
Abstract
A scavenger is an animal that feeds on the carcass or remains of any dead animal which it did not participate in its killing. Scavenging is pervasive across the animal kingdom and almost all predator species use carrion to a certain extent in both terrestrial and aquatic ecosystems. There is a group of animals, the obligate scavengers, which rely (almost) entirely on carrion. Among vertebrates, only birds have evolved into obligate scavengers, namely vultures, which suggests that the costs of adaptation to obligate scavenging are high. Obligate and facultative scavengers exhibit a wide array of adaptations to locate and exploit carrion across systems, including inexpensive locomotion to find the unpredictable carrion on savannas, caching carrion in cold tundra or chemotaxis in aquatic systems. Traditionally viewed as an opportunistic process, particularly for facultative scavengers, carrion consumption by vertebrates often follows complex and structured patterns and is crucial in maintaining the stability and structure of food webs.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
- Adaptations
- Aquatic ecosystems
- Facultative scavenging
- Forests
- Nestedness
- Obligate scavenging
- Savannah
- Steppe
- Tundra
Facultative Versus Obligate Scavengers
In 1877, Sanborn Tenney, a professor of natural history at Williams College in Massachusetts, USA, wrote about scavengers as those animals using or removing what others have ignored or discarded (Tenney 1877). Scavenging is likely to have been common since the very first origins of vertebrates. Many theropod dinosaurs, which dominated terrestrial ecosystems for more than 160 million years, likely gained an important part of their energetic requirements by feeding on carrion (Kane et al. 2016). In the Pleistocene, when mammalian megafauna was abundant (Martin 1989; Ruxton and Houston 2003), scavenging was common among carnivores, and even among early hominins, which were better scavengers than hunters (Blumenschine and Cavallo 1992; Moleón et al. 2014a).
Later on, with large carcasses becoming scarcer (Barnosky et al. 2004), the temporal patchiness of carrion availability might have inhibited evolution towards strict specialisation for scavenging in most vertebrates. As a result, obligate scavengers, i.e. species that rely entirely or near entirely on carrion as food resource, are rare. In the recent past, no reptiles or mammals have evolved into obligate scavengers (DeVault et al. 2003). Some benthic scavengers (e.g. hagfish: family Myxinidae) rely widely on carrion and may be considered obligate scavengers (Smith and Baco 2003; Beasley et al. 2015). However, among terrestrial vertebrates, only vultures (families Accipitridae and Cathartidae) have evolved into obligate scavengers (DeVault et al. 2003; Beasley et al. 2015; Moleón et al. 2014b).
Even so, facultative scavenging is fairly common and scavenging activity appears to be pervasive across the animal kingdom (DeVault et al. 2003; Wilson and Wolkovich 2011; Beasley et al. 2015; Mateo-Tomás et al. 2015). Facultative scavengers are animals that scavenge at variable rates but that can subsist on other food resources in the absence of carrion. All mammalian predators (e.g., raccoons Procyon lotor, wolves Canis lupus, bears -family Ursidae-, foxes Vulpes spp., hyenas Hyaena spp., lions Panthera leo), birds of prey (e.g., kites, eagles, buzzards-family Accipitridae) and corvids (e.g., ravens, crows-family Corvidae) are facultative scavengers (DeVault et al. 2003; Selva 2004; Wilson and Wolkovich 2011; Moleón et al. 2014b; Pereira et al. 2014). Some species, typically regarded as strict predators, also scavenge frequently, like the northern goshawk (Accipiter gentilis), the weasel (Mustela nivalis, Selva 2004) and the European otter (Lutra lutra, O’Sullivan et al. 1992). Some omnivores which are not strict predators (e.g. wild boar Sus scrofa) also consume carrion frequently (Selva et al. 2005). Woodpeckers and tits feed on carcasses during winter, and other passerines exploit resources provided by carcasses, like maggots or hairs to build nests (Moreno-Opo and Margalida 2013). An amazing spectrum of animals will consume carrion if the opportunity arises, including the hippopotamus (Hippopotamus amphibius), white-tailed deer (Odocoileus virginianus), and many snake species (DeVault and Krochmal 2002; Beasley et al. 2015). Carrion is by far the most important food for some reptiles (e.g. the lace monitor, Varanus varius, Guarino 2001).
Morphological, Physiological and Behavioral Adaptations to Scavenging
Terrestrial Scavengers
The adaptations shown by vultures includes morphological characters facilitating the dismembering of and access to the interior of large corpses, like long necks, strong beaks and scarce feather coverage on head and neck to prevent fouling (Kruuk 1967; Houston 1975, 1979). Vultures have excellent visual abilities, and some species like vultures from the genus Cathartes also possess a well-developed sense of smell to locate carrion (Houston 1986; Ruxton and Houston 2004). Their large body size allows vultures to cope with periods of food scarcity. In addition, vultures reduce the potential lethal effects of toxins and pathogens due to a number of physiological and immunological adaptations, including an acid digestive pH (Houston and Cooper 1975). Vultures have also developed foraging strategies to exploit randomly distributed resources, i.e. to explore large areas by gliding, and the transmission, active or passive, of information (Houston 1974; Buckley 1996; Ruxton and Houston 2004; Jackson et al. 2008). These adaptations make vultures very efficient scavengers compared to mammalian carnivores, which show a more limited mobility due to territoriality and the higher energetic costs of walking (Houston 1979; Ruxton and Houston 2004; Moleón et al. 2014b). Similar adaptations are shown by avian facultative scavengers. Corvids, and common ravens Corvus corax, in particular, actively recruit each other to carcasses, and share information about carrion location at their communal roosts (Heinrich 1988; Marzluff and Heinrich 1991; Heinrich et al. 1993; Marzluff et al. 1996; Wilmers et al. 2003a; Wright et al. 2003).
A scavenging lifestyle often involves watching and following others, both at the intra- and inter-specific levels. The transfer of social information within the scavenging guild is quite fluid (Moleón et al. 2014b). Avian scavengers may follow mammalian predators to feed on their kills. Vultures have been observed circling over large carnivores before hunting and being attracted to sounds of hyaenas (Attwell 1963; Kruuk 1967, 1972). Ravens associate with wolves (Stahler et al. 2002), polar bears Ursus maritimus (Gilchrist and Robertson 2000), and Siberian tigers Panthera tigris altaica (Matyushkin 1974) to feed on their kills. Ravens also react to gunshots and have learned to exploit gut piles left by hunters (White 2005). Mesopredators also follow large predators to feed on their kills, e.g. red Vulpes vulpes and arctic foxes Vulpes lagopus are reported to follow wolves and polar bears, respectively (Stirling and Archibald 1977; Jędrzejewska and Jędrzejewski 1998). Large scavenging birds, and particularly vultures and eagles, observe the activities of smaller birds, like crows or magpies, which are usually first to discover carcasses (König 1974; Prior and Weatherhead 1991; Vogeley 1999). Flocks of corvids are an unequivocal sign of carrion presence and often attract eagles and other raptors (Knight and Knight 1983; Selva 2004). Raptors often act as producers of information, while vultures are information scroungers (Kane et al. 2014). Griffon vultures Gyps fulvus use socially acquired information and they follow vultures that sink vertically to land to increase carcass location success (Cortés-Avizanda et al. 2014). Mammals also respond to aggregations of scavenging birds; hyenas and jackals react to alighting vultures by running towards the place where they have come down (Kruuk 1967).
Mammalian scavengers, Cathartes vultures and reptiles use the sense of smell to locate carcasses. In the case of small carrion items, chemical cues are the dominant stimuli used by scavengers to locate them (DeVault and Rhodes 2002). In snakes, olfaction also seems to be the principal sensory modality to detect carrion (DeVault and Krochmal 2002). They often consume carcasses of small animals like rodents, birds, fish, frogs and other snakes, which are inaccessible to avian and large mammalian scavenging competitors. Many snake species are attracted to decomposing odors and carrion (DeVault and Krochmal 2002). Scavenging is also widespread among crocodiles. They detect carrion, even concealed carcasses and/or at large distances, by chemical cues in the water and air, and promote olfaction by gular pumping (Weldon et al. 1990; Weldon and Ferguson 1993). Like vultures, crocodiles also possess a low stomach pH that inhibits bacteria from putrefying meat (Diefenbach 1975, in Platt et al. 2007) and can survive long periods without food.
Many major scavengers are social species. Vultures and corvids aggregate in flocks at carcasses and are commensal with other species. Some reptiles, like the Komodo dragon Varanus komodoensis, often feed communally at large carcasses (Bull et al. 2010). Within avian scavenger guilds, interspecific competition has been reduced by divergent eco-morphological characters and dominance hierarchies that lead to an ordered resource partitioning (Kruuk 1967; Wallace and Temple 1987; Houston 1988; Hertel 1994). The costs of adaptation to obligate scavenging are high, as evidenced by the small portion of terrestrial vertebrates relying exclusively on carrion (DeVault et al. 2003). Probably the most critical adaptations are related to detection (including the costs of locomotion) and detoxification of carrion (Shivik 2006). However, scavenging seems highly beneficial as an occasional energy acquisition strategy, given the fact that a large number of terrestrial vertebrates are facultative scavengers (DeVault et al. 2003; Wilson and Wolkovich 2011).
Aquatic Scavengers
There are numerous fish species which scavenge. Across a depth range of 800–5000 m, around 22% of the fish assemblage is thought to be scavengers and this proportion is relatively consistent across this range (Yeh and Drazen 2011). Scavenging requires a different set of sensory and swimming performance characteristics than are required for herbivorous and predatory fish. Food resources are less predictable, requiring some degree of endurance to allow fish to reach their next meal, but can be large (e.g. whale carcasses) when obtained (see chapter “Carrion Availability in Space and Time”). Carrion does not attempt to escape and so there is no requirement for burst swimming performance, though animals may need to compete for, or defend, carrion. Unlike on land, carrion may move with the current, but in general scavengers must detect this resource without the aid of prey movement as a signal. The characteristics of marine carrion shape the physiology, behavior and morphology of swimming scavengers. Scavenging fish have been studied in the deep sea for decades, in spite of the difficulty of working in this environment. Observations of scavenging fish in the abyss by Isaacs and Schwartzlose (1975) changed forever the scientific view of the deep ocean by showing that “active animals” were present even in this environment with few food resources.
