received little investigation. As a result, specific knowledge gaps remain including how the components of sublimation will individually and collectively respond to changes in forcing mechanisms, in addition to changing snow accumulation and melt dynamics. Process-based snow models that can integrate responses and feedbacks of the snow energy balance offer the ability to evaluate sublimation responses to these changing land cover and climate conditions, which is critically important for the understanding of the water balance in snow-dominated regions.
3 Climate Warming and ET Change
The terrestrial cryosphere covers approximately 66 million km2 (~52.5%) of the global land area where water is either permanently or seasonally frozen. Each year, most of the terrestrial cryosphere undergoes a seasonal transition between predominantly frozen and non-frozen (i.e., thawed) landscape conditions; these transitional events are relatively abrupt and analogous to a biospheric and hydrological on/off switch between active ecosystem processes during the growing season and largely dormant conditions during the seasonal frozen period. The cold temperatures and seasonal frozen conditions pose strong constraints to vegetation productivity; surface hydrological processes, surface energy fluxes, and land–atmosphere trace gas exchange in these regions. In seasonally frozen environments, vegetation photosynthetic activity and associated ET are constrained by low temperatures and chemical unavailability of water as a result of being frozen.
Recent warming has altered freeze–thaw (FT) processes in the northern regions, resulting in substantial changes in terrestrial ecological and hydrological processes, including generally earlier onset and lengthening of potential vegetation growing seasons, thawing and degradation of permafrost, earlier spring snowmelt and increased groundwater to stream discharge from permafrost thawing. Warming and associated lengthening of the potential growing season has led to positive impacts on ecosystems of more cold-limited regions of the northern high latitudes, including increases in vegetation structure, greenness, productivity, and net CO2 exchange. Recent warming trends also have been associated with negative ecosystem impacts within more water-limited regions of the northern latitudes by increasing water stress, which has been attributed to be a major cause of recent vegetation browning and productivity reductions, higher tree mortality rates, and increasing fire frequency.
Seasonal transitions of the landscape between predominantly frozen and thawed conditions are analogous to a biospheric and hydrological on/off switch, with marked differences in ET, vegetation productivity, and other biological activity between largely dormant winter and active summer conditions. By analyzing independent FT and ET records derived from satellite remote sensing, Zhang et al. (2011) investigated changes in FT seasons and ET from 1983 to 2006 and their connections in the northern cryosphere. They found that annual ET shows predominantly negative correlations with the timing of primary seasonal thaw (Tthaw) but has positive correlations with the timing of primary seasonal freeze (Tfreeze) in most areas of the domain (Fig. 4.6). The correlations between annual ET and Tthaw are generally higher than that between ET and Tfreeze, indicating that the onset of the non-frozen season has a relatively larger impact on annual ET than its cessation. This is likely because incident solar radiation available for evaporation is generally higher during the spring Tthaw period than the autumn Tfreeze period; the Tthaw period also coincides with the general onset of the active growing season in northern, boreal, and Arctic environments, whereas vegetation photosynthetic activity and transpiration is reduced in autumn when plants are transitioning to winter dormancy (Kimball et al. 2004). Annual ET is positively correlated with the annual number of non-frozen days (DNF) over most of the domain, with stronger correlations at higher latitudes than in lower latitude areas.

Correlation maps a between annual ET and Tthaw, b between annual ET and Tfreeze and c between annual ET and DNF from 1983 to 2006; the insets show the frequency distributions of corresponding correlation coefficients, (Zhang et al. 2011)
In contrast to the prevailing pattern, some regions in the southern portion of the Arctic region show positive correlations between annual ET and Tthaw (Fig. 4.6a), where earlier than the normal onset of non-frozen period corresponds with less-than-normal annual ET. Almost all of these regions are located in areas where ET is predominantly limited by water supply. This suggests that an advancing non-frozen period may not promote additional increases in annual ET in these water supply-limited regions.
At northern high latitudes, cold temperatures and limited photoperiod largely constrain the potential growing season to a relatively brief period during the spring and summer months. Vegetation is largely dormant during the winter frozen period and coupled with frozen temperatures, seasonally low solar radiation loads, and limited day length strongly constrain ET. Low temperatures also inhibit plant photosynthesis and respiration by decreasing enzyme activity and protein synthesis in plant cells (Raich and Schlesinger 1992). Because transpiration is a consequence of plant photosynthesis, the constraint of low temperatures on photosynthesis also functions as a constraint on ET. Low temperatures also restrict photosynthesis and canopy gas exchange by limiting water supply and mobility in roots and xylem, leading to canopy stomatal closure (Woodward and Kelly 1997). Moreover, the atmospheric capacity to hold moisture increases exponentially with increasing temperature according to the well-known Clausius–Clapeyron relationship. Low temperatures, therefore, suppress ET by reducing atmospheric moisture demand. In addition, vegetation in cold boreal and Arctic regions are well adapted to the characteristic harsh environmental conditions and recover photosynthesis and respiration at relatively high rates following snowmelt and the new release of water in the landscape (Havranek and Tranquillini 1995; Suni et al. 2003; Kimball et al. 2004). Earlier onset of springtime thaw facilitates earlier photosynthesis and transpiration and generally enhances plant growth and transpiration when plant-available moisture is not limiting. During later stages of the non-frozen season in autumn, plants begin to enter the stage of senescence and dormancy, coinciding with seasonal reductions in temperatures, solar radiation, and photoperiod; thus, plant photosynthesis and transpiration are substantially reduced. After several months of vigorous ET, surface water storage accessible to vegetation would be substantially depleted without sufficient recharge. As a result, an extended non-frozen period in autumn would have less impact on ET than a similar extension in the spring.
The results of the positive response of ET to an advancing and lengthening non-frozen season vary spatially and are even reversed in warmer, southern regions of the domain where water supply is a leading constraint for ET (Zhang et al. 2011). These results also are consistent with recent reports indicating increasing regional water stress (Angert et al. 2005; Schindler and Donahue 2006; Hogg et al. 2008) and associated vegetation browning and productivity reduction (Goetz et al. 2005; Zhang et al. 2009; Beck et al. 2011), higher tree mortality rates (Kurz et al. 2008; Mantgem et al. 2009), and increasing fire frequency (Westerling et al. 2006).
