
Abstract
Metabolism is defined as “1. The chemical processes occurring within a living cell or organism that are necessary for the maintenance of life. 2. The processing of a specific substance within the living body.” This definition simplifies a process that occurs at the cellular level in every living being and is the driving process of our existence. The consumption of energy is the basis of life; an innate and evolutionarily honed drive to maintain homeostasis and fulfill the needs for energy and cellular function. Derangements in metabolism are present with every disease process and may even be the cause. The quest to understand the exchange of energy at the cellular level and to develop novel techniques to manipulate, restore, or control this exchange is as old as medicine itself. The goal of this chapter is to review the history of our understanding of metabolic processes, to discuss normal cellular metabolism in a healthy subject, and to identify ways in which metabolism is altered in injury and illness.
Fasting is a great remedie of fever.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Acute Respiratory Distress Syndrome
- Critical Illness
- Lean Body Mass
- Total Energy Expenditure
- Respiratory Quotient
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
John Withals in “A shorte dictionarie most profitable for yong beginners the second tyme corrected, and augmented, with diuerse phrasys, and other thinges necessarie therevnto added: by Lewys Euans.”
Published 1574 by [In Paules churchyarde, at the signe of the Lucreece, by Thomas Purfoote] in Imprinted at London. Ref: www.openlibrary.org
Introduction
Metabolism is defined as “1. The chemical processes occurring within a living cell or organism that are necessary for the maintenance of life. 2. The processing of a specific substance within the living body” [1]. This definition simplifies a process that occurs at the cellular level in every living being and is the driving process of our existence. The consumption of energy is the basis of life; an innate and evolutionarily honed drive to maintain homeostasis and fulfill the needs for energy and cellular function.
Derangements in metabolism are present with every disease process and may even be the cause. The quest to understand the exchange of energy at the cellular level and to develop novel techniques to manipulate, restore, or control this exchange is as old as medicine itself. The goal of this chapter is to review the history of our understanding of metabolic processes, to discuss normal cellular metabolism in a healthy subject, and to identify ways in which metabolism is altered in injury and illness.
History of Metabolism
For centuries, man has struggled to understand the basis of life, often coming up with partially correct or wildly fanciful ways to explain the disease in front of them. Many of the basic explanations the ancients propagated about this concept of metabolism and disease came down to foods that cure or harm. Even today, our fascination with food as a way to manipulate the basis processes of life continues—a simple internet search produces countless reports of foods curing disease, reducing impotence, adding muscle, or losing weight. Yet, to end this never-ending search for the “magic” food source or the fountain of youth, the precisely orchestrated chemical and genetically determined metabolic processes at the cellular level must be fully elucidated. Only then can we find specific nutrients or foods that could “cure.”
The historical foundation of surgical metabolism over the centuries has constructed our current understanding. Dating back to the beginning of the Common Era, both the Ayurvedic and Greek “Aristotelian” philosophy of health and disease was based on the states of earth, water, fire and air or some combination of such, and was intimately tied to emotions/senses. This so-called humor physiology was pervasive for millenia [2]. Extensive catalogues of medicinal herbs and behaviors were employed to alter the balance between these humors to promote health and abate disease. Avicenna, a Persian philosopher and physician, applied the concept of humors across the spectrum of health and disease, treating excesses of one humor with removal or reduction of its counterpart. Galen is credited with the letting of blood to release the humors, a practice that lasted through the eighteenth century. In 1628, a new philosophy arose. William Harvey published Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus (An Anatomical Study of the Motion of the Heart and of the Blood in Animals), where he first described the cardiovascular system as a unit with the heart at the center pumping blood to the remainder of the body. This significant break with the long held practice of humoralism championed by the likes of Hippocrates and Galen was coupled with Harvey’s discovery of circulation and Santorio Sanctorius framework of metabolic balance. Sanctorius’ meticulous experiments over years illustrate the difference in weight of ones’ total intake compared to the reduced weight of what one excreted. This dissipation of quantity and mass he attributed to a theory called “insensible perspiration” [3]. These concepts were key in creating the understanding of physiologic conservation of energy and the body’s transformation of nutrients to functional energy through the bloodstream.
Crucial to the understanding of basic metabolism is the concept of oxygen consumption and cellular respiration. Joseph Priestly is credited with the discovery of oxygen gas. In his paper “Observations on Respiration and the Use of the Blood,” Priestley was the first to suggest a relationship between blood and air. Lavoisier, widely regarded as the “father of modern chemistry,” extrapolated on these findings of combustion and respiration in experiments done in collaboration with Pierre-Simon Laplace [4]. The pair not only devised an ice calorimeter to measure the amount of heat created during combustion or respiration, but calculated the carbon dioxide produced by the guinea pig and compared it to the generative process to create carbon dioxide. This characterized respiration as combustion, and this concept was later published in “On Heat.” The process of deranged metabolism in response to disease was also a concept that was delineated slowly over time. Louis Pasteur and Fredrich Henle first espoused the germ theory and Lister followed through on the notion of infectious organisms leading to suppuration and sepsis. This basic understanding of the pathogenesis of disease would later become crucial to the understanding of the metabolic response to illness. It would not be until the early twentieth century, however, before metabolism was brought first to the cellular level.
In the twentieth century, the concepts of metabolism and energy exchange in response to stress states were introduced. Sir David Cuthbertson published notable work on the metabolic and nutritional demands of critically ill patients, which still serves as the foundation for current nutritional therapy for the burn, septic, and other critically ill/injured patients [5]. Based on the findings of Lavoisier, indirect calorimetry is a commonly utilized method to estimate energy metabolism through respiratory gas exchange measurements. The carbon dioxide and nitrogen waste from individual’s consumption of oxygen is correlated back to the heat energy generation [6]. This technique is noninvasive, and can be applied to investigate numerous aspects of nutrition, thermogenesis, exercise, and the pathogenesis of metabolic diseases.
