
Abstract
Here we review sled dogs as a sentinel monitoring species of ecosystem health across the Arctic focusing on environmental changes including pollution, climate change, and infectious diseases. Studies on environmental contaminants have been carried out mostly in Alaska and Greenland. While the majority of reports focus on mercury exposure and health effects, a major classical case-controlled study of exposure and effects from persistent organic pollutants (POPs) has been carried out on Greenland sled dog bitches and their pups. Altogether, the studies show that mercury and POPs affect multiple health endpoints across physiological systems, including reproductive, endocrine, and immune systems, that ultimately affect systems such as the liver and kidney. Therefore, sled dogs have proved to be a good model for assessing the health effects from contaminant exposure of top predators and Northerners in the Arctic. Furthermore, they are widely distributed across the Arctic and show similar correlations to important health indicators reported in Northerners and polar bears. With respect to climate change and disease dynamics of zoonosis, most studies have taken place in Canada. However, at present sled dogs are not utilized in monitoring studies of zoonotic diseases. Such an inclusion will increase the understanding of environmental changes, pollution, and diseases dynamics in Northerners and wildlife. We therefore recommend that ecosystem health assessments in the Arctic including that of Northerners start to include analyses of sled dogs combined with modeling tools. Doing so in a circumpolar perspective will further increase our understanding and monitoring possibilities of ecosystem health and Northerners exposure to contaminants, diseases, and climate change in the Arctic.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Arctic
- Arctic fox
- Climate change
- Contaminants
- Diseases
- Ecosystem
- Endocrine
- Energetics
- Genetics
- Histopathology
- Hormones
- Immune
- Inuits
- Mercury
- OHCs
- PCB
- One Health
- Organohalogen
- PBPK
- Persistent organic pollutants
- Polar bears
- POPs
- Sentinels
- Sled dogs
- Vitamins
- Zoonosis
1 Introduction: The Sled Dog
The domesticated sled dog (Canis familiaris) is widely used by indigenous people in Greenland, Alaska, Canada, and Russia to pull dog sledges (Fig. 2.1). It hence represents a unique mode of transport allowing, e.g., hunters and tourist guides to explore, hunt, and transport their items and food in over considerable distances during the winter where the sea ice and snow coverage serve as the “road” for this ancient and original form of transport. The use of sled dogs as an animal engine of transport as compared to modern forms of transport in the Arctic, like the snowmobile or outboard driven engines, is of course slower, but much more reliable and does not require expensive investments and expensive gasoline which may limit the range and cause functional problems. In addition, the dogs can warn the hunters against polar bears (Ursus maritimus) in the night. They are even used during the bear hunt where a few dogs will be cut loose from the dragging team and will catch up with and distract the bear. Sled dogs are excellent in helping the driver to avoid unstable ice conditions, and the sledge is extremely suitable for crossing difficult landscape routes due to its high flexibility. Finally, the dogs are more silent and reliant compared to, for example, a noisy snowmobile that may break down, which make dogs very suitable for hunting in remote areas.

The dog sledge represents a unique transport medium allowing the hunters to explore, hunt, and transport their items and food over considerable distances. Northwest Greenland, spring 1984. (Photo: R. Dietz)
In Greenland, the number of sled dogs has decreased by ca. 50% over the last three decades (Sonne et al. 2018; Statistics Greenland 2017, Fig. 2.2). The reason for this is a substantial loss of sea ice, which has reduced the ability of locals to go hunting and fishing using sledges and ice as their transport platform. Such changes influence lifestyle and the way of living as well as dietary habits of Northerners (Cavalieri and Parkinson 2012).

Trends in the Greenland sled dogs population size from 1990 to 2016. The total (red) and West Greenland (blue) numbers refer to the Y-axis, while the East Greenland (yellow) numbers refer to the Z-axis. (Source: Statistics Greenland (2017))
2 The Sled Dog as Circumpolar Biomonitoring Species
The sled dog is equally distributed in the circumpolar areas of Europe, North America, and Asia, and despite some physiological differences, it may therefore be an ideal biomonitoring species for Northerners and predator health in the Arctic (Burger and Gochfeld 2001; Sonne 2010; Sonne et al. 2017a). Sled dogs live in proximity to Northerners, and since they are fed a local food web diet, sled dogs are ideal to monitor contaminants and diseases in a One Health perspective (Sonne et al. 2017a). Relatively few studies have used sled dogs as a sentinel species for One Health monitoring in the Arctic (Sonne et al. 2017a). Given that sled dogs have a known history and are easy to access, they are suitable as additional biomonitoring species to polar bears and Arctic fox (Vulpes lagopus).
3 Pollution in the Arctic
Pollution has been extensively monitored in the Arctic due the high exposure of Inuit populations appearing from high trophic consumption of marine mammals (AMAP assessment 2018; AMAP 2015). Since the 1940s, large amounts of lipophilic organohalogen compounds (OHCs) have been released into the environment and transported to the Arctic (AMAP 2014, Rigét 2019; de Wit et al. 2010; Butt et al. 2010; Hoferkamp et al. 2010; Hung et al. 2010). This transport is mainly caused by the so-called grasshopper effect being an evaporation-precipitation mechanism due to the relatively low vapor pressure of these pollutants. The OHCs include polychlorinated biphenyls (PCBs) , organochlorine pesticides (OCPs), brominated flame retardants (BFRs), and proteinophilic per- and polyfluoroalkyl substances (PFASs), such as perfluorinated carboxylic acids (PFCAs) and perfluorinated sulfonates (PFSAs). These chemicals typically originate from industrial and household use of insulating fluids and coolants in electric and electronic equipment and machinery, agricultural pest control, textiles, construction material, and firefighting foams (de Wit et al. 2010). Many of these compounds are persistent in the environment and organisms, undergo long-range transport by atmospheric and seawater currents, and ultimately end up in the cold Arctic environment (Rigét et al. 2016; Letcher et al. 2010, 2018). Arctic fish and wildlife rely on energy-rich fatty and waxy tissues as their main energy source (Butt et al. 2010; Dietz et al. 2013; Houde et al. 2011). Such fatty tissues typically accumulate various natural fat-soluble compounds, such as specific vitamins (A and D) and endogenous steroid hormones depending on the species-specific fatty acid and lipid class composition (Sonne 2010; Sonne et al. 2014a, b; Kirkegaard et al. 2010a, b, Bechshøft et al. 2011). However, these tissues also easily absorb the lipophilic contaminants, which are consequently prone to biomagnify through the food web to top predators such as polar bears, Arctic fox, seals, whales, and seabirds and ultimately to indigenous Northerners and their pets that traditionally consume the aforementioned wildlife (Letcher et al. 2010, 2018; Rigét et al. 2016).
In addition to these substances, elevated emissions of mercury (Hg) since the 1850s (early industrialization) have led to a global spread and high concentrations of this heavy metal (Dietz et al. 2009). The Hg sources include burning of fossil fuels, metal production (ferrous and non-ferrous), large-scale gold production, artisanal and small-scale gold production, cement production, chlor-alkali industry, waste incineration, as well as release from dental amalgam upon body cremation (Outridge et al. 2011). Mercury is easily methylated by marine primary producers, biomagnifies, and thus poses health concerns for Arctic top predators and wildlife (AMAP 2015; Dietz et al. 2013; Outridge et al. 2011). In fact, predator mammals in East Greenland and Svalbard, such as sled dogs, polar bears, Arctic foxes, and killer whales (Orcinus orca), are among the most contaminated animals on our globe (Dietz et al. 2015; Letcher et al. 2010; Pedersen et al. 2015; Pedro et al. 2017). Several of the OHCs and Hg have been globally regulated through international treaties and conventions including the Stockholm and the Minamata Conventions, which initially resulted in environmental declines. However, over the last decade, the concentrations of the highly toxic PCBs and PFASs have remained essentially unchanged in polar bears inhabiting contaminant hot spots, such as East Greenland and Hudson Bay, due to climate-related changes in the food chains, generational transfer, and continued emissions (Dietz et al. 2013; Houde et al. 2011; Boisvert et al. 2019; Letcher et al. 2018). For Hg, body burdens even appear to be continuously increasing in most top predators in the central Arctic reaching up to 20-fold increases above baseline levels prior to the industrialization (Dietz et al. 2009, 2011; Rigét et al. 2011).
