
Abstract
The revised edition of this chapter has been carefully and entirely rewritten. As a general rule, all sections have been revised and updated with current knowledge, more recent references included, and some sections expanded. Of particular note is the incorporation of significant additional materials in both sections dealing with some biological activities of lichen-derived secondary metabolites with particular emphasis on antitumor and anti-cellular agents and antiviral activities of lichen secondary metabolites. Every other section has been thoroughly revised and enriched with more current references.
Section 7.1 deals with broad and specific definitions of lichens and antiviral agents. Here, we will focus briefly on viruses and antiviral agents: their nature, source, general characteristics, and effects. Attempt is made to briefly classify antiviral agents based on their mode of action and virus targets. Lichen as a broad group of useful source of phytochemical agents is briefly described with emphasis on their unique niche in natural product research.
Section 7.2 deals with issues related to secondary metabolites (phytochemicals) from lichen. Lichen compounds because they possess some biological effects will form also the focus of this section. Some classes of lichen compounds are presented as well as their molecular structures. How they influence biological host and agents would be described briefly in the latter part of this section with presentation of some that display antiviral activities. The exact sources of these compounds are fully elucidated. Attempt is also made to mention lichen-derived compounds yet-to-be-fully-validated antiviral property.
Section 7.3 will focus on conclusion beginning with a summary of narrated up-to-date available data of antiviral lichen compounds currently undergoing preliminary and extensive research and development. How these agents have been applied or would be applied to biomedical and pharmaceutical utility would be addressed. The pharmaceutical industry involvement in further product development would be mentioned to display existing link between basic research setting and industry development endpoint. Finally, statement is made speculating on the future of lichen research and the positive expected outcomes of improved research interest in this unique group of medicinal plants.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

7.1 Antiviral Agents and Lichens
7.1.1 Antiviral Agents
Antiviral agents refer to chemicals, moiety, natural products, and medical and therapeutic substances employed for the prevention and treatment of virus infections and diseases. Therefore, viruses which cause a number of infections and diseases in plants, animals, and humans are targeted by these agents. Viruses are very tiny infectious agents that cannot carry out metabolism or replicate exclusively on their own. Following infection of a host (e.g., a cell), it can direct the cell machinery to produce viral proteins and eventually more viruses. Viruses have either RNA or DNA as their genetic material. This genetic material or nucleic acid may be either single or double stranded (designated ss or ds). The whole intact virus (virion) consists of the nucleic acid and an outer shell of protein. Antiviral agents could target the different proteins (substructure) of the respective virus (virion), its nucleic acid, and other virus or host cell factors to terminate or control the putative viral infection/disease. Viruses in contrast to bacteria and fungi are notoriously intracellular parasites living inside the cell of their host and harness the metabolic and replicative ability of their host cell to drive their own multiplication and proliferation within the host, thereby making intervention against them difficult. Consequently, antiviral agents have been shown to act by one or more of the following processes:
-
1.
Inactivating extracellular virus particles
-
2.
Preventing viral attachment and/or entry
-
3.
Preventing functioning or replication of viral genome
-
4.
Preventing synthesis or functioning of specific viral protein
-
5.
Preventing assembly or release of new infectious virions
Extensive screening efforts have been made to find antiviral agents from synthetic and natural sources. Natural products from plants traditionally have provided the pharmaceutical industry with one of its most important sources of lead compounds, and over 50% of modern drugs are derived from natural sources (Jassim and Naji 2003; Gautam et al. 2007; Harvey 2008; Newman and Cragg 2012; Atanasov et al. 2015). The bioactive molecules occur in plants as secondary metabolites and as defense mechanisms against predation, herbivores, fungal attack, microbial invasion, and viral infection. During the past three decades, potent agents have become available against viral infections, and extensive studies have shown that medicinal plants of several parts of the world contain compounds active against viruses that cause human diseases (Semple et al. 1998; Kott et al. 1999; Sindambiwe et al. 1999; Akram et al. 2018). Not to be left out of these are lichens which are also known as promising sources of diverse secondary metabolites, the majority of which exhibit various biological properties including extensive antiviral properties (Pengsuparp et al. 1995; Cohen et al. 1996; Neamati et al. 1997; Esimone et al. 2007; Stocker-Worgotter 2008; Molnar and Farkas 2010; Lai et al. 2013; Basile et al. 2015; Paluszczak et al. 2018).
7.1.2 Lichens
A lichen is a stable, ecologically obligate, self-supporting mutualism between an exhabitant fungus and one or more inhabitant, extracellularly located unicellular or filamentous photoautotrophic partners (Farrar 1976; Hawksworth and Honegger 1994). Thus, lichens are symbiotic organisms of fungi (mycobiont) and algae or cyanobacteria (photobiont) (Voss et al. 1983). Lichen thalli are complex ecosystems rather than organisms (Farrar 1976; Lumbsch 1998). Lichens form the largest mutualistic group among fungi especially given that about 21% of all fungi are able to act as a mycobiont (Honegger 1991). It is estimated that about 40 fungi genera are involved as photobiont in lichen formation (Kirk et al. 2008). Lichens can be found in a wide range of habitats ranging from aquatic to terrestrial terrain (Brightman and Seaward 1977; Seaward 2008; Lisicka 2008; Nash 2008). They are well distributed all over the world and are able to survive in extreme environmental conditions (Nash 2008).
Lichens are known to synthesize a great variety of secondary metabolites, many of which are unique.