James Harris and Francis Benedict published A Biometric Study of Basal Metabolism in Man in 1919, which provided a method to estimate an individual’s basal metabolic rate and daily caloric requirements without quantitative calorimetric measurements. A regression equation was laboriously constructed. Even today, the Harris-Benedict equation allows healthcare professionals to better estimate and provide the metabolic needs of a patient in a practical and cost effective manner.
Though a significant tragedy, the 1942 Coconut Grove night club fire in Boston germinated a host of medical advances in the treatment of post-burn resuscitation, but more importantly illuminated the connection between fluid resuscitation and metabolic demands. The contributions of this body of literature have helped to recognize the stress response to injury, the importance of restoring hemodynamic stability to cellular metabolism, and began illustrating some of the sequela of resuscitation [7].
In the 1960s, Stanley Dudrick and Douglas Wilmore began investigations on the metabolic effects of intravenous nutrition supplementation and its effects on cellular metabolism, developing what they termed total parental nutrition (TPN). Initially, parenteral nutrition (PN) was designed to provide 100 % of an individual’s daily caloric needs, and was designed for those who suffered from short bowel syndrome [8]. The utilization of PN has expanded far past the treatment of short bowel syndrome, and it now has specific indications in any individual unable to tolerate enteral feeds. However, over time it was found that the metabolic derangements found particularly in the liver and gut were often detrimental to the patient, and the indications of PN usage have been narrowed appropriately. There were two major seminal papers in the early 1990s regarding PN. Kudsk’s work investigated the importance of the route of nutrition and its relationship with septic complications after trauma. His work demonstrated a lower incidence of septic morbidity in patients fed enterally as compared to with PN, with most of the significant complications occurring in the more severely injured patients [9]. The VA cooperative trial looked at the use of perioperative PN and its relationship with serious complications after major abdominal or thoracic surgical procedures in malnourished patients. That trial concluded that preoperative PN should be limited to only patients that are severely malnourished unless there are other specific indications for its’ need [10].
Since the 1970s, significant progress has been made toward the understanding of caloric deficit and protein malnutrition. Malnutrition and metabolic derangements from inadequate caloric provision were identified as common problems within hospitalized patients. The emphasis on caloric deficit gave way to an emphasis on protein energy malnutrition. In 1974, Bistrian and Blackburn published their groundbreaking work entitled Skeletons in the Closet. They identified a 50 % incidence of protein energy malnutrition in a modern Boston hospital. From that point forward, we began to better understand malnutrition as a reflection of metabolic derangements associated with stress [11]. Pierre Singer published a body of literature throughout the 1990s and early 2000s on protein catabolism and the turnover of lean body mass in stressed states and the impact this has on wound healing, infection rates, and mortality. Influencing metabolism by provision of goal directed nutritional therapy became the emphasis of many institutional protocols throughout the world.
Throughout the past decade, great strides have been made in the understanding of critical illness and the contributions that adequate, early, and goal directed nutrition can make toward the reduction of morbidity and mortality. In addition to adequate caloric provision, much was published on early goal directed therapy aimed to rapidly resuscitate patients with injury and illness in order to prevent and attenuate metabolic derangements. In 2001, the landmark paper from Rivers et al. entitled Early Goal Directed Therapy in the Treatment of Severe Sepsis and Septic Shock was published in the New England Journal of Medicine. Emphasis was placed on adequate fluid resuscitation, vasopressor support, maintenance of central venous pressure, and provision of stress dosed steroids in an attempt to support end organ perfusion and minimize the metabolic impact of critical illness [12]. In addition, recognition of the deleterious effects of stress induced hyperglycemia also was heightened. Through work such as Van den Berghe’s Intensive Insulin Therapy in Critically Ill Patients, published in the New England Journal in 2001, it was demonstrated that elevation of serum blood glucose levels, as well as profound hypoglycemia, were independent predictors of mortality and infectious complications [13]. The maintenance of serum glucose homeostasis helps to prevent this, demonstrating the importance of attenuating the metabolic response to stress.
The basic understanding of cellular metabolism and its effects on the patient is constantly evolving over time. It has far reaching clinical implications for the practitioner; the understanding of the stress response, of nutritional support, of the resuscitation of illness and injury and the manipulation of the gastrointestinal tract to improve disease outcomes all have their basis in cellular metabolism. As our understanding of the complex mechanisms behind these processes has broadened, we have bettered our ability to care for our patients.
Cellular Metabolism
Mitochondrial Metabolism
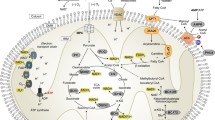
In order to maintain functional homeostasis in the body as a whole, metabolism must be regulated at the cellular level. Mitochondria are the main source of energy for eukaryotic cells. They divide by fission, which supports the theory that they originated as symbiotic bacteria. They are rod-shaped organelles approximately 0.2–0.5 μm in diameter, with an outer and inner membrane, each with unique characteristics. The outer membrane contains protein channels, or porins, which allow passage of molecules up to 10 kilodaltons (kDa). The inner membrane is composed almost entirely of proteins. Oxidative phosphorylation takes place here, which generates the electrochemical gradient necessary for proton transport and synthesis of adenosine triphosphate (ATP).
Critical metabolic pathways take place in the mitochondria. Oxidative phosphorylation, which generates the electrochemical gradient necessary for proton transport and synthesis of ATP, occurs within the mitochondrial membrane. This process is the culmination of the tricarboxylic acid cycle. This is a highly regulated process with numerous components, and it has been found that mitochondrial dysfunction plays a role as an early cellular event in critical illness. Targeted therapies during these times of metabolic stress have demonstrated improvement in mitochondrial function with regards to the electron transport system, oxidative phosphorylation, ATP production, and reduction of oxidative stress [14]. The clinical impact of these findings is only at the point of basic investigation.