The abovementioned OHCs and Hg pose a health threat to Arctic top predators and northern people (Northerners) because most of the compounds, or their biotransformation metabolites, have structural similarities to hormones and may act via non-endocrine pathways as well. This causes these xenobiotic environmental compounds to have negative effects on immune and neuro-endocrine functioning, growth and development, reproduction, and general fitness (Grandjean and Landrigan 2006; Letcher et al. 2010; Sonne 2010). Since these compounds target different organ-tissues, exposure manifests in several health effects (Sonne 2010). Furthermore, seasonal cycles of energy requirement for fasting, breeding, lactation, and migration lead to increased metabolism of adipose tissue causing release and pulsed exposure to bioavailable contaminants circulating in the blood stream (Polischuk et al. 2002; Tartu et al. 2017). In polar bears, for example, up to 70% of the total OHC body burden is transported from mother to offspring during lactation, resulting in cub adipose tissue concentrations that are approximately three times higher than in their mother (Dietz et al. 2004, 2007; Muir et al. 2006; Polischuk et al. 2002; Bytingsvik et al. 2012). There is evidence that such high exposure poses a great risk to neonatal individuals during their critical period of development. As for other mammals, a female polar bears’ first litter is particularly vulnerable to OHC effects on normal development and growth as a higher percentage of the total body burden of contaminants in the mother is excreted to blood and milk (Letcher et al. 2010; Sonne 2010; Sonne et al. 2012). In a meta-study, it has recently been modeled that chlorinated and brominated OHCs , singularly or collectively, were better predictors for declines in population densities in 14 polar bear subpopulations than were human population density, harvest rate, and sea ice extension (Nuijten et al. 2016). From a population conservation point of view, contaminants that reduce pregnancy, fecundity, and survival are among the most important to monitor and assess in different Arctic subpopulations of polar bears and other predators.
4 Diseases
Polar bears have received considerable focus since loss of sea ice and access to seals as main prey has projected the species to go extinct around year 2100 (for instance: Amstrup et al. 2010; Castro de la Guardia et al. 2013; Whiteman 2018; Hamilton et al. 2014; Molnár et al. 2011). The current threats against polar bears are however echoed for other marine and terrestrial animals in the Arctic. But while the degree of bioaccumulation of environmental contaminants discriminates between animal groups, i.e., terrestrial vs. marine and high vs. low trophic levels, the threat of diseases is a universal one. Some disease agents readily cross trophic levels and main habitats; an example of such diseases is the so-called zoonoses, i.e., diseases that can infect Northerners via animal vectors (such as rabies). Zoonoses make up approximately 60% of all human infectious diseases. Zoonoses are therefore not rare, but more the rule than the exception of human infectious diseases (Jones et al. 2008).
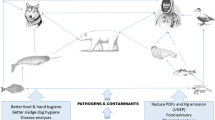
Diseases can be indicators of ecosystem health . For example, climate change is, in addition to posing an increased level of energetic stress (such as in the classic example of the polar bear and loss of sea ice and hunting grounds), also projected to increase the risk of both disease spread and virulence (Burek et al. 2008; Harvell et al. 2002; Shope 1992). This risk may be further accentuated by the concurrent exposure to environmental contaminants known to pose immunotoxic effects (Desforges et al. 2016). It applies to both climate change and environmental contaminants that their effects/presence is particularly accentuated in the Arctic (Bard 1999; ASIA 2004). There is as such a risk that disease-related mortality of Arctic mammals may increase – as well as a risk of zoonotic infections in Northerners – in the aftermath of anthropogenic environmental pollution and climate change (Fig. 2.3) (Jenssen et al. 2015; Sonne 2010). Regarding human zoonotic infections, this is a highlighted concern in many Arctic indigenous communities where it is still common practice to ingest wildlife foods raw and where wildlife game continues to be an important part of their diet (Chan et al. 1995). However, with infectious diseases, such as zoonoses, crossing species barriers and contrasting habitats, it points to the potential use of indicator species that they present a link between the environment, wildlife, and Northerners for monitoring the current challenges of overall ecosystem health: an indicator species such as the sled dog.

Often Northerners eat raw seafood, which increase the risk of disease transfer (zoonoses). Northwest Greenland, spring 1984. (Photo: R. Dietz)
Since the domestication of the dog 18,000–33,000 years ago (Thalmann et al. 2013; Wang et al. 2016), humans and dogs have not only shared food items, space, and security but to a high degree also pathogens. Today, more than 60 different pathogens, i.e., zoonoses , are known to be shared between dogs and people, and some of these have developed a synanthropic life cycle, i.e., a life cycle specialized for the human-dog relation (Craig et al. 2003). Large studies of zoonotic diseases in Northerners and wildlife have taken place in Canada and Alaska (Jenkins et al. 2013; Sonne et al. 2017a); however only a few studies have focused on zoonoses harbored by sled dogs (Salb et al. 2008), and sled dogs have not been included in a large-scale study of Arctic zoonoses and Northerners health risk before. Moreover, almost no studies have investigated the zoonotic infection pressure on Inuit of Greenland – Greenland still holds one of the largest populations of working sled dogs.
The status, character, and extent of Arctic zoonoses are still generally poorly elucidated; known diseases are moreover likely highly under-reported and other diseases are probably yet to be acknowledged/recognized (Parkinson et al. 2014; Gilbert et al. 2010). Some of the known diseases that are causes for significant human health concern are parasitic diseases such as trichinosis, echinococcosis, anisakiasis, toxoplasmosis, and toxocariasis, viral diseases like rabies, and bacterial infections such as brucellosis and clostridiosis (Magnaval et al. 2016; Jenkins et al. 2013; Rausch 1972 and others). The sled dog acts as a significant or potential host for most of these (Salb et al. 2008, Rausch 2003). The importance of dogs in the epidemiology of Arctic human zoonotic infections is exemplified by the parasitic disease echinococcosis. This was once an endemic disease in the Arctic affecting primarily those who hunted ungulates or were herders (Rausch 2003). Dogs act as host for all species of Echinococcus present in the Arctic and they are the definitive host for the dominant species causing human infections: E. granulosus (Gilbert et al. 2010, Rausch 2003). However, since the introduction of motorized snowmobiles and resultant decline in dog numbers, human cases of echinococcosis also declined drastically (Rausch 2003). The parasite and disease nevertheless still persist in the Arctic today, in particular where dogs are to be found, and in Canada, the highest human incidence of infection is found among Arctic indigenous communities (Gilbert et al. 2010; Himsworth et al. 2010).
As for infectious diseases of Arctic sled dogs per se, infectious epizootics of canine distemper virus (CDV, Morbillivirus) and parvoviral diarrhea probably top the list today – as they likely have since before the arrival of the Western societies. As such, canine distemper outbreaks have been recorded at least since 1860 in Greenland with concurrent reports of grave consequences for the owners and settlements that lost most of their dogs during such an event (Vernersen and Jensen 2018). These two viral diseases are not zoonotic; they are however carried by many other Arctic predators, in particular CDV which can be isolated from seals, polar bears, and foxes (Beineke et al. 2015). It is therefore unsurprising that there are strong indications of epizootics within dog populations being initiated by transmission from and further spread by wildlife (if not via main sled routes of travel) (Vernersen and Jensen 2018, Blixenkrone-Møller et al. 1989). This is an example of how the sled dogs may act as a mirror of Arctic ecosystem health via the health status of other top predators. Rabies is another example of a zoonosis with a broad host spectrum potentially including all Arctic mammals with the ability of causing mass die-offs among Inuit sled dog populations through transmission from wildlife (Tabel et al. 1974). Rabies is however no longer considered as great a concern as earlier with the advent of vaccines, general awareness, and control programs (Tabel et al. 1974). However, the occurrence of rabies is climate change sensitive, and problems may reoccur/rise, e.g., with the currently observed changes in rodent and fox populations (Parkinson and Butler 2005).
Summed up: Monitoring Arctic ecosystem health, including wildlife and human health, is an ongoing challenge of accelerating importance. The sled dogs represent an interesting link between Northerners, prey species, predators, and the Arctic environment. Sled dogs would be of value to achieve a high quantity and quality of samples from across the Arctic that could infer on the consequences and proper management of current environmental challenges that among others affect disease dynamics related to canine, wildlife, human, and overall ecosystem health.
5 Contaminants in Sled Dogs
Sporadic studies on sled dog health have mostly focused on Hg exposure (Harley et al. 2016; Lieske et al. 2011; Dunlap et al. 2007, 2011; Hansen and Danscher 1995; Hansen et al. 1989), wood smoke exposure (Montrose et al. 2015), hypospadias (penile malformation and PCB exposure) (Sonne et al. 2008), and infectious diseases which include zoonoses (Jenkins et al. 2013). At least six studies have been published on effects related to mercury exposure in sled dogs. These have been conducted in Alaska (Dunlap et al. 2007, 2011; Harley et al. 2016; Lieske et al. 2011) and Greenland (Hansen and Danscher 1995; Hansen et al. 1989). In Alaska, a cohort of sled dogs following a diet based on fish was investigated (Lieske et al. 2011). The study analyzed concentrations of Hg in blood and hair and found that sled dogs had a very high Hg exposure with a bioaccumulation similar to that seen in polar bears. Furthermore and based on concentrations in hair and hair-blood concentration ratios; the Hg toxicokinetics in sled dogs was found to be more similar to that of humans than that of laboratory rats, suggesting sled dogs as a potential biomonitoring species for exposure and effects of Hg in the Arctic environment. Another study by Dunlap et al. (2007) reported elevated Hg concentrations in hair of Arctic sled dogs as compared to other subpopulations of family dogs from temperate regions reflecting the high dietary intake of local Arctic communities. The study by Dunlap et al. (2011) is interesting as it showed that Yukon sled dogs suffer from Hg exposure since the dogs had antioxidant mechanisms that were induced due to high Hg concentrations, including the scavenging and neutralization of free radicals by vitamins. Such effects are known to lead to, e.g., vitamin deficiencies and immunotoxic effects affecting the overall health of the dogs. It is, however, important to study the confounding variables further before solid conclusions on toxicogenomics in sled dogs (and Arctic fox) can be drawn (Harley et al. 2016). Another study by Dunlap et al. (2012) showed that dietary habits of Yukon sled dogs are reflecting the ratio of healthy omega-3 and unhealthy omega-6 fatty acids as well as mercury exposure. This poses sled dogs as an ideal local biomonitoring species for human dietary exposure to fatty acids and mercury (Fig. 2.4).