There is strong evidence that carrion is usually detected from the odor plume emitted from the carrion. Observations at bait show fish arriving from downstream of the bait (Wilson and Smith 1984), indicating that the fish had either followed a trail of chemicals released from the carrion (chemotaxis) or swum generally up-current (rheotaxis) after detecting that carrion was in their vicinity. Because most studies of scavengers are conducted by still or video photography of bait, scientists seldom have information on how the scavengers behaved prior to arrival. Løkkeborg (1998) tracked cod Gadus morhua in fjords and observed fish turning towards bait and swimming to it. These results indicated that fish were active prior to bait detection at ranges of up to 700 m. Deep sea researchers also fed transponders to fish inside small bait packages at a photographic lander (a free-falling autonomous vehicle) to track them for periods of 3–9 h, at ranges of up to 500 m from the camera lander (Bagley et al. 1994, 2004). Photography at the lander allowed the species and size of the scavenger to be determined, while a current meter measured the direction and speed of currents. Evidence from the tracks of the fish suggested that abyssal grenadiers (Coryphaenoides spp.) swum perpendicular to the current direction. This behavior would maximize their chance of crossing an odor plume being emitted by a carrion item on the seafloor (Bagley et al. 2004).
Bailey and Priede (2002) used a simple mathematical model to predict the arrival time patterns of fish using different search strategies. These were “cross-current” scavenging, where fish swam across the current until detecting a plume and then turned upstream to swim to the bait; “sit and wait”, where the scavenger remained stationary unless an odor plume was detected; and “drifting”, where scavengers went with the current until coming close enough to the carrion to hear scavengers feeding or detect other evidence of activity (e.g. stimulated bioluminescence). Using a range of assumptions, the arrival patterns predicted that the “cross current” model was the best fit to real data for abyssal fish.
All models assumed that once in the vicinity of the bait the fish would localize it using sound, hydrodynamic disturbance or bioluminescence, but at that time the process had not been studied. Using video at bait showed deep sea fish “pole-dancing” around the lander, repeatedly circling the legs of the vehicle and other structures in the field of view. A combination of video analysis and anatomical investigation showed that the fish were using a chin barbell to taste their way around the vicinity of the bait. It appears that for these fish the odor plume got them close to the bait, after which the signal was not clear enough for them to locate it directly. Tasting the seabed until the carrion was obtained seemed to be the solution (Bailey et al. 2007).
In general, we know little about how fish respond to carrion, which is surprising as the scavenging behavior by fish is the basis of the long-line fishing and fish trapping industries (Stoner 2004). Whereas deep sea fish are often surprisingly tolerant of competitors around carrion, given how sparse these resources might be, this is definitely not the case for other taxa (Stoner 2004). Wolffish Anarhichas denticulatus have been observed aggressively defending bait (Godø et al. 1997) and preventing cod and haddock Melanogrammus aeglefinus from obtaining it. Aggression between shallow-water fish clearly reduces the numbers of fish able to feed on carrion (Dunlop et al. 2015) by scaring smaller individuals away from this food source. The extent of aggression around carrion was density dependent, with fish being more likely to interact aggressively at high densities. Carrion-defending and aggressive behaviors probably affect the importance of scavenging across fish assemblages, with some taxa and size ranges dominating the resource. Density-dependent aggression of the type observed by Dunlop et al. (2015) will tend to reduce the difference in catches between areas (and times) of high and low population density. As a result, the effects of spatial management, or trends with time, could be hidden by differences in fish behavior. This is a very practical example of how an understanding of scavenger ecology is essential to management.
Scavengers Across the World
Tundra and Polar Regions
The tundra is a relatively simple ecosystem inhabited by many endemic species adapted to low productivity and cold temperatures, like the Arctic fox and the polar bear. Tundra food webs are marked by complex temporal fluctuations; the best known example is the cycles of voles and lemmings (Killengreen et al. 2011 and references therein). In high latitudes, carrion is a key food resource used to survive periods of food scarcity and overcome harsh environmental conditions. Carrion resources, mostly of marine origin, are associated with seabird and seal colonies, carcasses of marine mammals washed ashore and other marine subsidies, like dead fish (Fig. 1).

Examples of scavengers in different ecosystems (from top to bottom): polar bears congregate at a whale carcass in the Arctic (photo by Alexander Gruzdev); wolves are often followed by raven flocks to feed on their kills (photo by Dan Hartman); the scavenger community of the African savanna is one of the richest (photo by Sarah C. Jones); condors and other vulture species rely often on livestock carcasses in Patagonian steppes (photo by Manuel de la Riva)
Giant petrels Macronectes halli and M. giganteus are the principal scavenging seabirds in the Antarctic ecosystems, and feed mainly on penguin and seal carrion. Seal carrion is highly important for giant petrels during the breeding season, particularly for males. These species also scavenge on floating carcasses during pelagic trips (Hunter 1985; González-Solís et al. 2002). Giant petrels are also frequent scavengers at penguin colonies, together with kelp gulls Larus dominicanus, lesser sheathbills Chionis minor and sub-Antarctic skuas Catharacta lonnbergi (Hunter 1991). The populations of some seal species in the Antarctic have expanded, promoting an increase in carrion and placenta availability, which has benefited scavengers like the skuas (Phillips et al. 2004).
In the northern polar regions, carrion may come additionally from carcasses of large herbivores, like the reindeer Rangifer tarandus or muskox Ovibos moschatus, and from polar bear kills. Polar bears kill mostly ringed seals Phoca hispida and bearded seals Erignathus barbatus, and leave behind a substantial portion of the carcasses, which are scavenged afterwards by Arctic foxes and eventually also by other polar bears (Stirling and Archibald 1977). Polar bears can also kill bigger prey, like beluga whales Delphinapterus leucas, even in high numbers when they are entrapped in ice. Lowry et al. (1987) documented the killing of more than 40 belugas in an ice entrapment and observed 15 polar bears feeding on them. Twelve days later, carrion was distributed widely and at least 30 polar bears were recorded feeding. Such bear aggregations have been recorded scavenging on large whale carcasses (Bentzen et al. 2007; Fig. 1). Ivory gulls Pagophila eburnea and glaucous gulls Larus hyperboreus are important avian scavengers in Svalbard (Gjertz and Lydersen 1986). In the Canadian Arctic, ravens, gulls, arctic foxes, and even wolves have been also recorded scavenging on polar bear kills (Stirling and Archibald 1977).
The Arctic fox is the mammalian scavenger per excellence in the tundra and polar habitats. Their use of carrion and marine subsidies is highest during winter and lemming population declines (Roth 2002). Carrion storage, and food caching in general, is particularly common among Arctic foxes as a response to trophic pulses and fluctuating environments (Careau et al. 2007; Samelius et al. 2007). With the shrinking ice cover and tundra under global warming, the influx of predator-scavengers from neighboring ecosystems (e.g. red foxes from boreal forests) is increasing competition and eventually could exclude Arctic foxes from carcasses (Killengreen et al. 2011, 2012).
Temperate and Boreal Forests
Temperate and boreal forests are complex ecosystems where scavenging is widespread. Although there are very few obligate scavengers (e.g. turkey vultures Cathartes aura), all predators are scavengers to some extent. Most important are avian and mammalian scavengers. For instance, predators such as the goshawk and the weasel, or insectivores like shrews and woodpeckers also consume carrion when the opportunity arises. More than 30 species of birds and mammals were recorded consuming carrion in the forests of Bialowieza, northeastern Poland (Selva 2004). Typically, the bulk of the carrion found in these forests are ungulates, mainly provided by large predators such as wolves, by harsh winters, or by human-caused mortality like hunting (Houston 1978; Wilmers et al. 2003a, b; Selva and Fortuna 2007; Mateo-Tomás et al. 2015). Carrion is a very important food resource for vertebrates in winter and early spring, and support hibernating species like the brown bear Ursus arctos (Green et al. 1997). Scavengers, and particularly the brown and black bear Ursus americanus, also rely on carrion pulses from spawning salmon that arrive every year in some rivers of these forests (Fortin et al. 2007; Levi et al. 2015, see chapter “Ecological Functions of Vertebrate Scavenging”). Caching food (i.e. storing it in locations hidden from the sight of conspecifics and other species) is common among the main scavengers like corvids and canids.
The most efficient and frequent scavenger in northern forests is probably the common raven (Wilmers et al. 2003b; Selva et al. 2005; Wikenros et al. 2013; Fig. 1). Other corvids like jays, magpies, crows and nutcrackers are frequent visitors to carcasses (Wilmers et al. 2003b; Selva et al. 2005; Wikenros et al. 2013; Ray et al. 2014). Among raptors, the common buzzard Buteo buteo, as well as eagles (e.g. white-tailed eagle Haliaeetus albicilla, golden eagle Aquila chrysaetos and bald eagle Haliaeetus leucocephalus) rely on carrion during some periods and efficiently exploit hunter-kills, often concentrated in space and time and exposed in more open areas within the forest (Wilmers et al. 2003a; Selva et al. 2005; Mateo-Tomás et al. 2015; Fig. 1).
Mammalian scavengers in temperate and boreal forests are also well represented. As in other biomes, canids such as red foxes, coyotes Canis latrans, raccoon dogs Nyctereutes procyonoides, and wolves are among the most frequent scavengers (Selva et al. 2005; Wikenros et al. 2013). Wolves not only subsidize other scavengers with carrion, but also facilitate scavenging of large ungulate carcasses by opening them (Selva et al. 2003). Some mustelids, like martens Martes spp. and wolverines Gulo gulo, are quite dependent on carrion, particularly in years when alternative sources of prey (e.g. rodents or hares) are scarce (Jędrzejewski et al. 1993; Koskela et al. 2013). Felids are not frequent users of carrion, and often suffer kleptoparasitism by larger species like the brown bear, the wild boar, and the wolverine, with a limited or significant effect on lynx predation rates depending on the area (Selva 2004; Mattisson et al. 2011; Krofel et al. 2012).
