
Abstract
Reconstruction of ancient environmental conditions based on traditional analyses such as pollen and other proxies has provided understanding of climate variation and its influence on vegetation. These studies, however, are restricted to site-specific scales and have overseen the spatial understanding of past vegetation patterns. Hence, reconstruction of spatial distribution patterns of vegetation is yet a pending issue in paleoecological studies. The objective of this chapter is to develop a methodological integration of landscape approaches (paleoecological, bioclimatic, and geographical) to reconstruct spatial distribution patterns of vegetation in the Purepecha region in central Mexico. Correlation of climatic patterns, pollen rain, paleoecological data, and physical landscape components was jointly analyzed with the aid of a geographic information system. Result from this integrated approach indicated that during ca. 1000 B.C. (Preclassic period), climatic conditions were relatively more humid than the current climate. The dry climatic conditions were preferably dominant on valleys, plains, and footslopes. On the contrary, humid conditions were preferably distributed in hills and mountain landforms. Our outcomes provide a reproducible integrated methodology for reconstructing spatial patterns of vegetation. We, further, document for the first time the past spatial vegetation patterns in the core of the Purepecha culture.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
Introduction
Vegetation is a key component and one of the most recognizable elements on the Earth surface (Zonneveld 1989). Vegetation distribution and composition are determined by climatic, geomorphological, soil, and ecological factors, which interact simultaneously in long periods of time (Velázquez et al. 2016). While deep knowledge of the description and dynamics of local and regional vegetation is constantly rising (Rzedowski and Calderon 2013), there are still limited methodological contributions to reconstruct vegetation patterns, especially during the last millennium.
There are several approaches focusing at explaining the dynamic nature of vegetation. The paleoecological approach is based on the interpretation of fossil pollen and other proxies (e.g., diatoms, ostracods, charcoal), an objective view of climatic variations (Israde et al. 2010). The approach also provides valuable information on the impact of ancient societies (Islebe et al. 2016). The bioclimatological approach focuses on studying climate–vegetation relationship (Rivas-Martínez et al. 2011). This approach has been recently used as a tool for spatial reconstruction of the native vegetation (Castro-López and Velázquez 2018). The geographical approach relies upon landforms and soils and other physical attributes to delineate vegetation patterns. This approach uses geographic information systems (GIS) as a main integrative tool (Mas et al. 2004; Velázquez et al. 2016) enabling mapping and monitoring of a group of individuals, populations, and plant communities with different spatial resolution (Pedrotti 2013). Although there are numerous investigations to quantitatively reconstruct spatial patterns of vegetation, outcomes are yet far from being completed. Attempts to formalize the approaches have resulted in a great deal of interrelated and practical terminology under multiple forms (Reed et al. 2014). For this reason, the problem persists due to the lack of an integrative methodology that combines different approaches, namely, paleoecological, bioclimatic, and geographical, for contributing to the historical mapping of the vegetation. Any attempt to build a holistic approach will lead to a thorough understanding of the spatial distribution of vegetation. Some integrative methods have been proposed to reconstruct past scenarios from different perspectives. Some take as basis models to estimate the source area of the pollen and their deposits (Sugita et al. 2010). Others come to integrate landscape elements examined from a GIS (Etter et al. 2008; Brouwer 2013). In the same way, Caseldine and Fyfe (2006) integrate specific pollen parameters, vegetation mosaic, and landscape elements to simulate the distribution of pollen in a specific position within the landscape. Carrillo-Bastos et al. (2012) used fossil pollen interpolation methods to reconstruct the vegetation cover and infer precipitation variations. More comprehensive studies are reported by Gopar and Velázquez (2016) under a Boolean prediction framework giving emphasis to bioclimatic zoning. While the results are encouraging, they still offer limited outcomes. Reed et al. (2016) stated that the alternative of developing a holistic approach will tend toward providing detailed solutions to the problems of structure, composition, and spatial distribution of the past, present, and future vegetation. The integration is paramount to the relevance of contribution to understand the spatial-specific impacts of climate change. However, this integration is still, to our knowledge, absent in scientific literature.