Studies have revealed elevated Hg concentrations in hair of Arctic sled dogs as compared to other subpopulations of family dogs, which may cause them to suffer from Hg exposure health effects. Northwest Greenland, summer 2015. (Photo: R. Dietz)
In polar bears, high Hg concentrations have been associated with biological endpoints (Sonne 2010). It is likely that such impacts are also relevant for the health of Northerners as a similar tissue distribution and partitioning of Hg can be anticipated (AMAP 2015; Basu et al. 2009; Dietz et al. 2013; Sonne 2010). Sled dogs were also used as a sentinel model organism for Inuit exposure to investigate effects of local pollution from wood smoke (particles) on DNA hypomethylation and the risk of immunologically related gene expression (Montrose et al. 2015). The authors compared a wide range of dog kennels in order to reflect different exposure scenarios. Using dust trackers, the authors reported significant differences in smoke-related particle exposure among kennels, but there were no effects on DNA methylation and the functional expression of immunologically related canine genes. It is hard to conclude on this; however, the number of epigenetic studies is increasing for humans, and there are newer techniques that in the future may provide additional insights to such cause and effect relationships.
With respect to organic environmental contaminants; as part of a field survey in Tasiilaq on the southwest coast of Greenland, blood was sampled from four sled dogs in September 2014. The blood was analyzed for various PFASs (all PFSAs and PFCAs), and the concentrations of PFOS were by far the greatest of the analyzed compounds. This was in accordance with analyses of East Greenland polar bears and local Inuit (Dietz et al. 2008; Long et al. 2012; AMAP 2015) as well as in Hudson Bay polar bears (Boisvert et al. 2019; Letcher et al. 2018). From an ecological One Health point of view, it is therefore reasonable that sled dogs are good sentinel species for monitoring Arctic ecosystem health including that of Northerners (Sonne et al. 2017a).
6 Developmental Effects
A study of a male sled dog in Scoresby Sound in East Greenland in the year 2000 revealed that it was suffering from hypospadias which is a mal-closure of the ventral part of penis and urethra being part of the testicular dysgenesis syndrome (TDS). The TDS is characterized by hypospadias, testicular malign neoplasm (cancer), and cryptorchidism as described for humans by, e.g., Skakkebæk et al. (2001). The male dog from Scoresby Sound is the only reported case of hypospadias in Arctic mammals including polar bears and seals indicating that this is not a regular occurring event.
Previous studies have shown TDS being associated with exposure to environmental endocrine-disrupting chemicals, including PCBs, DDTs and genetics, which disrupt the testosterone production by Leydig cells as well as the sperm production by Sertoli cells (Edwards et al. 2006; Skakkebæk et al. 2001). Such effects are of course vital if occurring in high frequencies in, for example, male sled dogs, in polar bears, or in Inuit populations. In humans, however, surgical reconstruction is possible in the less severe cases. The authors did not have a chance to examine the dam for other causes such as organ pathology or tumors that could explain the case study of hypospadias in its offspring. In Greenland Inuit, the prevalence of hypospadias is four times lower than in, e.g., the USA (approximately 0.08%) despite Inuit peoples carrying high body burdens of endocrine-disrupting chemicals (AMAP 2015; Giwercman et al. 2006; Sonne et al. 2013). The reason for this is unknown; however, one explanation could be that the genotype of the Greenland Inuit is less sensitive to develop hypospadias (Giwercman et al. 2006).
Local Inuit people and researchers of High-Arctic Canada have previously reported that female sled dogs in Iqaluit (Nunavut) that were fed seal blubber vs. traditional dog pellets had a higher incidence of females in their litters (Sonne 2010). As part of an interview investigation of polar bear hunters in Scoresby Sound in East Greenland, similar observations of a higher female-male ratio were reported (Dietz et al. 2001; Sonne 2010). Reports of skewed offspring sex-ratio indicate an endocrine-disrupting feminization of the prenatal environment and fetuses, which may increase the female-male ratio of sled dogs as previously suggested for humans (Taylor et al. 2007; Tiido et al. 2006).
7 Controlled Studies
7.1 Immune Effects
In the literature, one large cohort study of sled dogs has been published with the aim to complement field studies of polar bears with relevant controlled studies in order to unravel how contaminant exposure affects health endpoints. For that purpose, the Greenland sled dog was selected as the model species of OHC exposure and health effects as its nutrition physiology is similar to that of polar bears (Sonne 2010). The study was performed in Aasiaat (Egedesminde) in West Greenland during year 2004–2006 in order to include the specific Arctic environmental physical parameters (Sonne 2010).
The parental P generation of dogs was eight sister pairs obtained from Inuit hunters (Fig. 2.5). The exposed group of dogs were fed minke whale (Balaenoptera acutorostrata) blubber with high OHC levels to mimic polar bear exposure according to Sonne (2010), while the control group was fed pig (Sus scrofa) fat with low OHC levels. Overall, effects from exposure to the complex mixture of OHCs can be divided into immune and endocrine effects, organ pathology, and effects on vitamin concentrations and distribution. It has been shown that the cocktail of environmental contaminants in the Arctic affected cellular, humoral, and complement parts of the immune system of the dogs in the controlled study (Sonne 2010) which has been supported by a recent review of marine mammals across taxa (Desforges et al. 2016). Such reduction in, e.g., lymphocyte proliferations and antibody production (IgG) may affect the ability to respond to intruding infectious pathogens as previously suggested for polar bears (Letcher et al. 2010) and Inuit and other northern peoples (AMAP 2015).

One of the female sled dogs included in the controlled Aasiaat study over the period 2004–2006. (Photo: C. Sonne)
7.2 Endocrinology
Several steroids (including sex steroid) and thyroid hormones were analyzed from the experimental study on Greenland sled dogs. These were analyzed in both the mothers (P generation) and the pup offspring (F1 generation). For the P generation, an increase was observed for all major steroid hormones in the group of females exposed to minke whale blubber, indicating slight overcompensation of the negative feedback system (Sonne et al. 2014b). In the F1 generation of pups, there seemed to be reduced concentrations of testosterone and testes size although that was based on a very low sample size (Sonne 2010). With respect to reproductive steroid hormones, Svalbard polar bears are exposed to similar OH-PCB levels as the sled dogs, which caused concentration-dependent reduction of plasma concentrations of pregnenolone and androstenedione in female polar bears (Gustavson et al. 2015). The authors suggested that CYP17 might be a potential target enzyme for these effects of OH-PCBs. Similarly for sled dogs the upregulated CYP activities may affect the concentrations of testosterone in the F1 generation (Sonne 2010). Analyses of thyroid hormones showed that concentrations of free T3, total T3, and T4 were lowest in the exposed female sled dogs after 10 months of age and that total T3 was lowest in the exposed group of F1 pups (Kirkegaard et al. 2011). Such relationships have also been found in Svalbard and East Greenland polar bears (Sonne 2010; Jenssen et al. 2015) as well as Inuits (AMAP 2015; Dallaire et al. 2008). Thus, in conclusion the sex steroid and thyroid hormone system appears to be affected by environmental contaminants in Arctic wildlife and in local Inuit.
7.3 Organ Pathology and Vitamins
Organ pathology has been investigated in Greenland polar bears for nearly two decades focusing on the liver, kidney, and thyroid gland (Sonne 2010; Sonne et al. 2011). Since the studies were of correlative nature, organ morphology was enrolled as an important parameter of the sled dog study. The results showed that lesions in the exposed group were similar to those of East Greenland polar bears and that up to 14% of the liver and ca. 60% of the kidney lesions could be ascribed to exposure to the environmental contaminants of the minke whale blubber (Sonne 2010). The liver and kidneys are very important organs for the overall metabolism and physiology, including metabolism and excretion of contaminants, and a reduced function is likely to have negative effects on mammals. Based on this it can be hypothesized that similar effects may exist in Inuit people eating at the same trophic level as polar bears and sled dogs. With respect to vitamins in the liver and kidney of the sled dogs, vitamins A, E, and D seemed to be affected by the exposure to environmental contaminants (Kirkegaard et al. 2010b; Sonne 2010; Sonne et al. 2014b). Similar findings have been reported for polar bears and Northerners (Sonne 2010; AMAP 2015) which are clear indices that there is a risk of suffering from hypovitaminosis with potential effects on immune system, reproduction, and development (Letcher et al. 2010; Sonne 2010).
