
Abstract
This chapter describes the efficiency of bumble bees as biocontrol agents and also, which factors should be taken into account when, using them. Bumble bee mediated biocontrol of grey mould (Botrytis cinerea Pers.:Fr.) decreased the infection up to three times in open field strawberry. The presence of other flower visitors foraging on strawberry aid the dissemination of BCAs. The open conditions set some obstacles, which must be overcome. The flowering stage of target crop and availability of alternative food sources affect the bumble bee foraging. Knowing the bumble bee foraging behaviour helps to find out right solutions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Management of bumble bees for delivering biocontrol agents has been studied for more than 20 years (Peng et al. 1992; Yu and Sutton 1997). Most of the research, however, has been done mainly in laboratory or greenhouse conditions (Kevan et al. 2003; Mommaerts et al. 2011). The reason for using bees as vectors for biocontrol agents (BCA) stays in their morphological and behavioural characteristics. Bumble bees have relatively large body surface covered with branched hair, which aids trapping and transporting pollen grains (Free and Williams 1972; Batra et al. 1973). Similarly to pollen, the spores of microorganisms can stick to the fur, which characteristic has been put in use in entomovector technology to deliver BCAs to the target crop (Fig. 1). The commercial availability of bumble bee colonies has enabled the increase in usage of the buff-tailed bumble bee Bombus terrestris L. in Europe and the common eastern bumble bee B. impatiens Cresson (Hymenoptera: Apidae) in North America not only in greenhouse (Mommaerts et al. 2011) but also in field conditions (Kovach et al. 2000; Dedej et al. 2004; Karise et al. 2016a). In this chapter the potential of bumble bees as vectors of BCAs in open field conditions will be considered. In addition, we talk also about several aspects, which have to be taken into account if harnessing bumble bees as vectors in open fields.

A bumble bee covered all over the body with a large amount of Prestop Mix powder, at the time of homing all the powder was gone (Photo: Reet Karise)
2 Bumble Bee Efficacy in Open Fields
2.1 Grey Mould Suppression
The data, collected during BICOPOLL project, confirmed that the bumble bees proved to be effective in mediating biofungicide Prestop-Mix (Gliocladium catenulatum Strain J1446 as active organism, Verdera OY, Finland) in open field conditions. Prestop Mix is a biological preparation, which is safe both to humans and beneficial organisms visiting the fields (Verdera 2015). The infection rate decreased approximately 1.5–3 times when pathogen pressure was light or moderate, but no change was seen when there was high pathogen pressure due to heavy rain and cool temperature conditions (Fig. 2) (Karise et al. 2016a). Higher rainfall and colder temperatures during the fruit maturing period create particularly good conditions for the pathogen B. cinerea (Wilcox 1994; Cota et al. 2009) by which the infection rate on berries might rise up to 70–80%. In these conditions, also chemical control most likely could fail without proper decision supporting systems (Evenhuis and Wilms 2009). The efficacy of honey and bumble bee-vectored biocontrol has been found to be comparable to synthetic fungicide spraying (Kovach et al. 2000).

Grey mould infection rate on control plots (not treated) and plots treated with bumble bee-mediated Prestop-Mix. The means with standard error bars are presented. Asterisks (∗∗∗) indicate a significant decrease (P < 0.001) in the infection rate during the years with low pathogen pressure (from Karise et al. 2016a)
2.2 Inoculum Dissemination: Safety and Efficacy
The basis for dissemination task is the ability of the preparations to adhere to and get released from the hair of a bee easily. The amounts of powdery preparations, stuck to the hairs of the bees exiting the nest box, are variable. Some bees are covered only ventrally, some others however all over their bodies, although the larger amounts of the powders may disturb the vectoring bees, they lose most of it within first 60 s (Mommaerts et al. 2010). Both bumble bees and honey bees might suffer some-what when covered with larger amounts of the powders containing kaolin in closed experimental conditions (Karise et al. 2016b, 2018). Though, the effect was never noticed on the field nor in greenhouses when performing the BCA vectoring task. In addition, the commercial bumble bee hives have short life-span and are not meant to function longer than the time needed for pollination of the target crop. Even with the shortened individual life-span, the number of days a bee lived, was suitable to fulfil the pollination and vectoring tasks (Fig. 3). Commercial bumble bee hives are well suitable also with bioinsecticides, despite some of the entomopathogenic fungi can infect also the vectoring bees (reviewed by Mänd et al. 2010).