Enzyme Function
Enzymes are the workhorse of metabolic pathways. They are necessary to catalyze the innumerable biochemical processes that are vital for the living cell and organism as a whole. Their proper function is dependent on multiple elements including temperature, pH, availability of substrates, and presence of cofactors. The physiologically stressed patient has derangements in virtually all of these variables that can lead to enzyme dysfunction, such as fever, acidosis or severe malnutrition. Additionally, malabsorption secondary to gastrointestinal (GI) tract dysfunction or disease states amplifies the issue for already nutritionally deficient individual by further decreasing substrates and cofactor availability. Further considerations must be made for the patient with enzyme deficiencies, such as those who are lactose intolerant. This can lead to added substrate and cofactor perturbations as well as enzyme dysfunction.
Metabolism of the Body as a Whole
In the stressed state, the body undergoes various physiologic and metabolic changes. A hypermetabolic state ensues after a stress response, such as surgery, trauma, or sepsis, which leads to insulin resistance and hyperglycemia. There is increased oxidative stress at the cellular level and systemic activation of various inflammatory processes that lead to complex immunologic changes. There is a poor adaptation to starvation, and accelerated turnover of energy stores [15].
Basics of Whole Body Metabolism
The building blocks of the human body are carbohydrates, proteins and fats. These components are used in various ways and with various levels of efficiency by all aerobic cells. Glucose, the preferred fuel of cellular metabolism, is utilized in the well-fed state. Fat is stored for later oxidation when readily available stores of glucose are depleted. Protein, viewed largely structural in nature rather than used for energy purposes, is used as an endogenous energy source during stressed states and periods of altered metabolism. Glucose is the driving force behind energy production, and maintenance of a readily available source of glucose is of primary importance. Glycogen, the immediately available storage form of glucose, is found primarily in the liver and is available for approximately 12–24 h of fasting. After these stores are depleted, the body will then turn to protein and fat stores to produce glucose through gluconeogenesis. If starvation persists, other metabolic changes occur to provide alternate fuels to sustain cellular metabolism. In the stressed state, energy generation is deranged even further, in conjunction with greatly increased metabolic demand.
Energy Expenditure
Total energy expenditure (TEE) is divided into three components in healthy subjects: resting energy expenditure or metabolic rate (REE), energy required for digestion, and energy expenditure associated with activity. Various other things contribute to TEE, including the ambient temperature, vasoconstriction, shivering or stressed states [16]. To measure energy expenditure, one must have a basic knowledge of what each of these three components contribute to the total.
REE, or the basic metabolic rate in the fasting state at rest, represents anywhere from 60 % to 75 % of TEE. REE can be measured using various methods. In general, this measurement is determined by body size and body composition. Gender, age, and race also affect the basal metabolic rate to varying degrees [17].
Digestion is a thermogenic process that contributes around 5 % to 10 % of the TEE. The magnitude of this contribution is determined by a number of factors, including the size of the meal, the components of the meal, the time of day it is eaten, age, and the amount of caffeine or spice within the meal [18].
Physical activity is the most variable component of TEE and is therefore the most difficult to predict. In hospitalized patients, it is generally considered to compose around 5 % to 10 % of TEE. However, this is incredibly variable and can be affected by the activity of the patient, shivering, “thrashing” behavior, or fever [19].
There are various ways to measure energy expenditure. Because the components of TEE are so variable and unpredictable, predicting this reliably and for an extended period of time remains rather difficult in actuality. There are over 200 predictive equations to estimate energy expenditure, and they are often inaccurate or incomplete, resulting in over or underestimation of energy expenditure, especially in stressed states. Measuring energy expenditure frequently is simply not practical or possible.
Estimating Energy Expenditure
Direct Calorimetry
Direct calorimetry measures the exchange and release of heat from a patient. Heat is the basic byproduct of energy metabolism, and is therefore considered a direct measurement of energy expenditure. Direct calorimeters are complicated and sophisticated machines that measure the change in temperature of air or water that is circulated through the walls of a chamber housing the patient. Although most accurate, this technique is not practical in a clinical setting.
Indirect Calorimetry
Indirect calorimetry (IC) is considered by many to be the clinical “gold standard” of the measurement of energy expenditure. IC is certainly the most accurate for determining resting energy expenditure in the critically ill patient. “Metabolic carts” are available in most institutions, and today are often a component of modern computer driven ventilators. IC is most useful in specific disease states in which predictive equations are demonstrated to be unreliable or inaccurate, such as chronic respiratory distress syndrome, acute respiratory distress syndrome, or obesity. IC is also useful to help understand why patients are failing nutritional therapy when fed by standard predictive equations, as in the case of poor wound healing or delayed ventilator weaning, as it provides directed information on whether appropriate energy is being provided. IC is an effective tool in evaluating whether a patient is being over or underfed by measuring respiratory quotients.
Indirect calorimetry calculates REE and the respiratory quotient (RQ) by measuring whole body oxygen and carbon dioxide gas exchange. The accuracy of IC is dependent on many variables, and these must be manipulated to achieve a steady state (see Table 1.1) [20]. Once obtained, the REE gives a good approximation of the patients energy needs at that time.
The RQ helps direct the amount of energy provided, as well as the component composition of carbohydrate, protein, and lipid that is being provided. An RQ of less than 0.82 implies lipid catabolism, usually from underfeeding, and an increase in caloric delivery is warranted. A RQ of greater than 1 indicates pure carbohydrate oxidation and the potential for lipogenesis, usually from overfeeding. This increases the respiratory demand, and the total caloric and carbohydrate delivery should be reduced. A RQ of 0.85 indicates appropriately mixed substrate delivery, which is a sign of appropriate caloric and component provision. Although useful in clinical settings, care must be exercised when interpreting the RQ, as conditions such as diabetes, ketoacidosis, or hypermetabolism from inflammatory states can alter the value significantly [21, 22].
