
Abstract
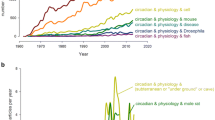
Organisms use various clocks in order to adapt to the daily, tidal, monthly, and annual cycles of the environment. This chapter deals with circadian (daily) clocks and the role light plays in synchronizing them with the 24 h cycles in the environment. We will first characterize these different clocks, their functions, their properties, how to model them, and how light affects them and mention their adaptive significance (Sect. 18.1). The main part of the chapter describes how the circadian system of several organisms is synchronized by light: Synechococcus and Synechocystis are chosen as representatives of cyanobacteria (Sect. 18.2), Ostreococcus and Chlamydomonas as examples of unicellular algae (Sect. 18.3), Arabidopsis as a plant (Sect. 18.4), the ascomycete Neurospora as a fungus (Sect. 18.5), Drosophila as an insect (Sect. 18.6), and rodents (Sect. 18.7) and man (Sect. 18.8) as mammals. These examples show that circadian rhythms occur in almost all organisms. The similarities and differences of their clock mechanisms and the way they are entrained by light are pointed out.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
18.1 Biological Clocks
The daily revolutions of the earth around its axis are responsible for day and night and its annual orbit around the sun for the seasons with their fluctuations in day length. Most organisms have adapted to these diurnal and annual cycles. The strategies and mechanisms used are quite delicate and complicated.
It came as a surprise that photosynthesis and many other processes are, however, additionally controlled by internal clocks. Thus, photosynthesis fluctuates not only during the daily light-dark cycle (=LD; see the List of Abbreviations and http://www.circadian.org/dictionary.html) but also when the plants are kept under LL and constant temperature (Hennessey and Field 1991). However, the period length (period for short) of this rhythmic event then deviates from exactly 24 h and is therefore called circadian (from Latin circa, about, and dies, day). If in the absence of LD and temperature cycles other 24 h time cues (also called zeitgeber, German for time giver) would control the rhythm, it should show an exact 24 h rhythm. This is not the case, demonstrating the endogenous nature of a clock that is locked to light signals (see Fig. 18.1).

Top: petal movement of a single flower of Kalanchoe blossfeldiana released from 12:12 h LD cycles in DD for 7 days shows free run of 22 h (whereas in LD the period is 24 h, not shown). Maximal (short red arrows) and minimal (short blue arrows) opening of flowers, period length τ (red double-headed arrow), phase Φ and amplitude (vertical red line) are indicated. Second diagram: rhythm annihilating light pulse (1st LP, red arrow), which, if given at the proper time (in minimum) and strength, induces arrhythmicity. A second LP (2nd LP, red arrow) starts the oscillation again. Insets show open and closed petals of flowers; see also a time-lapse movie (links: http://nbn-resolving.de/urn:nbn:de:bsz:21-opus-66695 and http://nbnresolving.de/urn:nbn:de:bsz:21-opus-66709)
18.1.1 Spectrum of Rhythms
Endogenous rhythms of organisms are not only tuned to the daily cycle of 24 h. The range of rhythms found in organisms covers ultradian (with periods of several hours to very short ones), circadian, and annual (with periods of about a year) rhythms. Other rhythms such as tidal, 14-day, and monthly ones cope with influences of the moon on the earth, mainly on the water movements of the oceans, and they are therefore found in organisms at the coasts and in the sea. Annual rhythms interact with the day-length changes during the year (see below). There are furthermore rhythms with periods covering several years. The following discussion of a “biological clock” is restricted to circadian rhythms. Even they are often not just composed of one clock type but form a “circadian system” consisting of two or more clocks with different properties which are or are not coupled mutually (Rosbash 2009; Bell-Pedersen et al. 2005; Panda and Hogenesch 2004).
18.1.2 Function of Circadian Clocks
The term “clock” usually implies a time-measuring device or function. For instance, the day length (or night length) can be determined by an organism. Since day length is a function of the time of the year (long days in summer, short days in winter), it can be used to time certain events such as flowering or tuber formation of a plant or breeding of birds and mammals during the most appropriate season. These processes are denoted photoperiodism (see Chap. 19).
However, a clock can also be used to set a certain temporal order. For instance, the circadian control of our sleep-wake cycle ensures that we rise in the morning and fall asleep in the evening at a preferred time. Food intake and digestion are likewise controlled by this clock and gated to certain times of the day (Silver et al. 2011; Duguay and Cermakian 2009; Forsgren 1935). The circadian clock will time these events also under constant conditions.
Furthermore, circadian clocks can serve as alarm clocks. They tell the organism important times of the day. For instance, the alarm clocks of insects such as bees allow them to visit the flowers of a plant at the time they offer nectar and/or pollen. From the standpoint of the plant, attracting certain insects is more efficient if timed to their active period. If flowers open at night, pollination by moths or bats is facilitated. Evolution has worked on the plant and the pollinator to bring about this delicate interplay controlled by circadian clocks. This phenomenon is often termed synchronization to the environment.
Alarm clocks might also exist in man. Some humans are able to wake up at a certain time of the night without external help by relying on a “head clock” (Clauser 1954). Although not tested yet, this alarm device might be based on a circadian clock.
Circadian clocks can also be used by insects (Heinze and Reppert 2012; Homberg et al. 2011; Merlin et al. 2011; Reppert et al. 2010; Collett 2008), birds (Muheim 2011), lizards (Foà et al. 2009), fishes (Leis et al. 2011), bats (Holland et al. 2010), and other animals (Ugolini et al. 2007) for navigation and orientation using the direction of the sun. These animals have to take the changing position of the sun or the polarization pattern of the sky during the day into account, and the circadian clock is used as an internal time reference for this sun compass orientation (Able 1995; Schmidt-Koenig 1975). Some birds fly at night and orient by using a star compass, whereby the changing night sky is compensated for (Dacke et al. 2011; Ugolini et al. 2005).
18.1.3 Properties and Formal Structure of the Circadian System
Besides being found in almost all living beings, from prokaryotes to higher organisms, circadian clocks possess a number of formal properties. The clocks:
-
Have a period of roughly 24 h (about 18 and 28 h in extreme cases) under constant conditions
-
Are synchronized by time cues (mainly daily light and temperature changes) to 24 h function on the cellular level and are heritable
-
Are of advantage to the clock bearer
-
Have a period that is only slightly dependent on temperature (if constant)
If, for instance, the plant Kalanchoe blossfeldiana is kept under constant weak green light conditions, the period of the opening and closing of the four petals of the flowers amounts to 22 h at a temperature of 22 °C (Fig. 18.1). If exposed to an LD 12:12, the flowers open during the light period and close during the dark period. The period of the cycle is now exactly 24 h. Under constant conditions the “free-run” period is 21.9 h at 15 °C, 22.3 h at 20 °C, and 21.3 h at 25 °C (Oltmanns 1960). The differences in period are quite small compared to the influence temperature normally has on chemical and biochemical reactions.
Mutants of organisms are known which differ in clock properties. For instance, the locomotor activity rhythm of the Drosophila mutant pers has a period of 19.5 h compared to 24.4 h for the wild type, and the period of the mutant perl amounts to 28.6 h. Another mutant (per0) is arrhythmic.
Any useful model of a circadian system has to take the general properties above into account and has to offer mechanisms which lead to the circadian period of about 24 h, to the low temperature dependence of period, to ways of synchronizing the rhythms to the 24 h time cues, etc. We will first discuss modeling of a circadian system (see Sect. 18.1.4) and then focus on some features of the light action on the clocks (see Sect. 18.1.5). Clocks in different organisms will be treated in subsequent sections.
18.1.4 Modeling Circadian Clocks
Published models of circadian systems are of different kinds. Some are purely mathematical ones, describing the variables in, usually, differential equations; others are presented as block diagrams based on concepts from control theory. They often use numerical methods to simulate the circadian behavior. A third form of models describes reactions in words and figures without deriving or attempting quantitative relations (see the selected examples in Sects. 18.2, 18.3, 18.4, 18.5, 18.6, and 18.7).
Ultimately, the models should give precise qualitative and quantitative descriptions and predictions at the molecular, the cellular, and the organism level.
It is important to model not only the circadian system proper but also the inputs particularly the light pathways – and also the outputs of the clock.
The Light Input to the Clock
The detailed way in which light affects a circadian system is important for a model. Light signals from the environment are perceived in photoreceptor molecules and organs which might differ widely between organisms. These photoreceptors have to be identified for each system. After photons are absorbed, the excitation energy affects the clock via a signal chain. The details of these pathways have to be known and the way in which the transformed light signal enters the clock has to be determined. Modeling requires specific knowledge for each circadian system under study.
The Circadian System and the Feedback Concept
In models for circadian systems, the concepts of positive and negative feedback and of time delay are frequently used. Feedback simply means that a signal in the system is fed back to one or several points in the system and affects the production or the destruction of the signal itself. Control theory tells that feedback in a system might lead to oscillations, in particular if the signal in the feedback loop is delayed in a suitable way.
In several relevant models the feedback links can easily be visualized: often this “circular process” is denoted TTFL (for transcription-translation feedback loop) in the present context. The time delays which exist in the system could be due to transcription, translation, transport, and production or decomposition of clock-related components (see Sects. 18.2, 18.3, 18.4, 18.5, 18.6, and 18.7).
A simple description of a feedback oscillator is as follows: let c(t) represent the concentration of an oscillating central variable in the clock (e.g., the protein FRQ). In a feedback model the signal c(t) in the loop is delayed in a suitable way before feeding back to reinforce (amplify) an already existing signal and induce oscillations. If we assume that the substance is produced at time t according to the concentration of the same substance c(t) at a certain earlier time c(t − t 0), we have a simple feedback system with delay t 0. The situation can be expressed as
Here K is a positive constant and the negative sign indicates that production is decreased if the concentration was high t 0 hours earlier, while it is increased if concentration was low t 0 hours earlier (inhibition occurs if concentration was high, activation occurs if concentration was low at some time units earlier).
The approach can describe sustained oscillations in the variable c if the delay t 0 and the feedback signal are large enough (i.e., if K is large enough). Furthermore, the period of the oscillations will be about four times the delay time introduced. Circadian oscillations would thus need a delay of about 6 h in the example in order to end up with a 24 h period. Interestingly, experimental results pointing at an explicit delay of about 6 h in a molecular feedback chain of the clock in Drosophila has recently been published (see page 39 and Meyer et al. 2006).
Simple models based on explicit feedback and time delay concepts (but using nonlinearities that are always present in biological systems and needed to create sustained and limited concentrations in the systems) have been used to simulate features of circadian rhythms (Kalanchoe petal rhythm, Johnsson et al. 1973; Karlsson and Johnsson 1972), photoperiodic flowering in Chenopodium (Bollig et al. 1976), and activity rhythm in the New Zealand weta (Lewis 1999). The specifications of the TTFL began later with the mechanisms of interaction between mRNA and protein levels (Hardin et al. 1990), and several molecular models have been published (Leloup and Goldbeter 2008; Dunlap et al. 2007; Loros et al. 2007; Mackey 2007; Lema and Auerbach 2006).
Many models of the circadian clockwork have been published, emphasizing different aspects of the oscillating system (Dalchau 2012; Beersma 2005) and different approaches such as used in systems biology (Hogenesch and Ueda 2011; Yamada and Forger 2010; Ukai and Ueda 2010; Hubbard et al. 2009). For the history see Tyson et al. (2008) and Roenneberg et al. (2008).
Output Signals from the Clock
It is also important to model reaction sequences downstream of the clock. The period of the circadian system will be reflected in the reactions driven by the clock. Amplitude and phase of the driven reactions might change, but the final reactions that are observable – the hands of the clock – have the same period as the clock. This is stressed since environmental light signals might affect the downstream reactions directly, thereby changing, for instance, their amplitude. Such changes should not be mistakenly ascribed to light effects on the circadian system itself.
The photoreception can be clock controlled by feedback links that change the properties of light receptor systems (for instance, control of the iris muscle in mammals, Fig. 18.10, and leaf position in plants). In addition, light adaptation and other changes of sensitivity to light might increase the level of complexity in modeling the light-induced effects on the circadian clock. Detailed modeling of the light reactions relies of course on experimental investigations of the light perception and transduction of the various organisms.
Posttranscriptional feedback loops (PTFL) have acquired much interest in recent studies, e.g., with respect to KaiC phosphorylation, Sect. 18.2, and also protein oxidation processes (Brown et al. 2012). In such a system the parameters of the overall system will then be dependent on the PTFL and more complex output signals can be found.
Single and Multi-oscillator Models
Several important features of circadian systems are modeled on the assumption that one single oscillator controls the clock. A one-oscillator model does not preclude the presence of many cellular oscillators. It only assumes that they are so strongly coupled to each other that they (in most cases) behave as one single unit (a “lumped” model).
However, in multi-oscillator models the circadian system can have new features that are not explainable under the assumption that the system consists of one single oscillator. The circadian system of humans is an example which is often modeled by two interacting oscillators. One of them is then assumed to have its strongest influence on (among other rhythms) the activity rhythm and the other one on (among others) the body temperature rhythm. Usually the two oscillators are coupled and oscillate in phase, but the dual nature of the system can show up in, for example, isolation experiments (without time cues) where the rhythms might display different periods (Oishi et al. 2001; Kronauer et al. 1982; Wever 1979).
Modeling often starts with a simple one-oscillator assumption, an approach that eventually turns out to be too simple. Many circadian systems should be modeled as multi-oscillatory systems, even on a single cell level (Daan et al. 2001; Roenneberg and Mittag 1996). In the case of Drosophila, several oscillators are nowadays implicated in more detailed modeling (see Sect. 18.6.3).
It is interesting that after a period of intensive experimental studies of the molecular mechanisms that underlie circadian rhythms, formal modeling of circadian rhythms and their light reactions has gained impact. Many models of the circadian clockwork have been published, emphasizing different aspects of the oscillating system. Not all of them focus on the light perception and the light reactions. We, therefore, do not mention all models here but refer to some papers (Beersma 2005; Ruoff and Rensing 2004; Leloup and Goldbeter 1999; Forger et al. 1999; Lakin-Thomas and Johnson 1999; Jewett et al. 1999a, b; Leloup and Goldbeter 1998; Deacon and Arendt 1996; Goldbeter 1995; Diez-Noguera 1994).
18.1.5 Comments on Light, Photoreceptors, and Circadian Models
As has been emphasized, light is the most important input signal to a circadian system, and there are several general features that must be handled by models such as:
-
Entrainment: Repetitive light pulses entrain the circadian rhythm (“entrainment,” “synchonization,” “phase locking”). The external light cycle will function as a synchronizer. This general property of circadian systems has also to be simulated by models of circadian systems. The range of entrainment can be used to test models. Besides light, temperature changes are also entraining circadian rhythms. At the same time, the speed of the circadian clock is only marginally affected by the environmental temperature, because they are “temperature compensated.” Models should take care of both facts (see, e.g., Ruoff and Rensing 2004, 1996).
-
Single light pulses: Single light pulses given to an organism during free-run phase shift, the rhythm and a phase response curve describes its time course. A model should handle this and the light signal pathway into the clock in detail.
-
Acclimation: Photoreceptors function over a huge range of light intensities. As an example, the human eye covers nine orders of magnitude. Still, the eye senses a contrast ratio of only 1,000. The reason is that the eye adapts to a light level that is interpreted as darkness. It can shift across six orders of magnitude. It takes 20–30 min to adapt from bright sunlight to complete darkness and about 5 min to adapt to bright sunlight from darkness.
-
Masking: Masking is an immediate response to stimuli such as light and other environmental influences that overrides the influence of the circadian system on behavior and physiology of an organism. Masking effects differ from entrainment of the clock, and techniques can be used to distinguish between both (Rietveld et al. 1993). Nocturnal animals respond to darkness by becoming more active (positive masking) and to light by becoming less active (negative masking). Diurnal animals show the opposite response (Pendergast and Yamazaki 2011). In fruit flies (Kempinger et al. 2009) and primates (Erkert et al. 2006), nocturnal light can shift the circadian clock and increase nocturnal activity independent of the clock.
-
Damping: Circadian rhythms might damp out under certain environmental conditions such as LL and/or DD or at too high or too low temperatures. There is apparently a permissive range allowing circadian rhythms to occur.
-
Stopping the clock by light pulses: In many models phase shifts and amplitude changes brought about by light pulses are concomitant features. Under certain conditions an external light pulse can reduce the amplitude completely, thus stopping the oscillation, and models must cope with this feature. Arrythmicity was indeed found (see Winfree 1970) for certain combinations of irradiance and pulse durations in Drosophila (Chandrashekaran and Engelmann 1973), modeled by Leloup and Goldbeter (2001), in Culex mosquito (Peterson 1981a, b), and in Kalanchoe (Engelmann et al. 1978). The phase at which such an arrhythmicity can be induced was fairly restricted (subjective midnight point; the strength of the pulse has to be such that it is just between evoking a strong or a weak phase response curve). A mathematically and biologically interesting question arises: will a circadian system start oscillating spontaneously again after having been sent into the nonoscillatory state or is it stable? The so-called singularities, limit cycles, etc., have attracted interest with respect to the mathematical structure of the circadian systems. An interesting case of arrhythmicity in the Siberian hamster was reported by Steinlechner et al. (2002) (see page 51).
The necessary synchronization of a circadian clock to the environmental cycle is most frequently achieved by using the LD cycle as time cue, but temperature rises or temperature drops can also function as zeitgeber. In animals non-photic zeitgeber such as feeding, social cues, and other signals can entrain (Silver et al. 2011; Honma and Honma 2009; Satoh et al. 2006; Mistlberger and Skene 2005; Stephan 2002).
If a Kalanchoe plant is kept for some days in an air-conditioned chamber with 12:12 h LD and after the last 12 h of light transferred to DD, the circadian opening and closing of the flowers will continue to run with its characteristic period going through subjective day and night cycles. A light pulse would shift this rhythm or not, depending on the phase of the clock at which the pulse is applied. If given before the subjective midnight point, the rhythm will be delayed, if given after this point, the rhythm will be advanced. During the subjective day period, there is normally a “dead zone” where a light pulse is without effect on the rhythm. These phase shifts can be plotted with respect to magnitude and direction by a phase response curve. They are based on experiments with light pulses administered at different phases (see Fig. 18.2).

(a) Eclosion rhythm of Drosophila pseudoobscura. Pupae kept in LD 12:12 h and released in DD at time 0 on first day. Curves in upper part show eclosion rate of a control population, the phase-shifting effect of a 25 sec blue-light exposure at two different phases (2 and 6 h after onset of DD, marked by blue vertical arrow) leading to a delay (→) and an advance (←), respectively (differences between the time of maximal eclosions of control (blue vertical dashed lines) and eclosion peaks of light-treated groups (red arrows). (b) A phase response curve plots the magnitude and direction (−values, delay; +values, advance) of the phase shifts against the phase at which the pupae were illuminated. Left curve (blue) for weak phase responses, right curve (red) for strong responses. CT is circadian time, CT 0 is the time at which the light period would begin if the LD12:12 would have continued. Inset shows a fly eclosing from the puparium (see also the time-lapse movies, links: [http://nbn-resolving.de/urn:nbn:de:bsz:21-opus-66660] and [http://nbn-resolving.de/urn:nbn:de:bsz:21-opus-66676])
The dominant role of light in this entrainment might be due to the high reliability of light as zeitgeber, whereas temperature changes during day and night are less reliable. However, the beginning of the light period and correspondingly of the dark period does not occur at the same time of the day during the course of the year. During the summer the light period is longer than during the winter, which is quite obvious at higher latitudes. This fact has to be taken into account by the organisms if light is the entraining agent.
Photoperiodic Induction
The Bünning hypothesis, according to which the circadian clock is used by organisms to measure day (night) length and initiate photoperiodic events accordingly (Bünning 1936), has been modeled. LD conditions in combination with variations of the circadian clock could be used to predict, e.g., flower induction and hibernation. The modeling thus involves the proper treatment of the light perception for the induction of the photoperiodic events and the light perception for the phasing and entrainment of the clock. The two perception mechanisms can of course be unified – only experiments can verify the models proposed. Even simple approaches can in some cases model photoperiodic events fairly precisely, e.g., flower induction in Chenopodium as described by Bollig et al. (1976).
Photoreceptors
Depending on the organism the photopigments and photoreceptors for resetting the clocks can be quite diverse (Collin et al. 2009; Cermakian and Boivin 2009; Foster et al. 2007 and Table 18.1). In many unicellulars, such as yeast or most algae such as Ostreococcus (Sect. 18.3), no special receptor structures have been found (Gotow and Nishi 2008). Instead pigment molecules in the cells are changed by light and a transduction chain finally resets the clock. In animals specialized light receptive organs are used such as the vertebrate eyes or the compound eyes in insects. But often extraretinal photoreceptors serve to perceive the synchronizing light either in addition to or instead of the usual eyes. For instance, in birds the pineal organ is light sensitive and synchronizes the circadian rhythm if the eyes are obscured or denervated or removed. In Drosophila flies, the circadian clock neurons in the brain are light sensitive on their own via the blue-light pigment cryptochrome. Furthermore the Hofbauer-Buchner eyelets are extraretinal structures in the brain and serve as additional devices for synchronization (Sect. 18.6).
There are several reasons why organisms use multiple photopigments and photoreceptors (Foster and Helfrich-Förster 2001; Roenneberg and Foster 1997); see also page 39, among them:
-
Natural LD cycles do not simply consist of light steps. Instead, light is increasing and decreasing slowly during the twilight of the day.
-
If organisms use certain light intensities during twilight as the onset, respectively, end of the day, the day length can be measured accurately and reliably and independently of daily weather conditions.
-
During twilight at dusk and dawn, not only the intensity of light changes but also its spectral composition. Different qualities of the environmental light can be used by a set of different photoreceptors.
-
Entrainment by dawn and dusk is more effective than lights on/off programs in all animals tested so far including man Fleissner and Fleissner (2001).
-
The signal-to-noise ratio is reduced if several inputs are used. Photopigments like PHY (Auldridge and Forest 2011), CRY (Chaves et al. 2011), opsins (Foster et al. 2007), and others synchronize circadian rhythms. Properties and functions of relevant pigments are described under the examples for organisms with circadian rhythms (Sects. 18.2, 18.3, 18.4, 18.5, 18.6, and 18.7). Depending on the kind of photoreceptor, different spectral wavelengths are more or less effective in resetting the circadian clock. Using varying fluence rates of colored light, action spectra can be obtained (see Chap. 8) which tell us how many photons of the different wavelengths are needed in order to evoke the same effect (see page 35). The effect of light depends, however, not only on the wavelength and the fluence rate but also on the phase of the circadian clock at which the light was given (see Sect. 18.1.5) and on the duration and shape of the pulses.
18.1.6 Adaptive Significance and Evolutionary Aspects of Circadian Clocks
The adaptive significance of possessing a circadian clock (Johnson 2005) has been demonstrated in cyanobacteria by using mutants with different periods in competition with each other and with the wild strain (Woelfle et al. 2004; Gonze et al. 2002; Johnson and Golden 1999; Ouyang et al. 1998): see also page 15, in Arabidopsis (Hut and Beersma 2011; Yerushalmi et al. 2011; Michael et al. 2003; Green et al. 2002), in Drosophila (Xu et al. 2011; Rosato and Kyriacou 2011; Kumar et al. 2005; Beaver et al. 2002; Fleury 2000; Klarsfeld and Rouyer 1998), and in mammals (Daan et al. 2011; Tauber et al. 2004; Sharma 2003; Hurd and Ralph 1998; DeCoursey and Krulas 1998).
The different functions of circadian clocks just mentioned are surely not the only reasons why they evolved. Winfree (1986) and others have discussed that early in evolution circadian clocks might have served to protect organisms from adverse effects of light. Circadian timing and light reception might have coevolved and even preceded the evolution of specialized photoreceptors and eyes. Homologies between pacemaking molecules and ancient photopigments from fungi to mammals suggest an evolutionary link between modern clock proteins and ancient light sensing proteins (Tauber et al. 2004; Sharma 2003; Crosthwaite et al. 1997). However, this link is difficult to prove. An interesting example is CRY, which is used as a clock protein in mammals but as a photopigment in Drosophila’s clock neurons in the brain. In peripheral clocks of Drosophila, CRY appears to fulfill both roles. Furthermore, in higher animals (vertebrates, insects) the retina is not only a photoreceptor organ but harbors at the same time (peripheral) clocks (see Sect. 18.7.4). It would be interesting to know whether primitive eyes (for instance, eye spots) contain circadian clock cells. Among vertebrates, retinal clocks seem to be quite ancient (lamprey Menaker et al. 1997).
Vertebrates show a wide evolutionary variety in their circadian system. They possess a so-called circadian axis (retina, pineal, suprachiasmatic nucleus) with circadian oscillators. In mammals, the pineal as part of this axis does not contain a circadian oscillator. Mammals also lack extraretinal circadian photoreceptors (in the pineal) in contrast to other vertebrates (Bertolucci and Foà 2004). A “nocturnal bottleneck” that could have led to the evolution of mammals and their exceptional circadian system is discussed by Menaker et al. (1997).
18.1.7 Current Concepts and Caveats
To understand how circadian clocks are synchronized by light and other time cues, the mechanisms of circadian oscillators have to be known, as well as the photoreceptors and pigments involved in the entrainment. The clock mechanisms are currently intensively studied (see Sect. 18.2 and the following ones). The prevailing opinion is that feedback loops (TTFLs) between clock gene products acting on the promoters of their genes are at the heart of these clocks (Hardin 2005). Transcription and translation are thus involved in modeling the clock.
However, the picture is probably more complicated, and cautions have been raised (Lakin-Thomas 2006). For instance, these TTFLs might not be the core clocks, but elements between the environmental inputs and the clock mechanism proper (Merrow et al. 1999). Other cases have been reported which make it difficult to accept the presently favored concept of a circadian clock mechanism as a general one. Enucleated Acetabularia still has a circadian rhythm of oxygen production (Karakashian and Schweiger 1976), dry seeds of bean plants show circadian rhythms in respiration (Bryant 1972), and some enzymes of human erythrocytes fluctuate in a circadian way (O’Neill and Reddy 2011; Ashkenazi et al. 1975). What is common to the two last mentioned systems is the complete lack of nucleic acid metabolism. This is an important issue, since several of the recently proposed models of circadian systems use feedback systems in transcriptional and translational events. It might therefore be wise to keep an open eye on alternative mechanisms underlying the circadian oscillators. Of course, there is no guarantee that all circadian clocks use the same mechanism, although their properties are often quite similar.
Proteins could, for instance, be involved in timing mechanisms. We refer to the circadian clock mechanism in Synechococcus in Sect. 18.2, to Ostreococcus in Sect. 18.3, and to a report of Meyer et al. (2006) on the Drosophila clock (page 39). Another interesting case is the diapause of embryos in the eggs of silk moths (Bombyx mori), which is broken by exposure to low temperature. The duration of the chilling period is measured by esterase A4 complexing with another enzyme, PIN. After 14 days it dissociates from PIN, the conformation of the esterase A4 changes, and it becomes suddenly active. This enzyme is thus a kind of molecular timer (although here not on a 24 h basis, Kai et al. 1999).
Membranes and electrical activities might also be involved in the clock mechanism. Colwell suggested that neural activities in the suprachiasmatic nucleus (SCN) are required to generate rhythms in gene expression (Colwell 2011); see also results of Nitabach et al. (2005) in Drosophila.
Even if TTFLs are not at the heart of all clock mechanisms, it is still possible to build models on the general concept of feedback as discussed on page 6 – the delays and the molecular mechanisms have then to be found among other cellular reactions.
Photic phase response curves are similar in all organisms and this is true for mammals, nocturnal as well as diurnal, including man. However, the amplitude and duration of the advance and delay portion and the presence and length of a dead zone (see page 10) might vary in different species (Rusak and Zucker 1979). This allows for adjustment of the phase and period of the circadian clock to the 24 h day.
We will now discuss the circadian clocks and photoreceptors in selected examples.
18.2 Clocks and Light in Cyanobacteria
The simplest organisms known to possess a circadian clock are cyanobacteria. These prokaryotes are among the smallest, albeit most abundant, organisms on earth and were for a long time not thought to possess a circadian clock. It was assumed that a cell dividing several times per day (e.g., Synechococcus elongatus once every 5–6 h, Mori et al. 1996) would have no use for a circadian timing mechanism and that a nucleus is needed.
However, Stal and Krumbein (1985a, b) observed in cyanobacteria a circadian nitrogenase activity in reducing atmospheric nitrogen to ammonia. This enzyme is inhibited by oxygen and has therefore to be protected against oxygen produced during the day by photosynthesis. Evolution solved this dilemma in two ways, by either separating the processes in space or in time (Mitsui et al. 1986). Later it turned out that gene expression, metabolism, and cell division are all driven by a circadian clock (Johnson 2010). Besides Synechococcus (Mackey et al. 2011), the circadian rhythms of other cyanobacteria such as Synechocystis (Layana and Diambra 2011), Cyanothece (McDermott et al. 2011; Bradley and Reddy 1997), and Prochlorococcus (Mullineaux and Stanewsky 2009; Axmann et al. 2009; Zinser et al. 2009) were also studied. Synechococcus and Prochlorococcus dominate the picophytoplankton of the oceans, the latter being probably the most abundant photosynthetic organism on earth. In the following the main properties of the circadian clock, clock-driven processes, and the present view of the molecular clockwork and its light resetting in cyanobacteria are presented. How circadian clocks in cyanobacteria might have evolved is discussed by Johnson et al. (2011), Hut and Beersma (2011), and Simons (2009).
The necessary temperature independence of the period length (Sect. 18.1.5) was indeed found, even in the thermophilic cyanobacterium Thermosynechococcus elongatus tested in a temperature range between 35 and 55 °C (Onai et al. 2004). Since chemical reactions are usually temperature dependent with a Q10 often around two to three, meaning that the reaction is twice or three times as fast at a temperature 10 °C higher, mechanisms are needed to compensate the temperature effects (proposals; see Hatakeyama and Kaneko 2012; Akiyama 2012; Murakami et al. 2008; Kotov et al. 2007).
The circadian clock should be accurate despite a noisy environment inside and outside the cell. The individual oscillators in cyanobacteria are indeed quite stable. The stability in a population could be due to intercellular coupling, but this has been shown to be negligible theoretically and experimentally (Amdaoud et al. 2007). The high stability of individual oscillators in cyanobacteria must therefore be based on genetical and metabolic grounds.
In spite of the high precision of the clock, it has to be synchronized with the 24 h environment. The main environmental time cues are light and temperature. Lin et al. (1999) showed for Synechococcus that temperature pulses in addition to light entrain the circadian clock, but light was the most efficient time cue under the experimental conditions chosen. The rhythm continues if the cultures are transferred to LL or DD conditions, but as in other diurnal (i.e., day active versus nocturnal) organisms, the circadian period is shorter at higher light intensities and longer under lower intensities (25 h in DD, 22.6 h in LL, Aoki et al. 1997; Kondo et al. 1993).
Light phase shifts the rhythm (Golden et al. 2007), and a preliminary action spectrum has been determined (Inouye et al. 1998). It resembles the absorption spectrum of chlorophyll, indicating that photosynthesis in the thylakoids is responsible for the entrainment and phase shifting of the rhythm. At least, no other photoreceptor as an essential input pathway has been found in screens for phase-resetting mutants, although seven blue-light candidates have been predicted (Mackey et al. 2009). Instead, the phase of the clock seems to depend directly on the energy state of the cell, and the metabolic changes (caused by light) synchronize the clock (Mackey et al. 2011). This was tested by changing the ATP/(ADP + ATP) ratio in an oscillating in vitro system consisting of KaiA, KaiB, and KaiC. The metabolic effects of darkness were simulated by adding ADP to reduce the ratio of ATP/(ADP + ATP). To simulate the return to light, pyruvate kinase was added to convert the ADP to ATP. Phase shifts in the phosphorylation rhythm resulted, and the phase response curve obtained in vitro was similar to the one in vivo (Rust et al. 2011). A model by Rust et al. (2007) mimicked the phase response curve obtained from in vitro experiments. There seems to be a direct sensing of the electron flow by electron carriers of photosynthesis and respiration which synchronize the clock by affecting the ATP/ADP ratio and the oxidative state of the plastochinon pool (see Fig. 18.3 and its legend).

Model of the circadian clock of the cyanobacterium Synechococcus elongatus (inset, kindly supplied by Kondo: A cell). The hexameric KaiC protein (KaiC6m, red) consists of 6 monomeric KaiC proteins (KaiC1m with N- and C-terminal domains at very right) and undergoes during the course of a day (thick green arrows, night and day indicated by N and D in black circle) conformational changes (top and right, condensed as indicated by small dashed ovals; bottom, relaxed with wider dashed ovals, both with double-headed arrow indicating the width). The cycling of this loop is governed by Mg-ATP phosphorylation during the day due to the KaiA protein (green structure; genes in italics). Mg-ATP competes with the hexameric KaiB (KaiB4m, brown circles) for KaiC. Later the phosphorylated (P’s) KaiC becomes dephosphorylated, and the KaiB and KaiA are dissociated by mid-morning (relaxed conformation). Still later, under the influence of KaiA, assembly begins again and a new cycle starts. A rhythmic output (~bottom right, upper black arrow) pathway transduces temporal information from the oscillator to the genome by switching the coiling structure of the chromosome DNA (blue waves) from supercoiled (top, day) to relaxed (bottom, night): this affects metabolism via high and low amplitude expressing genes. There is furthermore an increase (→) in transcription via SasA and RpaA (~bottom right, lower black arrows). Red and blue light (flashes, top left) is absorbed by chlorophyll a in the thylakoid membrane (as in the following figures light is an L in a yellow circle and its absorption indicated in yellow). The photosynthetic electron transport and the respiratory electrons determine the plastoquinone ratio between the reduced (blue circle Qred) and oxidized (red circle Qox) form. Oxidized plastoquinone binds to KaiA (green structure), which aggregate (not shown) and degrade (light green oval, broken border) (After Mutoh et al. (2013), Kim et al. (2012), Mackey et al. (2011), Johnson et al. (2011), Qin et al. (2010))
The clock mechanism has been studied intensively, and the results are reviewed in a number of papers, such as Johnson et al. (2011), Mackey et al. (2011), Dong et al. (2010a), Loza-Correa et al. (2010), Taniguchi et al. (2010), Brunner et al. (2008), and Iwasaki and Kondo (2004). In the following we will briefly sketch it and its properties and the inputs and outputs.
The circadian system of cyanobacteria was supposed to consist of a negative feedback loop where the products of a gene cluster of three open reading frames KaiA, KaiB, and KaiC influence the transcription of their genes (Ishiura et al. 1998). It turned out, however, that these Kai proteins form a basic timing process of the circadian clock which, in contrast to the circadian clock mechanisms in eukaryotic organisms, persists even without transcription and translation (Nakajima et al. 2005).
The properties of the circadian rhythm are not ascribed to the Kai promoters, but to the Kai proteins. Specific regulation of the KaiBC promoter is not essential for the oscillation; even an Escherichia coli-derived promoter could do, provided the promoter supports sufficient RNA polymerase activity. A functional clock can be assembled from the KaiA, KaiB, and KaiC proteins in the presence of ATP in a test tube and exhibit its regular circadian period without damping for at least three cycles autonomously. The in vitro rhythm is furthermore temperature compensated, and it reflects the period if proteins from mutations affecting period length are used. The in vivo phase-resetting effect of light can be mimicked in vitro by adding ATP. The KaiABC clock has been studied biochemically, biophysically, and structurally (Murayama et al. 2011; Johnson et al. 2011).
How this protein clockworks is depicted in Fig. 18.3, which has been simplified by concentrating on the mechanisms which are used for the light entrainment of the clock (Kim et al. 2012). For details of the KaiC loop and its phosphorylation and dephosphorylation, see Qin et al. (2010). The players are the KaiC, KaiB, and KaiA proteins. KaiC consists of six monomers of two duplicated domains, CI (=N-terminal ring) and CII (=C-terminal ring). They form a homohexamer, which can be observed under the electron microscope (Mori et al. 2002). Twelve ATP molecules bound between its N- and C-terminal domains. Both domains possess ATPase activity, and the interfaces between CII domains are sites of phosphotransferase activities (Egli et al. 2012). KaiA enhances phosphorylation of KaiC, and KaiB inhibits it. The histidine kinase SasA interacts with KaiC and is necessary for a robust circadian rhythm (Iwasaki et al. 2000). KaiC contains two ATP-/GTP-binding domains which play an important role in the rhythm generation (Nishiwaki et al. 2000).
KaiC phosphorylation is the molecular timer for the circadian rhythm in Synechococcus. The energy consumed per day amounts to 15 ATPs only (net, the absolute numbers of ATP molecules hydrolyzed and synthesized over the daily cycle are unknown). The period is mainly determined by the KaiC, since period mutations (ranging from 14 to 60 h (Kondo et al. 1994)) consist of single amino acid substitutions in the KaiC protein.