Lethal effects of exposure to kaolin, Prestop-Mix, BotaniGard and wheat flour on the survival on bumblebees (B. terrestris). A and B present the longevity of bumblebees (days) and the survival probability (%) at 18 °C, while C and D the respective data at 28 °C. The longevity data are expressed as box plots with the minimum, lower quartile, median, upper quartile and maximum values. The numbers upon the boxes denote medians and different letters indicate statistically significant differences between groups at p < 0.05. From Karise et al. 2016b
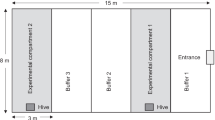
The efficient biocontrol can be achieved only when the BCAs are spread evenly over the entire field. In this purpose, it is important to study the bumble bee density on the flowers of the target crop. According to BICOPOLL project results, the bumble bee dispersal over the field was equable over a distance of 100 m (Karise et al. 2016a). The second important aspect is the dispersal rate of the BCAs. The results showed also the even distribution of the G. catenulatum on flowers. Indeed, this study was conducted on relatively small strawberry fields, but the good distribution of Clonostachys rosea (Link) Schroers by B. impatiens has been observed up to 150 m (Reeh et al. 2014). Wolf and Moritz (2008) have shown the mean foraging distance of B. terrestris being 267 m, whereas 40% of bumble bees foraged within the radius of 100 m. Thus the distance between bumble bee hives on larger fields could be around 200–300 m to guarantee the good visitation rate for the strawberry.
2.3 The Secondary Transmission of BCA
The greenhouse experiments of BICOPOLL project indicate, that bumble bees lose about 81% of Prestop-Mix already within the first 60 s of their flight (Mommaerts et al. 2010). The sequential visits of any other flower visitors aid the transportation of BCAs from flower to flower (Maccagnani et al. 2009). This phenomenon has been called the secondary inoculation and may play important role in efficacy of the technology (Nuclo et al. 1998). The rate of newly opened flowers and weather conditions, which favour insect visitation of flowers, affect the efficacy of secondary transmission of BCAs (Nuclo et al. 1998; Maccagnani et al. 2009). In the northern regions, the strawberry flowering occurs at the time when there are almost no other numerous wild bumble bees foraging, since the newly emerged overwintered queens are establishing their colonies yet. Still, at that time the numbers of foraging honey bees, several solitary bee species and also different species of dipterans might be quite high. The BICOPOLL field study showed, that the most abundant groups in Estonia were dipterans including syrphid flies which formed 49% of the number of all flower visitors, followed by honey bees 29% and solitary bees 13%. Ahrenfeldt et al. (2015) showed that the wild bee species diversity and community composition on strawberry has a north-south gradient from Mid-Norway through Denmark to Germany. The diversity of these insects is higher in the southernmost regions. So, it is possible that the effect of the secondary dissemination on the entomovectoring is higher in regions with higher insect abundance and diversity.
2.4 Additive Value from Pollination
The additive value from applying the entomovector technique comes through enhancing the pollination of target crop. The direct benefit from bee-pollination depends on the crop species and cultivar. There are plant species, which give almost no yield without animal pollination: for instance, blueberries, raspberries, apples, cherries and plums benefit largely from insect pollination. Strawberries, however, belong to those species by which the effect from pollination depends on the cultivar (Klatt et al. 2014a, b; Tuohimetsä et al. 2014). There are more than 200 stigmas in the strawberry flowers (Free 1993; Perkins-Veazie 1995) and each of these needs to be fertilized in order to achieve high quality fruits. In some strawberry cultivars the pollen grains can be released from the anthers with the help of wind. Other cultivars need a high functional insect diversity to get the flowers fully pollinated (Klein et al. 2007). Fruits from properly pollinated flowers are not only greater but also have longer shelf-life and highest commercial value (Klatt et al. 2014a, b). In BICOPOLL experiment, still, no increase was found in fruit weight due to insect pollination (Karise et al. 2016a). This suggests that the cultivar ‘Sonata’ used in those experiments can cope with wind pollination.
3 Bumble Bee Foraging Behaviour
When using bumble bees on open fields it is very important to understand basics of bumble bee foraging behaviour in order to find out right solutions.
