
Abstract
Research in the late 1900s has established that birds are theropod dinosaurs, with the discovery of feather preservation in non-avian theropods being the last decisive evidence for the dinosaur origin of this group. Partially due to the great interest in the origin of birds, more phylogenetic analyses of non-avian theropod dinosaurs have probably been published than any other group of fossil vertebrates. Despite a lot of uncertainty in the exact placement of many taxa and even some major clades, there is a remarkable consensus about the hierarchical position of birds (here used for the total group, Avialae) within theropod dinosaurs. Thus, birds are part of Paraves, together with such well-known theropod groups as dromaeosaurids and troodontids; Paraves are part of Maniraptora, which furthermore include Oviraptorosauria, Therizinosauria, and Alvarezsauroidea; Maniraptora belong to Maniraptoriformes, which also include Ornithomimosauria; Maniraptoriformes are a subclade of Coelurosauria, to which Tyrannosauroidea and some other basal taxa also belong; Coelurosauria are part of Tetanurae, together with Allosauroidea and Megalosauroidea; finally, Tetanurae are a subclade of Theropoda, which also include Ceratosauria and Coelophysoidea.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
3.1 Introduction
The transition of one major animal “bauplan” into another and the origin of evolutionary novelty has captured the interest and imagination of scientists and the general public alike, ever since the advent of evolutionary thought in the nineteenth century and its acceptance following the publication of Darwin’s epochal book The Origin of Species (1859). Birds are arguably the most extremely divergent example of a tetrapod bauplan, as they seem fundamentally different from their living reptilian relatives, crocodiles, turtles and lepidosaurs, in almost any respect, from their anatomy, via neurology and physiology to their behaviour. Although recent research has shown that some of these differences are less marked than originally thought (e.g. important aspects of the avian-type flow-through lung are already present in lepidosaurs and crocodiles; Farmer and Sanders 2010; Schachner et al. 2013; see also Cieri and Farmer 2016), many of these evolutionary novelties of birds require complex and changing functionary scenarios to explain their selective advantages, especially if many must be seen as exaptations to flight, rather than as consequences of this drastic change in locomotor behaviour. Thus, it is not surprising that the question of the origin of birds and the evolutionary history of their novelties has been a “hot topic” in evolutionary biology and palaeontology in the past 150 years. Understanding these evolutionary events requires a good idea of the interrelationships of bird ancestors, the origin of birds, and the phylogenetic relationships between early members of this clade.
The discovery of the first Mesozoic “bird”, Archaeopteryx lithographica, only 2 years after the publication of Darwin’s book (von Meyer 1861a, b) marks a milestone in our quest of understanding bird origins (although, interestingly, both initial descriptions of this animal came from anti-Darwinists, who came to diametrically opposite conclusions: Andreas Wagner (1862) came to the conclusion that this animal clearly represents a somewhat odd lizard, whereas Richard Owen (1863) concluded that the fossil undoubtedly represented a bird). Especially the preservation of feathers in the limestone slab that contained the skeleton of this taxon was taken as a clear indication that this animal was a transitional fossil of importance for the question of the origin of birds, and soon after the discovery of Archaeopteryx (and the discovery of the small theropod dinosaur Compsognathus in the same year; Wagner 1861), Darwin’s “bulldog” Thomas Henry Huxley published the hypothesis that birds were derived from small theropod dinosaurs (Huxley 1868). However, although this hypothesis fell on fertile ground in the beginning, other possibilities for the ancestry of birds were proposed subsequently, such as the Early Triassic basal archosauromorph Euparkeria (Broom 1913).
In his very influential book The Origin of Birds, the Danish artist Gerhard Heilmann (1926) summarized the current knowledge on this topic. Although he clearly noticed the many similarities between dinosaurs and birds (especially in Archaeopteryx), Heilmann came to the conclusion that birds cannot be derived from dinosaurs, as all dinosaurs then known lacked clavicles, whereas the furcula in birds is generally considered to be derived from a fusion of these bones, which are present in reptiles ancestrally. Heilmann’s very detailed and well-illustrated book had a lasting impact on the field and formed the basis for the common consensus for 50 years that birds were derived from some still unknown, probably arboreal, Triassic “Proavis”.
In the wake of the “dinosaur renaissance” in the 1960s and 1970s, especially the discovery of the dromaeosaurid Deinonychus in North America (Ostrom 1969a, b), and a comparison of this taxon with newly discovered (Ostrom 1970, 1972; Wellnhofer 1974) and already known specimens of Archaeopteryx led Ostrom (1973, 1976) to revive the hypothesis of the dinosaur origin of birds. Not surprisingly, the hypothesis was met with scepticism (e.g. Martin et al. 1980; Tarsitano and Hecht 1980; Martin 1983), and a sometimes heated debate ensued over the following two and a half decades (see Witmer 2002; Prum 2002 for a summary).
One important aspect that led to the acceptance of the hypothesis of the theropod origin of birds was the advent of new phylogenetic methods, following the publication of Hennig’s book Phylogenetic Systematics in 1966. The cladistic methodology outlined in this work first found acceptance in vertebrate palaeontology in the 1980s, and in a very influential paper published in 1986, Jacques Gauthier listed a total of 84 nested synapomorphies that supported the inclusion of birds in the theropod dinosaurs. Gauthier’s paper was the first of a long list of phylogenetic analyses that support the inclusion of birds in the Theropoda, and our knowledge of this transition and the successive acquisition of avian characters has considerably increased since (see Chiappe 2009; Brusatte et al. 2015; Cau 2018; Agnolín et al. 2019).
The final push for the theropod hypothesis, however, came from the discovery of abundant feathered dinosaurs in the Cretaceous of China, starting in the late 1990s (Chen et al. 1998; Ji et al. 1998; Xu et al. 1999a, b, 2001), and the subsequent realization that filamentous integumentary structures are widely distributed not only in theropod dinosaurs (Rauhut et al. 2012) but are even found in ornithischians (Zheng et al. 2009; Godefroit et al. 2014, 2020). In some instances, the interpretation of integumentary structures as feathers has been questioned, and the most detailed conflicting analyses interpreted these structures as degraded dermal collagen fibres (e.g. Lingham-Soliar 2003a, b, 2012; Lingham-Soliar et al. 2007) or other tissues (e.g. Lingham-Soliar 2010). However, these studies have been criticised on taphonomic, structural, and methodological grounds (e.g. Mayr 2010; Smith et al. 2015; Smithwick et al. 2017), and thus cannot be sustained. Furthermore, the vast array of taxa in which feathers have now been reported plus the great variety of feather types identified (e.g. Xu and Guo 2009) make these alternative interpretations untenable.
Although the opponents of the theropod origin of birds have questioned the cladistic methodology altogether (e.g. Feduccia 1996, 2013), there is no other hypothesis for avian origins that has been formulated in any comparable detail (see Xu et al. 2014; Brusatte et al. 2015; Cau 2018), and the criticism seems to be rather ideological than scientific (Prum 2003; Smith et al. 2015). Thus, in the absence of contrary evidence, the theropod origin of birds can now be regarded as being firmly established, and it is on this background that we will discuss the current consensus and controversies surrounding the origin of birds. For recent reviews of the overwhelming evidence that birds are theropods see, e.g., Xu et al. (2014), Brusatte et al. (2015), Smith et al. (2015), Mayr (2017), Cau (2018), and Agnolín et al. (2019).
3.2 Current Consensus on the Phylogeny of Theropod Dinosaurs and the Origin of Birds
Since the pioneering work of Gauthier (1986), numerous phylogenetic analyses of the interrelationships of theropod dinosaurs have been published (e.g. Novas 1992; Holtz 1994, 1998; Sereno 1997, 1999; Forster 1999; Rauhut 2003; Smith et al. 2007; Xu et al. 2009; Choiniere et al. 2010; Rauhut et al. 2010; Novas et al. 2015; Wang et al. 2017; Cau 2018), with many more analyses focusing on the different subclades of this clade. Interestingly, several of the main phylogenetic findings of Gauthier (1986) have consistently been confirmed, both in terms of tree topology as well as general taxonomic composition of several major clades, despite widely differing taxon and character sampling. Thus, these aspects of theropod phylogeny can be considered well established and largely uncontroversial.
All phylogenetic analyses including theropod dinosaurs agree that this clade is monophyletic, although there is some controversy as to whether certain basal taxa (e.g., Eoraptor, Herrerasaurus) are members of Theropoda or not. Nevertheless, at least the monophyly of Neotheropoda (the clade including Coelophysis and modern birds [Sereno 1998]; the classical Theropoda before the discovery of a number of basal taxa; see Colbert 1964) has never been questioned. Within Theropoda, a number of mainly Late Triassic and Early Jurassic taxa (sometimes included in a single lineage named Coelophysoidea), but also the clade Ceratosauria, which reached the Cretaceous/Paleogene boundary, are consistently found as basal forms outside a more derived clade which was named Tetanurae by Gauthier (1986). The interrelationships of these basal forms are still debated; although basically all phylogenetic analyses agree in the existence of two monophyletic clades, the Coelophysoidea (Fig. 3.1a) and the Ceratosauria (Fig. 3.1b), the referral of numerous taxa to either one of these clades remains controversial. Furthermore, whereas many early phylogenetic analyses recovered Coelophysoidea and Ceratosauria in a monophyletic clade (for which Gauthier [1986] used the name Ceratosauria; see, e.g., Gauthier 1986; Holtz 1994, 1998; Sereno 1997, 1999; but also Allain et al. 2007 as a more recent example), there is an emerging consensus that Ceratosauria represent the sister-taxon to Tetanurae to the exclusion of Coelophysoidea (e.g. Rauhut, 1998, 2003; Forster 1999; Carrano et al. 2002; Smith et al. 2007; Xu et al. 2009; Novas et al. 2015; Wang et al. 2017; Cau 2018) in a clade that was named Averostra by Paul (2002; see also definition by Ezcurra and Cuny 2007).

