
Abstract
The nutritional value of food resources represents one of the crucial facets of habitat quality that has to be investigated to predict primate persistence in forest fragments. Mantled howler monkeys (Alouatta palliata) are known to be tolerant to habitat fragmentation, although the boundaries of this flexibility are not fully understood. In some fragmented areas, howler monkey densities are among the highest ever recorded for a primate species. As a possible consequence of high competition and resource depletion over-crowding effects are predicted to alter the time-budget and decrease the diet quality of howler monkeys living in small fragments. To test this, we studied the feeding ecology of four howler monkey groups at La Suerte Biological Field Station, North-eastern Costa Rica, over three consecutive summers. Two groups occurred in a 20-ha forest fragment at high density, while the other two groups lived in a larger forest patch at lower density. We collected behavioural data via 5-min Focal Instantaneous Sampling to estimate time-budget, habitat use and diet. Food samples were also collected and analysed to evaluate nutritional contents. Ranging areas (Kernel analysis) and daily path-length were estimated via Ranges 8. Overall, our results show that the nutritional quality of the monkeys' diet in the high-density area was not significantly different than that recorded in the less populated fragment. Although the monkeys in the high-density forest did not alter their time-budget substantially they used much smaller home-ranges. Our results do not support the hypothesis that the monkeys living in the high-density forest are experiencing a drop in diet quality. Differences in forest structure between the two fragments may explain the similarities between the nutritional ecology of the groups.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
22.1 Introduction
Habitat fragmentation and its consequences are currently the main threat to primate conservation (Cowlishaw and Dunbar 2000), and to biodiversity worldwide (Laurance and Peres 2006). This phenomenon, caused primarily by human population growth, leads to archipelagos of habitats, which in turn implies reduction and isolation of wildlife populations (Malcolm 1997; Robinson 1996). The increased perimeter to area ratio of fragments also causes easier human exploitation of forests (e.g. logging, hunting), and a decrease of suitable habitat for species unable to persist in edge environments (Laurance et al. 2002). Populations living in fragments are exposed to genetic flow reduction that causes genetic erosion and makes them more vulnerable (Lande and Barrowclough 1987; Spielman et al. 2004).
In order to better estimate ecological viability and to develop effective management actions, it is essential to identify habitat features that determine species persistence in fragmented habitats. Island biogeography factors such as fragment size, age, and isolation have been often identified as strong predictors of species persistence (Whittaker and Fernandez-Palacios 2007). However, flexible species able to reduce or expand home ranges, move through the matrix, and/or adjust their diet are likely to persist in areas no longer identified as suitable by models (Estrada and Coates-Estrada 1996; Lovejoy et al. 1986; Onderdonk and Chapman 2000). In primates, for example, fragment size and isolation have not been always found to be good predictors of species presence/absence . In fact, while in Madagascar these factors seem to govern primate persistence (Ganzhorn et al. 2000), the same is not true for other areas (Onderdonk and Chapman 2000). Primates density as well, which is strongly affected by habitat degradation (Bowers and Matter 1997), does not always relate to obvious fragment features (Peres 1997).
The howler monkeys, genus Alouatta, which includes 14 species (International Union for Conservation of Nature (IUCN) 2012), are generally considered very flexible (Lovejoy et al. 1986; Bicca-Marques 2003; Garber et al. 2006). These monkeys often occur in fragments where other primates (e.g. Ateles sp.) cannot persist (Estrada and Coates-Estrada 1996; Gilbert 2003; Arroyo-Rodrìguez and Dias 2010). In particular, howler monkeys’ persistence in fragmented habitats seems to be based on their ability to adapt diet to food availability (Onderdonk and Chapman 2000; Bicca-Marques 2003; Rivera and Calmè 2006), increase leaf intake (Rodrìguez-Luna et al. 2003; Asensio et al. 2007), consume secondary or exotic plant species (Onderdonk and Chapman 2000), and also epiphytic or parasitic plants (Rodrìguez-Luna et al. 2003; Asensio et al. 2007). However, they are also very selective with regard to food resources and the monthly diversity of their diet is generally low, with only few plant species regularly included (Estrada et al. 1999). Food selection is based on nutritional contents, vegetative stage, and secondary compounds (Glander 1982), and it is vital in fragments where preferred food resources are scarce or scattered (Chiarello 2003; Juan et al. 2000). Howler monkeys are also able to rearrange their ranging pattern (Estrada and Coates-Estrada 1996; Onderdonk and Chapman 2000), and their time budget (Juan et al. 2000; Silver and Marsh 2003; Cristòbal-Azkarate and Arroyo-Rodrìguez 2007). Finally, in small or degraded fragments they can even use a fission–fusion feeding strategy to decrease intra-group feeding competition (Leighton and Leighton 1982; Asensio et al. 2007).