How this chemical clock could work has been discussed by Naef (2005), and models have been proposed by Kurosawa et al. (2006). Since no distinct phase element was found, the question is, how this timing mechanism enables global circadian gene expression. Apparently, the clock regulates the compaction (condensation or super-coiling status) of the chromosome and in this way controls the access to promoter elements and expression of genes globally, leading to circadian oscillation in many parts of metabolism and physiology (Woelfle and Johnson 2006; Cervený and Nedbal 2009; Nakahira et al. 2004; Smith and Williams 2006; Mackey et al. 2011) and this has been modeled (e.g., Miyoshi et al. 2007). There are at least two classes of clock-regulated genes: about 80 % of the assayed promoters are active during the day with a maximum near the end of day. In the smaller group expression has an opposite phase and is maximal at dawn and at night when the chromosome is compacted and minimal at dusk. These genes may encode, for instance, oxygen-sensitive enzymes, and they perform best at night, when photosynthesis is absent. Chromosome dynamics or DNA topology may thus be phase determining (Min et al. 2004). The global modulation of promoter activity as a result of circadian changes in the topology of chromosomes was termed oscillating nucleoid or oscilloid model (Woelfle et al. 2004).
This simple protein clock explains, also, why and how a circadian timing mechanism can function in cyanobacteria with generation times of 8 h or less (Kondo et al. 1997) and how division can still be gated by the circadian clock (Mori et al. 1996).
Using a bacterial luciferase gene as a reporter of clock-regulated promoter activity allowed continuous video recording of the amount of emitted light from many clones on a medium in Petri dishes (Kondo et al. 1993). First, the promoter for the psbAI gene (one of three psbA genes encoding a critical photosystem II reaction center protein, D1) was found to be controlled by the circadian clock. Then it was discovered that virtually all promoters in the genome are regulated by the circadian system. Most of them are activated during subjective day; some, such as the purF promoter (purF synthesis), are activated during the night. Division is also controlled by the circadian clock, even if occurring faster than 24 h (Johnson 2010; Dong et al. 2010b).
With low-light-level microscopy, the rhythm in single Synechococcus cells could be monitored (Mihalcescu et al. 2004), and recently confocal fluorescence correlation spectroscopy was used to study the dynamics of underlying processes (Goda et al. 2012).
How are all the outputs mentioned coupled to the circadian clock? The temporal information is transmitted from the Kai oscillator to RpaA via the SasA-dependent positive pathway. A further pathway is the LabA-dependent negative pathway. It is responsible for feedback regulation of KaiC. However, the labA/sasA double mutant has still a circadian kaiBC expression rhythm. That indicates a third circadian output pathway, which is CikA dependent. It acts as a negative regulator of kaiBC expression independent of the LabA-dependent pathway. The labA/sasA/cikA triple mutant is almost arrhythmic, in spite of a preserved circadian KaiC phosphorylation rhythm. A model was proposed in which temporal information from the Kai oscillator is transmitted to gene expression through these three separate output pathways (Taniguchi et al. 2010).
18.3 Algal Clocks: From Simple to Complex
Circadian rhythms are also found in eukaryotic algae such as Euglena (Mittag 2001), Chlamydomonas (Schulze et al. 2010), Acetabularia (Yang and de Groot 1992), and Lingulodinium (Wagner et al. 2005). As an example, we will select an extreme, the smallest known, eukaryote Ostreococcus tauri and mention additionally the more complicated and 30 times larger Chlamydomonas.
Ostreococcus tauri belongs to the Chlorophyta (class Prasinophyceae) and was discovered in 1994 in the picoplankton of the Thau lagoon in Southern France by Courties and Chretiennot-Dinet (1994). The coccoid cells are haplonts, only about 1 μm small, and the ultrastructure is very simple: they lack a cell wall and contain only a single chloroplast, mitochondrion, and Golgi body besides the nucleus. Other Ostreococcus species live in many oceanic regions. High-light and low-light adapted ecotypes of Ostreococcus in the Pacific Ocean and the subtropical and tropical North Atlantic have been described by Demir-Hilton et al. (2011). In the marine environment the long wavelengths are absorbed within the first meters of the water. Therefore, specific blue-light receptors such as aureochromes are used to absorb the short wavelengths (Djouani-Tahri et al. 2011a).
The genome has been completely sequenced and annotated (which means that after identification of the elements on the genome, biological information has attached to the sequences). For further information, see also link [http://www.geneontology.org/GO.evidence.shtml||geneontology] and Corellou et al. (2009). It is tiny (13 Mb) and about 20 chromosomes are densely packed (Grimsley et al. 2010; Keeling 2007) containing only ~8,000 genes. A genome-wide analysis of gene expression was conducted under LD conditions by Monnier et al. (2010) and showed that almost all were rhythmic. Transcriptional regulation of the main processes in the nucleus and the organelles, such as DNA replication, mitosis, and photosynthesis, was found to a high extent. Genes involved in handling oxidative stress and DNA repair allow Ostreococcus tauri to grow under a wide range of light intensities.
Ostreococcus can be manipulated and propagated easily. Transcriptional and translational luciferase reporter lines are available and allow to record the expression of individual clock genes in vivo and to differentiate between effects on transcriptional and posttranscriptional processes (Djouani-Tahri et al. 2011b; Corellou et al. 2009).
Work by Thommen et al. (2010) and Corellou et al. (2009) suggests that its circadian clock is a simplified Arabidopsis clock (see Sect. 18.4.1 and Fig. 18.5). Models with only one feedback loop were proposed by Pfeuty et al. (2012) and Troein et al. (2011) to describe the Ostreococcus clock (see Fig. 18.4). The reason for using such a simple model is the finding that this alga possesses only two of the clock genes known in land plants such as Arabidopsis thaliana (which expresses five homologues of TOC1 and eight of CCA1; see Fig. 18.5). The model is based on a negative transcriptional feedback loop between TOC1and CCA1. The time delay is brought about by the timing of the expression of TOC1 and CCA1. CCA1 represses TOC1 expression during most of the day except during a couple of h before dusk by binding to an evening element sequence (EE) in the TOC1 promoter (Morant et al. 2010). It afterward induces CCA1 transcription, so that CCA1 is expressed in the night and early morning (Corellou et al. 2009; Harmer et al. 2000).

Ostreococcus tauri cell (inset) and molecular model of its circadian clock and light inputs. The clock consists of a negative transcriptional/translational feedback loop (TTFL) between TOC1 and CCA1. Transcription and translation of TOC1 (green oval) activates (thick green arrows) transcription of the CCA1 protein which represses (thick red – I) TOC1 expression. CCA1 is degraded by proteasomes (red oval, broken border) with a maximum during the day. Degradation of TOC1 (green oval, broken border) by proteasomes peaks in the dark and is diurnally regulated. This transcription/translation feedback loop (TTFL, thick arrows/lines) drives a posttranslational circadian feedback loop (PTFL, ring of differently colored arrows, mechanism unknown) and an output of it is seen in the circadian rhythm of the redox state of peroxiredoxin (PRX, not shown). Normally the TTFL and PTFL are coupled, but in DD transcription ceases and the TTFL stops. However, the PRX rhythm persists, because it is driven by the PTFL. Long- (red flash) and short-wavelength (blue flash) light (L in yellow circle) are absorbed by a rhodopsin histidine kinase (Rhod-HK, oval), respectively, LOV histidine kinase (LOV-HK, oval) and affect via histidine phosphotransfer (HPT) TOC1. There is furthermore a feedback (yellow arrows) from the TTFL to the photoreceptors (After Pfeuty et al. (2012), McClung (2011) and Troein et al. (2011). Inset courtesy of François-Yves Bouget and Marc Lefranc)

Arabidopsis clock and light resetting: Red light (red flash) is absorbed by PHY, and PIF3 links the light signal to the transcription factors CCA1 (red oval) and LHY (red oval). Blue light (blue flash) is absorbed by CRY1 and CRY2, which interact with CCA1 and LHY. In the photoperiodic induction of flowering (right part of figure), blue light is absorbed by FKF1 which, together with GI, degrades CDF1 thereby releasing under long day conditions the inhibiting effect of CO. Long days are recognized by a coincidence mechanism based on clock regulation and leads to the formation of the flower hormone FT and flowering. Another clock output, the regulation of the hypocotyl growth via the evening complex EC and its inhibition of the PIF4 and 5 transcription is shown at bottom right. Central loop (thick green arrows and thick red – I) consists of clock genes CCA1 and LHY (black boxes with dented arrows, TAIR nomenclature for Arabidopsis), their products CCA1 and LHY (red – I), and the clock gene TOC1 and its product TOC1 (green oval). TOC1 represses (thick red a) CCA1 and LHY. The products (small green ovals) of ELF3, ELF4, and LUX form the evening complex EC and are the major elements of the evening loop. In a third loop EC represses the morning genes PRR7 and PRR9 (red ovals). Their products PRR7 and PRR9 inhibit the expression of CCA1 and LHY, and they regulate PRR7 and PRR9 positively. In a fourth loop GI (small green oval) induces TOC1 in the evening and is negatively regulated by TOC1, CCA1, and LHY. A fifth loop links the circadian clock to the dynamic chromatin remodeling via JMT30 (not shown). Inset shows an Arabidopsis thaliana plant exposed at two different times in a space experiment (Johnsson et al. 2009) (After Brown et al. (2012), Pokhilko et al. (2012), Sawa et al. (2007))
With this model light responses like those resulting from changing the light period from 2 to 22 h in a 24 h day, or skeleton photoperiods (a short light pulse at the beginning and at the end simulates the LD period; see Pittendrigh 1964), have been successfully simulated (Troein et al. 2011; Thommen et al. 2012).
The Ostreococcus clock is insensitive to fluctuations in light intensities (clouds, different depths in the water). This is due to the phase response curve (see Sect. 18.1.5), which possesses a broad dead zone in which light does not shift the phase of the rhythm. Only when light hits the oscillator outside the dead zone, it is sensed and re-entrains the oscillator. While synchronized with the LD cycle, the oscillator is blind to light. The light intensities might fluctuate considerably without affecting the clock (Pfeuty et al. 2012).
In Ostreococcus tauri a histidine kinase LOV-HK was found as a new class of eukaryotic blue-light receptor (Djouani-Tahri et al. 2011a). It is related to the large family of LOV-histidine kinases found in prokaryotes. It senses blue light and is under circadian control. But it is also important for the function of the circadian clock under blue light independent of its blue-light-sensing property. Another histidine kinase, rhodopsin-HK (Rhod-HK), probably senses longer wavelengths than LOV-HK (Pfeuty et al. 2012). Using long- and short-wavelength photoreceptors (see Fig. 18.4) allows the cells to discriminate light variations due to depth changes from those due to the day/night cycling.
Cryptochromes are further blue-light receptors in Ostreococcus. Five genes of the Cry/photolyase family (CPF) were identified by Heijde et al. (2010). All five CPF members are regulated by light, and CPF1 and CPF2 display photolyase activity. CPF1 is furthermore involved in the maintenance of the Ostreococcus circadian clock.
The molecular basis of light-dependent control of cell division in Ostreococcus was studied by Moulager et al. (2010, 2007). They found that the clock regulates directly cell division independently of the metabolism. The transcription of the main cell cycle genes such as cyclins and kinases was under circadian control.
Studies by van Ooijen et al. (2011) and O’Neill et al. (2011) in animals and plants revealed that posttranslational events such as rhythmic protein modifications are also involved in circadian timing. Non-transcriptional mechanisms are able to sustain circadian timing in Ostreococcus, although normally it functions together with transcriptional components. Targeted protein degradation in the circadian mechanism seems to play a central role. It was proposed that the oldest oscillator components are non-transcriptional, as in cyanobacteria, and conserved across the plant and animal kingdoms.
Another much studied unicellular green alga is Chlamydomonas reinhardtii (Chlorophyta > Chlorophyceae > Volvocales > Chlamydomonadaceae). It is of 14–22 μm size and found worldwide in freshwater but also in the soil. It is used as a model organism for molecular biology; for studies of flagellar motility, chloroplast dynamics, biogenesis, and genetics; and also for its circadian clock. Phototactic movement (swimming towards light) is driven by the flagellae and controlled by a circadian clock (Bruce 1972; Gaskill et al. 2010). This clock furthermore controls UV sensitivity (Nikaido and Johnson 2000), chemotaxis (Byrne et al. 1992), adherence to glass, cell division (Goto and Johnson 1995; Bruce 1970), and starch and nitrogen metabolism. The period length is temperature compensated, as in all circadian rhythms.
Light pulses with a certain fluence rate and wavelength at the breakpoint between delay shifts and advance shifts reduce the amplitude of the rhythm to such a degree that the clock stops its oscillation and reaches a “singularity.” The results are interpreted in the context of limit cycle models of circadian clocks and are used to suggest new strategies for measuring action spectra of light-induced clock resetting (Johnson et al. 1992).
The phase-shifting effect of light pulses was studied by Johnson et al. (1991) and Kondo et al. (1991). Blue and red light resets the circadian clock. PHY homologues of higher plants are not present, but homologues of cyanobacterial PHY (CHLAMYOPSIN5 and MIXED LINEAGE PROTEIN KINASE) and the cyanobacterial kinase CikA and two CRYs with significant homology to the CRYs from plants and animals were found (Mittag et al. 2005). Whether photoreceptor proteins ChR-1 and ChR-2 in the eyespot of Chlamydomonas reinhardtii at the outer chloroplast envelope are involved in the phase shift is not known (Hegemann 2008).
The genome of Chlamydomonas reinhardtii is entirely sequenced (Matsuo and Ishiura 2011). Subproteome and phosphoproteome analysis were and are used for finding photoreceptors (Boesger et al. 2009). Much work is devoted to the clock network and to clarify how clock-related factors are interconnected. System biology approaches are used for this means, metabalance (May et al. 2009; Manichaikul et al. 2009) and functional proteomics (Wagner and Mittag 2009). Based on elementary flux mode analysis, Schäuble et al. (2011) combined sequence information with metabolic pathway analysis and included circadian regulation. They are able to predict changes in the metabolic state and hypothesize on the physiological role of circadian control in nitrogen metabolism. Review articles of the circadian rhythm of Chlamydomonas are by Schulze et al. (2010), Brunner et al. (2008), Mittag and Wagner (2003), Werner (2002), Suzuki and Johnson (2002), and Mittag (2001), and for modeling the Chlamydomonas clock, see Jacobshagen et al. (2008), Matsuo and Ishiura (2011), and Breton and Kay (2006).
The germination efficiency of zygospores of Chlamydomonas reinhardtii depends on the photoperiod and is higher in long days and lower in short days (Mittag et al. 2005; Suzuki and Johnson 2002). A CO homologous gene (see Sect. 18.4.3) is influenced by day length and by the circadian clock, being more expressed in short photoperiods. Under these conditions algae accumulate more starch and express genes which coordinate cell growth and division (Romero and Valverde 2009). CO orthologs might represent ancient regulators of photoperiodic events. They arose early in the evolutionary lineage leading to flowering plants (Serrano et al. 2009).
18.4 Light Effects on Circadian Clocks in Plants: Arabidopsis
To grow and develop successfully, it is essential for plants to synchronize metabolism and physiology with the environment and the seasonal changes. Quite a number of plants can also synchronize the opening of their petals with the activity of visiting insects such as bees and butterflies, which in 1751 led Linnaeus to construct a flower clock: various plants are planted as a kind of clock circle in a round garden bed in such a way that their flowers open or close at the corresponding day- and nighttime. A circadian clock is often responsible for it, and this clock allows also to measure day length to anticipate changes in the surrounding, be it daily or seasonal ones. In both cases, light signals are the main pathways to transfer these transitions to the plant.
Many processes in plants are directly affected by light such as photosynthesis (Chaps. 16 and 17), photomorphogenesis, and photoperiodism (Chap. 19) and stomatal movements. The biological clock processes do not immediately show their light dependence but are synchronizing the plants to the external light program and its period (or rhythm). In general, multiple photoreceptors sense the quality and quantity of light in the environment (Chaps. 12, 13, and 14). Red light is sensed by phytochromes (PHYs), blue light by cryptochromes (CRYs) and ZTL of the ZTL family (ZTL and FKF1). Furthermore phototropins (absorbing in the UV-A/blue, Heijde and Ulm 2012) and UVR8 absorbing in UV-B (Rizzini et al. 2011) are used by plants. This system of different photoreceptors with partially antagonistic functions and overlapping action spectra detects radiation of different wavelengths over a wide spectral range. The PHY and CRY photoreceptors interact with each other and respond at different expression levels and localizations, allowing a simultaneous response to two or more environmental parameters (van Zanten et al. 2012; Kami et al. 2010).
Arabidopsis thaliana plants are well suited for studying the influence of light on circadian clocks, since many biochemical, physiological, and developmental events are under their control. Furthermore, numerous mutants are known, among them quite a number which affect the clockwork, clock inputs, and clock outputs. Besides mutants in which the function of the photoreceptors is affected, others are known, in which the transfer of the light-induced signals is changed (Strasser et al. 2010; Yanovsky et al. 2001).
For continuous recording it has been of much advantage to use a construct of the firefly luciferase gene with a promoter of the cab2 gene which is under control of the circadian clock. The method allows monitoring of circadian rhythms in whole plants but also in different tissues of the plant by recording the bioluminescence with a sensitive camera. It also makes screening of mutations in the clock easy by looking for aberrant temporal patterns of luciferase expression (Millar et al. 1995).
Plants contain circadian clocks in each cell. The clock components consist of interwoven feedback loops, outputs to the clock-controlled genes and driven processes, and inputs from the synchronizing time cues such as light (Fig. 18.5 and Sect. 18.4.1). In addition, posttranscriptional and posttranslational events contribute to the generation and maintenance of the rhythms (McWatters and Devlin 2011; Harmer 2009). Multiple photoreceptors are used by the plants to synchronize the circadian clock. The transduction pathways from light perception to the clock are apparently quite closely linked to the clock mechanism.
Whereas in animals a hierarchy of clock units with circadian centers is the rule, in plants cellular circadian clocks run autonomously in the different tissues and organs and are synchronized by the LD environment. Plant hormones might coordinate these local clocks. More details will be presented in Sect. 18.4.2.
Temperature compensation and the mechanism, by which the circadian clock of Arabidopsis avoids changes in period length under different environmental temperatures, has been discussed by Troncoso-Ponce and Mas (2012), Salomé et al. (2010), and Portolés and Más (2010); see also Hatakeyama and Kaneko (2012), Bodenstein et al. (2011), and Eckardt (2010).
18.4.1 Clock Mechanism and Clock-Controlled Genes
The Arabidopsis clock is a multi-feedback system with various coupled loops, which receives external inputs such as light and dark signals and possesses output pathways to control transcription, translation, and many physiological and metabolic processes in the plant. The interlocked loops are thought to make the clock more robust, more accurate, and less affected by disturbances of the environment but allow also more flexible inputs in different climates (Harmer 2009; Michael et al. 2003). Figure 18.5 shows a model of the clock (Pokhilko et al. 2012; Lu et al. 2011; Nakamichi 2011) including light inputs (Kolmos et al. 2011; Kim et al. 2007). From the various outputs of the clock (Kami et al. 2010), only the pathway to flowering and hypocotyl growth are indicated (Lu et al. 2012; Kunihiro et al. 2010).
The central loop consists of the clock genes CCA1 and LHY and their products CCA1 and LHY and the clock gene TOC1 and its product TOC1, a DNA-binding transcription factor. CCA1 and LHY are expressed with peaks shortly after dawn and repress TOC1 expression by binding to the evening element in its promoter. This is the morning oscillator. TOC1 is a DNA-binding transcription factor which peaks at dusk (Gendron et al. 2012). It represses (not activates, as shown by Pokhilko et al. 2012) CCA1 and LHY.
The ELF3, ELF4, and LUX genes and their products ELF3, ELF4, and LUX, forming the evening complex EC, regulate clock gene expression at night. LUX binds directly to the promoters of the target genes CCA1 and LHY, but ELF3 and ELF4 proteins are important for the function of the EC complex. The CCA1 and LHY proteins inhibit the expression of the ELF3, ELF4, and LUX genes thus closing the loop (Chow et al. 2012; Herrero et al. 2012; Nakamichi et al. 2012; Troncoso-Ponce and Mas 2012).
In a third loop, EC is also connected to the morning genes PRR7 and PRR9 by repressing them. PRR7 and PRR9 inhibit the expression of CCA1 and LHY by binding to their promoters. CCA1 and LHY in turn regulate PRR7 and PRR9 positively.
In a fourth loop, GI induces TOC1 in the evening and is negatively regulated by TOC1, CCA1, and LHY. TOC1 is rhythmically and light dependent degraded by combining with GI and ZTL, and ZTL serves as a light receptor. There is also a dark-dependent protein degradation (Adams and Carré 2011).
A fifth loop was proposed based on the finding that about 90 % of the transcripts of Arabidopsis do cycle. This large number is supposed to be due to a dynamic chromatin remodeling by the circadian clock via jumonji C (JmjC), a domain-containing protein, acting as histone demethylase (Lu and Tobin 2011).
The various clock genes, their products, and their interactions are studied intensively, and a final model has not yet been gained. Furthermore, the molecular composition of circadian clocks can differ between various cell and organ types. Thus, PRR3 modulates TOC1 stability in vasculature cell types, but not in others; CCA1 and LHY are not able to inhibit TOC1 expression in dark-grown roots (further examples in Harmer 2010; Hotta et al. 2008).
A circadian clock provides evolutionary advantage by increasing fitness and adaptation. This is shown in Arabidopsis thaliana plants which are arrhythmic (by overexpressing CCA1) by a reduced viability under extreme short days (Green et al. 2002) and a higher susceptibility to pathogen infections (Wang et al. 2011). Furthermore, the ztl mutant with a longer period and the toc1 mutant with a shorter period than the wild type contain more chlorophyll and produce more carbohydrates and biomass when grown under LD matching the length of their circadian period (Dodd et al. 2005).
18.4.2 Light as the Main Time Cue of the Circadian Clock
Whereas photoreceptors and the light-regulated responses including gene expression have been intensively studied, the signal transduction components are much less known. A large number of signaling components must exist, which are affected by external and internal factors. Both genetical and biochemical approaches are used to clarify these transduction pathways and modes.
Light is the most important time cue for synchronizing the circadian clock of Arabidopsis with the environmental 24 h day. The phase of the clock can be shifted by applying short light pulses in plants kept under constant conditions. The rhythm is delayed, if the light is applied in the first part of the subjective night, and is advanced, if the light is applied in the second part. The phase shifts can be plotted as a phase response curve (Covington et al. 2001). In a similar way, LD cycles synchronize the clock by phase shifting. Red and blue light are most effective, suggesting that PHY and CRY photoreceptors are involved, as indicated by the flashes in Fig. 18.5.
In addition to resetting the phase, light modulates also the period of the clock. Under LL conditions, but not under DD, phy and cry mutants have a longer period. This indicates that they are within the light input pathway and not part of the central oscillator itself. Otherwise the longer period should also show up under DD. The same applies for mutations in PRR7 and PRR9, where only LL causes a long period, apparently by affecting pathways. However, the ztl mutant has a longer period both under LL and DD, which suggests that ZTL plays a role within the central clock. Phototropin1 mutants do not affect the circadian rhythm.
Light signals can also entrain or reset the circadian clock at several points at the transcriptional, posttranscriptional, and posttranslational levels in the various clock loops by affecting the expression, activity, stability, or localization of oscillator components (Kozma-Bognár and Káldi 2008). Transcription of the clock genes LHY, CCA1, PRR7, PRR9, and GI is upregulated by red, far-red, and blue light. However, the corresponding mRNAs of LHY and CCA1 are degraded by light. In this way timing and entrainment are improved (Jones 2009; Yakir et al. 2007). Translation of LHY is induced by light, and its level depends on the available mRNA, which peaks at dawn. Cyclic removal of clock proteins is crucial for oscillator function, as mentioned before. Thus, proteolysis plays an important role and preferentially takes place in the dark. TOC1 degradation is controlled by the F-box protein ZTL, which binds to it light independent, but the degradation rate is increased in the dark. The mechanism of GI degradation is still unclear.
Which intermediate factors link these photoreceptors with the circadian components and how they do it is not well understood. For instance, the TOC1 protein in the first FBL is stabilized by light and degraded in darkness by an ubiquitin ligase complex, which is regulated by PRR3 and GI. The accumulation of ZTL over the day decreases TOC1 protein levels at the onset of night and increases the robustness of the transcriptional feedback loops of TOC1. Likewise, PRR5 protein accumulates in the evening before it is degraded by ZTL. It is suggested that PRR5, TOC1, GI, and ZTL form a functional unit in the evening loop (see Fig. 18.5, Harmer 2010; Jones 2009). Other candidates for light inputs are ELF3 and ELF4 in one of the feedback loops.
Many questions concerning the Arabidopsis clock and how it interacts with light are still open. For instance, most components of the clock act in the nucleus as transcriptional regulators and are together with light-signaling proteins co-localized in nuclear foci (figure 4 in Herrero and Davis 2012) generating rhythmic transcript accumulation. Further information and references can be found in Jones (2009) and Harmer (2010).
Output of the circadian system affects gene expressions, but gene expression is also directly affected by light and not only via the clock (immediate light effects). The clock output can furthermore gate the light input according to the phase of the clock by, e.g., affecting the phy and cry genes at the level of transcription (Toth et al. 2001) or by other types of rhythmic regulation such as the regulation of leaf position. This feature has been termed “Zeitnehmer” (German for time taker, in contrast to “zeitgeber,” time giver or time cue).
Under natural conditions the changing day lengths during the course of the year has to be taken into account. That is, neither dawn nor dusk drive the rhythm, but at least two signals must be used. For more information see Chap. 19 and Sect. 18.4.3.
Non-photic time cues are also used by the plant. Temperature cycles entrain the clock. Imbibition of Arabidopsis seeds sets a circadian clock which is insensitive to light during the first 60 h. From the 36th hour onward, light initiates a second rhythm which runs independently of the imbibition rhythm (i.e., the output, namely, CAB2 and CAT2, shows the two rhythms superimposed). Light applied after the 60th hour synchronizes the two rhythms (Kolar et al. 1998).
18.4.3 Photoperiodism
Many plants use the seasonal change in day length as a signal for growth (Niwa et al. 2009) and flowering (Millar 1999). In contrast to the effect of light on the circadian clock of Arabidopsis, the mechanism of photoperiodic induction of flowering in this plant is known in considerable detail (Srikanth and Schmid 2011; Amasino 2010; Imaizumi 2010; de Montaigu et al. 2010; Michaels 2009).
Plants measure day length in the leaves by a circadian clock. Depending on the type (long-day, short-day, long-short-day, short-long-day plant), the photoperiodic effect occurs under long days; short days; first long days, then short days; or first short day, then long days, respectively. In this way, a developmental switch from the vegetative to the reproductive stage is activated.
Day-length sensing in Arabidopsis occurs by an external coincidence mechanism, as predicted already in 1936 by Bünning (1936). It operates by the circadian and light regulation of CO in the leaves, which under appropriate day length induces FT expression, the long-searched florigen (see Fig. 18.5). It is transported to the apical meristem in the shoot, where it promotes flowering. FT combines at the apex with FD, which is present there, but inactive without FT. The FT-FD complex initiates reproductive development (flower evocation). Flower meristem identity genes are activated and flowers are induced according to the ABC (DE) model (Litt and Kramer 2010). Both CO and FT expressions are controlled by a group of transcription factors with overlapping functions (Lu et al. 2012; Imaizumi 2010).
A further prerequisite for flowering of some plants is vernalization, by which a prolonged cold period results in meristem competence to flower through the epigenetic repression of the floral repressor FLOWERING LOCUS C (Michaels 2009).
The CO-FT interaction is conserved among plants (Srikanth and Schmid 2011). The photoperiodic responses are conferred by the same genetic pathway in the long-day plant Arabidopsis thaliana and the short-day plant rice Oryza sativa. But the functions differ (Hayama and Coupland 2003).
18.5 Fungal Clocks and Light Resetting: Neurospora
Neurospora crassa (fungi: Ascomycota, Ascomycetes, Sordariales, Sordariaceae) was originally thought to be a tropical fungus (see page 34) but is nowadays found all over the world. It is a model organism for genetic and physiological studies, because it is easy to grow and has a haploid life cycle, which facilitates analysis of genetic recombination. The genome of the seven chromosomes is entirely sequenced (43 megabases long, includes approximately 10,000 genes), and strains of knockouts are available for most identified genes (see Colot et al. 2006 and link: [http://www.fgsc.net/]). A large collection of mutants is available and continuously updated (for further information see link: [http://www.fgsc.net/2000compendium/NewCompend.html||Collectionofmutants]). Transformation methods are routinely used, and molecular genetics methods such as the use of an inducible promoter for dosage control and RNAi for gene silencing are available (Ziv and Yarden 2010). Imaging techniques are applied (Larrondo et al. 2012; Castro-Longoria et al. 2010; Gooch et al. 2008) including the use of luciferase, GFP, and mCherry. For an overview of modern molecular biological approaches used in Neurospora studies, see Jinhu and Yi (2010) and Dunlap and Loros (2005).
Neurospora crassa was used already since the 1950s for studying circadian rhythms (Pittendrigh et al. 1959). The formation of aerial hyphae and asexual macroconidia (generation cycle in Neurospora; see Springer 1993) is under circadian control but manifested in many other functions. Rhythmic conidiation shows up in bands which are formed while the mycelium grows over the agar surface in “running tubes” (see link: http://geiselmed.dartmouth.edu/dunlaploros/research/media.php). The period and phase shifts can be determined simply by using a ruler and time markings at the growth front, but more accurate and elaborate imaging methods are also applied (Hogenesch and Ueda 2011; Dunlap and Loros 2005; Morgan et al. 2003).
About 20 % of the genes of Neurospora are clock controlled (Smith et al. 2010). This allows modulation of numerous biochemical and physiological processes in a circadian fashion. Pharmacological and genetical approaches have been used in order to unravel the circadian system which underlies overt rhythms such as conidiation (Lakin-Thomas et al. 1990). By the way, in Neurospora cytoplasm and nuclei stream through the colony (called syncytium) and the circadian rhythm stays in synchrony.
In the following the circadian system of Neurospora and its constituents will be described first (see Sect. 18.5.1 and Baker et al. 2012 for a recent review and a historical account, further reviews by Lakin-Thomas et al. 2011, Jinhu and Yi 2010, Brunner and Káldi 2008, de Paula et al. 2007, Dunlap et al. 2007, Loros et al. 2007). Thereafter (Sect. 18.5.2) it is shown how the circadian system is entrained (Merrow and Roenneberg 2007) by temperature (Brunner and Schafmeier 2006; Diernfellner et al. 2005) and light (Schafmeier and Diernfellner 2011; Price-Lloyd et al. 2005; Kozma-Bognár and Káldi 2008) and which photoreceptors are used (Chen et al. 2010). The outputs of the clock are described (Sect. 18.5.3) and finally a photoperiodic reaction of Neurospora is briefly mentioned (Sect. 18.5.4).
18.5.1 The Circadian System of Neurospora and Models
The circadian system of Neurospora consists of interwoven negative and positive feedback loops made up by a complicated interplay of various factors which affect the expression and function of the core clock components transcriptionally and posttranscriptionally. Phosphorylations and dephosphorylations of clock components ensure the robustness, precision, and entrainment of the circadian system and account for the complexities in rhythmic behavior (Baker et al. 2012; Schafmeier and Diernfellner 2011; Lakin-Thomas et al. 2011), modeling: Tseng et al. (2012).
A transcription-translation oscillator (TTO) has been proposed which possesses all the formal properties of a true circadian oscillator with light entrainment and temperature compensation. The molecular mechanism has been studied intensively (review: Vitalini et al. 2010; Dunlap et al. 2007; Loros et al. 2007). The gene frq, its mRNA, and product FRQ are essential components and belong to a negative limb of a feedback loop. In this loop frq expression is inhibited by the transcription factor WCC (white color complex). Details are given in Fig. 18.6 and a time line of the clock events in figure 4 of Baker et al. (2012).

Model of the biochemical and molecular processes of the Neurospora clock. The main feedback loop (thick lines) consists of frq (black box with dented arrow indicating transcription) which expresses FRQ (red oval) in the late night and early morning while activated by a WCC transcription factor (consisting of WC-1 and WC-2, yellow ovals). This is the positive arm in the feedback loop. In the negative arm FRQ dimerizes and forms a complex with FRH (a RNA helicase, not shown), which inhibits WCC (thick red – I). From noon to evening kinases phosphorylate WCC thereby inactivating it in the early night (not shown). In the late night FRQ interacts with ubiquitin ligase FWD-1 (blue oval) and is degraded (light red oval with dashed border) in proteasomes. Now the frq promoter can transcribe again being reactivated by newly synthesized WCC. In a second feedback loop (thin lines) VVD expressed by vvd inhibits (thin red – I) WCC. This feedback loop is responsible for adaptation under longer blue-light exposure (L-adaptation). Phase shifting of the circadian rhythm by light pulses and other light effects (e.g., carotenoid formation) occur via the blue-light receptor WCC (L-activation) (Modified from Brown et al. (2012))
In addition to the FRQ-WCC oscillator, there might be a FRQ-less oscillator (FLO) which is independent of FRQ and WCC (Lakin-Thomas et al. 2011) and coupled to the feedback loop shown in Fig. 18.6. Strains lacking FRQ (frq 0) or lacking WC-1 (wc0) still exhibit circadian rhythms in a choline-requiring strain depleted of choline. The same has been reported for nitrate reductase activity under DD or LL conditions. Apparently in both cases an FLO is the cause for this nutritionally induced rhythm in the absence of an intact FRQ protein.
Furthermore, a FRQ-less oscillator which requires WC (WFLO) but is independent of FRQ was proposed by de Paula et al. (2006) and Correa and Bell-Pedersen (2002). This oscillator requires WC-1 and WC-2 for activity. The WC-1 level is rhythmic in the absence of FRQ, indicating that this WFLO generates the rhythm of WC-1. The rhythm can be observed under DD and LL conditions. In contrast to the FRQ-WCC oscillator, its rhythm is apparently not inhibited by high or low light levels. This oscillator and the FRQ-WCC oscillator may interact with each other through their common WC proteins.
How to integrate these diverse oscillators that do not fit the TTO mechanism? This has been discussed in the context of the circadian system by Lakin-Thomas et al. (2011) (see their figure 3.2). FLOs might represent more than a dozen metabolic oscillators, which are not connected to the circadian system. Or, according to de Paula et al. (2007), there are multiple FLOs, which together with the FRQ/WCC oscillator form a network of coupled oscillators. But individual FLOs may drive a particular output. The FLOs may also function upstream or downstream of the FRQ/WCC oscillator. They need to bypass FRQ/WCC to drive the conidiation rhythm and biochemical rhythms when FRQ and WCC are disabled (Roenneberg and Merrow 1998). Or a single FLO is the central rhythm generator, which is mutually coupled to the FRQ/WCC, supplying stability, period control, and rhythmic input (Li and Lakin-Thomas 2010).
Brody et al. (2010) proposed that the many conditions and mutations leading to FLOs may converge on a pathway that includes ROS and the activation of a RAS-cAMP protein kinase. A cAMP pathway and metabolic regulation is involved also in other organisms (Bass and Takahashi 2010).
These findings suggest that the FRQ/WCC oscillation is not independent of the FLOs, which should be taken into account with respect to the TTO shown in Fig. 18.6, where FLO and WFLO are omitted.
Another question is how typical the molecular composition of the Neurospora clock is among the fungi. Whereas WC-1 and WC-2 are conserved, FRQ is less so, and other proteins might replace it as a negative element but work in a similar manner (Salichos and Rokas 2010; Dunlap and Loros 2006).
18.5.2 Entrainment and Photoreceptors of the Circadian System
The clock components described above can explain the molecular bases of the inputs and outputs. Temperature changes and light pulses or LD cycles are able to entrain the clock. We will concentrate on light but mention briefly temperature effects.
Moderate temperature changes of 1–2 °C are already sufficient for entrainment (Liu et al. 1998). The amount of FRQ depends on the phase of the oscillator and on the environmental temperature. Changing temperature corresponds to shifts in clock time, because the amount of FRQ is immediately changed within the clock mechanism.
As is usual in circadian rhythms, the period is only slightly dependent on a constant environmental temperature. This temperature compensation is valid in Neurospora between 30 and 36 °C. Many different gene products influence temperature compensation, but frq plays a major role: FRQ stability is involved, and CK2 by its phosphorylation of FRQ. Other kinases and phosphatases of the clockwork are not involved (Mehra et al. 2009). The temperature compensation is seen at the posttranslational level and due to the ratio and abundance of a small and a large isoform of FRQ, which are expressed in a temperature-dependent fashion. At lower temperatures (20 °C) the small and large isoforms are equal, at higher temperatures (28 °C) more of the large isoform as well as more FRQ is present (Diernfellner et al. 2007).
WC-1 is not only a clock component but also a blue-light photoreceptor: it is responsible for all light responses in Neurospora. There are three types of light-induced circadian responses observed: first, LL suppresses the circadian modulation of conidiation. Instead, conidia are formed all the time. In DD or safelight such as red, conidiation occurs in a circadian pattern (period length 22 h). Second, a single brief pulse of light applied in DD phase shifts the conidiation rhythm either by advancing or by delaying it. An action spectrum of light which phase shifts the rhythm shows maximal effects at 465 nm (Dharmananda 1980). A light pulse applied at late subjective day and early subjective night delays the rhythm; a light pulse at late subjective night and early subjective morning advances the rhythm. Third, LD cycles (or periodic light pulses) entrain the circadian clock. Below it will be shown how these effects of light – the rhythm annihilating one of LL, the phase shifting one of pulses, and the entraining one – are related in terms of molecular events. The light input pathways are also well understood.