3.1 Flower Selection
Bumble bee flower preference is based on several aspects. Food plant choice of bees depend on the amount and quality of nectar and pollen provided by the flowers present in the foraging area. The nectar production of flowers can vary between the cultivars (Bertazzini and Forlani 2016) and also depends on the weather conditions (Nicolson and Nepi 2005). For example, in moist conditions the nectar sugar content is low and larger amounts of nectar are needed to satisfy the colony needs (Shackleton et al. 2016). The effect of cultivars varies across the years, since different weather conditions favour different properties of the cultivars, best seen in hybrid lucerne (Karise et al. 2006). The food resource quantity and quality may also be manipulated by farmers, as demonstrated by Viik et al. (2012): proper fertilization increases both the numbers of flowers and the amounts of nectar per flower, which serve as main attractants for bees. In case of toxic compounds, e.g. pesticides in bee feed, the behaviour of foragers can be changed (Koskor et al. 2009).
The other important characteristic is the tongue length of the bee, which influences the flower selection and in turn the breadth of the diet (Teräs 1985). Short-tongued bumble bees usually have wider diet compared to long-tongued bumble bees, who prefer flowers with narrower and deeper corollas (Alford 1975; Goulson et al. 2008). Bees with short tongues do not pollinate properly flowers of red clover and field bean for instance, however open flowers of strawberry have no demands on the tongue length of the pollinator. The commercial bumble bee B. terrestris has relatively short tongue, however, the bee is able to feed on different flower types. In order to get access to the nectaries hidden in the bottom of deep corollas, B. terrestris often bites holes into corolla tubes and thus robs nectar without pollinating the flower. This bumble bee species very easily broadens their forage plant selection to non-native garden plants and mass-flowering crops (Goulson et al. 2002). The wide diet of B. terrestris enables them easily to react on changes in agricultural landscape and human activity. This also provides us the possibility to force bumble bees to forage on plant species not naturally in their diet. Some crops flower in the time, when there is no B. terrestris foragers available naturally.
3.2 Flower and Forage Patch Constancy
Bumble bee B. terrestris is a food generalist visiting several (2–4) plant species within the same foraging trip (Carvell et al. 2006; Parmentier et al. 2014; Somme et al. 2014). Bumble bees have no innate plant species preference. Unlike honey bees they constantly search for new profitable flower types, even in case of plentiful food resources, as it happens on large fields. It is suggested, that flower constancy may have emerged to save energy and/or time of the foragers (Free 1970; Dukas 1995; Gegear and Thomson 2004). Compared to honey bees the probability of spreading the BCAs on the non-target crop is quite high with B. terrestris.
Bumble bees generally prefer larger patches, and at the same time avoid less rewarding patches regardless of size (Makino et al. 2007). They also have shown strong constancy on sites, at which they already have found nectar and pollen resources (Osborne and Williams 2001; Cartar 2004), and visit same patches repeatedly until the site is still rewarding (Makino and Sakai (2007). In purpose of intensification of the forage effort, bumble bees recruit their nest mates to beneficial food sources by touching each other and releasing pheromone signals (Dornhaus and Chittka 1999, 2001; Ayasse and Jarau 2014). Beside the recruitment behaviour, the foragers systematically search for new flower resources. The distribution of foragers among patches depends on the relationship of recruitment rate and patch size and also on how long the individual forager spends in the particular patch (Renner and Nieh 2008).
3.3 Colony Size and Foraging Range
Colony size and flight range of the foragers affects the number of flower visiting bumble bees. In commercial crop production, it is very important to gain as much pollinating individuals as possible. The large colonies with lots of brood have higher need for pollen and nectar. According to this B. terrestris is one of the best succeeding bumble bee species with up to 400 workers in a colony, whereas in the colonies of B. muscorum and B. sylvarum e.g. only 20–100 workers are present (Benton 2006).
Bumble bees are central place foragers, which means that they collect food from the surroundings of their nest. B. terrestris has the maximum detected foraging range compared to other bumble bee species. Depending on different study methods, the observed foraging distances of the workers vary greatly even up to 4 km from the nest (Goulson and Stout 2001). The mass-marking experiments (Osborne et al. 2008) and radio-tracking (Hagen et al. 2011) have shown that B. terrestris workers can forage up to 2.5 km, although generally it remains between 500 and 1750 m from the nests (Walther-Hellwig and Frankl 2000; Westphal et al. 2006). The species with small colonies have usually foraging areas up to 500 m in radii (Benton 2006). In context of entomovectoring, however, there is concern, that longer foraging distance of B. terrestris can cause dispersing of bumble bees on larger areas, which in turn will affect the efficiency of the technology.