The ecology of Alouatta has been comprehensively studied, although the factors predicting their persistence in different forest fragments are not clear yet (Arroyo-Rodrìguez and Dias 2010). Individual density may increase at the initial stages of the fragmentation process due to predator disappearance (Estrada et al. 2002; Rosales-Meda et al. 2007), but this is also dependent on the quality and availability of food resources (Nagy and Milton 1979). In some areas, highly productive secondary vegetation seems to favour high-density populations, although this may cause resource depletion in the long term, leading to possible local extinctions in small fragments (Lovejoy et al. 1986; Arroyo-Rodrìguez and Dias 2010).
This study aims at investigating the consequences of high population density on the activity budget, ranging pattern, and nutritional ecology of the mantled howler monkey (Alouatta palliata) population inhabiting two forest fragments at La Suerte Biological Field Station, north-eastern Costa Rica. The two forest fragments are connected by a narrow corridor but differ in terms of size and forest history. A. palliata individual density in the smaller of those fragments is very high compared to the other area (150 vs. 57 ind/km2; Adams et al., unpublished) and to the species average (Campbell et al. 2011). Longitudinal demographic data do not show a population decline over the last decade (Pruetz and Leasor 2002; Adams et al., unpublished). We collected eco-ethological data and food samples on two groups of howler monkeys in each forest fragment over three summers (July–August) from 2009 to 2011. We hypothesize that if the small fragment is overcrowded, this would cause high competition and resource depletion, altering the time budget, and decreasing the quality of the diet of howler monkeys compared to animals living in the non-crowded forest. To test this hypothesis, we analysed behavioural, nutritional, and ranging data.
22.2 Methods
22.2.1 Study Site
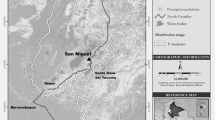
The study site was located in north-eastern Costa Rica, at La Suerte Biological Field Station (10° 26′ 15″ N; 86° 43′ 30″ W; Fig. 22.1). The site is 50 m above mean sea level and is covered by a lowland rainforest, with annual rainfall around 3900 mm and mean temperature of 24 °C (Bezanson et al. 2008). It includes two forest fragments, which are embedded in a matrix of abandoned fields, grazed pastures, and coconut plantations. La Suerte River connects both fragments, functioning as a corridor of riparian forest. The smaller forest (SF) fragment is about 20 ha and is one of the remaining stands of primary vegetation in the area. The larger forest (LF) fragment, to the north, covers 250 ha of secondary forest that was exploited for logging and grazing until the late seventies (Renee Molina, personal communication). Three primate species inhabit the area: mantled howler monkey (A. palliata), white-headed capuchin (Cebus capucinus), and Geoffroy’s spider monkey (Ateles geoffroyi). According to Adams et al. (unpublished), the two fragments are structurally similar but the SF has greater tree species diversity and larger crown volume of emergent trees. Howler monkeys’ density is considerably higher in the SF, 150 ind/km2 in 1999 and 2010, than in the LF, 30 ind/km2 in 1999 and 57 ind/km2 in 2010 (Adams et al., unpublished; Pruetz and Leasor, 2002).

Study site
22.2.2 Study Species
A. palliata is the second largest neotropical primate (Zingeser 1973), and its distribution encompasses all the Central American forests (Estrada and Coates-Estrada 1994). This monkey is diurnal, and largely arboreal, exploiting the higher canopy layers (Silva and Da1981), where it feeds mainly on leaves and fruits (less on flowers, buds, petioles, etc.; Milton 1980). Despite the high-fibre diet, it does not possess anatomical or physiological specializations to maximize cellulose digestion (Chivers and Hladik 1980). On average, A. palliata groups are composed of 3–4 males, 7–10 females, and the number of offspring and subadults equals to females number (Carpenter 1934). Its activity pattern is characterised by long resting period interrupted essentially by feeding/foraging bouts (Smith 1977).