In contrast to plants, Neurospora is sensitive to the blue range only and is blind to light beyond 520 nm. Light activates hundreds of genomic regions, about 20 % of all genes (Schafmeier and Diernfellner 2011; Smith et al. 2010). At least 3 % of the genes of Neurospora are light inducible, shown by microarray analysis (Lewis et al. 2002). WCC controls the expression of 24 transcription factor genes including those of the circadian oscillator.
Blue light induces frq, and wc-1 and wc-2 are required for it (the double mutant white color wc-1 and wc-2 is blind for light). The light is received by the FAD-associated LOV domain of WC-1 and changes its conformation allowing WCC to bind to LREs in promoters of immediate light-induced genes, i.e., induction begins in no more than 5 min (Lewis et al. 2002). The WCC-LRE complex enhances the capacity of WCC to activate transcription. Two different WC-1/WC-2 complexes are formed, a smaller one which binds to LREs in the dark and activates frq expression in the dark (and light exposure reduces its binding) and a larger one which replaces the smaller one after light exposure. The larger one is responsible for the light-induced activation of frq transcription.
This light-induced WC/LRE binding corresponds to the light-induced entrainment of the clock and phase resetting (Baker et al. 2012; Crosthwaite et al. 1997). Since frq expression cycles, the effect of light on the oscillator depends on the phase: when FRQ levels are low or rising before subjective dawn, a light pulse increases frq mRNA and advances the clock phases with highest frq expression in the later morning. If FRQ levels decrease, light will again increase frq mRNA, which will now delay the phase.
The blue-light chromophore that mediates binding of the larger WCC to frq LREs is FAD (and not FMN, discussed in Liu 2003). A mutant in which the LOV domain is removed does not show light responses anymore (He et al. 2002) and is arrhythmic in LD cycles and in DD. The circadian clock of this mutant cannot be entrained by light, but temperature cycles do entrain it. Deleting the WC-1 LOV domain has thus separated the light and dark function of WC-1.
VVD is another photoreceptor in Neurospora (Belozerskaya et al. 2012; Zoltowski et al. 2007). It is not essential for clock function, but modulates all its light responses. In vvd mutants light-induced gene expression is elevated (leading to, e.g., higher carotenoid synthesis and giving the mutant a vivid orange color), the phase of the circadian clock altered, and light adaptation partially lost. Furthermore, circadian gating of light induction of gene expression is affected in vvd (induction is higher in the subjective morning).
VVD is responsible for photoadaptation in Neurospora by reverting the elevated levels of light-induced gene transcription within 2–4 h to preinduction levels. In this way Neurospora can detect changes in light intensity and not just lights on or lights off (Schwerdtfeger 2003). VVD interacts in the nucleus with the WC-1 in the WCC and reduces its ability to activate transcription. Increasing light yields more VVD and stronger inactivation of the newly activated WCC (Chen et al. 2010; Hunt et al. 2010). The expression of vvd is clock controlled and it gates the input to the clock (Zeitnehmer). VVD allows the clock to take phase cues from dusk (Elvin et al. 2005), to avoid any WCC induction by moonlight (Malzahn et al. 2010), and it contributes to temperature compensation of the clock (Hunt et al. 2007).
There might be further photoreceptors as suggested by the genome sequence of Neurospora (Belozerskaya et al. 2012). It indicates a putative cry gene, two phy-like genes, and two genes of the Archean rhodopsin. Their functions are unknown. The finding of Dragovic (2002) that under certain circumstances (high light intensities) the conidiation of wc-2 mutants is still driven by the LDcycle suggests the existence of a wc-independent photoreceptor.
18.5.3 Outputs of the Circadian System
How the time information of the clock is used to regulate the various metabolic, physiological, and developmental overt rhythms in the cell is not yet well understood at the molecular level (Vitalini et al. 2006). The central clock affects numerous genes, which are not part of the clock, but controlled by it (clock-controlled genes, Loros et al. 2007). Primary clock-controlled genes are directly regulated by the WCC, and secondary clock-controlled genes are further downstream from the clock. An additional regulation can occur at the mRNA level through message stability rather than production (Guo et al. 2010). Chromatin remodeling, posttranscriptional, translational, and posttranslational mechanisms could provide further control of circadian rhythmicity.
Neurospora as a pioneer organism for biochemical investigations offers much information on metabolic pathways (Davis 2000), and chronobiologists take advantage of it by studying the circadian influences (Bass and Takahashi 2010; Harrisingh and Nitabach 2008; Hastings et al. 2008). They are using a systems biology approach to monitor rhythmicity in RNA, proteins, and the metabolism (for transcriptome, proteome, and metabolome, see Dong et al. 2008) under different lighting conditions and under specific genetic backgrounds. Further circadian outputs concern development (e.g., cell cycle and conidiation, Correa and Bell-Pedersen 2002), organelles, transport and signaling pathways (Baker et al. 2012), and osmotic stress (Vitalini et al. 2007).
Screens for clock-controlled genes were performed in Neurospora crassa and so far over 400 (Dong and Golden 2008; Correa et al. 2003; for functional categories see table 1 in Dunlap and Loros 2004) have been found by using different methods (review Bell- Pedersen 2000). They are being characterized at the molecular level (Lakin-Thomas et al. 2011). As much as 25 % of the Neurospora transcriptome is under clock control. Most of the ccg expression peaks just before dawn, but there are others which show maximal expression at other phases of the day (Correa et al. 2003).
If each of the 24 transcription factors regulates about 20 downstream genes and if WCC directly binds to regulate rhythms in about 180 additional morning-specific targets, the various phases of all of the known clock-controlled genes would be taken care of.
The outputs of the clock can feed back to input pathways, as exemplified by the clock-controlled component VVD on light input (Elvin et al. 2005). Such output to input loops can provide certain time-of-day-specific gates.
18.5.4 Photoperiodism
The circadian system seems to control also photoperiodic propagation and reproduction (conidiation, protoperithecia formation) of Neurospora crassa (Rémi et al. 2010; Tan et al. 2004; Roenneberg and Merrow 2001). Without FRQ photoperiod cannot be measured, indicating the role of the circadian system in the photoperiodic time measurement. The strains of Neurospora crassa have been isolated mainly from tropical areas. The survival vale of photoperiodic reactions in such strains is doubtful. However, Jacobson et al. (2004) found strains as far north as Alaska. A temporal segregation of asexual and sexual reproduction with conidiation in March and perithecia in July was described by Pandit and Maheshwari (1994).
18.6 How Light Affects Drosophila’s Circadian System
We are now coming to animals, which differ fundamentally from the examples treated so far. They possess a central nervous system that controls behavior and central circadian clocks brain. If light is going to synchronize these clocks, it either has to reach the clocks directly in translucent specimens or indirectly via specialized photoreceptor organs or via both pathways.
The fruit fly Drosophila is an example of an insect using both pathways. It has many advantages as an experimental animal such as easy rearing and a short generation time. It is well known genetically and a large number of mutants is available. Drosophila is amenable to genetic and molecular methods. For these and other reasons, this insect was and is used also for studying circadian rhythms, especially eclosion of the flies out of the puparium (a case produced in the last larval stage, in which metamorphosis from the larva to the fly takes place) and locomotor activity of the adults. Many mutations affecting the circadian clock, the photoreceptors, and the photoreception are known. Therefore, the effects of light on the circadian system could be studied intensively and successfully.
General reviews on the circadian clocks of Drosophila (Allada and Chung 2010; Tomioka and Matsumoto 2010), their genetics (Hardin 2011), molecular mechanism (Duvall and Taghert 2011; Hardin 2011; Weber et al. 2011), and their location and neurobiology (Yoshii et al. 2010, 2012; Hermann et al. 2012; Rieger et al. 2009) are available. Special reviews on the effect of light on the circadian rhythm and the pathways to the circadian system are by Peschel and Helfrich-Förster (2011), Barth et al. (2010), and Choi and Nitabach (2010). The output pathways of Drosophila’s circadian system are reviewed by Tomioka et al. (2012), Helfrich-Förster et al. (2011), and Frenkel and Ceriani (2011).
18.6.1 Circadian Eclosion
After completing several larval stages, Drosophila forms a puparium in which pupation and metamorphosis into the adult stage takes place. Eclosion from the puparium occurs under the daily LD cycles in a restricted time window (gate) only during the early morning hours. A fly which is not yet ready to eclose uses the next gate on the following day. If a culture of Drosophila flies is transferred into constant conditions of darkness, eclosion occurs still rhythmically. This shows that eclosion in a population of flies is not just the response to the onset of light, but under control of a circadian clock.
The eclosion rhythm can be entrained by an LD cycle and phase shifted by a single light pulse. Therefore, photoreceptors must exist which transfer the signal evoked by light to the oscillator controlling eclosion. An action spectrum for phase shifting the eclosion rhythm with a single light pulse shows a broad maximum in the blue (457 nm) and further maxima at 375, 435, and 473 nm. Light of wavelengths above 540 nm is ineffective (Klemm and Ninnemann 1976). The responsible photoreceptors for eclosion are (1) the larval eyes (a pair of Bolwig organs close to the mouth hook, each consisting of 12 light sensitive cells which are retained in the adult eyelet) using rhodopsin (Malpel et al. 2002) and (2) lateral neurons (LNs) using CRY (Kaneko et al. 2000). Eclosion of mutants which lack extraretinal photoreception but possess functional larval eyes is still entrained. Mutants lacking CRY and the visual system cannot be entrained by light, but temperature cycles do entrain, demonstrating a functional oscillator system (Malpel et al. 2004).
Whereas the visual sensitivity of the compound eyes of flies reared on a carotenoid-free diet is decreased by three orders of magnitude, the photosensitivity of the circadian eclosion rhythm is not affected. Furthermore, the eclosion rhythm of mutants lacking compound eyes was still synchronized by light. The compound eyes in the metamorphosed fly in the puparium are thus not needed to phase shift and entrain the eclosion rhythm.
How the circadian signals which control eclosion arise and reach their targets is reviewed by Helfrich-Förster (2005a, b). Recently, experiments were performed which try to simulate more natural conditions such as an LD pattern with gradually changing light intensities and varying day lengths. Under these conditions the eclosion rhythm of Drosophila seems to be more robust and shows seasonal variations (De et al. 2012).
18.6.2 Locomotor Activity and Sleep Are Controlled by Several Circadian Oscillators
Drosophila flies, like other animals, possess a multi-oscillatory system to control different events in a circadian way (Stanewsky et al. 1997). Circadian oscillators seem to be widespread throughout the different tissues and cells: using a construct in which the luciferase gene luc is fused to the regulatory upstream region of per (which is under circadian control), luminescence rhythms of the whole fly, of parts of the fly, and of cultured tissue can be monitored. Even the rhythms of cultured body parts are synchronized by LD cycles (Plautz et al. 1997), showing that the underlying peripheral oscillators are cell autonomous and photo responsive. Light entrains these oscillators directly, probably via CRY (Ivanchenko et al. 2001; Ito et al. 2008) (see Sect. 18.6.4).
However, behavior such as eclosion of the flies out of the puparium, adult locomotor activity, and sleep is driven by circadian centers in the brain. Adult activity rhythms can already be entrained by light applied in the first larval stage. The clock is apparently running at that time and throughout larval and pupal development and is resettable by light (Sehgal et al. 1992).
In an LD cycle, the flies are mainly active during the light period and sleep during the night. Sleep is defined as a quiescent state with reduced responsiveness to external stimuli (Bushey and Cirelli 2011). In order to sleep flies retreat to a preferred location, become immobile for periods up to 2.5 h, and do hardly respond to stimuli. Sleep is more abundant in young flies than in old ones and can be modulated by stimulants and hypnotics as in other animals (Hendricks et al. 2000; Shaw et al. 2000). Preventing sleep experimentally leads to a sleep rebound on the following night or day. During the day flies usually take a nap during midday. As a consequence, most activity occurs during the morning and evening. In DD the bimodal activity patterns are less pronounced, because morning and evening activity bouts come closer together and sometimes merge into one main activity bout. This is different under LL of low fluence rate, where both activity bouts are separated by a pronounced time of inactivity (nap) (Bachleitner et al. 2007; Yoshii et al. 2012). LL of low fluence rate does furthermore lengthen the free-running period of the flies in a dose-dependent manner (Konopka et al. 2007). At higher light intensities the flies become arrhythmic.
The clocks timing activity and sleep reside in about 150 clock neurons in the brain consisting of seven major groups, namely, three groups of dorsal neurons DNs (DN1−3) and of four groups of LNs (LNd, l-LNv, s-LNv, LPN) expressing different peptides/proteins (see Peschel and Helfrich-Förster 2011 and Fig. 18.7). Based on cell-specific ablation (Stoleru et al. 2004), respectively, targeted expression of PER (Grima et al. 2004), it was suggested that the ventral subset of the small LN cells (sLNs) is responsible for the morning activity, and the dorsal set of LNs (LNds) for the evening activity, thus providing a neuronal basis for morning and evening oscillators. The two oscillators were thought to be functionally coupled. Two oscillators could allow the fly to adapt to the seasonal changes of day length. A model of Pittendrigh and Daan (1976) predicts that the period of the morning oscillator M is shortened and that of the evening oscillator E is lengthened by light. As a consequence, M and E activity bouts are close together under short days and DD, but far apart under long days and under LL of low fluence rate. With increasing fluence rate of LL, the two activity bouts are predicted to free run with short and long periods, respectively, until the flies finally become arrhythmic (Daan et al. 2001).

Photoreceptors and neuronal clockwork of Drosophila melanogaster: looking toward the front of the brain from anterior, rhythm-relevant neurons and photoreceptors are shown. Light (L in yellow circle) for synchronization is received by the compound eye, the Hofbauer-Buchner (H-B) eyelets, ocelli, and via cryptochrome by some clock neurons themselves (each marked by a yellow point). The clock neurons consist of lateral neurons, LN (4 l-LNv, 4 s-LNv, one 5th s-LNv, 6 LNd, and 3 LPN), and of dorsal Nneurons (~15 DN1, 2 DN2, 30 DN3). Clock neurons marked in red are located in the anterior brain, the ones marked in blue in the posterior brain. Lateral and dorsal neurons as well as the clock neurons of the two brain hemispheres are connected with each other (gray arrows) and the majority of them project into the accessory medulla (aMe) – a small neuropil at the base of the medulla (pink area) that was first described in cockroaches as the site of the circadian clock (Homberg et al. 2003). The compound eyes express five different rhodopsins (see text) and are sensitive to blue, green, yellow, and red light (colored flashes). They transfer light information via the optic lobe (here visible: lamina and medulla) to the central brain (black arrow). The ocelli express only rhodopsin 2, which is blue-light sensitive. The H-B eyelets consist of four photoreceptor cells expressing rhodopsin 6, which has its absorption maximum in the green but responds also to red light. The H-B eyelets project toward the LNs. Direct synaptic contact was shown between the precursors of the H-B eyelets and the larval s-LNv (Wegener et al. 2004) but may exist also for the adult LNs and perhaps also for the DNs that project into the aMe. Cryptochrome in the clock neurons themselves is maximally responsive to blue light. The s-LNv (except the fifth) and l-LNv express the neuropeptide pigment-dispersing factor, PDF, that seems to be released in a rhythmic manner and is an important communication signal within the clock network. The l-LNv appears to play a special role in the light input pathway to the clock as well as in arousal and sleep (details see text) (After Peschel and Helfrich-Förster (2011), and Helfrich-Förster (2005a, b))
This was indeed found (Rieger et al. 2006). However, it is more complicated than expected. First, the blue-light photopigment CRY has to be knocked out to clearly see the internal desynchronization into the two free-running components. Second, the activity component with a short period did not only start from the M activity bout but in addition from the E activity bout, suggesting that the M cells may also control aspects of the E activity. The same is true for the E cells that can provoke M activity under certain conditions (Rieger et al. 2009; Sheeba et al. 2010). Third, the original simple assumption that the s-LNv controls the M component and the LNd controls the E component had to be refined. The two activity components are most likely controlled by variable subsets of DN and LN that interact in a complex manner depending on the environmental light and temperature conditions (reviewed by Yoshii et al. 2012). In spite of this obvious complexity, it is quite convincing that the brain clock of Drosophila is composed of M and E clock neurons that respond differently to light (and temperature) and control different aspects of behavior. M and E components are also involved in the circadian system of mammals (see Sect. 18.7.1).
Sleep occurring during the midday nap and the night appears to be controlled by the s-LNv (that are neither M nor E oscillators) and by sleep centers in the midbrain of the fly (Shaw et al. 2000). During neuronal activity the l-LNv promotes arousal. They are activated by the neuromodulators dopamine and octopamine and by light and inhibited by GABA. GABAergic inhibition of the l-LNv is important for sleep (McCarthy et al. 2011; Chung et al. 2009; Lear et al. 2009; Agosto et al. 2008; Parisky et al. 2008).
18.6.3 Molecular Mechanisms of Circadian Clock
The circadian oscillators which control activity and eclosion are supposed to consist of one main molecular feedback loop plus additional ones (Tomioka et al. 2012; Duvall and Taghert 2011; Hardin 2011). These are generated by interactions of several clock genes, the products of which activate or repress transcription, alter the stability of proteins or degrade them, and change subcellular localization. Transcription is activated by the transcription factors CLK, CYC, and PDP1#. Transcription is repressed by PER, TIM, and VRI. Protein stability and subcellular localization depends on the kinase DBT, CK2, SGG, and PP2a. SLIMB targets phosphorylated PER for degradation in the proteasome. Which role these components may play in each circadian oscillator cell of Drosophila is shown in Fig. 18.8.

Model of the circadian Drosophila clock based on a feedback loop involving transcriptional and translational events in the clock cells. The feedback loop has the following structure: CLK and CYC are transcription factors. They form heterodimers and bind to an E-box in the promoters of per and tim (and clock-controlled genes ccg) in the nucleus activating the transcription of per and tim. mRNA levels increase until they reach high levels early in the evening. The proteins PER and TIM reach maximal levels with a lag in late evening. Degradation of PER by DBT is counteracted by rapid binding of PER and TIM in the cytoplasm and accumulation in foci. After 6 h the complexes abruptly dissociate, PER and TIM move rapidly into the nucleus. DBT and SGG (not shown) in the cytoplasm interact with nuclear entrance or phosphorylate PER and destabilizes it. In the nucleus PER and TIM repress their own transcription by directly interacting with CLK-CYC transcriptional activators. TIM and PER are phosphorylated and degraded. This stops transcriptional repression at the end of the circadian cycle. Due to the lag between mRNAs and proteins, and due to a 6 h timer in the PER:TIM complex, this negative feedback results in a stable cycling in per and tim mRNA and protein levels. Entrainment of the clock by light (L in yellow circle) functions by affecting (blue flash) the blue-sensitive CRY. It stimulates CRY:TIM interaction, which triggers TIM degradation and prevents PER:TIM binding. TIM is phosphorylated, ubiquinated, and degraded in proteasomes. Other photoreceptors (rhodopsin, see text) and their interaction with the molecular gears of the clock are not shown (After Hardin (2005), Cyran et al. (2005). Insert by Dennis Pauls and ChristianWegener, Würzburg)
Results (Meyer et al. 2006; Cyran et al. 2005) from experiments in which the FRET signals of PER and TIM (tagged with two different fluorescent proteins) were measured in S2 cells (Schneider 2 cells, derived from a culture of late stage of Drosophila melanogaster embryos) show that PER and TIM bind rapidly in the cytoplasm and accumulate in foci. After 6 h the complexes abruptly dissociate, PER and TIM move independently and rapidly into the nucleus. This speaks in favor of a timer in the foci, perhaps similar to the circadian timing in Cyanophyceae (Sect. 18.2).
In this connection, it should also be mentioned that in many insects PER is not found in the nucleus and thus cycling of per mRNA might not always be necessary for PER cycling. Instead posttranscriptional mechanisms might be involved and the negative feedback of clock proteins on their own expression could be optional (Helfrich-Förster 2005a, b).
18.6.4 Photoreceptors of the Clock
Drosophila uses multiple photoreceptors for entraining its circadian system (for reasons see page 10). The rhythm of activity in adult flies can be entrained by (1) the compound eyes and (2) ocelli as external photoreceptors and by the (3) Hofbauer-Buchner eyelets behind the compound eyes (Veleri et al. 2007; Helfrich-Förster et al. 2002) and (4) LNv and DN neurons in the brain as internal photoreceptors (see Fig. 18.7 and reviews by Rieger et al. 2003; Helfrich-Förster et al. 1998). Whereas in (1), (2), and (3), rhodopsins serve as photopigments, in (4) this is CRY (Klarsfeld et al. 2004). Only when all photoreceptors are eliminated, fruit flies are unable to entrain their activity and sleep rhythm to LD cycles (Helfrich-Förster et al. 2001), suggesting that the multiple photoreceptors fulfill partially redundasnt roles and each single one is capable to reset the molecular feedback loop.
The following basic light effects must be explained:
-
1.
Phase shifting by light pulses
-
2.
Entrainment by LD cycles
-
3.
Attenuation of the rhythm by LL of high fluence rate
-
4.
Internal desynchronization (period lengthening/shortening in subpopulations) by LL of low fluence rate
-
5.
Adaptation of the activity pattern to different day lengths
According to the model (Fig. 18.8), all these effects are achieved through light-dependent degradation of TIM. Because TIM is not light sensitive by itself, the light signal must be transduced to TIM. As indicated in Fig. 18.7, this occurs via the blue-light-absorbing photopigment CRY. Photochemical changes in its flavin chromophore allow CRY to interact with TIM in the cytoplasm and the nucleus. This leads to TIM phosphorylation and its subsequent degradation in the proteasome (Naidoo et al. 1999) preventing the PER/TIM complex from participating in the negative feedback loop (Ceriani et al. 1999).
If a light pulse hits during the phases of the rising TIM concentration, TIM is reduced before it enters the nucleus and builds up again after the end of the pulse. The following peaks in TIM concentration are thus delayed (see Fig. 18.8). Because cytoplasmic PER is degraded if not protected by TIM, the PER peak is also delayed. If the light pulse hits at peaking TIM concentrations or afterward, nuclear TIM (bound in the PER/TIM complex) is degraded earlier, followed by PER degradation. As a consequence per and tim transcription starts earlier and the subsequent buildup of TIM and PER is also advanced. Thus, the following peaks in TIM and PER are advanced. This explains point 1.
The entrainment by LD cycles is the result of advancing and delaying phase shifts. These will keep the circadian oscillation in a certain phase relationship to the LD cycle. This explains point 2. LL of high fluence rate keeps the TIM level permanently extremely low (close to zero). As a consequence also PER cannot accumulate, per and tim mRNA remain at an intermediate level, and finally the clock genes and proteins stop to oscillate. This explains point 3.
LL of low fluence rate slows down the accumulation of TIM, but does not prevent it. As a consequence TIM and PER accumulation is permanently delayed, and this results in a period lengthening. This explains the first part of point 4.
The period shortening of some clock neurons under dim LL is more difficult to explain by the model, and the same is true for adaptation of the activity pattern to different day lengths since this includes a phase advance (period shortening) of the M oscillators (see page 37).
Thus, the model can explain the first three light effects mentioned as well as period lengthening (point 4). There is also experimental evidence: TIM degradation induced by light pulses can be measured in the LNs (see Fig. 18.7). It correlates well with the amount of phase shifts of the activity rhythm elicited by light pulses. Furthermore, the spectral response curves for TIM degradation and for phase shifts of the activity rhythm display a maximum between 400 and 450 nm, and this matches the absorption spectrum of CRY (Berndt et al. 2007), clearly indicating that these events are causally related.
Most interestingly, CRY levels are light controlled: CRY levels are maximal in the early morning and decrease over the course of the day reaching a minimum in the early night before they increase again during the night (Emery et al. 1998). This fact explains why the circadian clock is most sensitive to light pulses of low fluence rate in the early morning (about 3 h before lights on), when CRY level is maximal (Emery et al. 1998). Nevertheless, CRY is not the only factor that mediates phase shifts and entrains the clock to LD cycles:
-
(a)
Not all clock neurons contain CRY (Yoshii et al. 2008; Benito et al. 2008)
-
(b)
Entrainment of the molecular feedback loop upon light does also occur in mutants without functional CRY, at least in some clock neurons (Helfrich-Förster et al. 2001)
-
(c)
CRY-less flies can still entrain their sleep/wake rhythms to LD cycles and respond with phase shifts to light pulses, although with strongly reduced magnitude (Kistenpfennig et al. 2012)
-
(d)
Wild-type flies show a more pronounced phase-delay zone than advance zone in their PRC, although CRY is at its minimum during the early night when phase delays occur
-
(e)
The activity of wild-type flies can still be entrained by red light (600 nm), although CRY does not respond to light of wavelengths above 540 nm. The clock of eyeless flies lacks sensitivity to long wavelengths completely (Helfrich-Förster et al. 2002) and is much less sensitive to light of all wavelengths as compared to wild-type flies (1 by a magnitude of 1,000 (Hirsh et al. 2010) and not 10, as wrongly stated in Helfrich-Förster et al. 2002 due to a calculation error of the wild-type sensitivity).
This all emphasizes that CRY-independent pathways contribute to the light responses of the fruit fly’s clock.
Indeed, the still missing points of the basic light effects can only be explained by photoreception via the compound eyes and perhaps the H-B eyelets (point 4, period shortening during internal desynchronization under LL of low fluence rate; point 5, adaptation of the activity pattern to different day lengths). Only flies that lack these photoreceptor organs are unable to shorten period under LL and to adapt to different photoperiods (Rieger et al. 2006, 2003).
The most difficult point to explain is the period shortening of some clock neurons in response to LL. According to the model in Fig. 18.8, permanent TIM degradation can only slow down the feedback loop and finally stop it but hardly accelerate it. This implies that the input via the photoreceptor organs to the clock neurons does not lead to TIM degradation. This is in agreement with the fact that flies that retain the compound eyes and H-B eyelets but lack functional CRY do not become arrhythmic under LL, not even at high fluence rates (Stanewsky et al. 1998; Emery et al. 2000; Helfrich-Förster et al. 2001; Yoshii et al. 2004; Rieger et al. 2003). Moreover, the PER/TIM feedback loop runs with high amplitude in these flies under LL conditions without any sign for permanent TIM reduction (Rieger et al. 2006).
So far, it is completely unknown by which transduction cascade the light signal from the photoreceptor organs is transferred to the molecular clock and whether there are parallels to the mammalian system, where light input via the retinohypothalamic tract results in an increase of Ca2+ and cAMP in certain clock neurons finally leading to the activation of cAMP-responsive binding element (CREB). CREB binds to CREs in the promoters of per1 and per2 genes and activates their transcription. It is not yet established whether there are functional CRE sequences in the Drosophila per upstream region, but it was shown that mutations in the Drosophila CREB gene affect per expression (Belvin et al. 1999). Alternatively, per expression may be affected indirectly, for example, by the CREB-binding protein (CBP) that influences the transcriptional activity of the CLK/CYC heterodimer (Lim et al. 2007; Hung et al. 2007).
Although not yet proven, it is imaginable that activation of per transcription by light can provoke period shortening under LL conditions in some neurons as was observed by Rieger et al. (2006). Another possibility is that the period of some neurons is shortened via neuronal communication within the clock network.
The l-LNv neurons, which are responsible for arousal and sleep (see above), seem to play a crucial role in transferring the light information from the compound eyes to the network of clock neurons. Upon light the LNv increases neuronal activity (Sheeba et al. 2008b), releases the pigment-dispersing factor PDF, and increases arousal of the flies (Sheeba et al. 2008a; Shang et al. 2008).
But PDF acts also on the clock neurons themselves. Most clock neurons express the PDF receptor (Im and Taghert 2010), and they respond to PDF either by shortening or lengthening the period of their molecular clock (Yoshii et al. 2009). An increase of PDF in the dorsal brain does lead to internal desynchronization of the free-running activity rhythm into two components as does LL (Helfrich-Förster et al. 2000; Wülbeck et al. 2008). Furthermore, the same clock neurons seem to free run with a short and long periods, respectively, as observed under LL (Yoshii et al. 2009). This suggests that PDF is the factor that accelerates the speed of the M cells and decelerates the speed of the E cells.
Most interestingly, a recent study suggests that M and E clock cells express different adenylate cyclases, which may enable the two types to respond differentially to PDF (Duvall and Taghert 2012). In accordance with this, neither did pdf-null mutants show internal desynchronization upon light nor are they able to adapt M and E peaks to different photoperiods – very similar to eyeless mutants (Yoshii et al. 2009, 2012).
Several rhodopsins may be responsible for the observed light effects on the circadian clock: Rh1, Rh3, Rh4, and Rh5 which are expressed in the compound eyes, Rh6 which is expressed in the compound eyes and in the H-B eyelets, and Rh2 which is found in the ocelli (summarized in Szular et al. 2012). Among these Rh1 and Rh6 have been shown to be responsible for entraining fruit flies to red light (Hanai et al. 2008), whereas Rh1, Rh5, and Rh6 (plus CRY) are essential for entrainment to green and yellow light (Hanai and Ishida 2009).
In summary, the fly has at least two principle light-input pathways to the clock – one working via CRY on TIM degradation directly in the clock neurons and the other working via the photoreceptor organs and the neuropeptide PDF. The CRY pathway enables the fly clock to respond quickly and strongly to light. The photoreceptor input pathway seems to be more subtle but necessary for adapting the flies activity to different day lengths.
18.7 Light and Circadian Clocks in Mammals
Among vertebrates, the circadian clocks of mammals are the best studied. For experimental reasons, rodents are favored and among these mice (Ripperger et al. 2011) and rats, because they can be reared easily, are small and have a short generation time. However, most rodents used are night active, which has to be taken into account, if connections to the human circadian system are made. Day-active rodents are, e.g., the sand rat Psammomys obesus and the Nile grass rat Arvicanthis niloticus.
During the night diurnal and nocturnal species are sensitive to light at the same time. The molecular mechanisms of light resetting are also comparable. But animals in LL exposed to darkness would reset the SCN clock during their resting period, that is, at night in diurnal and during the day in nocturnal species. Arousal-independent cues (melatonin and GABA) shift the clock in day- and night-active animals at the same circadian time. Arousal-dependent zeitgeber (serotonin: its cerebral levels follow activity pattern) phase shift only during resting and have thus opposite effects in diurnal and nocturnal species (Challet 2007).
Several hands of the clock can be recorded such as locomotor activity, body temperature, melatonin secretion (see Sect. 18.7.6), or expression of clock genes or clock-driven genes. The locomotor activity is measured by running wheels or by infrared light beams (Jud et al. 2005). The records are used to construct actograms which allow to determine period and phase shifting of the rhythm.
Further advantages of using mice are that the genome was mapped and sequenced (see Müller and Grossniklaus 2010 for historical aspects), that many mutants are available, and that genetic and molecular biological methods are applicable. However, among the vertebrates mammals are special in several respects. Thus, peripheral clocks of mammals are not directly entrainable by light, and the pineal is not photoreceptive in contrast to most other vertebrates. In zebrafish, for instance, peripheral clocks are directly entrained by light, which resembles the situation in Drosophila, and the pineal is photoreceptive and contains a circadian oscillator as the central clock (see Sect. 18.7.6). These pineal cells are specialized photoreceptor cells resembling structurally and functionally retinal photoreceptors. They synthesize rhythmically the hormone melatonin with high levels at night and low levels during the day (Idda et al. 2012).
In the following we discuss the clock centers in the SCN and its network (Sect. 18.7.1), the clock mechanism driving these rhythms (Sect. 18.7.2), the circadian photoreceptors in the eye and the inputs to the SCN (Sect. 18.7.3), the circadian clocks in the eyes (Sect. 18.7.4), peripheral clocks (Sect. 18.7.5), and the function of the pineal organ (Sect. 18.7.6).
18.7.1 SCN and Its Network
The paired SCN is a center of circadian timing in mammals. It is situated in the anterior part of the hypothalamus at the ventral part of the third ventricle just above the optic chiasma (Fig. 18.10; Mohawk and Takahashi 2011; Welsh et al. 2010) and each SCN in a Rhesus monkey consists of about 10,000 neuron cells and additionally 15,000 glia cells (Roberts et al. 2012). Reorganization of neuron-astrocyte interactions and synaptic connectivity within the SCN is discussed by Jackson (2011), Marpegan et al. (2011), Ng et al. (2011), and Girardet et al. (2010). The SCN consists of a dorsomedial shell (dmSCN) and a ventrolateral core (vlSCN) with diverse afferents and efferents. Their neurons use different neuropeptides: in the shell VIP, GRP, and GABA, in the core AVP and PK2 (Mohawk and Takahashi 2011). For further details of structure and function of the SCN, see Tonsfeldt and Chappell (2012), Lowrey and Takahashi (2011), Dibner et al. (2010), Golombek and Rosenstein (2010), and Welsh et al. (2010); for the genetics of the SCN oscillators, its transcriptional loops, and how epigenetic mechanisms contribute to the control of circadian gene expression, see, e.g., Lowrey and Takahashi (2011), Colwell (2011), Kwon et al. (2011), Ripperger and Merrow (2011), and Bellet and Sassone-Corsi (2010).
The SCN is a circadian center and not just a place that transfers information of the LD cycle from the eye to an oscillator situated somewhere else. If this were the case, the various circadian rhythms would not disappear upon destruction of the SCN, but would not be synchronized anymore and would free run. However, the animals became arrhythmic.
There is further evidence for the SCN being a master oscillator: in organotypic slice cultures, in which the dorsal/ventral architecture is preserved (Silver and Schwartz 2005), metabolism, electrophysiological, and molecular events are still rhythmic. A particular strong evidence is that neural grafts of fetal SCN reestablish the circadian rhythms in SCN lesioned and thus arrhythmic recipients with the characteristic circadian properties of the donor.
To work properly as coordinated clocks, the oscillations must be coupled and form a network. Coupling of the neurons increases strength and precision of the rhythms, they resist better perturbations, and the range of entrainment is narrowed (Abraham et al. 2010). The overall period of the SCN is the average of the periods of the single cells. However, the intercellular coupling has to reach a certain strength, which is not present in dispersed SCN cell cultures.
The SCN oscillators must furthermore be synchronized with the 24 h environment. This occurs mainly by retinal inputs which generate action potentials. These travel along the retinohypothalamic tract (RHT) to the core of the SCN. There they activate the firing rate in the SCN neurons (see Sect. 18.7.3).
Since the core receives retinal inputs directly, the molecular oscillators respond immediately to the retinal inputs. The information from the core to the shell is transmitted in a so far unknown way and occurs delayed, so that the shell neurons react more slowly.
About 60–70 % of the SCN neurons show a circadian rhythm in action potentials which can last in single neurons for 4–6 h. The neurons switch between a hyperpolarized downstate during the night when they are silent and a depolarized upstate during the day when they are active. In this active state they respond to synaptic inputs that reduce their firing, but are not responsive to excitatory signals. During the night they are silent but respond to stimulation. How the change from day to night states occurs is unknown. The molecular oscillators in the SCN neurons influence the membranes and ion channels of the neuronal cells by a second messenger system which leads to circadian firing. It is highest during the day in diurnal as well as in nocturnal animals (Colwell 2011).
Under LL the rhythm of the locomotor activity of mammals can occasionally show a bimodal rhythm with a phase difference between the two components (Watanabe et al. 2007, 2006) or splitting with different periods (Butler et al. 2012; Indic et al. 2008; Helfrich-Förster 2004). Splitting is caused by the left and right halves of the SCN which oscillate in antiphase to each other (Mendoza et al. 2009; Tavakoli-Nezhad and Schwartz 2005). Arrhythmicity results, if the neurons within the SCN decouple from each other. The rhythmicity of individual cells remains, however, intact.
Internal desynchronization by non-24 h LD cycles (e.g., 11:11 h, called forced desynchrony protocol) can dissociate the molecular rhythms in the shell and core. The core stays entrained to the LD, but the shell is entrained for certain periods only. In between the rhythm free runs. As a result, two components of the locomotor activity, the body temperature and the slow-wave sleep, are displayed with different period lengths.
Splitting, internal desynchronization, and arrhythmicity are simulated by two groups of oscillators which are weakly coupled mutually but strongly coupled inside the group (Schroder et al. 2012). Light increases transcription of clock genes, which alters the circadian properties of individual cells. In DD weak coupling in the groups leads to oscillations with a single bout of activity per day. In LL, synchrony in a group occurs only under strong coupling. With increasing light intensity the rhythms of the two groups are in antiphase, which shows up as bimodal activity. At high light intensity arrhythmicity is found (Butler et al. 2012). Another model based on the Goodwin oscillator describes also splitting (Gu et al. 2011).
Many core neurons terminate on shell cells and their interplay leads to the circadian output to other hypothalamic regions. VIP modulates light-induced phase shifting and shifts the locomotor activity. If applied to the SCN in vitro, it shifts also the phase. Loss of VIP signaling leads to desynchrony among SCN neurons. Other neurotransmitters such as GRP and GABA phase shift the oscillations in SCN and the locomotor activity rhythm and synchronize asynchronous SCN neurons. Ca2+ signaling and cAMP activation, and perhaps other neuropeptides, are involved in synchronization by VIP.
Even in the intact SCN, the oscillations are not completely in synchrony, but their phase shows wavelike gradients at the various axes (shown by imaging, figure 3 in Mohawk et al. 2012), and the structure and synaptic connections of the SCN neurons and glia cells can change within a few hours. In order to find out how this occurs, various electrophysiological techniques were applied to individual SCN neurons and to SCN slices (Schaap et al. 2003). They allow monitoring of SCN neurons for longer periods. In organotypic slices from transgenic mice in which a luciferase reporter (Yamaguchi et al. 2003) or a fluorescent protein reporter (Quintero et al. 2003) drives a clock gene promoter, time-lapse imaging showed in horizontally cut slices two oscillating components, which might reflect the activity of morning and evening oscillators (de la Iglesia et al. 2004; Jagota et al. 2000). They had been inferred already earlier from behavioral studies (Pittendrigh and Daan 1976).