3.4 Compatibility With Temperate Climate Conditions
Probably commercial bumble bees are most effective at the time there are few other pollinators available and the flowering occurs early in the season, when the daily temperatures stay cool. Bumble bees are pollinators, who are able to forage in cool temperatures. The heat generation ability in bumble bees is different from most of insects. During warming-up, bumble bees use their flight muscles without moving the wings (Heinrich 1979), thick body hair insulates the temperature into the thorax allowing to keep the heat (Newsholme et al. 1972; Peat et al. 2005). Some arctic bumble bees are able to forage even when the air temperature is below zero. They can forage even with light rain or fog if needed. In temperate regions, the bumble bee foraging starts at 5 degrees if there are no food supplies in hives left. A bumble bee colony stores very little nectar in the hive (Alford 1975) and this forces bumble bees to forage whenever it is possible. They even expand the daily foraging also into early morning and late evening periods.
4 Steering Bumble Bees to the Target Crop
Bringing bees as extra pollinators to open fields always holds the risk that they prefer some other plants species over the target crop. Steering them to any crop needs some certain knowledge on the behaviour and requirements of the particular bee species. Most often honey bees are used as extra pollinators due to their bigger and longer-living colonies. Commercial bumble bees, on the contrary, are easier to use, their colonies are smaller and perish soon after the pollination task is fulfilled. Bumble bees are usually not as aggressive in protection their hive compared to honey bees.
4.1 Handling of Bumble Bees
The handling of commercial bumble bees is easy. Bumble bee hive is small and compact. The producers supply the hives suitable for outdoor using: hives are placed in waterproof and insulaedt boxes. The entrances of the hives can be closed and opened, so that the bees can only enter or move in- and out-wards. For example, Biobest Flying Doctors Hive (B. terrestris) has already built-in compartment for the BCA preparation.
The commercial bumble bee hives are supplied with liquid sugar solution. When taking the hives on field there are pros and cons on eliminating the sugar syrup. Eliminating it would encourage bumble bees to forage more actively. However, the extra food might be necessary in case of unsuitable weather conditions.
4.2 Synchronizing Pollination Service with Crop Flowering
Using commercial bumble bee colonies allows to synchronize the availability of flowers of target crops and insects, by taking the hives onto the fields exactly after flowering has started. This is important, because forager bees develop flower constancy and may stick on other plant species, which were in flower before the target crop. For strawberry, it is suggested that bumble bees should be taken to the fields when 5–10% of flowers are open, thus there is enough available forage for them. When there are too few flowers available, bees start searching alternative food source and learn to forage elsewhere. For the strawberry grower, it is also important to have bees on fields in a very right time, since the first flowers are those giving fruits with the highest quality and the protective effect of bumble bee-mediated Prestop-Mix was highest for the yield from the first flowers (Karise et al. 2016a).
4.3 Crop and Cultivar Selection
The attractiveness of flowers of different crops vary from cultivar to cultivar. In addition, the attractiveness of a crop to bees depends on which alternative food plants are available in the foraging area. To guarantee higher visitation rate, it is suggested to grow different cultivars within the same field. Varying sugar concentration of the nectar among cultivars has been seen to affect the bumble bee visitation rate (Yu and Sutton 1997; Kovach et al. 2000; Escande et al. 2002).
It is not needed to endeavour a 100% flower constancy. Our results indicate that effective disease control was achieved with about 22% strawberry pollen (Figs. 4 and 5) gathered by bumble bees (Karise et al. 2016a). Taking into account the bumble bee habit to try other plant species for food, it could be suggested to provide even some flowering plants nearby the strawberry field. During the 3 years of BICOPOLL project, the bumble bee preference for simultaneously gathered pollen, which was collected beside strawberry, varied between the years and places depending on which plants were available. The caragana, white clover, white nettle or Rosaceae species were gathered during the same foraging trips. The regional differences in environmental conditions within Europe are huge and therefore the region-specific data is needed for successive agricultural practice. The winter oilseed rape as a very attractive forage plant is usually suspected to draw bumble bees away from other crops. In case of strawberry this effect was not observed. The amount of strawberry pollen in bumble bee corbiculae was 20–25% each year independent of surrounding plant communities.

Strawberry pollen grain. (Photo: Märt Rahi)

A bumble bee carrying pollen into the hive. (Photo: Reet Karise)
5 Conclusion
The success of using bumble bees in outdoor conditions depends on several interacting factors. However, knowing the specific behavioural aspects of the bee species used and the characteristics of target crop, make it possible to achieve reliable control of the disease.