22.2.3 Behavioural, Nutritional, and Ranging Data
Behavioural data were collected via a 5-min focal instantaneous sampling (Altmann 1974) during July and August 2009, 2010, and 2011. Two groups were studied in each forest fragment, and each group was observed between 3 and 4 consecutive days per season from dawn to dusk. A total of 480 observation hours were collected. The ethogram included moving (moving on the same tree and travelling), feeding (all the actions related to food processing), resting, and other (social behaviours, self-grooming, scratching, stretching, drinking, defecating, and urinating). Only adult individuals were observed and used to estimate daily activity budget. Data collection also included posture, food item, feeding (or resting) tree’s code and global positioning system (GPS) coordinates. Feeding trees were marked and identified to species level on a following day with the aid of a local botanist. Samples of food items (leaves or fruits) eaten during behavioural observations were also collected from the same tree where the monkeys were observed eating. GPS coordinates were recorded every 30 min and analysed via RANGES 8 to estimate the home range size (via Kernel analysis; Worton 1989) and the daily path length.
22.2.4 Biochemical Analyses
Food samples were dried in the sun at La Suerte, and later analysed for their nutritional content at the laboratory of the Department of Veterinary Science, University of Pisa, and at the Department of Animal Ecology and Conservation, University of Hamburg following the methods outlined in Donati et al. (2011). Dry matter was used to determine the amount of the following nutritional compounds: crude protein (CP), crude fat (CF), neutral detergent fibre (NDF), acid detergent fibre (ADF), ashes (Donati et al. 2007). Nonstructural carbohydrates (NSCs) were assessed with the formula: NSC = 100-(%CP + %CF + %NDF + %ashes)(Milton 2008; Norconk et al. 2009). Metabolizable energy (ME) was estimated according to Conklin and Wrangham (1994) as follows: ME (kcal/100 g d.m.) = (4× CP×0.89) + (4×NDF×0.41) + (4×NSC) + (9×CF). Dried food samples collected in 2009 were also analysed with atomic absorption spectroscopy in order to get the concentration of mineral elements, which included macroelements (calcium, phosphorus, magnesium, sodium) and microelements (iron, copper, manganese, zinc; Williams-Guillèn 2003).
22.2.5 Data Analyses
Daily percentages of feeding, moving, and resting as well as daily path lengths of each group of monkeys were averaged per fragment and compared using the Mann–Whitney nonparametric U test. The diet diversity of each group was evaluated with the Shannon’s diversity index and averaged per fragment. The comparisons of the nutritional contents of the food items eaten in each fragment were also undertaken via the U test. For the comparisons, we showed both the difference between the food samples per se and the difference between the weighted diets using the proportion of feeding time as coefficient.
22.3 Results
22.3.1 Activity budget
At the La Suerte Biological Station, resting was the most represented activity in howler monkeys, consisting in 65.67 % ± 1.49 (n = 33 days) of the daily activity budget, while feeding and moving were the other main activities, 12.27 ± 1.71 and 18.52 ± 1.80, respectively. There was no significant difference in the monkey time budget between the two forest fragments (Table 22.1).
22.3.2 Ranging Pattern
The mean home range of SF groups was considerably smaller than the areas used by the monkeys in the LF (Fig. 22.2a). As for daily path length (Fig. 22.2b), SF groups moved significantly less than LF groups (U = 30.0, p = 0.02, n = 11 days SF, n = 13 days LF).

Ranging patterns: Comparison among mean home ranges (a) and mean daily path length (b) of the howler monkey groups in the two forest fragments
22.3.3 Nutritional quality of the diet
We collected a total of 28 different food samples during behavioural observations; 12 from the SF and 16 from the LF. The Shannon diversity index (H’) for the diet of the two groups did not differ appreciably (SF groups: 0.90; LF groups: 0.87).
Despite food samples collected in the SF contained substantially lower values of fibre fractions (Table 22.2), statistical analyses did not show any significant difference. The weighted diet (Table 22.3) did not show any significant difference as well between the dietary regime of the monkeys in the two forest fragments.
The comparison between the two fragments in terms of mineral content of food items showed that in SF samples calcium values were almost double than those from the LF, as supported by a strong statistical tendency, while phosphorus and magnesium showed similar values in both fragments (Table 22.4). Overall food samples of howler monkeys contained very low amounts of sodium. Microelements’ contents were similar between the samples of the two fragments, except for manganese, which was significantly higher in the LF samples (Table 22.4).
22.4 Discussion
Overall, the howler monkeys’ groups living in the two forest fragments of La Suerte did not differ in terms of activity and diet, while their ranging pattern was found to be substantially dissimilar. The activity budget of the groups in La Suerte is not only similar between the two fragments but also to the values reported in more extensive behavioural studies on this species (Carpenter 1934; Milton 1980; Williams-Guillèn 2003). Inactivity was prevailing, with no differences among the groups. Regarding the other two main activities, feeding and moving, there were no significant differences as well. Milton (1998) suggested that high values of inactivity in Alouatta are the consequence of large quantity of leaves passing through the intestinal tract and slowing down digestion due to cellulosic material. Other studies do not support this hypothesis and indicate that inactivity is rather a phylogenetic trait of the genus, independent from the diet (Pavelka and Knopff 2004). In La Suerte, inactivity was not correlated to obvious forest/fragment features or monkey density, in line with previous studies (Cristobal-Azkarate and Arroyo-Rodrìguez 2007).