Predictive Equations
There are many other ways to measure or estimate resting energy expenditure. Body composition measurements can help to estimate this value via water displacement and whole body air displacement plethysmography (ADP) in commercially available machines. These methods are reserved and utilized for outpatient, healthy individuals wishing to gain a better understanding of their metabolic footprint, and has extremely limited usefulness in the inpatient clinical setting.
Numerous equations exist to estimate energy expenditure, and understanding the data needed and the reliability of each formula is important to calculating a sound therapeutic goal. The reverse Fick equation estimates oxygen consumption from the content of oxygen within arterial and mixed venous blood multiplied by cardiac output measurements. However, this requires the placement of a pulmonary artery catheter to obtain data points, and is biased toward underestimation, both contributing to its limited usefulness in the clinical setting [23]. The Harris-Benedict equation predicts the basal metabolic rate, factoring in variables such as sex and age. This equation is used very commonly; however is limited because it does not take into consideration factors such as body mass index or age and racial diversity of current patient populations. The formulas are as follows [24]:
- Men:
-
Energy expenditure = 66 + 13.75 (wt in kg)+ 5 (ht in cm) – 6.8 (age)
- Women:
-
Energy expenditure = 655 + 9.6 (wt in kg) + 1.8 (ht in cm) – 4.7 (age)
It is common practice to use “adjusted body weight” parameters within the Harris-Benedict equation:
This practice will underestimate energy expenditure in most obese patients.
To combat this underestimation of the Harris-Benedict equation in the obese, as well as to take into consideration the changes in metabolism that occur in critical illness, many other equations have been put forth. The most commonly used of these are the Penn State and the Ireton–Jones equations. These equations use modifiers for illness severity, inflammatory states, respiratory demands, body weight and height, and include fluid factors like body temperature and minute ventilation. This improves reliability in critical ill patients.
Ireton Jones for spontaneously breathing patients [25]
-
Energy expenditure = 629 – 11 (age) + 25 (actual body weight) – 609 (BMI >27 1 = present, 0 = absent)
Ireton Jones for ventilated patients
-
Energy expenditure = 1784 – 11 (age) + 5 (actual body weight) + 244 (sex, male = 1, female =0) + 239 (Diagnosis of trauma 1 = present, 0 = absent) + 804 (BMI >27 1 = present, 0 = absent)
Penn State [26]
-
REE (kcal/day) = Harris-Benedict equation (0.85) + minute ventilation (33) + Maximum body temperature in 24 h (175) – 6433
To make estimation of energy needs less cumbersome for the bedside practitioner, the American Society of Parenteral and Enteral Nutrition have devised a simple calculation using kcal/kg based on the findings of the various estimation methods. Generally, a range of 20–35 kcal/kg/day is recommended for adults [27]. These requirements can be under or overestimated in patients who are severely critically ill, obese, or failing to respond to energy provided, but are quick and easy to use at the bedside.
Provision of Energy Requirements
Once an accurate estimation of the total energy requirements a patient needs, the next step is to determine what components to provide. A healthy subject generally meets their REE with a carbohydrate: lipid: protein ratio of approximately 50 %: 30 %: 20 %. This ratio changes based on the stress the individual is under and what illnesses or comorbidities they are experiencing [28]. There is also a robust philosophy that protein calories are included in meeting TEE and the limitations on provision of lipids is held at 30 % of TEE and carbohydrates to less than 400 g/day.
Carbohydrates
Carbohydrates are generally the most efficient and readily available form of glucose. The minimal requirement for carbohydrate intake is determined by the brain’s need for glucose. A minimum of 50 g of carbohydrates/day is necessary to avoid ketogenesis. The liver produces approximately 50 % of serum glucose via glycogenolysis and 50 % of serum glucose from gluconeogenesis [29, 30]. The main function of glycogenolysis and gluconeogenesis is to maintain plasma glucose homeostasis, usually in the range of 80–120 mg/dL. This level is necessary to supply a constant state of substrate provision to the tissues and organs. The liver plays an integral role in the maintenance of glucose homeostasis during fasting and fed states via production of lipoproteins and glycogen for storage, as well as the breakdown of these storage products to release glucose. Normal blood glucose levels are hormonally regulated through the interplay of cortisol, insulin, and various catecholamines. In illness or injury, peripheral glucose oxidation is slowed. This, coupled with increased circulating levels of stress hormones, causes a stress induced hyperglycemia, contributing to increased protein breakdown from muscles and blunting immune responses [31, 32].
Carbohydrate provision should usually make up no more than 50 % of caloric provision in patients, though this percentage should be altered in specific disease states. Dextrose is the most common carbohydrate form provided in enteral nutrition therapy.
Proteins
While all macronutrients are affected by the metabolic changes associated with the hypermetabolic state, protein catabolism is disproportionately increased [33].
A negative energy balance in critical illness correlates to morbidity [34] and most acutely with loss of lean body mass [33]. Heightened rates of protein catabolism in the stressed state lead to loss of lean body mass. Increased turnover of protein remains unabated after a stress response despite nutritional support that would be considered adequate for a non-stressed individual [33].
There are many factors leading to this increased loss of protein, including mobilization of peripheral stores for gluconeogenesis, production of acute phase reactants, promotion of wound healing via production of cells like fibroblasts, and restoration of acid base balance in the kidney [33]. Support of the immune system via maintenance of gut mucosal integrity is another important source of protein turnover and utilization [35]. Protein turnover is also increased by states such as starvation and prolonged immobilization, both of which are common during critical illness. This heightened and exaggerated catabolism of protein is disproportionate to the overall increase in metabolism as well; we know that provision of nonprotein calories in amounts sufficient to meet energy demands does little to preserve lean body mass and improve nitrogen sparing. In fact, protein requirements can increase to 15–20 % of total calories during times of physiologic stress [36].