The objective of this chapter is to provide an integrative methodology of approaches (paleoecological, bioclimatic, and geographical) to express the changes of spatial distribution patterns of vegetation in different time periods. This chapter focuses on the Purepecha region between the boundaries of Michoacan and Guanajuato. The region was limited because it has high plant and climatic diversity (Rzedowski and Calderon 2013).
Methods
Study Area
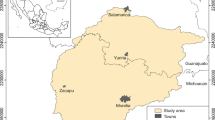
The area of study is located in the central part of the Michoacan-Guanajuato volcanic field (MGVF) (Cano-Cruz and Carrasco-Núñez 2008; Fig. 7.1). The MGVF is controlled by the fault system with east–west direction (Garduño-Monrroy et al. 2011). Volcanic activity has had significant impact on landscape formation from Oligocene to Holocene (Gómez-Vasconcelos et al. 2015). Rocks that emerge on the surface are basalts, andesites, rhyolites, ignimbrites, and granites (Cano-Cruz and Carrasco-Núñez 2008; Gómez-Vasconcelos et al. 2015).

Study area with the most important locations
The altitudinal gradient oscillates from 1700 m.a.s.l. (meters above sea level) in the plains to 3400 m.a.s.l. in the mountain area. Precipitation is controlled by the West winds carrying high precipitation and is concentrated, mainly, in the mountain zone (Reyes et al. 1994). Precipitation varies from 1200 to 600 mm with an average temperature of 20 °C (INEGI 2010). In elevations above 2000 m, it is recurrent to find vegetation types such as Abies, Pinus, Quercus, Arbutus, Alnus, Buddleja, Crataegus, and Clethra (Pérez-Calix 1996). In elevations less than 2000 m, the predominant genera are Acacia, Bursera, Cedrela, Condalia, Euphorbia, Erythrina, Eysenhardtia, Forestiera, and Yucca and in the proximity of the rivers Salix, Taxodium, Alnus, and Fraxinus (Rzedowski and Calderon 2013). Over the last few decades, the plant diversity has experienced high degree of disturbance, especially on foot slopes, valleys, and plains (Villaseñor and Ortiz 2012).
Bioclimatic Analysis
The bioclimatic analysis was carried out using the databases of the Digital Climatic Atlas of Mexico (version 2.0) (http://uniatmos.atmosfera.unam.mx/ACDM/). The layers are in raster format with pixel of 1 km2. The raster contains average temperature and precipitation information from 1902 to 2011. Raster layers were imported into the ArcGis to calculate bioclimatic indexes. The indexes used were the annual Ombrothermic Index (Io = (Pp/Tp) ∗ 10), Ombrothermic Index of the driest bimester of the driest quarter of the year (Iod2 = (Ppd2/Tpd2) ∗ 10), and Thermicity Index (It = (T + M + m) ∗ 10). The results of the indexes were compared with bioclimatic keys developed by Rivas-Martínez et al. (2011) to identifying bioclimates, thermotypes, and ombrotypes. The generated information was transformed to vector format to facilitate the editing of the data and to improve its spatial expression. The minimum cartographic area was 1 km2 to maintaining consistency in the reading of the map. Our study only presents the thermotypes and ombrotypes to scale 1:250,000 with projection to NAD 1927 UTM 14 N Zone.
Pollen Rain Analysis
In total, 57 samples of moss and surface water were collected following a climatic gradient from dry tropical to wet temperate conditions. Samples were collected within different types of vegetation: temperate pine–oak forests and tropical dry forests. Pollen grains were extracted using the acetolysis technique of Erdtman (1952). Subsequently, the samples were mounted with glycerin for identification. An optical microscope was used to identify pollen grains until reaching a minimum of 300 grains of arboreal taxa. Pollen taxa that presented more than 2% were used for canonical correspondence analysis (CCA) to examine whether they were related to climatic patterns. The CCA is a statistical technique for exploring relationships between two multivariate sets of variables (Gauch 1982). The analysis was done with the PC-ORD 5 Software (McCune and Mefford 2006).