In photoperiodic reactions such evening and morning oscillators are supposed to measure day length (Jagota et al. 2000) (see Sect. 18.7.6). The photoperiod (day length) changes the pattern of the clock gene expression: long days broaden the clock gene expression of the SCN and lengthen the time of its neuronal activity in the dmSCN and along the rostral-caudal axis of the SCN (see also Sect. 18.4.3).
Outputs of the SCN: Intercellular communications in the mammalian clock system are not restricted to the clock cells in the SCN. Primary targets of the SCN outputs are predominantly located in the hypothalamus and the thalamus (see Fig. 18.10 and references in Li et al. (2012)). How does the circadian information of the SCN reach the other brain areas?
The signals of the SCN are spread as synchronized nerve impulses to central parasympathetic nuclei (e.g., the dorsal motor nucleus of the vagus, which innervates gastrointestinal and respiratory organs) and central sympathetic nuclei (e.g., intermediolateral cell column of the spinal cord). Sympathetic signals to the adrenal gland are converted to hormonal (glucocorticoid) signals. They are released into the bloodstream and bind to glucocorticoid receptors of peripheral organs, activate the mammalian Per1 gene in systemic cells, and reset clocks all over the body (Tonsfeldt and Chappell 2012; Okamura 2007).
Locomotor activity is driven by cyclic releasing factors acting on receptors in the hypothalamus at the wall of the third ventricle. Circadian signaling factors are PK2, AVP, and CLC. They influence also other behavioral and physiological rhythms (Li et al. 2012; Dibner et al. 2010; Klein et al. 1991) such as thermal regulation (Kräuchi et al. 2006; Ruby et al. 2002), sleep-wake cycle, functions of the circulatory and gastrointestinal system (Bass and Takahashi 2010; Green et al. 2008; Gachon et al. 2004), and endocrine events (Vollrath 2002). The synthesis and secretion of melatonin is also controlled by the SCN (see Sect. 18.7.6 and Simonneaux and Ribelayga 2003).
If the SCNs are destroyed, the circadian control of these functions disappears. The close connection between SCN and metabolism shows how the circadian system is integrated with physiology (Hardie et al. 2012; Froy 2011; Bellet and Sassone-Corsi 2010). AMP-activated protein kinase (AMPK) is an energy sensor of the cell. It is activated, if the energy status falls, promotes ATP production, and conserves ATP by switching off biosynthetic pathways. It furthermore regulates the energy balance of the body via the hypothalamus that promotes also metabolism and feeding behavior. Circadian rhythms and metabolism are closely linked via the activating (CLOCK-BMAL1) and repressive (REV-ERB-a-REV-ERB-b) transcriptional complexes, the coordinate actions of which generate rhythmic gene expression (Cho et al. 2012).
18.7.2 Mechanism of the Mammalian Clock
The molecular basis of the mammalian master clock in the SCN has been studied intensively using various methods, among them systems biology which is used to identify the circadian system and its components, to analyse and measure them, to control the system, and to put it together from its parts (Hogenesch and Ueda 2011; Ukai and Ueda 2010; Baggs and Hogenesch 2010). The clock mechanism presently known involves three basic helix-loop-helix transcription factors (Clock, Npas2, and Bmal1), two period genes (Per1 and Per2), two cryptochrome genes (Cry1 and Cry2), CKIε and δ, and two orphan nuclear hormone receptors (RevErbα and Rorα). Gene expression is regulated by transcriptional factors (Dbp, Tef, Hfl, and Nfl3; Bmal2, Bhlhb2, and Bhlhb3; one period-related gene Per3; RevErbβ, Rorβ, and Rorγ Ukai and Ueda 2010).
How the clock genes and clock-related genes interact with each other is at least partly clarified (see Fig. 18.9 and 18.10). CLOCK and BMAL1 dimerize and activate transcription of Per and Cry through E-box elements. The PER and CRY proteins accumulate in the cytosol, are phosphorylated, and then are translocated into the nucleus. There they inhibit the activity of Clock and BMal1. After the turnover of PER and CRY, a new cycle of activation by CLOCK and BMAL1 begins. This clock oscillator influences physiological and metabolic processes.

The circadian clock of mammals consists of a main feedback loop in which the products (PERs and CRYs, thick green arrows) of the Per1, Per2, and Per3 genes and of the Cry1 and Cry2 genes inhibit (thick red – I) the expression of their genes (Per1–3 and Cry1–2). A kinase (CK1ε) is responsible for the destruction of the PERs and CRYs (blue ovals with broken borders). Further feedback loops (REVERBa, RORa, and RORb) interact with Bmal1 which via BMAL1/CLK affects the Per and Cry genes. Light (L and yellow and blue flash) affects the Per and Cry genes (After Kim et al. (2012)). Insets: The female author of this chapter and her coauthors during a circadian experiment in Spitsbergen (see link: http://nbn-resolving.de/urn:nbn:de:bsz:21-opus-53405)

Circadian centers of mammals and their pathways: the environmental L/D is received by the retina (bottom left and insert A) in ipRGC (dark blobs in A) and in the rods (PRs, short structures) and cones (long structures) and reach via action potentials (AP, insert C) in the RHT the SCN (insert B). The APs (insert C) open Ca2+ channels (Ca) which leads to discharge of neurotransmitters VIP, GRP, and GABA from the presynaptic neurons into the synapse and clock gene activation in the postsynaptic neurons via receptors (only AC shown), cAMP, PKA, and CREB. The signals are received by clock neurons cells (circles with ~, insert B) in the core of the SCN, which are entrained by them and by mutual interactions (⇌). They in turn synchronize clock cells in the shell of the SCN which interact with each other (⇌) and with other cells (circles with -). They communicate with target tissue/organs via neurotransmitters like AVP, PK2, VIP. Signals from the SCN reach the pineal (PIN) via sympathetic innervation with synapses in the PVN, IML, and SCG. Insert D shows part of a pinealocyte where the signals trigger via norepinephrine (NE) the release of melatonin (Mel) into the blood vessels (bl.v.) of the brain. The signal cascade involves Ca2+ and PKC which increase cAMP. PKA phosphorylates CREB in the nucleus, activating genes involved in melatonin synthesis. Melatonin reaches via the blood different targets, among them SCN cells and cells of the pars tuberalis (PT; main figure), both of which contain numerous melatonin receptors. The PT controls photoperiodic events. Another direct target of the ipRGC in the retina is the olivary pretectal nucleus (OPN) linking the pupillary light reflex via EW, CG, and the iris muscles (After Berson (2003), Hastings and Herzog (2004), Meijer and Schwartz (2003), Lincoln (2006), Pévet et al. (2006), Welsh et al. (2010), Dibner et al. (2010), and Schmidt et al. (2011))
The rhythmic expression of Bmal1 mRNA is also clock regulated. The Bmal1 promoter contains RRE instead of an E-box, and its activities are controlled by the rhythmically expressed transcriptional repressor REVERBα and the activator RORα. CRY inhibits by feedback the CLOCK/BMAL1 complex and is responsible for the functioning of the mammalian clock. Critical for the function is also the E-box-mediated transcriptional/posttranscriptional loop.
The circadian clock of mammals possesses also posttranscriptional regulation. Using newer technologies, several protein kinase, adenylate cyclase, and proteasome inhibitors were found which lengthen the period of the clock. The decisive step seems to be the CKIε/δ-dependent phosphorylation of PER2.
Circadian clocks must be protected against fluctuations in the environment such as temperature and food. They are therefore temperature compensated. Theoretical studies of Isojima et al. (2009) show that the effect of CKIε and CKIδ activity on PER2 determines period and is temperature insensitive. Circadian oscillators of mammals are also resistant against fluctuations of the transcription rate (Dibner et al. 2009).
It was mentioned already on page 9 that a special light pulse at a critical phase of the circadian oscillator might lead to arrhythmicity. This feature has been used by Ukai and Ueda (2010) and Pulivarthy et al. (2007) to find out whether the arrhythmicity is due to a stop of the oscillators in a singular state or whether the oscillators continue to oscillate but desynchronized to each other. In mammalian cell cultures made photo responsive by coupling αρ protein to melanopsin, it was shown that desynchronization of the cellular clocks and not arrhythmicity was responsible (Ukai and Ueda 2010; Pulivarthy et al. 2007). In light-responsive immortalized fibroblasts of mice, such a critical light pulse reduces the amplitude of the rhythm by 40 % but desynchronizes simultaneously the cells (Pulivarthy et al. 2007). In rats it was shown in vivo that desynchronization is responsible for the low amplitude of the locomotor activity after such a critical light pulse (Ukai et al. 2007).
Using a tunable oscillator (see Tigges et al. 2009) in mammalian cells, it could be clarified how the phases of expression of oscillating genes are determined by the three CCEs (E/E-box, D-box, and RRE). The transcriptional activator DBP activates gene expression via the D-box; the E4BP4 represses gene expression. Dbp is regulated by the E-box, the morning control element. E4bp4, however, is regulated by the RRE, the nighttime control element. RRE activators are expressed during the day phase through the D-box. The RRE repressors are influenced by a morning element (E-box). A daytime activator and a morning repressor determine the nighttime transcription through the RRE (Ukai-Tadenuma et al. 2008). Rhythms did not have a strong amplitude, if only the morning activator or only the night repressor was expressed. Morning activation and night repression are responsible for the control during the day. Various combinations of transcriptional regulators with CCEs for the three basic circadian phases (morning, day, and night) result in other phases and show that transcriptional regulation of upstream transcription factors can determine the phase of the downstream output. The period length of the clock is largely determined by posttranscriptional circuits.
18.7.3 Circadian Photoreceptors in the Eye
As mentioned, the SCN of mammals is only synchronized by light perceived via the eyes: enucleated animals free run in LD cycles (Meijer et al. 1996).
Whereas the rods and cones of the outer retina are responsible for normal vision and communicate with the brain via the optic nerve, a small subset of intrinsically photosensitive retinal ganglion cells (ipRGCs; see Fig. 18.9) in the inner nuclear layer of the retina controls the circadian rhythm in the SCN. These cells regulate also masking effects of the light, the immediate melatonin suppression, and the pupil reflexes (Hughes et al. 2012; Lucas et al. 2012). They occur in small numbers only (in humans 0.2–0.8 % of all ganglion cells in the retina) but spread widely as a network across the entire retina (Moore et al. 1995) and are strongly arborized (Hannibal et al. 2002). They detect the average illuminance of the day and integrate it over long intervals (Do and Yau 2010; Pickard and Sollars 2010; Husain 2005; Warren et al. 2003). In addition they receive input from the rod/cone circuitry. Plachetzki et al. (2005) provide a historical overview.
There are five types of ipRGCs, M1–M5, which differ in connection and function. The Brn3b-negative M1 cells innervate the SCN and entrain the clock cells, and the Brn3b-positive M1 cells innervate the shell of the OPN. From there the pupillary light response is controlled. The other M cells innervate the core of the OPN, the SC, the dLGN, and other regions, but their specific functions are not yet known (Hughes et al. 2012).
The LD cycles of the environment are sensed by the Brn3b-negative M1 ipRGCs and transferred as electrical signals via their monosynaptic axons in the RHT to the SCN. Melanopsin serves as the photopigment. G protein is activated, thereafter PLC and finally an influx of Ca2+ leads to action potentials. As a result, Glu and PACAP are released postsynaptically at the SCN neurons. Ca2+ influx activates protein kinases and CREB is phosphorylated (Golombek and Rosenstein 2010). It binds to CREs in promoters of many clock genes such as the Per1 and Per2 genes and activates transcription. Depending on the time at night at which light occurred, the increased PER advances or delays the locomotor activity rhythm.
The transduction cascade of the melanopsin-expressing ipRGCs differs fundamentally from the one of the rod and cone photoreceptors, since a different G protein is involved and melanopsin activation results in membrane depolarization rather than hyperpolarization as in rods and cones. It thus resembles more closely the phototransduction cascade of invertebrates (Palczewski 2012; Bailes and Lucas 2010). Whereas mammals have just one melanopsin (Opn4m), fish, birds, and amphibians possess two, Opn4m and Opn4x (Bellingham et al. 2006).
All inputs of the accessory light perception use the rod-cone and the melanopsin system (Hattar et al. 2003). However, the quality of the information differs. Cones signal rapid changes in light intensity, and rods and melanopsin gradual modulations during the day at dim and brighter intensities. Mice possess a small population of short-wavelength-sensitive S-cones which could explain the UV sensitivity (Provencio and Foster 1995). The SCN is thus able to encode environmental light over a wide range of intensities and temporal frequencies (Lucas et al. 2012).
After the light has been absorbed in the retina and the signals have reached the SCN, immediate early genes for transcription regulator proteins and furthermore nitrogen monoxide come into action. The latter is required for light-induced phase shifts of behavioral rhythms (reviewed by Rea 1998).
Under DD, circadian rhythms are maintained by the clock cells of the SCN. But the eyes, which also contain autonomous circadian clocks (see Sect. 18.7.4), can also modulate circadian rhythms in the SCN even in the absence of light. Removal of the eyes abolishes circadian rhythms in some cells of the SCN; they were apparently driven by inputs from the eyes. However, removal of the eyes can also amplify a normally dampened circadian rhythm in other cells of the SCN (Beaule and Amir 2003).
18.7.4 Retinal Clocks in the Eye
After presenting the circadian photoreceptors in the retina (Sect. 18.7.3), we have to point out that the retina of the eye contains circadian clocks itself. The retina, its cells and tissues, and their interactions are well studied. It consists of several tissue layers with five classes of neurons (photoreceptors, horizontal cells, bipolar cells, amacrine cells, and ganglion cells) and glia (Guido et al. 2010; Tosini et al. 2008). There are three nuclear layers (outer and inner nuclear layer, ganglion cell layer; see inset A in Fig. 18.9). Light has to pass through these layers and their neurons before reaching the photoreceptors.
The outer segments of rods and cones absorb light with various opsin photopigments in stacked discs. The disks are shed at the tip of the segments, phagocytosed and digested in the retinal pigment epithelium (Bobu and Hicks 2009; Strauss 2005; Grace et al. 1999; Young 1976). At the base the segments are renewed by forming new disks. This internal renewal is important for the functional integrity of rods and cones, because they cannot be replaced by new cells. The shedding of the outer disks occurs in the rods in the morning and in the cones in the evening, that is, at times, when they are not used anymore.
Several photopigments with diverse functions are found in the various cell layers (Peirson et al. 2009). They synchronize the circadian rhythms (Iuvone et al. 2005; Green and Besharse 2004; Devlin 2002).
Circadian clocks in the mammalian eye were predicted by Reme et al. (1991). They have later been found in the retinas of all classes of vertebrates. The retina contains besides the normal visual system a circadian system with receptors for synchronizing light and with many mutually coupled clocks (Ruan et al. 2006). Different aspects of retinal physiology such as sensitivity to light (differences of more than six orders of magnitude), neurohormone synthesis, rod disk shedding, intracellular signaling, and gene expression are controlled by these clocks (Green and Besharse 2004; Tosini and Fukuhara 2002; Tosini and Menaker 1996). The retinal clock cells control also the circadian melatonin expression in the eye and are partly responsible difference in the sensitivity to light.
There are indications that in the retina two or more oscillator types control the local physiology. Melatonin and dopamine in the ganglion cells of the inner retina and a part of the inner nuclear layer and/or the photoreceptors are regulated in antiphase (Thompson et al. 2003; Hattar et al. 2003).
This close connection between circadian clocks and photoreceptors is known also from invertebrates and Neurospora.
The molecular components and genetic basis of the retinal and other peripheral clocks of the body are the same as the one in the SCN cells (Yamazaki et al. 2000), and they possess the same properties such as shorter periods in corresponding mutants. These shorter periods show up also in the output of the eye clocks (Grace et al. 1996). There are, however, variations in molecular detail (Steenhard and Besharse 2000).
The retinal clock cells control the local physiology and are responsible also for a circadian output of melatonin in the eye. This changes the sensitivity of the retina to light, covering a range of more than six orders of magnitude.
Similarities and differences in the molecular mechanisms of the retinal versus the SCN oscillators are discussed by Green and Besharse (2004). They also examined the interactions between the retinal and the SCN clocks. Light and dopamine phase shift the retinal clock (Steenhard and Besharse 2000). The circadian release of melatonin in the eye is responsible for the rhythmic adaptation of phototransduction, for the recycling of biochemical components in the retina, and for other aspects of the retinal physiology. Other rhythmic events in the eye such as visual resolution (Tassi et al. 2000), ERG, intraocular pressure (Nickla et al. 1998), cats: Sole et al. (2007), choroid thickening, and eye growth might be or are also under circadian control.
18.7.5 Peripheral Clocks and Their Entrainment
In the brain, clock genes are not only expressed in the SCN but also in other parts of the central nervous system (e.g., cerebellum, hippocampus, arcuate nuclei, paraventricular hypothalamic nuclei, piriform and cerebral cortices, olfactory bulbs, amygdala, retina Guilding and Piggins 2007). Furthermore, rhythms of clock gene and/or protein expression have been observed in peripheral tissues and organs such as liver, pancreas, fat tissue, gut, lung, and heart (references, also for the following, in Pévet and Challet 2011); Bass and Takahashi (2010). These rhythms persist in culture, that is, without the influence of the SCN cells. However, they dampen after a few cycles. Imaging has shown that individual cells continue to express rhythms, but are not synchronized anymore. In contrast to the SCN, they do not react to external time cues such as the LD cycle of the environment and cannot communicate with and entrain each other. Peripheral oscillators can be entrained by temperature cycles as exhibited in core body temperature. Thus, rhythmic gene expression can be driven by both local intracellular clocks and by extracellular systemic cues.
Metabolism is linked with the circadian system not only via the SCN but also via peripheral tissues, endocrine, and local signals. Feeding time and certain drug treatments can generate behavioral rhythms in the absence of the SCN (Honma and Honma 2009). The liver clock is reset by feeding, and the clock genes and protein expression shift their phase accordingly (references in Mohawk et al. 2012).
18.7.6 Pineal Organ, Melatonin, and Photoperiodism
The pineal resembles a tiny pine cone and is an endocrine gland in the center of the vertebrate brain between the hemispheres (see Fig. 18.9). It is a part of the “photoneuroendocrine system” (retina, suprachiasmatic nucleus, pineal) and consists mainly of pinealocytes, which produce and secrete melatonin. In lower vertebrates, they contain functional circadian oscillators, but in mammals they do not (Falcón et al. 2009). Furthermore, mammals lack extraretinal circadian photoreceptors in the pineal (Bertolucci and Foà 2004). Instead, light is perceived in photoreceptors of the rods, cones, and melanopsin-expressing retinal ganglion cells (for the exceptional circadian system of mammals and its evolution, see Menaker et al. 1997; Heesy and Hall 2010; Davies et al. 2012). The light signals are converted to electric signals in this photoneuroendocrine system (Do and Yau 2010). The electric signals are propagated via the RHT neurons to the SCN. From the SCN the light signaling is conveyed to the pineal via the paraventricular nuclei (PVN) of the hypothalamus, the sympathetic preganglionic neurons of the intermediolateral cell column of the spinal cord (IML), and noradrenergic sympathetic neurons to the superior cervical ganglion (SCG) (see Fig. 18.9 and Pévet and Challet 2011 for the neurotransmitters involved). The SCN output stimulating the PVN derives from two populations of SCN neurons, one active during daytime and the other during nighttime.
The pinealocytes in the pineal synthesize melatonin (=N-acetyl-5-methoxytryptamine Reiter et al. 2010) during the night upon release of noradrenaline from sympathetic pineal nerve terminals (for the neurotransmitters used in the pineal, see Stehle et al. 2011). Melatonin is immediately released into the blood circulation but also into the cerebrospinal fluid and rapidly degraded in the liver. Therefore, its plasma concentration precisely reflects its synthesis.
Central and local mechanisms regulate amplitude and rhythmic timing of melatonin synthesis from serotonin. Although the pinealocytes possess – like most cell types – the basic clock machinery, they are not able to produce a melatonin rhythm. Instead the rhythm is controlled by the SCN and without its sympathetic input there is no rhythm (Borjigin et al. 2012).
Melatonin transfers the temporal cues received from the SCN to the structures within the brain or the periphery that express melatonin receptors (Pévet and Challet 2011) (but note that circadian information can be sent from the SCN also via nervous or other hormonal signals). In some of them such as the pars tuberalis of the hypophysis, the melatonin signals drive rhythms; in other targets they synchronize peripheral oscillators, such as in the fetal adrenal gland. Some of these sites may not be involved in circadian control, but instead in photoperiodic responses (see page 56 and Chap. 19).
Besides melatonin, the pineal secretes also N-acetylserotonin (NAS) rhythmically. At night the levels are even higher than those of melatonin. NAS as an antioxidant is even more effective than melatonin. It may thus not only serve as the precursor of melatonin (Jang et al. 2010).
The rhythm of pineal melatonin is a very reliable marker of the circadian clock. But the phase of melatonin secretion varies widely between individuals and strains, as shown in rats. This wide variation of melatonin onset in an animal model is paralleled by a wide range of circadian chronotypes in humans.
Light affects melatonin synthesis not only by its resetting effect on the circadian system but has also an immediate effect (“masking”) if applied at night. One lux is already sufficient in golden hamster to inhibit melatonin synthesis (Brainard et al. 1983). For humans, a fluence response curve for immediate inhibition by blue light, which is most effective, was determined by Brainard et al. (2008).
Melatonin is produced not only in the pineal but also in the Harderian gland, the gastrointestinal tract, and the retina but is there only of local importance (see Sect. 18.7.4 and Hardeland et al. 2011). It is found not only in animals but also in bacteria, unicellular eukaryotes, and plants serving various tasks (Dibner et al. 2010).
Melatonin acts at the central level as well as at the periphery: it does not only affect the gonads and other centrally controlled events but also other tissues of the body and the brain (Pévet et al. 2006). Being a small molecule, melatonin can pass the placenta and convey circadian and seasonal information to the fetus (Stehle et al. 2011).
Melatonin modulates sleep propensity (Cajochen et al. 2010; Pandi-Perumal et al. 2006; Turek 2005; Gillette and Abbott 2005), vascular tone, immune function, controls seasonal reproduction (Sellix and Menaker 2011; Reiter et al. 2010; Revel et al. 2009), seasonal thermoregulation including torpor and hibernation (Saarela and Reiter 1994; Heldmaier and Steinlechner 1981; Chap. 12 in Heldmaier and Werner 2004), metabolism, energy balance (body weight regulation!), and immune responses (references in Barrenetxe et al. 2004).
Phase shifting of the SCN clockwork is the central and best characterized effect of melatonin (Shimomura et al. 2010). Treatment with exogenous melatonin can synchronize the SCN (Pévet and Challet 2011). Orally or injected melatonin pulses advance or delay the circadian rhythm depending on the phase of application. The phase response curve to melatonin pulses is similar to that of light pulses but displaced by 180° (Lewy et al. 1996). Therefore, melatonin can be used in a similar way as light pulses – if properly phased, to shift the circadian system. Circadian phase disorders can be treated in this way (Lewy and Sack 1997).
Melatonin shows connections to human diseases as discussed in Sects. 18.8.3, 18.8.4, and 18.8.5 and by Hardeland et al. (2012) and Pandi-Perumal et al. (2013). It is used to cure sleep disturbances and insomnia (for instance, in elderly people, Fiorentino and Martin 2010), depression, jet lag, and shift-work-related sleep cycle disorders (see Sect. 18.8). The antioxidant properties of melatonin (Reiter et al. 1999; Pandi-Perumal et al. 2012) might protect the skin against UV (Reiter et al. 2004).
In seasonally breeding mammals the melatonin production in the pineal plays a decisive role (reviews Ikegami and Yoshimura 2012; Hut and Beersma 2011; Walton et al. 2011; Revel et al. 2009; Morgan and Hazlerigg 2008). The duration and magnitude of the melatonin secretion depends on the length of the night, which changes during the year especially in higher latitudes. In this way seasonal changes are sensed by photoperiodically responsive mammals and control reproduction (stimulating or inhibiting, depending on the species (Stehle et al. (2001), Hoffmann (1981)) and pelage color, among others. The length of the night is coded by the duration of the nocturnal peak of melatonin (Revel et al. 2009; Morgan and Hazlerigg 2008). Clock genes in calendar cells of the pars tuberalis of the hypophysis, where melatonin receptors are strongly expressed, regulate prolactin release (Johnston et al. 2006; Lincoln et al. 2003). These cells contain circadian clocks, and the phase relationship between the expression of the clock genes Cry and Per1 is set by melatonin at dusk, respectively, dawn.
Whereas the photoperiodic timing in the SCN for the melatonin secretion in the pineal uses external coincidence, the timing in the pars tuberalis seems to follow an internal coincidence (see Sect. 18.4.3). As a consequence the transcription of downstream genes (prolactin releasing factor?) leads to either a long-day cell state or a short-day cell state (Lincoln et al. 2002).
Pinealectomy abolishes the photoperiodic response.
18.8 Light and the Human Circadian System
The effect of light on the circadian system of mammals has been discussed already in Sect. 18.7. A few peculiarities of the human circadian system and its responses to light are mentioned in the following.
The circadian system governs not only the sleep-wake cycle, body temperature, alertness, and efficiency but also many other metabolic, physiological, and behavioral events such as enzymatic activities in organs, hormonal secretion, and so on. There is a long list of circadian clock-driven events in man (Minors and Waterhouse 1981).
The sleep/wake cycle (Dijk and von Schantz 2005), body temperature, and urine amount and its composition can be monitored easily and have therefore often been used as hands of the circadian system. Melatonin concentration in the blood is a particularly useful measure because it is not much disturbed by activities, in contrast to the body temperature rhythm. Light has, however, an immediate suppressing effect on melatonin concentration (Pévet and Challet 2011; Reiter et al. 2010).
The circadian system shows up clearly under isolation from external time cues. In a cave or in an isolation facility, a person who has no information of the outside time will sleep and wake according to its internal circadian clock. By continuously measuring the body temperature, the sleep time, and the locomotor activity, the period length of these parameters can be determined under the light conditions given (see, e.g., Johnsson et al. 1979).
However, the design of the recording conditions has to be taken into account. Wever (1979) and others (e.g., Weitzman et al. 1981; Siffre 1975; Mills 1964) determined an average “free run” of about 25 h, but this estimate was too high, as discussed by Czeisler and Gooley (2007): the subjects were able to self-select their LD cycle. They exposed themselves to light during most of the delay portion of the phase response curve during wakefulness and to darkness during most of the phase-advance portion. As a result of this and due to the light intensities used, the free-running periods were overestimated (Khalsa et al. 2003; Honma et al. 2003; Klerman et al. 1996). Using other protocols such as “forced desynchrony,” which had been used already in 1938 by Kleitman (1963), period could be measured without the influence of the self-selected LD cycles (Duffy and Wright 2005; Czeisler et al. 1999) and turned out to be much closer to 24 h, namely, 24.2 h as an average. To entrain the circadian clock of a person with such a period to the 24 h day, the rhythm has to be advanced daily by about 0.2 h (12 min). The period of about 25 % of tested persons is less than 24.0 h, requiring a daily delay, whereas in the remaining individuals period is longer, thus requiring a daily advance of the rhythm. The interindividual variation in period length in humans is much smaller than assumed before (0.55 % instead of 30 %, references in Czeisler and Gooley 2007) and corresponds now more to the values of other mammals such as hamster and mouse. Entrainment thus requires daily shifts of less than 1 h.
In the forced desynchrony protocols, the LD cycles used were outside of the range of entrainment (e.g., an LD cycle of 28 h). If non-24 h cycles inside the range of entrainment are used, the observed period length shows a so-called aftereffect, which depends on the imposed cycle length. Subjects after a 24.65 h cycle displayed a longer period than subjects after entrainment by a 23.5 h cycle and demonstrate the plasticity of the human circadian system (Scheer et al. 2007). Blind individuals exhibit a free-run period closer to 24.5 h, and the shorter period in sighted subjects might represent an aftereffect of the entrained life to 24 h.
18.8.1 Light Synchronizes the Human Circadian System
Which time cues synchronize the human circadian system? The free-running rhythm of humans can be synchronized to the 24 h day by knowing the time of day and by external time cues such as light, temperature, noise, or social contact (for entrainment by non-photic signals, see Mistlberger and Skene 2005). Light plays a much more important role in humans than claimed by, e.g., Wever (1979) in earlier studies, and shorter exposures (Laakso et al. 1993) and lower intensities than assumed before are able to entrain the human circadian system (Wright et al. 2001).
In man, as in other mammals, the eyes seem to be the only places harboring photoreceptors which are able to synchronize or phase shift the circadian rhythms in the SCN, the master clock in vertebrates. Findings of Campbell and Murphy (1998) that extraretinal photoreception can phase shift the circadian rhythm of body temperature and melatonin concentration by illuminating the backside of the knees could not be verified in later experiments; see Rüger et al. (2003) with further references. Recently, Timonen et al. (2012) claimed to have influenced the circadian clock by light administered via the ear. This needs to be confirmed. That the eyes are the only sites for the photoreception is supported by the following:
All humans with bilateral enucleation and 20 % of the remaining blind people exhibit free-running circadian rhythms (“blind free runners,” Emens et al. 2005). In the rest either the blindness affects only normal vision of images, but not the circadian vision based on the ipRGCs, or other time cues are used for synchronization (Mistlberger and Skene 2005). Occasionally, free run is observed even in people with intact vision living in a normal environment (Giedke et al. 1983; Miles et al. 1977). It is not known why light (and other time cues) are ineffective in these people.
As in other mammals, single light pulses are able to phase shift circadian rhythms in humans. The phase response curve is of the strong or weak type (see page 10), depending on the strength and length of the light exposure (Khalsa et al. 2003; Minors et al. 1991) (questioned, however, by Duffy and Wright 2005 with further references). The human circadian system is responsive to light throughout the daytime (and of course much more during the nighttime, but under normal conditions the daytime light exposure synchronizes the human circadian system) without a “dead zone” (see page 10 and Jewett et al. 1997). A dose-response curve for phase shifting the melatonin rhythm by white light exists. Exposure to a light intensity of 2,000 lx for 1–3 h increased the magnitude of light-induced delays, whereas shorter exposures with higher intensities (4,000 and 8,000 lx) do not increase the shift further (Dewan et al. 2011). Light exposures of as low as 1.5 lx are sufficient to keep the circadian rhythm entrained (Czeisler and Gooley 2007). Phase shifting the rhythm by intermittent light is more effective than is a continuously applied light pulse.
Besides phase the amplitude of the rhythm is an important parameter. A first light pulse has been claimed to reduce the amplitude of the circadian rhythm in man, which would render the system more sensitive toward the phase-shifting effect of a second light pulse (Czeisler et al. 1989). A light pulse of a critical strength applied at a critical phase point (subjective midnight) induces arrhythmicity by pushing the clock into a singular point (references in Czeisler and Gooley (2007); see also page 9).
In order to find out which wavelengths are effective in shifting the phase of the circadian rhythm in man, action spectra (see Chap. 8) were determined by using the suppression of plasma melatonin. The results of those experiments are compiled and discussed by Brainard and Hanifin (2005). Light in the short-wavelength range (459–484 nm) is most effective. This differs from the spectral sensitivity of the visual system and points to special circadian photoreceptors as discussed in Sect. 18.7.3. The circadian rhythms of some people completely blind for vision are still entrained, and this applies also for suppressing melatonin by light. Responsible are the retinal ganglion cells which project directly to the SCN. Rods and cones are apparently dispensable but serve additionally for entrainment. Longer wavelengths are effective in resetting especially at low light intensities. The issue of photoreception and circadian entrainment has been discussed in Sect. 18.7.4 (for humans see also Kronauer et al. 1999). A detailed action spectrum which could help to identify the photoreceptors is still missing. Spectral sensitivity of the retinal ganglion cells to light seems to change during the night (Figueiro et al. 2005). The background light and the history of previous illumination affect the resetting properties of light, but this issue needs more studies.
18.8.2 Significance of Light and the Circadian Clock in Shift Work and Jet Lag
The circadian rhythm of modern man is often delayed with respect to the natural LD cycle. He uses electric light and can therefore stay up during the winter time much longer than natural daylight would otherwise permit (Cardinali 1998). This independence or even insulation from the natural light easily leads to permanent sleep deprivation. In addition, modern society expects full range services throughout the 24 h. Traffic, economy, health service, and security have to rely on shift work or night work by a considerable part of the workers (about 20 % in the industrialized nations, half of them on night or rotating shifts).
Normally the circadian clock is in synchrony with the daily cycling of external 24 h cycles, and the body can adapt to the cyclic demands. Night work and shift work clashes with our circadian clock and disrupts this synchrony. The internal coherence among oscillations is lost and problems may arise. Sleep disturbances and effects on health (Reed 2011; Erren et al. 2010) and on safety of workers and drivers (Philip and Akerstedt 2006; Barger et al. 2005) are due to shift work (Kecklund et al. 2012; Saksvik et al. 2011; Arendt 2010; Folkard 2008) and jet lag (Arendt 2009; Auger and Morgenthaler 2009; Coste and Lagarde 2009).
About 10 % of shift workers suffer from shift work disorder. The resulting internal desynchrony brings behavioral, hormonal, and metabolic rhythms out of phase and increases the risk of gastrointestinal problems, cancer, depression, heart disease, sleep disturbances, and accidents; it furthermore affects reproductive cycles, menstruation, and pregnancy (Mahoney 2010; Su et al. 2008) and decreases productivity (Roth 2012).
The synchronizing effect of light on the circadian system of man is one of the problems of shift work (Monk 2000). For instance, the high fluence rate of outdoor light in the morning after a night shift prevents the phase shift of the circadian system needed for optimal adjustment of the night workers clock (Horowitz et al. 2001). Wearing dark goggles is advisable in this case (Eastman et al. 1994). On the other hand, light can be used also for adjusting the clock to the shift work schedule, if properly applied (Turek 2005; Crowley et al. 2003). Models are used successfully for constructing LD cycles which phase shift the rhythm in such a way that they align better with shift work and day sleep schedules (Jewett et al. 1999b; Martin and Eastman 1998). More empirical data from shift work effects on the circadian rhythms are, however, needed for detailed simulations of this kind (Åkerstedt 1998). Other counteractions consist of light exposures at certain times of the circadian cycle (Boivin and James 2005) and of using chronobiotics such as melatonin (see Sect. 18.7.6 and Arendt (2005)). In using combinations of light and melatonin, it should be taken into account that the phase-shifting effect of light pulses and melatonin pulses is 180° out of phase (Skene 2003).
Whether the internal desynchrony occurs at a peripheral or at a central level has been studied by Salgado-Delgado et al. (2008) in a rat model. Working and feeding during the sleeping period uncouples metabolic functions from the biological clock which remains fixed to the LD cycle. The data suggest that in night workers the combination of work and eating during working hours may cause internal desynchronization. In this connection the quality of sleep and the cognitive performance of shift workers and workers with day shift only were studied in offshore fleet workers (Hansen et al. 2010). Morning types (for testing the chronobiological phase type, see link [http://nbn-resolving.de/urn:nbn:de:bsz:21-opus-37910] ||Chronobiological phase type, page 97) are under higher health risk even if treated with bright light (Griefahn and Robens 2010).
This internal desynchrony occurs already at the level of the first output from the SCN, namely, relaying nuclei of the hypothalamus such as the arcuate and dorsomedial nucleus, both associated with metabolism and regulation of the sleep/wake cycle. They transmit temporal signals to other brain areas and to the periphery. The SCN itself and the paraventricular nucleus stay, however, locked to the LD cycle (Salgado-Delgado et al. 2010). Desynchronization between clock and the LD cycle is thus unfavorable or even increases mortality (Park et al. 2012).
Jet lag is another problem. The circadian clock cannot rapidly adapt to a new LD cycle and this leads to desynchrony. Overviews and practical considerations are provided by Kolla and Auger (2011), Arendt (2009), Auger and Morgenthaler (2009), Coste and Lagarde (2009), and Revell and Eastman (2005). Symptoms can be reduced by proper avoidance or exposure to light (Zee and Goldstein 2010). Based on the human phase response curve to light pulses, one should avoid morning light and expose oneself to evening light in flying westward but expose oneself to morning light and avoid evening light in flying eastward (however, flight time and duration influence also adaptation to the new time zone). Chronobiotics (melatonin) and behavioral therapies are alternatives (Srinivasan et al. 2008; Touitou and Bogdan 2007). A discrepancy between biological and social timing leads to chronic sleep shortage and jet lag symptoms (social jet lag). It contributes to weight-related pathologies such as obesity (Roenneberg et al. 2012).
18.8.3 Light, Sleep, and Sleep Disorders
The sleep/wake cycle and its influence on performance is an interplay of the external LD and social cycles and of internal oscillators, namely, a circadian clock and a homeostatic oscillator. Sleep homeostasis implies that sleep deficit increases the intensity and duration of sleep and excessive sleep reduces sleep propensity. The circadian system determines sleep propensity, timing of sleep, and sleep structure and consolidates sleep and wakefulness, but the homeostatic oscillator contributes also to sleep timing and duration, to REM and slow-wave sleep, and furthermore to performance parameters such as attention and memory (Dijk and von Schantz 2005). The homeostatic oscillator interacts with the clock or its outputs according to the circadian principle the longer we are awake, the shorter we sleep and according to the homeostatic regulation of sleep the longer we are awake, the deeper our sleep (Bjorvatn and Pallesen 2009).