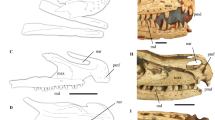
Skulls of representatives of different theropod clades. (a) Coelophysid Coelophysis bauri (NMNH P-42200; photo courtesy Jörg Schneider) in left dorsolateral view. (b) Ceratosaur Ceratosaurus nasicornis (USNM 4735) in right lateral view. (c) Allosauroid Allosaurus sp. (MOR 693; photo courtesy Serjoscha Evers) in left lateral view. (d) Alvarezsauroid Shuvuuia deserti (IGM 100/1001) in right lateral view. (e) Oviraptorosaurid Citipati osmolskae (IGM 100/978) in right lateral view. (f) Dromaeosaurid Tsaagan mangas (IGM 100/1015) in right lateral view. All scale bars are 10 cm
Whereas Coelophysoidea seems to represent the first successful radiation of theropod dinosaurs in the Triassic and includes both taxa from the Late Triassic and Early Jurassic, the earliest averostrans are Early Jurassic in age (see Dal Sasso et al. 2018), and there is growing evidence that an explosive radiation of this clade in the latest Early to Middle Jurassic might have been triggered by the Pliensbachian/Toarcian extinction event (Pol and Rauhut 2012; Rauhut and Carrano 2016; Rauhut et al. 2016).
The Tetanurae are the main clade of theropod dinosaurs that include most of the well-known forms and also recent birds. They first occur in the fossil record in the earliest Middle Jurassic, but the clade obviously experienced an explosive radiation soon after its origin, as all major clades, including avialans, are established by the Late Jurassic (Rauhut et al. 2010, 2016; Xu et al. 2010). Basically all recent phylogenetic analyses agree that Tetanurae split into three major lineages early in their evolutionary history, the Megalosauroidea (Spinosauroidea in older literature), Allosauroidea (Fig. 3.1c) and Coelurosauria, although the exact taxonomic composition of the three clades somewhat differs, especially in respect to inclusion or exclusion of basal taxa (e.g. Holtz 1998; Allain 2002; Rauhut 2003; Rauhut and Xu 2005; Smith et al. 2007; Benson 2010; Benson et al. 2010; Choiniere et al. 2010; Carrano et al. 2012; Novas et al. 2015; Wang et al. 2017; Cau 2018).
Megalosauroids include mainly large-bodied and often heavily built megapredators, such as Megalosaurus and Torvosaurus, that thrived during the Middle and Late Jurassic (Benson 2010; Carrano et al. 2012; Rauhut et al. 2016), but also the highly specialized gigantic spinosaurids of the Cretaceous, which include the largest theropod known, Spinosaurus, which probably reached a length of 18 m and up to 10 t in body mass (Stromer 1915; Therrien and Henderson 2007; Hone and Holtz 2017). Likewise, allosauroids were also generally large-bodied and megapredatory theropods that originated in the Middle Jurassic and thrived to at least the early Late Cretaceous, culminating in the gigantic carcharodontosaurids (e.g. Brusatte and Sereno 2008; Benson et al. 2010; Carrano et al. 2012).
In contrast to these two major lineages of tetanurans, the third major clade, the Coelurosauria, includes both large and small forms and saw repeated changes in trophic ecology (Zanno and Makovicky 2011). Coelurosaurs include such iconic animals as Tyrannosaurus rex or Velociraptor mongoliensis, and more phylogenetic analysis of this clade have probably been published than most other fossil animals (e.g. Makovicky and Sues 1998; Norell et al. 2001, 2006; Xu et al. 2002, 2011, 2015; Makovicky et al. 2003; Senter 2007; Zhang et al. 2008; Choiniere et al. 2010, Choiniere et al. 2014; Turner et al. 2012; Agnolín and Novas 2013; Godefroit et al. 2013a, b; Brusatte et al. 2014; Foth et al. 2014; Cau et al. 2015, 2017). Although there are considerable differences in the placement of many taxa and even whole clades within coelurosaurs (see below), there also exists some consensus about the general topology of the coelurosaur family tree. Thus, basically all analyses of the last 20 years agree that Tyrannosauroidea are one of the most basal clades. Tyrannosauroidea have recently been found to include not only the well-known, gigantic megapredators of the Late Cretaceous, such as Tyrannosaurus, but also several other lineages, reaching back to the Middle Jurassic, such as the rather small-bodied, obviously fleet-footed proceratosaurids (Brusatte et al. 2010; Rauhut et al. 2010; Brusatte and Carr 2016).
Tyrannosauroidea and several other basal taxa and clades are outside a derived clade of coelurosaurs that Holtz (1996) named Maniraptoriformes. The most basal group within this clade are the Ornithomimosauria, generally small to medium-sized theropods with small skulls, long necks and elongate hindlimbs. All derived members of this clade are toothless and were probably omnivorous. Although most members of the Ornithomimosauroidea do not exceed 5–6 m in length and weighed less than 600 kg (Benson et al. 2018), the clade also includes the giant Deinocheirus that reached more than 11 m in length and more than six tons in weight (Lee et al. 2014).
An important clade within coelurosaurs is the Maniraptora. The clade was originally coined by Gauthier (1986) to include birds (Avialae) and theropods that share characters, especially in the manus that are not present in ornithomimosaurs. The clade has been phylogenetically defined by Holtz (1996) as all coelurosaurs that share a more recent ancestor with birds than with ornithomimids. Ever since the analysis of Gauthier (1986), several clades were consistently found to be maniraptorans, including Oviraptorosauria (Figs. 3.1e and 3.2a), Troodonotidae (Fig. 3.2b), Dromaeosauridae (Fig. 3.1f), and Avialae (including modern birds), together with some taxa that do not seem to be included in a larger clade, such as the genus Ornitholestes. A number of clades that have only more recently been recognized (or firmly established as theropodan, in the case of therizinosaurs), including Alvarezsauroidea (Fig. 3.1d), Therizinosauria, Scansoriopterygidae, and Anchiornithidae, are usually also found within Maniraptora, although their detailed relationships differ widely between different analyses (see below).

Non-avian theropod skeletons documenting avian-like behaviour. (a) Postcranial skeleton of the oviraptorosaurid Citipati osmolskae in a brooding position on a nest of eggs (IGM 100/1004). (b) Troodontid Mei long in an avian-like sleeping position (IVPP V12733). Scale bars are 10 cm
Within Maniraptora, the Troodontidae, Dromaeosauridae, and Avialae are united in the clade Paraves, defined as all maniraptorans that are more closely related to extant birds than to Oviraptor (Sereno 1997, 1998). Whereas the recently recognized Anchiornithidae (Xu et al. 2016; Foth & Rauhut 2017) have always also been recovered as Paraves (e.g. Xu and Fucheng 2005; Hu et al. 2009, 2018; Xu et al. 2009, 2011; Godefroit et al. 2013a, b; Foth and Rauhut 2017), the Alvarezsauroidea and Scansoriopterygidae have been found to be Paraves only in some, but not all phylogenetic analyses.
Nested within Paraves is the clade Avialae, which is the most-inclusive clade containing extant birds, but not Dromaeosauridea or Troodontidae (Maryańska et al. 2002). Basel members of this clade are Archaeopteryx and Alcmonavis from the Late Jurassic of Germany (see Rauhut et al. 2019) and the groups Jeholornithidae, Sapeornithidae, and Confuciusornithidae, which are all known from the Early Cretaceous of China (e.g. Mayr 2017; Wang and Zhou 2017). The clade that embraces Confuciusornithidae and extant birds including all their descendants is the Pygostylia (Chiappe 2002). This clade contains two major groups, the Enantiornithes and Ornithuromorpha (Euornithes), which are summarized as Ornithothoraces (Sereno 1998; Chiappe 2002). The Enantiornithes are small-bodied, toothed Avialae, which represent the most successful group of stem birds during the Cretaceous in terms of species richness as well as temporal and geographic range (Mayr 2017; Wang and Zhou 2017). According to the foot morphology they were primarily arboreal (O’Connor et al. 2011a), but as indicated by differences in the snout shape, tooth morphology, and pedal claw geometry, a certain degree of ecological specializations was present. This includes, for instance, the long-snouted Longipterygidae (O’Connor et al. 2011b) or the raptorial Bohaiornithidae (Li et al. 2014).