Radiocarbon Data and Pollen Diagrams
The paleoecological approach was used to have a chronological framework in the lake sediments (Israde et al. 2010). Research outcomes often lack calibrations, so that dating uncertainties remain (Giesecke et al. 2014). We conducted a review of the paleoecological studies conducted in the study area for C14 dating. The database was calibrated to calendar years using Calib 14 (version 7.1) (Stuiver et al. 2018). All the lakes located within the study area were evaluated in the present survey. A first approximation ca. 1000 B.C. showed a greater relationship in the chronological dating. On the dating indicated, percentage data of pollen and the interpretations of the authors were collected for the model. In the same way, the minimum, average, and maximum values of the taxa were collected throughout the diagram.
Bioclimatic Approach
The isobioclimates were used to build a frame of reference in relation to the vegetation cover. The isobioclimates are the combination of bioclimates, thermotypes, and ombrotypes. All correlations were estimated in GIS. Landscape components were overlapped to find correlation patterns (Fig. 7.2). Subsequently, we placed the percentage values of pollen fossil in relation to the reference frame. In this test, CCA results were also used to validate the information. The isobioclimates that were empty subsequently were re-labeled based on the climatic gradient expressed by Gopar et al. (2015).

Methodological processes for the reconstruction of the vegetal landscape. Each step must be done independently
Statistical Analysis
Data under study that come from different approaches were subjected to canonical correspondence analysis to identify patterns of climatic, plant species assemblages and landscape components (Velázquez et al. 2016). Best ordinations axes with significant values to depict patterns were chosen, and from there plant species assemblages and its distribution were defined.
Results
The integration of landscape approaches (paleoecological, bioclimatic, and geographical) under a baseline allowed the methodological construction for the reconstruction of climatic patterns and vegetation cover. The current bioclimatic analysis was determined by three thermotypes (temperature gradient) and three ombrotypes (precipitation gradient). The mesotropical thermotypes occupied the largest area, about 84%, whereas the thermotropical about 14%, and the supratropical about 2% (Fig. 7.3).

Distribution of current thermotypes (temperature gradient)
The ombrotypes is composed of the dry ombrotype with 49% of the total surface, followed by the subhumid ombrotype with 40% and the humid ombrotype around 11% (Fig. 7.4).

Current distribution ranges of ombrotypes (precipitation gradient)
The CCA showed that the environmental variables showed a high relation with pollen (Fig. 7.5). The first axis contributed 11.2% with positive regression coefficient between humid climates to subhumid and temperate pollen. Dry climates showed negative regression in relation to tropical pollen and grasslands. The third axis contributed 6.4% of the variance explained. In this axis, the positive regression coefficient was associated with tropical and temperate taxa. On the other hand, pollen grasslands showed negative values. It should be also added that the Pearson correlation for axis 1 and axis 3 was 0.76 and 0.75 close to 1. Thus, our results indicate a good segregation of the data. The tropical pluviseasonal mesotropical humid (TPMH) isobioclimate presented a greater relation to Pinus, Quercus, and Alnus. The tropical pluviseasonal mesotropical subhumid (TPMSH) showed not relationship with any taxa specific.

Organizational chart of pollen taxa in relation to current isobioclimates. Environmental variables (isobioclimates) are displayed with arrows. TPSH tropical pluviseasonal supratropical humid, TPMH tropical pluviseasonal mesotropical humid, TPMSH tropical pluviseasonal mesotropical subhumid, TXMD tropical xeric mesotropical dry, and TXTD tropical xeric thermotropical dry
The calibration of C14 dating revealed that six sites have corresponded to 1000 B.C., with a variation of ±200 years. Figure 7.6 compares the paleoecological records collected within the study area. The figure only shows the quantitative data of the proxies. For this dating, the study area was represented by mesotropical thermotype and humid ombrotype in most of the surface. On the other hand, a dry isobioclimate was observed (Fig. 7.7).