In contrast to time budget, ranging patterns showed large differences between the howler monkey groups. In particular, the monkeys in the LF had larger home ranges and travelled further. This is in agreement with previous reports showing that ranging size is affected by fragment size (Cristobal-Azkarate and Arroyo-Rodrìguez 2007). It is possible that in the high-density SF howler monkey groups moved less to avoid interactions with conspecific groups. In contrast, in the LF the howler groups did not simply move further but their size was smaller than in the SF, as a consequence of a more obvious fission–fusion strategy (Occhibove 2010). This behaviour is typically used to reduce intra-group competition when resources are limited (Arroyo-Rodrìguez and Dias 2010). Thus, the behavioural evidence seems to indicate, contrary to our expectation, that in the LF preferred food resources may have been more limited. The structure of the two forest fragments may help to explain this finding. In fact, vegetation plots indicate that the LF had low floristic diversity and smaller average food tree size compared to the SF, which may make the former area less suitable for the howler monkeys and may limit its carrying capacity (Adams et al., unpublished).
According to Chapman et al. (2002), folivorous primate biomass is strongly related to the protein-to-fibre ratio of the leaves used as food. Although the biological relevance of these measures is a matter of debate (DeGabriel et al. 2013; Wallis et al. 2012), the index has been used to evaluate food quality of herbivorous, since nitrogen is considered a limiting nutrient in many terrestrial ecosystems (White 1993). In fact, monkeys often prefer leaves with higher protein and low fibre ratios (Chapman et al. 2002; Ganzhorn 2002; Hanya and Bernard 2012; Milton 1979) also because fibres require microbial fermentation and contain components (e.g. cellulose and hemicellulose) that are only partially digestible (McNab 2002). In La Suerte, nutritional data indicated that NDF and ADF were higher in food samples from LF groups, with a clear statistical trend. Nevertheless, CP/NDF and CP/ADF ratios were not significantly different between the food samples from the two fragments. The monkey-weighted diet echoed the results from food samples with even slighter nutritional differences, suggesting that animal feeding has further compensated for the variation in nutrient availability between the two fragments. Thus, overall the monkeys living in the two fragments did not show major difference in terms of macronutrient intake. It is important to mention, however, that analyses based on feeding time can yield biased results and they do not always correlate with actual intake, due to the differences in food size, density, and processing time (Rothman et al. 2011). However, feeding time has been shown to provide acceptable proxies of actual food intake in case of leaves’ consumption (Kurland and Gulin 1987).
We used the National Research Council (NRC) (2003) tables for nonhuman primates to examine the nutritional composition of the food samples in relation to the monkeys’ nutritional requirements. NDF and ADF percentages of all the samples were higher than the minimum suggested to keep gastrointestinal health (10–30 % and 5–15 %, respectively). The proportion of protein, in all samples, was in line with the lower limit of the range suggested by the NRC (2003). Milton (1998) suggested that leaves eaten by howlers have to contain at least 11 % of CP (on dry matter) to cover their needs. Our food samples were all above this value.
Although primates prioritize proteins as source of energy (Felton et al. 2009b; Rothman et al. 2011), overall they do not require high proteinaceous diets (Oftedal 1992). Mature leaves, basically the only food category consumed in both fragments in La Suerte during our study period, were found to be the main source of energy and to contain more sugars than young leaves (Occhibove 2010). Mature leaves, however, also contained higher fibre and lower protein concentrations than young leaves, thus making in principle this food less palatable (Behie and Pavelka 2012). Behie and Pavelka (2012) propose that food selection in folivorous primates may be driven by energy intake rather than protein intake maximization, since the protein amount is above the minimum requirements in both mature and young leaves. This idea is supported by similar results in a gorilla population at Bwindi National Park, Uganda (Ganas et al. 2009). Our samples of leaves from La Suerte were, in fact, well above the minimum protein requirements and contained comparable sugars in both fragments. This seems to suggest that in the high-density area, howler monkeys do not feed on fallback food resources, despite the predicted potential high inter and intra-group competition.