4 Regional/Basin ET and Impact on Water Balance
ET plays an important role in linking the water, energy, and carbon cycles and represents over 60% of precipitation over the global land area (Oki and Kanae 2006). A better understanding of recent changes in the Arctic regions requires linking ET with other hydrological components including precipitation and river discharge. Precipitation and river discharge measurements are currently available from Arctic observation networks (Yang et al. 2005; McClelland et al. 2004). Evapotranspiration (ET) is highly heterogeneous both spatially and temporally due to strong vegetation canopy control on transpiration. Relatively sparse measurements of these variables across the northern high latitudes make an accurate assessment of ET a challenge. Remotely sensed data, especially from polar-orbiting satellites, provide relatively frequent and spatially contiguous monitoring of surface biophysical variables affecting ET, including albedo, biome type, and vegetation density. Satellite-based ET products have been produced at regional and global scales with varying accuracy (Suzuki et al. 2018; Zhang et al. 2009; Cleugh et al. 2007; Mu et al. 2007; Fisher et al. 2008).
Zhang et al. (2009) developed an evapotranspiration (ET) algorithm driven by satellite remote sensing inputs, including AVHRR GIMMS NDVI, MODIS land cover, and NASA/GEWEX solar radiation and albedo, and regionally corrected NCEP/NCAR Reanalysis daily surface meteorology. The algorithm was used to assess spatial patterns and temporal trends in ET over the pan-Arctic basin and Alaska from 1983 to 2005 (Fig. 4.7). The annual ET patterns are spatially complex but show generally reduced ET with increasing latitude. The spatial pattern of ET also corresponds to distributions of the major biome types. Boreal forest regions have the highest annual ET (277.35 ± 68.89 mm yr−1) among the three major biome types followed by grassland (248.40 ± 76.94 mm yr−1) and Arctic tundra (158.98 ± 37.61 mm yr−1) c water bodies resolved by the 1-km resolution global land cover classification cover only 6.4% of the pan-Arctic domain, they are distributed across a wide geographic range, with relatively large magnitude and spatial variation in annual ET (410.35 ± 142.58 mm yr−1) relative to vegetated land areas. Meanwhile, both the GPCP and GPCC sources show similar multiyear mean annual precipitation patterns; though the GPCP precipitation rates are much larger, averaging 1.31 (±0.38) times the GPCC precipitation rates. Both precipitation datasets show portions of Southern Alaska, Northeastern Canada near Hudson Bay, and Western Eurasia having the largest precipitation, with relatively arid polar tundra areas showing the least precipitation.

Maps of multi-year (1983–2005) mean annual calculated ET a and precipitation derived from b GPCP and c GPCC sources (Zhang et al. 2009)
The pan-Arctic region has a small positive trend in annual P for the 1983–2005 period (Fig. 4.8), indicated by both GPCP (7.4 mm decade-1) and GPCC (4.3 mm decade-1) sources, while ET shows a significant positive trend of 3.8 mm decade-1 for this period coinciding with regional warming (Trenberth et al. 2007) and vegetation greening trends (e.g., Goetz et al. 2005; Zhang et al. 2008). On average, ET represents 44.4% (±1.6) and 54.7% (±1.8) of P for the pan-Arctic domain relative to the GPCP and GPCC sources, respectively. Both P and ET show positive trends so the net effect on P–ET is reduced and annual P–ET shows an insignificant wetting trend indicated by both GPCP and GPCC data. Both GPCP- and GPCC-derived results show P–ET interannual variability of approximately ±6% relative to the long-term mean. Time series plots of annual P–ET anomalies for the domain show that the years 1984–1985, 1995–1996, 1998–2001, and 2003 are dry relative to the long-term mean. Both regional average P and ET show seasonal changes during the 23-year period (Table 4.1). The GPCP and GPCC data show positive regional P trends in spring, summer, and autumn, with slightly decreasing P trends in winter. The ET results show significant (P < 0.1) positive trends for all four seasons, with the largest ET increases in spring and summer, similar to P. These results suggest that regional warming and associated lengthening of the seasonal non-frozen period are promoting increases in annual ET, while positive P trends during this period are counteracting these evaporative water losses and associated changes to the annual water balance. On a seasonal basis, larger positive trends in P relative to ET in spring, summer, and autumn imply that the pan-Arctic domain is becoming wetter during the growing season. In contrast, decreasing P and slightly positive ET trends indicate that the pan-Arctic domain is becoming drier during the winter period.

Annual time series of precipitation (P) derived from GPCP and GPCC data sources; ET derived from the NDVI-based ET algorithm, and corresponding annual P–ET anomalies from the long-term (23-year) mean. Dashed lines show linear trends for the time series. The two P series from GPCP and GPCC sources are highly correlated (r = 0.91; p < 0.001). The resulting P–ET anomalies are also highly correlated (r = 0.89; p < 0.001). (Zhang et al. 2009)
Similar interannual changes were found by Suzuki et al. (2018) for the Mackenzie River, the Lena River, and the Yukon River basin. Comparisons were made among monthly precipitation (P) from GLDAS-2 and river runoff (R) for each river basin (Fig. 4.9). The maximum monthly P occurred in July for the Lena and Mackenzie River basins and in June or July for the Yukon River basin. The Lena River exhibits the largest amount of river runoff (R), followed by the Yukon River and the Mackenzie River. The peak value of R occurred in June for all three rivers. Using monthly climatology of river runoff data, the peak snowmelt occurs between April and June, accounting for approximately 40%, 36%, and 28% of the annual R in the Lena River, Yukon River, and Mackenzie River basins, respectively. E increased steadily from 2002 to 2016 with slopes of 1.2–4.1 mm y−1. The increase in E from GLDAS-2 dataset can explain most of the decrease in water storage. Thus, evapotranspiration driven by increasing summer temperatures may be the primary factor controlling water storage. This suggests that future warming might further decrease the water storage at high latitudes in the Arctic circumpolar region.

Temporal variations in monthly water balance components from 2002 to 2016 in the a Lena River basin, b Yukon River basin, and c Mackenzie River basin (Suzuki et al. 2018, open access, no special permission is required to reuse all or part of the article published by MDPI, including figures and tables)
Evapotranspiration of the major Arctic river basins simulated by a land surface model, CHANGE (Park et al. 2011), shows larger interannual variability, with large regional differences (Fig. 4.10), representing differences in climate, soil, vegetation, and landscape. Interestingly, the interannual variability of ET is larger in relatively warm basins (i.e., Ob and Mackenzie) than cold (Yenisey and Lena). A considerable area of the latter is underlaid by permafrost relative to the first. The recent warming temperature derives the warming of permafrost and thereby wetting soil that reduces moisture stress to ET. In other words, the wetted soil decreases the variability of ET under the warming temperature (Ohta et al. 2014). In reality, observations have identified the expansion of thermokarst lake resulted from permafrost degradation in the eastern Siberia (Ulrich et al. 2017), and consecutive positive anomalies of soil moisture were observed at the same region in the recent decade due to abnormally high winter precipitation (Iijima et al. 2010).