Diagram of proxies located within the study area. The dotted horizontal line indicates the worked age. Patz1 (Watts and Bradbury 1982), Patz2 (Metcalfe et al. 2007), Zirah1 (Ortega et al. 2010), Zirah2 (Torres-Rodríguez et al. 2012), Zacap1 (Lozano-Garcia and Xelhuantzi-López 1997), Zacap2 (Leng et al. 2005), Zacap3 (Arnauld et al. 1997), Yurir (Metcalfe et al. 1989), and San Nicolas de Parangueo (Brown 1984) (∗) interpolated data

Diversity climates in the area of study ca. 1000 B.C. Climate in “X” is not available in the study area. Climate of the hatched area is located at latitudes above 3100 m.a.s.l. ∗ Data interpolated
Discussion
The integration of landscape approaches provides a simple and useful tool for reconstructing past scenarios. A first approximation of ca. 1000 B.C. indicates that the humid ombrotype were located in the areas of hills and foot of mountains and dry ombrotype in the valleys. Torres-Rodríguez et al. (2012) report at the Lake Zirahuen humid conditions with elements of mesophilic forest before this data, and subsequent to this period, a decrease in humidity persists. Rivas-Martínez et al. (2011) argues that climates are positively associated with the altitudinal gradient, thermal and pluviometrical, where its regional distribution follows clear and uniform altitudinal gradients in systematic order. This would reinforce the argument that climatic strips showed a displacement following this climatic gradient.
The methodological integration of landscape approaches (paleoecological, bioclimatic and geographical) enabled the recognition of the distribution of climatic patterns. This type of research is widely documented by paleoecological studies on a punctual scale. However, their understanding and spatial expression have been limited due to the scarcity of suitable sites. Therefore, they contribute to understanding part of the process and not the regional complexity. The integration of approaches gives a clear and concise view of the chorological changes in climatic patterns and plant communities in different temporal scales.
GIS is a powerful tool for integrating, analyzing, and managing input and output data from the implemented models (Devereux et al. 2004). Leverington et al. (2002) argue that the use of GIS lies in the rapidity to generate quantitative and georeferenced databases. It is used as an efficient tool to integrate local and spatial database and to express the scenarios of the historical landscape (Gaudin et al. 2008; Carrillo-Bastos et al. 2012; Brouwer 2013). It must be also added that an adequate integration of landscape components, hierarchically analyzed, will allow recognizing the different patterns that give subsistence to the vegetation types (Gopar and Velázquez 2016) and their different degrees of transformation (Etter et al. 2008) and provide sustainable natural resource solutions on multiple timescales (Reed et al. 2016). The results can be even more promising if more emphasis is given to bioclimatic zones. Gopar and Velázquez (2016) reported that bioclimatic analysis is an excellent tool to identify the climate-vegetation relationship, especially in regions with high geological, ecological, and human complexity. Currently, there is a rise in spatializing the historical landscape. However, the validation and calibration of historical maps has been hampered by the lack of more precise spatial data (Etter et al. 2008). By virtue of this, biophysical characteristics provide an alternative on the conditions of habitat, and their projection with the same physical characteristics reflected the complement of species associated to them (Simonson et al. 2018).
Overall, the analyses here used have proven to be an effective tool to help reconstructing vegetation distribution patterns. Nonetheless, pending uses remain, for example the association of the pollen spectrum with the climate and their relationship with vegetation patterns were not fully completed. It is necessary to recalibrate dates to have a complete chronological framework of the lake sediments. Furthermore, limitations of the study should be considered for future research; in particular, it can be used if topography has not changed significantly. García-Quintana et al. (2016) report that during the last 3000 years B.P., no major changes have occurred compared to what happened during the Pleistocene and the beginning of the Holocene. It should be also mentioned that the difference between lake sedimentation rates makes difficult having a linear chronological framework (Israde et al. 2010). In the same way, it is also necessary to incorporate the orientation of slopes to have a regional overview, since some types of vegetation are confined to certain slope orientations favoring their establishment and development (Rzedowski 2006).