Another concept to consider when thinking about protein requirements is the idea of “conditionally essential” amino acids, as mentioned earlier. Certain amino acids which are not considered essential in the non-stressed state become essential during critical illness because of increased requirements for that particular amino acid. For example, high amounts of tryptophan, phenylalanine, and tyrosine are seen in acute phase proteins, and the increased synthesis of these proteins requires proportionately higher need for these amino acids [37]. Similarly, glutamine, which is normally 50 % of the body’s intracellular free amino acid pool, serves as fuel for rapidly dividing cells and as a nitrogen shuttle between visceral organs and muscle tissue, rapidly decreases in concentration in certain states and exceeds the body’s ability to synthesize it [38, 39]. This alteration in substrate utilization can lead to breakdown of muscle tissue in order to supply these “conditionally essential” amino acids, further enhancing the loss of lean body mass.
Loss of lean body mass and negative nitrogen balance is seen most dramatically in the first 9–12 days following an insult, but even continues to exceed the loss that would be expected based on the increase in resting energy expenditure through the first 21 days [40]. In fact, 16 % of total body protein is lost in that first 21-day period, 67 % of which comes from muscle stores [40]. Loss of lean body mass in critical illness is linked to increased rates of pneumonia, impaired wound healing and prolonged rehabilitation, pressure ulcers, and even mortality [41].
Minimizing the loss of lean body mass and preservation of a positive nitrogen balance is obviously important to reduce the negative impacts of stress catabolism. The question that remains is how do we best achieve this goal? Every patient is different, and different conditions affect this catabolic state differently. For example, minor trauma or surgery has less of an effect on protein turnover than severe sepsis or burns. There are many calculations and methods to assess protein metabolism and nitrogen balance, including assessing nitrogen balance, measuring amino acids directly by sampling directly across tissue beds, measurement of tracers to assess protein degradation, and turnover of individual components, such as urea.
Of these methods, nitrogen balance is the most widely used [33].
Nitrogen balance is best assessed through the quantification of urine urea nitrogen.
-
Nitrogen balance = Nitrogen intake – Nitrogen output
-
Nitrogen output (g/day) = urinary urea nitrogen (mg/100 mL) × urinary volume (L/day)/100 + 20% of urinary urea losses + 2 g [42]
Calculating this equation requires a 24-h urine collection. In addition, this equation must be adjusted to account for losses from renal dysfunction, stool, ostomy, and fistula losses [15]. These losses, plus incomplete urine collections can make this equation somewhat erratic and cumbersome to use at the bedside. However, in general it is a reliable tool to assess the adequacy of protein delivery.
But, there is a paucity of data to suggest there are clinical benefits related to reduction of net N losses through the provision of protein-rich diets. Additionally, since there is some data implying high amounts of protein may in fact be harmful, there is a significant amount of uncertainly about the optimum amount of protein to provide. Current guidelines from the American Society for Parenteral and Enteral Nutrition (ASPEN), the European Society for Parental and Enteral Nutrition (ESPEN), and the Society of Critical Care Medicine (SCCM) recommend 1.5–2.0 g protein/kg/day to patients with moderate to severe stress as a result of trauma, sepsis, or surgery [43–45]. Patients with severe sepsis or burns may require up to 2.5–3 g protein/kg/day [46]. Ongoing assessments of adequate protein delivery, such as urine urea nitrogen and documentation of wound healing, should be performed.
While there is much discussion about the potential benefits of protein provision in the stressed state, there is also the question of whether or not protein intake should be limited in certain disease states. There is concern about of the amount of protein to provide patients with chronic liver disease. The concern for hepatic encephalopathy in this patient subset prompts many to restrict dietary protein. This practice can worsen protein calorie malnutrition that is already present in many of these patients and, in turn, worsen outcomes [47]. Most of these patients can tolerate protein without exacerbation of hepatic encephalopathy, and while telologically sensible, branched chain amino acid diets have not significantly impacted management [47].
Protein provision in renal failure is misunderstood. The restriction of protein in these patients has been the topic of debate for many years. The results of clinical trials are varied, some claiming benefit and others claiming none. The most recent data seems to suggest that a protein-restricted diet could delay the need for hemodialysis and has no harmful effects on nutritional status [48]. Once hemodialysis is started, there is no role for protein restriction.
Fats
Lipids are essential molecules that serve important functions throughout the human body, and are a major form of caloric support, comprising 15–30 % of recommended total energy intake. Depending on the chemical configuration, lipids provide a dense form of calories, at 8.3 kcal/g for medium chain and 9 kcal/g for long chain fatty acids. Similarly, the chemical composition of fatty acids has different effects on inflammation and immune response [49–55]. During times of critical illness or stress, provision of certain formulas can curb the immune response, lower infection, and improve aspects of patient outcomes [56].
Long chain fatty acids (LCFA) are typically composed of 14 or more carbon atoms and are described as saturated or unsaturated based on the degree of hydrogenation and double bonding of the carbon molecules in the chain. Monounsaturated fats have one double bond, where polyunsaturated fatty acids (PUFA) have more than one. As the number of double bonds increase and the number of carbon molecules decrease, the more liquid the fat is at room temperature. Thus, animal fats, which are longer, and tend to be less saturated, are solids at room temperature (i.e., lard) [57]. Location of the double bonds also has a major role in the metabolic end-products putative effects.
Trace Elements/Minerals
Most commercially available formulas provide Daily Recommended Intake (DRI) for vitamins and trace elements (see Table 1.2). Patients with severe malnutrition, high losses as with enteric fistulae, bypass procedures, and/or malabsorption may require additional supplementation. Many vitamins act as cofactors for various metabolic processes, including a vital role in the production of ATP. Deficiencies lead to detrimental physiologic conditions.