The circadian clock gates sleep and wakefulness in such a way, that it occurs in synchrony with the LD cycle of the environment. The clock stops the production of melatonin and increases cortisol secretion and heart rate 2–3 h prior to waking up (Cajochen et al. 2010).
The effects of light on sleep have been reviewed by Sadeghniiat-Haghighi et al. (2011), Zamanian et al. (2010), and Czeisler and Gooley (2007), and various treatments of sleep disorders are discussed by Thorpy (2011), Dodson and Zee (2010), Bjorvatn and Pallesen (2009), and Blythe et al. (2009).
Common sleep disorders are (Reid et al. 2011; Barion and Zee 2007; Fahey and Zee 2006):
-
The rapid time zone change syndrome (jet lag) with excessive sleepiness and a reduced alertness during daytime in people who travel across time zones (Auger and Morgenthaler 2009; Dean et al. 2009).
-
The shift work sleep disorder in people who frequently rotate shifts or work at night (Kolla and Auger 2011).
-
The delayed sleep phase syndrome; affected people (typically adults) fall asleep at very late times and have difficulties in waking up in time (Rahman et al. 2009; Okawa and Uchiyama 2007).
-
The advanced sleep phase syndrome, in which sleep is advanced (frequent in the elderly); affected people are sleepy in the evening; sleep onset and waking up is early (Caruso and Hitchcock 2010). This syndrome is associated with a mutation in Per2, a clock gene (Xu et al. 2005; Toh et al. 2001).
-
The non-24 h sleep-wake disorder, in which an individual has a normal sleep pattern but a period deviating from 24 h (Okawa and Uchiyama 2007). This syndrome might be caused by weakened or missing stimuli. In blind people, this disorder is more frequently found (Skene and Arendt 2007; Das et al. 2006). Timed melatonin or Zopiclone treatment and/or bright light treatment was successful in this syndrome but is often not used (fewer than 15 % of the 3,000 blind and visually impaired New Zealanders have been prescribed melatonin Warman et al. 2011).
-
Irregular sleep-wake rhythm disorder, in which a circadian pattern is lacking. It occurs in aging and with neurodegenerative diseases, such as Alzheimer’s disease, but also in traumatic brain injury and in mentally retarded persons (Zee and Vitiello 2009). Causes are degeneration or decreased neuronal activity of SCN neurons, decreased response of the clock to zeitgeber such as light and activity, too weak exposure to bright light, lack of social contact and physical activity during the day. This disorder is treated by consolidating sleep during the night and wakefulness during the day mainly by restoring or enhancing SCN zeitgeber. Pharmacologic treatments were negative or inconsistent,
-
Sleep disturbance in psychiatric disorders (Harvey et al. 2011). Clock genes and dopamin/serotonin have been linked to a range of disorders.
Sleep protection and sleep hygiene are important for the well being of adults, children and babies including premature newborns. For the development of the latter and a relaxed condition and good clinical conditions, oral feeding and a close parent-infant relationship are important. A correct sleeping-awaking pattern is a key factor for the development of the brain. To protect sleep of newborns, the environment has to be modified in such a way, that the noise level is reduced, a dark (or at least a semi-dark) period is offered and the face of the infant protected from direct light (Colombo and Bon 2011).
According to Bruni and Novelli (2010) about 20–30 % of young children are affected by sleep disorders (problems getting to sleep = dyssomnias, sleep terrors and sleepwalking = parasomnias). Besides medication such as melatonin, light therapy and scheduled waking is used as a safe and protective intervention for parasomnias (Montgomery and Dunne 2007).
Gradisar et al. (2011) point to the high incidence of sleep disturbance in the youth and in adults (period 1999–2010 reviewed). Sleep onset is delayed in school students, and this delay increases with age of students, which shortens night sleep and increases daytime sleepiness. Begin of weekend sleep is 2 h later and longer. A worldwide delayed sleep-wake behavior pattern exists and delayed sleep phase disorder is wide spread. A fixed advanced sleep/wake schedule with morning blue-light exposure advances circadian phase (Sharkey et al. 2011). How important short-wavelength light in the morning is has been shown by Figueiro and Rea (2010) and is relevant to lighting practice in schools. Wolfson and Carskadon (2003) discuss the effects of advanced sleep disorder on performance and schoolwork. Kohyama (2011) reports that more than 50 % of interviewed Japanese children complained of daytime sleepiness, 25 % of insomnia, and some of both. It is due to asynchronization, brought about by a combination of nighttime light exposure disturbing the clock and decreased melatonin secretion, and a lack of morning light exposure, which prevents synchronization of the clock to the 24 h cycle and reduces the activity of the serotonergic system. An early phase of asynchronization results from inadequate sleep hygiene. It can easily be resolved by a regular sleep-wake cycle. Without adequate intervention the symptoms worsen and become chronic.
Sleep disorders are common in millions of older adults, which might be partly due to the aging processes of the eye lens. As a result the transmission of blue light to the retina is reduced. The entrainment of the circadian clock is hampered and the risk of sleep disturbances increases. Kessel et al. (2011) showed a positive correlation between lens aging and sleep disorders and propose that this results from a disturbed entrainment.
Additionally, environmental and physiological conditions weaken synchronization in the aged. A regular 24 h LD cycle promotes synchronization. Evening light exposure benefits older adults with early evening sleepiness and does not influence the quality of the subsequent sleep (Münch et al. 2011); low irradiances and 90 min exposure are already sufficient (Figueiro et al. 2011). Further studies are needed (Schmoll et al. 2011; Figueiro et al. 2009; Gammack 2008).
In treating sleep disorders, shiftwork and jet lag, light therapy and melatonin administration are effectively used, whereby the timing of the light exposure are important (Gooley 2008; Lack and Wright 2007; Dagan and Borodkin 2005); for practical advices see Dumont and Beaulieu (2007).
Subjects in polar regions, where suboptimal light conditions prevail during the dark part of the year were studied by Arendt (2012). Their health is affected, and the same is true for people in temperate zones with insufficient light exposure. If the intensity of light exposure was increased, circadian phase advanced and sleep improved. Light rich in blue is more effective than white light. In polar regions at the south base personnel adapt their circadian system to night work within a week, whereas in temperate zones this rarely occurs. The same was found on high-latitude North Sea oil installations. In both cases conflicts with the environmental light are absent, which could explain the faster adaptation.
18.8.4 Shift Work and Cancer
Shift work has increased worldwide and was classified as a potential cancer risk by the International Agency for Research on Cancer in 2007; for background and practical information, see Erren et al. (2010) and Erren (2010). Sufficient evidence from animal experiments exist that light exposure during the biologic night increases tumor growth. There is some evidence that night-shift work causes breast cancer (Hansen and Stevens 2011), for which the light exposure during the night could be responsible by reducing melatonin production (Reed 2011).
If night-shift work should indeed increase breast cancer rate, blindness should lower the risk and nighttime illumination in communities should increase it. This was confirmed by studies (Stevens 2009b). Furthermore, clock genes have been related to breast cancer risk, particularly a Per3 mutation. The same mutation predicts a chronobiological morning type and shorter sleep duration. Epigenetic influences on clock genes might be caused by night light (Stevens 2009a).
However, phase shift of the circadian rhythm, sleep disruption, lifestyle factors such as diet, less physical activity and a high BMI and lower vitamin D as well as endocrine influences due to a disturbed clock (Fritschi et al. 2011; Costa et al. 2010) could be additional factors increasing cancer risk (Humble 2010; Bertone-Johnson 2009). Independent of melatonin there is an interaction between sleep and the immune system: Sleep disturbances can suppress the immune system and increase cancer-stimulatory cytokines. However, the causes of cancer are complex and manyfold. An undisturbed and good sleep without light interruption could be a way of reducing the cancer risk (Blask 2009).
18.8.5 Affective Disorders, Endogenous Depressions
Circadian rhythms are not influenced by jet lag, shift work and sleep disorders only, but also by several psychiatric disorders. They include affective disorders, in which the amplitude and phase of several rhythms are altered. It has therefore been suggested that the clock is changed or its sensitivity to zeitgeber such as light or social cues. Non-pharmacological (light therapy, sleep deprivation) and pharmacological (lithium, antidepressants, agomelatine, vitamin D) (Parker and Brotchie 2011, and Chapter 23) therapies of affective disorders influence circadian rhythms, which might indicate that they play a role in these disorders (Schulz and Steimer 2009).
There are several connections between endogenous depression and circadian rhythms (Chellappa et al. 2009; Germain and Kupfer 2008; Lamont et al. 2007; McClung 2007), the causes of which are not well understood. Animal models could help here (Kronfeld-Schor and Einat 2012; Workman and Nelson 2011; Ashkenazy et al. 2009a, b). In depressed patients the phase relationship of the circadian rhythm to the light sensitivity could be anomalous due to some defect in the retina (Steiner et al. 1987). Treatments which affect the circadian rhythms and the sensitivity of the retina to light have a therapeutic effect (Even et al. 2008; Terman and Terman 1999).
A number of disorders in humans are caused by circadian desynchrony as a result of improper light schedules. The physiological responses are discussed by Antle et al. (2009). Clinical aspects of human circadian rhythms are described by Klerman (2005) and practical aspects of chronotherapeutics and chronopharmacological aspects by Wirz- Justice and Terman (2012), Ohdo et al. (2011), Levi and Schibler (2007), and Benedetti et al. (2007).
Four subtypes of depression can be self-treated using agents without prescription: neuroticism with Hypericum perforatum and antihistamines, malaise (fatigue, aching, etc.) in cases of an activated immune system with analgesics, demotivation with energizing agents, and SAD with bright morning light. Melancholia, however, cannot be self-treated and requires hospitalization, if severe (Charlton 2009).
18.8.5.1 SAD: A Recurrent Depression
A special type of depression is the seasonal affective disorder (SAD). It was described by Rosenthal et al. (1984), and his book (Rosenthal 2006) is a standard introduction. Patients show typical symptoms of major depressive disorder, but the depressive episodes occur at a characteristic time of the year. It affects 0.4–2.9 % of the US population. The figures vary between different studies (Howland 2009a; Westrin and Lam 2007a; Winkler et al. 2006), countries, and cultures (Kasof 2009). SAD is found also in children (Giedd et al. 1998), but not as commonly as in young adults, and is more frequent in women. SAD occurs at various latitudes (Kegel et al. 2009; Hansen et al. 2008; Mersch et al. 1999) and in both hemispheres (Brancaleoni et al. 2009; Teng et al. 1995). In polar regions SAD is rare, but a milder form is reported (Magnusson and Partonen 2005).
SAD occurs usually during fall or winter (Madsen et al. 2012). The cognitive and emotional symptoms are as in other types of depression, but the vegetative symptoms are reversed, namely, longer sleep and increased appetite. During spring the symptoms disappear due to the daylight or a light therapy. The latter is effective both in seasonal and in nonseasonal depression (Fischer et al. 2012; Pail et al. 2011; Howland 2009b; Shirani and Louis 2009; Prasko 2008; Even et al. 2008; Terman 2007; Michalak et al. 2007; Levitan 2005 and Golden et al. 2005), but it has, however, to be continued during the winter. It is recommended against SAD in Canadian, American, and international clinical guidelines.
Bright light is more effective than dim light (Rastad et al. 2011), although room light does work in mild cases (Rastad et al. 2008). An exposure of 10 min is already sufficient (Tanaka et al. 2011) and even the light of light-emitting diodes of a “litebook” screen (Desan et al. 2007). Effective doses depend on the individual and range from 10,000 lx for 30 min/day for 8 weeks to 3,000 lx for 2 h/day for 5 weeks. Patients sit comfortably in front of the light box and glance occasionally at it. The amount of light and the wavelength was studied by Anderson et al. (2009). Blue light (Gordijn et al. 2012; Pail et al. 2011; Strong et al. 2009) or blue-light-enriched white light (Meesters et al. 2011) is more effective than standard treatment and red light (Gagné et al. 2011).
Light therapy works for 20–50 % of SAD patients only (Terman et al. 1996). Therefore, alternatives or additional methods are used such as cognitive-behavioral therapy (Rohan et al. 2009), negative air ions (Flory et al. 2010), and pharmacological treatments using antidepressants (Westrin and Lam 2007b; Winkler et al. 2006). Second-generation antidepressants (fluoxetine, escitalopram, duloxetine, reboxetine) are, however, not effective (Thaler et al. 2011). An alternative way of applying light for SAD treatment via the ear canal was used by Timonen et al. (2012). Predictors of response to light therapy were studied by Privitera et al. (2010). Light might exert its effect via the retina of SAD patients, but this has not been verified yet (Lavoie et al. 2009).
According to the phase shift hypothesis, SAD patients become depressed, because the circadian clock is delayed with respect to the sleep/wake cycle. The severity of symptoms does indeed correlate with the degree of misalignment. As a therapy, light exposure in the morning phase advances the clock and restores the correct phase relationship. An alternative treatment is afternoon/evening low-dose melatonin application.
A subgroup of SAD patients is phase advanced, and light should be administered in the evening and melatonin in the morning, but the other type is predominant (Lewy et al. 2006a, b, 2007, 2009).
The duration of melatonin secretion may be influenced in SAD (Salva et al. 2011; Srinivasan et al. 2006), and therefore, melatonin might work as a therapy (Lewy 2007; Lewy et al. 2006a, b). The binding of monoaminergic ligands in the brain changes seasonally, and brain monoamine transmission is involved in many psychiatric disorders. Seen from a phylogenetic standpoint, monoamines adapt the organisms and cells to seasonal changes in the environment such as light, temperature, and energy resources (Praschak-Rieder and Willeit 2012; Ciarleglio et al. 2011; Willeit et al. 2008).
In this connection it has been discussed, whether SAD might reflect some kind of photoperiodic reaction in humans (Howland 2009b; Levitan 2007; Bronson 2004; Wehr 2001; Roenneberg and Aschoff 1990). Photoperiodism in primates is well known (Cayetanot et al. 2005; Oster et al. 2002; Wehr 2001; Di Bitetti and Janson 2000). Sleep, body weight, mood, and behavior in humans are under seasonal control (Cizza et al. 2011). However, the influence of temperature fluctuations and day length is nowadays much lower due to temperature control and the use of artificial light. Sleep has shortened, more food is consumed, and the long-term weight has increased. Especially in women, who are more predisposed to seasonality, SAD might be analogous to hibernation. It is characterized by depressed mood, hypersomnia, weight gain, and carbohydrate craving during the winter.
Hereditary factors seem also to be involved in SAD, as evidenced by family history, twin studies, and molecular genetics studies (Howland 2009a). The mutated gene expressing melanopsin is supposed to increase the risk of SAD (Roecklein et al. 2009), reactions of the hypothalamus to light might be abnormal (Vandewalle et al. 2011), and clock genes might be involved in mental disorders (Lamont et al. 2007; McClung 2007).
There are many questions unanswered, before the light/SAD/clock relations are settled (Klerman 2005). General (Partonen and Magnusson 2001) and special literature on SAD are by Praschak-Rieder and Willeit (2012), Lewy et al. (2006a, b), and two articles in Touitou (1998), and for management of SAD, see Author (2009) and Lam and Levitan (2000).
18.9 Final Remarks and Acknowledgements
In this chapter, we tried to give an overview of circadian rhythms and their entrainment by light by selecting examples from various organisms. The choices are subjective but cover the widespread occurrence of these rhythms ranging from cyanobacteria to human beings.
The reader will have noticed that the field is a wide one and developing fast, for instance, by comparing the present edition of the book with the second (2008) and first one (2008). It is not easy to keep up with new results and sometimes difficult to judge in cases of controversial findings and opinions. We have tried to cover results published in a large number of papers (more than 700 cited). To restrict the number of references, we have preferred to cite reviews and relatively recent papers. If more detailed references are needed, contact the third author at engelmann@uni-tuebingen.de.
We acknowledge help of many colleagues providing literature, information, and critique, especially Lars Olof Björn, Jonathan Emens, Susan Golden, Carl Johnson, Patricia Lakin-Thomas, Stephan Michel, Takao Kondo, Peter Ruoff, Ueli Schibler, Dorothea Staiger, Stephan Steinlechner, Michael Young, Hinrich Lüning, Påvel Nicklasson, François-Yves Bouget, and Marc Lefranc and Dennis Pauls and Christian Wegener, Würzburg.
References
Able KP (1995) Orientation and navigation: a perspective on fifty years of research. Condor 97:592–604
Abraham U, Granada AE, Westermark PO, Heine M, Kramer A, Herzel H (2010) Coupling governs entrainment range of circadian clocks. Mol Syst Biol 6:438
Adams S, Carré IA (2011) Downstream of the plant circadian clock: output pathways for the control of physiology and development. Essays Biochem 49:53–69
Agosto J, Choi JC, Parisky KM, Stilwell G, Rosbash M, Griffith LC (2008) Modulation of GABAA receptor desensitization uncouples sleep onset and maintenance in Drosophila. Nat Neurosci 11:354–359
Åkerstedt T (1998) Is there an optimal sleep-wake pattern in shift work? Scand J Work Environ Health 24(Suppl 3):18–27
Akiyama S (2012) Structural and dynamic aspects of protein clocks: how can they be so slow and stable? Cell Mol Life Sci 69:2147–2160
Allada R, Chung BY (2010) Circadian organization of behavior and physiology in Drosophila. Annu Rev Physiol 72:605–624
Amasino R (2010) Seasonal and developmental timing of flowering. Plant J 61:1001–1013
Amdaoud M, Vallade M, Weiss-Schaber C, Mihalcescu I (2007) Cyanobacterial clock, a stable phase oscillator with negligible intercellular coupling. Proc Natl Acad Sci U S A 104:7051–7056
Anderson JL, Glod CA, Dai J, Cao Y, Lockley SW (2009) Lux vs. wavelength in light treatment of seasonal affective disorder. Acta Psychiatr Scand 120:203–212
Antle MC, Smith VM, Sterniczuk R, Yamakawa GR, Rakai BD (2009) Physiological responses of the circadian clock to acute light exposure at night. Rev Endocr Metab Disord 10:279–291
Aoki S, Kondo T, Wada H, Ishiura M (1997) Circadian rhythm of the cyanobacterium Synechocystis sp. strain PCC 6803 in the dark. J Bacteriol 179:5751–5755
Arendt J (2005) Melatonin: characteristics, concerns, and prospects. J Biol Rhythms 20:291–303
Arendt J (2009) Managing jet lag: some of the problems and possible new solutions. Sleep Med Rev 13:249–256
Arendt J (2010) Shift work: coping with the biological clock. Occup Med 60:10–20
Arendt J (2012) Biological rhythms during residence in polar regions. Chronobiol Int 29:379–394
Ashkenazi I, Hartman H, Strulovitz B, Dar O (1975) Activity rhythms of enzymes in human red blood cell suspension. J Interdisc Cycle Res 6:291–301
Ashkenazy T, Einat H, Kronfeld-Schor N (2009a) Effects of bright light treatment on depression- and anxiety-like behaviors of diurnal rodents maintained on a short daylight schedule. Behav Brain Res 201:343–346
Ashkenazy T, Einat H, Kronfeld-Schor N (2009b) We are in the dark here: induction of depression- and anxiety-like behaviours in the diurnal fat sand rat, by short daylight or melatonin injections. Int J Neuropsychopharmacol 12:83–93
Auger RR, Morgenthaler TI (2009) Jet lag and other sleep disorders relevant to the traveler. Travel Med Infect Dis 7:60–68
Auldridge ME, Forest KT (2011) Bacterial phytochromes: more than meets the light. Crit Rev Biochem Mol Biol 46:67–88
Anonymous (2009) Management of seasonal affective disorder. Drug Ther Bull 47:128–132
Axmann IM, Dühring U, Seeliger L, Arnold A, Vanselow JT, Kramer A, Wilde A (2009) Biochemical evidence for a timing mechanism in Prochlorococcus. J Bacteriol 191:5342–5347
Bachleitner W, Kempinger L, Wülbeck C, Rieger D, Helfrich-Förster C (2007) Moonlight shifts the endogenous clock of Drosophila melanogaster. Proc Natl Acad Sci U S A 104:3538–3543
Baggs JE, Hogenesch JB (2010) Genomics and systems approaches in the mammalian circadian clock. Curr Opin Genet Dev 20:581–587
Bailes HJ, Lucas RJ (2010) Melanopsin and inner retinal photoreception. Cell Mol Life Sci 67:99–111
Baker CL, Loros JJ, Dunlap JC (2012) The circadian clock of Neurospora crassa. FEMS Microbiol Rev 36:95–110
Barger LK, Cade BE, Ayas NT, Cronin JW, Rosner B, Speizer FE, Czeisler CA (2005) Extended work shifts and the risk of motor vehicle crashes among interns. N Engl J Med 352:125–134
Barion A, Zee PC (2007) A clinical approach to circadian rhythm sleep disorders. Sleep Med 8:566–577
Barrenetxe J, Delagrange P, Martinez JA (2004) Physiological and metabolic functions of melatonin. J Physiol Biochem 60:61–72
Barth M, Schultze M, Schuster CM, Strauss R (2010) Circadian plasticity in photoreceptor cells controls visual coding efficiency in Drosophila melanogaster. PLoS One 5:e9217
Bass J, Takahashi JS (2010) Circadian integration of metabolism and energetics. Science 330:1349–1354
Beaule C, Amir S (2003) The eyes suppress a circadian rhythm of fos expression in the suprachiasmatic nucleus in the absence of light. Neuroscience 121:253–257
Beaver LM, Gvakharia BO, Vollintine TS, Hege DM, Stanewsky R, Giebultowicz JM (2002) Loss of circadian clock function decreases reproductive fitness in males of Drosophila melanogaster. Proc Natl Acad Sci USA 19:2134–2139
Beersma DGM (2005) Why and how do we model circadian rhythms? J Biol Rhythms 20:304–313
Bell-Pedersen D (2000) Understanding circadian rhythmicity in Neurospora crassa: from behavior to genes and back again. Fungal Genet Biol 29:1–18
Bell-Pedersen D, Cassone V, Earnest D, Golden S, Hardin P, Thomas T, Zoran M (2005) Circadian rhythms from multiple oscillators: lessons from diverse organisms. Nat Rev Genet 6:544–556
Bellet MM, Sassone-Corsi P (2010) Mammalian circadian clock and metabolism - the epigenetic link. J Cell Sci 123:3837–3848
Bellingham J, Chaurasia S, Melyan Z, Liu C, Cameron M, Tarttelin E, Iuvone P, Hankins M, Tosini G, Lucas R (2006) Evolution of melanopsin photoreceptors: discovery and characterization of a new melanopsin in nonmammalian vertebrates. PLoS Biol 4:e254
Belozerskaya TA, Gessler NN, Isakova EP, Deryabina YI (2012) Neurospora crassa light signal transduction is affected by ROS. J Signal Transduct 2012:791963
Belvin MP, Zhou H, Yin JC (1999) The Drosophila dCREB2 gene affects the circadian clock. Neuron 22:777–787
Benedetti B, Barbini B, Colombo C, Smeraldi E (2007) Chronotherapeutics in a psychiatric ward. Sleep Med Rev 11:509–522
Benito J, Houl JH, Roman GW, Hardin PE (2008) The blue-light photoreceptor cryptochrome is expressed in a subset of circadian oscillator neurons in the Drosophila cns. J Biol Rhythms 23:296–307
Berndt A, Kottke T, Breitkreuz H, Dvorsky R, Hennig S, Alexander M, Wolf E (2007) A novel photoreaction mechanism for the circadian blue light photoreceptor Drosophila cryptochrome. J Biol Chem 282:13011–13021
Berson DM (2003) Strange vision: ganglion cells as circadian photoreceptor. Trends Neurosci 26:314–320
Bertolucci C, Foà A (2004) Extraocular photoreception and circadian entrainment in nonmammalian vertebrates. Chronobiol Int 21:501–519
Bertone-Johnson ER (2009) Vitamin D and the occurrence of depression: causal association or circumstantial evidence? Nutr Rev 67:481–492
Di Bitetti MS, Janson CH (2000) When will the stork arrive? Patterns of birth seasonality in neotropical primates. Am J Primatol 50:109–130
Bjorvatn B, Pallesen S (2009) A practical approach to circadian rhythm sleep disorders. Sleep Med Rev 13:47–60
Blask DE (2009) Melatonin, sleep disturbance and cancer risk. Sleep Med Rev 13:257–264
Blythe J, Doghramji PP, Jungquist CR, Landau MB, Valerio TD, Ancoli-Israel S, Auerbach SH (2009) Screening & treating patients with sleep/wake disorders. JAAPA Suppl Sleep, 1–17; quiz 19
Bobu C, Hicks D (2009) Regulation of retinal photoreceptor phagocytosis in a diurnal mammal by circadian clocks and ambient lighting. Invest Ophthalmol Vis Sci 50:3495–3502
Bodenstein C, Heiland I, Schuster S (2011) Calculating activation energies for temperature compensation in circadian rhythms. Phys Biol 8:056007
Boesger J, Wagner V, Weisheit W, Mittag M (2009) Analysis of flagellar phosphoproteins from Chlamydomonas reinhardtii. Eukaryot Cell 8:922–932
Boivin DB, James FO (2005) Light treatment and circadian adaptation to shift work. Ind Health 43:34–48
Bollig I, Chandrashekaran M, Engelmann W, Johnsson A (1976) Photoperiodism in Chenopodium rubrum – an explicit version of the Bünning hypothesis. Int J Chronobiol 4:83–96
Borjigin J, Zhang LS, Calinescu AA (2012) Circadian regulation of pineal gland rhythmicity. Mol Cell Endocrinol 349:13–19
Bradley RL, Reddy KJ (1997) Cloning, sequencing, and regulation of the global nitrogen regulator gene ntca in the unicellular diazotrophic cyanobacterium Cyanothece sp. strain BH68K. J Bacteriol 179:4407–4410
Brainard GC, Hanifin JP (2005) Photons, clocks, and consciousness. J Biol Rhythms 20:314–325
Brainard GC, Richardson BA, King TS, Matthews SA, Reiter RJ (1983) The suppression of pineal melatonin content and N-acetyltransferase activity by different light irradiances in the Syrian hamster: a dose-response relationship. Endocrinology 113:293–296
Brainard GC, Sliney G, Hanifin JP, Glickman G, Byrne B, Greeson JM, Jasser S, Gerner E, Rollag MD (2008) Sensitivity of the human circadian system to short-wavelength (420-nm) light. J Biol Rhythms 23:379–386
Brancaleoni G, Nikitenkova E, Grassi L, Hansen V (2009) Seasonal affective disorder and latitude of living. Epidemiol Psichiatr Soc 18:336–343
Breton G, Kay SA (2006) Circadian rhythms lit up in Chlamydomonas. Genome Biol 7:215
Brody S, Oelhafen K, Schneider K, Perrino S, Goetz A, Wang C, English C (2010) Circadian rhythms in Neurospora crassa: downstream effectors. Fungal Genet Biol 47:159–168
Bronson FH (2004) Are humans seasonally photoperiodic? J Biol Rhythms 19:180–192
Brown SA, Kowalska E, Dallmann R (2012) (Re)inventing the circadian feedback loop. Dev Cell 22:477–487
Bruce VG (1970) The biological clock in Chlamydomonas reinhardi. J Protozool 17:328–334
Bruce VG (1972) Mutants of the biological clock in Chlamydomonas reinhardi. Genetics 70:537–548
Bruni O, Novelli L (2010) Sleep disorders in children. Clin Evid (Online)
Brunner M, Káldi K (2008) Interlocked feedback loops of the circadian clock of Neurospora crassa. Mol Microbiol 68:255–262
Brunner M, Schafmeier T (2006) Transcriptional and post-transcriptional regulation of the circadian clock of cyanobacteria and Neurospora. Genes Dev 20:1061–1074
Brunner M, Simons MJP, Merrow M (2008) Lego clocks: building a clock from parts. Genes Dev 22:1422–1426
Bryant TR (1972) Gas exchange in dry seeds: circadian rhythmicity in the absence of DNA replication, transcription, and translation. Science 178:634–636
Bushey D, Cirelli C (2011) From genetics to structure to function: exploring sleep in Drosophila. Int Rev Neurobiol 99:213–244
Butler MP, Rainbow MN, Rodriguez E, Lyon SM, Silver R (2012) Twelve-hour days in the brain and behavior of split hamsters. Eur J Neurosci 36:2556–2566
Byrne TE, Wells MR, Johnson CH (1992) Circadian rhythms of chemotaxis to ammonium and of methylammonium uptake in Chlamydomonas. Plant Physiol 98:879–886
Bünning E (1936) Die endonome Tagesrhythmik als Grundlage der photoperiodischen Reaktion. Ber Deut Bot Ges 54:590–607
Cajochen C, Chellappa S, Schmidt C (2010) What keeps us awake? The role of clocks and hourglasses, light, and melatonin. Int Rev Neurobiol 93:57–90
Campbell SS, Murphy PJ (1998) Extraocular circadian phototransduction in humans. Science 279:396–399
Cardinali D (1998) The human body circadian: how the biological clock influences sleep and emotion. Cienc Cult 50:172–177
Caruso CC, Hitchcock EM (2010) Strategies for nurses to prevent sleep-related injuries and errors. Rehabil Nurs 35:192–197
Castro-Longoria E, Ferry M, Bartnicki-Garcia S, Hasty J, Brody S (2010) Circadian rhythms in Neurospora crassa: dynamics of the clock component frequency visualized using a fluorescent reporter. Fungal Genet Biol 47:332–341
Cayetanot F, van Someren EJW, Perret M, Aujard F (2005) Shortened seasonal photoperiodic cycles accelerate aging of the diurnal and circadian locomotor activity rhythms in a primate. J Biol Rhythms 20:461–469
Ceriani M, Darlington T, Staknis D, Mas P, Petti A, Weitz C, Kay S (1999) Light-dependent sequestration of TIMELESS by CRYPTOCHROME. Science 285:553–568
Cermakian N, Boivin DB (2009) The regulation of central and peripheral circadian clocks in humans. Obes Rev 10(Suppl 2):25–36
Cervený J, Nedbal L (2009) Metabolic rhythms of the cyanobacterium Cyanothece sp. ATCC 51142 correlate with modeled dynamics of circadian clock. J Biol Rhythms 24:295–303
Challet E (2007) Minireview: entrainment of the suprachiasmatic clockwork in diurnal and nocturnal mammals. Endocrinology 148:5648–5655
Chandrashekaran MK, Engelmann W (1973) Early and late subjective night phase of the Drosophila rhythm require different energies of blue light for phase shifting. Z Naturforsch 28c:750–753
Charlton BG (2009) A model for self-treatment of four sub-types of symptomatic ‘depression’ using non-prescription agents: neuroticism (anxiety and emotional instability); malaise (fatigue and painful symptoms); demotivation (anhedonia) and seasonal affective disorder ‘SAD’. Med Hypotheses 72:1–7
Chaves I, Pokorny R, Byrdin M, Hoang N, Ritz T, Brettel K, Essen L-O, van der Horst GTJ, Batschauer A, Ahmad M (2011) The cryptochromes: blue light photoreceptors in plants and animals. Annu Rev Plant Biol 62:335–364
Chellappa SL, Schröder C, Cajochen C (2009) Chronobiology, excessive daytime sleepiness and depression: is there a link? Sleep Med 10:505–514
Chen CH, Dunlap JC, Loros JJ (2010) Neurospora illuminates fungal photoreception. Fungal Genet Biol 47:922–929
Cho H, Zhao X, Hatori M, Yu RT, Barish GD, Lam MT, Chong L-W, Di- Tacchio L, Atkins AR, Glass CK, Liddle C, Auwerx J, Downes M, Panda S, Evans RM (2012) Regulation of circadian behaviour and metabolism by REV-ERB-a and REV-ERB-b. Nature 485:123–127
Choi C, Nitabach MN (2010) Circadian biology: environmental regulation of a multioscillator network. Curr Biol 20:R322–R324
Chow BY, Helfer A, Nusinow DA, Kay SA (2012) ELF3 recruitment to the PRR9 promoter requires other evening complex members in the Arabidopsis circadian clock. Plant Signal Behav 7:170–173
Chung BY, Kilman VL, Keath JR, Pitman JL, Allada R (2009) The GABA(A) receptor RDL acts in peptidergic PDF neurons to promote sleep in Drosophila. Curr Biol 19:386–390
Ciarleglio CM, Resuehr HES, McMahon DG (2011) Interactions of the serotonin and circadian systems: nature and nurture in rhythms and blues. Neuroscience 197:8–16
Cizza G, Requena M, Galli G, de Jonge L (2011) Chronic sleep deprivation and seasonality: implications for the obesity epidemic. J Endocrinol Invest 34:793–800
Clauser C (1954) Die Kopfuhr. Ferdinand Enke, Stuttgart
Collett TS (2008) Insect navigation: visual panoramas and the sky compass. Curr Biol 18:R1058–R1061
Collin SP, Davies WL, Hart NS, Hunt DM (2009) The evolution of early vertebrate photoreceptors. Philos Trans R Soc Lond B Biol Sci 364:2925–2940
Colombo G, Bon GD (2011) Strategies to protect sleep. J Matern Fetal Neonatal Med 24(Suppl 1):30–31
Colot HV, Park G, Turner GE, Ringelberg C, Crew CM, Litvinkova L, Weiss RL, Borkovich KA, Dunlap JC (2006) A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc Natl Acad Sci U S A 103:10352–10357
Colwell CS (2011) Linking neural activity and molecular oscillations in the SCN. Nat Rev Neurosci 12:553–569
Corellou F, Schwartz C, Motta J-P, Djouani-Tahri EB, Sanchez F, Bouget F-Y (2009) Clocks in the green lineage: comparative functional analysis of the circadian architecture of the picoeukaryote Ostreococcus. Plant Cell 21:3436–3449
Correa A, Bell-Pedersen D (2002) Distinct signaling pathways from the circadian clock participate in regulation of rhythmic conidiospore development in Neurospora crassa. Eukaryot Cell 1:273–280