Mediterranean Regions
Scavengers in these regions have been traditionally associated with cattle farming. Carrion has been mostly supplied through livestock carcasses, hunting of wild ungulates, and more recently, artificial feeding stations or vulture restaurants and dumps (Oro et al. 2013). The scavenger species at these carrion supplies are the same, but the number of individuals of these species, as well as their time of detection and consumption is very different (Cortés-Avizanda et al. 2010, 2012, 2014, Moreno-Opo et al. 2015a, b, 2016). Within the Mediterranean region, Spain is home to >90% of the obligate European scavengers (Eurasian griffon vulture Gyps fulvus, Egyptian vulture Neophron percnopterus, bearded vulture Gypaetus barbatus and cinereous vulture Aegypius monachus, Fig. 1) and of a rich community of facultative scavengers which includes some threatened species like the Spanish imperial eagle Aquila adalberti, the red kite Milvus milvus, and the Iberian lynx Lynx pardinus. The guild of obligate avian scavengers in other Mediterranean countries, like Italy, France, Portugal or Greece, constitutes less than 10% of the vulture population in the European Union (Deinet et al. 2013; Margalida et al. 2014). This has promoted several reintroductions to recover the scarce populations of some avian scavengers, like the bearded vulture in the Alps (Austria, Italy, France and Switzerland) and Massif Central (France), the Eurasian griffon vulture in Italia, Israel and France or the cinereous vulture in France (see reviews in Terrasse 2001; Margalida and Heredia 2005; Eliotout 2007; Donázar et al. 2009).
Among avian scavengers, Eurasian griffon vultures and common ravens are the most habitual species scavenging carrion resources in the Mediterranean (Cortés-Avizanda et al. 2010; Mateo-Tomás et al. 2015; Moreno-Opo et al. 2015a, 2016). In this sense, griffon vultures play important ecosystem services eliminating ca. 10 thousand tons of meat per year, and, in the case of bearded vultures, 200 tons of bones (Margalida and Colomer 2012; Margalida et al. 2012). Other corvids that frequently visit carrion resources are the carrion crow Corvus corone, magpie Pica pica and Eurasian jay Garrulus glandarius. Among raptors other than vultures, the common buzzard, the red and black kite, the golden and Spanish imperial eagles and, more occasionally goshawks, obtain additional food resources from carrion subsidies (Sánchez et al. 2009; Sánchez-Zapata et al. 2010; Mateo-Tomás et al. 2015; Moreno-Opo et al. 2016; Margalida et al. 2017). In particular, non-adult individuals of golden and Spanish imperial eagles are prone to use scavenging as a feeding strategy in winter, coinciding with low levels of their main natural prey and the occupancy of low quality habitats (Sánchez et al. 2009; Sánchez-Zapata et al. 2010; Margalida et al. 2017). With respect to mammalian scavengers in Mediterranean areas, the most frequent are the red fox, the stone marten Martes foina and the wild boar (Mateo-Tomás et al. 2015).
Savannas
The earliest studies on scavengers were mostly conducted in the African savanna, which encompasses the majority of the savanna regions on the planet. This is not surprising because this continent is home of probably the most emblematic scavengers: vultures and hyaenas (Fig. 1). Classic studies by Petrides (1959), Attwell (1963), Kruuk (1966, 1967, 1972) and Houston (1974, 1975, 1979) on these species have inspired, and still do, many scavenging ecologists worldwide. The scavenger communities of savanna regions of South America, Asia and Australia are poorly known, so here we focus on African scavengers. However, the general characteristics of the scavenging fauna are probably pervasive among continents, with the exception that there are no obligate scavengers in Australia.
The savanna regions of the African continent support the richest community of vultures in the world, including ten resident and one wintering species, eight of which are endemic or near-endemic (Del Hoyo 1994). This species array ranges from consumers of meat and other soft tissues (e.g. Rüppell’s vulture Gyps rüppellii) to bone eaters (bearded vulture) and even one species that rarely scavenges and has evolved to exploit palm-nuts and capture small prey (palm-nut vulture Gypohierax angolensis). Foraging habits among African vultures are also variable, including species that forage solitarily (e.g. white-headed vulture Aegypius occipitalis) and species that forage in pairs (e.g. lappet-faced vulture A. tracheliotos) and in groups (e.g. white-headed vulture Gyps africanus). The morphological, ecological and behavioral diversity of African vultures, together with dominance hierarchies at carcasses, results in an ordered resource partitioning that likely reduces competition for carrion (Kruuk 1967; Hertel and Lehman 1998; Kendall et al. 2012; Kendall 2013). Such a variety has been related to the high diversity of large mammalian herbivores, whose carcasses provide major feeding resources for scavengers. Indeed, Houston (1983) suggested that the evolutionary origin of vultures was closely related to the presence of long-distance migratory ungulates, which still survive in a few African regions.
The community of facultative scavengers inhabiting the African savanna is also diverse and includes omnivorous species such as corvids (e.g. white-necked raven Corvus albicollis), medium-sized predators such as birds of prey (e.g. tawny eagle Aquila rapax) and mammalian carnivores (e.g. side-striped jackal Canis adustus), and apex predators such as hyaenas and large cats (Fig. 1). In areas with resident herbivore populations, spotted hyaenas (Crocuta crocuta) and lions can outcompete vultures and consume most of the ungulate carrion biomass (Moleón et al. 2015). The brown hyena (Parahyaena brunnea), i.e. the mammal species most specialized to use carrion (up to 90% of its diet; Skinner and Chimimba 2005), is found in the driest savannas of Southern Africa.
Steppes
Steppes are located in several regions of the world, mainly Eurasia, North America and South America. This cold, dry grassland is an important biome for scavengers.
In the Americas there are seven species of obligate scavengers, all of them from the family Cathartidae (known as vultures from the New Word). These vultures occur from high mountains to the coastline and from woodlands to the steppes. Apart from them, other facultative scavengers, both mammals and birds, aggregate and compete in carcasses found in those open areas.
In South American steppes, three are the species of obligate scavengers, the Andean condor Vultur gryphus, the turkey vulture and the black vulture Coragyps atratus (Fig. 1). Other bird species that facultatively consume carcasses are the southern Caracara plancus and chimango Milvago chimango caracaras, and the black-chested buzzard eagle Geranoaetus melanoleucus (Travaini et al. 1998; Lambertucci et al. 2009a). Mostly during the dark hours, carcasses are also consumed by mammals such as pumas Puma concolor, foxes Lycalopex spp., skunks Conepatus spp., and armadillos Zaedyus pichiy, and also by passerines and lizards (Travaini et al. 1998; Elbroch and Wittmer 2012). Currently, all these species scavenge mainly on carcasses from domestic animals since steppes have been historically used for extensive livestock raising, which has produced a sharp decline in the populations of native herbivores (Fig. 1). Scavengers also feed on carcasses of exotic wild species, such as the red deer Cervus elaphus and the European hare Lepus europaeus. Native species such as guanacos Lama guanicoe, and rheas Rhea pennata that once were the main supply of carrion are currently much less consumed in many areas due to their low numbers which brought them to an ecological extinction (Novaro et al. 2000; Lambertucci et al. 2009b).
As in other biomes, competition for carrion is important and seems to relate to the size of the species. Generally, bigger species (condors) dominate over the carcasses (Wallace and Temple 1987). However, when the number of individuals of other species strongly increases, the size-related hierarchy could be alternated and smaller species can dominate at carcasses (Carrete et al. 2010). In South American steppes this is the case of black vultures, that can reach very high densities in the surroundings of human settlements and might oust Andean condors at carcasses where generally they would be dominant (Carrete et al. 2010; Barbar et al. 2015).
Scavenger composition is also influenced by the size of the carcass, with few individuals of few species feeding on hare carcasses (Travaini et al. 1998) and more species in larger numbers feeding on larger carcasses like sheep, guanaco or horse (e.g. Lambertucci et al. 2009a). The place where the carcass is located can modify the order of arrival and consumption patterns. Carcasses close to roads are mainly used by smaller bird scavengers as caracaras, while larger species as condors and turkey vultures are the first to eat at carcasses far from roads (Lambertucci et al. 2009a). Moreover, bird scavenger composition can also be affected by weather conditions, and for instance, in gusty or very windy days carcasses are mainly used by larger species (Shepard and Lambertucci 2013). This is particularly relevant in Patagonian steppes which have cold-temperate climate with strong winds.
Regarding mammals, pumas, the largest carnivore in the area, can be observed scavenging, but they are more important as suppliers of carcasses for other species. In fact, they can provide large amounts of meat which are used by at least 12 species in the Patagonian steppes. Pumas need to increase their kill rate to compensate for the losses due to scavengers (Elbroch and Wittmer 2013). Two species of fox (Lycalopex cualpaeus and Lycalopex griseus) are commonly observed displacing smaller species in order to feed on puma kills.
Tropical Forests
In tropical forests Cathartidae and Accipitridae vultures constitute important members of the scavenger community (Houston 1979), but they differ substantially in their distribution and the habitats they occupy. For example, in the tropical forests of Asia and Africa, obligate vulture species are rare when compared to neotropical forests, where species such as Cathartidae vultures are well represented. Two New World vulture species, the turkey vulture and the greater yellow-headed vulture Cathartes melambrotus, are the most common vultures of neotropical forests. In these forests, mammalian carnivores are rather inefficient at locating carcasses. This is a consequence of their limitations in the daily foraging areas covered and the trade-off between the energy invested to search for food and the distance travelled to find temporal and unpredictable food resources. Thus, mammalian carnivores are probably outcompeted by avian scavengers and consume carrion just when the chance appears. Avian scavengers, which use little energy in soaring (Pennycuick 1983) and can quickly cover large areas of forest, are more efficient carrion foragers (Houston 1984, 1986).
With respect to obligate vulture species, the consumption of carrion in forest habitats is possible thanks to the well-developed olfactory lobe and sense of smell which is used for finding food in forested areas (Chapman 1929; Houston 1984, 1986). The three Cathartes species—the turkey, the greater yellow-headed and the lesser yellow-headed Cathartes burrovianus—differ only in their wing and tail shape and in the color of their heads, having the remarkable ability to locate food by smell (Houston 2001). American black vultures and king vultures Sarcoramphus papa are also frequent on tropical forests but have no sense of smell; they often rely on Cathartes vultures behavior to find food.
Agricultural and Other Human-Modified Landscapes
Despite human domination over much of the planet (Waters et al. 2016), scavenging ecology often has been studied in relatively natural environments (e.g., Houston 1974; Wilmers et al. 2003b; DeVault et al. 2004; Selva et al. 2005). However, studies conducted in agricultural and other human-dominated environments suggest that vertebrate scavengers are also very active in such habitats. In this section, we briefly review studies investigating carrion use by vertebrates in heavily-altered landscapes, focusing on agricultural habitats, a major land use across the world where diversity is often reduced and vertebrate communities are dominated by a few generalist species (Swihart et al. 2003).