Interannual variability of ET in the Arctic major river basins, simulated by a land surface model CHANGE using WATCH forcing dataset. The dot lines and numbers represent the ET trend in individual river basins
The four major basins show an increasing trend of ET over the period of 1979–2015 (Fig. 4.10). However, the trends evidently indicate large regional differences; the Siberian basins have significantly high increasing rates of ET (+0.72–+1.01 mm yr−1) compared to Mackenzie. The significant increases in the Siberian basins are due to the recent warming temperature under the wetted soil moisture mentioned above because the temperature is a major deriving factor of ET (Park et al. 2008). The warming temperature also derives increases in vegetation biomass, so that the increased leaf area enhances transpiration and canopy interception to precipitation water. In contrast, the higher leaf area reduces solar radiation reaching on soil surface and thereby less soil evaporation. However, most soil evaporation occurs in the spring season when solar radiation is the strongest, and leaf opening is not initiated (Park et al. 2008). In the Mackenzie River, warming was consistent with Siberia, while there was soil drought due to less precipitation during the recent period (Park et al. 2013). It suggests that the lower ET in the Mackenzie River for the recent decade is likely because of the influence of the drought. The comparison of ET between the river basins concludes that the differences in regional ETs mainly resulted from different climate and soil moisture conditions.
5 The Net Surface Moisture Flux (P-E)
Great hydrologic consequences are arising from the variations in the net surface moisture flux, which is the difference between precipitation (P) and evapotranspiration (E), over the large region and watersheds. For simplicity, here we use E to represent evapotranspiration over terrestrial surfaces and evaporation over ocean surfaces. In this respect, a key underlying question is: Will Arctic landscapes become wetter or drier as climate changes? Over the ocean, a corresponding question is: Will the ocean surface gain or lose freshwater to the atmosphere? While scaling issues complicate the answer to these questions, the fundamental uncertainty surrounds the surface moisture budget and the relationship P and E. If P (including both rain and snow) exceeds E over a period of time, the excess goes into runoff or storage. If E exceeds P, the surface moisture deficit leads to drying over land unless there is sufficient recharge from below. A drying surface leads to decreased water supplies, increased wildfire risk, and moisture stress on vegetation, all of which have consequences for terrestrial ecosystems and human activities. Over the ocean, the consequences of an imbalance of P and E will be a change of the near-surface stratification.
The trajectory of Arctic surface wetness is confounded by observations of decreasing soil moisture in the Arctic (Hinzman et al. 2013) and in subarctic Swedish basins where precipitation has been increasing (Destouni and Varrot 2014). Earth system model results also show a reduction in wetland extent in higher latitudes, largely associated with permafrost thaw (Avis et al. 2011). The most comprehensive assessment of trends of P-E based on historical data and model simulations appears to be that of Rawlins et al. (2010), who used a variety of precipitation datasets, atmospheric reanalyses, land surface model output, and global climate models. Rawlins et al. (2010) used various approaches, such as average of nine global climate models (GCMs), average of five land surface models (LSMs), the surface energy balance and remote sensing (RS) method, the Variable Infiltration Capacity (VIC) model, and the ERA-Interim reanalysis, to synthesizing data, and found increases of E over Arctic terrestrial regions in recent decades. More generally, Rawlins et al. also found that trends of P, P-E, and river discharge were generally positive in the observational data, for which record lengths ranged from 20 to 50 years. However, trends of P-E, computed as differences between historical P datasets and satellite-derived (AVHRR GIMMS) E, showed no significant trend. The nine global climate models examined by Rawlins et al. (2010) showed statistically significant trends of terrestrial pan-Arctic P-E over the period 1950–1999 in eight of the nine cases, and in all nine cases for the period 1950–2049. Trends for the historical period were smaller than for the future period in the climate model output. All results were for annual means. The positive trends in annual mean P-E contrast with the expectation that longer and warmer summers will increase E sufficiently to favor summer drying. Anticipated increases of high-latitude wildfire activity (Partain et al. 2016; Flannigan et al. 2015) are consistent with this expectation, highlighting the mixed picture of future surface wetness trends in the Arctic. For the Arctic marine areas, Bintanja and Selton’s (2014) analysis of model output found that increased open water drove an increase of E that, in turn, was a major contributor to future increases of P over the Arctic Ocean in climate model simulations. However, compelling demonstrations of increases of E over the Arctic Ocean based on historical data are generally lacking.
For the future, one of the earlier examinations of pan-Arctic hydrology changes projected by climate models found that Arctic P-E increased in model simulations of the twenty-first century (Kattsov et al. 2007), implying a wetter Arctic surface in the future. However, that study presented changes in only the annual pan-Arctic P-E and did not consider differences between ocean and land areas nor between different terrestrial subregions. Model-based studies summarized below are unanimous in projecting future increases of river discharge in the Arctic. The coarse resolution of these models and their rudimentary treatment of permafrost and vegetative processes make it questionable to base conclusions about Arctic surface wetness trends on results from these simulations. A more recent evaluation of 25 CMIP5 global climate model projections, although global rather than the Arctic in scope, distinguished summer and winter changes of E (Laine et al. 2014). The results presented by Laine et al. (2014) highlight the challenge of assessing future changes in high-latitude surface wetness. As shown in Fig. 4.11 (Laine et al. 2014), The projected changes of P and E show the expected pattern, with increases of E over Arctic land areas, primarily during summer, and strong increases over the subarctic seas during winter in areas of sea ice loss (consistent with Bintanja and Selten 2014). However, the middle panels of Fig. 4.11 show that changes of P-E over the Arctic are much less spatially coherent and less robust than changes of P and E separately. In particular, summer P-E is projected to decrease much of Canada, increase over Alaska, decrease over the western and northern Eurasian subarctic, and increase over parts of northeastern Russia, including Chukotka. Over most of these areas, the sign of the projected change is not robust across the models at the 95% confidence level. Figure 4.11e’s spatial pattern over high-latitude land areas is very consistent with the projected changes in soil moisture obtained by Dirmeyer et al. (2013) using 15 of the same models. Dirmeyer et al. (2013) showed reductions of summer soil moisture over northern Canada and north-central Russia but not over Alaska and eastern Siberia, from the preindustrial to the twentieth century. However, decreases of summer soil moisture do spread to southern and central Alaska in the twenty-first century (Dirmeyer et al. 2013). The results of these two studies highlight the uncertainty in the trajectory of surface wetness in Arctic land areas in the present generation of global climate models.