References
Ahrenfeldt E, Klatt B, Arildsen J, Trandem N, Andersson G, Tscharntke T, Smith H, Sigsgaard L (2015) Pollinator communities in strawberry crops-variation at multiple spatial scales. Bull Entomol Res 105:497–506
Alford DV (1975) Bumblebees. Davis-Poynter, London
Ayasse M, Jarau S (2014) Chemical ecology of bumble bees. Annu Rev Entomol 59:299–319
Batra LR, Batba SWT, Bohart GE (1973) The mycoflora of domesticated and wild bees (Apoidea). Mycopathol Mycol Appl 49:13–44
Benton T (2006) Bumblebees. Collins New Naturalist Series, London
Bertazzini M, Forlani G (2016) Intraspecific variability of floral nectar volume and composition in rapeseed (Brassica napus L. var. oleifera). Front Plant Sci 7:288
Cartar RV (2004) Resource-tracking by bumble bees: responses to plant-level differences in quality. Ecology 85:2764–2771
Carvell C, Westrich P, Meek WR, Pywell RF, Nowakowski M (2006) Assessing the value of annual and perennial forage mixtures for bumblebees by direct observation and pollen analysis. Apidologie 37:326–340
Cota LV, Maffia LA, Mizubuti ESG, Macedo PEF (2009) Biological control by Clonostachys rosea as a key component in the integrated management of strawberry gray mold. Biol Control 50:222–230
Dedej S, Delaplane KS, Scherm H (2004) Effectiveness of honey bees in delivering the biocontrol agent Bacillus subtilis to blueberry flowers to suppress mummy berry disease. Biol Control 31:422–427
Dornhaus A, Chittka L (1999) Insect behaviour: evolutionary origins of bee dances. Nature 401:38
Dornhaus A, Chittka L (2001) Food alert in bumblebees (Bombus terrestris): possible mechanisms and evolutionary implications. Behav Ecol Sociobiol 50:570–576
Dukas R (1995) Transfer and interference in bumblebee learning. Anim Behav 49:1481–1490
Escande AR, Laich FS, Pedraza MV (2002) Field testing of honeybee-dispersed Trichoderma spp. to manage sunflower head rot (Sclerotinia sclerotiorum). Plant Pathol 51:346–351
Evenhuis A, Wilms JAM (2009) Designing strategies to control grey mould in strawberry cultivation using decision support systems. Acta Hortic 842:247–250
Free JB (1970) The flower constancy of bumblebees. J Anim Ecol 39:395–402
Free JB (1993) Insect Pollination of Crops, 2nd edn. Academic, London
Free JB, Williams IH (1972) Hoarding by honey bees (Apis mellifera L.). Anim Behav 20:327–334
Gegear RJ, Thomson JD (2004) Does the flower constancy of bumble bees reflect foraging economics? Ethology 110:793–805
Goulson D, Stout JC (2001) Homing ability of the bumblebee Bombus terrestris (Hymenoptera: Apidae). Apidologie 32:105–112
Goulson D, Hughes WOH, Derwent LC, Stout JC (2002) Colony growth of the bumblebee, Bombus terrestris, in improved and conventional agricultural and suburban habitats. Oecologia 130:267–273
Goulson D, Lye GC, Darvill B (2008) Diet breadth, coexistence and rarity in bumblebees. Biodivers Conserv 17:3269–3288
Hagen M, Wikelski M, Kissling WD (2011) Space use of bumblebees (Bombus spp.) revealed by radio-tracking. PLoS One 6(5):e19997
Heinrich B (1979) Bumblebee economics. Harvard University Press, Cambridge
Karise R, Mänd M, Ivask M, Koskor E, Bender A (2006) The effect of pollen amount and its caloric value in hybrid lucerne (Medicago x varia) on its attractiveness to bumble bees (Bombus terrestris). Agron Res 4:211–2016
Karise R, Dreyersdorff G, Jahani M, Veromann E, Runno-Paurson E, Kaart T, Smagghe G, Mänd M (2016a) Reliability of the entomovector technology using Prestop-Mix and Bombus terrestris L. as a fungal disease biocontrol method in open field. Sci Rep 6:31650
Karise R, Muljar R, Smagghe G, Kaart T, Kuusik A, Dreyersdorff G, Williams IH, Mänd M (2016b) Sublethal effects of kaolin and the biopesticides Prestop-Mix and BotaniGard on metabolic rate, water loss and longevity in bumble bees (Bombus terrestris). J Pest Sci 89:171–178