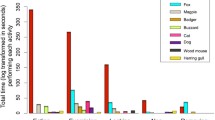
In an intensively farmed region of northern Indiana, USA, small carrion items, such as mouse carcasses, were scavenged extensively by two species of generalist mesopredators: raccoons and Virginia opossums Didelphis virginiana (DeVault et al. 2011). The high scavenging efficiency by vertebrates in the agricultural region was attributed to elevated densities of raccoons and Virginia opossums, and the reduced search area for carrion in the sparsely distributed forested habitat islands (DeVault et al. 2011). For larger carcasses in the same northern Indiana agricultural area, Olson et al. (2016) found that 87% of raccoon, Virginia opossum, and rabbit Oryctolagus spp. carcasses were fed upon by 16 species of vertebrate scavengers, and included more bird scavengers (seven species) than was found for the smaller mouse carcasses in the earlier study (DeVault et al. 2011). Other studies have investigated scavenging efficiency by vertebrates on carcasses (usually birds) found in agricultural fields themselves. Scavenging efficiency ranged from 38% to 92% for small bird carcasses (Balcolm 1986; Tobin and Dolbeer 1990; Linz et al. 1997; Kostecke et al. 2001) and was 96% for large birds (mallards Anas platyrhynchos; Peterson et al. 2001).
Wide-ranging scavengers that use highly altered, agricultural habitats as well as more natural habitats have been shown to adjust foraging strategies based upon available resources. For example, in the USA, turkey vultures fed extensively on domestic farm animal carcasses in an agricultural region (Coleman and Fraser 1987), whereas their diet was almost exclusively wild vertebrates in a heavily forested area (Kelly et al. 2007). Likewise, in Bulgaria, the diet of the facultative scavenging golden jackal Canis aureus included a large percentage of domestic animals in an agricultural area, whereas in a forested region wild ungulates were the dominant food items (Raichev et al. 2013).
Scavenging efficiency by vertebrates varies widely across human-dominated landscapes, and appears to be influenced by many factors including habitat, composition of the vertebrate and invertebrate community, and climate. Broadly, it appears that carcass use by vertebrates in agricultural regions is equivalent to, or in some cases higher than that found in more natural habitats. Smaller carcasses produced in agricultural habitats are usually removed by mammalian scavengers, whereas bird scavengers often use larger carcasses.
Deserts
In arid and semiarid environments, water availability is often the limiting factor in the decomposition process. Carcasses can desiccate before they are completely decomposed in some circumstances (Payne 1965; Parmenter and MacMahon 2009), and dried-out carcasses are generally less attractive to vertebrate scavengers than those that retain some water (Parmenter and MacMahon 2009). Water loss from carcasses is strongly influenced by surface area-to-volume ratios, with smaller carcasses having higher ratios and, thus, greater water loss rates (Parmenter and MacMahon 2009).
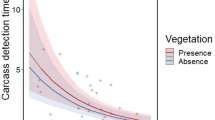
Despite water limitations, vertebrate scavengers are active in arid environments and usually feed upon available carcasses, especially when temperatures are cool (Stoddart 1970; Travaini et al. 1998; Parmenter and MacMahon 2009). Furthermore, carcass use by facultative scavengers can actually increase following water shortages because of a relative lack of alternative food resources. Brown et al. (2006) showed that carcass use by vertebrate scavengers increased following a period of below-average rainfall coincident with the El Niño Southern Oscillation in semi-arid southeastern Australia.
Like in other ecosystems, a variety of mammal and bird species are prolific scavengers in arid and semiarid environments. Stoddart (1970) studied the fate of 45 black-tailed jackrabbit Lepus californicus carcasses in northwestern Utah, USA. Of these, 60% were scavenged by birds (species undetermined). In arid southern Australia, corvids (Corvus coronoides and C. bennetti), wedge-tailed eagles Aquila audax and red foxes fed extensively on kangaroo offcuts left in the field after butchering (Read and Wilson 2004). Considering smaller carcasses, Rogers et al. (2014) found that common ravens, kit foxes Vulpes macrotis, coyotes, and greater roadrunners Geococcyx californianus found and fed upon 20 out of 25 carcasses of small migrating birds that died from colliding with power lines in the Sonoran Desert (California, USA).
Coastal Systems
The world’s coastline is an estimated 1,634,701 km long, ranging across all the climatic zones, from the polar, through the temperate, to the tropical regions. Coastal ecosystems represent the interface between land and sea and provide carrion mainly via colonies of mammals, seabirds and reptiles or through deposition of carcasses originated at sea (Polis and Hurd 1996; Moss 2017). Vertebrate scavengers inhabiting the coastal surface and exploiting carcasses provided by pinniped and seabird colonies include mostly birds and mammals. The procellariids southern and northern giant petrels opportunistically scavenge on dead seals and penguins in Antarctica (Bruyn and Cooper 2005). In Southern Chile, black vultures, kelp gull and feral dogs Canis familiaris compete for carrion occurring in a sea lion Otaria flavescens colony (Pavés et al. 2008). At temperate latitudes, the great black-backed gull Larus marinus and the red fox consume placenta and dead pups at grey seal Halichoerus grypus breeding colonies (Twiss et al. 2003; Culloch 2012; Quaggiotto et al. 2018; Fig. 1). In Namibia (Africa) brown hyenas restrict their range to the coastal area to scavenge on carrion produced during pupping season of the Cape fur seal Arctocephalus pusillus (Kuhn et al. 2008).
Similarly, marine inputs deposited on the coastline of California attracted coyotes to the shore, causing a numerical response in the population density which increased between 2.4 and 13.7 times in relation to the adjacent desert (Rose and Polis 1998). In experimental conditions, fish carcasses on a sandy beach in Australia determined the formation of consumers’ aggregation around the carrion patches. In this sub-tropical coastal environment not only the opportunistic Torresian crows Corvus orru and silver gull Chroicocephalus novaehollandiae, but also the birds of prey like the white-bellied sea eagle Haliaeetus leucogaster, whistling kite Haliastur sphenurus and brahminy kite Haliastur indus, and the lizard lace monitor were observed scavenging on the fish carcasses (Huijbers et al. 2015). Marine carrion is crucial in environments such as deserts, sandy beaches and tundra, where productivity is low (Spiller et al. 2010).
On urbanized beaches in Australia the scavenging community feeding on fish carcasses included rats Rattus spp., domestic cats Felis catus, dogs and red foxes, but not raptors. Raptors occurred only in rural areas, where they completely removed carrion within 24 h (Huijbers et al. 2013). Mammals scavenging at urban locations did not cover the functional role of raptors, as they could not compensate the extent of scavenging activity of the avian counterpart (Huijbers et al. 2015). By changing species composition of the coastal scavenging community, urbanization has also an impact on the scavenging processes occurring in these systems. Furthermore, the effect of invasive species, such as the red fox in Australia, can be detrimental to the endemic scavengers. Red foxes, for instance, can remove a large number of carcasses during the night and before the diurnal raptors even start foraging (Brown et al. 2016).
Carrion consumption by coastal scavengers may vary because of human activity (e.g. Lambertucci et al. 2018). Ronconi et al. (2014) found that great black-backed gulls living on Sable Island (Canada) rely more on seal carrion as food source than forty years ago. An increase of the seal population, changes in fish availability, reduction of fishery discards and alterations in the surrounding ecosystem could all have led to this shift in gull’s diet.
Marine and Freshwater Systems
In marine systems, carrion is exploited by many invertebrates at any depth, while vertebrate scavenging species are mostly found closer to the deep-sea floor. In shallow waters, fish observed visiting experimental marine mammal carcasses were found to be feeding on the amphipods colonising the carcass (Jones et al. 1998). Whale falls in the bathyal zone attract a large number of sleeper sharks Somniosus pacificus, hagfish Eptatretus deani and Mixine circifrons, and macrourids which scavenge the soft tissue of the whale carcass. These species characterize the first stage of the whale consumption, called mobile-scavenger stage, which lasts from months to years. Around 4400 m depth, grenadiers Coryphaenoides armatus were observed approaching experimental cetacean carcasses, feeding on the amphipods at the carcass, rather than scavenging (Jones et al. 1998). Floating whale carcasses attract scavenging elasmobranchs, such as the great white shark Carcharodon carcharias, the tiger shark Galeocerdo cuvier and the blue shark Prionace glauca (Long and Jones 1996; Dudley et al. 2000; Curtis et al. 2006). Although it is thought that scavenging behavior in sharks is common, this was occasionally observed and rarely quantified (Fallows et al. 2013). These large predatory fish congregate or solitarily arrive at carcasses attracted by the olfactory cues released by the decaying whale and transported by wind (Fallows et al. 2013). Juvenile individuals also scavenge on dead whales (Dicken 2008). However, adults are the most frequent attendants of such banquet, which are hypothesized to trigger mating between mature individuals (Fallows et al. 2013). Scavenging by sharks is common at whale calving grounds where both carrion and predatory opportunities increase (Bonfil et al. 2010). There is evidence that tiger sharks also scavenge on bird dies-offs at sea (Gallagher et al. 2011).
Notoriously, killer whales Orcinus orcas, together with sharks, were the main vertebrate scavengers of whale carcasses and their remains produced by the whaling industry. It is likely that killer whales adopted this opportunistic behavior and specialized in scavenging on such nutritious resources when these were patchy and temporally predictable. Before (and possibly after) the whaling era, instead, scavenging represented one of the foraging alternatives used opportunistically by these apex predators (Whitehead and Reeves 2005). Scavenging seabirds, such as kelp gulls, Antarctic skuas Catharacta antarctica, black browed albatrosses Thalassarche melanophris and different species of petrels often associate with hunting killer whales to easily gain some pieces of food from the remains of the killed prey (Williams et al. 1990).
Carrion in freshwater ecosystems is mainly derived from fish or aquatic birds, and it is used by a diverse array of scavengers, even Eurasian otters (O’Sullivan et al. 1992). Some freshwater systems experience a carrion pulse every year derived from fish migrating to their spawning areas, like salmon (see chapter “Ecological Functions of Vertebrate Scavenging”). Such food pulses support high-density populations of brown bears (i.e. Hilderbrand et al. 1999) and other mammals such as wolves, marten, mink, and coyotes, and a diverse array of avian scavengers including bald eagles, ravens, jays, mergansers, gulls, and even owls (Levi et al. 2015). Some of these predator-scavengers of salmon, like the glaucous-winged gulls (Larus glaucescens) and brown bears shifted their spatial distribution together with salmon, thus closely tracking the phenology of their main food resource (Schindler et al. 2013). Mobile scavengers, like gulls, by transporting nutrients derived from fish carrion to distant locations, have an important impact on local aquatic communities (Payne and Moore 2006).