Correa A, Lewis ZA, Greene AV, March IJ, Gomer RH, Bell-Pedersen D (2003) Multiple oscillators regulate circadian gene expression in Neurospora. Proc Natl Acad Sci U S A 100:13597–13602
Costa G, Haus E, Stevens R (2010) Shift work and cancer – considerations on rationale, mechanisms, and epidemiology. Scand J Work Environ Health 36:163–179
Coste O, Lagarde D (2009) Clinical management of jet lag: what can be proposed when performance is critical? Travel Med Infect Dis 7:82–87
Courties C, Chretiennot-Dinet MJ (1994) Smallest eukaryotic organism. Nature 370:255
Covington MF, Pandab S, Liu XL, Strayer CA, Wagner DR, Kay SA (2001) ELF3 modulates resetting of the circadian clock in Arabidopsis. Plant Cell 13:1305–1316
Crosthwaite SK, Dunlap JC, Loros JJ (1997) Neurospora wc-1 and wc-2: transcription, photoresponses, and the origins of circadian rhythmicity. Science 276:763–769
Crowley SJ, Lee C, Tseng CY, Fogg LF, Eastman CI (2003) Combinations of bright light, scheduled dark, sunglasses, and melatonin to facilitate circadian entrainment to night shift work. J Biol Rhythms 18:513–523
Cyran S, Yiannoulos G, Buchsbaum A, Saez L, Young M, Blau J (2005) The double-time protein kinase regulates the subcellular localization of the Drosophila clock protein period. J Neurosci 25:5430–5437
Czeisler CA, Gooley JJ (2007) Sleep and circadian rhythms in humans. Cold Spring Harb Symp Quant Biol 72:579–597
Czeisler C, Kronauer R, Allan J, Duffy J, Jewett M, Brown E, Ronda J (1989) Bright light induction of strong (type 0) resetting of the human circadian pacemaker. Science 244:1328–1333
Czeisler CA, Duffy JF, Shanahan TL, Brown EN, Mitchell JF, Rimmer DW, Ronda JM, Silva EJ, Allan JS, Emens JS, Dijk DJ, Kronauer RE (1999) Stability, precision, and near-24-hour period of the human circadian pacemaker. Science 284:2177–2181
Daan S, Spoelstra K, Albrecht U, Schmutz I, Daan M, Daan B, Rienks F, Poletaeva I, dell’Omo G, Vyssotski A, Lipp HP (2011) Lab mice in the field: unorthodox daily activity and effects of a dysfunctional circadian clock allele. J Biol Rhythms 26:118–129
Daan SA, Albrecht U, van der Horst GTJ, Illnerova H, Roenneberg T, Wehr TA, Schwartz WJ (2001) Assembling a clock for all seasons: are there M and E oscillators in the genes? J Biol Rhythms 16:105–116
Dacke M, Byrne MJ, Baird E, Scholtz CH, Warrant EJ (2011) How dim is dim? Precision of the celestial compass in moonlight and sunlight. Philos Trans R Soc Lond B Biol Sci 366:697–702
Dagan Y, Borodkin K (2005) Behavioral and psychiatric consequences of sleep-wake schedule disorders. Dialogues Clin Neurosci 7:357–365
Dalchau N (2012) Understanding biological timing using mechanistic and black-box models. New Phytol 193:852–858
Das A, Sasmal NK, Deb RK, Bera NK, Sanyal D, Chatterjee SS, Bhaduri G (2006) A study of sleeping disorders in blind patients. J Indian Med Assoc 104:619–621, 626
Davies WIL, Collin SP, Hunt DM (2012) Molecular ecology and adaptation of visual photopigments in craniates. Mol Ecol 21:3121–3158
Davis RH (2000) Neurospora: contributions of a model organism. Oxford University Press, Oxford
De J, Varma V, Sharma VK (2012) Adult emergence rhythm of fruit flies Drosophila melanogaster under seminatural conditions. J Biol Rhythms 27:280–286
Deacon S, Arendt J (1996) Adapting to phase shifts. I. An experimental model for jet lag and shift work. Physiol Behav 59:665–673
Dean DA, Forger DB, Klerman EB (2009) Taking the lag out of jet lag through model-based schedule design. PLoS Comput Biol 5:e1000418
DeCoursey P, Krulas J (1998) Behavior of SCN-lesioned chipmunks in a natural habitat: a pilot study. J Biol Rhythms 13:229–244
Demir-Hilton E, Sudek S, Cuvelier ML, Gentemann CL, Zehr JP, Worden AZ (2011) Global distribution patterns of distinct clades of the photosynthetic picoeukaryote Ostreococcus. ISME J 5:1095–1107
Desan PH, Weinstein AJ, Michalak EE, Tam EM, Meesters Y, Ruiter MJ, Horn E, Telner J, Iskandar H, Boivin DB, Lam RW (2007) A controlled trial of the litebook light-emitting diode (LED) light therapy device for treatment of seasonal affective disorder (SAD). BMC Psychiatry 7:38
Devlin PF (2002) Signs of the time: environmental input to the circadian clock. J Exp Bot 53:1535–1550
Dewan K, Benloucif S, Reid K, Wolfe LF, Zee PC (2011) Light-induced changes of the circadian clock of humans: increasing duration is more effective than increasing light intensity. Sleep 34:593–599
Dharmananda S (1980) Studies on the circadian clock of Neurospora crassa: lightinduced phase shifting. Ph.D. thesis, University of California, Santa Cruz
Dibner C, Sage D, Unser M, Bauer C, d’Eysmond T, Naef F, Schibler U (2009) Circadian gene expression is resilient to large fluctuations in overall transcription rates. EMBO J 28:123–134
Dibner C, Schibler U, Albrecht U (2010) The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol 72:517–549
Diernfellner AC, Schafmeier T, Merrow MW, Brunner M (2005) Molecular mechanism of temperature sensing by the circadian clock of Neurospora crassa. Genes Dev 19:1968–1973
Diernfellner A, Colot HV, Dintsis O, Loros JJ, Dunlap JC, Brunner M (2007) Long and short isoforms of neurospora clock protein frq support temperature compensated circadian rhythms. FEBS Lett 581:5759–5764
Diez-Noguera A (1994) A functional model of the circadian system based on the degree of intercommunication in a complex system. Am J Physiol 267:1118–1135
Dijk D-J, von Schantz M (2005) Timing and consolidation of human sleep, wakefullness, and performance by a symphony of oscillators. J Biol Rhythms 20:279–290
Djouani-Tahri E-B, Christie JM, Sanchez-Ferandin S, Sanchez F, Bouget F-Y, Corellou F (2011a) A eukaryotic LOV-histidine kinase with circadian clock function in the picoalga Ostreococcus. Plant J 65:578–588
Djouani-Tahri EB, Sanchez F, Lozano J-C, Bouget F-Y (2011b) A phosphate regulated promoter for fine-tuned and reversible overexpression in Ostreococcus: application to circadian clock functional analysis. PLoS One 6:e28471
Do MTH, Yau K-W (2010) Intrinsically photosensitive retinal ganglion cells. Physiol Rev 90:1547–1581
Dodd AN, Salathia N, Hall A, Kévei E, Tóth R, Nagy F, Hibberd JM, Millar AJ, Webb AAR (2005) Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 309:630–633
Dodson ER, Zee PC (2010) Therapeutics for circadian rhythm sleep disorders. Sleep Med Clin 5:701–715
Dong G, Golden SS (2008) How a cyanobacterium tells time. Curr Opin Microbiol 11:541–546
Dong G, Kim Y-I, Golden SS (2010a) Simplicity and complexity in the cyanobacterial circadian clock mechanism. Curr Opin Genet Dev 20:619–625
Dong G, Yang Q, Wang Q, Kim Y-I, Wood TL, Osteryoung KW, van Oudenaarden A, Golden SS (2010b) Elevated ATPase activity of KaiC applies a circadian checkpoint on cell division in Synechococcus elongatus. Cell 140:529–539
Dong W, Tang X, Yu Y, Nilsen R, Kim R, Griffith J, Arnold J, Schüttler H-B (2008) Systems biology of the clock in neurospora crassa. PLoS One 3:e3105
Dragovic Z (2002) Light reception and circadian behavior in ‘blind’ and ‘clock-less’ mutants of Neurospora crassa. EMBO J 21:3643–3651
Duffy JF, Wright KP (2005) Entrainment of the human circadian system by light. J Biol Rhythms 20:326–338
Duguay D, Cermakian N (2009) The crosstalk between physiology and circadian clock proteins. Chronobiol Int 26:1479–1513
Dumont M, Beaulieu C (2007) Light exposure in the natural environment: relevance to mood and sleep disorders. Sleep Med 8:557–565
Dunlap J, Loros J (2004) The Neurospora circadian system. J Biol Rhythms 19:414–424
Dunlap J, Loros J (2005) Analysis of circadian rhythms in Neurospora: overview of assays and genetic and molecular biological manipulation. Methods Enzymol 393:3–22
Dunlap JC, Loros JJ (2006) How fungi keep time: circadian system in Neurospora and other fungi. Curr Opin Microbiol 9:579–587
Dunlap JC, Loros JJ, Colot HV, Mehra A, Belden WJ, Shi M, Hong CI, Larrondo LF, Baker CL, Chen C-H, Schwerdtfeger C, Collopy PD, Gamsby JJ, Lambreghts R (2007) A circadian clock in Neurospora: how genes and proteins cooperate to produce a sustained, entrainable, and compensated biological oscillator with a period of about a day. Cold Spring Harb Symp Quant Biol 72:57–68
Duvall LB, Taghert PH (2011) Circadian rhythms: biological clocks work in phospho-time. Curr Biol 21:R305–R307
Duvall LB, Taghert PH (2012) The circadian neuropeptide PDF signals preferentially through a specific adenylate cyclase isoform AC3 in M pacemakers of Drosophila. PLoS Biol 10:e1001337
Eastman CI, Stewart KT, Mahoney MP, Liu L, Fogg LF (1994) Dark goggles and bright light improve circadian rhythm adaptation to night-shift work. Sleep 17:535–543
Eckardt NA (2010) Temperature compensation of the circadian clock: a role for the morning loop. Plant Cell 22:3506
Egli M, Mori T, Pattanayek R, Xu Y, Qin X, Johnson CH (2012) Dephosphorylation of the core clock protein KaiC in the cyanobacterial KaiABC circadian oscillator proceeds via an ATP synthase mechanism. Biochemistry 51:1547–1558
Elvin M, Loros JJ, Dunlap JC, Heintzen C (2005) The PAS/LOV protein VIVID supports a rapidly dampened daytime oscillator that facilitates entrainment of the Neurospora circadian clock. Genes Dev 19:2593–2605
Emens JS, Lewy AJ, Lefler BJ, Sack RL (2005) Relative coordination to unknown ‘weak zeitgebers’ in free-running blind individuals. J Biol Rhythms 20:159–167
Emery P, So W, Kaneko M, Hall J, Rosbash M (1998) CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 95:669–679
Emery P, Stanewsky R, Hall J, Rosbash M (2000) Drosophila cryptochrome – a unique circadian-rhythm photoreceptor. Nature 404:456–457
Engelmann W, Johnsson A, Kobler H, Schimmel M (1978) Attenuation of the petal movement rhythm of Kalanchoe with light pulses. Physiol Behav 43:68–76
Erkert HG, Gburek V, Scheideler A (2006) Photic entrainment and masking of prosimian circadian rhythms (Otolemur garnettii, primates). Physiol Behav 88:39–46
Erren TC (2010) Shift work, cancer and “white-box” epidemiology: association and causation. Epidemiol Perspect Innov 7:11
Erren TC, Falaturi P, Morfeld P, Knauth P, Reiter RJ, Piekarski C (2010) Shift work and cancer: the evidence and the challenge. Dtsch Arztebl Int 107:657–662
Even C, Schröder CM, Friedman S, Rouillon F (2008) Efficacy of light therapy in nonseasonal depression: a systematic review. J Affect Disord 108:11–23
Fahey CD, Zee PC (2006) Circadian rhythm sleep disorders and phototherapy. Psychiatr Clin N Am 29:989–1007; abstract ix
Falcón J, Besseau L, Fuentès M, Sauzet S, Magnanou E, Boeuf G (2009) Structural and functional evolution of the pineal melatonin system in vertebrates. Ann N Y Acad Sci 1163:101–111
Figueiro MG, Rea MS (2010) Lack of short-wavelength light during the school day delays dim light melatonin onset (DLMO) in middle school students. Neuro Endocrinol Lett 31:92–96
Figueiro MG, Bullough JD, Parsons RH, Rea MS (2005) Preliminary evidence for a change in spectral sensitivity of the circadian system at night. J Circadian Rhythm 3:14
Figueiro MG, Bierman A, Bullough JD, Rea MS (2009) A personal light-treatment device for improving sleep quality in the elderly: dynamics of nocturnal melatonin suppression at two exposure levels. Chronobiol Int 26:726–739
Figueiro MG, Lesniak NZ, Rea MS (2011) Implications of controlled short wavelength light exposure for sleep in older adults. BMC Res Notes 4:334
Fiorentino L, Martin JL (2010) Awake at 4 AM: treatment of insomnia with early morning awakenings among older adults. J Clin Psychol 66:1161–1174
Fischer R, Kasper S, Pjrek E, Winkler D (2012) On the application of light therapy in German-speaking countries. Eur Arch Psychiatry Clin Neurosci 262:501–505
Fleissner G, Fleissner G (2001) Perception of natural Zeitgeber signals. In: Kumar V (ed) Biological rhythms. Narosa Publishing House, New Delhi
Fleury F (2000) Adaptive significance of a circadian clock: temporal segregation of activities reduces intrinsic competitive inferiority in Drosophila parasitoids. Proc Biol Sci 267:1005–1010
Flory R, Ametepe J, Bowers B (2010) A randomized, placebo-controlled trial of bright light and high-density negative air ions for treatment of seasonal affective disorder. Psychiatry Res 177:101–108
Folkard S (2008) Do permanent night workers show circadian adjustment? a review based on the endogenous melatonin rhythm. Chronobiol Int 25:215–224
Forger D, Jewett M, Kronauer R (1999) A simpler model of the human circadian pacemaker. J Biol Rhythms 14:532–537
Forsgren E (1935) Über die Rhythmik der Leberfunktion, des Stoffwechsels und des Schlafes. Gumperts Bokhandel, Göteborg
Foster R, Helfrich-Förster C (2001) Photoreceptors for circadian clocks in mice and fruit flies. Philos Trans R Soc Lond B Biol Sci 356 B:1779–1789
Foster RG, Hankins MW, Peirson SN (2007) Light, photoreceptors, and circadian clocks. Methods Mol Biol 362:3–28
Foà A, Basaglia F, Beltrami G, Carnacina M, Moretto E, Bertolucci C (2009) Orientation of lizards in a Morris water-maze: roles of the sun compass and the parietal eye. J Exp Biol 212:2918–2924
Frenkel L, Ceriani MF (2011) Circadian plasticity: from structure to behavior. Int Rev Neurobiol 99:107–138
Fritschi L, Glass DC, Heyworth JS, Aronson K, Girschik J, Boyle T, Grundy A, Erren TC (2011) Hypotheses for mechanisms linking shiftwork and cancer. Med Hypotheses 77:430–436
Froy O (2011) The circadian clock and metabolism. Clin Sci (Lond) 120:65–72
Gachon F, Nagoshi E, Brown SA, Ripperger J, Schibler U (2004) The mammalian circadian timing system: from gene expression to physiology. Chromosoma 113:103–112
Gagné A-M, Lévesque F, Gagné P, Hébert M (2011) Impact of blue vs red light on retinal response of patients with seasonal affective disorder and healthy controls. Prog Neuropsychopharmacol Biol Psychiatry 35:227–231
Gammack JK (2008) Light therapy for insomnia in older adults. Clin Geriatr Med 24:139–149, viii
Gaskill C, Forbes-Stovall J, Kessler B, Young M, Rinehart CA, Jacobshagen S (2010) Improved automated monitoring and new analysis algorithm for circadian phototaxis rhythms in Chlamydomonas. Plant Physiol Biochem 48:239–246
Gendron JM, Pruneda-Paz JL, Doherty CJ, Gross AM, Kang SE, Kay SA (2012) Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc Natl Acad Sci U S A 109:3167–3172
Germain A, Kupfer DJ (2008) Circadian rhythm disturbances in depression. Hum Psychopharmacol 23:571–585
Giedd JN, Swedo SE, Lowe CH, Rosenthal NE (1998) Case series: pediatric seasonal affective disorder. A follow-up report. J Am Acad Child Adolesc Psychiatry 37:218–220
Giedke H, Engelmann W, Reinhard P (1983) Free running circadian rest-activity cycle in normal environment. A case study. Sleep Res 12:365
Gillette M, Abbott S (2005) Basic mechanisms of circadian rhythms and their relation to the sleep/wake cycle. In: Cardinali DP, Perumal SR (eds) Neuroendocrine correlates of sleep/wakefullness. Springer, New York
Girardet C, Becquet D, Blanchard M-P, François-Bellan A-M, Bosler O (2010) Neuroglial and synaptic rearrangements associated with photic entrainment of the circadian clock in the suprachiasmatic nucleus. Eur J Neurosci 32:2133–2142
Goda K, Ito H, Kondo T, Oyama T (2012) Fluorescence correlation spectroscopy to monitor Kai protein-based circadian oscillations in real time. J Biol Chem 287:3241–3248
Goldbeter A (1995) A model for circadian oscillations in the Drosophila period protein (PER). Proc Biol Sci 261:319–324
Golden RN, Gaynes BN, Ekstrom RD, Hamer RM, Jacobsen FM, Suppes T, Wisner KL, Nemeroff CB (2005) The efficacy of light therapy in the treatment of mood disorders: a review and meta-analysis of the evidence. Am J Psychiatry 162:656–662
Golden SS, Cassone VM, LiWang A (2007) Shifting nanoscopic clock gears. Nat Struct Mol Biol 14:362–363
Golombek DA, Rosenstein RE (2010) Physiology of circadian entrainment. Physiol Rev 90:1063–1102
Gonze D, Roussel MR, Goldbeter A (2002) A model for the enhancement of fitness in Cyanobacteria based on resonance of a circadian oscillator with the external lightdark cycle. J Theor Biol 214:577–597
Gooch VD, Mehra A, Larrondo LF, Fox J, Touroutoutoudis M, Loros JJ, Dunlap JC (2008) Fully codon-optimized luciferase uncovers novel temperature characteristics of the Neurospora clock. Eukaryot Cell 7:28–37
Gooley JJ (2008) Treatment of circadian rhythm sleep disorders with light. Ann Acad Med Singapore 37:669–676
Gordijn MCM, t Mannetje D, Meesters Y (2012) The effects of blue-enriched light treatment compared to standard light treatment in seasonal affective disorder. J Affect Disord 136:72–80
Goto K, Johnson CH (1995) Is the cell division cycle gated by a circadian clock? The case of Chlamydomonas reinhardtii. J Cell Biol 129:1061–1069
Gotow T, Nishi T (2008) Simple photoreceptors in some invertebrates: physiological properties of a new photosensory modality. Brain Res 1225:3–16
Grace MS, Wang LA, Pickard GE, Besharse JC, Menaker M (1996) The tau mutation shortens the period of rhythmic photoreceptor outer segment disk shedding in the hamster. Brain Res 735:93–100
Grace MS, Chiba A, Menaker M (1999) Circadian control of photoreceptor outer segment membrane turnover in mice genetically incapable of melatonin synthesis. Vis Neurosci 16:909–918
Gradisar M, Gardner G, Dohnt H (2011) Recent worldwide sleep patterns and problems during adolescence: a review and meta-analysis of age, region, and sleep. Sleep Med 12:110–118
Green CB, Besharse JC (2004) Retinal circadian clocks and control of retinal physiology. J Biol Rhythms 19:91–102
Green CB, Takahashi JS, Bass J (2008) The meter of metabolism. Cell 134:728–742
Green RM, Tingay S, Wang ZY, Tobin EM (2002) Circadian rhythms confer a higher level of fitness to Arabidopsis plants. Plant Physiol 129:576–584
Griefahn B, Robens S (2010) Alterations of the cortisol quiescent period after experimental night work with enforced adaptation by bright light and its relation to morningness. Eur J Appl Physiol 108:719–726
Grima B, Chelot E, Xia R, Rouyer F (2004) Morning and evening peaks of activity rely on different clock neurons of the Drosophila brain. Nature 431:869–873
Grimsley N, Péquin B, Bachy C, Moreau H, Piganeau G (2010) Cryptic sex in the smallest eukaryotic marine green alga. Mol Biol Evol 27:47–54
Grone BP, Chang D, Bourgin P, Cao V, Fernald RD, Heller HC, Ruby NF (2011) Acute light exposure suppresses circadian rhythms in clock gene expression. J Biol Rhythms 26:78–81
Gu C, Wang J, Wang J, Liu Z (2011) Mechanism of phase splitting in two coupled groups of suprachiasmatic-nucleus neurons. Phys Rev E Stat Nonlin Soft Matter Phys 83:046224
Guido ME, Garbarino-Pico E, Contin MA, Valdez DJ, Nieto PS, Verra DM, Acosta-Rodriguez VA, de Zavalía N, Rosenstein RE (2010) Inner retinal circadian clocks and non-visual photoreceptors: novel players in the circadian system. Prog Neurobiol 92:484–504
Guilding C, Piggins HD (2007) Challenging the omnipotence of the suprachiasmatic timekeeper: are circadian oscillators present throughout the mammalian brain? Eur J Neurosci 25:3195–3216
Guo J, Cheng P, Liu Y (2010) Functional significance of FRH in regulating the phosphorylation and stability of Neurospora crassa circadian clock protein FRQ. J Biol Chem 285:11508–11515
Hanai S, Ishida N (2009) Entrainment of drosophila circadian clock to green and yellow light by rh1, rh5, rh6 and cry. Neuroreport 20:755–758
Hanai S, Hamasaka Y, Ishida N (2008) Circadian entrainment to red light in Drosophila: requirement of rhodopsin 1 and rhodopsin 6. Neuroreport 19:1441–1444
Hannibal J, Hindersson P, Knudsen SM, Geor B, Fahrenkrug J (2002) Melanopsin is expressed in PACAP-containing retinal ganglion cells of the human retinohypothalamic tract. Invest Ophthalmol Vis Sci 45:4202–4209
Hansen J, Stevens RG (2012) Case-control study of shift-work and breast cancer risk in Danish nurses: impact of shift systems. Eur J Cancer 48:1722–1729
Hansen JH, Geving IH, Reinertsen RE (2010) Offshore fleet workers and the circadian adaptation of core body temperature, blood pressure and heart rate to 12-h shifts: a field study. Int J Occup Saf Ergon 16:487–496
Hansen V, Skre I, Lund E (2008) What is this thing called “SAD”? a critique of the concept of seasonal affective disorder. Epidemiol Psichiatr Soc 17:120–127
Hardeland R, Cardinali DP, Srinivasan V, Spence DW, Brown GM, Pandi- Perumal SR (2011) Melatonin–a pleiotropic, orchestrating regulator molecule. Prog Neurobiol 93:350–384
Hardeland R, Madrid JA, Tan D-X, Reiter RJ (2012) Melatonin, the circadian multioscillator system and health: the need for detailed analyses of peripheral melatonin signaling. J Pineal Res 52:139–166
Hardie DG, Ross FA, Hawley SA (2012) AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol 13:251–262
Hardin PE (2005) The circadian timekeeping system of Drosophila. Curr Biol 15:R714–R722
Hardin PE (2011) Molecular genetic analysis of circadian timekeeping in Drosophila. Adv Genet 74:141–173
Hardin PE, Hall JC, Rosbash M (1990) Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 343:536–540
Harmer SL (2009) The circadian system in higher plants. Annu Rev Plant Biol 60:357–377
Harmer S (2010) Plant biology in the fourth dimension. Plant Physiol 154:467–470
Harmer S, Hogenesch L, Straume M, Chang H, Han B, Zhu T, Wang X, Kreps J, Kay S (2000) Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 290:2110–2113
Harrisingh MC, Nitabach MN (2008) Circadian rhythms. Integrating circadian timekeeping with cellular physiology. Science 320:879–880
Harvey AG, Murray G, Chandler RA, Soehner A (2011) Sleep disturbance as transdiagnostic: consideration of neurobiological mechanisms. Clin Psychol Rev 31:225–235
Hastings MH, Herzog ED (2004) Clock genes, oscillators, and cellular networks in the suprachiasmatic nuclei. J Biol Rhythms 19:400–413
Hastings MH, Maywood ES, O’Neill JS (2008) Cellular circadian pacemaking and the role of cytosolic rhythms. Curr Biol 18:R805–R815
Hatakeyama TS, Kaneko K (2012) Generic temperature compensation of biological clocks by autonomous regulation of catalyst concentration. Proc Natl Acad Sci U S A 109:8109–8114
Hattar S, Lucas RJ, Mrosovsky N, Thompson S, Douglas RH, Hankins MW, Lem J, Biel M, Hofman F, Foster RG (2003) Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature 424:75–81
Hayama R, Coupland G (2003) Shedding light on the circadian clock and the photoperiodic control of flowering. Curr Opin Plant Biol 6:13–19
He Q, Cheng P, Yang Y, Wang L, Gardner K, Liu Y (2002) White collar-1, a DNA binding transcription factor and a light sensor. Science 297:840–843
Heesy CP, Hall MI (2010) The nocturnal bottleneck and the evolution of mammalian vision. Brain Behav Evol 75:195–203
Hegemann P (2008) Algal sensory photoreceptors. Annu Rev Plant Biol 59:167–189
Heijde M, Ulm R (2012) UV-B photoreceptor-mediated signalling in plants. Trends Plant Sci 17:230–237
Heijde M, Zabulon G, Corellou F, Ishikawa T, Brazard J, Usman A, Sanchez F, Plaza P, Martin M, Falciatore A, Todo T, Bouget F-Y, Bowler C (2010) Characterization of two members of the cryptochrome/photolyase family from Ostreococcus tauri provides insights into the origin and evolution of cryptochromes. Plant Cell Environ 33:1614–1626
Heinze S, Reppert SM (2012) Anatomical basis of sun compass navigation i: the general layout of the monarch butterfly brain. J Comp Neurol 520:1599–1628
Heldmaier G, Steinlechner S (1981) Seasonal control of energy requirements for thermoregulation in the Djungarian hamster (Phodopus sungorus), living in natural photoperiod. J Comp Physiol B 142:429–437
Heldmaier G, Werner D (2004) Environmental signal processing and adaptation, vol 110. Blackwell Synergy
Helfrich-Förster C (2005a) Neurobiology of the fruit fly’s circadian clock. Genes Brain Behav 4:65–76
Helfrich-Förster C, Stengl M, Homberg U (1998) Organization of the circadian system in insects. Chronobiol Int 15:567–594
Helfrich-Förster C, Winter C, Hofbauer A, Hall J, Stanewsky R (2001) The circadian clock of fruit flies is blind after elimination of all known photoreceptors. Neuron 30:249–261
Helfrich-Förster C, Edwards T, Yasuyama K, Schneuwly S, Meinertzhagen I, Hofbauer A (2002) The extraretinal eyelet of Drosophila: development, ultrastructure and putative circadian function. J Neurosci 22:9255–9266
Helfrich-Förster C (2004) The circadian clock in the brain: a structural and functional comparison between mammals and insects. J Comp Physiol A 190:601–613
Helfrich-Förster C (2005b) Organization of endogenous clocks in insects. Biochem Soc Trans 33:957–961
Helfrich-Förster C, Täuber M, Park JH, Mühlig-Versen M, Schneuwly S, Hofbauer A (2000) Ectopic expression of the neuropeptide pigment-dispersing factor alters behavioral rhythms in Drosophila melanogaster. J Neurosci 20:3339–3353
Helfrich-Förster C, Nitabach MN, Holmes TC (2011) Insect circadian clock outputs. Essays Biochem 49:87–101
Hendricks JC, Finn SM, Panckeri KA, Chavkin J, Williams JA, Sehgal A, Pack AI (2000) Rest in Drosophila is a sleep-like state. Neuron 25:129–138
Hennessey TL, Field CB (1991) Circadian rhythms in photosynthesis. Plant Physiol 96:831–836
Hermann C, Yoshii T, Dusik V, Helfrich-Förster C (2012) Neuropeptide F immunoreactive clock neurons modify evening locomotor activity and free-running period in Drosophila melanogaster. J Comp Neurol 520:970–987
Herrero E, Davis SJ (2012) Time for a nuclear meeting: protein trafficking and chromatin dynamics intersect in the plant circadian system. Mol Plant 5:554–565
Herrero E, Kolmos E, Bujdoso N, Yuan Y, Wang M, Berns MC, Uhlworm H, Coupland G, Saini R, Jaskolski M, Webb A, Gonçalves J, Davis SJ (2012) EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell 24:428–443
Hirsh J, Riemensperger T, Coulom H, Iché M, Coupar J, Birman S (2010) Roles of dopamine in circadian rhythmicity and extreme light sensitivity of circadian entrainment. Curr Biol 20:209–214
Hoffmann K (1981) The role of the pineal gland in the photoperiodic control of seasonal cycles in hamsters. In: Follett D, Follett B (eds) Biological clocks in seasonal reproductive cycles. Wright, Bristol, pp 237–250
Hogenesch JB, Ueda HR (2011) Understanding systems-level properties: timely stories from the study of clocks. Nat Rev Genet 12:407–416
Holland RA, Borissov I, Siemers BM (2010) A nocturnal mammal, the greater mouse-eared bat, calibrates a magnetic compass by the sun. Proc Natl Acad Sci U S A 107:6941–6945
Homberg U, Reischig T, Stengl M (2003) Neural organization of the circadian system of the cockroach Leucophaea maderae. Chronobiol Int 20(4):577–591
Homberg U, Heinze S, Pfeiffer K, Kinoshita M, el Jundi B (2011) Central neural coding of sky polarization in insects. Philos Trans R Soc Lond B Biol Sci 366:680–687
Honma KI, Honma S (2009) The SCN-independent clocks, methamphetamine and food restriction. Eur J Neurosci 30:1707–1717
Honma S, Hashimoto S, Nakao M, Kato Y, Honma K-I (2003) Period and phase adjustments of human circadian rhythms in the real world. J Biol Rhythms 18:261–270
Horowitz T, Cade B, Wolfe J, Czeisler C (2001) Efficacy of bright light and sleep/darkness scheduling in alleviating circadian maladaptation to night work. Am J Physiol 281:384–391
Hotta CT, Xu X, Xie Q, Dodd AN, Johnson CH, Webb AA (2008) Are there multiple circadian clocks in plants? Plant Signal Behav 3:342–344
Howland RH (2009a) An overview of seasonal affective disorder and its treatment options. Phys Sportsmed 37:104–115
Howland RH (2009b) Somatic therapies for seasonal affective disorder. J Psychosoc Nurs Ment Health Serv 47:17–20
Hubbard KE, Robertson FC, Dalchau N, Webb AAR (2009) Systems analyses of circadian networks. Mol Biosyst 5:1502–1511
Hughes S, Hankins MW, Foster RG, Peirson SN (2012) Melanopsin phototransduction: slowly emerging from the dark. Prog Brain Res 199:19–40
Humble MB (2010) Vitamin D, light and mental health. J Photochem Photobiol B 101:142–149
Hung H-C, Maurer C, Kay SA, Weber F (2007) Circadian transcription depends on limiting amounts of the transcription co-activator nejire/cbp. J Biol Chem 282:31349–31357
Hunt SM, Elvin M, Crosthwaite SK, Heintzen C (2007) The PAS/LOV protein VIVID controls temperature compensation of circadian clock phase and development in Neurospora crassa. Genes Dev 21:1964–1974
Hunt SM, Thompson S, Elvin M, Heintzen C (2010) VIVID interacts with the WHITE COLLAR complex and FREQUENCY-interacting RNA helicase to alter light and clock responses in Neurospora. Proc Natl Acad Sci U S A 107:16709–16714
Hurd MW, Ralph MR (1998) The significance of circadian organization for longevity in the golden hamster. J Biol Rhythms 13:430–436
Husain M (2005) The neural retina: three channels of light detection. Adv Clin Neurosci Rehabil 5:22–23
Hut RA, Beersma DGM (2011) Evolution of time-keeping mechanisms: early emergence and adaptation to photoperiod. Philos Trans R Soc Lond B Biol Sci 366:2141–2154
Idda ML, Bertolucci C, Vallone D, Gothilf Y, Sánchez-Vázquez FJ, Foulkes NS (2012) Circadian clocks: lessons from fish. Prog Brain Res 199:41–57
de la Iglesia HO, Cambras T, Schwartz WJ, Diez-Noguera A (2004) Forced desynchronization of dual circadian oscillators within the rat suprachiasmatic nucleus. Curr Biol 14:796–800
Ikegami K, Yoshimura T (2012) Circadian clocks and the measurement of daylength in seasonal reproduction. Mol Cell Endocrinol 349:76–81
Im SH, Taghert PH (2010) PDF receptor expression reveals direct interactions between circadian oscillators in Drosophila. J Comp Neurol 518:1925–1945
Imaizumi T (2010) Arabidopsis circadian clock and photoperiodism: time to think about location. Curr Opin Plant Biol 13:83–89
Indic P, Schwartz WJ, Paydarfar D (2008) Design principles for phase-splitting behaviour of coupled cellular oscillators: clues from hamsters with ‘split’ circadian rhythms. J R Soc Interface 5:873–883
Inouye C, Okamoto K, Ishiura M, Kondo T (1998) The action spectrum of phase shift by light signal in the circadian rhythm in cyanobacterium. Plant Cell Physiol 39(Suppl):S82
Ishiura M, Kutsuna S, Aoki S, Iwasaki H, Andersson C, Tanabe A, Golden S, Johnson C, Kondo T (1998) Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science 281:1519–1523
Isojima Y, Nakajima M, Ukai H, Fujishima H, Yamada RG, Masumoto KH, Kiuchi R, Ishida M, Ukai-Tadenuma M, Minami Y, Kito R, Nakao K, Kishimoto W, Yoo S-H, Shimomura K, Takao T, Takano A, Kojima T, Nagai K, Sakaki Y, Takahashi JS, Ueda HR (2009) CKiepsilon/delta-dependent phosphorylation is a temperature-insensitive, period-determining process in the mammalian circadian clock. Proc Natl Acad Sci U S A 106:15744–15749
Ito C, Goto SG, Shiga S, Tomioka K, Numata H (2008) Peripheral circadian clock for the cuticle deposition rhythm in Drosophila melanogaster. Proc Natl Acad Sci U S A 105:8446–8451
Iuvone PM, Tosini G, Pozdeyev N, Haque R, Klein DC, Chaurasia SS (2005) Circadian clocks, clock networks, arylalkylamine n-acetyltransferase, and melatonin in the retina. Prog Retin Eye Res 24:433–456
Ivanchenko M, Stanewsky R, Giebultowicz JM (2001) Circadian photoreception in Drosophila: functions of cryptochrome in peripheral and central clocks. J Biol Rhythms 16:205–215
Iwasaki H, Kondo T (2004) Circadian timing mechanism in the prokaryotic clock system of cyanobacteria. J Biol Rhythms 19:436–444
Iwasaki H, Williams S, Kitayama Y, Ishiura M, Golden S, Kondo T (2000) A kaiC-interacting sensory histidine kinase, SasA, necessary to sustain robust circadian oscillation in cyanobacteria. Cell 101:223–233
Jackson FR (2011) Glial cell modulation of circadian rhythms. Glia 59:1341–1350
Jacobshagen S, Kessler B, Rinehart CA (2008) At least four distinct circadian regulatory mechanisms are required for all phases of rhythms in mrna amount. J Biol Rhythms 23:511–524
Jacobson D, Powell A, Dettman J, Saenz G, Barton M, Hiltz M, Dvorachek W Jr, Glass N, Taylor J, Natvig D (2004) Neurospora in temperate forests of western North America. Mycologia 96:66–74
Jagota A, de la Iglesia H, Schwartz W (2000) Morning and evening circadian oscillations in the suprachiasmatic nucleus in vitro. Nat Neurosci 3:372–376
Jang S-W, Liu X, Pradoldej S, Tosini G, Chang Q, Iuvone PM, Ye K (2010) Nacetylserotonin activates TrkB receptor in a circadian rhythm. Proc Natl Acad Sci U S A 107:3876–3881
Jewett M, Rimmer D, Duffy J, Klerman E, Kronauer R, Czeisler C (1997) Human circadian pacemaker is sensitive to light throughout subjective day without evidence of transients. Am J Physiol 273:R1800–R1809
Jewett M, Borbely A, Czeisler C (1999a) Special issue: proceedings workshop on biomathematical models of circadian rhythmicity, sleep regulation and neurobehavioral function in humans, May 18–21 1999 Dedham, Mass. J Biol Rhythm 14
Jewett M, Kronauer R, Megan E (1999b) Interactive mathematical models of subjective alertness and cognitive throughput in humans. J Biol Rhythms 14:588–597
Jinhu G, Yi L (2010) Molecular mechanism of the Neurospora circadian oscillator. Protein Cell 1:331–341
Johnson CH (2005) Testing the adaptive value of circadian systems. Methods Enzymol 393:818–837
Johnson CH (2010) Circadian clocks and cell division: what’s the pacemaker? Cell Cycle 9:3864–3873
Johnson CH, Golden SS (1999) Circadian programs in Cyanobacteria: adaptiveness and mechanism. Annu Rev Microbiol 53:389–409
Johnson CH, Kondo T, Hastings JW (1991) Action spectrum for resetting the circadian phototaxis rhythm in the CW15 strain of Chlamydomonas: II. Illuminated cells. Plant Physiol 97:1122–1129
Johnson CH, Kondo T, Goto K (1992) Circadian rhythms in Chlamydomonas. In: Honma K, Honma S, Hiroshige T (eds) Circadian clocks from cell to human: proc Fourth Sapporo symp biol rhythms. Hokkaido Univ. Hokkaido University Press, Sapporo, pp 139–155
Johnson CH, Stewart PL, Egli M (2011) The cyanobacterial circadian system: from biophysics to bioevolution. Annu Rev Biophys 40:143–167
Johnsson A, Karlsson H, Engelmann W (1973) Phase shifts in the Kalanchoe petal rhythm, caused by light pulses of different duration. A theoretical and experimental study. J Chronobiol 1:147–156