8 Sled Dog Modeling
8.1 PBPK Modeling
Sonne et al. (2015, 2016) used a PBPK (physiologically based pharmacokinetic) model, also adapted for polar bears by Dietz et al. (2015), to calculate potential effects from contaminants on the sled dogs. This allowed for estimating risk quotients (RQs) based on critical daily dose (CDD) and critical body residues (CBR) obtained from controlled studies. In PBPK models, the body is subdivided into anatomical compartments representing individual organs or tissue groups. The transport of chemicals in the body is described by mass balance differential equations that incorporate blood flows, partitioning into compartments and tissue volumes. Numerous specific PBPK models have been used for modeling fate and disposition of a certain drug or for cancer risk assessment of industrial chemicals. However, only a few studies have reported on specific PBPK models for Arctic organisms (Cropp et al. 2014; Hickie et al. 2013). The PBPK model applied to the sled dogs for estimating fate, distribution, and CBR of contaminants has been presented by Gustavson et al. (2008) and Sonne et al. (2009, 2015).
The modeling showed that RQs in exposed, but not control, dogs exceeded immune effect thresholds, corroborating results of in vivo immunotoxicity in the sled dog cohort studies (Sonne et al. 2016). The modeling also showed that the exposed dogs were in risk of having effects on reproduction, which to some extent is supported by data in Kirkegaard et al. (2010a) and Sonne et al. (2016). In addition, PBPK modeling of East Greenland polar bears exposed to similar cocktail of environmental contaminants suggests that these are in risk of effects on immune and reproductive systems or even carcinogenic effects as supported by empirical field data (Dietz et al. 2015; Letcher et al. 2010; Jenssen et al. 2015). For Inuit people, there are no publications showing similar effects on reproduction, while there are studies that support effects on the immune system (AMAP 2015; Dallaire et al. 2004, 2006).
8.2 Energetics and Health Effects Modeling
As mentioned above, the most important contaminant effects in terms of individual and population-level health are those that affect demographic-related parameters such as reproduction, growth, morbidity, and mortality. Changes in these parameters are often difficult to measure directly in wildlife populations; thus biomarkers are used to study various health endpoints. The problem is that molecular and cellular biomarkers of contaminant effects are often difficult to link to broader and observable health implications in individuals and provide only a descriptive snapshot of a given endpoint measured at that particular time (Jager et al. 2010). Biology-based and process-driven approaches are therefore necessary to describe the physiological mechanisms that underpin the effects of stressors in animals. Dynamic energy budget (DEB) models may be useful in this context as these provide a framework to understand the flow of energy through living organisms as it relates to important physiological processes such as growth, development, reproduction, and maintenance (Kooijman and Bedaux, 1996). DEB models are particularly useful in ecotoxicology since they can explain life-history traits over the entire life cycle of organisms as a function of their environment and ecology, as well as anthropogenic stressors.
There have been no mechanistic or individual-based modeling studies performed to date using sled dogs. However, a recent study using DEB theory to model environmental contaminant exposure in mink (Mustela vison) provides an example of the type of modeling necessary to understand the implication of contaminants for organism health (Desforges et al. 2017). The model of physiological energy demands over the entire lifetime of the animal accurately predicted growth, development, and reproductive output when compared to captive mink studies. The model also included the toxicokinetics and dynamics of PCBs in mink, accurately replicating dose-response relationships for effects on growth and reproduction resulting from changes in energy allocation during ontogeny. DEB models have also been used to describe and predict fat accumulation in polar bears and how it can relate to starvation, reproduction, and population effects with increased nutritional stress due to climate change (Molnár et al. 2011, 2010). When linked to individual-based models (IBMs), energy budget models can be used to extrapolate contaminant effects to the population levels (Martin et al. 2013). This approach can be easily applied to sled dogs as their food intake, growth, and reproductive output can be closely monitored thereby providing the necessary data for DEB modeling. Lastly, since the model is based on physiology and the mechanisms underpinning contaminant effects, the results can be more confidently extrapolated to other top predator species such as polar bears, Arctic foxes, and Northerners exposed to the same contaminants.
8.3 Modeling of Disease and Contaminant Source
Based on the above sections, sled dogs can be employed in answering the need to biomonitor selected Arctic communities for effects of contaminants and diseases on the hormone and immune systems and general health and development. Doing so, sources and pathways of contaminant and disease exposure, mainly through the diet, should be identified. Since the exposure to contaminants and zoonotic diseases mainly occurs through the diet, understanding the dietary physiology and ecology of the sled dogs is an important aim, best done in a quantitative manner in order to allow for extrapolation to those wildlife species for which it acts as surrogate.
The analysis for stable carbon and nitrogen isotopes, typically in muscle and keratinous tissues such as hair, has become the backbone of quantitative chemical investigations of the foraging region and trophic position of mammalian top predators (Boecklen et al. 2011; Jardine et al. 2006). It has been successfully used to show how spatiotemporal plasticity in dietary habits influences the sources and intensity of contaminant exposure in polar bears (Cardona-Marek et al. 2009, McKinney et al. 2009, 2010, 2011, Routti et al. 2012) and Arctic foxes (Fuglei et al. 2007). At the present day, stable isotope mixing models (Parnell et al. 2013) are a powerful modeling tool that can help elucidate how the diet of an individual sled dog is composed of different feed items or species. Hence, it will allow identifying dietary items and habits that lie at the source of increased exposure and risk to specific contaminants or diseases of interest. Furthermore, this chemical methodology of measuring stable isotopes in bulk tissues, i.e., a homogenized tissue, has recently been refined to the precise measurement of individual compounds, such as the measurement of stable carbon isotopes of individual fatty acids and stable carbon and nitrogen isotopes of individual amino acids (Iverson et al. 2004; McMahon et al. 2013). This approach has in fact already showed to be promising and further increase the accuracy and power of reconstructive investigations of temporal variation in polar bear dietary habits and its impact on contaminant exposure (McKinney et al. 2013).
Despite the above promising conceptual quantitative framework and its suggested applicability for using sled dogs as sentinel species for monitoring Arctic ecosystem health, there have been no efforts to date to use either bulk or compound-specific stable isotope measurements for quantitative diet reconstructions in sled dogs. Nonetheless, these approaches have large potential to provide the quantitative ecological input required for effect-oriented modeling, such as the above-mentioned PBPK and DEB modeling.
8.4 Genetic Modeling
Studies of genetic makers or even full nuclear genomes have revolutionized the field of evolutionary biology by shedding light over phylogeny and population structure. However, gene and genomic information of an individual does not only constitute the specific individual’s ancestry, but it also first codes for an individual’s abilities and physiological operation. In the genome, the exome coding regions are throughout life expressed as RNA transcription, to maintain and regulate specific cell and overall body functions. The transcription of messenger RNA and microRNA, respectively, facilitates protein expression in a cell and various regulation of expression, and these expressions can vary as response to external influences and stress (Wasaki et al. 2003; Feder and Walser 2005; Todgham and Hofmann 2009; Jozefczuk et al. 2010; Chapman et al. 2011; Pujolar et al. 2012; Lemay et al. 2013). The relatively new field of transcriptomics made accessible by sequencing technology targeting RNA molecules is increasingly expanding (Wang et al. 2009; Martin and Wang 2011). Although no environmental change or stress-related transcriptomic investigation of sled dogs has been made to date, it is a potent methodology for future work.
9 Other Biomonitoring Species: The Arctic Fox
In a similar study to the one on sled dogs in West Greenland, domesticated Arctic foxes , also fed minke whale blubber, were used to study biological effects from environmental contaminants in the Arctic as well (Hallanger et al. 2012; Pedersen et al. 2015; Rogstad et al. 2017; Sonne 2010; Sonne et al. 2017b). Similar for the sled dogs, the foxes were exposed to the complex mixture of organic contaminants, and effects on organ pathology, endocrine system, and vitamin concentrations and distribution were investigated (Hallanger et al. 2012; Rogstad et al. 2017; Sonne et al. 2017b). In the exposed group of Arctic foxes, higher hepatic CYP activity may have led to increased testosterone metabolism reducing the blood concentrations and combined with increased oxidative stress also lowered vitamin E (Helgason et al. 2013; Sonne et al. 2017b). Similar liver and kidney lesions as those found in the sled dogs were found and ascribed to the chemical cocktails of environmental contaminants in the minke whale blubber (Sonne 2010). In addition, lesions were found in the thyroid glands, which may be related to changes in thyroid hormone concentrations that again can be a support for the study of thyroid lesions in East Greenland polar bears (Sonne 2010; Sonne et al. 2011).
10 Inuit People and One Health
Due to their reliance on the marine food resources , Inuit people are heavily exposed to environmental chemicals and mercury (AMAP 2015; Bonefeld-Jørgensen 2010). This is of extreme concern and it is therefore important that exposure to toxic anthropogenic pollutants is closely monitored continuously in order to discover if there are significant changes. This is undertaken via AMAP showing that, e.g., East Greenland hunters are exposed to high PCB concentrations due to ingestion of seals, polar bears, killer whales, and narwhals (AMAP 2011; Bonefeld-Jørgensen 2010; Dietz et al. 2013; Sonne et al. 2013). As a supplement to the AMAP monitoring program, a PBPK model has been built as a first step to estimate potential health effects in Greenlanders (Sonne et al. 2014c). For the Greenlanders, significant correlations were found between chemically analyzed contaminant blood concentrations and calculated daily intake of OHCs; and the PBPK model predicted blood concentrations of a factor 2–3 within the actual measured values. Furthermore, the model itself estimated that the most important excretion route for higher chlorinated PCBs in Greenlanders was in fact via alveolar excretion and not as previously suggested via feces or urine. This combination of PBPK modeling, monitoring of Inuit people, and studies of sled dogs would be a very strong combination of tools to further understand and monitor human exposure to contaminants in the Arctic .