Structure of Scavenger Communities
Interactions among obligate scavengers have always been recognized as highly organized and driven by competition (Kruuk 1967; Wallace and Temple 1987), whereas scavenging has traditionally been viewed as a more random and opportunistic process for facultative scavengers (Hiraldo et al. 1991). In the last decade, however, many studies have demonstrated that vertebrate scavenger communities can show complex patterns that deviate from random processes (Selva and Fortuna 2007). We can consider that an assemblage is structured if it is more ordered than expected by chance. In random communities, interactions are the result of opportunistic encounters (i.e. a facultative scavenger finding a carcass without an active search for it). Structured communities show identifiable patterns and are a consequence of one or several interacting processes shaping which interactions are possible and which more likely (Selva and Fortuna 2007).
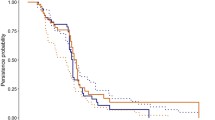
One of the patterns that has been used to study the structure of vertebrate scavenger communities is nestedness. A community of interacting species is nested if the specialist species interact with a subset of the species that interact with the more generalist species (Bascompte et al. 2003). This concept has been widely identified in interacting communities, especially mutualistic interactions (Bascompte and Jordano 2007), with important implications for the stability and persistence of mutualistic networks (Bastolla et al. 2009; Rohr et al. 2014). A nested pattern in scavenger communities emerges when the species feeding on carcasses visited by few consumers are subsets of the assemblage of species feeding on carcasses visited by more consumers (Fig. 2). Selva and Fortuna (2007) were the first authors that identified nestedness in a scavenger assemblage. Since then, we know that nestedness is not universal (i.e. it does not appear in all the scavenger communities), and that it depends on many interacting factors, such as the type of carcass, composition of the scavenger assemblage, and the competition degree of the community (Sebastián-González et al. 2013; Allen et al. 2014; Moleón et al. 2015).

Schematic representation of a perfectly nested scavenger community. Light gray circles represent carcasses, dark grey circles represent scavengers. A carcass is linked to a scavenger if it is consumed by that species. Scavengers are from top to bottom: vulture, fox, wild boar, raven and magpie. In this example, the species feeding on carcasses visited by few consumers (i.e. vulture) are subsets of the assemblage of species feeding on carcasses visited by more consumers (i.e. first carcass on the top, consumed by all the scavengers). Picture performed by E. Sebastián-González. Silhouettes from The Noun Project https://thenounproject.com, authors: S. Laing, A. Bearne, M. Turan Ercan, H. Richir, Bluetip Design and P. Lehmann
The type of carcass is one of the factors affecting the structure of scavenger communities. For example, the contribution to nestedness (i.e. how much a species or a carcass follows the nested pattern) was high in carcasses originated from human hunting, it was medium for carcasses from predator kills and lower from animals dead from disease or starvation (Selva and Fortuna 2007). This pattern is related to the spatio-temporal distribution and predictability of the resources. Hunter kills are normally aggregated in an area (i.e. hunting spots) and in time (i.e. hunting season). This can lead to a super-saturation of the resource, allowing the exploitation of carcasses by more consumers (Wilmers et al. 2003a, but see Cortés-Avizanda et al. 2012 for a different perspective of a similar problem). In contrast, death by disease or starvation is a much more random process that can occur at any time and location, and these carcasses are mainly consumed by carrion specialists. Other carcass characteristic found to be very important for assemblage structure is size. Scavenger communities consuming larger carcasses (>10 kg) were organized under a nested pattern, while small carcasses showed a more random organization (Sebastián-González et al. 2013; Moleón et al. 2015). Larger carcasses may be partitioned by a higher number of species, promoting both competitive and facilitative processes, which can lead to more structured communities. Small carcasses, however, are often consumed by a single individual, increasing the likelihood of opportunistic encounters and the randomness in the interactions.
Independently of the characteristics of the carcass, the composition of the scavenger community also has important implications for the structure of the assemblage. Birds presented a higher contribution to nestedness than mammals, indicating that they are the main responsible for the nested structure of the community (Selva and Fortuna 2007). Moreover, Sebastián-González et al. (2016) studied scavenger communities in Spain and found that those assemblages including obligate scavengers (i.e. vultures) were nested, while those lacking vultures presented a more random organization (see also Sebastián-González et al. 2013). Two facilitation mechanisms may be behind the role of key species in the structure of the community: carcass location and skin opening. Avian scavengers in general, and vultures in particular, are more adapted to scavenging because of their low energy-demanding search strategies (Selva et al. 2005; Jackson et al. 2008). Thus, birds are able to encounter and consume most of the carcasses available in an area and can act as indicators of the presence of the carcass for other species. If the ability to locate the resource differs among scavenger species, the facilitation of carcass location may lead to an increase in the nestedness of the assemblage. Vultures can also facilitate carcass use to other species by opening thick-skin animals. Many scavengers are unable to consume carcasses from intact (i.e. unopened) animals because they lack the tools (e.g. strong beak) or the strength to break the skin. The same mechanism has been proposed for hyenas (Moleón et al. 2015), pumas (Allen et al. 2014) and wolves (Selva et al. 2003). Limitation of the trophic resources by competitive interactions can also increase nestedness through interspecific dominance hierarchies. For example, the presence of black bears in a Californian scavenger community increased nestedness by decreasing complementary use of carrion by other species (Allen et al. 2014).
Nestedness has been found to be high under stressful conditions in several animal communities (Worthen et al. 1998; Baber et al. 2004; Cook et al. 2004). Theoretical studies on mutualistic interaction networks suggest that nestedness can reduce competition among species and enhance coexistence (Bastolla et al. 2009; Rohr et al. 2014). Scavenger communities seem to mirror other animal communities in their relationship among nestedness and competition. Selva and Fortuna (2007) found that the scavenger community in the cold season, when food is scarcer and weather is more extreme, was more nested than at the warm season. Moreover, both species richness and nestedness are characteristic of scavenger networks that were robust to high levels of interspecific competition for carrion (Sebastián-González et al. 2016). Thus, nestedness may favor the coexistence of interacting species by counterbalancing the effects of high competition.
Thus, we conclude that scavenger assemblages show non-random nested organizations that can emerge when carcasses are not monopolized by a single consumer (e.g. because they are small). In general, nestedness in scavenger communities is the consequence of three interacting processes: (1) predictability in the availability of the resource, (2) facilitative processes provided by key species, and (3) differences in the abilities of the scavenger species to compete for the carrion.
Conclusions and Future Perspectives
Although there are few obligate scavengers, carrion consumption is a foraging strategy widely spread among vertebrates in all biomes. The scavenger species, the number of obligate scavengers, and even the adaptations to locate and consume carrion differ among ecosystems. However, everywhere scavenging is a profitable strategy to acquire energy, even if sporadic, at the individual, population and community level, and an important path for energy and nutrient cycling.
Recent advances in technology, such as automatic cameras or satellite telemetry, have exponentially improved the amount and quality of collected data, which have greatly contributed to obtain a complete picture of many scavenger communities and a better understanding of their functioning. This has been particularly important for ecosystems where direct observations have been difficult or impossible, like polar or abyssal regions. Although important achievements in scavenging ecology have been made in the last decades, there are still pending tasks, such as the identification of all scavenger species in some ecosystems and their inter- and intra-specific interactions at carrion, or the assessment of the role of carrion in the ecology and population dynamics of many obligate and facultative scavenger species.
Box 1 Vultures in Tibet and Sky Burials
The Himalayan range is the mountain region in the world where more species of vultures exist. However, the information on species population numbers and ecology for that area is rather limited. Most of the research in southern Asia comes from India, and is particularly focused on the vulture crisis suffered there (Green et al. 2004). As much as nine species of vultures are found in the southern portion of the Himalayan range. Eight of those species are present in China, seven of which are present in the Tibetan plateau as well, one of the richest vulture regions in the world (MaMing and Xu 2015).
In the Tibetan plateau, which is over 4000 m asl, vultures are the main scavengers and feed not only on wildlife and livestock carrion, but also on human corpses. A common cultural practice in this area are the sky burials. In this type of funeral the dead human body is placed on a platform on a mountain from where scavengers (mainly vultures) consume it. The remains are disposed for scavengers in as generous a way as possible (Fig. 3). The Himalayan griffon vulture Gyps himalayensis is the most common species at sky burials, and can be counted at hundreds in a single funeral; cinereous and bearded vultures are also observed at sky burials (MaMing et al. 2016). Tibetans believe that the cessation of breathing is only a first stage of death. Tibetan Buddhists view death as the journey from this life to the next one, and consider vultures as sacred animals aiding humans in this journey. This makes vultures highly important for both their environment and culture.