25-model mean changes in precipitation P (upper panels), P-E (middle panels), and evapotranspiration E (lower panels) for Dec–Feb (left panels) and Jun–Aug (right panels). In dotted regions, the sign of change is robust among the different models at the 95% confidence level. Hatching in middle and lower panels indicates where a change in P-E is dominated by P and E, respectively (Laine et al. 2014, with open access)
River discharge data, which can be viewed as proxies for P-E integrated over river basins, generally show increases from the mid-twentieth century to the early 2000s, with varying degrees of statistical significance (Yang et al. 2004a, b; Shiklomanov and Lammers 2009; Overeem and Syvitski 2010; Holmes et al. 2013; Bring and Destouni 2014). Model-based studies consistently show that projected changes in atmospheric forcing will drive twenty-first-century increases in high-latitude river discharge (e.g., Vliet et al. 2013; Koirala et al. 2014; Bring et al. 2015).
Various factors contribute to the uncertainty in the projected changes in Arctic surface wetness. Internal variability, which can affect trends over decadal and multidecadal timescales, is clearly one consideration. However, studies such as those of Rawlins et al. (2010), Laine et al. (2014) and Dirmeyer et al. (2013) have utilized multimodel ensembles that tend to average out internal variations, and these studies indeed contained robust signals in the future changes of P and E, although not the hydrologically critical difference, P-E. Across-model differences in process formulations, especially the formulations in the terrestrial modules, almost certainly are key contributors to the uncertainty. Because of their coarse (~100–200 km) resolution, global climate models are often unable to resolve terrain and vegetation variations that characterize the Arctic landscape. The terrestrial modules of many global models include crude treatments of permafrost, with an inadequate representation of soil horizons and resolution of the changing active layer. Important differences in Arctic vegetation such as tundra types (e.g., heath vs. tussock), forest composition, and effects of small lakes and ponds are generally not well considered. Also not included in most global models are local topography, subgrid-scale vegetation distributions, and permafrost hydrology, which are all important determinants of soil moisture, drainage, and, to at least some extent, land surface evaporation.
6 Conclusion and Discussion
Evapotranspiration (ET) is a key component in global water and energy cycles. In the northern regions where water is either permanently or seasonally frozen, ET is influenced by both hydrological and thermal conditions of the land surface through complex physical processes and feedbacks to the surrounded environment. This chapter presents and discusses recent research advances about the ET in northern regions. ET in northern regions tends to increase with the decrease of latitude. The largest ET typically appears in the forest ecosystem, while the grasslands and shrublands have the small ET. While the seasonal variations in ET are usually high, the interannual variability in annual ET is usually low over the Arctic regions, reflecting the dominance of cold temperature constraints on northern ecosystem processes. It should be noted that the sublimation from snow cover accounts for about 15% (in Siberia) to 25% (in western Canada) of winter precipitation. This is certainly a non-negligible hydrological component that affects the end of winter snow mass, river discharge, and regional water resources across the broad northern regions. Many factors, such as soil moisture, vegetation type, and productivity, and ecosystem features affect ET over the northern regions. In addition, precipitation plays a key role in impacting ET. ET is more sensitive to precipitation in the early growing season than in the late growing season. Furthermore, changes in freeze–thaw processes due to warming also affect land surface conditions and the ET processes. Such an effect is different between the southern and northern parts of the Arctic because of the water supply in the soil. Therefore, annual ET shows positive correlations with the timing of primary seasonal thaw in the northern Arctic region, in which soil water is relatively large. This is because the earlier onset of springtime thaw facilitates earlier photosynthesis and transpiration and generally enhances plant growth and transpiration when plant-available moisture is high. However, the reverse relationship is witnessed in the southern Arctic regions where the water supply is usually limited. From 1983 to 2005, ET increased significantly in the Arctic region with a rate of 3.8 mm decade−1 because of regional warming and vegetation greening. Such an increase in ET may exert significant impacts on the regional hydrology and water resources. Advanced models can simulate past ET change over the large northern watersheds. Remote sensing has proved new ET data and information that support climate and hydrology research and applications. It should be noted that most available ET studies in the Arctic region rely heavily on the models and/or satellite observations, while the ground measurements of ET in such a harsh but hydroclimatically important regions are yet extremely sparse. It is therefore highly recommended that the community should focus more on improving our understanding of ET processes and its feedbacks to climate by integrating a wide range of in situ observations and physical-based modeling approach. Finally, it is a key underlying question: Will Arctic landscapes become wetter or drier as climate changes? Global models suggest increases of E over Arctic terrestrial regions in recent decades. Rawlins et al. also found that trends of P, P-E, and river discharge were generally positive in the observational data. For the future, summer P-E is projected to decrease much of Canada, increase over Alaska, decrease over the western and northern Eurasian subarctic, and increase over parts of northeastern Russia. Over most of these areas, the sign of the projected change is not robust across the models at the 95% confidence level. Many factors contribute to the uncertainty in the projected changes in Arctic surface wetness. There is certainly a need to quantify and narrow the uncertainties in global models over the large northern regions.