Vitamin B6 deficiency is associated with hyperhomocysteinemia and hyperglycemic states in surgical intensive care unit patients [59, 60]. High dose supplementation increased immune response of critically ill patients [61]. While signs and symptoms are nonspecific, vitamin B1 (thiamine) deficiency can go unrecognized in the intensive care unit. Early and appropriate supplementation can prevent negative consequences [62]. Vitamin B2 (riboflavin) is a key factor for flavin adenine dinucleotide (FAD) and flavin mononucleotide (FMN). A large proportion of critically ill individuals have suboptimal vitamin B2 status, which can be significantly improved with supplementation. However, this improvement is transient, and deteriorates with discontinuation of the intake [63, 64].
Outcome specific data is lacking in regard to certain vitamin deficiencies, however it should be the goal of the physician to provide adequate substrate and cofactors necessary to support the metabolic demands of the stressed patient.
Specific Nutrients and Their Effects on Metabolism
Omega 3 Oils
Omega 3 oils are long chain n-3 PUFA most commonly found in fish and canola oils. They are considered essential fatty acids, as humans cannot synthesize de novo. Two biologically active examples include eicosapentaenoic acid (EPA) (Fig. 1.1), a 20-carbon fatty acid, and docosahexaenoic acid (DHA) (Fig. 1.2), a 22-carbon molecule. Although found in much smaller amounts, docosapentaenoic acid (DPA) (Fig. 1.3), a 22-carbon fatty acid, also has metabolic importance. These fatty acids (FA) are absorbed from the gastrointestinal tract, transported to the liver, and released into the circulation in lipoproteins and plasma phospholipids. They are incorporated into cell membranes throughout the body. Following initial ingestion of fish oils, there is a nonlinear increase in the concentration of EPA and DHA in membranes within days, and with continued consumption, greatest inclusion around 2 weeks. Upon removal from the diet, elimination from plasma lipids occurs over a few days, while cellular membrane concentrations can remain elevated for up to 2 months [49].

EPA

DHA

DPA
When incorporated into cellular membranes, n-3 PUFAs will increase in proportion to n-6 FAs (such as arachidonic acid). This has clinical importance during stress as metabolism of n-6 FAs leads to proinflammatory and pro-thrombotic eicosanoid derivatives which impair T-cell function, cytokine secretion, leukocyte migration, and reticuloendothelial system function [50]. Conversely, n-3 phospholipid metabolism leads to less active and even anti-inflammatory molecules [51]. Omega-6 fatty acids are predominantly immune-suppressing by means of their products of metabolism, namely through the elongation to arachidonic acid. In large doses, they tend to inhibit the response of the reticuloendothelial system. They alter the function of neutrophils and macrophages [65]. They also affect signal transduction and immune mediation. The entities responsible for these actions are prostaglandins, leukotrienes, and thromboxanes—all eicosanoid mediators of inflammation that are derived from arachidonic acid metabolism [56]. High intake of omega-6 fatty acids has been associated with a prothrombotic and proaggregatory state that can be manifested as increased blood viscosity, decreased bleeding time, vasospasm, and vasoconstriction [66, 67].
Patients both at risk for and those with the diagnosis of acute lung injury are found to have decreased concentrations of n-3 PUFAs of 25 % and as low as 6 % of normal, respectively [51]. While some studies have found a beneficial effect of supplementation with n-3 enriched formulas [52–54], others have failed to show any benefit even with twice day supplementation [55].
In contrast to other omega-6 fatty acids, gamma-linolenic acid acts favorably on the immune system by modifying the cellular lipid composition and altering eicosanoid biosynthesis. The end products of its metabolism are anti-inflammatory and can affect the breakdown of arachidonic acid into its proinflammatory constituents [68, 69]. It has been added to enteral formulas for decades and is found naturally in borage oil [56].
The use of omega-3 fatty acids in inflammatory states has been supported in multiple prospective, randomized studies [68]. Found in oils such as flaxseed, safflower, canola as well as in fatty, cold-water fish (salmon, swordfish), omega-3 fats have been studied for decades secondary to their beneficial effects in critical illness. They have anti-inflammatory and neuroprotective properties from stimulating resolvins, protectins and maresins (lipid mediators), docosatrienes, and neuroprotectins, which are potent effectors of the resolution of inflammation [56]. Resolvins function by regulating polymorphonuclear neutrophil transmigration. Neuroprotectin decreases neutrophil infiltration, proinflammatory gene signaling, and nuclear factor-ĸB binding. Furthermore, EPA and DHA alter cellular level metabolism and response by changing cell membrane phospholipid ratio, affecting gene expression and endothelial expression of intercellular adhesion molecule-1 (ICAM-1), E-selectin and other endothelial receptors that regulate vascular integrity [69, 70]. EPA has also been found to help prevent the loss of diaphragm function in sepsis [71]. It may also enhance resistance to gram-negative pathogens, such as Pseudomonas, although this has thus far only been demonstrated in mice [72].
Beginning in 1999, the use of omega-3 fatty acids in acute respiratory distress syndrome (ARDS) and Acute Lung Injury (ALI) showed good results and outcomes [73]. Using EPA, DHA, gamma-linolenic acid (GLA), and antioxidants in ARDS improved ventilation, oxygenation, and intensive care unit (ICU) length of stay. Although with some mixed results, subsequent studies have confirmed the trend toward the beneficial use of these dietary lipids in ARDS and ALI, with modest morbidity and mortality improvement [54, 55, 74]. In many critical care practices it is common to use anti-inflammatory lipid formulas as pharmacologic agents to attenuate inflammatory states, both in passive modulation of the inflammatory response, and the active resolution of inflammation [56].