To conclude, the integration of paleoecological, bioclimatic, and geographical approaches showed promising results for understanding past vegetation distribution patterns. This information may be a key to quantitatively assess in a region with a great cultural value.
References
Arnauld C, Metcalfe SE, Petrequin P (1997) Holocene climatic change in the Zacapu Lake Basin, Michoacan: synthesis of results. Quat Int 43(97):173–179
Brouwer M (2013) Reconstructing total paleo-landscapes for archaeological investigation: an example from the central Netherlands. J Archaeol Sci 40(5):2308–2320. https://doi.org/10.1016/j.jas.2013.01.008
Brown RB (1984) The paleoecology of the northern frontier of mesoamerica (pollen, mexico, archaeology). Dissertation, University of Arizona, USA
Cano-Cruz M, Carrasco-Núñez G (2008) Evolución de un cráter de explosión (maar) riolítico: Hoya de Estrada, Campo Volcánico Valle de Santiago, Guanajuato, México. Revista Mexicana de Ciencias Geológicas 25(3):549–564
Carrillo-Bastos A, Islebe GA, Torrescano-Valle N (2012) Geospatial analysis of pollen records from the Yucatán Peninsula, Mexico. Veg Hist Archaeobotany 21(6):429–437. https://doi.org/10.1007/s00334-270012-0355-1
Caseldine C, Fyfe R (2006) A modelling approach to locating and characterizing elm decline/landnam landscapes. Quat Sci Rev 25(5–6):632–644. https://doi.org/10.1016/j.quascirev.2005.07.015
Castro-López V, Velazquez A (2018) Reconstruction of native vegetation based upon integrated landscape approaches. Biodivers Conserv. https://doi.org/10.1007/s10531-018-1655-2
Devereux BJ, Devereux LS, Lindsay C (2004) Modelling the impact of traffic emissions on the urban environment: a new approach using remotely sensed data. In: Kelly REJ, Drake NA, Barr SL (eds) Spatial modelling of the terrestrial environment. Wiley, Chichester, pp 227–242
Erdtman OS (1952) Pollen morphology and plant taxonomy, angiosperms, (an introduction to palynology). Alqmus and Wiksell, Stockholm
Etter A, McAlpine C, Possingham H (2008) Historical patterns and drivers of landscape change in Colombia since 1500: a regionalized spatial approach. Ann Assoc Am Geogr 98(1):2–23. https://doi.org/10.1080/00045600701733911
García-Quintana A, Goguitchaichvili A, Morales J et al (2016) Datación magnética de rocas volcánicas formadas durante el Holoceno: caso de flujos de lava alrededor del Lago de Pátzcuaro (campo volcánico Michoacán-Guanajuato). Revista Mexicana de Ciencias Geológicas 33(2):209–220
Garduño-Monroy VH, Soria-Caballero DC, Israde-Alcántara I, Hernández-Madrigal VM, Rodríguez-Ramírez A, Ostroumov M, Rodríguez-Pascua MA, Chacon-Torres AC, Mora-Chaparro JC (2011) Evidence of tsunami events in the Paleolimnological record of Lake Pátzcuaro, Michoacán, México. Geofis Int 50(2):147–161
Gauch HG (1982) Multivariate analysis in community ecology. Cambridge University Press, Cambridge, NY
Gaudin L, Dominique M, Lanos P (2008) Correlation between spatial distributions of pollen data, archaeological records and physical parameters from North-Western France: a GIS and numerical analysis approach. Veg Hist Archaeobotany 17(5):585–595. https://doi.org/10.1007/s00334-008-0172-8
Giesecke T, Davis B, Brewer S et al (2014) Towards mapping the late quaternary vegetation change of Europe. Veg Hist Archaeobotany 23(1):75–86. https://doi.org/10.1007/s00334-012-0390-y