7.2 Lichen Secondary Metabolites
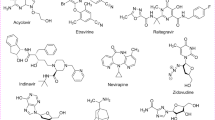
Lichens are known to produce tremendous amounts of chemically diverse group of relatively low molecular weight secondary metabolites, ranging from aliphatic compounds like caperatic acid to aromatic like lobaric acid (Turk et al. 2003). Many examples of lichen secondary metabolites (LSM) abound in literature; a few are shown in Fig. 7.1. For instance, ramalin (γ-glutamyl-N′-(2-hydroxyphenyl) hydrazide) is a secondary metabolite isolated from Ramalina terebrata (Suh et al. 2017; Paudel et al. 2011). Other examples include norstictic acid, a depsidone derivative which has been isolated from different lichen genera including Usnea and Ramalina (Ebrahim et al. 2016). Atranorin is another LSM, a derivative of β-orcinol which is widely distributed in many lichen families (Studzinska-Sroka et al. 2017). Physodic acid from Hypogymnia physodes, squamatic acid from Cladonia uncialis, and salazinic acid and caperatic acid from Platismatia glauca (Paluszczak et al. 2018) are all examples of LSM. Furthermore, physciosporin from Pseudocyphellaria coriacea (Yang et al. 2015), lobaric acid and lobarstin from Stereocaulon alpnum (Hong et al. 2018), depsides from Stereocaulon evolutum (Vu et al. 2015), and usimines A–C, usnic acid derivatives, and stereocalpin A (Seo et al. 2008a, b) from Stereocaulon alpnum are other examples of secondary metabolites isolated from lichens.

Structures of some LSM showing both long aliphatic and aromatic compounds
Lichen has three major pathways that yield secondary metabolites. They include the shikimic pathway, the mevalonic acid pathway, and acetate-polymalonate pathway. The shikimic pathway yields pulvinic acid derivatives, while the mevalonic pathway yields the terpenoids and sterols. The acetate-polymevalonate yields the depside, depsidones, depsipeptides, depsone, etc. Most of the compounds mentioned in this subsection are products of acetate-polymalonate pathway.
7.2.1 Some Biological Activities of Lichen-Derived Secondary Metabolites
LSM have been used in lichen chemotaxonomy (Xu et al. 2016) and systematics (Molnar and Farkas 2010). They have also been studied for bioactivity. Lichen secondary metabolites have been reported to have several bioactive properties that have been demonstrated in the laboratories, including antitumor, antibacterial, anti-herbivore, antioxidant, and antiviral actions. Usnic acid which is wildly available from the Usnea genera has been reported to decrease proliferation of human breast cancer cells and human lung cancer cells without any DNA damage (Mayer et al. 2005). Many of the secondary metabolites from the acetate-polymalonate pathway have been known for different antitumor properties. For instance, ramalin from Ramalina terebrata has strong multiple point inhibition of colorectal cancer replication, migration, invasion, and proliferation (Suh et al. 2017). Norstictic acid inhibited the proliferation, migration, and invasion of breast cancer cells in vitro as well as in vivo invasion growth (Ebrahim et al. 2016). Caperatic acid and physodic acid have also been reported to potently inhibit Wnt signaling in colorectal cancer (Paluszczak et al. 2018). Wnt signaling is a major aberration associated with colorectal cancer cell survival, proliferation, and migration. Kwon et al. (2016) recently reported the inhibition of VCAM-1 expression in TNF-α-stimulated vascular smooth muscle cells via modulation of NF-kB and MAPK signaling pathways by lobaric acid. VCAM-1 expression is important in the development of atherosclerosis making lobaric a potential agent in the treatment inflammatory diseases. Physciosporin, a lichen secondary metabolite, has been reported to inhibit cell motility in lung cancer models A549, H1650, and H1975 (Yang et al. 2015). Atranorin also inhibits cell motility and tumorigenesis in cancer cell model (Zhou et al. 2017; Tian et al. 2017). The depside sphaerophorin isolated from Sphaerophorus globosus and the depsidone pannarin isolated from Psoroma pholidotoides, P. pulchrum, and P. pallidum also displayed anticancer activity against the M14 human melanoma cells (Russo et al. 2008).
It was reported by Bezivin et al. (2004) that (−)-usnic acid, the enantiomer of usnic acid isolated from Cladonia convoluta, showed obvious cytotoxic activities against cancer cell lines, such as murine Lewis lung carcinoma, human chronic myelogenous leukemia, human brain metastasis of a prostate carcinoma, human breast adenocarcinoma, and human glioblastoma, while its enantiomer (+)-usnic acid was reported to inhibit non-small cancer cell motility (Yang et al. 2015). Strong anti-larvicidal activity against the third and fourth instar larvae of the house mosquito (Culex pipiens) and against the polyphagous larvae of the herbivorous insect Spodoptera littoralis were observed with the two enantiomers of usnic acid and vipiric acids (Cetin et al. 2008; Emmerich et al. 1993).
Lichen have also been found to contain a variety of secondary lichen substances with strong antioxidant activity scavenging toxic free radicals (Jha et al. 2017; Thadhani and Karunaratne 2017; Kumar et al. 2014). Vulpinic acid, usnic acid, and pinastric acids from lichen species were reported to have UV blocking and thus photo-protecting properties (Legouin et al. 2017). The common existence of phenolic groups among lichens could account for this. The depsides atranorin isolated from Placopsis sp. and divaricatic acid isolated from Protousnea malacea and the depsidones pannarin isolated from Psoroma pallidum and 10-chloropannarin isolated from Erioderma chilense displayed some antioxidant properties (Hidalgo et al. 1994). Similarly, both sphaerophorin and pannarin (a depsidone) inhibited superoxide anion formation in vitro (Russo et al. 2008).
Lichen is also known to provide sources of metabolites with notable antimicrobial properties. Good antimicrobial properties against a wide range of microbes by parietin isolated from Xanthoria parietina have been reported (Basile et al. 2015). Also, isodivaricatic acid, divaricatinic acid, and usnic acid isolated from Protousnea poeppigii were also reported to have displayed antifungal action against Microsporum gypseum, Trichophyton mentagrophytes, and T. rubrum (Schmeda-Hirschmann et al. 2008). Moderate to strong antifungal action against several fungal species was displayed by extracts from Alectoria sarmentosa and Cladonia rangiferina (Ranković and Mišić 2007) and Evernia prunastri and Hypogymnia physodes (Halama and Van Haluwin 2004).