References
Angert A, Biraud S, Bonfils C, Henning CC, Buermann W, Pinzon J, Tucker CJ, Fung I (2005) Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proc Natl Acad Sci USA 102(31):10823–10827
Avis CA, Weaver AJ, Meissner KJ (2011) Reduction in areal extent of high-latitude wetlands in response to permafrost thaw. Nat Geosci 4(7):444–448
Baldocchi D (2014) Measuring fluxes of trace gases and energy between ecosystems and the atmosphere - the state and future of the eddy covariance method. Glob Change Biol 20(12):3600–3609
Baldocchi DD, Xu LK, Kiang N (2004) How plant functional-type, weather, seasonal drought, and soil physical properties alter water and energy fluxes of an oak-grass savanna and an annual grassland. Agric For Meteorol 123(1):13–39
Barry R, Prévost M, Stein J, Plamondon AP (1990) Simulation of snowmelt runoff pathways on the Lac Laflamme watershed. J Hydrol 113(1–4):103–121
Beck PSA, Horning N, Goetz SJ, Loranty MM, Tape KD (2011) Shrub Cover on the North Slope of Alaska: a circa 2000 Baseline Map. Arct Antarct Alp Res 43(3):355–363. https://doi.org/10.1657/1938-4246-43.3.355
Berg NH (1986) Blowing snow at a Colorado Alpine site: measurements and implications. Arct Alp Res 18(2):147–161
Betts AK, Ball JH, Viterbo P (1999) Basin-scale surface water and energy budgets for the Mississippi from the ECMWF reanalysis. J Geophys Res Atmos 104(D16):19293
Biederman JA, Brooks PD, Harpold AA, Gochis DJ, Gutmann E, Reed DE et al (2014) Multiscale observations of snow accumulation and peak snowpack following widespread, insect-induced lodgepole pine mortality. Ecohydrology 7:150–162. https://doi.org/10.1038/nclimate219810.1002/eco.1342
Bintanja R, Selten FM (2014) Future increases in Arctic precipitation linked to local evaporation and sea-ice retreat. Nature 509(7501):479–482
Boon S (2012) Snow accumulation following forest disturbance. Ecohydrology 5:279–285. https://doi.org/10.1002/eco.212
Bring A, Asokan SM, Jaramillo F, Jarsjö J, Levi L, Pietroń J, Prieto C, Rogberg P, Destouni G (2015) Implications of freshwater flux data from the CMIP5 multimodel output across a set of Northern Hemisphere drainage basins. Earths Future 3(6):206–217
Bring A, Destouni G (2014) Arctic climate and water change: model and observation relevance for assessment and adaptation. Surv Geophys 35(3):853–877
Brutsaert W (1982) Evaporation into the atmosphere: theory, history, and applications. Springer, New York
Brutsaert W (2013) Use of pan evaporation to estimate terrestrial evaporation trends: the case of the Tibetan Plateau. Water Resour Res 49(5):3054–3058
Calder IR (1998) Water use by forests, limits and controls. Tree Physiol 18(8–9):625–631. https://doi.org/10.1093/treephys/18.8-9.625
Chang L-L, Dwivedi R, Knowles J, Fang Y-H, Niu G-Y, D Pelletier J, Rasmussen C, Durcik M, Barron-Gafford G, Meixner T (2018) Why do large-scale land surface models produce a low ratio of transpiration to evapotranspiration? J Geophys Res: Atmos
Cleugh HA, Leuning R, Mu Q, Running SW (2007) Regional evaporation estimates from flux tower and MODIS satellite data. Remote Sens Environ 106(3):285–304. https://doi.org/10.1016/j.rse.2006.07.007
Coendersgerrits AMJ, Ent RJVD, Bogaard TA, Wangerlandsson L, Hrachowitz M, Savenije HHG (2014) Uncertainties in transpiration estimates. Nature 506(7487):E1
Davis RE, Hardy JP, Ni W, Woodcock C, McKenzie JC, Jordan R, Li X (1997) Variation of snow cover ablation in the boreal forest: a sensitivity study on the effects of conifer canopy. J Geophys Res Atmos 102(D24):29389–29395. https://doi.org/10.1029/97jd01335
Destouni G, Verrot L (2014) Screening long-term variability and change of soil moisture in a changing climate. J Hydrol 516(1):131–139
Dirmeyer PA, Jin Y, Singh B, Yan X (2013) Trends in land-atmosphere interactions from CMIP5 simulations. J Hydrometeorology 14(3):829–849
Evaristo J, Jasechko S, Mcdonnell JJ (2015) Global separation of plant transpiration from groundwater and streamflow. Nature 525(7567):91
Fischer M, Trnka M, Kučera J, Deckmyn G, Orság M, Sedlák P, Žalud Z, Ceulemans R (2013) Evapotranspiration of a high-density poplar stand in comparison with a reference grass cover in the Czech-Moravian Highlands. Agric For Meteorol 181:43–60
Fisher JB, Melton F, Middleton E, Hain C, Anderson M, Allen R, Mccabe MF, Hook S, Baldocchi D, Townsend PA (2017) The future of evapotranspiration: Global requirements for ecosystem functioning, carbon and climate feedbacks, agricultural management, and water resources. Water Resour Res 53(4):2618–2626
Fisher JB, Tu KP, Baldocchi DD (2008) Global estimates of the land–atmosphere water flux based on monthly AVHRR and ISLSCP-II data, validated at 16 FLUXNET sites. Remote Sens Environ 112(3):901–919. https://doi.org/10.1016/j.rse.2007.06.025
Flannigan MD, Wotton BM, Marshall GA, de Groot WJ, Johnston J, Jurko N, Cantin AS (2015) Fuel moisture sensitivity to temperature and precipitation: climate change implications. Clim Change 134(1–2):59–71
Goetz S, Bunn A, Fiske G, Houghton R (2005) Satellite-observed photosynthetic trends across boreal North America associated with climate and fire disturbance. Proc Nat Acad Sci USA 102(38):13521–13525
Good, S.P., Noone, D. and Bowen, G. (2015) WATER RESOURCES. Hydrologic connectivity constrains partitioning of global terrestrial water fluxes. Science 349(6244), 175–177
Harpold AA, Biederman JA, Condon K, Merino M, Korgaonkar Y, Nan TC et al (2014) Changes in snow accumulation and ablation following the Las Conchas Forest Fire, New Mexico, USA. Ecohydrology 7:440–452. https://doi.org/10.1002/eco.1363