The food mineral content was very similar between the two forest fragments, except for calcium and manganese, more represented in the SF and LF samples, respectively. It is well documented that minerals are fundamental to the health (Robbins 1993) and mineral availability has been proposed as a limiting factor to population growth of frugivorous primates (Rode et al. 2006). However, little is known regarding the role of minerals in primate diet selection (Felton et al. 2009a). In fact, few authors support the idea that minerals influence primate diet selection, partly because minerals can be obtained from nonfood sources (e.g. salt licks, soil, eggshells). Moreover, some studies indicate that mineral intake of wild primates eating natural diets exceeds established requirements of humans and deficiencies seem unlikely (Milton 2003; Rothman et al. 2006). Our preliminary results from La Suerte indicated that dietary calcium may show large differences even between contiguous areas. Previous work suggested that some mature leaves were consumed more than young leaves as important sources of minerals , especially of calcium, iron, and manganese (Silver et al. 2000, 2003). Felton et al. (2009b) also found that spider monkeys (Ateles chamek) ate large amounts of figs probably because they contained high concentrations of minerals, especially available calcium (O’Brien et al. 1998; Silver et al. 2000), which is crucial for maintenance and reproduction (Robbins 1993). Data on a larger sample of food collected over a longer period of time are necessary to properly test the potential role of mineral availability in La Suerte.
Looking at the NRC tables (2003) for mineral requirements in primates, the leaves from the SF showed an amount of calcium far higher than the recommended value, while in the LF samples the values were comparable. Similarly to Williams-Guillèn (2003), phosphorus concentration was found to be half of the value indicated in the NRC (2003). In both fragments, magnesium and iron were higher than the recommended value of the NRC (2003), suggesting that howlers’ requirements were met for these elements. According to previous studies on the nutritional ecology of wild A. palliata (Williams-Guillèn 2003) and to the nutritional content of the most common food of other herbivorous (e.g. forage), sodium concentration was very low, that is four to five times lower than the suggested values by the NRC (2003). This seems to be usual for folivorous primates and it is not clear what the implications of it are. In contrast, manganese concentration in both fragments was much higher than the NRC (2003) recommended value, especially in the LF where it exceeded by around 20 times the proposed concentration. This amount may cause neurologic toxicity (Burton and Guilarte 2009). Also Williams-Guillèn (2003) in Nicaraguan A. palliata found higher amounts of manganese than the NRC (2003), but we found much higher values in La Suerte. In fact, hyper-abundance of some elements rather than deficiency may also play a role in determining the distribution of these monkeys and more research needs to be focussed on mineral elements.
In conclusion, in both forest fragments of the La Suerte Biological Station, howler monkeys seem able to reach their nutritional requirements, adjusting their feeding strategy and ranging pattern to the size and resource availability in each area. No sign of major diet quality drop due to monkey high density and possible overcrowding were identified in this study. This demonstrates the excellent ability of A. palliata to cope with different environments. Nevertheless, the long-term resource depletion due to overcrowding in the SF may still represent a risk (Estrada et al. 1999; Pruetz and Leasor 2002). Therefore, it is necessary to improve the connection between the fragments in order to minimize this risk and maximize gene flow between the two small populations. It is also important to note, however, that our data covers only 2 months per year, and this may lead to biased conclusions, since food availability varies seasonally and supra-annually (Cant 1980). More detailed investigations over a full year are needed to get a clear picture of the demographic and ecological constraints for A. palliata in La Suerte.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behavior 49:227–267
Arroyo-Rodríguez V, Dias PA (2010) Effects of habitat fragmentation and disturbance on howler monkeys: a review. Am J Primatol 72:1–16