Vultures in the Tibet feed on human corpses at the called “sky burials”. Photo credit: Lama Rinczen
References
Allen ML, Elbroch LM, Wilmers CC et al (2014) Trophic facilitation or limitation? Comparative effects of pumas and black bears on the scavenger community. PLoS ONE 9(7):e102257
Attwell RIG (1963) Some observations on feeding habits, behavior and inter-relationships of Northern Rhodesian vultures. Ostrich 34:235–247
Baber MJ, Fleishman E, Babbitt KJ et al (2004) The relationship between wetland hydroperiod and nestedness patterns in assemblages of larval amphibians and predatory macroinvertebrates. Oikos 107:16–27
Bagley P, Smith A, Priede I (1994) Tracking movements of deep demersal fishes in the Porcupine seabight, north-east Atlantic Ocean. J Mar Biol Assoc 74:473–480
Bagley P, Priede I, Amieson A et al (2004) Lander techniques for deep-ocean biological research. Int J Soc Underw Technol 26:3–12
Bailey D, Priede I (2002) Predicting fish behaviour in response to abyssal food falls. Mar Biol 141:831–840
Bailey D, Wagner H, Jamieson A et al (2007) A taste of the deepsea: the roles of gustatory and tactile searching behaviour in the grenadier fish Coryphaenoides armatus. Deep Sea Res Part I Oceanogr Res Pap 54:99–108
Balcolm R (1986) Songbird carcasses disappear rapidly from agricultural fields. Auk 103:817–820
Barbar F, Werenkraut V, Morales JM et al (2015) Emerging ecosystems change the spatial distribution of top carnivores even in poorly populated areas. PLoS One 10(3):e0118851
Barnosky AD, Koch PL, Feranec RS et al (2004) Assessing the causes of late Pleistocene extinctions on the continents. Science 306:70–75
Bascompte J, Jordano P (2007) Plant-animal mutualistic networks: the architecture of biodiversity. Annu Rev Ecol Evol Syst 38:567–593
Bascompte J, Jordano P, Melián CJ et al (2003) The nested assembly of plant-animal mutualistic networks. Proc Natl Acad Sci U S A 100:9383–9387
Bastolla U, Fortuna MA, Pascual-García A et al (2009) The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature 458:1018–1020
Beasley JC, Olson ZH, DeVault TL (2015) Ecological role of vertebrate scavengers. In: Benbow ME, Tomberlin J, Tarone A (eds) Carrion ecology, evolution, and their application. CRC Press, Boca Raton, pp 107–128
Bentzen TW, Follman EH, Amstrup SC et al (2007) Variation in winter diet of southern Beaufort Sea polar bears inferred from stable isotope analysis. Can J Zool 85:596–608
Blumenschine RJ, Cavallo JA (1992) Scavenging and human evolution. Sci Am 267:70–76
Bonfil R, Francis MP, Duffy C et al (2010) Large-scale tropical movements and diving behavior of white sharks Carcharodon carcharias tagged off New Zealand. Aquat Biol 8:115–123
Brown OJF, Field J, Letnic M (2006) Variation in the taphonomic effect of scavengers in semi-arid Australia linked to rainfall and the El Niño Southern Oscillation. Int J Osteoarchaeol 16:165–176
Brown M, Schlacher T, Schoeman D et al (2016) Invasive carnivores alter ecological function and enhance complementarity in scavengers assemblages on ocean beaches. Ecology 96:2715–2725
Bruyn PJN, Cooper J (2005) Who’s the boss? Giant petrel arrival times and interspecific interactions at a seal carcass at sub-Antarctic Marion Island. Polar Biol 28:571–573
Buckley NJ (1996) Food finding and the influence of information, local enhancement, and communal roosting on foraging success of North American vultures. Auk 113:473–488
Bull JJ, Jessop TS, Whiteley M (2010) Deathly drool: evolutionary and ecological basis of septic bacteria in Komodo dragon mouths. PLoS ONE 5(6):e11097
Careau V, Giroux JF, Berteaux D (2007) Cache and carry: hoarding behaviour of Arctic fox. Behav Ecol Sociobiol 62:87–96
Carrete M, Lambertucci SA, Speziale K et al (2010) Winners and losers in human-made habitats: interspecific competition outcomes in two Neotropical vultures. Anim Conserv 13:390–398
Chapman FM (1929) My tropical air castle. D Appleton-Century Co, New York
Coleman JS, Fraser JD (1987) Food habits of black and turkey vultures in Pennsylvania and Maryland. J Wildl Manag 51:733–739
Cook RR, Angermeier PL, Finn DS et al (2004) Geographic variation in patterns of nestedness among local stream fish assemblages in Virginia. Oecologia 140:639–649
Cortés-Avizanda A, Carrete M, Donázar JA (2010) Managing supplementary feeding for avian scavengers: guidelines for optimal design using ecological criteria. Biol Conserv 143:1707–1715
Cortés-Avizanda A, Jovani R, Carrete M et al (2012) Resource unpredictability promotes species diversity and coexistence in an avian scavenger guild: a field experiment. Ecology 93:2570–2579
Cortés-Avizanda A, Jovani R, Donázar JA et al (2014) Bird sky networks: how do avian scavengers use social information to find carrion? Ecology 95:1799–1808
Culloch R (2012) Short note: observations from video footage of red fox (Vulpes vulpes) activity within a grey seal (Halichoerus grypus) breeding colony on the UK Mainland. Aquat Mamm 38:81–85
Curtis TH, Kelly JT, Menard KL et al (2006) Observations on the behavior of white sharks scavenging from a whale carcass at Point Reyes, California. Calif Fish Game 92:113–124
da Diefenbach CO (1975) Gastric function in Caiman crocodilus (Crocodylia: Reptilia)—I. Rate of gastric digestion and gastric motility as a function of temperature. Comp Biochem Physiol A 51(2):259–265
Deinet S, Ieronymidou C, McRae L et al (2013) Wildlife comeback in Europe: the recovery of selected mammal and bird species. Final report to Rewilding Europe by ZSL, BirdLife International and the European Bird Census Council, London
Del Hoyo J (1994) In: Elliott A, Sargatal J (eds) New world vultures to guineafowl. Handbook of the birds of the world, vol 2. Lynx Edicions, Barcelona
DeVault TL, Krochmal AR (2002) Scavenging by snakes: an examination of the literature. Herpetologica 58:429–436
DeVault TL, Rhodes OE (2002) Identification of vertebrate scavengers of small mammal carcasses in a forested landscape. Acta Theriol 47:185–192
DeVault TL, Rhodes OE Jr, Shivik JA (2003) Scavenging by vertebrates: behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 102:225–234
DeVault TL, Brisbin IL, Rhodes OE (2004) Factors influencing the acquisition of rodent carrion by vertebrate scavengers and decomposers. Can J Zool 82:502–509
DeVault TL, Olson ZH, Beasley JC et al (2011) Mesopredators dominate competition for carrion in an agricultural landscape. Basic Appl Ecol 12:268–274
Dicken ML (2008) First observations of young of the year and juvenile great white sharks (Carcharodon carcharias) scavenging from a whale carcass. Mar Freshw Res 59:596–602
Donázar JA, Margalida A, Campión D (eds) (2009) Vultures, feeding stations and sanitary legislation: a conflict and its consequences from the perspective of conservation biology, Munibe 29 (Suppl). Sociedad de Ciencias Aranzadi, San Sebastián
Dudley SF, Anderson-Reade MD, Thompson GS et al (2000) Concurrent scavenging off a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish Bull Ocean Atmos Adm 98:646–649
Dunlop K, Scott M, Parsons D et al (2015) Do agonistic behaviours bias baited remote underwater video surveys of fish? Mar Ecol 36:810–818
Elbroch LM, Wittmer HU (2012) Table scraps: inter-trophic food provisioning by pumas. Biol Lett 8:776–779
Elbroch LM, Wittmer HU (2013) Nuisance ecology: do scavenging condors exact foraging costs on pumas in Patagonia? PLoS ONE 8(1):e53595
Eliotout B (2007) Le vautour fauve. Delachaux et niestlé, Lausanne
Fallows C, Gallagher A, Hammerschlag N (2013) White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS ONE 8(4):e60797
Fortin JK, Farley SD, Rode KD et al (2007) Dietary and spatial overlap between sympatric ursids relative to salmon use. Ursus 18:19–29
Gallagher A, Jackson T, Hammerschlag N (2011) Occurrence of tiger shark (Galeocerdo cuvier) scavenging on avian prey and its possible connection to large-scale bird die-off in the Florida Keys. Fla Sci 4:264–269
Gilchrist HG, Robertson GJ (2000) Observations of marine birds and mammals wintering at polynyas and ice edges in the Belcher Islands, Nunavut, Canada. Arctic 53:61–68
Gjertz I, Lydersen C (1986) Polar bear predation on ringed seals in the fast-ice of Hornsund, Svalbard. Polar Res 4:65–68
Godø O, Huse I, Michalsen K (1997) Bait defence behaviour of wolffish and its impact on long-line catch rates. ICES J Mar Sci 54:272–275
González-Solís J, Croxall JP, Briggs DR (2002) Activity patterns of giant petrels, Macronectes spp., using different foraging strategies. Mar Biol 140:197–204
Green GI, Mattson DJ, Peek JM (1997) Spring feeding on ungulate carcasses by grizzly bears in Yellowstone National Park. J Wildl Manag 61:1040–1055
Green RE, Newton I, Shultz S et al (2004) Diclofenac poisoning as a cause of vulture population declines across the Indian subcontinent. J Appl Ecol 41:793–800
Guarino F (2001) Diet of a large carnivorous lizard, Varanus varius. Wildl Res 28:627–630
Heinrich B (1988) Winter foraging at carcasses by three sympatric corvids, with emphasis on recruitment by the raven, Corvus corax. Behav Ecol Sociobiol 23:141–156
Heinrich B, Marzluff JM, Marzluff CS (1993) Common ravens are attracted by appeasement calls of food discoverers when attacked. Auk 110:247–254
Hertel F (1994) Diversity in body size and feeding morphology within past and present vulture assemblages. Ecology 75:1074–1084
Hertel F, Lehman N (1998) A randomized nearest-neighbor approach for assessment of character displacement: the vulture guild as a model. J Theor Biol 190:51–61
Hilderbrand GV, Schwartz CC, Robbins CT et al (1999) The importance of meat, particularly salmon, to body size, population productivity, and conservation of North American brown bears. Can J Zool 77:132–138
Hiraldo F, Blanco JC, Bustamante J (1991) Unspecialized exploitation of small carcasses by birds. Bird Study 38:200–207
Houston DC (1974) The role of griffon vultures Gyps africanus and Gyps rüpellii as scavengers. J Zool 172:35–46
Houston DC (1975) Ecological isolation of African scavenging birds. Ardea 63:55–64