Additionally, it was reported (Ranković and Mišić 2008; Ranković et al. 2008) that physodic acid from Hypogymnia physodes, stictic acid from Xanthoparmelia conspersa, protocetraric acid from Flavoparmelia caperata, atranorin sourced from Physcia aipolia, fumarprotocetraric acid isolated from Cladonia furcata, 140 gyrophoric acid from Umbilicaria polyphylla, lecanoric acid from Ochrolechia androgyna, and usnic acid from Flavoparmelia caperata all showed relatively strong antimicrobial effects against some bacteria and fungi pathogens.
7.2.2 Antiviral Activities of Lichen Secondary Metabolites
Lichens have proven a useful source of antiviral agents. Investigation of the antiviral properties of atranorin showed that it exhibited strong inhibitory properties against hepatitis C virus (Vu et al. 2015). Usnic acid enantiomers and derivatives showed significant antiviral properties against influenza virus in in vitro studies (Sokolov et al. 2012). Established antiviral activities have been attributed to some other lichen secondary metabolites including parietin, anthraquinones, hypericin, lichenan, etc. For instance, lichenan, which is widely distributed in lichens, is demonstrated to inhibit the tobacco mosaic virus (Stubler and Buchenauer 1996). Similarly, some workers (Cohen and Towers 1995a, b; Cohen et al. 1996) reported the antiviral activity of some naturally occurring anthraquinones, bianthrones, and hypericin derivatives isolated from the lichens Nephroma laevigatum and Heterodermia obscurata. They particularly reported their activities against the virus herpes simplex virus type 1 (HSV-1) where 6 out of 13 compounds at not more than 2 μg/ml could completely inactivate HSV-1 in an endpoint cytopathic effect (CPE) assay. The compounds include emodin, 7-chloroemodin, and 7-chloro-1-O-methylemodin, and 5,7-dichloroemodin being the most active anthraquinone (0.25 μg/ml). The hypericin derivatives 7,70 dichlorohypericin and hypericin equally showed comparable inhibitory activity against HSV-1 with complete inactivation at less than 0.06 μg/ml. Thus, the potential of these compounds as possible tools for further development against HSV-1 is not in question. Moreover, these metabolites if tested against a wider array of viruses may be quite effective as is reflected in the study by Andersen et al. (1991) who showed that hypericin and emodin isolated from Hypericum perforatum were inhibitory against vesicular stomatitis virus (VSV), HSV-2, parainfluenza virus, and vaccinia virus. It is therefore potentially useful to promote a more broad-based extensive screening of potential antiviral metabolites against a wide array of viruses.
In a screening exercise involving a total of 69 lichen species from New Zealand by Perry et al. (1999), they identified the three species Cladia retipora, Pseudocyphellaria glabra, and P. homoeophylla with antiviral activity. These antiviral activities were later traced to the usnic acid constituent following a bioactivity-guided isolation process. Viruses strongly inhibited include the herpes simplex virus type 1 (HSV-1) and the polio type 1 virus. It is also well known that usnic acid has multifaceted biological activities. So its antiviral activity equally does not come as a surprise.
In another related study, investigation of some Icelandic lichens resulted in the identification of two antiviral compounds active against three viruses: respiratory syncytial virus RSV, HSV-1, and HSV-2. Activity against HSV 1 was less potent than activity against RSV. The compounds were the depsidone salazinic acid (IC50 11.9 μg/ml) from Parmelia saxatilis (L.) Ach. and the benzyl depside alectorialic acid (IC50 17.0 μg/ml) from Alectoria nigricans (Ach.) (Omarsdottir et al. 2006). In the year following, Fazio et al. (2007) reported their findings where they described the antiviral activity of parietin isolated from Teloschistes chrysophthalmus against some Junin and Tacaribe arenaviruses.
Extracts of the lichen species Xanthoria parietina and Xanthoparmelia tinctina were evaluated and shown to be active against human parainfluenza virus type 2 (HPIV-2) (Karagöz and Aslan 2005). Other investigations have reported antiviral activities of lichen extracts against the human cytomegalovirus (Wood et al. 1990), HIV (Hirabayashi et al. 1989), and Epstein-Barr virus (Yamamoto et al. 1995).
The search for lichen metabolites with antiviral properties continued with some investigations from African lichens. In some studies involving lichen species from West Africa, Esimone et al. (2007) tested some extracts of the lichen Parmelia perlata against some RNA viruses: yellow fever, infectious bursal disease virus, and polio virus. While the water and acetone extracts showed no activity against the viruses when tested at concentrations below the cytotoxic level, the crude polysaccharide fraction on the other hand showed activity against yellow fever virus with an IC50 of 15 μg/ml. Following the outcome of further investigation for mechanism of action profile, they proposed that the target of the inhibitory activity is possibly mediated against the envelope of the yellow fever virus. Furthermore, Esimone and coworkers screened derivatives from the lichen Ramalina farinacea against lentiviruses and adenoviruses. Their reported work (Esimone et al. 2005) initially showed that the ethyl acetate-soluble fraction (ET4) from R. farinacea inhibited the infectivity of lentiviral and adenoviral vectors, as well as HIV-1. Recorded antiviral activity was about 20 μg/ml. Preliminary mechanistic studies based on the addition of the extracts at different time points in the viral infection cycle (kinetic studies) led to the suggestion that early steps in the lentiviral or adenoviral replication cycle could be the major target of ET4.