Asensio N, Cristobal-Azkarate J, Dias PA, Vea JJ, Rodriguez-Luna E (2007) Foraging habits of Alouatta palliata mexicana in three forest fragments. Folia Primatol 78:141–153
Behie AM, Pavelka MSM (2012) Food selection in the black howler monkey following habitat disturbance: implications for the importance of mature leaves. J Trop Ecol 28:153–160
Bezanson M, Garber PA, Murphy JT, Premo LS (2008) Patterns of subgrouping and spatial affiliation in a community of mantled howling monkeys (Alouatta palliata). Am J Primatol 70:282–293
Bicca-Marques JC (2003) How do howler monkeys cope with habitat fragmentation? In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer/Plenum, New York. pp 283–303
Bowers MA, Matter SF (1997) Landscape ecology of mammals: relationships between density and patch size. J Mammal 78:999–1013
Burton NC, Guilarte TR (2009) Manganese neurotoxicity: lessons learned from longitudinal studies in nonhuman primates. Environ Health Perspectives 117:325–332
Campbell CJ, Fuentes A, MacKinnon KC (2011) Primates in perspectives. Oxford University Press, Oxford Cant JH (1980) What limits primates? Primates 21(4):538–544
Carpenter C R (1934) A field study of the behavior and social relations of howling monkeys (Alouatta palliata). Comp Psychol Monogr 10: 1–168
Chapman CA, Chapman LJ, Bjorndal KA, Onderdonk DA (2002) Application of protein-to-fiber ratios to predict colobine abundance on different spatial scales. Int J of Primatol 23:283–310
Chiarello AG (2003) Primates in the Brazilian Atlantic Forest: the influence of forest fragmentation on survival. In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer/Plenum, New York. pp 99–121
Chivers DJ, Hladik CM (1980) Morphology of the gastrointestinal tract in primates: comparisons with other mammals in relation to diet. J Morphol 166:337–86
Conklin NL, Wrangham RW (1994) The value of figs to a hindgut fermenting frugivores: a nutritional analysis. Biochem Syst Ecol 22:137–151
Cowlishaw G, Dunbar R (2000) Primate conservation biology. University of Chicago Press, Chicago
Cristóbal-Azkarate J, Arroyo-Rodríguez V (2007) Diet and activity pattern of howler monkeys (Alouatta palliata) in Los Tuxtlas, Mexico: effects of habitat fragmentation and implications for conservation. Am J Primatol 69:1013–1029
DeGabriel JL, Moore BD, Felton AM, Ganzhorn JU, Stolter C, Wallis IR, Johnson CN, Foley, WJ (2013) Translating nutritional ecology from the laboratory to the field: milestones in linking plant chemistry to population regulation in mammalian browsers. Oikos 123:298–308
Donati G, Bollen A, Borgognini-Tarli SM, Ganzhorn JU (2007) Feeding over the 24-h cycle: dietary flexibility of cathemeral collared lemurs (Eulemur collaris). Behav Ecol Sociobiol 61:1237–1251 Donati G, Kesch K, Ndremifidy K, Schmidt SL, Ramanamanjato JB, Borgognini SM, Ganzhorn JU (2011) Better few than hungry: flexible feeding ecology of collared lemurs Eulemur collaris in littoral forest fragments. PLoS One 6(5):e19807. doi:10.1371/journal.pone.0019807
Estrada A, Coates-Estrada R (1994) Non flying mammals and landscapes changes in the tropical rain forest region of Los Tuxtlas, Mexico. Ecography 17:229–241
Estrada A, Coates-Estrada R (1996) Tropical rain forest fragmentation and wild populations of primates at Los Tuxtlas, Mexico. Int J Primatol 17:759–783
Estrada A, Juan-Solano S, Ortiz Martinez T, Coates-Estrada R (1999) Feeding and general activity pattern of a howler monkey (Alouatta palliata) troop living in a forest fragment at Los Tuxtlas, Mexico. Am J Primatol 48:167–183
Estrada A, Mendoza A, Castellanos L, Pacheco R, Van Belle S, Garcia Y, Munoz D (2002) Population of the black howler monkey (Alouatta pigra) in a fragmented landscapes in Palenque, Chiapas, Mexico. Am J Primatol 58:45–55
Felton AM, Felton A, Lindenmayer DB, Foley WJ (2009a) Nutritional goals of wild primates. Funct Ecol 23:70–78
Felton AM, Felton A, Raubenheimer D, Simpson SJ, Foley WJ, Wood JT, Wallis IR, Lindenmayer DB (2009b) Protein content of diets dictates the daily energy intake of a free-ranging primate. Behav Ecol 20:685–690
Ganas J, Ortmann S, Robbins MM (2009) Food choices of the mountain gorilla in Bwindi Impenetrable National Park, Uganda: the influence of nutrients, phenolics and availability. J Trop Ecol 25:125–134
Ganzhorn J U (2002) Distribution of a folivorous lemur in relation to seasonally varying food resources: integrating quantitative and qualitative aspects of food characteristics. Oecologia 131: 427–435.