Despite these encouraging findings, there remain a number of points of concern regarding the widespread use of these agents ubiquitously in the care of the stressed individual. In the early studies, the control formulations used were “proinflammatory” (i.e., Pulmocare—a high fat (n-6), low carbohydrate formula). At that time, little was known of the proinflammatory tendency of this feeding. To date, further studies with a more neutral fat composition have not been added as a control to verify the results. It can be interpreted that at least partial benefit was seen secondary to the high omega-6 formula used as control, rather than entirely due to the beneficial effect of the study elements. Additionally, the definition of ARDS and ALI used in some studies was based on prior criteria, rather than contemporary definitions. The benefit seen in these studies may be called into question secondary to selection biases.
Further questions about adding gamma-linolenic acid in ICU setting have been raised. While there are theoretical benefits of supplemental use in the ICU setting [69], isolated benefit has only been shown in animal models [75–77]. Added concerns revolve around the optimal dosage and timing of supplementation, as well as the route of administration. For example, if parenteral nutrition is necessary, formulations with fish oil, olive oils, medium chain triglycerides, or structured lipids are not available in the United States, where soy based preparations are the only option [78]. Finally, many studies showing beneficial in conjunction with omega-3 fatty acids have also been conducted with other favorable agents such as glutamine, arginine, or nucleic acids. It is difficult to delineate the contribution of each agent toward the improved clinical metrics [79].
Valid concerns regarding supplementation with omega 3 FAs also revolve around drug interactions, the bioavailability in diabetics or schizophrenics and fasting hyperglycemia in diabetic patients. Despite a concern for increased risk of bleeding, there are no randomized controlled trials demonstrating this effect. Additional apprehensions center around the “fishy” aroma and the development of gas, bloating, eructation, and diarrhea. Time released preparations can help reduce these effects.
Amino Acids
During times of stress, as in the perioperative period, critical illness or following trauma, specific amino acids and their supplementation deserve consideration. A promising area on the horizon for protein delivery is the idea of specific amino acid delivery and the potential benefits of this practice. Specific amino acids, such as glutamine and arginine are becoming an ever-increasing part of so-called immunonutrition. Immunonutrition is the concept that specific therapeutic agents like amino acids, antioxidants, or fish oils can have powerful therapeutic benefits. We know that certain amino acids exert pharmacologic activity when given during critical illness, and that these actions can have wide and varied clinical results.
The amino acids that are the most promising as therapeutic targets are those that are conditionally essential. There is an overwhelming amount of data to support the claim that these immune modulating formulas impact outcomes (see Table 1.3). It is difficult, however, to know which portion of the formula is causing the clinical effect. Specialty formulas designed to enhance the immune system are costly, and many practitioners express concern that in the absence of hard data to support their use we cannot rationalize the expense. However, ASPEN and SCCM both recommend the use of immune modulating formulas containing arginine, glutamine, nucleic acids, omega-3 fatty acids, and antioxidants are recommended for patients undergoing major elective surgery, trauma, burns, head and neck cancer, and critically ill patients on mechanical ventilation. This is not a panacea however with caution being exercised in those with severe sepsis [45].
Glutamine
Glutamine (Fig. 1.4) is the most abundant free amino acid in the human body, making up more than 50 % of the intracellular free amino acid pool. It has been studied in great detail over the past 15 years, and its effects and uses are wide and varied. Glutamine is an important fuel source for rapidly dividing cells, such as fibroblasts and reticuloendothelial cells, as well as the gut mucosa of the small bowel [33]. In addition, it carries two nitrogen moieties per molecule, and serves as a shuttle for nitrogen between muscle tissue and visceral organs [15]. Glutamine obviously plays an important role in cell homeostasis and organ metabolism. Because of these functions, glutamine stores rapidly deplete and decreased circulating concentrations are seen in exercise, sepsis, and trauma [80]. When the circulating concentration of glutamine exceeds the body’s ability to synthesize de novo glutamine, atrophy of intestinal mucosa, impairment of immune function, and decreased protein synthesis are seen [81]. Decreased levels of plasma glutamine have also correlate with increased mortality [82].

Glutamine
Glutamine exhibits a direct antioxidant effect and reduces oxidative stress on cells by attenuating the iNOS pathway in sepsis. It also acts directly on the tissue by preserving mitochondrial function and acting as a substrate for ATP synthesis. In addition, it attenuates cytokine expression [81]. But perhaps the most important manner in which glutamine exerts its actions on the immune system is by the upregulation of heat shock proteins. Heat shock proteins function to refold misfolded proteins coming off the endoplasmic reticulum, help eliminate irreversibly damaged proteins by marking them for apoptosis, stimulate the innate immune system, and attenuate proinflammatory cytokines [83]. Heat shock proteins are induced by a stress response, and this is enhanced by glutamine administration, heightening the response [84, 85]. The proposed mechanism is attenuation of the inflammatory response by binding and inhibiting nuclear factor kappa-light-chain-enhancer of activated B cells (NF-ĸΒ), thus limiting interleukin (IL)-6 and tumor necrosis factor-α (TNF-α), which occurs as a proinflammatory response to injury/sepsis [86].
Glutamine supplementation in critical illness has been shown to decrease mortality, length of stay, and infectious morbidity. Interestingly, there data that suggest there is no effect from glutamine supplementation. These studies are very heterogeneous with respect to patient population, route of glutamine administration, and amount of glutamine administration [86]. It seems that high doses given via a parenteral route demonstrate greater effect than low doses given by the enteral route. It is recommended to give high dose oral glutamine at levels greater than 0.2–0.3 g/kg/day (or, roughly, 20–40 g/day). There is more data for intravenous (IV) than oral (PO) glutamine; however PO in high doses seems to be efficacious [85].