Later, in another related study (Esimone et al. 2009), they determined the antiviral activity of ET4 against other wild-type viruses, including the herpes simplex virus type1 (HSV-1) and the respiratory syncytial virus (RSV). They found ET4 to inhibit HSV-1 and RSV potently with IC50 of 6.09 and 3.65 μg/ml, respectively. Attempt to elucidate the possible mechanism of action of the ET4 revealed through the time-of-addition studies suggests that both entry and post entry steps of the HIV-1 replication cycle and the entry step of the RSV replication cycle are targeted, respectively. Thus, ET4 is expected to inhibit HIV-1 at some point around the viral envelope, while RSV is inhibited downstream beyond the viral envelope. Additionally, they reported that ET4 inhibited HIV-1 reverse transcriptase with an IC50 of 0.022 μg/ml. Further activity-guided fractionation of ET4 yielded two subfractions rfO and rfM with varied activities. While rfM demonstrated activity against HSV-1 (DNA virus) but not against the lentiviral vector, rfO was active against lentiviral vector and HIV-1 (RNA viruses) but not against HSV-1.
In a follow-up continuity study by Lai et al. (2013), further investigation involving the ethyl acetate-soluble fraction (ET4) of R. farinacea led to the isolation of 13 phenolic compounds including 1 new hydroquinone depside (5-hydroxysekikaic acid) and 1 new orsellinic acid derivative (2,3-dihydroxy-4-methoxy-6-pentylbenzoic acid). All 13 compounds displayed varying degrees of antiviral activity against the respiratory syncytial virus (RSV) with one of them sekikaic acid showing the most potent inhibition toward a recombinant strain of the respiratory syncytial virus rgRSV (IC50 5.69 μg/ml) and the clinical RSV A2 strain (IC50 7.73 μg/ml). The time-of-addition assay for mechanistic elucidation revealed that sekikaic acid clearly interferes with viral replication at a viral post-entry step.
7.2.3 Experimental Methods in the Discovery of Antiviral Lichen Compounds
Viral infection could be evaluated by direct cytological and histological examination under the light microscope for signs of viral-induced cytopathic effect (CPE) (Esimone et al. 2008; Lai et al. 2013), through antigen detection by immunological and serological assays (Odimegwu 2018), immunohistochemistry (Lai et al. 2013), immunofluorescence (Sloan et al. 2015), viral proteomics (Tavalai et al. 2008; Schreiner et al. 2010), and viral genome detection by hybridization with specific nucleic acid probe polymerase chain reaction (PCR) (Ternette et al. 2007). They could also be further indirectly detected by hemabsorption (Drake et al. 2016), hemagglutination (Motohashi et al. 2013; Li et al. 2015), and, in the case of egg viral cultivation pocks on chorioallantoic membrane (CAM) (Wang et al. 2011), inclusion bodies (Sloan et al. 2015). Enzyme-linked immunosorbent assay (ELISA) (Ternette et al. 2007), complement fixation tests (CFT) (Ronalds et al. 1983), protein immunoblot (Western blot) (Kohlmann et al. 2009), and other serological assays are equally useful toward viral detection. Moreover, disease or death could also confirm viral infection especially in animal models. Additionally, there are various tools and vehicles for antiviral screening.
Cell Lines and Medium
Antiviral screening is largely done by in vitro methods requiring the use of cell lines of mammalian origin. Therefore, cell lines utilized are greatly widespread and commonly include Vero (a monkey kidney-derived primary cell line), Hep2 (of human airway epithelial lineage), Hela (human cervical carcinoma cell line), human tsA201 cell (a derivative of human embryonic kidney cell line 293), 293A cell line (a permanent line of primary human embryonal kidney transformed by sheared human adenovirus type 5 (Ad 5) DNA), and TZM-bl cell (a transformed cell line of cervical cancer origin expressing the classical HIV-1 entry receptors). Madin-Darby canine kidney (MDCK) cells and several other cell lines may be utilized in various antiviral screenings. These cells may for instance usually be grown in Dulbecco’s Modified Eagle Medium (DMEM) consisting of high glucose and 2 mM L-glutamine and supplemented with heat-inactivated fetal calf serum and a mixture of penicillin and streptomycin (Pen-Strep) (Ternette et al. 2007; Esimone et al. 2008; Odimegwu et al. 2018). There are other media which are enriched to accommodate additional requirements of certain specialized cells utilized in the antiviral screening.
Viral Particles and Vectors
Viruses are usually utilized in these screenings since the effects of antiviral compounds of lichen origin must be assayed in the presence of the target virus. In this case, the virus could be utilized live and unaltered (e.g., RSV), or the virus is slightly genetically modified without necessarily altering its infectivity while imparting maybe some sort of regulatory or reporter protein property (an example is the rgRSV expressing the green fluorescent protein) (Ternette et al. 2007; Lai et al. 2013). In some cases, viruses can be wholly redesigned to produce a viral particle with clearly transformed property referred to as viral vectors. Viral vectors are designed to act as the virus and to effectively deliver the viral genome into susceptible host cell. They can be genetically engineered to either delete unwanted genes/antigen/structures or to introduce new foreign genes/antigen/structures from another virus to create pseudo-typed viruses (vectors) (Esimone et al. 2005, 2009). They find great utility in the evaluation of antiviral lichen compounds.
Antiviral Assays
A variety of assay types exist to enable the effective evaluation of antiviral agents. The preliminary stage of screening of novel antiviral compound from lichen may begin with an initial simple screening of crude or broad-based fraction of the lichen containing the supposed antiviral substance. Screening is completed through utilization of various bioassays presented sometimes in the format of a bioassay-guided fractionation process.
Bioassay-Guided Fractionation
Bioassay-guided isolation involves the use of some well-defined protocol for the isolation of pure compounds with defined biological activity from a crude extract of a natural product which could be plant or animal parts or a cocktail of mixture (Esimone et al. 2008). It involves step-by-step separation of fractions/constituents contained within the crude extracts or mixture based on their physicochemical activities and through assaying for biological activity. Several rounds of separation of fractions/constituents and bioassay may be required to finally identify pure molecular hit compound(s).