11 Conclusions
Based on the accumulated scientific data, sled dogs have proved to be a good model for assessing the health of top predators and Northerners in the Arctic. Sled dogs are widely distributed and when compared to polar bears and Inuit people they are similarly exposed and show similar correlations to important health indicators. We therefore recommend that ecosystem health assessments in the Arctic, including that of Northerners, should also be monitored using sled dogs combined with modeling tools such as PBPK and DEB. Performing this in a circumpolar perspective will further increase our understanding and monitoring possibilities of ecosystem health and human exposure to contaminants, diseases, and climate change in the Arctic.
References
AMAP (2011) What are the toxicological effects of mercury in Arctic biota? In: AMAP assessment 2011: mercury in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo, pp 113–138
AMAP (2014) Trends in Stockholm convention persistent organic pollutants (POPs) in arctic air, human media and biota. AMAP technical report to the Stockholm convention. AMAP Technical Report No. 7 (2014) Arctic Monitoring and Assessment Programme (AMAP), Oslo
AMAP (2015) Human health in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo. vii + 165 pp
AMAP assessment (2018) Biological effects of contaminants on arctic wildlife and fish. Arctic Monitoring and Assessment Programme (AMAP), Tromsø. vii+84pp
Amstrup SC, Deweaver ET, Douglas DC, Marcot BG, Durner GM, Bitz CM, Bailey DA (2010) Greenhouse gas mitigation can reduce sea-ice loss and increase polar bear persistence. Nature 468:955–958
ASIA, Assessment ACI (2004) Impacts of a warming Arctic-Arctic climate impact assessment by Arctic climate impact assessment pp. 144. ISBN 0521617782. Cambridge University Press, Cambridge, UK
Bard SM (1999) Global transport of anthropogenic contaminants and the consequences for the Arctic marine ecosystem. Marine Poll Bull 38(5):356–379
Basu N, Scheuhammer AM, Sonne C, Letcher RJ, Dietz R (2009) Is dietary mercury of neurotoxicological concern to wild polar bears (Ursus maritimus)? Environ Toxicol Chem 28:133–140
Bechshøft TØ, Jakobsen J, Sonne C, Dietz R (2011) Distribution of vitamin A (retinol) and E (α-tocopherol) in polar bear kidney: implications for biomarker studies. Sci Total Environ 409:3508–3511
Beineke A, Baumgärtner W, Wohlsein P (2015) Cross-species transmission of canine distemper virus—an update. One Health 1:49–59
Blixenkrone-Møller M, Bohm J, Lund E (1989) Udbrud af hundesyge blandt slædehunde i Nordgrønland. Dansk Veterinær Tidsskrift 72:488–497
Boecklen WJ, Yarnes CT, Cook BA, James AC (2011) On the use of stable isotopes in trophic ecology. Ann Review Ecol Evolut System 42:411–440
Boisvert G, Sonne C, Rigét FF, Dietz R, Letcher RJ (2019) Bioaccumulation and biomagnification of perfluoroalkyl acids and precursors in East Greenland polar bears and their ringed seal prey. Environ Pollut Part B 252:1335–1343
Bonefeld-Jørgensen C (2010) Biomonitoring in Greenland: human biomarkers of exposure and effects - a short review. Rural Remote Health 10:1362
Burek KA, Gulland FMD, O’Hara TM (2008) Effects of climate change on Arctic marine mammal health. Ecol Appl 18:126–134
Burger J, Gochfeld M (2001) On developing bioindicators for human and ecological health. Environ Monit Assessment 66:23–46
Butt CM, Berger U, Bossi R, Tomy GT (2010) Levels and trends of poly- and perfluorinated compounds in the Arctic environment. Sci Total Environ 408:2936–2965
Bytingsvik J, Lie E, Aars J, Derocher AE, Wiig Ø, Jenssen BM (2012) PCBs and OH-PCBs in polar bear mother-cub pairs: a comparative study based on plasma levels in 1998 and 2008. Sci Total Environ 417-418:117–128
Cardona-Marek T, Knott KK, Meyer BE, O’Hara TM (2009) Mercury concentrations in southern Beaufort Sea polar bears: variation based on stable isotopes of carbon and nitrogen. Environ Toxicol Chem 28:1416–1424
Castro de la Guardia L, Derocher AE, Myers PG, Terwisscha van Scheltinga AD, Lunn NJ (2013) Future sea ice conditions in Western Hudson Bay and consequences for polar bears in the 21st century. Glob Chang Biol 19:2675–2687
Cavalieri DJ, Parkinson CL (2012) Arctic Sea ice variability and trends, 1979–2010. Cryosphere 6:881–889
Chan HM, Kim C, Khoday K, Receveur O, Kuhnlein HV (1995) Assessment of dietary exposure to trace metals in Baffin Inuit food. Environ Health Perspec 103(7–8):740
Chapman RW, Mancia A, Beal M, Veloso A, Rathburn C, Blair A, Holland AF, Warr GW, Didinato GU, Sokolova IM, Wirth EF (2011) The transcriptomic responses of the eastern oyster, Crassostrea virginica, to environmental conditions. Mol Ecol 20:1431–1449
Craig PS, Rogan MT, Campos-Ponce M (2003) Echinococcosis: disease, detection and transmission. Parasitology 127:S5–S20
Cropp R, Nash SB, hawker D (2014) A model to resolve organochlorine pharmacokinetics in migrating humpback whales. Environ Toxicol Chem 9999:1–12
Dallaire F, Dewailly E, Muckle G, Vezina C, Jacobson SW, Jacobson JL, Ayotte P (2004) Acute infections and environmental exposure to organochlorines in Inuit infants from Nunavik. Environ Health Perspect 112:1359–1364
Dallaire F, Dewailly E, Vezina C, Muckle G, Weber JP, Bruneau S, Ayotte P (2006) Effect of prenatal exposure to polychlorinated biphenyls on incidence of acute respiratory infections in preschool Inuit children. Environ Health Perspect 114:1301–1305
Dallaire R, Dewailly E, Ayotte P, Muckle G, Laliberte C, Bruneau S (2008) Effects of prenatal exposure to organochlorines on thyroid hormone status in newborns from two remote coastal regions in Quebec, Canada. Environ Res 108:387–392
de Wit CA, Herzke D, Vorkamp K (2010) Brominated flame retardants in the Arctic environment--trends and new candidates. Sci Total Environ 408:2885–2918
Desforges JPW, Sonne C, Levin M, Siebert U, De Guise S, Dietz R (2016) Immunotoxic effects of environmental pollutants in marine mammals. Environ Int 86:126–139
Desforges JP, Sonne C, Dietz R (2017) Using energy budgets to combine ecology and toxicology in a mammalian sentinel species. Sci Rep 7:46267. https://doi.org/10.1038/srep46267
Dietz R, Sonne-Hansen C, Born EW, Sandell HT, Sandell B (2001) Aberrant polar bears in East Greenland. An interview investigation, 1999 (with english summary). National Environmental Research Institute, Technical Report no. 359, pp. 50. www.neri.dk
Dietz R, Rigét FF, Sonne C, Letcher RJ, Born EW, Muir DCG (2004) Seasonal and temporal trends in Polychlorinated biphenyls and Organochlorine Pesticides in East Greenland polar bears (Ursus maritimus), 1990-2001. Sci Total Environ 331:107–124
Dietz R, Rigét FF, Sonne C, Muir DCG, Backus S, Born EW, Kirkegaard M, Letcher RJ (2007) Age and seasonal variability of polybrominated diphenyl ethers in free-ranging East Greenland polar bears (Ursus maritimus). Environ Pollut 146:177–184
Dietz R, Bossi R, Rigét FR, Sonne C, Born EW (2008) Increasing perfluorinated acids in East Greenland polar bears (Ursus maritimus) - a new toxic threat to the Arctic bears. Environ Sci Technol 42:2701–2707
Dietz R, Outridge PM, Hobson KA (2009) Anthropogenic contribution to mercury levels in present-day Arctic animals – a review. Sci Total Envirion 407:6120–6131
Dietz R, Born EW, Rigét FF, Sonne C, Aubail A, Basu N (2011) Temporal trends and future predictions of mercury concentrations in Northwest Greenland polar bear (Ursus maritimus) hair. Environ Sci Technol 45:1458–1465
Dietz R, Basu N, Braune B, O’Hara T, Scheuhammer TM, Sonne C, Andersen M, Andreasen C, Andriashek D, Asmund G, Aubail A, Baagøe H, Born EW, Chan HM, Derocher AE, Grandjean P, Knott K, Kirkegaard M, Lunn N, Messier F, Obbard M, Olsen MT, Peacock E, Renzoni A, Rigét F, Skaare JU, Stern G, Stirling I, Taylor M, Wiig Ø, Aars J (2013) What are the toxicological effects of mercury in Arctic biota? Sci Total Environ 443:775–790