Karise R, Raimets R, Dreyersdorff G, Mänd M (2018) Using respiratory physiology techniques in assessments of pesticide effects on bees. In: Oomen P, Pistorius J (Ed.) Hazards of pesticides to bees: 13th International Symposium of the ICP-PR Bee Protection Group; October 18–20 (2017) València (Spain) – Proceedings, Julius Kühn-Institut, p 61–66
Kevan PG, Al-Mazra’awi MS, Sutton JC, Tam L, Boland G, Broadbent B, Thompson SV, Brewer GJ (2003) Using pollinators to deliver biological control agents against crop pests. In: Downer RA, Mueninghoff JC, Volgas GC (eds) Pesticide formulations and delivery systems: meeting the challenges of the current crop protection industry. American Society Testing and Materials, W Conshohocken, pp 148–153
Klatt BK, Holzschuh A, Westphal C, Clough Y, Smit I, Pawelzik E, Tscharntke T (2014a) Bee pollination improves crop quality, shelf life and commercial value. Proc R Soc B 281:20132440
Klatt BK, Klaus F, Westphal C, Tscharntke T (2014b) Enhancing crop shelf life with pollination. Agric Food Secur 3:14
Klein A-M‚ Vaissiere BE‚ Cane JH‚ Steffan-Dewenter I‚ Cunningham SA‚ Kremen C‚ Tscharntke T (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc Lond B Biol Sci 274(1608): 303–313
Koskor E, Muljar R, Drenkhan K, Karise R, Bender A, Viik E, Luik A, Mänd M (2009) The chronic effect of the botanical insecticide Neem EC on the pollen forage of the bumble bee Bombus terrestris L. Agron Res 7:341–346
Kovach J, Petzoldt R, Harman GE (2000) Use of honeybees and bumble bees to disseminate Trichoderma harzianum 1295–22 to strawberries for Botrytis control. Biol Control 18:235–242
Maccagnani B, Giacomello F, Fanti M, Gobbin D, Maini S, Angeli G (2009) Apis mellifera and Osmia cornuta as carriers for the secondary spread of Bacillus subtilis on apple flowers. BioControl 54:123–133
Makino TT, Sakai S (2007) Experience changes pollinator responses to floral display size: from size-based to reward-based foraging. Funct Ecol 21:854–863
Makino TT, Ohashi K, Sakai S (2007) How do floral display size and density of surrounding flowers influence the likelihood of bumble bee revisitation to a plant? Funct Ecol 21:87–95
Mänd M, Williams IH, Viik E, Karise R (2010) Oilseed rape, bees and integrated pest management. In: Williams IH (ed) Biocontrol-based integrated management of oilseed rape Pests. Springer, London, pp 357–379
Mommaerts V, Put K, Vandeven J, Jans K, Sterk G, Hoffmann L, Smagghe G (2010) Development of a new dispenser for microbiological control agents and evaluation of dissemination by bumblebees in greenhouse strawberries. Pest Manag Sci 66:1199–1207
Mommaerts V, Put K, Smagghe G (2011) Bombus terrestris as pollinator-and-vector to suppress Botrytis cinerea in greenhouse strawberry. Pest Manag Sci 67:1069–1075
Newsholme EA, Crabtree B, Higgins SJ, Thornton SD, Start C (1972) The activities of fructose diphosphatase in flight muscles from the bumble-bee and the role of this enzyme in heat generation. Biochem J 128:89–97
Nicolson SW, Nepi M (2005) Dilute nectar in dry atmospheres: nectar secretion patterns in Aloe castanea (Asphodelaceae). Int J Plant Sci 166:227–233
Nuclo RL, Johnson KB, Stockwell VO, Sugar D (1998) Secondary colonization of pear blossoms by two bacterial antagonists of the fire blight pathogen. Plant Dis 82:661–668
Osborne JL, Williams IH (2001) Site constancy of bumble bees in an experimentally patchy habitat. Agric Ecosyst Environ 83:129–141
Osborne JL, Martin AP, Carreck NL, Swain JL, Knight ME, Goulson D, Hale RJ, Sanderson RA (2008) Bumblebee flight distances in relation to the forage landscape. J Anim Ecol 77:406–415
Parmentier L, Meeus I, Cheroutre L, Mommaerts V, Louwye S, Smagghe G (2014) Commercial bumblebee hives to assess an anthropogenic environment for pollinator support: a case study in the region of Ghent (Belgium). Environ Monit Assess 186:2357–2367