In contrast to Enantiornithes, Ornithuromorpha possesses an enormous ecological diversification in terms of habitat and diet preferences, while their species diversity is lower when compared to their sister taxon. The ecological diversity includes semi-to-fully aquatic, but also ground-dwelling, and even secondary flightless taxa. In contrast to other Avialae, they also show a higher degree of tooth reduction and adaptation to piscivory, omnivory, insectivory, and granivory (Mayr 2017; Wang and Zhou 2017). The most successful group of Ornithuromorpha are the Aves (Neornithes), which represent the crown-group of extant birds, and are the only theropod branch that survived the K/T extinction event. The Aves already originated in the Late Cretaceous, showing an initial diversification of the clades Palaeognathae, Galloanseres, and Neovaves (Clarke et al. 2005; Brown et al. 2008; Prum et al. 2015; Mayr 2017). However, the actual radiation of crown group birds happened during the early Cenozoic, after the K/T event (Mayr 2009; Prum et al. 2015).
3.3 Remaining Controversies
Although there is a remarkable consensus in the general hierarchy of theropod interrelationships and the hierarchy levels that most clades belong to, there are numerous controversies about the exact phylogenetic position of numerous taxa and some entire clades. In non-tetanuran theropods, these uncertainties mainly concern the taxonomic composition of the basal clade Coelophysoidea and the question whether Coelophysoidea and Ceratosauria are united in a clade or whether Ceratosauria are closer to Tetanurae. Concerning Coelophysoidea, current hypotheses reach from uniting basically all Late Triassic and the vast majority of Early Jurassic neotheropods in this clade (e.g. Carrano et al. 2005; Allain et al. 2007) to the possibility that a number of Late Triassic and Early Jurassic taxa are more closely related to averostrans (e.g. Rauhut 2003; Smith et al. 2007; Ezcurra and Brusatte 2011; Langer et al. 2014; Ezcurra 2017; Martínez and Apaldetti 2017), including the possibility of another clade of mainly Early Jurassic theropods, the Dilophosauridae (e.g. Smith et al. 2007). In respect to the phylogenetic position of the Ceratosauria, there seems to be a growing consensus that this clade is united with the Tetanurae in a monophyletic Averostra, with the last formal analysis finding a Coelophysoidea-Ceratosauria clade being that of Allain et al. (2007), whereas all more recent phylogenies found support for Averostra.
Within basal tetanurans, there is some disagreement on the relationships between the three major lineages, Megalosauroidea, Allosauroidea, and Coelurosauria. Thus, whereas most recent analyses found Allosauroidea and Coelurosauria to be sister taxa in a clade named either Avetheropoda or Neotetanurae (see Carrano et al. 2012), to the exclusion of Megalosauroidea (e.g. Allain 2002; Smith et al. 2007; Benson 2010; Benson et al. 2010; Rauhut et al. 2010, 2016; Carrano et al. 2012; Novas et al. 2015; Wang et al. 2017), some analyses found Allosauroidea and Megalosauroidea to be united in a clade called Carnosauria to the exclusion of Coelurosauria (e.g. Rauhut 2003; Rauhut et al. 2012; Cau 2018).
Apart from uncertainties of the placements of several genera within their respective clades, a further important discrepancy in phylogenetic hypotheses of basal tetanurans concerns the placement of the only recently recognized Megaraptora. The first representatives known of this clade were represented by very fragmentary material (Novas 1998; Azuma and Currie 2000; Calvo et al. 2004), and so their recognition as belonging to a monophyletic clade and an analysis of their phylogenetic relationships only became possible after more complete remains had been found (Sereno et al. 2008; Hocknull et al. 2009). The first work to recognize a monophyletic Megaraptora was Benson et al. (2010), who recovered Megaraptora as part of the Neovenatoridae, the sister taxon to Carcharodontosauridae within the Allosauroidea. This phylogenetic placement was supported by several subsequent analyses (e.g. Carrano et al. 2012; Rauhut et al. 2016), but Novas et al. (2013) argued that Megaraptora were basal coelurosaurs and, more specifically, a mainly Gondwanan radiation of tyrannosauroids. Coelurosaur affinities have been supported by several more recent analyses using new material (Porfiri et al. 2014; Aranciaga Rolando et al. 2019), but the exact position of this interesting clade is still uncertain (e.g. Apesteguía et al. 2016; Coria and Currie 2016; Novas et al. 2016).
Within basal Coelurosauria, an important early clade of uncertain phylogenetic position are the Compsognathidae. This clade might be an early radiation of coelurosaurian theropods that originated in the Late Jurassic at the latest and reached a wide distribution in the Early Cretaceous, although the exact taxonomic composition of the group is also still debated. One problem with the current concept of the Compsognathidae might be that several taxa included in this clade are juveniles (e.g. Juravenator: Chiappe and Göhlich 2010; Scipionyx: Dal Sasso and Maganuco 2011), and some of the characteristics supporting compsognathid monophyly might be ontogenetically variable (see Rauhut et al. 2012). Thus, Compsognathus and its closest relatives are found as the most basal larger clade of coelurosaurs in some analyses (e.g. Rauhut 2003; Holtz et al. 2004; Rauhut et al. 2010; Cau 2018), as sister taxon to Maniraptoriformes (e.g. Senter 2007; Smith et al. 2007; Xu et al. 2009, Xu et al. 2015; Choiniere et al. 2014; Rauhut et al. 2019), or as basal Maniraptora (e.g. Choiniere et al. 2010; Foth et al. 2014).
Another problematic clade within coelurosaurs are the Alvarezsauroidea. Originally thought to be basal birds (e.g. Perle et al. 1994; Novas 1996; Chiappe et al. 1998; Chiappe 2002), most more recent phylogenies have placed these animals as basal maniraptorans (e.g. Clark et al. 2002; Senter 2007; Choiniere et al. 2010, 2014; Foth et al. 2014; Xu et al. 2018), and Sereno (1999) suggested that alvarezsauroids were the sister taxon to Ornithomimosauria. The problem with alvarezsauroids was that most first discoveries of this clade were of highly derived members that have a very aberrant morphology (e.g. Perle et al. 1994; Novas 1996, 1997; Chiappe et al. 1998), making their placement within theropods problematic. However, with the recent discovery of more basal forms (Choiniere et al. 2010, 2014; Xu et al. 2018), our understanding of alvarezsauroid anatomy, phylogeny, and evolution is rapidly improving, and a consensus of this clade being basal maniraptorans seems to be emerging, although the exact phylogenetic position at the base of Maniraptora remains unstable.
A similar problem has affected the Therizinosauria. As with alvarezsauroids, the first discoveries of therizinosaurs were of highly derived forms (e.g. Maleev 1954; Perle 1979, 1982; Barsbold and Perle 1980), and even the placement in one of the principal clades of dinosaurs of these animals was at first unclear (see Paul 1984). Only with the discovery of more basal forms did the theropod affinities of therizinosaurs become firmly established (Russell and Dong 1993). Since then, this clade has repeatedly been found as the sister taxon of the Oviraptorosauria within the Maniraptora (e.g. Makovicky & Sues 1998; Holtz 1998; Clark et al. 2002; Rauhut 2003; Holtz et al. 2004; Xu et al. 2007; Choiniere et al. 2014; Cau 2018), although most recent analyses have favoured a more basal position of therizinosaurs, outside the Pennaraptora (Oviraptorosauria + Paraves; e.g. Senter 2007; Zanno 2010; Xu et al. 2011, 2017; Turner et al. 2012; Agnolín and Novas 2013; Brusatte et al. 2014; Foth et al. 2014; Foth and Rauhut 2017; Hu et al. 2018). This problem remains currently unresolved, as highlighted by the analyses presented by Rauhut et al. (2019): whereas an unweighted analysis found a Therizinosauria-Oviraptorosauria clade, an implicit weight analysis of the same data matrix found the therizinosaurs outside the Pennaraptora.
Another only recently recognized clade of interesting, bird-like theropods are the Scansoriopterygidae. These animals are so far only known from the early Late Jurassic Yanliao Biota of north-eastern China, from where at least four different taxa have been described (Zhang et al. 2002, 2008; Xu et al. 2015; Wang et al. 2019). Scansoriopterygids are small, bird-like theropods that included volant forms with membranous wings (Xu et al. 2015; Wang et al. 2019). The clade was originally regarded as a radiation of basal avialans (Zhang et al. 2008), and this has been supported by some subsequent analyses (e.g. Xu et al. 2011; Foth et al. 2014), whereas a number of more recent analyses regarded scansoriopterygids as basal paravians, outside a Avialae-Deinonychosauria split (Xu et al. 2015; Wang et al. 2019). On the other hand, Agnolín & Novas (2013) recovered scansoriopterygids as basal oviraptorosaurs, which was supported by Brusatte et al. (2014) and Rauhut et al. (2019). Thus, more finds and more detailed studies of the known taxa are necessary to resolve the relationships of these interesting animals.
Another area of conflict concerns basal paravian phylogeny. In most analyses of coelurosaur interrelationships, Troodontidae and Dromaeosauridae are recovered as sister groups, forming the monophyletic Deinonychosauria (e.g. Sereno 1997, 1999; Holtz 1998; Clark et al. 2002; Rauhut 2003; Senter 2007; Turner et al. 2012; Rauhut et al. 2019). In contrast, several recent analyses found Troodontidae and Avialae as sister taxa to the exclusion of Dromaeosauridae (e.g., Godefroit et al. 2013b; Choiniere et al. 2014; Foth et al. 2014; Cau 2018). This uncertainty reflects the great similarity of many of these bird-like dinosaurs and is mirrored by the uncertain phylogenetic position of some other basal paravians, such as the anchiornithids, which are considered to be troodontids in some analyses (e.g. Hu et al. 2009; Turner et al. 2012; Godefroit et al. 2013b; Brusatte et al. 2014), basal deinonychosaurs (e.g. Xu et al. 2011, 2015; Wang et al. 2019), or avialans more basal than Archaeopteryx (e.g. Agnolín and Novas 2013; Godefroit et al. 2013a; Foth et al. 2014; Rauhut et al. 2019), apart from other occasional placements within Paraves (e.g. as basal taxon outside the Deinonychosauria-Avialae split; Lefèvre et al. 2017).