Dietz R, Gustavson K, Sonne C, Desforges JP, Rigét FF, McKinney MA, Letcher RJ (2015) Physiologically-based pharmacokinetic modelling of immune, reproductive and carcinogenic effects from contaminant exposure in polar bears (Ursus maritimus). Environ Res 140:45–55
Dunlap KL, Reynolds AJ, Bowers PM, Duffy LH (2007) Hair analysis in sled dogs (Canis lupus familiaris) illustrates a linkage of mercury exposure along the Yukon River with human subsistence food systems. Sci Total Environ 385:80–85
Dunlap KL, Reynolds AJ, Gerlach SC, Duffy LK (2011) Mercury interferes with endogenous antioxidant levels in Yukon River subsistence-fed sled dogs. Environ Res Lett 6:044015
Dunlap KL, Reynolds AJ, Duffy LK, Gerlach SC, Loring PA, Cleroux M, Godin JP (2012) Selected plasma fatty acid levels in subsistence fed sled dogs along the Yukon River: a pilot study for biomonitoring. Polar Rec 48:177–183
Edwards TM, Moore BC, Guillette LJ (2006) Reproductive dysgenesis in wildlife: a comparative view. Int J Androl 29:109–121
Feder ME, Walser JC (2005) The biological limitations of transcriptomics in elucidating stress and stress responses. J Evol Biol 18(4):901–910
Fuglei E, Bustnes JO, Hop H, Mørk T, Bjornfoth H, van Bavel B (2007) Environmental contaminants in Arctic foxes (Alopex lagopus) in Svalbard: relationships with feeding ecology and body condition. Environ Pollut 146:128–138
Gilbert NL, Dare OK, Libman MD, Muchaal PK, Ogden NH (2010) Hospitalization for trichinellosis and echinococcosis in Canada, 2001-2005: the tip of the iceberg? Can J Public Health 101:337–340
Giwercman YL, Kleist KE, Giwercman A, Giwercman C, Toft G, Bonde JP, Pedersen HS (2006) Remarkably low incidence of hypospadias in Greenland despite high exposure to endocrine disrupters; possible protective effect of androgen receptor genotype. Pharmacogenet Genomics 16:375–377
Grandjean P, Landrigan PJ (2006) Review: developmental neurotoxicity of industrial chemicals. Lancet 16:2167–2178
Gustavson K, Petersen G, Sonne C, Dietz R, Birkved M, Rigét FF (2008) Environmental contaminants in East Greenland polar bears: an evaluation of the potential adverse toxic effects (Ursus maritimus) by use of critical body burden residue concept. Secretary for Arctic Monitoring and Assessment Programme and Danish EPA. www.dhigroup.com
Gustavson L, Ciesielski TM, Bytingsvik J, Styrishave B, Hansen M, Lie E, Aars J, Jenssen BM (2015) Hydroxylated polychlorinated biphenyls decrease circulating steroids in female polar bears (Ursus maritimus). Environ Res 138:191–201
Hallanger IG, Jorgensen EH, Fuglei E, Ahlstrom O, Muir DCG, Jenssen BM (2012) Dietary contaminant exposure affects plasma testosterone, but not thyroid hormones, vitamin a, and vitamin E, in male juvenile foxes (Vulpes lagopus). J Toxicol Environ Health. 75A 75:1298–1313
Hamilton SG, Guardia LCDL, Derocher AE, Sahanatien V, Tremblay B, Huard D (2014) Projected polar bear sea ice habitat in the Canadian Arctic Archipelago. PLoS One 9:e113746
Hansen JC, Danscher G (1995) Quantitative and qualitative distribution of mercury in organs from arctic sled dogs: an atomic absorption spectrophotometric and histochemical study of tissue samples from natural long-termed high dietary organic mercury-exposed dogs from Thule, Greenland. Pharmacol Toxicol 77:189–195
Hansen JC, Reske-Nielsen E, thorlacius-ussing O, Rungby J, Danscher G (1989) Distribution of dietary mercury in a dog. Quantitation and localization of total mercury in organs and central nervous system. Sci Total Environ 78:23–43
Harley JR, Bammler TK, Farin FM, Beyer RP, Kavanagh TJ, Dunlap KL, Knott KK, Ylitalo GM, O’Hara TM (2016) Using domestic and free-ranging Arctic canid models for environmental molecular toxicology research. Environ Sci Technol 50:1990–1999
Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld RS, Samuel MD (2002) Climate warming and disease risks for terrestrial and marine biota. Science 296:2158–2162
Helgason LB, Wolkers H, Fuglei E, Ahlstrom O, Muir DGC, Jørgensen EH (2013) Seasonal emaciation causes tissue redistribution and an increased potential for toxicity of lipophilic pollutants in farmed Arctic fox (Vulpes lagopus). Environ Toxicol Chem 32:1784–1792
Hickie BE, Cadieux MA, Riehl KN, Bossart GD, Alava JJ, Fair PA (2013) Modeling PCB-bioaccumulation in the Bottlenose Dolphin (Tursiops truncatus): estimating a dietary threshold concentration. Environ Sci Technol 47:12314–12324
Himsworth CG, Jenkins E, Hill JE, Nsungu M, Ndao M, Andrew Thompson RC, Covacin C et al (2010) Emergence of sylvatic Echinococcus Granulosus as a parasitic zoonosis of public health concern in an indigenous Community in Canada. Am J Tropical Med Hygiene 82:643–645
Hoferkamp L, Hermanson MH, Muir DGC (2010) Currently used pesticides in arctic media, 2000-2007. Sci Total Environ 408:2985–2994
Houde M, De Silva AO, Muir DCG, Letcher RJ (2011) An updated review of monitoring and accumulation of perfluorinated compounds in aquatic biota. Environ Sci Technol 45:7962–7973
Hung H, Kallenborn R, Breivik K, Su YS, Brorstrom-Lunden E, Olafsdottir K, Thorlacius JM, Leppanen S, Bossi R, Skov H, Mano S, Patton GW, Stern G, Sverko E, Fellin P (2010) Atmospheric monitoring of organic pollutants in the Arctic under the Arctic Monitoring and Assessment Programme (AMAP): 1993–2006. Sci Total Environ 408:2854–2873
Iverson SJ, Field C, Bowen WD, Blanchard W (2004) Quantitative fatty acid signature analysis: a new method of estimating predator diets. Ecol Monogr 74:211–235
Jager T, Vandenbrouck T, Baas J, De Coen WM, Kooijman SALM (2010) A biology-based approach for mixture toxicity of multiple endpoints over the life cycle. Ecotoxicology 19:351–361
Jardine TD, Kidd KA, Fisk AT (2006) Applications, considerations, and sources of uncertainty when using stable isotope analysis in ecotoxicology. Environ Sci Technol 40:7501–7511
Jenkins EJ, Castrodale LJ, de Rosemond SJC, Dixon BR, Elmore SA, Gesy KM, Hoberg EP, Polley L, Schurer JM, Simard M, Thompson RCA (2013) Tradition and transition: parasitic zoonoses of people and animals in Alaska, northern Canada, and Greenland. Adv Parasitol 82:33–204
Jenssen BM, Dehli Villanger G, Gabrielsen KM, Bytingsvik J, Ciesielski TM, Sonne C, Dietz R (2015) Anthropogenic flank attack on polar bears: interacting consequences of climate warming and pollutant exposure. Frontiers Ecol Evol 3:1–7
Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, Daszak P (2008) Global trends in emerging infectious diseases. Nature 451:990–993
Jozefczuk S, Klie S, Catchpole G, Szymanski J, Cuadros-Inostroza A, Steinhauser D, Selbig J, Willmitzer L (2010) Metabolomic and transcriptomic stress response of Escherichia coli. Mol Systems Biol 6:364
Kirkegaard M, Sonne C, Dietz R, Jenssen BM, Leifsson PS, Jensen JEB, Letcher RJ (2010a) Testosterone concentrations and male genital organ morphology in Greenland sled dogs (Canis familiaris) dietary exposed to organohalogen contaminants. Toxicol Environ Chem 92:955–967
Kirkegaard M, Sonne C, Dietz R, Jenssen BM, Letcher RJ (2010b) Organohalogens in a whale blubber supplemented diet reduce hepatic retinol and renal tocopherol concentrations in Greenland sled dogs. J Toxicol Environ Health Part A 73:773–786
Kirkegaard M, Sonne C, Dietz R, Letcher RJ, Jensen AL, Jenssen BM, Grandjean P (2011) Alterations in thyroid hormone status in Greenland sled dogs exposed to naturally OHC contaminated whale blubber. Ecotoxicol Environ Safe 74:157–163
Kooijman S, Bedaux JJM (1996) The analysis of aquatic toxicity data. VU University Press, Amsterdam