Johnsson A, Engelmann W, Klemke W, Ekse AT (1979) Free-running human circadian rhythms in Svalbard. Z Naturforsch C 34C:470–473
Johnsson A, Solheim BGB, Iversen T-H (2009) Gravity amplifies and microgravity decreases circumnutations in Arabidopsis thaliana stems: results from a space experiment. New Phytol 182:621–629
Johnston J, Tournier B, Andersson H, Masson-Pevet M, Lincoln G, Hazlerigg D (2006) Multiple effects of melatonin on rhythmic clock gene expression in the mammalian Pars tuberalis. Endocrinology 147:959–965
Jones MA (2009) Entrainment of the Arabidopsis circadian clock. J Plant Biol 52:202–209
Jud C, Schmutz I, Hampp G, Oster H, Albrecht U (2005) A guideline for analyzing circadian wheel-running behavior in rodents under different lighting conditions. Biol Proced Online 7:101–116
Kai H, Arai T, Yasuda F (1999) Accomplishment of time-interval activation of esterase A4 by simple removal of pin fraction. Chronobiol Int 16:51–58
Kami C, Lorrain S, Hornitschek P, Fankhauser C (2010) Light-regulated plant growth and development. Curr Top Dev Biol 91:29–66
Kaneko M, Hamblen M, Hall J (2000) Involvement of the period gene in developmental time-memory: effect of the per short mutation on phase shifts induced by light pulses delivered to Drosophila larvae. J Biol Rhythms 15:13–30
Karakashian M, Schweiger H (1976) Circadian properties of the rhythmic system in individual nucleated and enucleated cells of Acetabularia mediterranea. Exp Cell Res 97:366–377
Karlsson H, Johnsson A (1972) A feedback model for biological rhythms. II. Comparisons with experimental results, especially on the petal rhythm of Kalanchoe. J Theor Biol 36:175–194
Kasof J (2009) Cultural variation in seasonal depression: cross-national differences in winter versus summer patterns of seasonal affective disorder. J Affect Disord 115:79–86
Kecklund G, Milia LD, Axelsson J, Lowden A, Akerstedt T (2012) 20th International symposium on shiftwork and working time: biological mechanisms, recovery, and risk management in the 24-h society. Chronobiol Int 29:531–536
Keeling PJ (2007) Ostreococcus tauri: seeing through the genes to the genome. Trends Genet 23:151–154
Kegel M, Dam H, Ali F, Bjerregaard P (2009) The prevalence of seasonal affective disorder (SAD) in Greenland is related to latitude. Nord J Psychiatry 63:331–335
Kempinger L, Dittmann R, Rieger D, Helfrich-Forster C (2009) The nocturnal activity of fruit flies exposed to artificial moonlight is partly caused by direct light effects on the activity level that bypass the endogenous clock. Chronobiol Int 26:151–166
Kessel L, Siganos G, Jørgensen T, Larsen M (2011) Sleep disturbances are related to decreased transmission of blue light to the retina caused by lens yellowing. Sleep 34:1215–1219
Khalsa S, Jewett M, Cajochen C, Czeisler C (2003) A phase response curve to single bright light pulses in human subjects. J Physiol 549:945–952
Kim W-Y, Fujiwara S, Suh S-S, Kim J, Kim Y, Han L, David K, Putterill J, Nam HG, Somers DE (2007) ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 449:356–360
Kim Y-I, Vinyard DJ, Ananyev GM, Dismukes GC, Golden SS (2012) Oxidized quinones signal onset of darkness directly to the cyanobacterial circadian oscillator. Proc Natl Acad Sci U S A 109:17765–17769
Kistenpfennig C, Hirsh J, Yoshii T, Helfrich-Förster C (2012) Phase-shifting the fruit fly clock without cryptochrome. J Biol Rhythms 27:117–125
Klarsfeld A, Rouyer F (1998) Effects of circadian mutations and LD periodicity on the life span of Drosophila melanogaster. J Biol Rhythms 13:471–478
Klarsfeld A, Malpel S, Michard-Vanhee C, Picot M, Chelot E, Rouyer F (2004) Novel features of cryptochrome-mediated photoreception in the brain circadian clock of Drosophila. J Neurosci 24:1468–1477
Klein D, Moore R, Reppert S (1991) Suprachiasmatic nucleus: the mind’s clock. Oxford University Press, New York
Kleitman L (1963) Sleep and wakefulness. University of Chicago Press, Chicago
Klemm E, Ninnemann H (1976) Detailed action spectrum for the delay shift in pupae emergence of Drosophila pseudoobscura. Photochem Photobiol 24:369–371
Klerman EB (2005) Clinical aspects of human circadian rhythms. J Biol Rhythms 20:375–386
Klerman EB, Dijk DJ, Kronauer RE, Czeisler CA (1996) Simulations of light effects on the human circadian pacemaker: implications for assessment of intrinsic period. Am J Physiol 270:R271–R282
Kohyama J (2011) Sleep health and asynchronization. Brain Dev 33:252–259
Kolar C, Fejes E, Adam E, Schaefer E, Kay S, Nagy F (1998) Transcription of Arabidopsis and wheat Cab genes in single tobacco transgenic seedlings exhibits independent rhythms in a developmentally regulated fashion. Plant J 13:563–569
Kolla BP, Auger RR (2011) Jet lag and shift work sleep disorders: how to help reset the internal clock. Cleve Clin J Med 78:675–684
Kolmos E, Herrero E, Bujdoso N, Millar AJ, Tóth R, Gyula P, Nagy F, Davis SJ (2011) A reduced-function allele reveals that EARLY FLOWERING3 repressive action on the circadian clock is modulated by phytochrome signals in Arabidopsis. Plant Cell 23:3230–3246
Kondo T, Johnson C, Hastings J (1991) Action spectrum for resetting the circadian phototaxis rhythm in the CW15 strain I: cells in darkness. Plant Physiol 95:197–205
Kondo T, Strayer C, Kulkarni R, Taylor W, Ishiura M, Golden S, Johnson C (1993) Circadian rhythms in prokaryotes: luciferase as a reporter of circadian gene expression in cyanobacteria. Proc Natl Acad Sci U S A 90:5672–5676
Kondo T, Tsinoremas N, Golden S, Johnson C, Kutsuna S, Ishiura M (1994) Circadian clock mutants of cyanobacteria. Science 266:1233–1236
Kondo T, Mori T, Lebedeva NV, Aoki S, Ishiura M, Golden SS (1997) Circadian rhythms in rapidly dividing cyanobacteria. Science 275:224–227
Konopka RJ, Pittendrigh C, Orr D (2007) Reciprocal behaviour associated with altered homeostasis and photosensitivity of Drosophila clock mutants. J Neurogenet 21:243–252
Kotov NV, Baker RE, Dawidov DA, Platov KV, Valeyev NV, Skorinkin AI, Maini PK (2007) A study of the temperature dependence of bienzyme systems and enzymatic chains. Comput Math Methods Method 8:93–112
Kozma-Bognár L, Káldi K (2008) Synchronization of the fungal and the plant circadian clock by light. Chembiochem 9:2565–2573
Kräuchi K, Cajochen C, Pache M, Flammer J, Wirz-Justice A (2006) Thermoregulatory effects of melatonin in relation to sleepiness. Chronobiol Int 23:475–484
Kronauer RE, Czeisler CA, Pilato SF, Moore-Ede MC, Weitzman ED (1982) Mathematical model of the human circadian system with two interacting oscillators. Am J Physiol 242:3–17
Kronauer RE, Forger DB, Jewett ME (1999) Quantifying human circadian pacemaker response to brief, extended, and repeated light stimuli over the phototopic range. J Biol Rhythms 14:500–515
Kronfeld-Schor N, Einat H (2012) Circadian rhythms and depression: human psychopathology and animal models. Neuropharmacology 62:101–114
Kumar S, Mohan A, Sharma V (2005) Circadian dysfunction reduces lifespan in Drosophila melanogaster. Chronobiol Int 22:641–653
Kunihiro A, Yamashino T, Mizuno T (2010) PHYTOCHROME-INTERACTING FACTORS PIF4 and PIF5 are implicated in the regulation of hypocotyl elongation in response to blue light in Arabidopsis thaliana. Biosci Biotechnol Biochem 74:2538–2541
Kurosawa G, Aihara K, Iwasa Y (2006) A model for the circadian rhythm of cyanobacteria that maintains oscillation without gene expression. Biophys J 91:2015
Kwon I, Choe HK, Son GH, Kim K (2011) Mammalian molecular clocks. Exp Neurobiol 20:18–28
Laakso M, Hätönen T, Stenberg D, Alila A, Smith S (1993) The human circadian response to light -strong and weak resetting. J Biol Rhythms 8:351–360
Lack LC, Wright HR (2007) Treating chronobiological components of chronic insomnia. Sleep Med 8(6):637–644
Lakin-Thomas PL (2006) Transcriptional feedback oscillators: maybe, maybe not. J Biol Rhythms 21:83–92
Lakin-Thomas P, Johnson H (1999) Commentary: molecular and cellular models of circadian systems. J Biol Rhythms 14:486–489
Lakin-Thomas P, Cote G, Brody S (1990) Circadian rhythms in Neurospora crassa: biochemistry and genetics. Crit Rev Microbiol 17:365–416
Lakin-Thomas PL, Bell-Pedersen D, Brody S (2011) The genetics of circadian rhythms in Neurospora. Adv Genet 74:55–103
Lam R, Levitan R (2000) Pathophysiology of seasonal affective disorder: a review. J Psychiatr Neurosci 25:469–480
Lamont EW, Harbour VL, Barry-Shaw J, Diaz LR, Robinson B, Stewart J, Amir S (2007) Restricted access to food, but not sucrose, saccharine, or salt, synchronizes the expression of Period2 protein in the limbic forebrain. Neuroscience 144:402–411
Larrondo LF, Loros JJ, Dunlap JC (2012) High-resolution spatiotemporal analysis of gene expression in real time: in vivo analysis of circadian rhythms in Neurospora crassa using a FREQUENCY-luciferase translational reporter. Fungal Genet Biol 49:681–683
Lavoie M-P, Lam RW, Bouchard G, Sasseville A, Charron M-C, Gagné A-M, Tremblay P, Filteau M-J, Hébert M (2009) Evidence of a biological effect of light therapy on the retina of patients with seasonal affective disorder. Biol Psychiatry 66:253–258
Layana C, Diambra L (2011) Time-course analysis of cyanobacterium transcriptome: detecting oscillatory genes. PLoS One 6:e26291
Lear BC, Zhang L, Allada R (2009) The neuropeptide PDF acts directly on evening pacemaker neurons to regulate multiple features of circadian behavior. PLoS Biol 7:e1000154
Leis JM, Siebeck U, Dixson DL (2011) How Nemo finds home: the neuroecology of dispersal and of population connectivity in larvae of marine fishes. Integr Comp Biol 51:826–843
Leloup JC, Goldbeter A (1998) A model for circadian rhythms in Drosophila incorporating the formation of a complex between the PER and TIM proteins. J Biol Rhythms 13:70–87
Leloup JC, Goldbeter A (1999) Chaos and biorhythmicity in a model for circadian oscillations of the PER and TIM proteins in Drosophila. J Theor Biol 198:445–459
Leloup JC, Goldbeter A (2001) A molecular explanation for the long-term suppression of circadian rhythms by a single light pulse. Am J Physiol 280:1206–1212
Leloup J-C, Goldbeter A (2008) Modeling the circadian clock: from molecular mechanism to physiological disorders. Bioessays 30:590–600
Lema G, Auerbach A (2006) Modes and models of GABA A receptor gating. J Physiol 572:183
Levi F, Schibler U (2007) Circadian rhythms: mechanisms and therapeutic implications. Annu Rev Pharmacol Toxicol 47:593–628
Levitan RD (2005) What is the optimal implementation of bright light therapy for seasonal affective disorder (SAD)? J Psychiatry Neurosci 30:72
Levitan RD (2007) The chronobiology and neurobiology of winter seasonal affective disorder. Dialogues Clin Neurosci 9:315–324
Lewis R (1999) Control system models for the circadian clock of the New Zealand Weta, Hemideina thoracica (Orthoptera: Stenopelmatidae). J Biol Rhythms 14:480–485
Lewis ZA, Correa A, Schwerdtfeger C, Link K, Xie X, Gomer R, Thomas T, Ebbole D, Bell-Pedersen D (2002) Overexpression of White Collar-1(WC-1) activates circadian clock-associated genes, but is not sufficient to induce most light-regulated gene expression in Neurospora crassa. Mol Microbiol 45:917–931
Lewy A, Sack R (1997) Exogenous melatonin’s phase-shifting effects on the endogenous melatonin profile in sighted humans: a brief review and critique of the literature. J Biol Rhythms 12:588–594
Lewy AJ (2007) Melatonin and human chronobiology. Cold Spring Harb Symp Quant Biol 72:623–636
Lewy AJ, Ahmed S, Sack RL (1996) Phase shifting the human circadian clock using melatonin. Behav Brain Res 73:131–134
Lewy AJ, Emens J, Jackman A, Yuhas K (2006a) Circadian uses of melatonin in humans. Chronobiol Int 23:403–412
Lewy AJ, Lefler BJ, Emens JS, Bauer VK (2006b) The circadian basis of winter depression. Proc Natl Acad Sci U S A 103:7414–7419
Lewy AJ, Rough JN, Songer JB, Mishra N, Yuhas K, Emens JS (2007) The phase shift hypothesis for the circadian component of winter depression. Dialogues Clin Neurosci 9:291–300
Lewy AJ, Emens JS, Songer JB, Sims N, Laurie AL, Fiala SC, Buti AL (2009) Winter depression: integrating mood, circadian rhythms, and the sleep/wake and light/dark cycles into a bio-psycho-social-environmental model. Sleep Med Clin 4:285–299
Li J-D, Hu W-P, Zhou Q-Y (2012) The circadian output signals from the suprachiasmatic nuclei. Prog Brain Res 199:119–127
Li S, Lakin-Thomas P (2010) Effects of prd circadian clock mutations on FRQ-less rhythms in Neurospora. J Biol Rhythms 25:71–80
Lim C, Lee J, Choi C, Kim J, Doh E, Choe J (2007) Functional role of creb-binding protein in the circadian clock system of Drosophila melanogaster. Mol Cell Biol 27:4876–4890
Lin R, Chou H, Huang T (1999) Priority of light/dark entrainment over temperature in setting the circadian rhythms of the prokaryote Synechococcus RF-1. Planta 209:202–206
Lincoln GA (2006) Melatonin entrainment of circannual rhythms. Chronobiol Int 23:301–306
Lincoln GA, Messager S, Andersson H, Hazlerigg D (2002) Temporal expression of seven clock genes in the suprachiasmatic nucleus and the pars tuberalis of the sheep: evidence for an internal coincidence timer. Proc Natl Acad Sci U S A 99:13890–13895
Lincoln GA, Andersson H, Loudon A (2003) Clock genes in calendar cells as the basis of annual timekeeping in mammals–a unifying hypothesis. J Endocrinol 179:1–13
Litt A, Kramer EM (2010) The ABC model and the diversification of floral organ identity. Semin Cell Dev Biol 21:129–137
Liu Y (2003) Molecular mechanisms of entrainment in the Neurospora circadian clock. J Biol Rhythms 18:195–205
Liu Y, Merrow M, Loros J, Dunlap J (1998) How temperature changes reset a circadian oscillator. Science 281:825–829
Loros JJ, Dunlap JC, Larrondo LF, Shi M, Belden WJ, Gooch VD, Chen C-H, Baker CL, Mehra A, Colot HV, Schwerdtfeger C, Lambreghts R, Collopy PD, Gamsby JJ, Hong CI (2007) Circadian output, input, and intracellular oscillators: insights into the circadian systems of single cells. Cold Spring Harb Symp Quant Biol 72:201–214
Lowrey PL, Takahashi JS (2011) Genetics of circadian rhythms in mammalian model organisms. Adv Genet 74:175–230
Loza-Correa M, Gomez-Valero L, Buchrieser C (2010) Circadian clock proteins in prokaryotes: hidden rhythms? Front Microbiol 1:130
Lu SX, Tobin EM (2011) Chromatin remodeling and the circadian clock: jumonji C-domain containing proteins. Plant Signal Behav 6:810–814
Lu SX, Knowles SM, Webb CJ, Celaya RB, Cha C, Siu JP, Tobin EM (2011) The Jumonji C domain-containing protein JMJ30 regulates period length in the Arabidopsis circadian clock. Plant Physiol 155:906–915
Lu SX, Webb CJ, Knowles SM, Kim SHJ, Wang Z, Tobin EM (2012) CCA1 and ELF3 interact in the control of hypocotyl length and flowering time in Arabidopsis. Plant Physiol 158:1079–1088
Lucas RJ, Lall GS, Allen AE, Brown TM (2012) How rod, cone, and melanopsin photoreceptors come together to enlighten the mammalian circadian clock. Prog Brain Res 199:1–18
Mackey SR (2007) Biological rhythms workshop IA: molecular basis of rhythms generation. Cold Spring Harb Symp Quant Biol 72:7–19
Mackey SR, Ditty JL, Zeidner G, Chen Y, Golden SS (2009) Mechanisms for entraining the cyanobacterial circadian clock system with the environment. In: Ditty JL, Mackey SR, Johnson C (eds) Bacterial circadian programs. Springer, Berlin, pp 141–156
Mackey SR, Golden SS, Ditty JL (2011) The itty-bitty time machine genetics of the cyanobacterial circadian clock. Adv Genet 74:13–53
Madsen H, Dam H, Hageman I (2012) Study protocol: a cross-sectional survey of seasonal affective disorder in Danish populations with and without severe visual impairments. BMJ Open 2:e001020
Magnusson A, Partonen T (2005) The diagnosis, symptomatology, and epidemiology of seasonal affective disorder. CNS Spectr 10:625–634; quiz 1–14
Mahoney MM (2010) Shift work, jet lag, and female reproduction. Int J Endocrinol 2010:813764
Malpel S, Klarsfeld A, Rouyer F (2002) Larval optic nerve and adult extra-retinal photoreceptors sequentially associate with clock neurons during Drosophila brain development. Development 129:1443–1453
Malpel S, Klarsfeld A, Rouyer F (2004) Circadian synchronization and rhythmicity in larval photoperception-defective mutants of Drosophila. J Biol Rhythms 19:10–21
Malzahn E, Ciprianidis S, Káldi K, Schafmeier T, Brunner M (2010) Photoadaptation in Neurospora by competitive interaction of activating and inhibitory LOV domains. Cell 142:762–772
Manichaikul A, Ghamsari L, Hom EFY, Lin C, Murray RR, Chang RL, Balaji S, Hao T, Shen Y, Chavali AK, Thiele I, Yang X, Fan C, Mello E, Hill DE, Vidal M, Salehi-Ashtiani K, Papin JA (2009) Metabolic network analysis integrated with transcript verification for sequenced genomes. Nat Methods 6:589–592
Marpegan L, Swanstrom AE, Chung K, Simon T, Haydon PG, Khan SK, Liu AC, Herzog ED, Beaulé C (2011) Circadian regulation of ATP release in astrocytes. J Neurosci 31:8342–8350
Martin SK, Eastman CI (1998) Medium-intensity light produces circadian rhythm adaptation to simulated night-shift work. Sleep 21:154–165
Matsuo T, Ishiura M (2011) Chlamydomonas reinhardtii as a new model system for studying the molecular basis of the circadian clock. FEBS Lett 585:1495–1502
May P, Christian J-O, Kempa S, Walther D (2009) Chlamycyc: an integrative systems biology database and web-portal for Chlamydomonas reinhardtii. BMC Genomics 10:209
McCarthy EV, Wu Y, Decarvalho T, Brandt C, Cao G, Nitabach MN (2011) Synchronized bilateral synaptic inputs to Drosophila melanogaster neuropeptidergic rest/arousal neurons. J Neurosci 31:8181–8193
McClung CA (2007) Circadian genes, rhythms and the biology of mood disorders. Pharmacol Ther 114:222–232
McClung CA (2011) Circadian rhythms: lost in post-translation. Curr Biol 21:R400–R402
McDermott JE, Oehmen CS, McCue LA, Hill E, Choi DM, Stöckel J, Liberton M, Pakrasi HB, Sherman LA (2011) A model of cyclic transcriptomic behavior in the cyanobacterium Cyanothece sp. ATCC 51142. Mol Biosyst 7:2407–2418
McWatters HG, Devlin PF (2011) Timing in plants–a rhythmic arrangement. FEBS Lett 585:1474–1484
Meesters Y, Dekker V, Schlangen LJM, Bos EH, Ruiter MJ (2011) Low-intensity blue-enriched white light (750 lux) and standard bright light (10,000 lux) are equally effective in treating SAD. A randomized controlled study. BMC Psychiatry 11:17
Mehra A, Baker CL, Loros JJ, Dunlap JC (2009) Post-translational modifications in circadian rhythms. Trends Biochem Sci 34:483–490
Meijer J, Watanabe K, Detari L, deVries M, Albus H, Treep J, Schaap J, Rietveld W (1996) Light entrainment of the mammalian biological clock. Prog Brain Res 111:175–190
Meijer JH, Schwartz WJ (2003) In search of the pathways for light-induced pacemaker resetting in the suprachiasmatic nucleus. J Biol Rhythms 18:235–249
Menaker M, Moreira L, Tosini G (1997) Evolution of circadian organization in vertebrates. Braz J Med Biol Res 30:305–313
Mendoza J, Pévet P, Challet E (2009) Entrainment and coupling of the hamster suprachiasmatic clock by daily dark pulses. J Neurosci Res 87:758–765
Merlin C, Heinze S, Reppert SM (2011) Unraveling navigational strategies in migratory insects. Curr Opin Neurobiol 22(2):353–361
Merrow M, Roenneberg T (2007) Circadian entrainment of Neurospora crassa. Cold Spring Harb Symp Quant Biol 72:279–285
Merrow M, Brunner M, Roenneberg T (1999) Assignment of circadian function for the Neurospora clock gene frequency. Nature 399:584–586
Mersch P, Middendorp H, Bouhuys A, Beersma D, van den Hoofdakker R (1999) Seasonal affective disorder and latitude: a review of the literature. J Affect Disord 53:35–48
Meyer P, Saez L, Young M (2006) PER-TIM interactions in living Drosophila cells: an interval timer for the circadian clock. Science 311:226–229
Michael TP, Salome PA, Yu HJ, Spencer TR, Sharp EL, McPeek MA, Alonso JM, Exker JR, McClung CR (2003) Enhanced fitness conferred by naturally occurring variations in the circadian clock. Science 302:1049–1053
Michaels SD (2009) Flowering time regulation produces much fruit. Curr Opin Plant Biol 12:75–80
Michalak EE, Murray G, Wilkinson C, Dowrick C, Lam RW (2007) A pilot study of adherence with light treatment for seasonal affective disorder. Psychiatry Res 149:315–320
Mihalcescu I, Hsing W, Leibler S (2004) Resilient circadian oscillator revealed in individual cyanobacteria. Nature 430:81–85
Miles L, Raynal D, Wilson M (1977) Blind man living in normal society has circadian rhythm of 24.9 hours. Science 198:421–423
Millar AJ (1999) Tansley review no. 103 – biological clocks in Arabidopsis thaliana. New Phytol 141:175–197
Millar AJ, Carre IA, Strayer CA, Chua N-H, Kay AS (1995) Circadian clock mutants in Arabidopsis identified by luciferase imaging. Science 267:1161–1163
Mills JN (1964) Circadian rhythms during and after three months in solitude underground. J Physiol 174:217–231
Min H, Johnson CH, Golden SS (2004) Phase determination of circadian gene expression in Synechococcus elongatus PCC 7942. J Biol Rhythms 19:103–112
Minors DS, Waterhouse J (1981) Circadian rhythms and the human. Wright, Bristol
Minors D, Waterhouse J, Wirz-Justice A (1991) A human phase response curve to light. Neurosci Lett 133:36–40
Mistlberger RE, Skene DJ (2005) Nonphotic entrainment in humans? J Biol Rhythms 20:339–352
Mitsui A, Kumazawa S, Takahashi A, Ikemoto H, Cao S, Arai T (1986) Strategy by which nitrogen-fixing unicellular cyanobacteria grow photoautotrophically. Nature 323:720–722
Mittag M (2001) Circadian rhythms in microalgae. Int Rev Cytol 206:213–247
Mittag M, Wagner V (2003) The circadian clock of the unicellular eukaryotic model organism Chlamydomonas reinhardtii. Biol Chem 384:689–695
Mittag M, Kiaulehn S, Johnson CH (2005) The circadian clock in Chlamydomonas reinhardtii. What is it for? What is it similar to? Plant Physiol 137:399–409
Miyoshi F, Nakayama Y, Kaizu K, Iwasaki H, Tomita M (2007) A mathematical model for the kai-protein-based chemical oscillator and clock gene expression rhythms in cyanobacteria. J Biol Rhythms 22:69
Mohawk JA, Takahashi JS (2011) Cell autonomy and synchrony of suprachiasmatic nucleus circadian oscillators. Trends Neurosci 34:349–358
Mohawk JA, Green CB, Takahashi JS (2012) Central and peripheral circadian clocks in mammals. Annu Rev Neurosci 35:445–462
Monk TH (2000) What can the chronobiologist do to help the shift worker? J Biol Rhythms 15:86–94
Monnier A, Liverani S, Bouvet R, Jesson B, Smith JQ, Mosser J, Corellou F, Bouget F-Y (2010) Orchestrated transcription of biological processes in the marine picoeukaryote Ostreococcus exposed to light/dark cycles. BMC Genomics 11:192
de Montaigu A, Tóth R, Coupland G (2010) Plant development goes like clockwork. Trends Genet 26:296–306
Montgomery P, Dunne D (2007) Sleep disorders in children. Clin Evid (Online) 2007:pii
Moore R, Speh J, Card J (1995) The rhd originates from a distinct subset of retinal ganglion cells. J Comp Neurol 352:351–366
Morant P-E, Thommen Q, Pfeuty B, Vandermoere C, Corellou F, Bouget F-Y, Lefranc M (2010) A robust two-gene oscillator at the core of Ostreococcus tauri circadian clock. Chaos 20:045108
Morgan L, Greene A, Bell-Pedersen D (2003) Circadian and light-induced expression of luciferase in Neurospora crassa. Fungal Genet Biol 38:327–332
Morgan PJ, Hazlerigg DG (2008) Photoperiodic signalling through the melatonin receptor turns full circle. J Neuroendocrinol 20:820–826
Mori T, Binder B, Johnson C (1996) Circadian gating of cell division in Cyanobacteria growing with average doubling times of less than 24 hours. Proc Natl Acad Sci U S A 93:10183–10188
Mori T, Saveliev S, Xu Y, Stafford W, Cox M, Inman R, Johnson C (2002) Circadian clock protein KaiC forms ATP-dependent hexameric rings and binds DNA. Proc Natl Acad Sci U S A 99:17203–17208
Moulager M, Monnier A, Jesson B, Bouvet R, Mosser J, Schwartz C, Garnier L, Corellou F, Bouget F-Y (2007) Light-dependent regulation of cell division in Ostreococcus: evidence for a major transcriptional input. Plant Physiol 144:1360–1369
Moulager M, Corellou F, Vergé V, Escande M-L, Bouget F-Y (2010) Integration of light signals by the retinoblastoma pathway in the control of s phase entry in the picophytoplanktonic cell Ostreococcus. PLoS Genet 6:e1000957
Muheim R (2011) Behavioural and physiological mechanisms of polarized light sensitivity in birds. Philos Trans R Soc Lond B Biol Sci 366:763–771
Mullineaux CW, Stanewsky R (2009) The rolex and the hourglass: a simplified circadian clock in Prochlorococcus? J Bacteriol 191:5333–5335
Murakami R, Miyake A, Iwase R, Hayashi F, Uzumaki T, Ishiura M (2008) ATPase activity and its temperature compensation of the cyanobacterial clock protein KaiC. Genes Cells 13:387–395
Murayama Y, Mukaiyama A, Imai K, Onoue Y, Tsunoda A, Nohara A, Ishida T, Maéda Y, Terauchi K, Kondo T, Akiyama S (2011) Tracking and visualizing the circadian ticking of the cyanobacterial clock protein kaic in solution. EMBO J 30:68–78
Mutoh R, Nishimura A, Yasui S, Onai K, Ishiura M (2013) The ATP-mediated regulation of KaiB-KaiC interaction in the cyanobacterial circadian clock. PLoS One 8:e80200
Müller B, Grossniklaus U (2010) Model organisms–a historical perspective. J Proteomics 73:2054–2063
Münch M, Scheuermaier KD, Zhang R, Dunne SP, Guzik AM, Silva EJ, Ronda JM, Duffy JF (2011) Effects on subjective and objective alertness and sleep in response to evening light exposure in older subjects. Behav Brain Res 224:272–278
Naef F (2005) Circadian clocks go in vitro: purely post-translational oscillators in cyanobacteria. Mol Syst Biol 1:2005.0019
Naidoo N, Song W, Hunter-Ensor M, Seghal A (1999) A role for the proteasome in the light response of the timeless clock protein. Science 285:1737–1741
Nakahira Y, Katayama M, Miyashita H, Kutsuna S, Iwasaki H, Oyama T, Kondo T (2004) Global gene repression by KaiC as a master process of prokaryotic circadian system. Proc Natl Acad Sci U S A 101:881–885
Nakajima M, Imai K, Ito H, Nishiwaki T, Murayama Y, Iwasaki H, Oyama T, Kondo T (2005) Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro. Science 308:414–415
Nakamichi N (2011) Molecular mechanisms underlying the Arabidopsis circadian clock. Plant Cell Physiol 52:1709–1718
Nakamichi N, Kiba T, Kamioka M, Suzuki T, Yamashino T, Higashiyama T, Sakakibara H, Mizuno T (2012) Transcriptional repressor prr5 directly regulates clock-output pathways. Proc Natl Acad Sci U S A 109:17123–17128
Ng FS, Tangredi MM, Jackson FR (2011) Glial cells physiologically modulate clock neurons and circadian behavior in a calcium-dependent manner. Curr Biol 21:625–634
Nickla D, Wildsoet C, Wallman J (1998) Visual influences on diurnal rhythms in ocular length and choroidal thickness in chick eyes. Exp Eye Res 66:163–181
Nikaido SS, Johnson CH (2000) Daily and circadian variation in survival from ultraviolet radiation in Chlamydomonas reinhardtii. Photochem Photobiol 71:758–765
Nishiwaki T, Iwasaki H, Ishiura M, Kondo T (2000) Nucleotide binding and autophosphorylation of the clock protein KaiC as a circadian timing process of cyanobacteria. Proc Natl Acad Sci U S A 97:495–499
Nitabach MN, Sheeba V, Vera DA, Blau J, Holmes TC (2005) Membrane electrical excitability is necessary for the free-running larval Drosophila circadian clock. J Neurobiol 62:1–13
Niwa Y, Yamashino T, Mizuno T (2009) The circadian clock regulates the photoperiodic response of hypocotyl elongation through a coincidence mechanism in Arabidopsis thaliana. Plant Cell Physiol 50:838–854
Ohdo S, Koyanagi S, Matsunaga N, Hamdan A (2011) Molecular basis of chronopharmaceutics. J Pharm Sci 100:3560–3576
Oishi T, Yamao M, Kondo C, Haida Y, Masuda A, Tamotsu S (2001) Multiphotoreceptor and multioscillator system in avian circadian organization. Microsc Res Tech 53:43–47
Okamura H (2007) Suprachiasmatic nucleus clock time in the mammalian circadian system. Cold Spring Harb Symp Quant Biol 72:551–556
Okawa M, Uchiyama M (2007) Circadian rhythm sleep disorders: characteristics and entrainment pathology in delayed sleep phase and non-24-h sleep-wake syndrome. Sleep Med Rev 11:485–496
Oltmanns O (1960) Über den Einfluss der Temperatur auf die endogene Tagesrhythmik und die Blühinduktion bei der Kurztagpflanze Kalanchoe blossfeldiana. Planta 54:233–264
Onai K, Morishita M, Itoh S, Okamoto K, Ishiura M (2004) Circadian rhythms in the thermophilic cyanobacterium Thermosynechococcus elongatus: compensation of period length over a wide temperature range. J Bacteriol 186:4972–4977
O’Neill JS, Reddy AB (2011) Circadian clocks in human red blood cells. Nature 469:498–503
O’Neill JS, van Ooijen G, Dixon LE, Troein C, Corellou F, Bouget F-Y, Reddy AB, Millar AJ (2011) Circadian rhythms persist without transcription in a eukaryote. Nature 469:554–558
van Ooijen G, Dixon LE, Troein C, Millar AJ (2011) Proteasome function is required for biological timing throughout the twenty-four hour cycle. Curr Biol 21:869–875
Oster H, Maronde E, Albrecht U (2002) The circadian clock as a molecular calendar. Chronobiol Int 19:507–516
Ouyang Y, Andersson C, Kondo T, Golden S, Johnson C (1998) Resonating circadian clocks enhance fitness in Cyanobacteria. Proc Natl Acad Sci U S A 95:8660–8664
Pail G, Huf W, Pjrek E, Winkler D, Willeit M, Praschak-Rieder N, Kasper S (2011) Bright-light therapy in the treatment of mood disorders. Neuropsychobiology 64:152–162
Palczewski K (2012) Chemistry and biology of vision. J Biol Chem 287:1612–1619
Panda S, Hogenesch JB (2004) It’s all in the timing: many clocks, many outputs. J Biol Rhythms 19:374–387
Pandi-Perumal SR, Srinivasan V, Maestroni GJM, Cardinali DP, Poeggeler B, Hardeland R (2006) Melatonin: nature’s most versatile biological signal? FEBS J 273:2813–2838
Pandi-Perumal SR, Bahammam AS, Brown GM, Spence DW, Bharti VK, Kaur C, Hardeland R, Cardinali DP (2013) Melatonin antioxidative defense: therapeutical implications for aging and neurodegenerative processes. Neurotox Res 23:267–300
Pandit A, Maheshwari R (1994) Sexual reproduction by Neurospora in nature. Fungal Genet Newsl 41:67–68
Parisky KM, Agosto J, Pulver SR, Shang Y, Kuklin E, Hodge JJL, Kang K, Kang K, Liu X, Garrity PA, Rosbash M, Griffith LC (2008) PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 60:672–682
Park N, Cheon S, Son GH, Cho S, Kim K (2012) Chronic circadian disturbance by a shortened light-dark cycle increases mortality. Neurobiol Aging 33:1122.e11–1122.e22
Parker G, Brotchie H (2011) ‘D’ for depression: any role for vitamin D? ‘Food for Thought’ ii. Acta Psychiatr Scand 124:243–249
Partonen T, Magnusson A (2001) Seasonal affective disorder: practice and research. Oxford University Press, Oxford
de Paula RM, Lewis ZA, Greene AV, Seo K, Morgan L, Vitalini M, Bennett L, Gomer R, Bell-Pedersen D (2006) Two circadian timing circuits in Neurospora crassa cells share components and regulate distinct rhythmic processes. J Biol Rhythms 21:159–168
de Paula RM, Vitalini MW, Gomer RH, Bell-Pedersen D (2007) Complexity of the Neurospora crassa circadian clock system: multiple loops and oscillators. Cold Spring Harb Symp Quant Biol 72:345–351
Peirson SN, Halford S, Foster RG (2009) The evolution of irradiance detection: melanopsin and the non-visual opsins. Philos Trans R Soc Lond B Biol Sci 364:2849–2865
Pendergast JS, Yamazaki S (2011) Masking responses to light in period mutant mice. Chronobiol Int 28:657–663
Peschel N, Helfrich-Förster C (2011) Setting the clock–by nature: circadian rhythm in the fruitfly Drosophila melanogaster. FEBS Lett 585:1435–1442
Peterson E (1981a) Dynamic response of a circadian pacemaker. I. Recovery from extended light exposure. Biol Cybern 40:171–179
Peterson E (1981b) Dynamic response of a circadian pacemaker. II. Recovery from light pulse perturbations. Biol Cybern 40:181–194
Pévet P, Challet E (2011) Melatonin: both master clock output and internal time-giver in the circadian clocks network. J Physiol Paris 105:170–182
Pévet P, Agez L, Bothorel B, Saboureau M, Gauer F, Laurent V, Masson-Pévet M (2006) Melatonin in the multi-oscillatory mammalian circadian world. Chronobiol Int 23:39–51
Pfeuty B, Thommen Q, Corellou F, Djouani-Tahri EB, Bouget F-Y, Lefranc M (2012) Circadian clocks in changing weather and seasons: lessons from the picoalga Ostreococcus tauri. Bioessays 34:781–790
Philip P, Akerstedt T (2006) Transport and industrial safety, how are they affected by sleepiness and sleep restriction? Sleep Med Rev 10:347–356
Pickard GE, Sollars PJ (2010) Intrinsically photosensitive retinal ganglion cells. Sci China Life Sci 53:58–67
Pittendrigh CS (1964) The entrainment of circadian oscillations by skeleton photoperiods. Science 144:565
Pittendrigh C, Daan S (1976) A functional analysis of circadian pacemakers in nocturnal rodents. J Comp Physiol A106:333–355
Pittendrigh CS, Bruce BG, Rosensweig NS, Rubin ML (1959) Growth patterns in Neurospora. Nature 184:169–170
Plachetzki D, Serb J, Oakley T (2005) New insights into the evolutionary history of photoreceptor cells. Trends Ecol Evol 20:465–467
Plautz J, Kaneko M, Hall J, Kay S (1997) Independent photoreceptive circadian clocks throughout Drosophila. Science 278:1632–1635
Pokhilko A, Fernández AP, Edwards KD, Southern MM, Halliday KJ, Millar AJ (2012) The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops. Mol Syst Biol 8:574
Portolés S, Más P (2010) The functional interplay between protein kinase CK2 and CCA1 transcriptional activity is essential for clock temperature compensation in Arabidopsis. PLoS Genet 6:e1001201
Praschak-Rieder N, Willeit M (2012) Imaging of seasonal affective disorder and seasonality effects on serotonin and dopamine function in the human brain. Curr Top Behav Neurosci 11:149–167
Prasko J (2008) Bright light therapy. Neuro Endocrinol Lett 29(Suppl 1):33–64
Price-Lloyd N, Elvin M, Heintzen C (2005) Synchronizing the Neurospora crassa circadian clock with the rhythmic environment. Biochem Soc Trans 33:949–952
Privitera MR, Moynihan J, Tang W, Khan A (2010) Light therapy for seasonal affective disorder in a clinical office setting. J Psychiatr Pract 16:387–393