Garber PA, Estrada A, Pavelka MS (2006) New perspectives in the study of Mesoamerican primates: concluding comments and conservation priorities. In: Estrada A, Garber PA, Pavelka MS, Luecke L (eds) New perspectives in the study of Mesoamerican primates: distribution, ecology, behavior, and conservation. Springer, New York. pp 563–584
Gilbert KA (2003) Primates and fragmentation of the Amazon forest. In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer/Plenum, New York. pp 145–157
Glander KE (1982) The impact of plant secondary compounds on primate feeding behavior. Am J Phys Anthropol 25:1–18
Hanya G, Bernard H (2012) Fallback foods of red leaf monkeys (Presbytis rubicunda) in Danum Valley, Borneo. Int J Primatol 33:322–337
IUCN (2012) IUCN Red List of Threatened Species. Version 2012.2. www.iucnredlist.org. Downloaded on 13 March 2013
Juan S, Estrada A, Coates-Estrada R (2000) Contrastes y similitudines en el uso de recursos y patròn general de actividades en tropas de monos aulladores (Alouatta palliata) en fragmentos de selva de Los Tuxtlas, Mexico. Neotrop Primates 8:131–135
Kurland JA, Gaulin SJC (1987) Comparability among measures of primate diets. Primates 28:71–77
Lande R, Barrowclough GF (1987) Effective population size, genetic variation, and their use in population management. In: Lande R, Barrowclough GF, Soulé ME (eds) Viable populations for conservation. Cambridge University Press, Cambridge. pp 87–123
Laurance WF, Peres CA (2006) Emerging threats to tropical forests. University of Chicago Press, Chicago
Laurance WF, Lovejoy TE, Vasconcelos HL, Bruna EM, Dirham RK, Stoufer PC (2002) Ecosystem decay of Amazonian forest fragments: a 22 year investigation. Conserv Biol 16:605–618
Leighton M, Leighton DR (1982) The relationship of size of feeding aggregate to size of food patch: howler monkeys (Alouatta palliata) feeding in Trichilia cipo fruit trees on Barro Colorado Island. Biotropica 14:81–90
Lovejoy TE, Bierregaard RO Jr, Rylands AB, Malcolm JR, Quintela CE, Harper LH, Brown KS, Powell AH, Powell GVN, Schubart HO R, Hays MB (1986) Edge and other effects of isolation on Amazon forest fragments. In: Soulé ME (ed) Conservation Biology: the science of scarcity and diversity. Sinauer Associates, Sunderland, MA. pp 257–285.
Malcolm JR (1997) Insect biomass in Amazonian forest fragments. In: Stork NE, Adis J, Didham RK (eds) Canopy Arthropods. Chapman and Hall, London. pp 510–533
McNab BK (2002) The physiological ecology of vertebrates: a view from energetics. Cornell University Press, Cornell
Milton K (1979) Factors influencing leaf choice by howler monkeys: a test of some hypotheses of food selection by generalist herbivores. Am Nat 114:362–378
Milton K (1980) The Foraging Strategy of Howler Monkeys: a study in primate economics. Columbia University Press, New York
Milton K (1998) Physiological ecology of howlers (Alouatta): energetic and digestive considerations and comparison with the Colobinae. Int J Primatol 19:513–548
Milton K (2003) Micronutrient intakes of wild primates: are humans different? Comp Biochem Physiol A Mol Integr Physiol 136:47–59
Milton K (2008) Macronutrient Patterns of 19 Species of Panamanian Fruits from Barro Colorado Island. Neotrop Primates 15:1–7
Nagy KA, Milton K (1979) Energy metabolism and food consumption by wild howler monkeys (Alouatta palliata). Ecology 60:475–480
National Research Council (2003) Nutrient requirements of non-human primates. National Academy of Sciences, Washington D.C.
Norconk MA, Wright BW, Conklin-Brittain NL, Vinyard CJ (2009) Mechanical and nutritional properties of food as factors in platyrrhine dietary adaptations. In: Garber PA, Bicca-Marques JC, Estrada AE, Heymann EW, Strier KB (eds) South American Primates, developments in primatology: progress and prospects. Springer, New York, pp 279–319.
O’Brien TG, Kinnaird M, Dierenfeld ES (1998) What’s so special about figs? Nature 392:668
Occhibove F (2010) Sopravvivere alla frammentazione dell’habitat: l’ecologia alimentare di Alouatta palliata presso la stazione biologica “La Suerte”, Costa Rica. MSc Thesis. University of Pisa, Italy.