Lemay MA, Henry P, Lamb CT, Robson KM, Russello MA (2013) Novel genomic resources for a climate change sensitive mammal: characterization of the American pika transcriptome. BMC Genomics 14:311
Letcher RJ, Bustnes JO, Dietz R, Jenssen BM, Jørgensen EH, Sonne C, Verreault J, Vijayan MM, Gabrielsen GW (2010) Effects assessment of persistent organohalogen contaminants in Arctic wildlife and fish. Sci Total Environ 408:2995–3043
Letcher RJ, Morris AD, Dyck M, Sverko E, Reiner E, Blair DAD, Chu SG, Shen L (2018) Legacy and (re)emerging halogenated persistent organic pollutants in polar bears from a contamination hotspot in the Arctic, Hudson Bay Canada. Sci Total Environ 610-611:121–136
Lieske CL, Moses SK, Castellini JM, Klejka J, Hueffer K, O’Hara TM (2011) Toxicokinetics of mercury in blood compartments and hair of fish-fed sled dogs. Acta Vet Scand 53:66
Long M, Bossi R, Bonefeld-Jørgensen EC (2012) Levels and temporal trends of perfluoroalkyl acids in human serum from Greenland Inuit. Int J Circumpolar Health 71:17998
Magnaval JF, Leparc-Goffart I, Gibert M, Gurieva A, Outreville J, Dyachkovskaya P et al (2016) A serological survey about zoonoses in the Verkhoyansk area, Northeastern Siberia (Sakha Republic, Russian Federation). Vector-Borne Zoonotic Dis 16(2):103–109
Martin JA, Wang Z (2011) Next-generation transcriptome assembly. Nat Rev Genet 12:671–682
Martin BT, Jager T, Nisbet RM, Preuss TG, Hammers-Wirtz M, Grimm V (2013) Extrapolating ecotoxicological effects from individuals to populations: a generic approach based on Dynamic Energy Budget theory and individual-based modelling. Ecotoxicology 22:574–583
Mckinney MA, Peacock E, Letcher RJ (2009) Sea ice-associated diet change increases the levels of chlorinated and brominated contaminants in polar bears. Environ Sci Technol 43:4334–4339
McKinney MA, Stirling I, Lunn NJ, Peacock E, Letcher RJ (2010) The role of diet on long-term concentration and pattern trends of brominated and chlorinated contaminants in western Hudson Bay polar bears, 1991-2007. Sci Total Environ 408:6210–6222
McKinney MA, Letcher RJ, Aars J, Born EW, Branigan M, Dietz R, Evans TJ, Gabrielsen GW, Muir DCG, Peacock E, Sonne C (2011) Regional contamination versus regional diet differences: understanding geographic variation in brominated and chlorinated contaminant levels in polar bears. Environ Sci Technol 45:896–902
McKinney MA, Iverson SJ, Fisk AT, Sonne C, Rigét FF, Letcher RJ, Arts MT, Born EW, Rosing-Asvid A, Dietz R (2013) Global change effects on the long-term feeding ecology and contaminant exposures of East Greenland polar bears. Glob Chang Biol 19:2360–2372
McMahon KW, Hamady LL, Thorrold SR (2013) A review of ecogeochemistry approaches to estimating movements of marine animals. Limnol Oceanogr 58:697–714
Molnár PK, Derocher AE, Thiemann GW, Lewis MA (2010) Predicting survival, reproduction and abundance of polar bears under climate change. Biol Conserv 143:1612–1622
Molnár PK, Derocher AE, Klanjscek T, Lewis MA (2011) Predicting climate change impacts on polar bear litter size. Nat Commun 2:186
Montrose L, Noonan CW, Cho YH, Lee J, Harley J, O’Hara TM, Cahill C, Ward TJ (2015) Evaluating the effect of ambient particulate pollution on DNA methylation in Alaskan sled dogs: potential applications for a sentinel model of human health. Sci Total Environ 512–513:489–494
Muir DCG, Backus S, Derocher AE, Dietz R, Evans TJ, Gabrielsen GW, Nagy J, Norstrom RJ, Sonne C, Stirling I, Taylor MK, Letcher RJ (2006) Brominated flame retardants in polar bears (Ursus maritimus) from Alaska, the Canadian Arctic, East Greenland, and Svalbard. Environ Sci Technol 40:449–455
Nuijten RJM, Hendriks AJ, Jenssen BM, Schipper AM (2016) Circumpolar contaminant concentrations in polar bears (Ursus maritimus) and potential population-level effects. Environ Res 151:50–57
Outridge PM, Dietz R, Amyot M, Barkay T, Basu N, Berg T, Braune B, Carrie J, Chételat J, Cole A, Constant P, Dastoor A, Dommergue A, Donaldson SG, Douglas T, Durnford D, Evans M, Ferrari C, Gaden A, Gantner AK, Gantner N, Goodsite M, Hedman J, Hintelmann H, Hobson K, Johnson M, Kirk J, Kroer N, Krümmel E, Larose C, Lean D, Leech T, Letcher RJ, Loseto L, Macdonald RW, Muir DCG, Munthe J, Nielsen TG, O’Hara T, Pacyna J, Poissant L, Poulain A, Rigét F, Rognerud S, Ryzhkov A, Scheuhammer T, Skov H, Sonne C, Sørensen S, Steenhuisen F, Steffen A, Stern G, Stow J, Sundseth K, Travnikov O, Verta M, Wang F, Wängberg I, Wilson SJ, Zdanowicz C (2011) Mercury in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo. xiv + 193 pp
Parkinson AJ, Butler JC (2005) Potential impacts of climate change on infectious diseases in the Arctic. Int J Circumpolar Health 64(5):478–486
Parkinson AJ, Evengard B, Semenza JC, Ogden N, Børresen ML, Berner J et al (2014) Climate change and infectious diseases in the Arctic: establishment of a circumpolar working group. Int J Circumpolar Health 73(1):25163
Parnell AC, Phillips DL, Bearhop S, Semmens BX, Ward EJ, Moore JW, Jackson AL, Grey J, Kelly DJ, Inger J (2013) Bayesian stable isotope mixing models. Environmetrics 24:387–399
Pedersen KE, Styrishave B, Sonne C, Dietz R, Jenssen BM (2015) Accumulation and potential health effects of organohalogenated compounds in the Arctic fox (Vulpes lagopus) - a review. Sci Total Environ 502:510–516
Pedro S, Boba C, Dietz R, Sonne C, Rosing-Asvid A, Provatas A, McKinney MA (2017) Blubber-depth distribution and bioaccumulation of PCBs and organochlorine pesticides in Arctic-invading killer whales. Sci Total Environ 601-602:237–246
Polischuk SC, Norstrom RJ, Ramsay MA (2002) Body burdens and tissue concentrations of organochlorines in polar bears (Ursus maritimus) vary during seasonal fasts. Environ Pollut 118:29–39
Pujolar JM, Marino IA, Milan M, Coppe A, Maes GE, Capoccioni F, Ciccotti E, Bervoets L, Covaci A, Belpaire C, Cramb G (2012) Surviving in a toxic world: transcriptomics and gene expression profiling in response to environmental pollution in the critically endangered European eel. BMC Genomics 13:507
Rausch RL (1972) Observations on some natural-focal zoonoses in Alaska. Arch Environ Health 25(4):246–252
Rausch RL (2003) Cystic echinococcosis in the Arctic and Sub-Arctic. Parasitology 127:S73–S85
Rigét FF, Braune B, Bignert A, Wilson S, Aars J, Andersen M, Asmund G, Aubail A, Dam M, Dietz R, Evans M, Evans T, Gamberg M, Gantner N, Green N, Gunnlaugsdóttir H, Kannan K, Letcher RJ, Muir D, Ólafsdóttir K, Renzoni A, Roach P, Sonne C, Stern G, Wiig Ø (2011) Temporal trends of Hg in Arctic biota, an update. Sci Total Environ 409:3520–3526
Rigét F, Vorkamp K, Bossi R, Sonne C, Letcher RJ, Dietz R (2016) Twenty years of monitoring of persistent organic pollutants in Greenland biota. A review. Environ Pollut 217:114–123
Rigét FF, Bignert A, Brause B, Dam M, Dietz R, Evans M, Green N, Gunnlaugsdóttir H, Hoydal KS, Kucklick J, Letcher RJ, Muir DCG, Schuur S, Sonne C, Stern G, Tomy G, Vorkamp K, Wilson S (2019) Temporal trends of persistent organic pollutants in Arctic marine and freshwater biota. Sci Total Environ 649:99–110
Rogstad TW, Sonne C, Villanger GD, Ahlstrøm Ø, Fuglei E, Muir D, Jørgensen E, Jenssen BM (2017) Exposure to high PCB diet reduces alpha-tochopherol levels and disrupts thyroid hormone balance in blue foxes (Alopex lagopus). Environ Res 154:284–290
Routti H, Letcher RJ, Born EW, Branigan M, Dietz R, Evans TJ, McKinney MA, Peacock E, Sonne C (2012) Influence of carbon and lipid sources on variation of mercury and other trace elements in polar bears (Ursus maritimus). Environ Toxicol Chem 31:2739–2747
Salb AL, Barkema HW, Elkin BT, Thompson RA, Whiteside DP, Black SR et al (2008) Dogs as sources and sentinels of parasites in humans and wildlife, northern Canada. Emerg Infect Dis 14(1):60