These different phylogenetic hypotheses also affected the phylogenetic position of Archaeopteryx, which until today represents a yardstick for early bird evolution. Traditionally, Archaeopteryx is a basal member of the Avialae (e.g. Sereno 1999; Rauhut 2003, Senter 2007; Turner et al. 2012; Brusatte et al. 2014; see above), while some recent studies placed Archaeopteryx together with Anchiornis outside Avialae as sister taxon to Deinonychosauria (e.g. Xu et al. 2011, 2015; Xu and Pol 2014; Godefroit et al. 2013a; Hu et al. 2018; Wang et al. 2019). On the other hand, due to a high level of homoplasy in early Paraves, single studies classified Rahonavis, Balaur, or Microraptor to be basal Avialae (e.g. Agnolín and Novas 2011, 2013; Cau et al. 2015, 2017; Foth and Rauhut 2017; Lefèvre et al. 2017), while they are traditionally placed within Dromaeosauridae (see Turner et al. 2012; Brusatte et al. 2013). Further controversies remain regarding the exact relationships between Jeholornithidae, Sapeornithidae, Confuciusornithidae, and more derived Ornithothoraces. Many studies found the long-tailed Jeholornithidae to be the sister taxon of a monophyletic clade Pygostylia containing the short-tailed Sapeornithidae and Confuciusornithidae and more derived Ornithothoraces (e.g. Zhou et al. 2008; O’Connor et al. 2009, 2013; Zhang et al. 2014; Wang et al. 2015). This relationship represents the most parsimonious explanation for the tail evolution in the stem line of birds. However, other analyses found Sapeornithidae to be more basal than Jeholornithidae (e.g. Zhou et al. 2010; Turner et al. 2012; Cau et al. 2017; Foth and Rauhut 2017; Agnolín et al. 2019), which is more parsimonious, explaining the evolution of the pectoral girdle and sternum. In contrast to basal Avialae, the phylogenetic relationship of the main clades within Ornithothoraces are well supported by various phylogenetic analyses (e.g., Clarke et al. 2006; Zhou et al. 2008; Wang et al. 2015; O’Connor et al. 2016).
3.4 The Occurrence of Feathers in the Fossil Record of Theropod Dinosaurs
As feathers are not skeletal tissues, they might only be preserved under exceptional circumstances, such as in Konservat-Lagerstätten. It is therefore not surprising that the fossil record of feathers in general is rather poor, and this is especially also the case for Mesozoic theropods.
The first record of a feather from the Mesozoic was the original isolated feather of Archaeopteryx, which von Meyer (1861a) first announced. More importantly, a skeletal specimen including feather impressions was found in the same year (von Meyer 1861b), and it was primarily the feather impressions that led to the identification of this animal as a bird (Owen 1863). The importance of the feather impressions in this iconic fossil was such that their authenticity was questioned as recently as 1985 (Hoyle and Wickramasinghe 1985), although there is no reasonable doubt that these structures are real (see Rietschel 1985; Charig et al. 1986; Wellnhofer 2008).
For many decades, the feathers of Archaeopteryx were the only known fossil feathers from the Mesozoic. More importantly, although feathers were later occasionally found as carbonized traces in exceptional lagerstätten or preserved in amber (see Davis and Briggs 1995; Kellner 2002; Prado et al. 2016), these were isolated finds of feathers, which did not allow a taxonomic identification of the animal that they belonged to, and thus provided only limited data on the evolution of these structures. This changed drastically with the discovery of abundant feathered dinosaurs and early birds in the Lower Cretaceous Jehol Biota of China in the 1990s (e.g. Hou et al. 1995; Chen et al. 1998; Ji et al. 1998; Xu et al. 1999a, b). Since then, feathers have been reported from these deposits and the older (late Middle to early Late Jurassic) Yanliao biota for all major clades of maniraptoran theropods (see Xu 2020), and for some more basal coelurosaurian taxa, such as the compsognathid Sinosauropteryx (Chen et al. 1998) and the tyrannosauroid Yutyrannus (Xu et al. 2012).
Reports of feathers in non-coelurosaurian theropods, in contrast, are still exceedingly rare, mainly because no such taxa have been found in these exceptional lagerstätten. Most reports of the possible presence of feathers in non-coelurosaurian theropods are therefore debatable. Gierlinski (1997) reported feather-like impressions in a theropod resting trace from the Early Jurassic of North America, and this interpretation was more recently supported by Kundrát (2004). As there are no coelurosaurian theropods known from the Early Jurassic, these traces might not only represent the oldest evidence of feathers, but also indirect evidence for non-coelurosaurian feathers. Although Kundrát (2004) made a good case for these imprints to represent feathers, some uncertainty remains, and another problem is, of course, the difficulty in identifying the trackmaker.
Another indirect evidence for feathers in a non-coelurosaurian theropod was presented by Ortega et al. (2010), who reported bumps on the ulna of the carcharodontosaurid Concavenator, which they interpreted as feather quill knobs. However, as argued by Foth et al. (2014), these knobs are in a different position than the quill knobs found in some modern volant birds and are irregularly spaced, casting doubt on this interpretation. Although Cuesta et al. (2018), in a study of probable forelimb myology of Concavenator, did not find any evidence for these knobs representing attachments of interosseous ligaments, as suggested by Foth et al. (2014), their significance remains controversial.
Probably the best evidence of feathers in a non-coelurosaurian theropod is provided by the exceptionally preserved holotype specimen of Sciurumimus albersdoerferi from the Kimmeridgian Torleite Formation in southern Germany (Rauhut et al. 2012). This specimen does have abundant filament impressions above the base of the tail and shows numerous phosphatized filaments in different parts of the body under UV light (Rauhut et al. 2012; see also Foth et al. 2020). In the case of Sciurumimus, the question is thus not so much the presence of protofeathers, but there is some uncertainty about its phylogenetic position. Rauhut et al. (2012) recovered this taxon as a basal tetanuran, and probably a megalosauroid, based on an analysis of this taxon in three different phylogenetic matrices. Thus, in this hypothesis, the presence of protofeathers in this taxon extends the record of these structures to at least the base of Tetanurae. However, Godefroit et al. (2013a) recovered Sciurumimus as a basal coelurosaur, in which case the origin of protofeathers might well lie within this clade. However, as argued by Rauhut et al. (2012), we strongly suspect that possible coelurosaurian characters in Sciurumimus are due to the very early ontogenetic stage of the only known specimen, as heterochrony seems to have played an important role in the evolution of coelurosaurian theropods (e.g. Bhullar et al. 2012; Foth et al. 2016), and thus consider a basal tetanuran placement of this taxon to be more likely.
3.5 Conclusions and Outlook
Although there is still much debate about many details of the phylogenetic relationships of theropod dinosaurs and thus the origin of birds, there is a remarkable consensus on the backbone structure of the family tree of the ancestors of birds and the relative hierarchical placement of almost all major clades that constitute this tree (Fig. 3.3). Thus, disregarding the more problematic (and often smaller) groups, all recent analyses agree that avialans (“birds”) are members of the Paraves, together with dromaeosaurids and troodontids; Paraves are a subclade of Maniraptora, together with oviraptorosaurs, therizinosaurs, and alvarezsauroids; Maniraptora is part of Maniraptoriformes, together with ornithomimosaurs; Maniraptoriformes is a subclade of Coelurosauria, together with Tyrannosauroidea; Coelurosauria is part of Tetanurae, together with Megalosauroidea and Allosauroidea; Tetanurae is part of Neotheropoda, together with Ceratosauria and Coelophysoidea (and probably part of Averostra together with Ceratosauria). Thus, this phylogenetic hierarchy forms a solid base for improving our understanding of the evolution of the unique avian body plan (Fig. 3.4), as originally lined out by Gauthier (1986) and elaborated in more detail recently by Brusatte et al. (2014), Xu et al. (2014), and Cau (2018). With the discovery of abundant feathered dinosaurs, mainly from the Late Jurassic and Early Cretaceous of China, but also from other areas, the phylogenetic hierarchy outlined above helps us to extend such evolutionary scenarios to the evolution of feathers (Xu 2006, 2020; Xu and Guo 2009) and novel insights into the possible functional context in which these structures evolved.