Antiviral Cytopathic and Plaque Reduction Assays
Screening of novel antiviral constituent requires the presence of the screened substance in flask or system containing appropriate mammalian cells either adhered or in suspension within cell culture medium including the investigated lichen antiviral substance (Cohen et al. 1996; Kott et al. 1999; Esimone et al. 2009; Lai et al. 2013). Antibiotics are normally included in these media to prevent contamination. Normally, routine screening could take between 24 h and several days following which plaques representing regions of virus activity/replication are counted and reported relative to control setup excluding the antiviral substance. Viral-induced cytopathy or plaque formation may be observed directly under the light microscope or may be done following specific/immune staining of responsible viral protein present in the infected cell (Kott et al. 1999; Esimone et al. 2008). Where a fluorescent protein has been introduced into virus or vector, then observation could also be analyzed using fluorescent microscopy (Lai et al. 2013).
Additional antiviral assays to further define certain specific properties of the identified compound could still be analyzed employing the viral plaque reduction assay, for instance, assays investigating the mechanism of antiviral activity in a broader sense (e.g., time point of optimal antiviral effect during viral infectivity cycle), interference of compound with some measurable viral or host cell enzymes, and protein systems.
Viral Cytopathic Effect Measurement
As with viral plaque reduction assays above, a generalized viral cytopathic effect reduction by investigated antiviral lichen compound could be carried out to characterize the antiviral activity of the substance. The experimental setup remains essentially the same as above, but measurement of antiviral activity may be done by analyzing for the presence of cell pathology-indicating enzymes in cell culture supernatant or the measurement for any reduction in substance-treated cell oxidative activity for sign of cell pathology (Semple et al. 1998; Odimegwu 2018).
Further Antiviral Assays to Discover Antiviral Lichen Compounds
Additional antiviral assays to further define certain specific properties of the identified compound could still be analyzed employing the viral plaque reduction assay, for instance, assays investigating the mechanism of antiviral activity in a broader sense (e.g., time point of optimal antiviral effect during viral infectivity cycle) (Stubler and Buchenauer 1996; Lai et al. 2013) and interference of compound with some measurable viral or host cell enzymes and protein systems (Odimegwu 2018). In existence are other virological and immune-based assays that have been reportedly employed for the antiviral screening of lichen-derived compounds, which include virucidal effect (Andersen et al. 1991; Lai et al. 2013), protein immunoblot assay/Western blot analysis (Neamati et al. 1997), enzyme-linked immunosorbent assay (ELISA) (Semple et al. 1998), fluorescence-activated cell sorting assays (Odimegwu 2018), DNA and RNA polymerase inhibition assays (Pengsuparp et al. 1995), and some other antiviral assays. Clearly, antiviral assays could always be developed to fill the gap for any specific analysis of the candidate lichen-derived constituents/compounds.
7.3 Conclusion
Secondary metabolites of lichens with virus-inhibiting effects as explained so far (summarized in Table 7.1) constitute potent lead compounds for the development of novel antiviral agents. Thus, secondary metabolites of lichens represent a promising source of novel bioactive lead compounds for the medical and pharmaceutical industry in particular. Though many lichen compounds have been well characterized and proved to be efficacious against certain viruses, the potential of discovering more hitherto unknown antiviral compounds of lichen origin remains very promising.
Indeed, lichens have proven to be useful source of antiviral agents. There seems to have been a recent upsurge in the recognition of the potential of lichens as useful source of secondary metabolites with varied biological activities. And this is partly due to the increasingly fairly well-documented biological and antiviral properties of lichen secondary metabolites. And the fact that new compounds continue to be isolated from lichens makes them an attractive potential source of novel biologicals and antivirals of the future. Another obvious advantage is the relative ease of their growth and survival given the right environment; therefore, special biotechnological platform can be designed for their optimal cultivation and exploitation.
Although the rate of reportage of antiviral agents of lichen origin is low, it is equally important to understand perhaps why this is so. Perhaps, antiviral evaluation is not being pursued by many investigations on lichens, and where is it is carried out the virus focus is usually too narrowed on one a few viruses. Nevertheless, as currently available reports and data show, lichens widely exist waiting to be discovered and investigated for possible antiviral potential. So what is required is the need to speed up extensive screening of potential lichen candidates/metabolites. Screenings should equally be made broad-based to discover even the remotest virus that could be covered by any one single metabolite. Given the varied opportunities offered by lichens, it is expected that they would thus remain an interesting focus of our searchlight in the pursuit of effective antiviral remedies against the hordes of viral infections and diseases facing humanity.