Shope RE (1992) 1992. Impacts of global climate change on human health: spread of infectious disease. In: Majumdar SK, Kalkstein LS, Yarnal B, Miller EW, Rosenfeld LM, Easton PA (eds) Global climate change: implications, challenges and mitigation measures. The Pennsylvania Academy of Science, USA, pp 263–370
Skakkebæk NE, Rajpert-De Meyts E, Main KM (2001) Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects. Hum Reprod 16:972–978
Sonne C (2010) Health effects from long-range transported contaminants in Arctic top predators: an integrated review based on studies of polar bears and relevant model species. Environ Int 36:461–491
Sonne C, Dietz R, Born EW, Leifsson PS, Andersen S (2008) Is there a link between hypospadias and organochlorine exposure in East Greenland sled dogs (Canis familiaris)? Ecotoxicol Environ Saf 69:391–395
Sonne C, Gustavson K, Rigét FF, Dietz R, Birkved M, Letcher RJ, Muir DCG, Bossi R, Vorkamp K, Born EW, Petersen G (2009) Reproductive performance in East Greenland polar bears (Ursus maritimus) may be affected by organohalogen contaminants as shown by critical body residue modelling and risk quotients estimation. Chemosphere 77:1558–1568
Sonne C, Leifsson PS, Iburg T, Dietz R, Born EW, Letcher RJ, Kirkegaard M (2011) Thyroid gland lesions in organohalogen contaminated East Greenland polar bears (Ursus maritimus). Toxicol Environ Chem 93:789–805
Sonne C, Letcher RJ, Bechshøft TØ, Rigét FF, Muir DCG, Leifsson PS, Born EW, Hyldstrup L, Basu N, Kirkegaard M, Dietz R (2012) Two decades of biomonitoring polar bear health in Greenland: a review. Acta Vet Scand 54:S15
Sonne C, Dietz R, Letcher RJ (2013) Chemical cocktail party in East Greenland: a first time evaluation of human organohalogen exposure from consumption of ringed seal and polar bear tissues and possible health implications. Toxicol Environ Chem 95:853–859
Sonne C, Dietz R, Rigét FF, Letcher RJ, Munk Pedersen K, Styrishave B (2014a) Steroid hormones in blood plasma from Greenland sled dogs (Canis familiaris) dietary exposed to organohalogen polluted minke whale (Balaenoptera acuterostrata) blubber. Toxicol Environ Chem 96:273–286
Sonne C, Kirkegaard M, Jacobsen J, Jenssen BM, Letcher RJ, Dietz R (2014b) Altered 25-hydroxyvitamin D3 in liver tissue from Greenland sled dogs (Canis familiaris) dietary exposed to organohalogen polluted minke whale (Balaenoptera acuterostrata) blubber. Ecotoxicol Environ Saf 104:403–408
Sonne C, Gustavson K, Rigét FF, Krüger T, Dietz R, Bonefeld-Jørgensen E (2014c) Physiologically based pharmacokinetic modeling of contaminants in Greenlanders: a new tool for monitoring levels and possible health effects? Environ Int 64:91–97
Sonne C, Gustavson K, Dietz R, Letcher RJ (2015) Physiologically-based pharmacokinetic modelling and verification of distribution, elimination and bioaccumulation of persistent organic pollutants in Greenland sledge dogs (Canis familiaris). Environ Res 140:45–55
Sonne C, Gustavson K, Dietz R, Letcher RJ (2016) Risk evaluation of the Arctic environmental POP exposure based on critical body residue and critical daily dose using captive Greenland sledge dogs (Canis familiaris) as surrogate species. Environ Int 88:221–227
Sonne C, Letcher RJ, Jenssen BM, Desforges JP, Eulaers I, Andersen-Ranberg E, Gustavson K, Styrishave B, Dietz R (2017a) A veterinary perspective on one health in the arctic. Acta Vet Scand 59:84
Sonne C, Torjesen PA, Berg KA, Fuglei E, Muir DCG, Jenssen BM, Jørgensen E, Dietz R, Ahlstøm Ø (2017b) Exposure to persistent organic pollutants reduces testosterone concentrations and affects sperm viability and morphology during the mating peak-period in a controlled experiment on farmed Arctic foxes (Vulpes lagopus). Environ Sci Technol 51:4673–4680
Sonne C, Sonne C, Langebæk R, Dietz R, Andersen-Ranberg E, Houser G, Hansen AJ, Sinding MHS, Olsen MT, Egevang C, Gilbert MTP, Meldgaard M (2018) Greenland sled dogs at risk of extinction. Science 360:1080
Statistics Greenland (2017) Greenland in Figures 2017, 40 pp. Available at: http://www.stat.gl/dialog/main.asp?lang=en&version=2017&sc=GF&colcode=p
Tabel H, Corner AH, Webster WA, Casey CA (1974) History and epizootiology of rabies in Canada. Can Vet J 15(10):271
Tartu S, Bourgeon S, Aars J, Andersen M, Polder A, Thiemann GW, Welker JM, Routti H (2017) Sea ice-associated decline in body condition leads to increased concentrations of lipophilic pollutants in polar bears (Ursus maritimus) from Svalbard, Norway. Sci Total Environ 576:409–419
Taylor KC, Jackson LW, Lynch CD, Kostyniak PJ, Buck Louis GM (2007) Preconception maternal polychlorinated biphenyl concentrations and the secondary sex ratio. Environ Res 103:99–105
Thalmann O, Shapiro B, Cui P, Schuenemann VJ, Sawyer SK, Greenfield DL, Germonpré MB, Sablin MV, López-Giráldez F, Domingo-Roura X, Napierala H, Uerpmann HP, Loponte DM, Acosta AA, Giemsch L, Schmitz RW, Worthington B, Buikstra JE, Druzhkova A, Graphodatsky AS, Ovodov ND, Wahlberg N, Freedman AH, Schweizer RM, Koepfli KP, Leonard JA, Meyer M, Krause J, Pääbo S, Green RE, Wayne RK (2013) Complete mitochondrial genomes of ancient canids suggest a European origin of domestic dogs. Science 342:871–874
Tiido T, Rignell-Hydbom A, Jönsson BAG, Giwercman YL, Pedersen HS, Wojtyniak B, Ludwicki JK, Lesovoy V, Zvyezday V, Spano M, Manicardi GC, Bizzaro D, Bonefeld-Jørgensen EC, Toft G, Bonde JP, Rylander L, Hagmar L, Giwercman A, INUENDO (2006) Impact of PCB and p,p´-DDE contaminants on human sperm Y:X chromosome ratio: studies in three european populations and the Inuit population in Greenland. Environ Health Perspect 114:718–724
Todgham AE, Hofmann GE (2009) Transcriptomic response of sea urchin larvae Strongylocentrotus purpuratus to CO2-driven seawater acidification. J Exp Biol 212:2579–2594
Vernersen ES and Jensen J (2018) Hundesyge i Historisk Perspektiv. Bachelor’s thesis. Copenhagen University
Wang Z, Gerstein M, Snyder M (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10:57–63
Wang GD, Zhai W, Yang HC, Wang L, Zhong L, Liu YH, Fan RX, Yin TT, Zhu CL, Poyarkov AD, Irwin DM (2016) Out of southern East Asia: the natural history of domestic dogs across the world. Cell Res 26:21–33
Wasaki J, Yonetani R, Kuroda S, Shinano T, Yazaki J, Fujii F, Shimbo K, Yamamoto K, Sakata K, Sasaki T, Kishimoto N (2003) Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots. Plant Cell Environ 26:1515–1523
Whiteman JP, Harlow HJ, Durner GM, Regehr EV, Amstrup SC, Ben-David M (2018) Phenotypic plasticity and climate change: can polar bears respond to longer Arctic summers with an adaptive fast? Oecologia 186:369–381
Acknowledgments
Nordic Council of Ministers (NMR NORDEN) is acknowledged for financial support to the project entitled Infectious Zoonotic Diseases Transmissible from Harvested Wildlife to Humans in the European Arctic (ZORRO). We also acknowledge BONUS BALTHEALTH that has received funding from BONUS (Art. 185), funded jointly by the EU, Innovation Fund Denmark (Grants 6180-00001B and 6180-00002B), Forschungszentrum Jülich GmbH, German Federal Ministry of Education and Research (Grant FKZ 03F0767A), Academy of Finland (Grant 311966), and Swedish Foundation for Strategic Environmental Research (MISTRA).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Sonne, C. et al. (2020). Sled Dogs as Sentinel Species for Monitoring Arctic Ecosystem Health. In: Pastorinho, M., Sousa, A. (eds) Pets as Sentinels, Forecasters and Promoters of Human Health. Springer, Cham. https://doi.org/10.1007/978-3-030-30734-9_2
Download citation
DOI: https://doi.org/10.1007/978-3-030-30734-9_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-30733-2
Online ISBN: 978-3-030-30734-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)