Peat J, Darvill B, Ellis J, Goulson D (2005) Effects of climate on intra- and interspecific size variation in bumble-bees. Funct Ecol 19:145–151
Peng G, Sutton JC, Kevan PG (1992) Effectiveness of honey bees for applying the biocontrol agent Gliocladium roseum to strawberry flowers to suppress Botrytis cinerea. Can J Plant Pathol 14:117–129
Perkins-Veazie P (1995) Growth and ripening of strawberry fruit. In: Janick J (ed) Horticultural reviews, vol 17. Wiley, Oxford, pp 267–297
Reeh KW, Hillier NK, Cutler GC (2014) Potential of bumble bees as bio-vectors of Clonostachys rosea for Botrytis blight management in lowbush blueberry. J Pest Sci 87:543–550
Renner MA, Nieh JC (2008) Bumble bee olfactory information flow and contact-based foraging activation. Insect Soc 55:417–424
Shackleton K, Balfour NJ, Al Toufailia H, Gaioski R, de Matos Barbosa M, Silva CA de S, Bento JMS, Alves DA, Ratnieks FLW (2016) Quality versus quantity: foraging decisions in the honeybee (Apis mellifera scutellata) feeding on wildflower nectar and fruit juice. Ecol Evol 6:7156–7165
Somme L, Vanderplanck M, Michez D, Lombaerde I, Moerman R, Wathelet B, Wattiez R, Lognay G, Jacquemart A-L (2014) Pollen and nectar quality drive the major and minor floral choices of bumble bees. Apidologie 46:92–106
Teräs I (1985) Food plants and flower visits of bumblebees (Bombus: Hymenoptera, Apidae) in southern Finland. Acta Zool Fenn 179
Tuohimetsä S, Hietaranta T, Uosukainen M, Kukkonen S, Karhu S (2014) Fruit development in artificially self- and cross-pollinated strawberries (Fragaria × ananassa) and raspberries (Rubus idaeus). Acta Agric Scand Sect B Soil Plant Sci 64:408–415
Verdera: Safety Data. Available at: http://verdera.fi/en/products/horticulture/prestop-mix/safety-data/. Accessed 7 Dec 2015
Viik E, Mänd M, Karise R, Lääniste P, Williams IH, Luik A (2012) The impact of foliar fertilization on the number of bees (Apoidea) on spring oilseed rape. Žemdirbyste–Agriculture 99:41–46
Walther-Hellwig K, Frankl R (2000) Foraging distances of Bombus muscorum, Bombus lapidarius and Bombus terrestris (Hymenoptera, Apidae). J Insect Behav 13:239–246
Westphal C, Steffan-Dewenter I, Tscharntke T (2006) Bumblebees experience landscapes at different spatial scales: possible implications for coexistence. Oecologia 149:289–300
Wilcox WF (1994) Relationship between strawberry gray mold incidence, environmental variables, and fungicide applications during different periods of the fruiting season. Phytopathology 84:264–270
Wolf S, Moritz RFA (2008) Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 39:419–427
Yu H, Sutton JC (1997) Effectiveness of bumblebees and honeybees for delivering inoculum of Gliocladium roseum to raspberry flowers to control. Biol Control 10:113–122
Acknowledgements
The authors are thankful to Riin Muljar, Gerit Dreyersdorff, Anna Bontšutšnaja and other students for their assistance throughout the field work periods. This research was supported by the EU-ERANET activity of the CORE Organic Programme II project BICOPOLL, the Institutional Research Funding (IUT36-2) of the Estonian Ministry of Education and the Department Agriculture, and Fisheries of the Flemish government and the Flemish Research Organization IWT-Flanders.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Mänd, M., Karise, R., Smagghe, G. (2020). Bumble Bees and Entomovectoring in Open Field Conditions. In: Smagghe, G., Boecking, O., Maccagnani, B., Mänd, M., Kevan, P. (eds) Entomovectoring for Precision Biocontrol and Enhanced Pollination of Crops. Springer, Cham. https://doi.org/10.1007/978-3-030-18917-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-18917-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-18916-7
Online ISBN: 978-3-030-18917-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)