Oftedal OT (1992) The nutritional consequences of foraging in primates: the relationship of nutrient intakes to nutrient requirements. Philos Trans R Soc B Biol Sci 334:161–170
Onderdonk DA, Chapman CA (2000) Coping with forest fragmentation: the primates of Kibale National Park, Uganda. Int J Primatol 21:587–611
Pavelka MS, Knopff KH (2004) Diet and activity in black howler monkeys (Alouatta pigra) in southern Belize: does degree of frugivory influence activity level? Primates 45:105–111
Peres CA (1997) Effects of habitat quality and hunting pressure on Arboreal Folivore Densities in Neotropical Forests: a case study of Howler Monkeys (Alouatta spp. ). Folia Primatol 68:199–222
Pruetz JD, Leasor HC (2002) Survey of three primate species in forest fragments at La Suerte Biological Field Station, Costa Rica. Neotrop Primates 10:4–9
Rivera A, Calmè S (2006) Forest fragmentation and changes in the feeding ecology of black howlers (Alouatta pigra) from the Calakmul area in Mexico. In: Estrada A, Garber P, Pavelka M, Luecke L (eds) New perspectives in the study of Mesoamerican primates: distribution, ecology, behavior and conservation. Springer, U.S.A. pp 189–214
Robbins CT (1993) Wildlife feeding and nutrition (2nd ed). Academic, New York
Robinson JG (1996) Hunting wildlife in forest patches: an ephemeral resource. In: Schellas J, Greenberg R (eds) Forest patches in tropical landscapes. Island Press, London. pp 111–130
Rode KD, Chapman CA, McDowell LR, Stickler C (2006) Nutritional correlates of population density across habitats and logging intensities in redtail monkeys (Cercopithecus ascanius). Biotropica 38:625–634
Rodriguez-Luna E, Dominguez-Dominguez LE, Morales-Mavil JE, Martinez-Morales M (2003) Foraging strategy changes in a Alouatta palliata mexicana troop released on a island. In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer/Plenum Publishers, New York. pp 229–250
Rosales-Meda M, Estrada A, Lòpez JE (2007) Demographic survey of black howler monkey (Alouatta pigra) in the Lachuà Eco-Region in Alta Verapaz, Guatemala. Am J Primatol 70:231–237
Rothman JM, Dierenfeld ES, Molina DO, Shaw AV, Hintz HF, Pell AN (2006) Nutritional chemistry of foods eaten by gorillas in Bwindi Impenetrable National Park, Uganda. Am J Primatol 68:675–691
Rothman JM, Raubenheimer D, Chapman CA (2011) Nutritional geometry: gorillas prioritize non-protein energy while consuming surplus protein. Biol Lett 7:847–849
Silva EC, Da Jr (1981) A preliminary survey of brown howler monkeys (Alouatta fusca) at the Cantareira Reserve (São Paulo, Brazil). Rev Bras Biol 41:897–909
Silver S C, Marsh L K (2003) Dietary flexibility, behavioral plasticity, and survival in fragments: lessons from translocated howlers. In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer/Plenum, New York. pp 251–265
Silver SC, Ostro LET, Yeager CP, Dierenfeld ES (2000) Phytochemical and mineral components of foods consumed by black howler monkeys (Alouatta pigra) at two sites in Belize. Zoo Biol 19:95–109
Smith CC (1977) Feeding behaviour and social organization in howling monkeys. In: Clutton-Brock TH (ed) Primate ecology: studies of feeding and ranging behavior in lemurs, monkeys and apes. Academic, New York. pp 97–126
Spielman D, Brook BW, Briscoe DA, Frankham R (2004) Does inbreeding and loss of genetic diversity decrease disease resistance? Conserv Genet 5:439–448
Wallis I, Edwards M, Windley H, Krockenberger A, Felton A, Quenzer M, Ganzhorn J, Foley W (2012) Food for folivores: nutritional explanations linking diets to population density. Oecologia 169:281–291
White TCR (1993) The inadequate environment: nitrogen and the abundance of animals. Springer, Berlin
Williams-Guillèn K (2003) The behavioral ecology of mantled howling monkeys (Alouatta palliata) living in a Nicaraguan shade coffee plantation. PhD dissertation
Whittaker RJ, Fernandez-Palacios JM (2007) Island biogeography: ecology, evolution, and conservation. Oxford University Press, Oxford
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168
Acknowledgments
This work was carried out under the collaboration between the Department of Biology (Anthropology Unit) of University of Pisa, the Department of Veterinary Science of University of Pisa, and the Department of Animal and Ecology and Conservation of University of Hamburg. We acknowledge the staff of the above Departments for providing help during the chemical analyses and the data analyses. We are grateful to Renee Molina and the La Suerte Biological Field Station staff for their warm welcome and for providing help during the data and sample collection in the field. We thank all the students of the 3rd session of the summer course “Primate Behaviour and Ecology” (years 2009, 2010, and 2011) in La Suerte for their cooperation and precious help during data collection.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Occhibove, F. et al. (2015). Living in Islands of Forests: Nutritional Ecology of the Howler Monkey (Alouatta palliata) at La Suerte Biological Field Station, North-Eastern Costa Rica. In: Huettmann, F. (eds) Central American Biodiversity. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-2208-6_22
Download citation
DOI: https://doi.org/10.1007/978-1-4939-2208-6_22
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-2207-9
Online ISBN: 978-1-4939-2208-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)