Havranek WM, Tranquillini W (1995) 5–physiological processes during winter dormancy and their ecological significance. Ecophysiology of coniferous forests. p 95–124
Hogg EH, Brandt JP, Michaelian M (2008) Impacts of a regional drought on the productivity, dieback, and biomass of western Canadian aspen forests. Can J For Res 38(6):1373–1384. https://doi.org/10.1139/x08-001
Hinzman LD, Deal CJ, McGuire AD, Mernild SH, Polyakov IV, Walsh JE (2013) Trajectory of the Arctic as an integrated system. Ecol Appl 23(8):1837–1868
Holmes RM, Coe MT, Fiske GJ, Gurtovaya T, Mcclelland JW, Shiklomanov AI, Spencer RGM, Tank SE, Zhulidov AV (2013) Climate change impacts on the hydrology and biogeochemistry of arctic rivers. Wiley, Ltd
Iijima Y, Fedorov AN, Park H, Suzuki K, Yabuki H, Maximov TC, Ohata T (2010) Abrupt increases in soil temperatures following increased precipitation in a permafrost region, central Lena River basin. Russia Permafrost Periglac Process 21:30–41
Jasechko S, Sharp ZD, Birks SJ, Yi Y, Fawcett PJ (2013) Terrestrial water fluxes dominated by transpiration. Nature 496(7445):347
Kane DL, Hinzman LD, Gieck RE, Mcnamara JP, Youcha EK, Oatley JA (2008) Contrasting extreme runoff events in areas of continuous permafrost, Arctic Alaska. Hydrol Res 39(4):287–298
Kane DL, Hinzman LD, Mcnamara JP, Zhang Z, Benson CS (2000) An overview of a nested watershed study in Arctic Alaska. Nordic Hydrol 31(4–5):245–266
Kattelmann R, Elder K (1991) Hydrologic characteristics and water balance of an Alpine Basin in the Sierra Nevada. Water Resour Res 27(7):1553–1562
Kattsov VM, Walsh JE, Chapman WL, Govorkova VA, Pavlova TV, Zhang X (2007) Simulation and projection of arctic freshwater budget components by the IPCC AR4 global climate models. J Hydrometeorology 8(3):571–589. https://doi.org/10.1175/jhm575.1
Katul, GG, Oren R, Manzoni S, Higgins C, Parlange MB (2012) Evapotranspiration: a process driving mass transport and energy exchange in the soil‐plant‐atmosphere‐climate system. Rev Geophys 50(3)
Kimball JS, Mcdonald KC, Running SW, Frolking SE (2004) Satellite radar remote sensing of seasonal growing seasons for boreal and subalpine evergreen forests. Remote Sens Environ 90(2):243–258. https://doi.org/10.1016/j.rse.2004.01.002
Koirala S, Hirabayashi Y, Mahendran R, Kanae S (2014) Global assessment of agreement among streamflow projections using CMIP5 model outputs. Environ Res Lett 9(9):064017
Kool D, Agam N, Lazarovitch N, Heitman JL, Sauer TJ, Bengal A (2014) A review of approaches for evapotranspiration partitioning. Agric For Meteorol 184(1):56–70
Kurz, WA, Dymond, CC, Stinson, Rampley, GJ, Neilson, ET (2008) Mountain pine beetle and forest carbon feedback to climate change. Nature 45:987–990. https://doi.org/10.1038/nature06777
Laîné A, Nakamura H, Nishii K, Miyasaka T (2014) A diagnostic study of future evaporation changes projected in CMIP5 climate models. Clim Dyn 42(9–10):2745–2761
Leydecker A, Melack JM (1999) Evaporation from snow in the central Sierra Nevada of California. Hydrol Res 30(2):81–108
Liljedahl AK, Hinzman LD, Harazono Y, Zona D, Tweedie CE, Hollister RD, Engstrom R, Oechel WC (2011) Nonlinear controls on evapotranspiration in arctic coastal wetlands. Biogeosciences 8(11):3375–3389
Ma N, Szilagyi J, Zhang Y, Liu W (2019) Complementary-relationship-based modeling of terrestrial evapotranspiration across China during 1982–2012: validations and spatiotemporal analyses. J Geophys Res: Atmos 124. https://doi.org/10.1029/2018JD029580
Ma N, Wang N, Zhao L, Zhang Z, Dong C, Shen S (2014) Observation of mega-dune evaporation after various rain events in the hinterland of Badain Jaran Desert, China. Sci Bull 59(2):162–170
Mackay DS, Ewers BE, Cook BD, Davis KJ (2007) Environmental drivers of evapotranspiration in a shrub wetland and an upland forest in northern Wisconsin. Water Resour Res 43(3):W03442
Mantgem PJV, Stephenson NL, Byrne JC, Daniels LD, Franklin JF, Fule PZ, Harmon ME, Larson AJ, Smith JM, Taylor AH (2009) Widespread increase of tree mortality rates In the Western United States. Science 323(5913):521–524
Matheny AM, Bohrer G, Stoy PC, Baker IT, Black AT, Desai AR, Dietze MC, Gough CM, Ivanov VY, Jassal RS (2014) Characterizing the diurnal patterns of errors in the prediction of evapotranspiration by several land-surface models: an NACP analysis. J Geophys Res Biogeosci 119(7):1458–1473
Maxwell RM, Condon LE (2016) Connections between groundwater flow and transpiration partitioning. p 377
McClelland JW (2004) Increasing river discharge in the Eurasian Arctic: Consideration of dams, permafrost thaw, and fires as potential agents of change. J Geophys Res 109(D18). https://doi.org/10.1029/2004jd004583
Meiman JR, Grant LO (1974) Snow-air interactions and management on mountain watershed snowpack. In: Environmental Research Center. Colorado State University, Fort Collins, pp 3–5
Mendez J, Hinzman LD, Kane DL (1998) Evapotranspiration from a wetland complex on the arctic coastal plain of Alaska. Nordic Hydrol 29(4–5):303–330
Miralles DG, De Jeu RAM, Gash JH, Holmes TRH, Dolman AJ (2011) Magnitude and variability of land evaporation and its components at the global scale. Hydrol Earth Syst Sci 15(3):967–981
Mu Q, Heinsch FA, Zhao M, Running SW (2007) Development of a global evapotranspiration algorithm based on MODIS and global meteorology data. Remote Sens Environ 111(4):519–536. https://doi.org/10.1016/j.rse.2007.04.015
Mu QZ, Jones LA, Kimball JS, McDonald KC, Running SW (2009) Satellite assessment of land surface evapotranspiration for the pan-Arctic domain. Water Resour Res 45(20)