Gómez-Vasconcelos MG, Garduño-Monrroy VH, Macías JL et al (2015) The Sierra de Mil Cumbres, Michoacán, México: transitional volcanism between the Sierra Madre Occidental and the Trans-Mexican Volcanic Belt. J Volcanol Geotherm Res 301:128–147. https://doi.org/10.1016/j.jvolgeores.2015.05.005
Gopar-Merino LF, Velázquez A (2016) Componentes del paisaje como predictores de cubiertas de vegetación: estudio de caso del estado de Michoacán, México. Investigaciones Geográficas (90):75–88. https://doi.org/10.14350/rig.46688
Gopar-Merino LF, Velázquez A, Giménez de Azcárate J (2015) Bioclimatic mapping as a new method to assess effects of climatic change. Ecosphere 6(1):1–12. https://doi.org/10.1890/ES14-00138.1292
INEGI (2010) Compendio de información geográfica municipal, México http://www.inegi.org.mx/geo/contenidos/topografia/compendio.aspx. Accessed 1 July 2018
Islebe GA, Domínguez-Vázquez G, Espadas-Manrique C et al (2016) Cambio climático: contexto histórico, paleoecológico y paleoclimático. Tendencias actuales y perspectivas. In: Balvanera P, Arias-González JE, Rodríguez-Estrella R, Almeida-Leñero L, Schmitter-Soto JJ (eds) Una mirada al conocimiento de los ecosistemas de México. Universidad Nacional Autónoma de México (UNAM), Ciudad de México, pp 25–56
Israde-Alcántara I, Velázquez-Durán R, Lozano-García MS et al (2010) Evolución paleolimnológica del Lago Cuitzeo, Michoacán durante el Pleistoceno-Holoceno. Bol Soc Geol Mex 62(3):345–357. https://doi.org/10.1016/j.quaint.2004.10.022
Leng MJ, Metcalfe SE, Davies SJ (2005) Investigating Late Holocene climate variability in central Mexico using carbon isotope ratios in organic materials and oxygen isotope ratios from diatom silica within lacustrine sediments. J Paleolimnol 34(4):413–431. https://doi.org/10.1007/s10933-005-6748-8
Leverington DW, Teller JT, Mann JD (2002) A GIS method for reconstruction of late Quaternary landscapes from isobase data and modern topography. Comput Geosci 28(5):631–639. https://doi.org/10.1016/S0098-3004(01)00097-8
Lozano-García MS, Xelhuantzi-López MS (1997) Some problems in the Late Quaternary pollen records of Central Mexico: Basins of Mexico and Zacapu. Quat Int 43–44(97):117–123. https://doi.org/10.1016/S1040-6182(97)00027-X
Mas JF, Velázquez A, Díaz-Gallegosa JR et al (2004) Assessing land use/cover changes: a nationwide multidate spatial database for Mexico. Int J Appl Earth Obs Geoinf 5(4):249–261. https://doi.org/10.1016/j.jag.2004.06.002
McCune B, Mefford MJ (2006) PC-ORD, multivariate analysis of ecological data, version 5.31. MJM Software. Gleneden Beach, Oregon, EEUU
Metcalfe SE, Street-Perrott FA, Brown RB et al (1989) Late Holocene human impact on Lake basins in Central Mexico. Geoarchaeol Int J 4(2):119–141
Metcalfe SE, Davies SJ, Braisby JD et al (2007) Long and short-term change in the Pátzcuaro Basin, central Mexico. Palaeogeogr Palaeoclimatol Palaeoecol 247(3–4):272–295. https://doi.org/10.1016/j.palaeo.2006.10.018
Ortega B, Vázquez G, Caballero M et al (2010) Late Pleistocene: Holocene record of environmental changes in Lake Zirahuen, Central Mexico. J Paleolimnol 44(3):745–760. https://doi.org/10.1007/s10933-010-9449-x
Pedrotti F (2013) Plant and vegetation mapping. Springer, Berlin/Heridelberg
Pérez-Calix E (1996) Flora y vegetación de la cuenca del lago de Zirahuén Michoacán, México. Flora del Bajío y de regiones adyacentes, fascículo complementario XIII:1–73