Houston DB (1978) Elk as winter-spring food for carnivores in northern Yellowstone National Park. J Appl Ecol 15:653–661
Houston DC (1979) The adaptations of scavengers. In: Sinclair ARE, Griffiths MN (eds) Serengeti, dynamics of an ecosystem. University of Chicago Press, Chicago, pp 263–286
Houston DC (1983) The adaptive radiation of griffon vultures. In: Wilbur SR, Jackson JA (eds) Vulture biology and management. University of California Press, Berkeley, pp 135–152
Houston DC (1984) Does the king vulture Sarcoramphus papa use a sense of smell to locate food? Ibis 126:67–69
Houston DC (1986) Scavenging efficiency of turkey vultures in tropical forest. Condor 88:318–323
Houston DC (1988) Competition for food between neotropical vultures in forest. Ibis 130:402–417
Houston DC (2001) Vultures and condors. Clin Baxter Photography Ltd, Grantown-on-Spey
Houston DC, Cooper JE (1975) The digestive tract of the white-back griffon vulture and its role in disease transmission among wild ungulates. J Wildl Dis 11:306–313
Huijbers CM, Schlacher TA, Schoeman DS et al (2013) Urbanisation alters processing of marine carrion on sandy beaches. Landsc Urban Plan 119:1–8
Huijbers CM, Schlacher TA, Schoeman DS et al (2015) Limited functional redundancy in vertebrate scavenger guilds fails to compensate for the loss of raptors from urbanized sandy beaches. Divers Distrib 21:55–63
Hunter S (1985) The role of giant petrels in the Southern Ocean ecosystem. In: Siegfried WR, Laws RM, Condy PR (eds) Antarctic nutrient cycles and food webs. Springer, New York, pp 534–542
Hunter S (1991) The impact of avian predator-scavengers on king penguin Aptenodytes patagonicus chicks at Marion Island. Ibis 133:343–350
Isaacs J, Schwartzlose R (1975) Active animals of the deep-sea floor. Sci Am 233:85–91
Jackson AL, Ruxton GD, Houston DC (2008) The effect of social facilitation on foraging success in vultures: a modelling study. Biol Lett 4:311–313
Jędrzejewska B, Jędrzejewski W (1998) Predation in vertebrate communities. The Białowieża Primeval Forest as a case study. Springer, Berlin
Jędrzejewski W, Zalewski A, Jędrzejewska B (1993) Foraging by pine marten Martes martes in relation to food resources in Białowieża National Park, Poland. Acta Theriol 38:405–426
Jones E, Collins M, Bagley P et al (1998) The fate of cetacean carcasses in the deep sea: observations on consumption rates and succession of scavenging species in the abyssal north-east Atlantic Ocean. Proc R Soc B 265:1119–1127
Kane A, Jackson AL, Ogada DL et al (2014) Vultures acquire information on carcass location from scavenging eagles. Proc R Soc B 281:20141072
Kane A, Healy K, Ruxton GD et al (2016) Body size as a driver of scavenging in theropod dinosaurs. Am Nat 187:706–716
Kelly NE, Sparks DW, DeVault TL et al (2007) Diet of black and turkey vultures in a forested landscape. Wilson J Ornithol 119:267–270
Kendall C (2013) Alternative strategies in avian scavengers: how subordinate species foil the despotic distribution. Behav Ecol Sociobiol 67:383–393
Kendall C, Virani MZ, Kirui P et al (2012) Mechanisms of coexistence in vultures: understanding the patterns of vulture abundance at carcasses in Masai Mara National Reserve, Kenya. Condor 114:523–531
Killengreen ST, Lecomte N, Ehrich E et al (2011) The importance of marine vs. human induced subsidies in the maintenance of an expanding mesocarnivore in the Arctic tundra. J Anim Ecol 80:1049–1060
Killengreen ST, Strømseng E, Yoccoz NG et al (2012) How ecological neighbourhoods influence the structure of the scavenger guild in low arctic tundra. Divers Distrib 18:563–574
Knight SK, Knight RL (1983) Aspects of food finding by wintering bald eagles. Auk 100:477–484
König C (1974) On the behaviour of vultures on carcasses in Spain. J Ornithol 115:289–320
Koskela A, Kojola I, Aspi J et al (2013) Effect of reproductive status on the diet composition of wolverines (Gulo gulo) in boreal forests of eastern Finland. Ann Zool Fenn 50:100–106
Kostecke RM, Linz GM, Bleier WJ (2001) Survival of avian carcasses and photographic evidence of predators and scavengers. J Field Ornithol 72:439–447
Krofel M, Kos I, Jerina K (2012) The noble cats and the big bad scavengers: effects of dominant scavengers on solitary predators. Behav Ecol Sociobiol 66:1297–1304
Kruuk H (1966) Clan-system and feeding habits of spotted hyaenas (Crocuta crocuta Erxleben). Nature 209:1257–1258
Kruuk H (1967) Competition for food between vultures in East Africa. Ardea 55:171–193
Kruuk H (1972) The spotted hyaena. A study of predation and social behavior. University of Chicago Press, Chicago
Kuhn B, Wiesel I, Skinner J (2008) Diet of brown hyaenas (Parahyaena brunnea) on the Namibian coast. Trans R Soc S Afr 63:1–8
Lambertucci SA, Speziale KL, Rogers TE et al (2009a) How do roads affect the habitat use of an assemblage of scavenging raptors? Biodivers Conserv 18:2063–2074
Lambertucci SA, Trejo A, Di Martino S et al (2009b) Spatial and temporal patterns in the diet of the Andean condor: ecological replacement of native fauna by exotic species. Anim Conserv 12:338–345
Lambertucci SA, Navarro J, Sanchez Zapata JA, Hobson KA, Alarcón PAE, Wiemeyer G, Blanco G, Hiraldo F, Donázar JA (2018) Tracking data and retrospective analyses of diet reveal the consequences of loss of marine subsidies for an obligate scavenger, the Andean condor. Proc R Soc B Biol Sci 285(1879):20180550
Levi T, Wheat RE, Allen JM et al (2015) Differential use of salmon by vertebrate consumers: implications for conservation. PeerJ 3:e1157
Linz GM, Bergman DL, Bleier WJ (1997) Estimating survival of song bird carcasses in crops and woodland. Prairie Nat 29:7–13
Løkkeborg S (1998) Feeding behaviour of cod, Gadus morhua: activity rhythm and chemically mediated food search. Anim Behav 56:371–378
Long D, Jones R (1996) White shark predation and scaveging on cetaceans in the Eastern North Pacific Ocean. In: Kimley AP, Ainley DG (eds) Great white sharks: the biology of Carcharodon carcharias. Academic, San Diego, pp 293–307
Lowry LF, Burns JJ, Nelson RR (1987) Polar bear, Ursus maritimus, predation on belugas, Delphinapterus leucas, in the Bering and Chukchi Seas. Can Field Nat 101:141–146
MaMing R, Xu G (2015) Status and threats to vultures in China. Vulture News 68:3–24
MaMing R, Lee L, Yang X, Buzzard P (2016) Vultures and sky burials on the Qinghai-Tibet plateau. Vulture News 71:22–35
Margalida A, Colomer MA (2012) Modelling the effects of sanitary policies on European vulture conservation. Sci Rep 2:753
Margalida A, Heredia R (eds) (2005) Biología de la conservación del quebrantahuesos (Gypaetus barbatus) en España. Organismo Autónomo Parques Nacionales, Madrid
Margalida A, Carrete M, Sánchez-Zapata JA et al (2012) Good news for European vultures. Science 335:284
Margalida A, Bogliani G, Bowden C et al (2014) One health approach to use of pharmaceuticals. Science 346:1296–1298
Margalida A, Colomer MA, Sánchez R et al (2017) Behavioral evidence of hunting and foraging techniques by a top predator suggests the importance of scavenging for pre-adults. Ecol Evol 7:4192–4199
Martin PS (1989) Prehistoric overkill: the global model. In: Martin PS, Klein RG (eds) Quaternary extinctions. University of Arizona Press, Tucson, pp 354–403
Marzluff JM, Heinrich B (1991) Foraging by common ravens in the presence and absence of territory holders: an experimental analysis of social foraging. Anim Behav 42:755–770
Marzluff JM, Heinrich B, Marzluff CS (1996) Raven roosts are mobile information centres. Anim Behav 51:89–103
Mateo-Tomás P, Olea PP, Moleón M et al (2015) From regional to global patterns in vertebrate scavenger communities subsidized by big game hunting. Divers Distrib 21:913–924
Mattisson J, Andrén H, Persson J et al (2011) Influence of intraguild interactions on resource use by wolverines and Eurasian lynx. J Mammal 92:1321–1330
Matyushkin EN (1974) Large carnivores and scavengers of the middle Sikhote-Alin. Bulleten Moskovskogo Obshchestva Ispytatelei Prirody 79:5–21
Moleón M, Sánchez-Zapata JA, Margalida A et al (2014a) Humans and scavengers: the evolution of interactions and ecosystem services. Bioscience 64:394–403
Moleón M, Sánchez-Zapata JA, Selva N et al (2014b) Inter-specific interactions linking predation and scavenging in terrestrial vertebrate assemblages. Biol Rev 89:1042–1054
Moleón M, Sánchez-Zapata JM, Sebastián-González E et al (2015) Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 124:1391–1403
Moreno-Opo R, Margalida A (2013) Carcasses provide resources not exclusively to scavengers: patterns of carrion exploitation by passerine birds. Ecosphere 4:105
Moreno-Opo R, Trujillano A, Arredondo A et al (2015a) Manipulating size, amount and appearance of food inputs to optimize supplementary feeding programs for European vultures. Biol Conserv 181:27–35
Moreno-Opo R, Trujillano A, Margalida A (2015b) Optimization of supplementary feeding programs for European vultures depend on environmental and management factors. Ecosphere 6:127
Moreno-Opo R, Trujillano A, Margalida A (2016) Behavioral coexistence and feeding efficiency drive niche partitioning in European avian scavengers. Behav Ecol 27:1041–1052
Moss B (2017) Marine reptiles, birds and mammals and nutrient transfers among the seas and the land: an appraisal of current knowledge. J Exp Mar Biol Ecol 492:63–80
Novaro AJ, Funes MC, Walker RS (2000) Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biol Conserv 92:25–33
O’Sullivan WM, Sleeman DP, Murphy DM (1992) Otters Lutra lutra feeding on carrion. Ir Nat 24:140–143
Olson ZH, Beasley JC, Rhodes OE (2016) Carcass type affects local scavenger guilds more than habitat connectivity. PLoS ONE 11(2):e0147798
Oro D, Genovart M, Tavecchia G et al (2013) Ecological and evolutionary implications of food subsidies from humans. Ecol Lett 16:1501–1514
Parmenter RR, MacMahon JA (2009) Carrion decomposition and nutrient cycling in a semiarid shrub-steppe ecosystem. Ecol Monogr 79:637–661
Pavés H, Schlatter R, Espinoza C (2008) Scavenging and predation by black vultures Coragyps atratus at a South American sea lion breeding colony. Vulture News 58:4–15