References
Akram M, Tahir IM, Shah SMA, Mahmood Z, Altaf A, Ahmad K, Munir N, Daniyal M, Nasir S, Mehboob H (2018) Antiviral potential of medicinal plants against HIV, HSV, influenza, hepatitis, and coxsackievirus: a systematic review. Phytother Res 32(5):811–822
Andersen DO, Weber ND, Wood SG, Hughes BG, Murray BK, North JA (1991) In vitro virucidal activity of selected anthraquinones and anthraquinone derivatives. Antivir Res 16:185–196
Atanasov AG, Waltenberger B, Pferschy Wenzig EM, Linder T, Wawrosch C, Uhrin P, Temml V, Wang L, Schwaiger S, Heiss EH (2015) Discovery and resupply of pharmacologically active plant-derived natural products: a review. Biotechnol Adv 33:1582–1614
Basile A, Rigano D, Loppi S et al (2015) Antiproliferative, antibacterial and antifungal activity of the lichen Xanthoria parietina and its secondary metabolite parietin. Int J Mol Sci 16:7861–7875
Bezivin C, Tomasi S, Rouaud I, Delcros J, Boustie J (2004) Cytotoxic activity of compounds from the lichen: Cladonia convoluta. Planta Med 70:874–877
Brightman FH, Seaward MRD (1977) Lichens of man-made substrates. In: Seaward MRD (ed) Lichen ecology. Academic Press, London, pp 253–293
Cetin H, Tufan-Cetin O, Turk AO, Tay T, Candan M, Yanikoglu A, Sumbul H (2008) Insecticidal activity of major lichen compounds, (−)- and (+)-usnic acid, against the larvae of house mosquito, Culex pipiens L. Parasitol Res 102:1277–1279
Cohen PA, Towers GHN (1995a) Anthraquinones and phenanthroperylenequinones from Nephroma laevigatum. J Nat Prod 58:520
Cohen PA, Towers GHN (1995b) The anthraquinones of Heterodermia obscurata. Phytochemistry 40:911
Cohen PA, Hudson JB, Towers GHN (1996) Antiviral activities of anthraquinones, bianthrones and hypericin derivatives from lichens. Experientia 52:180–183
Drake MG, Bivins-Smith ER, Proskocil BJ et al (2016) Human and mouse eosinophils have antiviral activity against parainfluenza virus. Am J Respir Cell Mol Biol 55(3):387–394
Ebrahim HY, Elsayed HE, Mohyeldin MM et al (2016) Norstictic acid inhibits breast cancer cell proliferation, migration, invasion, and in vivo invasive growth through targeting C-met. Phytother Res 30:557–566
Emmerich R, Giez I, Lange OL, Proksch P (1993) Toxicity and antifeedant activity of lichen compounds against the polyphagous herbivorous insect Spodoptera littoralis. Phytochemistry 33:1389–1394
Esimone CO, Grunwald T, Wildner O, Nchinda G, Tippler B, Proksch P, Uberla K (2005) In vitro pharmacodynamic evaluation of antiviral medicinal plants using a vector-based assay technique. J Appl Microbiol 99(6):1346–1355
Esimone CO, Ofokansi KC, Adikwu MU, Ibezim EC, Abonyi DO, Odaibo GN, Olaleye DO (2007) In vitro evaluation of the antiviral activity of extracts from the lichen Parmelia perlata (L.) Ach. against three RNA viruses. J Infect Dev Ctries 1(3):315–320
Esimone CO, Eck G, Duong TN, Uberla K, Proksch P, Grunwald T (2008) Potential anti-respiratory syncytial virus lead compounds from Aglaia species. Pharmazie 63:1–6
Esimone CO, Grunwald T, Nworu CS, Kuate S, Proksch P, Uberla K (2009) Broad spectrum anti-viral fractions from the lichen Ramalina farinacea (L.) Ach. Chemotherapy 55:119–126
Farrar JF (1976) The lichen as an ecosystem: observation and experiment. In: Brown DH, Hawksworth DL, Bailey RH (eds) Lichenology: progress and problems, Special volume – The Systematics Association, vol 8. Academic Press, London, pp 385–406
Fazio AT, Adler MT, Bertoni MD, Sepulveda CS, Damonte EB, Maier MS (2007) Lichen secondary metabolites from the cultured lichen mycobionts of Teloschistes chrysophthalmus and Ramalina celastri and their antiviral activities. Z Naturforsch 62:543–549
Gautam R, Saklani A, Jachak SM (2007) Indian medicinal plants as a source of antimycobacterial agents. J Ethnopharmacol 110:200–234
Halama P, Van Haluwin C (2004) Antifungal activity of lichen extracts and lichenic acids. BioControl 49:95–107
Harvey AL (2008) Natural products in drug discovery. Drug Discov Today 13:894–901
Hawksworth DL, Honegger R (1994) The lichen thallus: a symbiotic phenotype of nutritionally specialized fungi and its response to gall producers. In: Williams MAJ (ed) Plant galls, Special volume – The Systematics Association, vol 49. Clarendon Press, Oxford, pp 77–98
Hidalgo ME, Fernandez E, Quilhot W, Lissi E (1994) Antioxidant activity of depsides and depsidones. Phytochemistry 37:1585–1587
Hirabayashi K, Iwata S, Ito M, Shigeta S, Narui T, Mori T, Shibata S (1989) Inhibitory effect of a lichen polysaccharide sulfate, GE-3-S, on the replication of human immunodeficiency virus (HIV) in vitro. Chem Pharm Bull 37:2410–2412
Honegger R (1991) Functional aspects of the lichen symbioses. Annu Rev Plant Physiol Plant Mol Biol 42:553–578
Hong JM, Suh SS, Kim TK et al (2018) Anti-cancer activity of lobaric acid and lobarstin extracted from the Antarctic lichen Stereocaulon alpnum. Molecules 23:658. https://doi.org/10.3390/molecules23030658. eCollection 2018 Mar
Jassim SAA, Naji MA (2003) Novel anti-viral agents: a medicinal plant perspective. J Appl Microbiol 95:412–427