Simplified cladogram of theropod relationships, showing common relationships between theropod dinosaurs and alternative phylogenetic positions for several problematic taxa (modified from Rauhut 2003)

Simplified time-calibrated theropod phylogeny showing the major events of character evolution along the theropod-bird transition. (1) Theropoda: bipedal locomotion∗, initial vertebral pneumatization and ventilatory air sacs∗, increased metabolic rate∗, thin bone walls, four-fingered hand (∗plesiomorphic characters); (2) Neotheropoda: widely arched furcula; (3) Averostra: extended vertebral pneumatization; (4) Tetanurae/Orionides: three-fingered hand with initial lateral folding mechanism; (5) Avetheropoda; (6) Coelurosauria; (7) Maniraptoriformes; (8) Maniraptora: semilunate carpal with partial lateral folding mechanism; (9) Pennaraptora: cerebral expansion, costosternal ventilator pump, V-shaped furcula, initial forelimb-flapping capabilities, increased manual lateral folding mechanism, two-layered eggshells, brooding behaviour; (10) Paraves/Eumaniraptora: extreme miniaturization, elaborated visual cortex, forelimb elongation and thickening, asymmetric egg shape, egg shells with low porosity and without ornamentation, potential third (external) layer in eggshell; (11) Avialae: aerial locomotion, asymmetric pennaceous feathers, lateral facing glenoid, forelimb elongation and thickening with increased flapping capabilities, shortened bony tail; (12) Pygostylia: crop, dorsolateral facing glenoid, strut-like coracoid, U-shaped furcula, fused sternum, rod-like pygostyle, posterior pubis orientation, one active ovary and oviduct; (13) Ornithothoraces: alula wing feather, sternal keel, synsacrum with 8 or more vertebrae; (14) Euornithes/Ornithuromorpha: kinetic skull, full forelimb-flapping capabilities and manual lateral folding mechanism, fused carpometacarpus, fusion of pelvic bones, metatarsal fully fused, increased egg size, ploughshare-shaped pygostyle. All silhouettes taken from (www.phylopic.org)
New discoveries and more detailed studies of key taxa will certainly help to further improve our understanding of theropod phylogeny. However, there seems to be a trend to increase “birdiness” in several lineages independently, including possible multiple experiments with flight in derived coelurosaurian theropods (e.g. Xu et al. 2003; Foth et al. 2014; Wang et al. 2019). Together with the incomplete preservation of many remains, this marked parallelism—which is also seen in other parts of the theropod family tree (e.g. Rauhut and Pol 2019)—will make detailed reconstructions of the phylogenetic relationships at the origin of birds, in which the relationships of all relevant taxa can be established, difficult, if not impossible. However, such a detailed understanding might not be necessary to improve our understanding of the origin of birds, as the general agreement on the placement of most taxa in the hierarchy leading towards birds provides ample anatomical and functional data for hypothesis formulation and testing.
Likewise, new discoveries of feathered dinosaurs, not only in the now famous Yanliao and Jehol Biota, but also other lagerstätten, such as the limestones of the Solnhofen Archipelago (Chiappe and Göhlich 2010; Rauhut et al. 2012) or in Mesozoic amber (e.g. Xing et al. 2016a, b, 2019), will certainly improve our understanding of feather diversity, evolution, and function. Furthermore, the use of novel techniques, such as laser-stimulated fluorescence (e.g. Kaye et al. 2015, 2019), new microscopic or chemical techniques (Schweitzer et al. 1999, 2008), investigations of the role of melanosomes for both feather colouring and structure (see Smithwick and Vinther 2020, and references therein), and further improvements of methods such as UV photography have great potential to provide new insights into the preservation and structure of feathers in fossil taxa.
References
Agnolín FL, Novas FE (2013) Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. Springer, Dordrecht
Agnolín FL, Motta MJ, Egli FB, Lo Coco G, Novas FE (2019) Paravian phylogeny and the dinosaur-bird transition: an overview. Front Earth Sci 6:252
Agnolín FL, Novas FE (2011) Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)? Anais da Academia Brasileira de Ciências 83:117–162
Allain R (2002) Discovery of megalosaur (Dinosauria, Theropoda) in the middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae. J Vertebr Paleontol 22:548–563
Allain R, Tykoski RS, Aquesbi N, Jalil N-E, Manbaron M, Russell DA, Taquet P (2007) An Abelisauroid (Dinosauria: Theropoda) from the early Jurassic of the High Atlas Mountains, Morocco, and the radiation of Ceratosaurs. J Vertebr Paleontol 27:610–624
Apesteguía S, Smith ND, Juárez Valieri R, Makovicky PJ (2016) An unusual new theropod with a didactyl manus from the Upper Cretaceous of Patagonia, Argentina. PLoS One 11:e0157793
Aranciaga Rolando AM, Novas FE, Agnolín FL (2019) A reanalysis of Murusraptor barrosaensis Coria & Currie (2016) affords new evidence about the phylogenetical relationships of Megaraptora. Cretac Res 99:104–127
Azuma Y, Currie PJ (2000) A new carnosaur (Dinosauria: Theropoda) from the Lower Cretaceous of Japan. Can J Earth Sci 37:1735–1753
Barsbold R, Perle A (1980) Segnosauria, a new infraoder of carnivorous dinosaurs. Acta Palaeontologica Polonica Pol 25:187–195
Benson RBJ (2010) A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods. Zool J Linnean Soc 158:882–935
Benson RBJ, Hunt G, Carrano MT, Campione N, Mannion P (2018) Cope’s rule and the adaptive landscape of dinosaur body size evolution. Palaeontology 61:13–48
Benson RBJ, Carrano MT, Brusatte SL (2010) A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic. Naturwissenschaften 97:71–78
Bhullar B-AS, Marugán-Lobón J, Racimo F, Bever GS, Rowe TB, Norell MA, Abzhanov A (2012) Birds have paedomorphic dinosaur skulls. Nature 487:223–226
Broom R (1913) On the South-African pseudosuchian Euparkeria and allied genera. Proc Zool Soc London 83:619–633
Brown JW, Rest JS, Garcia-Moreno J, Sorenson MD, Mindell DP (2008) Strong mitochondrial DNA support for a Cretaceous origin of modern avian lineages. BMC Biol 6:1–18
Brusatte SL, Carr TD (2016) The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Sci Rep 6:20252
Brusatte SL, Sereno PC (2008) Phylogeny of Allosauroidea (Dinosauria: Theropoda): comparative analysis and resolution. J Syst Palaeontol 6:155–182
Brusatte SL, Norell MA, Carr TD, Erickson GM, Hutchinson JR, Balanoff AM, Bever GS, Choiniere JN, Makovicky PJ, Xu X (2010) Tyrannosaur paleobiology: new research on ancient exemplar organisms. Science 329:1481–1485
Brusatte SL, Vremir M, Csiki-Sava Z, Turner AH, Watanabe A, Erickson GM, Norell MA (2013) The osteology of Balaur bondoc, an island-dwelling dromaeosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Romania. Bull Am Museum Nat Hist 374:1–100
Brusatte SL, Lloyd GT, Wang SC, Norell MA (2014) Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition. Curr Biol 24:2386–2392
Brusatte SL, O’Connor JK, Jarvis ED (2015) The origin and diversification of birds. Curr Biol 25:R888–R898
Calvo JO, Porfiri JD, Veralli C, Novas FE, Poblete F (2004) Phylogenetic status of Megaraptor namunhuaiquii Novas based on a new specimen from Neuquén, Patagonia, Argentina. Ameghiniana 41:565–575
Carrano MT, Sampson SD, Forster CA (2002) The osteology of Masiakasaurus knopfleri, a small Abelisauroid (Dinosauria, Theropoda) from the Late Cretaceous of Madagascar. J Vertebr Paleontol 22:510–534
Carrano MT, Hutchinson JR, Sampson SD (2005) New information on Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona. J Vertebr Paleontol 25:835–849
Carrano MT, Benson RBJ, Sampson SD (2012) The phylogeny of Tetanurae (Dinosauria: Theropoda). J Syst Palaeontol 10:211–300
Cau A (2018) The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana 57:1–25
Cau A, Brougham T, Naish D (2015) The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird? PeerJ 3:e1032
Cau A, Beyrand V, Voeten FAE, Fernandez V, Taffereau P, Stein K, Barsbold R, Tsogtbaatar K, Currie PJ, Godefroit P (2017) Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs. Nature 552:395–399
Charig AJ, Greenaway F, Milner AC, Walker CA, Whybrow PJ (1986) Archaeopteryx is not a forgery. Science 232:622–626
Chen P, Dong Z, Zhen S (1998) An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391:147–152
Chiappe LM (2002) Basal bird phylogeny: problems and solutions. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: Above the heads of dinosaurs. University of California Press, Berkeley, pp 448–472