Musselman KN, Clark MP, Liu CH, Ikeda K, Rasmussen R (2017a) Slower snowmelt in a warmer world. Nat Clim Change 7:214–219. https://doi.org/10.1038/nclimate3225
Musselman KN, Clark MP, Liu CH, Ikeda K, Rasmussen R (2017b) Slower snowmelt in a warmer world. Na Clim Change 7:214–219. https://doi.org/10.1038/nclimate3225
Ni W, Li X, Woodcock CE, Roujean JL, Davis R E (1997) Transmission of solar radiation in boreal conifer forests: measurements and models. J Geophys Res Atmos 102(D24):29555–29566. https://doi.org/10.1029/97jd00198
Oberbauer S, Dawson T (1992) Water relations of Arctic vascular plants Physiological ecology of arctic plants: implications for climate change
Oechel WC, Vourlitis GL, Hastings SJ, Ault RP, Bryant P (1998) The effect of water table manipulation and elevated temperature on the net CO2 flux of wet sedge tundra ecosystem. Glob Change Biol 4(1):77–90
Ohta T, Kotani A, Iijima Y, Maximov T, Ito S, Hanamura M, Kononov A, Maximov A (2014) Effects of waterlogging on water and carbon dioxide fluxes and environmental variables in a Siberian larch forest. Agri For Meteorol 188:64–75
Oki T, Kanae S (2006) Global Hydrological Cycles and World Water Resources. Science 313(5790):1068
Oren R, HSIEH C-I, Stoy P, Albertson J, Mccarthy HR, Harrell P, Katul GG (2006) Estimating the uncertainty in annual net ecosystem carbon exchange: spatial variation in turbulent fluxes and sampling errors in eddy-covariance measurements. Glob Change Biol 12(5):883–896
Overeem I, Syvitski JPM (2010) Shifting discharge peaks in Arctic rivers, 1977–2007. Geogr Ann 92(2):285–296
Park H, Yamazaki T, Yamamoto K, Ohta T (2008) Tempo-spatial characteristics of energy budget and evapotranspiration in the eastern Siberia. Agric For Meteorol 148:1990–2005
Park H, Iijima Y, Yabuki H, Ohta T, Walsh J, Kodama Y, Ohata T (2011) The application of a coupled hydrological and biogeochemical model (CHANGE) for modeling of energy, water, and CO2 exchanges over a larch forest in eastern Siberia. J Geophys Res 116:D15102. https://doi.org/10.1029/2010JD015386
Park H, Walsh J, Fedorov AN, Sherstiukov AB, Iijima Y, Ohata T (2013) The influence of climate and hydrological variables on opposite anomaly in active-layer thickness between Eurasian and North American watersheds. Cryosphere 7:631–645
Partain JL Jr, Alden S, Strader H, Bhatt US, Bieniek PA, Brettschneider BR, Walsh JE, Lader RT, Olsson PQ, Rupp TS (2016) An Assessment of the role of anthropogenic climate change in the alaska fire season of 2015. Bull Am Meteor Soc 97(12):S14–S18
Penn CA, Bearup LA, Maxwell RM, Clow DW (2016) Numerical experiments to explain multiscale hydrological responses to mountain pine beetle tree mortality in a headwater watershed. Water Resour Res 52(4):3143–3161. https://doi.org/10.1002/2015wr018300
Pomeroy JW, Gray DM (1995) Snowcover accumulation, relocation and management. Sci Rep 7:134. Natl Hydrol Res Inst Environ Can, Saskatoon, Canada
Pomeroy JW, Li L (1997) Development of the prairie blowing snow model for application in climatological and hydrological models. Proc W Snow Conf 65:186–197
Pomeroy JW, Parviainen J, Hedstrom N, Gray DM (1998) Coupled modelling of forest snow interception and sublimation. Hydrol Process 12(15):2317–2337
Potter KM, Conkling BL (2016) Forest health monitoring: national status, trends, and analysis 2015 (Gen Tech Rep SRS-213). Asheville, NC: US Department of Agriculture, Forest Service, Southern Research Station
Pugh E, Small E (2012) The impact of pine beetle infestation on snow accumulation and melt in the headwaters of the Colorado River. Ecohydrology 5:467–477. https://doi.org/10.1002/eco.239
Raich JW, Schlesinger WH (1992) The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus 44(2):81–89
Rawlins MA, Steele M, Holland MM, Adam JC, Cherry JE, Francis JA, Groisman PY, Hinzman LD, Huntington TG, Kane DL (2010) Analysis of the arctic system for freshwater cycle intensification: observations and expectations. J Clim 23(21):5715–5737
Raz-Yaseef N, Young-Robertson J, Rahn T, Sloan V, Newman B, Wilson C, Torn MS (2017) Evapotranspiration across plant types and geomorphological units in polygonal Arctic tundra. J Hydrol 553:816–825
Schindler DW, Donahue W (2006) A case study of the Saskatchewan river system. In: Fifth Biennial Rosenberg International Forum on Water Policy
Schlesinger WH, Jasechko S (2014) Transpiration in the global water cycle. Agric For Meteorol 189–190(6):115–117
Scott RL, Biederman JA (2017) Partitioning evapotranspiration using long‐term carbon dioxide and water vapor fluxes. Geophys Res Lett 44
Scott RL, Huxman TE, Cable WL, Emmerich WE (2006) Partitioning of evapotranspiration and its relation to carbon dioxide exchange in a Chihuahuan Desert shrubland. Hydrol Process 20(15):3227–3243
Shen M, Piao S, Jeong SJ, Zhou L, Zeng Z, Ciais P, Chen D, Huang M, Jin CS, Li LZ (2015) Evaporative cooling over the Tibetan Plateau induced by vegetation growth. Proc Natl Acad Sci USA 112(30):9299–9304
Shiklomanov AI, Lammers RB (2009) Record Russian river discharge in 2007 and the limits of analysis. Environ Res Lett 4(4):045015
Shukla J, Mintz Y (1982) Influence of Land-Surface Evapotranspiration on the Earth’s Climate. Science 215(4539):1498–1501
Suni T, Berninger F, Markkanen T, Keronen P, Rannik l, Vesala T (2003) Interannual variability and timing of growing-season CO2 exchange in a boreal forest. J Geophys Res Atmos 108(D9). https://doi.org/10.1029/2002jd002381
Suzuki K, Ohata T, Kubota J, Vasilenko N, Zhuravin S, Vulinsy V (2002) Winter hydrological processes in the Mogot experimental watershed, in the southern mountainous taiga, Eastern Siberia. In: Proceedings of the 6th water resources symposium, Tokyo, 525–530