Jha BN, Shrestha M, Pandey DP et al (2017) Investigation of antioxidant, antimicrobial and toxicity activities of lichens from high altitude regions of Nepal. BMC Complement Altern Med 17:282. eCollection 2017
Karagöz A, Aslan A (2005) Antiviral and cytotoxic activity of some lichen extracts. Biol Bratis 60:281–286
Kirk PM, Cannon PF, Minter DW, Stalpers JA (eds) (2008) Dictionary of the fungi, 10th edn. CAB International, Wallingford
Kohlmann R, Schwannecke S, Tippler B, Ternette N, Temchura V, Tenbusch M, Überla K, Grunwald T (2009) Protective efficacy and immunogenicity of an adenoviral vector vaccine encoding the codon-optimized F protein of respiratory syncytial virus. J Virol 83(23):12601–12610
Kott V, Barbini L, Cruanes M et al (1999) Anti-viral activity in Argentine medicinal plants. J Ethnopharmacol 64:79–84
Kumar J, Dhar P, Tayade AB et al (2014) Antioxidant capacities, phenolic profile and cytotoxic effects of saxicolous lichens from trans-Himalayan cold desert of Ladakh. PLoS One 9:e98696
Kwon IS, Yim JH, Lee HK et al (2016) Lobaric acid inhibits VCAM-1 expression in TNF-α-stimulated vascular smooth muscle cells via modulation of NF-ΰB and MAPK signaling pathways. Biomol Ther (Seoul) 24:25–32
Lai D, Odimegwu DC, Esimone C, Grunwald T, Proksch P (2013) Phenolic compounds with in vitro activity against respiratory syncytial virus from the Nigerian lichen Ramalina farinacea. Planta Med 79(15):1440–1446
Legouin B, Lohézic-Le Dévéhat F, Ferron S et al (2017) Specialized metabolites of the lichen Vulpicida pinastri act as photoprotective agents. Molecules 22:1162. https://doi.org/10.3390/molecules22071162. eCollection 2017 Jul
Li F, Ma C, Wang J (2015) Inhibitors targeting the influenza virus hemagglutinin. Curr Med Chem 22:1361–1382
Lisicka E (2008) Lichens on an acrylic-coated aluminium roof. Graph Scr 20:9–12
Lumbsch HT (1998) Taxonomic use of metabolic data in lichen-forming fungi. In: Frisvad JC, Bridge PD, Arora DK (eds) Chemical fungal taxonomy. Marcel Dekker, New York, pp 345–387
Mayer M, O’Neill MA, Murry KE, Santos-Magalhaes NS, Carneiro-Leao AMA, Thompson AM, Appleyard VCL (2005) Usnic acid: a non-genotoxic compound with anti-cancer properties. Anti-Cancer Drugs 16:805–809
Molnar K, Farkas E (2010) Current results on biological activities of lichen secondary metabolites: a review. Z Naturforsch 65c:157–173
Motohashi Y, Igarashi M, Okamatsu M et al (2013) Antiviral activity of stachyflin on influenza A viruses of different hemagglutinin subtypes. Virol J 10:118. https://doi.org/10.1186/1743-422X-10-118
Nash TH III (ed) (2008) Lichen biology, 2nd edn. Cambridge University Press, Cambridge
Neamati N, Hong H, Mazumder A, Wang S, Sunder S, Nicklaus MC, Milne GWA, Proksa B, Pommier Y (1997) Depsides and depsidones as inhibitors of HIV-1 integrase: discovery of novel inhibitors through 3D database searching. J Med Chem 40:942–951
Newman DJ, Cragg GM (2012) Natural products as sources of new drugs over the 30 years from 1981 to 2010. J Nat Prod 75:311–335
Odimegwu DC (2018) Low-dose Sekikaic acid modulates host immunity and protects cells from Respiratory Syncytial Virus infection. Biotechnol J Int 21(2):1–10
Odimegwu DC, Okoye FBC, Nworu CS, Esimone CO (2018) Anti-respiratory syncytial virus activities of leaf extracts of Alchornea cordifolia and Alchornea floribunda. Afr J Pharm Pharmacol 12(8):97–105
Omarsdottir S, Óladóttir AK, Árnadóttir T, Ingólfsdóttir K (2006) Antiviral compounds from Icelandic lichens. Planta Med 72. https://doi.org/10.1055/s-2006-949742
Paluszczak J, Kleszcz R, Studzińska-Sroka E et al (2018) Lichen-derived caperatic acid and physodic acid inhibit Wnt signaling in colorectal cancer cells. Mol Cell Biochem 441:109–124
Paudel B, Bhattarai HD, Koh HY et al (2011) Ramalin, a novel nontoxic antioxidant compound from the Antarctic lichen Ramalina terebrata. Phytomedicine 18:1285–1290
Pengsuparp T, Cai L, Constant H, Fong HHS, Lin LZ, Kinghorn AD, Pezzuto JM, Cordell GA, Ingolfsdöttir K, Wagner H, Hughes SH (1995) Mechanistic evaluation of new plant-derived compounds that inhibit HIV-1 reverse transcriptase. J Nat Prod 58:1024–1031
Perry NB, Benn MH, Brennan NJ, Burgess EJ, Ellis G, Galloway DJ, Lorimer SD, Tangney RS (1999) Antimicrobial, antiviral and cytotoxic activity of New Zealand lichens. Lichenologist 31(6):627–636
Ranković B, Mišić M (2007) Antifungal activity of extracts of the lichens Alectoria sarmentosa and Cladonia rangiferina. Mikol Fitopatol 41:276–281
Ranković B, Mišić M (2008) The antimicrobial activity of the lichen substances of the lichens Cladonia furcata, Ochrolechia androgyna, Parmelia caperata and Parmelia conspersa. Biotechnol Biotechnol Equip 22:1013–1016
Ranković B, Mišić M, Sukdolak S (2008) The antimicrobial activity of substances derived from the lichens Physcia aipolia, Umbilicaria polyphylla, Parmelia caperata and Hypogymnia physodes. World J Microbiol Biotechnol 24:1239–1242
Ronalds CJ, Hardiman AE, Griffiths PD (1983) Hotting up the complement fixation test. J Hyg (Lond) 90:127–134
Russo A, Piovano M, Lombardo L, Garbarino J, Cardile V (2008) Lichen metabolites prevent UV light and nitric oxide-mediated plasmid DNA damage and induce apoptosis in human melanoma cells. Life Sci 83:468–474