Provencio I, Foster RG (1995) Circadian rhythms in mice can be regulated by photoreceptors with cone-like characteristics. Brain Res 694:183–190
Pulivarthy SR, Tanaka N, Welsh DK, Haro LD, Verma IM, Panda S (2007) Reciprocity between phase shifts and amplitude changes in the mammalian circadian clock. Proc Natl Acad Sci U S A 104:20356–20361
Qin X, Byrne M, Xu Y, Mori T, Johnson CH (2010) Coupling of a core posttranslational pacemaker to a slave transcription/translation feedback loop in a circadian system. PLoS Biol 8:e1000394
Quintero J, Kuhlman S, McMahon D (2003) The biological clock nucleus: a multiphasic oscillator network regulated by light. J Neurosci 23:8070–8076
Rahman SA, Kayumov L, Tchmoutina EA, Shapiro CM (2009) Clinical efficacy of dim light melatonin onset testing in diagnosing delayed sleep phase syndrome. Sleep Med 10:549–555
Rastad C, Ulfberg J, Lindberg P (2008) Light room therapy effective in mild forms of seasonal affective disorder–a randomised controlled study. J Affect Disord 108:291–296
Rastad C, Ulfberg J, Lindberg P (2011) Improvement in fatigue, sleepiness, and health-related quality of life with bright light treatment in persons with seasonal affective disorder and subsyndromal SAD. Depress Res Treat 2011:543906
Rea M (1998) Photic entrainment of circadian rhythms in rodents. Chronobiol Int 15:395–423
Reed VA (2011) Shift work, light at night, and the risk of breast cancer. AAOHN J 59:37–45; quiz 46
Reid KJ, McGee-Koch LL, Zee PC (2011) Cognition in circadian rhythm sleep disorders. Prog Brain Res 190:3–20
Reiter RJ, Tan DX, Cabrera J, D’Arpa D, Sainz R, Mayo J, Ramos S (1999) The oxidant/antioxidant network: role of melatonin. Biol Signals Recept 8:56–63
Reiter RJ, Tan DX, Herman TS, Thomas CR (2004) Melatonin as a radioprotective agent: a review. Int J Radiat Oncol Biol Phys 59:639–653
Reiter RJ, Tan DX, Fuentes-Broto L (2010) Melatonin: a multitasking molecule. Prog Brain Res 181:127–151
Reme C, Wirz-Justice A, Terman M (1991) The visual input stage of the mammalian circadian pacemaking system: I. Is there a clock in the mammalian eye? J Biol Rhythms 6:5–29
Reppert SM, Gegear RJ, Merlin C (2010) Navigational mechanisms of migrating monarch butterflies. Trends Neurosci 33:399–406
Revel FG, Masson-Pévet M, Pévet P, Mikkelsen JD, Simonneaux V (2009) Melatonin controls seasonal breeding by a network of hypothalamic targets. Neuroendocrinology 90:1–14
Revell VL, Eastman CI (2005) How to trick mother nature into letting you fly around or stay up all night. J Biol Rhythms 20:353–365
Rieger D, Stanewsky R, Helfrich-Forster C (2003) Cryptochrome, compound eyes, Hofbauer-Buchner eyelets, and ocelli play different roles in the entrainment and masking pathway of the locomotor activity rhythm in the fruit fly Drosophila melanogaster. J Biol Rhythms 18:377–391
Rieger D, Shafer OT, Tomioka K, Helfrich-Forster C (2006) Functional analysis of circadian pacemaker neurons in Drosophila melanogaster. J Neurosci 26:2531–2543
Rieger D, Wülbeck C, Rouyer F, Helfrich-Förster C (2009) Period gene expression in four neurons is sufficient for rhythmic activity of Drosophila melanogaster under dim light conditions. J Biol Rhythms 24:271–282
Rietveld WJ, Minors DS, Waterhouse JM (1993) Circadian rhythms and masking: an overview. Chronobiol Int 10:306–312
Ripperger JA, Merrow M (2011) Perfect timing: epigenetic regulation of the circadian clock. FEBS Lett 585:1406–1411
Ripperger JA, Jud C, Albrecht U (2011) The daily rhythm of mice. FEBS Lett 585:1384–1392
Rizzini L, Favory J-J, Cloix C, Faggionato D, O’Hara A, Kaiserli E, Baumeister R, Schäfer E, Nagy F, Jenkins GI, Ulm R (2011) Perception of UV-B by the Arabidopsis uvr8 protein. Science 332:103–106
Roberts DE, Killiany RJ, Rosene DL (2012) Neuron numbers in the hypothalamus of the normal aging Rhesus monkey: stability across the adult lifespan and between the sexes. J Comp Neurol 520:1181–1197
Roecklein KA, Rohan KJ, Duncan WC, Rollag MD, Rosenthal NE, Lipsky RH, Provencio I (2009) A missense variant (P10L) of the melanopsin (OPN4) gene in seasonal affective disorder. J Affect Disord 114:279–285
Roenneberg T, Aschoff J (1990) Annual rhythm of human reproduction II: environmental correlations. J Biol Rhythms 5:217–240
Roenneberg T, Foster R (1997) Twilight times: light and the circadian system. Photochem Photobiol 66:549–561
Roenneberg T, Merrow M (1998) Molecular circadian oscillators: an alternative hypothesis. J Biol Rhythms 13:167–179
Roenneberg T, Merrow M (2001) Seasonality and photoperiodism in fungi. J Biol Rhythms 16:403–414
Roenneberg T, Mittag M (1996) The circadian program of algae. Semin Cell Dev Biol 7:753–763
Roenneberg T, Chua EJ, Bernardo R, Mendoza E (2008) Modelling biological rhythms. Curr Biol 18:R826–R835
Roenneberg T, Allebrandt KV, Merrow M, Vetter C (2012) Social jetlag and obesity. Curr Biol 22:939–943
Rohan KJ, Roecklein KA, Lacy TJ, Vacek PM (2009) Winter depression recurrence one year after cognitive-behavioral therapy, light therapy, or combination treatment. Behav Ther 40:225–238
Romero JM, Valverde F (2009) Evolutionarily conserved photoperiod mechanisms in plants: when did plant photoperiodic signaling appear? Plant Signal Behav 4:642–644
Rosato E, Kyriacou CP (2011) The role of natural selection in circadian behaviour: a molecular-genetic approach. Essays Biochem 49:71–85
Rosbash M (2009) The implications of multiple circadian clock origins. PLoS Biol 7:e62
Rosenthal NE (2006) Winter blues. Guilford Press, New York
Rosenthal NE, Sack DA, Gillin JC, Lewy AJ, Goodwin FK, Davenport Y, Mueller PS, Newsome DA, Wehr TA (1984) Seasonal affective disorder. A description of the syndrome and preliminary findings with light therapy. Arch Gen Psychiatry 41:72–80
Roth T (2012) Shift work disorder: overview and diagnosis. J Clin Psychiatry 73:e09
Ruan GX, Zhang DQ, Zhou T, Yamazaki S, McMahon DG (2006) Circadian organization of the mammalian retina. Proc Natl Acad Sci U S A 20:9703–9708
Ruby N, Dark J, Burns D, Heller H, Zucker I (2002) The suprachiasmatic nucleus is essential for circadian body temperature rhythms in hibernating ground squirrels. J Neurosci 22:357–364
Rüger M, Gordijn MCM, Beersma DGM, de Vries B, Daan S (2003) Acute and phase-shifting effects of ocular and extraocular light in human circadian physiology. J Biol Rhythms 18:409–419
Ruoff P, Rensing L (1996) The temperature-compensated goodwin model simulates many circadian clock properties. J Theor Biol 179:275–285
Ruoff P, Rensing L (2004) Temperature effects on circadian clocks. J Theor Biol 29:445–456
Rusak B, Zucker I (1979) Neural regulation of circadian rhythms. Physiol Rev 59:449–526
Rust MJ, Markson JS, Lane WS, Fisher DS, O’Shea EK (2007) Ordered phosphorylation governs oscillation of a three-protein circadian clock. Science 318:809–812
Rust MJ, Golden SS, O’Shea EK (2011) Light-driven changes in energy metabolism directly entrain the cyanobacterial circadian oscillator. Science 331:220–223
Rémi J, Merrow M, Roenneberg T (2010) A circadian surface of entrainment: varying T, t, and photoperiod in Neurospora crassa. J Biol Rhythms 25:318–328
Saarela S, Reiter R (1994) Function of melatonin in thermoregulatory processes. Life Sci 54:295–311
Sadeghniiat-Haghighi K, Yazdi Z, Jahanihashemi H, Aminian O (2011) The effect of bright light on sleepiness among rapid-rotating 12-hour shift workers. Scand J Work Environ Health 37:77–79
Saksvik IB, Bjorvatn B, Hetland H, Sandal GM, Pallesen S (2011) Individual differences in tolerance to shift work–a systematic review. Sleep Med Rev 15:221–235
Salgado-Delgado R, Angeles-Castellanos M, Buijs MR, Escobar C (2008) Internal desynchronization in a model of night-work by forced activity in rats. Neuroscience 154:922–931
Salgado-Delgado R, Nadia S, Angeles-Castellanos M, Buijs RM, Escobar C (2010) In a rat model of night work, activity during the normal resting phase produces desynchrony in the hypothalamus. J Biol Rhythms 25:421–431
Salichos L, Rokas A (2010) The diversity and evolution of circadian clock proteins in fungi. Mycologia 102:269–278
Salomé PA, Weigel D, McClung CR (2010) The role of the Arabidopsis morning loop components CCA1, LHY, PRR7, and PRR9 in temperature compensation. Plant Cell 22:3650–3661
Salva MAQ, Hartley S, Barbot F, Alvarez JC, Lofaso F, Guilleminault C (2011) Circadian rhythms, melatonin and depression. Curr Pharm Des 17:1459–1470
Satoh Y, Kawai H, Kudo N, Kawashima Y, Mitsumoto A (2006) Time-restricted feeding entrains daily rhythms of energy metabolism in mice. Am J Physiol Regul Integr Comp Physiol 290:R1276–R1283
Sawa M, Nusinow DA, Kay SA, Imaizumi T (2007) FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 318:261–265
Schaap J, Albus H, VanderLeest H, Eilers P, Detari L, Meijer J (2003) Heterogeneity of rhythmic suprachiasmatic nucleus neurons: implications for circadian waveform and photoperiodic encoding. Proc Natl Acad Sci U S A 100:15994–15999
Schafmeier T, Diernfellner ACR (2011) Light input and processing in the circadian clock of Neurospora. FEBS Lett 585:1467–1473
Scheer FAJL, Wright KP, Kronauer RE, Czeisler CA (2007) Plasticity of the intrinsic period of the human circadian timing system. PLoS One 2:e721
Schmidt TM, Chen S-K, Hattar S (2011) Intrinsically photosensitive retinal ganglion cells: many subtypes, diverse functions. Trends Neurosci 34:572–580
Schmidt-Koenig K (1975) Migration and homing in animals. Springer, Berlin/Heidelberg
Schmoll C, Lascaratos G, Dhillon B, Skene D, Riha RL (2011) The role of retinal regulation of sleep in health and disease. Sleep Med Rev 15:107–113
Schroder S, Herzog ED, Kiss IZ (2012) Transcription-based oscillator model for light-induced splitting as antiphase circadian gene expression in the suprachiasmatic nuclei. J Biol Rhythms 27:79–90
Schulz P, Steimer T (2009) Neurobiology of circadian systems. CNS Drugs 23(Suppl 2):3–13
Schulze T, Prager K, Dathe H, Kelm J, Kiessling P, Mittag M (2010) How the green alga Chlamydomonas reinhardtii keeps time. Protoplasma 244:3–14
Schwerdtfeger C (2003) Vivid is a flavoprotein and serves as a fungal blue light photoreceptor for photoadaptation. EMBO J 22:4846–4855
Schäuble S, Heiland I, Voytsekh O, Mittag M, Schuster S (2011) Predicting the physiological role of circadian metabolic regulation in the green alga Chlamydomonas reinhardtii. PLoS One 6:e23026
Sehgal A, Price J, Young MW (1992) Ontogeny of a biological clock in Drosophila melanogaster. Proc Natl Acad Sci U S A 89:1423–1427
Sellix MT, Menaker M (2011) Circadian clocks in mammalian reproductive physiology: effects of the “other” biological clock on fertility. Discov Med 11:273–281
Serrano G, Herrera-Palau R, Romero JM, Serrano A, Coupland G, Valverde F (2009) Chlamydomonas CONSTANS and the evolution of plant photoperiodic signaling. Curr Biol 19:359–368
Shang Y, Griffith LC, Rosbash M (2008) Light-arousal and circadian photoreception circuits intersect at the large PDF cells of the Drosophila brain. Proc Natl Acad Sci U S A 105:19587–19594
Sharkey KM, Carskadon MA, Figueiro MG, Zhu Y, Rea MS (2011) Effects of an advanced sleep schedule and morning short wavelength light exposure on circadian phase in young adults with late sleep schedules. Sleep Med 12:685–692
Sharma V (2003) Adaptive significance of circadian clocks. Chronobiol Int 20:901–919
Shaw PJ, Cirelli C, Greenspan RJ, Tononi G (2000) Correlates of sleep and waking in Drosophila melanogaster. Science 287:1834–1837
Sheeba V, Fogle KJ, Kaneko M, Rashid S, Chou Y-T, Sharma VK, Holmes TC (2008a) Large ventral lateral neurons modulate arousal and sleep in Drosophila. Curr Biol 18:1537–1545
Sheeba V, Gu H, Sharma VK, O’Dowd DK, Holmes TC (2008b) Circadian- and light-dependent regulation of resting membrane potential and spontaneous action potential firing of Drosophila circadian pacemaker neurons. J Neurophysiol 99:976–988
Sheeba V, Fogle KJ, Holmes TC (2010) Persistence of morning anticipation behavior and high amplitude morning startle response following functional loss of small ventral lateral neurons in Drosophila. PLoS One 5:e11628
Shimomura K, Lowrey PL, Vitaterna MH, Buhr ED, Kumar V, Hanna P, Omura C, Izumo M, Low SS, Barrett RK, LaRue SI, Green CB, Takahashi JS (2010) Genetic suppression of the circadian clock mutation by the melatonin biosynthesis pathway. Proc Natl Acad Sci U S A 107:8399–8403
Shirani A, Louis EKS (2009) Illuminating rationale and uses for light therapy. J Clin Sleep Med 5:155–163
Siffre M (1975) Six months alone in a cave. Natl Geogr Mag 147:426
Silver R, Schwartz W (2005) The suprachiasmatic nucleus is a functionally heterogeneous timekeeping organ. Methods Enzymol 393:451–465
Silver R, Balsam PD, Butler MP, LeSauter J (2011) Food anticipation depends on oscillators and memories in both body and brain. Physiol Behav 104:562–571
Simonneaux V, Ribelayga C (2003) Generation of the melatonin endocrine message in mammals: a review of the complex regulation of melatonin synthesis by norepinephrine, peptides, and other pineal transmitters. Pharmacol Rev 55:325–395
Simons MJP (2009) The evolution of the cyanobacterial posttranslational clock from a primitive “phoscillator”. J Biol Rhythms 24:175–182
Skene D (2003) Optimization of light and melatonin to phase-shift human circadian rhythms. J Neuroendocrinol 15:438–441
Skene DJ, Arendt J (2007) Circadian rhythm sleep disorders in the blind and their treatment with melatonin. Sleep Med 8:651–655
Smith RM, Williams SB (2006) Circadian rhythms in gene transcription imparted by chromosome compaction in the cyanobacterium Synechococcus elongatus. Proc Natl Acad Sci U S A 103:8564–8569
Smith KM, Sancar G, Dekhang R, Sullivan CM, Li S, Tag AG, Sancar C, Bredeweg EL, Priest HD, McCormick RF, Thomas TL, Carrington JC, Stajich JE, Bell-Pedersen D, Brunner M, Freitag M (2010) Transcription factors in light and circadian clock signaling networks revealed by genomewide mapping of direct targets for Neurospora white collar complex. Eukaryot Cell 9:1549–1556
Sole MJD, Sande PH, Bernades JM, Aba MA, Rosenstein RE (2007) Circadian rhythm of intraocular pressure in cats. Vet Ophthalmol 10:155–161
Springer M (1993) Genetic control of fungal differentiation: the three sporulation pathways of Neurospora crassa. Bioessays 15:365–374
Srikanth A, Schmid M (2011) Regulation of flowering time: all roads lead to Rome. Cell Mol Life Sci 68:2013–2037
Srinivasan V, Smits M, Spence W, Lowe AD, Kayumov L, Pandi-Perumal SR, Parry B, Cardinali DP (2006) Melatonin in mood disorders. World J Biol Psychiatry 7:138–151
Srinivasan V, Spence DW, Pandi-Perumal SR, Trakht I, Cardinali DP (2008) Jet lag: therapeutic use of melatonin and possible application of melatonin analogs. Travel Med Infect Dis 6:17–28
Stal LJ, Krumbein M (1985a) Nitrogenase activity in the non-heterocystous cyanobacterium Oscillatoria sp. grown under alternating light-dark cycles. Arch Microbiol 143:67–71
Stal LJ, Krumbein M (1985b) Oxygen protection of nitrogenase in the aerobically nitrogen fixing non-heterocystous cyanobacterium Oscillatoria sp. Arch Microbiol 143:72–76
Stanewsky R, Jamison C, Plautz J, Kay S, Hall J (1997) Multiple circadian-regulated elements contribute to cycling period gene expression in Drosophila. EMBO J 16:5006–5018
Stanewsky R, Kaneko M, Emery P, Beretta B, Wager-Smith K, Kay SA, Rosbash M, Hall JC (1998) The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila. Cell 95:681–692
Steenhard BM, Besharse JC (2000) Phase shifting the retinal circadian clock: xPer2 mRNA induction by light and dopamine. J Neurosci 20:8572–8577
Stehle JH, von Gall C, Schomerus C, Korf H (2001) Of rodents and ungulates and melatonin: creating a uniform code for darkness by different signaling mechanisms. J Biol Rhythms 16:312–325
Stehle JH, Saade A, Rawashdeh O, Ackermann K, Jilg A, Sebestény T, Maronde E (2011) A survey of molecular details in the human pineal gland in the light of phylogeny, structure, function and chronobiological diseases. J Pineal Res 51:17–43
Steiner M, Werstiuk E, Seggie J (1987) Dysregulation of neuroendocrine crossroads: depression, circadian rhythms and the retina–a hypothesis. Prog Neuropsychopharmacol Biol Psychiatry 11:267–278
Steinlechner S, Stieglitz A, Ruf T (2002) Djungarian hamsters: a species with a labile circadian pacemaker? Arrhythmicity under a light-dark cycle induced by short light pulses. J Biol Rhythms 17:248–258
Stephan FK (2002) The “other” circadian system: food as a Zeitgeber. J Biol Rhythms 17:284–292
Stevens RG (2009a) Light-at-night, circadian disruption and breast cancer: assessment of existing evidence. Int J Epidemiol 38:963–970
Stevens RG (2009b) Working against our endogenous circadian clock: breast cancer and electric lighting in the modern world. Mutat Res 680:106–108
Stoleru D, Peng Y, Agosto J, Rosbash M (2004) Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature 431:862–868
Strasser B, Sánchez-Lamas M, Yanovsky MJ, Casal JJ, Cerdán PD (2010) Arabidopsis thaliana life without phytochromes. Proc Natl Acad Sci U S A 107:4776–4781
Strauss O (2005) The retinal pigment epithelium in visual function. Physiol Rev 85:845–881
Strong RE, Marchant BK, Reimherr FW, Williams E, Soni P, Mestas R (2009) Narrow-band blue-light treatment of seasonal affective disorder in adults and the influence of additional nonseasonal symptoms. Depress Anxiety 26:273–278
Su S-B, Lu C-W, Kao Y-Y, Guo H-R (2008) Effects of 12-hour rotating shifts on menstrual cycles of photoelectronic workers in Taiwan. Chronobiol Int 25:237–248
Suzuki L, Johnson CH (2002) Photoperiodic control of germination in the unicell Chlamydomonas. Naturwissenschaften 89:214–220
Szular J, Sehadova H, Gentile C, Szabo G, Chou W-H, Britt SG, Stanewsky R (2012) Rhodopsin 5- and rhodopsin 6-mediated clock synchronization in drosophila melanogaster is independent of retinal phospholipase c-? signaling. J Biol Rhythms 27:25–36
Tan Y, Merrow M, Roenneberg T (2004) Photoperiodism in Neurospora crassa. J Biol Rhythms 19:135–143
Tanaka K, Takahashi M, Tanaka M, Takanao T, Nishinoue N, Kaku A, Kato N, Tagaya H, Miyaoka H (2011) Brief morning exposure to bright light improves subjective symptoms and performance in nurses with rapidly rotating shifts. J Occup Health 53:258–266
Taniguchi Y, Takai N, Katayama M, Kondo T, Oyama T (2010) Three major output pathways from the kaiabc-based oscillator cooperate to generate robust circadian kaibc expression in cyanobacteria. Proc Natl Acad Sci U S A 107:3263–3268
Tassi P, Pellerin N, Moessinger M, Hoeft A, Muzet A (2000) Visual resolution in humans fluctuates over the 24h period. Chronobiol Int 17:187–195
Tauber E, Last K, Olive P, Kyriacou C (2004) Clock gene evolution and functional divergence. J Biol Rhythms 19:445–458
Tavakoli-Nezhad M, Schwartz WJ (2005) c-Fos expression in the brains of behaviorally ‘split’ hamsters in constant light: calling attention to a dorsolateral region of the suprachiasmatic nucleus and the medial division of the lateral habenula. J Biol Rhythms 20:419–429
Teng C, Akerman D, Cordas T, Kasper S, Vieira A (1995) Seasonal affective disorder in a tropical country: a case report. Psychiatry Res 56:11–15
Terman M (2007) Evolving applications of light therapy. Sleep Med Rev 11:497–507
Terman J, Terman M (1999) Photopic and scotopic light detection in patients with seasonal affective disorder and control subjects. Biol Psychiatry 46:1642–1648
Terman M, Amira L, Terman J, Ross D (1996) Predictors of response and nonresponse to light treatment for winter depression. Am J Psychiatry 153:1423–1429
Thaler K, Delivuk M, Chapman A, Gaynes BN, Kaminski A, Gartlehner G (2011) Second-generation antidepressants for seasonal affective disorder. Cochrane Database Syst Rev 12:CD008591
Thommen Q, Pfeuty B, Morant P-E, Corellou F, Bouget F-Y, Lefranc M (2010) Robustness of circadian clocks to daylight fluctuations: hints from the picoeucaryote Ostreococcus tauri. PLoS Comput Biol 6:e1000990
Thommen Q, Pfeuty B, Corellou F, Bouget F-Y, Lefranc M (2012) Robust and flexible response of the Ostreococcus tauri circadian clock to light/dark cycles of varying photoperiod. FEBS J 279:3432–3448
Thompson CL, Rickman CB, Shaw SJ, Ebright JN, Kelly U, Sancar A, Rickman DW (2003) Expression of the blue-light receptor cryptochrome in the human retina. Invest Ophthalmol Vis Sci 44:4515–4521
Thorpy MJ (2011) Understanding and diagnosing shift work disorder. Postgrad Med 123:96–105
Tigges M, Marquez-Lago TT, Stelling J, Fussenegger M (2009) A tunable synthetic mammalian oscillator. Nature 457:309–312
Timonen M, Nissilä J, Liettu A, Jokelainen J, Jurvelin H, Aunio A, Räsänen P, Takala T (2012) Can transcranial brain-targeted bright light treatment via ear canals be effective in relieving symptoms in seasonal affective disorder? A pilot study. Med Hypotheses 78:511–515
Toh KI, Jones R, He Y, Eide EJ, Hinz WA, Virshup DM, Ptácek LJ, Fu YH (2001) An hper2 phosphorylation site mutation in familial advanced sleep phase syndrome. Science 291:1040–1043
Tomioka K, Matsumoto A (2010) A comparative view of insect circadian clock systems. Cell Mol Life Sci 67:1397–1406
Tomioka K, Uryu O, Kamae Y, Umezaki Y, Yoshii T (2012) Peripheral circadian rhythms and their regulatory mechanism in insects and some other arthropods: a review. J Comp Physiol B 182(6):729–740
Tonsfeldt KJ, Chappell PE (2012) Clocks on top: the role of the circadian clock in the hypothalamic and pituitary regulation of endocrine physiology. Mol Cell Endocrinol 349:3–12
Tosches MA, Bucher D, Vopalensky P, Arendt D (2014) Melatonin signaling controls circadian swimming behavior in marine zooplankton. Cell 159:46–57
Tosini G, Fukuhara C (2002) The mammalian retina as a clock. Cell Tissue Res 309:119–126
Tosini G, Menaker M (1996) Circadian rhythms in cultured mammalian retina. Science 272:419–421
Tosini G, Pozdeyev N, Sakamoto K, Iuvone PM (2008) The circadian clock system in the mammalian retina. Bioessays 30:624–633
Toth R, Kevei E, Hall A, Millar AJ, Nagy F, Kozma-Bognar L (2001) Circadian clock-regulated expression of phytochrome and cryptochrome genes in Arabidopsis. Plant Physiol 127:1607–1616
Touitou Y (1998) Biological clocks: mechanisms and applications, Proc Internat Congr Chronobiol Paris 7–11 September 1997. Elsevier, Amsterdam
Touitou Y, Bogdan A (2007) Promoting adjustment of the sleep-wake cycle by chronobiotics. Physiol Behav 90:294–300
Troein C, Corellou F, Dixon LE, van Ooijen G, O’Neill JS, Bouget F-Y, Millar AJ (2011) Multiple light inputs to a simple clock circuit allow complex biological rhythms. Plant J 66:375–385
Troncoso-Ponce MA, Mas P (2012) Newly described components and regulatory mechanisms of circadian clock function in Arabidopsis thaliana. Mol Plant 5:545–553
Tseng Y-Y, Hunt SM, Heintzen C, Crosthwaite SK, Schwartz J-M (2012) Comprehensive modelling of the Neurospora circadian clock and its temperature compensation. PLoS Comput Biol 8:e1002437
Turek FW (2005) Role of light in circadian entrainment and treating sleep disorders–and more. Sleep 28:548–549
Tyson JJ, Albert R, Goldbeter A, Ruoff P, Sible J (2008) Biological switches and clocks. J R Soc Interface 5(Suppl 1):S1–S8
Ugolini A, Boddi V, Mercatelli L, Castellini C (2005) Moon orientation in adult and young sandhoppers under artificial light. Proc Biol Sci 272:2189–2194
Ugolini A, Somigli S, Pasquali V, Renzi P (2007) Locomotor activity rhythm and sun compass orientation in the sandhopper Talitrus saltator are related. J Comp Physiol A 193:1259–1263
Ukai H, Ueda HR (2010) Systems biology of mammalian circadian clocks. Annu Rev Physiol 72:579–603
Ukai H, Kobayashi TJ, Nagano M, Masumoto KH, Sujino M, Kondo T, Yagita K, Shigeyoshi Y, Ueda HR (2007) Melanopsin-dependent photo-perturbation reveals desynchronization underlying the singularity of mammalian circadian clocks. Nat Cell Biol 9:1327–1334
Ukai-Tadenuma M, Kasukawa T, Ueda HR (2008) Proof-by-synthesis of the transcriptional logic of mammalian circadian clocks. Nat Cell Biol 10:1154–1163
Vandewalle G, Hébert M, Beaulieu C, Richard L, Daneault V, Garon M-L, Leblanc J, Grandjean D, Maquet P, Schwartz S, Dumont M, Doyon J, Carrier J (2011) Abnormal hypothalamic response to light in seasonal affective disorder. Biol Psychiatry 70:954–961
Veleri S, Rieger D, Helfrich-Förster C, Stanewsky R (2007) Hofbauer-Buchner eyelet affects circadian photosensitivity and coordinates TIM and PER expression in Drosophila clock neurons. J Biol Rhythms 22:29–42
Vijayan V, Zuzow R, O’Shea EK (2009) Oscillations in supercoiling drive circadian gene expression in cyanobacteria. Proc Natl Acad Sci U S A 106:22,564–22,568
Vitalini MW, de Paula RM, Park WD, Bell-Pedersen D (2006) The rhythms of life: circadian output pathways in Neurospora. J Biol Rhythms 21:432–444
Vitalini MW, de Paula RM, Goldsmith CS, Jones CA, Borkovich KA, Bell- Pedersen D (2007) Circadian rhythmicity mediated by temporal regulation of the activity of p38 MAPK. Proc Natl Acad Sci U S A 104:18223–18228
Vitalini MW, Dunlap J, Heintzen C, Liu Y, Loros J, Bell-Pedersen D (2010) Circadian rhythms. In: Borkovich KA, Ebbole DJ (eds) Cellular and molecular biology of filamentous fungi. ASM Press, Washington, DC, pp 442–466
Vollrath L (2002) Chronoendokrinologia- quo vadis? Ann Anat 184:583–593
Wagner V, Mittag M (2009) Probing circadian rhythms in Chlamydomonas rheinhardtii by functional proteomics. Methods Mol Biol 479:173–188
Wagner V, Gessner G, Mittag M (2005) Functional proteomics: a promising approach to find novel components of the circadian system. Chronobiol Int 22:403–415
Walton JC, Weil ZM, Nelson RJ (2011) Influence of photoperiod on hormones, behavior, and immune function. Front Neuroendocrinol 32:303–319
Wang W, Barnaby JY, Tada Y, Li H, Tör M, Caldelari D, Lee DU, Fu X-D, Dong X (2011) Timing of plant immune responses by a central circadian regulator. Nature 470:110–114
Warman GR, Pawley MDM, Bolton C, Cheeseman JF, Fernando AT, Arendt J, Wirz-Justice A (2011) Circadian-related sleep disorders and sleep medication use in the New Zealand blind population: an observational prevalence survey. PLoS One 6:e22073
Warren E, Allen C, Brown R, Robinson D (2003) Intrinsic light responses of retinal ganglion cells projecting to the circadian system. Eur J Neurosci 17:1727–1735
Watanabe T, Kojima M, Tomida S, Nakamura TJ, Yamamura T, Nakao N, Yasuo S, Yoshimura T, Ebihara S (2006) Peripheral clock gene expression in CS mice with bimodal locomotor rhythms. Neurosci Res 54:295–301
Watanabe T, Naito E, Nakao N, Tei H, Yoshimura T, Ebihara S (2007) Bimodal clock gene expression in mouse suprachiasmatic nucleus and peripheral tissues under a 7-hour light and 5-hour dark schedule. J Biol Rhythms 22:58–68
Weber F, Zorn D, Rademacher C, Hung H-C (2011) Post-translational timing mechanisms of the Drosophila circadian clock. FEBS Lett 585:1443–1449
Wegener C, Hamasaka Y, Nässel DR (2004) Acetylcholine increases intracellular Ca2+ via nicotinic receptors in cultured pdf-containing clock neurons of drosophila. J Neurophysiol 91:912–923
Wehr TA (2001) Photoperiodism in humans and other primates: evidence and implications. J Biol Rhythms 16:348–364
Weitzman ED, Czeisler CA, Zimmerman JC, Moore-Ede MC (1981) Biological rhythms in man: relationship of sleep-wake, cortisol, growth hormone, and temperature during temporal isolation. Adv Biochem Psychopharmacol 28:475–499
Welsh DK, Takahashi JS, Kay SA (2010) Suprachiasmatic nucleus: cell autonomy and network properties. Annu Rev Physiol 72:551–577
Werner R (2002) Chlamydomonas reinhardtii as a unicellular model for circadian rhythm analysis. Chronobiol Int 19:325–343
Westrin A, Lam RW (2007a) Long-term and preventative treatment for seasonal affective disorder. CNS Drugs 21:901–909
Westrin A, Lam RW (2007b) Seasonal affective disorder: a clinical update. Ann Clin Psychiatry 19:239–246
Wever RA (1979) The circadian system of man. Springer, New York
Willeit M, Sitte HH, Thierry N, Michalek K, Praschak-Rieder N, Zill P, Winkler D, Brannath W, Fischer MB, Bondy B, Kasper S, Singer EA (2008) Enhanced serotonin transporter function during depression in seasonal affective disorder. Neuropsychopharmacology 33:1503–1513
Winfree A (1970) Integrated view of resetting a circadian clock. J Comp Physiol A 28:327–374
Winfree AT (1986) The timing of biological clocks. Scientific American Books, Inc., New York
Winkler D, Pjrek E, Iwaki R, Kasper S (2006) Treatment of seasonal affective disorder. Expert Rev Neurother 6:1039–1048
Wirz-Justice A, Terman M (2012) Chronotherapeutics (light and wake therapy) as a class of interventions for affective disorders. Handb Clin Neurol 106:697–713
Woelfle M, Johnson C (2006) No promoter left behind: global circadian gene expression in cyanobacteria. J Biol Rhythms 21:419
Woelfle MA, Ouyang Y, Phanvijhitsiri K, Johnson CH (2004) The adaptive value of circadian clocks: an experimental assessment in Cyanobacteria. Curr Biol 14:1481–1486
Wolfson AR, Carskadon MA (2003) Understanding adolescents’ sleep patterns and school performance: a critical appraisal. Sleep Med Rev 7:491–506
Workman JL, Nelson RJ (2011) Potential animal models of seasonal affective disorder. Neurosci Biobehav Rev 35:669–679
Wright KP, Hughes RJ, kronauer RE, Dijk DJ, Czeisler CA (2001) Intrinsic near-24-hour pacemaker period determines limits of circadian entrainment to a weak synchronizer in humans. Proc Natl Acad Sci U S A 98:14027–14032
Wülbeck C, Grieshaber E, Helfrich-Förster C (2008) Pigment-dispersing factor (pdf) has different effects on Drosophila’s circadian clocks in the accessory medulla and in the dorsal brain. J Biol Rhythms 23:409–424
Xu Y, Padiath QS, Shapiro RE, Jones CR, Wu SC, Saigoh N, Saigoh K, Ptacek LJ, Fu YH (2005) Functional consequences of a CKIdelta mutation causing familial advanced sleep phase syndrome. Nature 434:640–644
Xu K, DiAngelo JR, Hughes ME, Hogenesch JB, Sehgal A (2011) The circadian clock interacts with metabolic physiology to influence reproductive fitness. Cell Metab 13:639–654
Yakir E, Hilman D, Harir Y, Green RM (2007) Regulation of output from the plant circadian clock. FEBS J 274:335–345
Yamada Y, Forger D (2010) Multiscale complexity in the mammalian circadian clock. Curr Opin Genet Dev 20:626–633
Yamaguchi S, Isejima H, Matsuo T, Okura R, Yagita K, Kobayashi M, Okamura H (2003) Synchronization of cellular clocks in the suprachiasmatic nucleus. Science 302:1408–1412
Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, Ueda M, Block GD, Sakaki Y, Menaker M, Tei H (2000) Resetting central and peripheral circadian oscillators in transgenic rats. Science 288:682–685
Yang XP, de Groot EJ (1992) Identification of two clock proteins in Acetabularia cliftonii and construction of cdna libraries from Acetabularia cliftonii and Acetabularia mediterranea. Int J Biochem 24:1141–1150
Yanovsky MJ, Mazzella MA, Whitelam GC, Casal JJ (2001) Resetting of the circadian clock by phytochromes and cryptochromes in Arabidopsis. J Biol Rhythms 16:523–530
Yerushalmi S, Yakir E, Green RM (2011) Circadian clocks and adaptation in Arabidopsis. Mol Ecol 20:1155–1165
Yoshii T, Funada Y, Ibuki-Ishibashi T, Matsumoto A, Tanimura T, Tomioka K (2004) Drosophila cryb mutation reveals two circadian clocks that drive locomotor rhythm and have different responsiveness to light. J Insect Physiol 50:479–488
Yoshii T, Todo T, Wülbeck C, Stanewsky R, Helfrich-Förster C (2008) Cryptochrome is present in the compound eyes and a subset of Drosophila’s clock neurons. J Comp Neurol 508:952–966
Yoshii T, Wülbeck C, Sehadova H, Veleri S, Bichler D, Stanewsky R, Helfrich-Förster C (2009) The neuropeptide pigment-dispersing factor adjusts period and phase of Drosophila’s clock. J Neurosci 29:2597–2610
Yoshii T, Hermann C, Helfrich-Förster C (2010) Cryptochrome-positive and – negative clock neurons in Drosophila entrain differentially to light and temperature. J Biol Rhythms 25:387–398
Yoshii T, Rieger D, Helfrich-Förster C (2012) Two clocks in the brain: an update of the morning and evening oscillator model in Drosophila. Prog Brain Res 199:59–82
Young R (1976) Visual cells and the concept of renewal. Invest Ophthalmol Vis Sci 15:700–725
Zamanian Z, Kakooei H, Ayattollahi SMT, Dehghani M (2010) Effect of bright light on shift work nurses in hospitals. Pak J Biol Sci 13:431–436
van Zanten M, Tessadori F, Peeters AJM, Fransz P (2012) Shedding light on large scale chromatin reorganization in Arabidopsis thaliana. Mol Plant 5:583–590
Zee PC, Goldstein CA (2010) Treatment of shift work disorder and jet lag. Curr Treat Options Neurol 12:396–411
Zee PC, Vitiello MV (2009) Circadian rhythm sleep disorder: irregular sleep wake rhythm type. Sleep Med Clin 4:213–218
Zhdanova I, Reebs S (2006) Circadian rhythms in fish. In: Sloman K, Wilson R, Balshine S (eds) Behavior and physiology of fish, vol 24, Fish physiology. Elsevier, Amsterdam, pp 197–238
Zinser ER, Lindell D, Johnson ZI, Futschik ME, Steglich C, Coleman ML, Wright MA, Rector T, Steen R, McNulty N, Thompson LR, Chisholm SW (2009) Choreography of the transcriptome, photophysiology, and cell cycle of a minimal photoautotroph, Prochlorococcus. PLoS One 4:e5135
Ziv C, Yarden O (2010) Gene silencing for functional analysis: assessing RNAi as a tool for manipulation of gene expression. Methods Mol Biol 638:77–100
Zoltowski BD, Schwerdtfeger C, Widom J, Loros JJ, Bilwes AM, Dunlap JC, Crane BR (2007) Conformational switching in the fungal light sensor Vivid. Science 316:1054–1057
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media New York
About this chapter
Cite this chapter
Johnsson, A., Helfrich-Förster, C., Engelmann, W. (2015). How Light Resets Circadian Clocks. In: Björn, L. (eds) Photobiology. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-1468-5_18
Download citation
DOI: https://doi.org/10.1007/978-1-4939-1468-5_18
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-1467-8
Online ISBN: 978-1-4939-1468-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)