Chiappe LM (2009) Downsized dinosaurs: the evolutionary transition to modern birds. Evol Educ Outreach 2:248–256
Chiappe LM, Göhlich UB (2010) Anatomy of Juravenator starki (Theropoda: Coelurosauria) from the Late Jurassic of Germany. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 258:257–296
Chiappe LM, Norell MA, Clark JM (1998) The skull of a relative of the stem-group bird Mononykus. Nature 392:275–278
Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y, Han F (2010) A basal alvarezsauroid theropod from the Early Jurassic of Xinjiang, China. Science 327:571–574
Choiniere JN, Clark JM, Forster CA, Norell MA, Eberth DA, Erickson GM, Chu H, Xu X (2014) A juvenile specimen of a new coelurosaur (Dinosauria: Theropoda) from the Middle–Late Jurassic Shishugou Formation of Xinjiang, People’s Republic of China. J Syst Palaeontol 12:177–215
Cieri RL, Farmer CG (2016) Unidirectional pulmonary airflow in vertebrates: a review of structure, function, and evolution. J Comp Psychol B 186:541–552
Clark JM, Norell MA, Makovicky PJ (2002) Cladistic approaches to the relationships of birds to other theropod dinosaurs. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley, pp 31–61
Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA (2005) Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 433:305–308
Clarke JA, Zhou Z, Zhang F (2006) Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui. J Anat 208:287–308
Colbert EH (1964) Relationships of the saurischian dinosaurs. Am Museum Novitates 2181:1–24
Coria RA, Currie PJ (2016) A new megaraptoran dinosaur (Dinosauria, Theropoda, Megaraptoridae) from the Late Cretaceous of Patagonia. PLoS One 11:e015797
Cuesta E, Ortega F, Sanz JL (2018) Appendicular osteology of Concavenator corcovatus (Theropoda; Carcharodontosauridae; Early Cretaceous; Spain). J Vertebr Paleontol 38:e1485153
Dal Sasso C, Maganuco S (2011) Scipionyx samniticus (Theropoda: Compsognathidae) from the Lower Cretaceous of Italy. Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 37:1–281
Dal Sasso C, Maganuco S, Cau A (2018) The oldest ceratosaurian (Dinosauria: Theropoda), from the Lower Jurassic of Italy, sheds light on the evolution of the three-fingered hand of birds. PeerJ 6:e5976
Darwin CR (1859) On the origin of species by means of natural selection. John Murray, London
Davis PG, Briggs DEG (1995) Fossilization of feathers. Geology 23:783–786
Ezcurra MD (2017) A new early coelophysoid neotheropod from the Late Triassic of northwestern Argentina. Ameghiniana 54:506–538
Ezcurra MD, Brusatte SL (2011) Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur C amposaurus arizonensis from the Late Triassic of North America. Palaeontology 54:763–772
Ezcurra MD, Cuny G (2007) The coelophysoid Lophostropheus airelensis, gen. nov.: a review of the systematics of ‘Liliensternus’ airelensis from the Triassic-Jurassic outcrops of Normandy (France). J Vertebr Paleontol 27:73–86
Farmer CG, Sanders K (2010) Unidirectional airflow in the lungs of alligators. Science 327:338–340
Feduccia A (1996) The origin and evolution of birds. Yale University Press, New Haven
Feduccia A (2013) Bird origins anew. Auk 130:1–12
Forster CA (1999) Gondwanan dinosaur evolution and biogeographic analysis. J Afr Earth Sci 28:169–185
Foth C, Rauhut OWM (2017) Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs. BMC Evol Biol 17:236
Foth C, Tischlinger H, Rauhut OWM (2014) New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 511:79–82
Foth C, Hedrick BP, Ezcurra MD (2016) Cranial ontogenetic variation in early saurischians and the role of heterochrony in the diversification of predatory dinosaurs. PeerJ 4:e1589
Foth C, Haug C, Haug JT, Tischlinger H, Rauhut OWM (2020) Two of a feather: a comparison of the preserved integument in the juvenile theropod dinosaurs Sciurumimus and Juravenator from the Kimmeridgian Torleite Formation of southern Germany. In: Foth C, Rauhut OWM (eds) The evolution of feathers: from their origin to the present. Springer, Cham
Gauthier JA (1986) Saurischian monophyly and the origin of birds. Memoirs Calif Acad Sci 8:1–55
Gierlinski G (1997) What type of feather could nonavian dinosaur have, according to an Early Jurassic ichnological evidence from Massachusetts? Przeglad Geologiczny 54:419–422
Godefroit P, Demuynck H, Dyke GJ, Hu D, Escuillié F, Claeys P (2013a) Reduced plumage and flight ability of a new Jurassic paravian theropod from China. Nat Commun 4:1394
Godefroit P, Cau A, Hu D, Escuillié F, Wu W, Dyke GJ (2013b) A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature 498:359–362
Godefroit P, Sinitsa SM, Dhouailly D, Bolotsky YL, Sizov AV, McNamara ME, Benton MJ, Spagna P (2014) A Jurassic ornithischian dinosaur from Siberia with both feathers and scales. Science 345:451–455
Godefroit P, Sinitsa SM, Cincotta A, McNamara ME, Reshetova SA, Dhouailly D (2020) Integumentary structures in Kulindadromeus zabaikalicus, a Basal Neornithischian Dinosaur from the Jurassic of Siberia. In: Foth C, Rauhut OWM (eds) The evolution of feathers: from their origin to the present. Springer, Cham
Heilmann G (1926) The origin of birds. Witherby, London
Hennig W (1966) Phylogenetic systematics. University of Illinois Press, Urbana
Hocknull SA, White MA, Tischler TR, Cook AC, Calleja ND, Sloan T, Elliot DA (2009) New Mid-Cretaceous (Latest Albian) Dinosaurs from Winton, Queensland, Australia. PLoS One 4:e6190
Holtz TRJ (1994) The phylogenetic position of the Tyrannosauridae: implications for theropod systematics. J Paleontol 68:1100–1117
Holtz TR (1996) Phylogenetic taxonomy of the Coelurosauria (Dinosauria: Theropoda). J Paleontol 70:536–538
Holtz TRJ (1998) A new phylogeny of the carnivorous dinosaurs. Gaia 15:5–61
Holtz TRJ, Molnar RE, Currie PJ (2004) Basal Tetanurae. In: Weishampel DB, Dodson P, Osmólska H (eds) The Dinosauria. University of California Press, Berkeley, pp 71–110
Hone DWE, Holtz TRJ (2017) A century of spinosaurs—a review and revision of the Spinosauridae with comments on their ecology. Acta Geologica Sinica 91:1120–1132
Hou L, Zhou Z, Martin LD, Feduccia A (1995) A beaked bird from the Jurassic of China. Nature 377:616–618
Hoyle F, Wickramasinghe C (1985) Archaeopteryx, the primordial bird: a case of fossil forgery. Christopher Davies, Swansea
Hu D, Hou L, Zhang L, Xu X (2009) A pre- Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461:640–643
Hu D, Clarke JA, Eliason CM, Qiu R, Li Q, Shawkey MD, Zhao C, D’Alba L, Jiang J, Xu X (2018) A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution. Nat Commun 9:217
Huxley TH (1868) On the animals which are most nearly intermediate between birds and reptiles. Ann Mag Nat Hist 4:66–75
Ji Q, Currie PJ, Norell MA, Ji S (1998) Two feathered dinosaurs from northeastern China. Nature 393:753–761
Kaye TG, Falk AR, Pittman M, Sereno PC, Martin LD, Burnham DA, Gong E, Xu X, Wang Y (2015) Laser-stimulated fluorescence in paleontology. PLoS One 10:e0125923
Kaye TG, Pittman M, Marugán-Lobón J, Martín-Abad H, Sanz JL, Buscalioni AD (2019) Fully fledged enantiornithine hatchling revealed by laser-stimulated fluorescence supports precocial nesting behavior. Sci Rep 9:5006
Kellner AWA (2002) A review of avian Mesozoic fossil feathers. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of Dinosaurs. University of California Press, Berkeley, pp 389–404
Kundrát M (2004) When did theropods become feathered? Evidence for pre-Archaeopteryx feathery appendages. J Exp Zool (MOL DEV EVOL) 302B:355–364
Langer MC, Rincón AD, Ramezani J, Solórzano A, Rauhut OWM (2014) New dinosaur (Theropoda, stem-Averostra) from the earliest Jurassic of the La Quinta Formation, Venezuelan Andes. R Soc Open Sci 1:140184
Lee Y-N, Barsbold R, Currie PJ, Kobayashi Y, Lee H-J, Godefroit P, Escuillié F, Tsogtbaatar C (2014) Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus. Nature 515:257–260
Lefèvre U, Cau A, Cincotta A, Hu D, Chinsamy A, Escuillié F, Godefroit P (2017) A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers. Sci Nat 104:74
Li Z, Zhou Z, Wang M, Clarke JA (2014) A new specimen of large-bodied basal Enantiornithine Bohaiornis from the early Cretaceous of China and the inference of feeding ecology in Mesozoic birds. J Paleontol 88:99–108
Lingham-Soliar T (2003a) The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers. Naturwissenschaften 90:563–567
Lingham-Soliar T (2003b) Evolution of birds: ichthyosaur integumental fibers conform to dromaeosaur protofeathers. Naturwissenschaften 90:428–432
Lingham-Soliar T (2010) Dinosaur protofeathers: pushing back the origin of feathers into the Middle Triassic. J Ornithol 151:193–200
Lingham-Soliar T (2012) The evolution of the feather: Sinosauropteryx, life, death and preservation of an alleged feathered dinosaur. J Ornithol 153:699–711