Schmeda-Hirschmann G, Tapia A, Lima B et al (2008) A new antifungal and antiprotozoal depside from the Andean lichen Protousnea poeppigii. Phytother Res 22:349–355
Schreiner S, Wimmer P, Sirma H, Everett RD, Blanchette P, Groitl P, Dobner T (2010) Proteosome-dependent degradation of Daxx by the Viral E1B-55K protein in human adenovirus-infected cells. J Virol 84(14):7029–7038
Seaward MRD (2008) Environmental role of lichens. In: Nash TH III (ed) Lichen biology, 2nd edn. Cambridge University Press, Cambridge, pp 274–298
Semple SJ, Reynolds GD, O’Leary GD, Flower RLP (1998) Screening of Australian medicinal plants for anti-viral activity. J Ethnopharmacol 60:163–172
Seo C, Sohn JH, Park SM et al (2008a) Usimines A-C, bioactive usnic acid derivatives from the Antarctic lichen Stereocaulon alpinum. J Nat Prod 71:710–712
Seo C, Yim JH, Lee HK et al (2008b) Stereocalpin A, a bioactive cyclic depsipeptide from the Antarctic lichen Stereocaulon alpinum. ChemInform 49:29–31
Sindambiwe JB, Calomme M, Cos P et al (1999) Screening of seven selected Rwandan medicinale plants for antimicrobial and anti-viral activities. J Ethnopharmacol 65:71–77
Sloan E, Tatham MH, Groslambert M, Glass M, Orr A, Hay RT, Everett RD (2015) Analysis of the SUMO2 Proteome during HSV-1 Infection. PLoS Pathog 11(7):e1005059. https://doi.org/10.1371/journal.ppat.1005059
Sokolov DN, Zarubaev VV, Shtro AA et al (2012) Anti-viral activity of (−)- and (+)-usnic acids and their derivatives against influenza virus A(H1N1)2009. Bioorg Med Chem Lett 22:7060–7064
Stocker-Worgotter E (2008) Metabolic diversity of lichen-forming ascomycetous fungi: culturing, polyketide and shikimate metabolite production, and PKS genes. Nat Prod Rep 25:188–200
Stubler D, Buchenauer H (1996) Antiviral activity of the glucan lichenan (poly-β{1 → 3, 1 →4} Danhydroglucose) 1. Biological activity in tobacco plants. J Phytopathol 144:37–43
Studzinska-Sroka E, Galanty A, Bylka W (2017) Atranorin - an interesting lichen secondary metabolite. Mini Rev Med Chem 17:1633–1645
Suh SS, Kim TK, Kim JE et al (2017) Anticancer activity of ramalin, a secondary metabolite from the antarctic lichen Ramalina terebrata, against colorectal cancer cells. Molecules 22:1361. https://doi.org/10.3390/molecules22081361. eCollection 2017 Aug
Tavalai N, Papior P, Rechter S, Stamminger T (2008) Nuclear domain 10 components promyelocytic leukemia protein and hDaxx independently contribute to an intrinsic antiviral defense against human cytomegalovirus infection. J Virol 82:126–137
Ternette N, Tippler B, Uberla K, Grunwald T (2007) Immunogenicity and efficacy of codon optimized DNA vaccines encoding the F-protein of respiratory syncytial virus. Vaccine 25:7271–7279
Thadhani VM, Karunaratne V (2017) Potential of lichen compounds as antidiabetic agents with antioxidative properties: a review. Oxid Med Cell Longev 2017:2079697
Tian Y, Li YL, Zhao FC (2017) Secondary metabolites from polar organisms. Mar Drugs 15:28. https://doi.org/10.3390/md15030028. eCollection 2017 Mar
Turk AO, Yilmaz M, Kivanc M, Turk H (2003) The antimicrobial activity of extracts of the lichen Cetraria aculeata and its protolichesterinic acid constituent. Z Naturforsch 58c:850–854
Voss EG, Burdet HM, Chaloner WG, Demoulin V, Hiepko P, Mcneill J, Meikle RD, Nicolson DH, Rollins RC, Silva PC, Greuter W (1983) International code of botanical nomenclature (Sydney Code). Regnum Veg 111:1–472
Vu TH, Le Lamer AC, Lalli C et al (2015) Depsides: lichen metabolites active against hepatitis C virus. PLoS One 10:e0120405
Wang X, Chi X, Wang M (2011) Structural characteristics and antiviral activity of multiple peptides derived from MDV glycoproteins B and H. Virol J 8:190
Wood S, Huffman J, Weber N, Andersen D, North J, Murray B, Sidwell R, Hughes B (1990) Antiviral activity of naturally occurring anthraquinones and anthraquinone derivatives. Planta Med 56:65–52
Xu M, Heidmarsson S, Olafsdottir ES et al (2016) Secondary metabolites from cetrarioid lichens: chemotaxonomy, biological activities and pharmaceutical potential. Phytomedicine 23:441–459
Yamamoto Y, Kinoshita Y, Kurokawa T, Yoshimura I, Ahmadjian V, Yamada Y (1995) Cell growth and pigment production in suspension cultures of a mycobiont isolated from the lichen Cladonia cristatella. Can J Bot 73:590–594
Yang Y, Park SY, Nguyen TT et al (2015) Lichen secondary metabolite, physciosporin, inhibits lung cancer cell motility. PLoS One 10:e0137889
Zhou R, Yang Y, Park SY et al (2017) The lichen secondary metabolite atranorin suppresses lung cancer cell motility and tumorigenesis. Sci Rep 7:8136. E Collection 2017
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Odimegwu, D.C., Ngwoke, K., Ejikeugwu, C., Esimone, C.O. (2019). Lichen Secondary Metabolites as Possible Antiviral Agents. In: Ranković, B. (eds) Lichen Secondary Metabolites. Springer, Cham. https://doi.org/10.1007/978-3-030-16814-8_7
Download citation
DOI: https://doi.org/10.1007/978-3-030-16814-8_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-16813-1
Online ISBN: 978-3-030-16814-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)