Payne JA (1965) A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology 46:592–602
Payne LX, Moore JW (2006) Mobile scavengers create hotspots of freshwater productivity. Oikos 115:69–80
Pennycuick CJ (1983) Thermal soaring compared in three dissimilar tropical bird species, Fregata magnificens, Pelecanus occidentalis and Coragyps atratus. J Exp Biol 102:307–325
Pereira LM, Owen-Smith N, Moleón M (2014) Facultative predation and scavenging by mammalian carnivores. Mammal Rev 44:44–55
Peterson CA, Lee SL, Elliott JE (2001) Scavenging of waterfowl carcasses by birds in agricultural fields of British Columbia. J Field Ornithol 72:150–159
Petrides GA (1959) Competition for food between five species of East African vultures. Auk 76:104–106
Phillips RA, Phalan B, Forster IP (2004) Diet and long-term changes in population size and productivity of brown skuas Catharacta antarctica lonnbergi at Bird Island, South Georgia. Polar Biol 27:555–561
Platt SG, Rainwater TR, Snider S et al (2007) Consumption of large mammals by Crocodylus moreletii: field observations of necrophagy and interspecific kleptoparasitism. Southwest Nat 52:310–317
Polis GA, Hurd SD (1996) Linking marine and terrestrial food webs: allochthonous input from the ocean supports high secondary productivity on small islands and coastal land communities. Am Nat 147:396–423
Prior KA, Weatherhead PJ (1991) Competition at the carcass: opportunities for social foraging by turkey vultures in southern Ontario. Can J Zool 69:1550–1556
Quaggiotto M-M, Barton PS, Morris C et al (2018) Seal carrion is a predictable resource for coastal ecosystems. Acta Oecol 88:41–51
Raichev EG, Tsunoda H, Newman C et al (2013) The reliance of the golden jackal (Canis aureus) on anthropogenic foods in winter in central Bulgaria. Mamm Study 38:19–27
Ray R–R, Seibold H, Heurich M (2014) Invertebrates outcompete vertebrate facultative scavengers in simulated lynx kills in the Bavarian Forest National Park, Germany. Anim Biodivers Conserv 37(1):77–88
Read JL, Wilson D (2004) Scavengers and detritivores of kangaroo harvest offcuts in arid Australia. Wildl Res 31:51–56
Rogers AM, Gibson MR, Pockette T et al (2014) Scavenging of migratory bird carcasses in the Sonoran desert. Southwest Nat 59:544–549
Rohr RP, Saavedra S, Bascompte J (2014) On the structural stability of mutualistic systems. Science 345:1253497
Ronconi RA, Steenweg RJ, Taylor PD et al (2014) Gull diets reveal dietary partitioning and ecosystem changes at a remote colony. Mar Ecol Prog Ser 514:247–261
Rose M, Polis G (1998) The distribution and abundance of coyotes: the effects of allochthonous food subsidies from the sea. Ecology 79:998–1007
Roth JD (2002) Temporal variability in arctic fox diet as reflected in stable carbon isotopes; the importance of sea ice. Oecologia 133:70–77
Ruxton GD, Houston DC (2003) Could Tyrannosaurus rex have been a scavenger rather than a predator? An energetics approach. Proc R Soc Lond B 270:731–733
Ruxton GD, Houston DC (2004) Obligate vertebrate scavengers must be large soaring fliers. J Theor Biol 228:431–436
Samelius G, Alisauskas RT, Hobson KA et al (2007) Prolonging the arctic pulse: long-term exploitation of cached eggs by Arctic foxes when lemmings are scarce. J Anim Ecol 76:873–880
Sánchez R, Margalida A, González LM et al (2009) Temporal and spatial differences in the feeding ecology of the Spanish imperial eagle Aquila adalberti during the non-breeding season: effects of the rabbit population crash. Acta Ornithol 44:53–58
Sánchez-Zapata JA, Eguía S, Blázquez M et al (2010) Unexpected role of ungulate carcasses in the diet of golden eagles Aquila chrysaetos in Mediterranean mountains. Bird Study 57:352–360
Schindler DE, Armstrong JB, Bentley KT et al (2013) Riding the crimson tide: mobile terrestrial consumers track phenological variation in spawning of an anadromous fish. Biol Lett 9:20130048
Sebastián-González E, Sánchez-Zapata JA, Donázar JA et al (2013) Interactive effects of obligate scavengers and scavenger community richness on lagomorph carcass consumption patterns. Ibis 155:881–885
Sebastián-González E, Moleón M, Gibert JP et al (2016) Nested species-rich networks of scavenging vertebrates support high levels of interspecific competition. Ecology 97:95–105
Selva N (2004) The role of scavenging in the predator community of Białowieża Primeval Forest (E Poland). PhD thesis, University of Sevilla, Spain
Selva N, Fortuna MA (2007) The nested structure of a scavenger community. Proc R Soc B 274:1101–1108
Selva N, Jędrzejewska B, Jędrzejewski W et al (2003) Scavenging on European bison carcasses in Białowieża Primeval Forest (eastern Poland). Ecoscience 10:303–311
Selva N, Jedrzejewska B, Jedrzejewski W et al (2005) Factors affecting carcass use by a guild of scavengers in European temperate woodland. Can J Zool 83:1590–1601
Shepard EL, Lambertucci SA (2013) From daily movements to population distributions: weather affects competitive ability in a guild of soaring birds. J R Soc Interface 10:20130612
Shivik JA (2006) Are vultures birds, and do snakes have venom, because of macro- and microscavenger conflict? Bioscience 56:819–823
Skinner JD, Chimimba CT (2005) The mammals of the Southern African subregion. Cambridge University Press, Cambridge
Smith CR, Baco AR (2003) Ecology of whale falls at the deep-sea floor. Oceanogr Mar Biol 41:311–354
Spiller DA, Piovia-Scorr J, Wright AN et al (2010) Marine subsidies have multiple effects on coastal food webs. Ecology 91:1424–1434
Stahler D, Heinrich B, Smith D (2002) Common ravens, Corvus corax, preferentially associate with grey wolves, Canis lupus, as a foraging strategy in winter. Anim Behav 64:283–290
Stirling I, Archibald WR (1977) Aspects of predation of seals by polar bears. J Fish Res Board Can 34:1126–1129
Stoddart LC (1970) A telemetric method for detecting jackrabbit mortality. J Wildl Manag 34:501–507
Stoner A (2004) Effects of environmental variables on fish feeding ecology: implications for the performance of baited fishing gear and stock assessment. J Fish Biol 65:1445–1471
Swihart RK, Gehring TM, Kolozsvary MB et al (2003) Response of ‘resistant’ vertebrates to habitat loss and fragmentation: the importance of niche breadth and range boundaries. Divers Distrib 9:1–18
Tenney S (1877) A few words about scavengers. Am Nat 11:129–135
Terrasse JF (2001) Le gypaète barbu. Delachaux et niestlé, Lausanne
Tobin ME, Dolbeer RA (1990) Disappearance and recoverability of songbird carcasses in fruit orchards. J Field Ornithol 61:237–242
Travaini A, Donazar JA, Rodriguez A et al (1998) Use of European hare (Lepus europaeus) carcasses by an avian scavenging assemblage in Patagonia. J Zool 246:175–181
Twiss SD, Duck C, Pomeroy PP (2003) Grey seal (Halichoerus grypus) pup mortality not explained by local breeding density on North Rona, Scotland. J Zool 259:83–91
Vogeley W (1999) Use of carcasses by Cape griffons Gyps coprotheres and food competition – observations at a vulture restaurant in south-eastern Botswana. Ornithol Beobachter 96:13–23
Wallace MP, Temple SA (1987) Competitive interactions within and between speies in a guild of avian scavengers. Auk 104:290–295
Waters CN, Zalasiewicz J, Summerhayes C et al (2016) The Anthropocene is functionally and stratigraphically distinct from the Holocene. Science 351:aad2622
Weldon PJ, Ferguson MWJ (1993) Chemoreception in crocodilians: anatomy, natural history, and empirical results. Brain Behav Evol 41:239–245
Weldon PJ, Swanson DJ, Olson JK et al (1990) The American alligator detects food chemicals in aquatic and terrestrial environments. Ethology 85:191–198
White C (2005) Hunters ring dinner bell for ravens: experimental evidence of a unique foraging strategy. Ecology 86:1057–1060
Whitehead H, Reeves R (2005) Killer whales and whaling: the scavenging hypothesis. Biol Lett 1:415–418
Wikenros C, Sand H, Ahlqvist P et al (2013) Biomass flow and scavengers use of carcasses after re-colonization of an apex predator. PLoS ONE 8(10):e77373
Williams AJ, Dyer BM, Randall RM, Komen J (1990) Killer whales Orcinus orca and seabirds “play” predation and association. Mar Ornithol 18:37–41
Wilmers CC, Stahler DR, Crabtree RL et al (2003a) Resource dispersion and consumer dominance: scavenging at wolf- and hunter- killed carcasses in Greater Yellowstone, USA. Ecol Lett 6:996–1003
Wilmers CC, Crabtree RL, Smith DW et al (2003b) Trophic facilitation by introduced top predators: grey wolf subsidies to scavengers in Yellowstone National Park. J Anim Ecol 72:909–916
Wilson R, Smith K (1984) Effect of near-bottom currents on detection of bait by the 4 abyssal grenadier fishes Coryphaenoides spp., recorded in situ with a video 5 camera on a free fall vehicle. Mar Biol 84:83–91
Wilson EE, Wolkovich EM (2011) Scavenging: how carnivores and carrion structure communities. Trends Ecol Evol 26:129–135
Worthen WB, Jones MT, Jettojn RM (1998) Community structure and environmental stress: desiccation promotes nestedness in mycophagous fly communities. Oikos 81:45–54
Wright J, Stone RE, Brown N (2003) Communal roosts as structured information centers in the raven, Corvus corax. J Anim Ecol 72:1003–1014
Yeh J, Drazen J (2011) Baited-camera observations of deep-sea megafaunal scavenger ecology on the California slope. Mar Ecol Prog Ser 424:145–156
Acknowledgements
The authors sincerely thank Alexander Gruzdev, Dan Hartman, Sarah C. Jones, Lama Rinczen and Manuel de la Riva for their pictures.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Selva, N. et al. (2019). Vertebrate Scavenging Communities. In: Olea, P., Mateo-Tomás, P., Sánchez-Zapata, J. (eds) Carrion Ecology and Management. Wildlife Research Monographs, vol 2. Springer, Cham. https://doi.org/10.1007/978-3-030-16501-7_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-16501-7_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-16499-7
Online ISBN: 978-3-030-16501-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)