Lingham-Soliar T, Feduccia A, Wang X (2007) A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. Proc R Soc Lond B 274:1823–1829
Makovicky PJ, Sues H-D (1998) Anatomy and phylogenetic relationships of the theropod dinosaur Microvenator celer from the Lower Cretaceous of Montana. Am Museum Novitates 3240:1–27
Makovicky PJ, Norell MA, Clark JM, Rowe TB (2003) Osteology and relationships of Byronosaurus jaffei (Theropoda: Troodontidae). Am Museum Novitates 3402:1–32
Maleev EA (1954) Noviy chyeryepoobrazniy yashschyer Mongolii [A new turtle-like reptile from Mongolia]. Priroda 1954:106–108
Martin LD (1983) The origin and the early radiation of birds. In: Brush AH, Clark GA (eds) Perspectives in ornithology. Cambridge University Press, Cambridge, pp 291–338
Martínez RN, Apaldetti C (2017) A late Norian-Rhaetian coelophysid neotheropod (Dinosauria, Saurischia) from the Quebrada Del Barro Formation, northwestern Argentina. Ameghiniana 54:488–505
Martin LD, Stewart JD, Whetstone KN (1980) The origin of birds: structure of the tarsus and teeth. Auk 97:86–93
Maryańska T, Osmólska H, Wolsan M (2002) Avialian status for Oviraptorosauria. Acta Palaeontologica Polonica 47:97–116
Mayr G (2009) Paleogene fossil birds. Springer, Berlin
Mayr G (2010) Response to Lingham-Soliar: Dinosaur protofeathers: pushing back to origin of feathers into the Middle Triassic. J Ornithol 151:523–524
Mayr G (2017) Avian evolution. Wiley, Chichester
Norell MA, Clark JM, Makovicky PJ (2001) Phylogenetic relationships among coelurosaurian theropods. In: Gauthier JA (ed) New perspectives on the origin and early evolution of birds: Proceedings of the international symposium in Honor of John H. Ostrom. Yale University, New Haven, pp 49–67
Norell MA, Clark JM, Turner AH, Makovicky PJ, Barsbold R, Rowe TB (2006) A new droameosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia). Am Museum Novitates 3545:1–51
Novas FE (1992) Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 35:51–62
Novas FE (1996) Alvarezsauridae, Cretaceous basal birds from Patagonia and Mongolia. Mem Queensland Mus 39:675–702
Novas FE (1997) Anatomy of Patagonykus puertai (Theropoda, Avialae, Alvarezsauridae), from the Late Cretaceous of Patagonia. J Vertebr Paleontol 17:137–166
Novas FE (1998) Megaraptor namunhuaiquii, gen. et so. nov., a large-clawed, Late Cretaceous theropod from Patagonia. J Vertebr Paleontol 18:4–9
Novas FE, Agnolín FL, Ezcurra MD, Porfiri JD, Canale JI (2013) Evolution of the carnivorous dinosaurs during the Cretaceous: the evidence from Patagonia. Cretac Res 45:174–215
Novas FE, Salgado L, Suárez M, Agnolín FL, Ezcurra MD, Chimento NR, de al Cruz R, Isasi MP, Vargas AO, Rubular-Rogers D (2015) An enigmatic plant-eating theropod from the Late Jurassic period of Chile. Nature 522:331–334
Novas FE, Aranciaga Rolando AM, Agnolín FL (2016) Phylogenetic relationships of the Cretaceous Gondwanan theropods Megaraptor and Australovenator: the evidence afforded by their manual anatomy. Mem of Mus Vic 74:49–61
O’Connor JK, Wang X, Chiappe LM, Gao C, Meng Q, Cheng X, Liu J (2009) Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species. J Vertebr Paleontol 29:188–204
O’Connor JK, Chiappe LM, Bell A (2011a) Pre-modern birds: avian divergences in the Mesozoic. In: Dyke GJ, Kaiser GW (eds) Living dinosaurs: the evolutionary history of modern birds. John Wiley, Chichester, pp 39–119
O’Connor JK, Chiappe LM, Gao C, Zhao B (2011b) Anatomy of the Early Cretaceous enantiornithine bird Rapaxavis pani. Acta Palaeontologica Polonica 56:463–475
O’Connor JK, Zhang Y, Chiappe LM, Meng Q, Quanguo L, Di L (2013) A new enantiornithine from the Yixian Formation with the first recognized avian enamel specialization. J Vertebr Paleontol 33:1–12
O’Connor JK, Wang M, Hu H (2016) A new ornithuromorph (Aves) with an elongate rostrum from the Jehol Biota, and the early evolution of rostralization in birds. J Syst Palaeontol 14:939–948
Ortega F, Escaso F, Sanz JL (2010) A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature 467:203–206
Ostrom JH (1969a) A new theropod dinosaur from the Lower Cretaceous of Montana. Postilla 128:1–17
Ostrom JH (1969b) Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum Nat Hist Bull 30:1–165
Ostrom JH (1970) Archaeopteryx: notice of a ‘new’ specimen. Science 170:537–538
Ostrom JH (1972) Description of the Archaeopteryx specimen in the Teyler Museum, Haarlem. Proceedings Koninklijk Nederlandse Akademie van Wetenschappen, B 75:289–305
Ostrom JH (1973) The ancestry of birds. Nature 242:136
Ostrom JH (1976) Archaeopteryx and the origin of birds. Biol J Linnean Soc 8:91–182
Owen R (1863) On the Archaeopteryx of von Meyer, with a description of the fossil remains of a long-tailed species from the lithographic stone of Solenhofen. Philos Trans R Soc Lond 153:33–47
Paul GS (1984) The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition. J Vertebr Paleontol 4:507–515
Paul GS (2002) Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. The John Hopkins University Press, Baltimore
Perle A (1979) Segnosauridae - a new family of theropods from the Late Cretaceous of Mongolia. Trudy Sovmestnay Sovetsko-Mongolskay Paleontologiyeskay Ekspeditsiy 8:45–55
Perle A (1982) A hind limb of Therizinosaurus from the Upper Cretaceous of Mongolia. Probl in Mongolian Geol 5:94–98
Perle A, Chiappe LM, Barsbold R, Clark JM, Norell MA (1994) Skeletal morphology of Mononykus olecranus (Theropoda: Avialae) from the Late Cretaceous of Mongolia. Am Mus Novit 3105:1–19
Pol D, Rauhut OWM (2012) A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proc R Soc B 279:3170–3175
Porfiri JD, Novas FE, Calvo JO, Agnolín FL, Ezcurra MD, Cerda IA (2014) Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation. Cretac Res 51:35–55
Prado GMEM, Anelli LE, Petri S, Romero GR (2016) New occurrences of fossilized feathers: systematics and taphonomy of the Santana Formation of the Araripe Basin (Cretaceous), NE, Brazil. PeerJ 4:e1916
Prum RO (2002) Perspectives in ornithology. Why ornithologist should care about the theropod origin of birds. Auk 119:1–17
Prum RO (2003) Are current critiques of the theropod origin of birds science? Rebuttal to Feduccia (2002). Auk 120:550–561
Prum RO, Berv JS, Dornburg A, Field DJ, Townsend JP, Moriarty Lemmon E, Lemmon AR (2015) A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526:569–573
Rauhut OWM (1998) Elaphrosaurus bambergi and the early evolution of theropod dinosaurs. J Vertebr Paleontol 18:71A
Rauhut OWM (2003) The interrelationships and evolution of basal theropod dinosaurs. Spec Pap Palaeontol 69:1–213
Rauhut OWM, Pol D (2019) Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs. Sci Rep 9:18826
Rauhut OWM, Carrano MT (2016) The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania. Zool J Linnean Soc 178:546–610
Rauhut OWM, Xu X (2005) The small theropod dinosaurs Tugulusaurus and Phaedrolosaurus from the Early Cretaceous of Xinjiang, China. J Vertebr Paleontol 25:107–118
Rauhut OWM, Milner AC, Moore-Fay S (2010) Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zool J Linnean Soc 158:155–195
Rauhut OWM, Foth C, Tischlinger H, Norell MA (2012) Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. Proc Natl Acad Sci USA 109:11746–11751
Rauhut OWM, Hübner TR, Lanser K-P (2016) A new megalosaurid theropod dinosaur from the late Middle Jurassic (Callovian) of north-western Germany: implications for theropod evolution and faunal turnover in the Jurassic. Palaeontologia Electronica 19:26A
Rauhut OWM, Tischlinger H, Foth C (2019) A non-archaeopterygid avialan theropod from the Late Jurassic of southern Germany. eLife 8:e43789
Rietschel S (1985) Feathers and wings of Archaeopteryx and the question of her fligth ability. In: Hecht MK, Ostrom JH, Viohl G, Wellnhofer P (eds) The beginnings of birds. Freunde des Jura-Museums, Eichstätt, pp 251–260
Russell DA, Dong Z (1993) The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People’s Republic of China. Can J Earth Sci 30:2107–2127
Schachner ER, Hutchinson JR, Farmer CG (2013) Pulmonary anatomy in the Nile crocodile and the evolution of unidirectional airflow in Archosauria. PeerJ 1:e60
Schweitzer MH, Watt JA, Avci R, Knapp LW, Chiappe LM, Norell MA, Marshall M (1999) Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous alvarezsaurid, Shuvuuia deserti. J Exp Zool (MOL DEV EVOL) 285:146–157
Schweitzer MH, Avci R, Collier T, Goodwin MB (2008) Microscopic, chemical and molecular methods for examining fossil perservation. C R Palevol 7:159–184
Senter P (2007) A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). J Syst Palaeontol 5:429–463