Suzuki K, Matsuo K, Yamazaki D, Ichii K, Iijima Y, Papa F, Yanagi Y, Hiyama T (2018) Hydrological variability and changes in the Arctic circumpolar tundra and the three largest Pan-Arctic river basins from 2002 to 2016. Remote Sens 10(3):402. https://doi.org/10.3390/rs10030402
Sexstone GA, Clow DW, Fassnacht SR, Liston GE, Hiemstra CA, Knowles JF, Penn CA (2018) Snow sublimation in mountain environments and its sensitivity to forest disturbance and climate warming. Water Resour Res 54:1191–1211. https://doi.org/10.1002/2017WR021172
Talsma CJ, Good SP, Jimenez C, Martens B, Fisher JB, Miralles DG, Mccabe MF, Purdy AJ (2018) Partitioning of evapotranspiration in remote sensing-based models. Agric For Meteorol s 260–261:131–143
Trenberth KE, Fasullo J, Dai A, Qian T, Smith L (2007) Estimates of the global water budget and Its annual cycle using observational and model data. J Hydrometeorology 8(4):758–769. https://doi.org/10.1175/jhm600.1
Trenberth KE, Fasullo JT, Kiehl J (2009) Earth’s global energy budget. Bull Am Meteor Soc 90(3):311–323
Ulrich M, Matthes H, Schirrmeister L, Schutze J, Park H, Iijima Y, Fedorov AN (2017) Differences in behavior and distribution of permafrost-related lakes in Central Yakutia and their response to climatic drivers. Water Resour Res 53. https://doi.org/10.1002/2016WR109267
Vliet MTHV, Franssen WHP, Yearsley JR, Ludwig F, Haddeland I, Lettenmaier DP, Kabat P (2013) Global river discharge and water temperature under climate change. Glob Environ Change-Hum Policy Dimensions 23(2):450–464
Wei Z, Yoshimura K, Wang L, Miralles DG, Jasechko S, Lee X (2017) Revisiting the contribution of transpiration to global terrestrial evapotranspiration. Geophys Res Lett 44:2792–2801
Westerling AL (2006) Warming and earlier spring increase Western U.S. forest wildfire activity. Science 313(5789):940–943. https://doi.org/10.1126/science.1128834
Wigmosta MS, Vail LW, Lettenmaier DP (1994) A distributed hydrology-vegetation model for complex terrain. Water Resour Res 30(6):1665–1679
Wilson KB, Baldocchi DD (2000) Seasonal and interannual variability of energy fluxes over a broadleaved temperate deciduous forest in North America. Agric For Meteorol 100(1):1–18
Woo M-K, Marsh P, Pomeroy JP (2000) Snow, frozen soils and permafrost hydrology in Canada, 1995–1998. Hydrol Process 14:1591–1611
Woodward FI, Kelly CK (1997) Plant functional types: towards a definition by environmental constraints. p 47–65
Wullschleger SD, Meinzer FC, Vertessy RA (1998) A review of whole-plant water use studies in tree. Tree Physiol (8–9):8–9
Xiao W, Zhongwang W, Wen X (2018) Evapotranspiration partitioning at the ecosystem scale using the stable isotope method—A review. Agric For Meteorol 263:346–361
Yamazaki T, Kondo J (1992) The snowmelt and heat balance in snow-covered forested areas. J Appl Meteorol 31(11):1322–1327
Yang D, Ye B, Kane D (2004a) Streamflow changes over Siberian Yenisei River Basin. J Hydrol 296(1–4):59–80. https://doi.org/10.1016/j.jhydrol.2004.03.017
Yang D, Kane D, Zhang Z, Legates D, Goodison B (2005) Bias corrections of long-term (1973–2004) daily precipitation data over the northern regions. Geophys Res Lett 32(19):n/a–n/a. https://doi.org/10.1029/2005gl024057
Yang D, Ye B, Shiklomanov A (2004b) Discharge characteristics and changes over the Ob river watershed in Siberia. J Hydrometeorol 5:595–610
Young-Robertson JM, Naama RY, Cohen LR, Brent N, Thom R, Victoria S, W C, W SD (2018) Evaporation dominates evapotranspiration on alaska's arctic coastal plain. Arct Antarct Alp Res 50(1)
Yuan W, Liu S, Liu H, Randerson JT, Yu G, Tieszen LL (2010) Impacts of precipitation seasonality and ecosystem types on evapotranspiration in the Yukon River Basin, Alaska. Water Res Res 46(2). https://doi.org/10.1029/2009wr008119
Zeng Z, Piao S, Li LZX, Zhou L, Ciais P, Wang T, Li Y, Lian X, Wood EF, Friedlingstein P (2017) Climate mitigation from vegetation biophysical feedbacks during the past three decades. Nat Clim Change 7(6):432–436
Zhang K, Kimball JS, Kim Y, Mcdonald KC (2011) Changing freeze-thaw seasons in northern high latitudes and associated influences on evapotranspiration. Hydrol Process 25(26):4142–4151
Zhang K, Kimball JS, Mu Q, Jones LA, Goetz SJ, Running SW (2009) Satellite based analysis of northern ET trends and associated changes in the regional water balance from 1983 to 2005. J Hydrol 379(1):92–110
Zhang Q, Manzoni S, Katul G, Porporato A, Yang D (2014) The hysteretic evapotranspiration—Vapor pressure deficit relation. J Geophys Res Biogeosci 119(2):125–140
Zhang Y, Ohata T, Kang E, Yao T (2003) Observation and estimation of evaporation from the ground surface of the cryosphere in eastern Asia. Hydrol Process 17:1135–1147
Zhang Y, Munkhtsetseg E, Kadota T, Ohata T (2005) An observational study of ecohydrology of a sparse grassland at the edge of the Eurasian cryosphere in Mongolia. J Geophys Res: Atmos 110. D14103. https://doi.org/10.1029/2004JD005474
Zhang Y, Ishikawa M, Ohata T, Oyunbaatar D (2008) Sublimation from thin snow cover at the edge of the Eurasian cryosphere in Mongolia. Hydrol Process 22:3564–3575
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Zhang, Y., Ma, N., Park, H., Walsh, J.E., Zhang, K. (2021). Evaporation Processes and Changes Over the Northern Regions. In: Yang, D., Kane, D.L. (eds) Arctic Hydrology, Permafrost and Ecosystems. Springer, Cham. https://doi.org/10.1007/978-3-030-50930-9_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-50930-9_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-50928-6
Online ISBN: 978-3-030-50930-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)