Reed J, Deakin L, Sunderland T (2014) What are integrated landscape approaches and how effectively have they been implemented in the tropics: a systematic map protocol. Environ Evid 4(2):1–7. https://doi.org/10.1186/2047-2382-4-2
Reed J, Van Vianen J, Deakin EL, Barlow J, Sunderland T (2016) Integrated landscape approaches to managing social and environmental issues in the tropics: learning from the past to guide the future. Glob Chang Biol 22:2540–2554. https://doi.org/10.1111/gcb.13284
Reyes S, Douglas MW, Maddox RA (1994) El monzón del suroeste de Norteamérica (TRAVASON/SWAMP). Atmósfera 7:117–137
Rivas-Martínez S, Rivas-Sáenz S, Penas A (2011) Worldwide bioclimatic classification system. Glob Geobot 1:1–634. https://doi.org/10.5616/gg110001
Rzedowski J (2006) Vegetación de México. 1ª Edición digital, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. http://www.biodiversidad.gob.mx/publicaciones/librosDig/pdf/VegetacionMx_Cont.pdf. Accessed 1 May 2018
Rzedowski J, Calderón de Rzedowski G (2013) Datos para la apreciación de la flora fanerogámica del bosque tropical caducifolio de México. Acta Botánica Mexicana 102:1–23
Simonson WD, Allen HD, Parham E et al (2018) Modelling biodiversity trends in the montado (wood pasture) landscapes of the Alentejo, Portugal. Landsc Ecol 33(5):811–827. https://doi.org/10.1007/s10980-018-0627-y
Stuiver M, Reimer PJ, Reimer RW (2018) CALIB 7.1 [WWW program] at http://calib.org. Accessed 20 May 2018
Sugita S, Parshall T, Calcote R, Walker K (2010) Testing the landscape reconstruction algorithm for spatially explicit reconstruction of vegetation in northern Michigan and Wisconsin. Quat Res 74(2):289–300. https://doi.org/10.1016/j.yqres.2010.07.008
Torres-Rodríguez E, Lozano-García S, Figueroa-Rangel BL et al (2012) Cambio ambiental y respuestas de la vegetación de los últimos 17,000 años en el centro de México: el registro del lago de Zirahuén. Revista Mexicana de Ciencias Geológicas 29(3):764–778
Velázquez A, Medina-García C, Durán-Medina E et al (2016) Standardized hierarchical vegetation classification. Mexican and global patterns. Springer Nature, Basel
Villaseñor JL, Ortiz E (2012) La familia Asteraceae en la flora del Bajío y de regiones adyacentes. Acta Botánica Mexicana 100:259–291
Watts WA, Bradbury JP (1982) Paleoecological studies at Lake Patzcuaro on the west-central Mexican Plateau and at Chalco in the basin of Mexico. Quat Res 17(1):56–70. https://doi.org/10.1016/0033-5894(82)90045-X
Zonneveld IS (1989) The Land Unit a fundamental concept in landscape ecology, and its applications. Landsc Ecol 3(2):67–86
Acknowledgments
We would like to thank the Mexican National Council of Science and Technology (CONACYT) for providing a PhD scholarship to the first author. We thank Rocio Aguirre, Fernando Gopar, and Dulce Bocanegra for their invaluable help and assistance in the fieldwork.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Castro-López, V., Velázquez, A., Domínguez-Vázquez, G. (2019). Integration of Landscape Approaches for the Spatial Reconstruction of Vegetation. In: Torrescano- Valle, N., Islebe, G., Roy, P. (eds) The Holocene and Anthropocene Environmental History of Mexico. Springer, Cham. https://doi.org/10.1007/978-3-030-31719-5_7
Download citation
DOI: https://doi.org/10.1007/978-3-030-31719-5_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-31718-8
Online ISBN: 978-3-030-31719-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)