Sereno PC (1997) The origin and evolution of dinosaurs. Annu Rev Earth Planetary Sci 25:435–489
Sereno PC (1998) A rationale for phylogenetic definitions with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210:41–83
Sereno PC (1999) The evolution of dinosaurs. Science 284:2137–2147
Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, Larsson HCE (2008) Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PLoS One 3:e3303
Smith ND, Makovicky PJ, Hammer WR, Currie PJ (2007) Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution. Zool J Linnean Soc 151:377–421
Smith NA, Chiappe LM, Clarke JA, Edwards SV, Nesbitt SJ, Norell MA, Stidham TA, Turner AH, van Tuinen M, Vinther J, Xu X (2015) Rhetoric vs. reality: a commentary on “Bird origins anew” by A. Feduccia. Auk 132:467–480
Smithwick F, Vinther J (2020) Review on color patterns of fossil feathers. In: Foth C, Rauhut OWM (eds) The evolution of feathers: from their origin to the present. Springer, Cham
Smithwick FM, Mayr G, Saitta ET, Benton MJ, Vinther J (2017) On the purported presence of fossilized collagen fibres in an ichthyosaur and a theropod dinosaur. Palaeontology 60:409–422
Stromer E (1915) Ergebnisse der Forschungsreisen Prof. Stromers in den Wüsten Ägyptens. II Wirbeltier-Reste der Baharîje-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften. Mathematisch-physikalische Klasse 28:1–32
Tarsitano S, Hecht MK (1980) A reconsideration of the reptilian relationships of Archaeopteryx. Zool J Linnean Soc 69:149–182
Therrien F, Henderson DM (2007) My theropod is bigger than yours...or not: estimating body size from skull length in theropods. J Vertebr Paleontol 27:108–115
Turner AH, Makovicky PJ, Norell MA (2012) A review of dromaeosaurid systematics and paravian phylogeny. Bull Am Museum Nat Hist 371:1–206
von Meyer H (1861a) Vogel-Federn und Palpipes priscus von Solnhofen. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde 1861:561
von Meyer H (1861b) Archaeopteryx lithographica und Pterodactylus. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde 1861:678–679
Wagner A (1861) Beiträge zur Kenntnis der urweltlichen Fauna des lithographischen Schiefers. 2. Abt. Schildkröten und Saurier. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften. Mathematisch-physikalische Klasse 9:67–124
Wagner A (1862) On a new fossil reptile supposed to be furnished with feathers. Ann Mag Nat Hist 9:261–267
Wang M, Zhou Z (2017) The evolution of birds with implications from new fossil evidences. In: Maina JN (ed) The biology of the avian respiratory system. Springer, Cham, pp 1–26
Wang M, Zheng X, O’Connor JK, Lloyd GT, Wang X, Wang Y, Zhang X, Zhou Z (2015) The oldest record of Ornithuromorpha from the early cretaceous of China. Nat Commun 6:6987
Wang S, Stiegler J, Amiot R, Wang X, Du G, Clark JM, Xu X (2017) Extreme ontogenetic changes in a ceratosaurian theropod. Curr Biol 27:1–5
Wang M, O’Connor JK, Xu X, Zhou Z (2019) A new Jurassic scansoriopterygid and the loss of membranous wings in theropod dinosaurs. Nature 569:256–259
Wellnhofer P (1974) Das fünfte Skelettexemplar von Archaeopteryx. Palaeontographica Abt. A 147:169–216
Wellnhofer P (2008) Archaeopteryx: Der Urvogel von Solnhofen. Verlag Dr Friedrich Pfeil, München
Witmer LM (2002) The debate on avian ancestry: phylogeny, function, and fossils. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley, pp 3–30
Xing L, McKellar RC, Wang M, Bai M, O’Connor JK, Benton MJ, Zhang J, Wang Y, Tseng K, Lockley MG, Li G, Zhang W, Xu X (2016a) Mummified precocial bird wings in mid-Cretaceous Burmese amber. Nat Commun 7:12089
Xing L, McKellar RC, Xu X, Li G, Bai M, Persons WS IV, Miyashita T, Benton MJ, Zhang J, Wolfe AP, Yi Q, Tseng K, Ran H, Currie PJ (2016b) A feathered dinosaur tail with primitive plumage trapped in mid-Cretaceous amber. Curr Biol 26:1–9
Xing L, McKellar RC, O’Connor JK, Niu K, Mai H (2019) A mid-Cretaceous enantiornithine foot and tail feather preserved in Burmese amber. Sci Rep 9:15513
Xu X (2006) Feathered dinosaurs from China and the evolution of major avian characters. Integr Zool 1:4–11
Xu X (2020) Filamentous integuments in nonavialan theropods and their kin: advances and future perspectives for understanding the evolution of feathers. In: Foth C, OWM R (eds) The evolution of feathers: from their origin to the present. Springer, Cham
Xu X, Fucheng F (2005) A new maniraptoran dinosaur from China with long feathers on the metatarsus. Naturwissenschaften 92:173–177
Xu X, Guo Y (2009) The origin and early evolution of feathers: insights from recent paleontological and neontological data. Vertebrata PalAsiatica 47:311–329
Xu X, Pol D (2014) Archaeopteryx, paravian phylogenetic analyses, and the use of probability-based methods for palaeontological datasets. J Syst Palaeontol 12:323–334
Xu X, Wang X, Wu X (1999a) A dromaeosaurid dinosaur with filamentous integument from the Yixian Formation of China. Nature 401:262–266
Xu X, Tang Z, Wang X (1999b) A therizinosauroid dinosaur with integumentary structures from China. Nature 399:350–354
Xu X, Zhou Z, Prum RO (2001) Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature 410:200–204
Xu X, Norell MA, Wang X, Makovicky PJ, Wu X (2002) A basal troodontid from the Early Cretaceous of China. Nature 415:780–784
Xu X, Zhou Z, Wang X, Kuang X, Zhang F, Du X (2003) Four-winged dinosaurs from China. Nature 421:335–340
Xu X, Tan Q, Wang J, Zhao X, Tan L (2007) A gigantic bird-like dinosaur from the Late Cretaceous of China. Nature 447:844–847
Xu X, Clark JM, Mo J, Choiniere JN, Forster CA, Erickson GM, Hone DWE, Sullivan C, Eberth DA, Nesbitt SJ, Zhao Q, Hernandez R, Jia C, Han F, Guo Y (2009) A Jurassic ceratosaur from China helps clarify avian digital homologies. Nature 459:940–944
Xu X, Ma Q, Hu D (2010) Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins. Chin Sci Bull 55:3971–3977
Xu X, You H, Du K, Han F (2011) An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475:465–470
Xu X, Wang K, Zhang K, Ma Q, Xing L, Sullivan C, Hu D, Cheng S, Wang S (2012) A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature 484:92–95
Xu X, Zhou Z, Dudley R, Mackem S, Chuong C, Erickson GM, Varricchio DJ (2014) An integrative approach to understanding bird origins. Science 346:1253293
Xu X, Zheng X, Sullivan C, Wang W, Xing L, Wang Y, Zhang X, O’Connor JK, Zhang F, Pan Y (2015) A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature 521:70–73
Xu X, Zhou Z, Sullivan C, Wang Y, Ren D (2016) An updated review of the Middle-Late Jurassic Yanliao Biota: chronology, taphonomy, paleontology and paleoecology. Acta Geol Sin 90:2229–2243
Xu X, Choiniere JN, Tan Q, Benson RBJ, Clark JM, Sullivan C, Zhao Q, Han F, Ma Q, He Y, Wang S, Xing H, Tan L (2018) Two Early Cretaceous fossils document transitional stages in alvarezsaurian dinosaur evolution. Curr Biol 28:1–8
Zanno LE (2010) A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora). J Syst Palaeontol 8:503–543
Zanno LE, Makovicky PJ (2011) Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proc Natl Acad Sci USA 108:232–237
Zhang F, Zhou Z, Xu X, Wang X (2002) A juvenil coelurosaurian theropod from China indicates arboreal habits. Naturwissenschaften 89:394–398
Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (2008) A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 455:1105–1108
Zhang Y, O’Connor JK, Di L, Meng Q, Sigurdsen T, Chiappe LM (2014) New information on the anatomy of the Chinese Early Cretaceous Bohaiornithidae (Aves: Enantiornithes) from a subadult specimen of Zhouornis hani. PeerJ 2:e407
Zheng X, You H, Xu X, Dong Z (2009) An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature 458:333–336
Zhou Z, Clarke JA, Zhang F (2008) Insight into diversity, body size and morphological evolution from the largest Early Cretaceous enantiornithine bird. J Anat 212:565–577
Zhou Z, Zhang Z, Li Z (2010) A new Lower Cretaceous bird from China and tooth reduction in early avian evolution. Proc R Soc B 277:219–227
Acknowledgements
This work was supported by the Volkswagen Foundation under grant I/84 640 (to OR) and the Swiss National Science Foundation under grant PZ00P2_174040 (to CF).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Rauhut, O.W.M., Foth, C. (2020). The Origin of Birds: Current Consensus, Controversy, and the Occurrence of Feathers. In: Foth, C., Rauhut, O. (eds) The Evolution of Feathers. Fascinating Life Sciences. Springer, Cham. https://doi.org/10.1007/978-3-030-27223-4_3
Download citation
DOI: https://doi.org/10.1007/978-3-030-27223-4_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-27222-7
Online ISBN: 978-3-030-27223-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)