Arginine
Arginine (Fig. 1.5) is considered a nonessential amino acid under normal physiologic conditions. l-Arginine is available to the host from endogenous synthesis via citrulline conversion in kidney, from endogenous protein breakdown, and from dietary protein sources [87]. Arginine plays an active role in cell growth and proliferation, wound healing, immune function and regulation, and waste ammonia disposal through the urea cycle [33]. Arginine also helps regulate vascular endothelial vasodilatation through the nitrogen oxygen synthase (NOS) pathway and the generation of nitric oxide (NO) [88]. NO is also a potent intracellular signaling molecule influencing virtually every mammalian cell type and is involved in the cytotoxicity of activated macrophages [87]. De novo synthesis, as well as dietary supplementation of arginine is diminished in critical illness, making it a conditionally essential amino acid.

Arginine
In critical illness, the cellular demand for arginine is increased due to the up regulation of arginase-1 in trauma and surgery [89]. In addition, plasma arginine levels diminish rapidly in trauma, critical illness, and sepsis [90]. When arginase pathways are upregulated, there is a resultant decrease in substrate availability for the opposing NOS pathway, thereby downregulating NO synthesis [87]. This inhibition of the NOS pathway is secondary to increased circulating levels of asymmetric dimethylarginine (ADMA), which may have a direct inhibitory effect on the NOS pathway [91, 92]. ADMA is then converted to citrulline, which is then use to synthesize arginine [19]. The question is whether this downregulation of the NOS pathway and resultant decrease in the levels of circulating NO is the source of the vasoconstriction and end organ malperfusion seen in sepsis, or if this downregulation is an adaptive strategy the body undergoes to prevent hypotension after critical illness [93].
This is the source of much controversy in the topic of arginine supplementation. Is the increased activity of arginase-1 involved in the increased levels of ADMA purely a mechanism by which to increase de novo synthesis of arginine in times of need? Does supplementation of arginine fuel the NOS pathway in preference to the arginase-1 pathway? In hemodynamically unstable critically ill patients, there was concern that vasodilatation related to increased nitric oxide could worsen systemic hypotension [94]. As arginine is a common constituent of many immune-enhancing formulas utilized in the critically ill patient, this concept led to growing concern about immunonutrition use in septic patients.
Making generalized statements about amino acid metabolism in critical care is extremely difficult because the “critically ill” population is not a homogeneous group. Making any generalized statements about the toxicity or benefits of any dietary supplement, let alone an amino acid with the metabolic complexity of arginine is not advisable as no study of these individual supplements can effectively be done in a clinical setting. Although theoretical concern is understood, both animal and human data are available to support arguments for and against the use of arginine [95, 96]. In light of this conflicting data, currently ASPEN recommends against the use of arginine containing formulas in the hemodynamically unstable patient regardless of the nature of the insult [45]. Further research as to the safety and efficacy of arginine in sepsis is needed.
In hemodynamically stable patients, the optimal dose of arginine to be delivered has yet to be determined. It seems that the 15–30 g of enteral supplemental arginine is safe and appears to meet the needs of the patient [97]. This is the amount that is commonly received at goal rates with immune modulating formulas.
Amino Acids on the Horizon
There are many other promising amino acid targets that are the fuel for ongoing research. Citrulline (Fig. 1.6) is one such amino acid. Citrulline is utilized in the de novo synthesis of arginine and is produced through conversion of ADMA, as previously discussed [92]. Studies demonstrate that citrulline availability is closely linked with arginine availability [95, 97]. Provision of citrulline may provide for de novo synthesis of arginine, thereby reducing the amounts of circulating ADMA and its deleterious effects.

Citrulline
Choline is another essential amino acid that is necessary for cell membrane structure. Choline is not a part of many parenteral nutritional formulations, and patients on long-term PN have impaired choline synthesis [33]. There is some evidence to support that this choline deficiency plays a role in the hepatic impairment seen in patients on long-term PN, and enteral repletion of choline to prevent or treat PN-induced hepatic failure is currently under research and shows promise [98].
States of Altered Metabolism
Metabolism in the Stressed State
One of the key ways in which metabolism impacts the surgical patient is catabolism. Surgical stress, trauma, burn injury, and critical illness all impact basic cellular metabolism, essentially creating a metabolic pathway leading to energy consumption and tissue breakdown. The inflammatory response is propagated by the release of inflammatory mediators, such as the numerous interleukins and tumor necrosis factor alpha [99]. The metabolic response to stress may be further altered by the presence of bacterial products, such as endotoxin. Even pain and anxiety can indirectly trigger a catabolic response through activation of the neuroendocrine system [100]. This catabolic response is directly proportional to the degree of stress that the host experiences, and can persist for weeks to months following the injury or illness [101].
The so-called “ebb phase” of the metabolic response to injury occurs immediately following the insult and lasts for several hours after the injury. It is characterized by elevated blood glucose levels with normal glucose production, elevated free fatty acid levels, low insulin levels, high levels of circulating catecholamines and glucagon, elevated serum lactate level, decreased oxygen consumption, and increased cardiac output [102]. The “flow phase” of metabolic response occurs next and can last weeks following injury, and is characterized by glucose intolerance, inflammation, and muscle wasting [102].
One of the most objective ways to measure the degree of response to stress in the host is to measure the alterations in resting energy expenditure (REE), via indirect calorimetry [103]. In minor trauma, such as long bone fracture, REE can increase from 10 % to 30 % over baseline. In sepsis, REE increases from 20 % to 25 % over baseline, while in severely stressed states, such as burn injury, REE can increase from 40 % to 100 % [104]. This increased need for substrates is due to the need for wound healing, synthesis of cellular and humoral immune components and acute phase reactants [33]. Amino acids are the major substrate for cellular metabolism during periods of catabolic stress, and lean body mass turnover is rapidly accelerated [33]. The body turns over muscle mass to provide alanine and glutamine as substrates to meet the increased energy needs [35, 36]. If exhausted or not appropriately supplemented, failure of gluconeogenesis in the liver, severe hypoglycemia, and amino acid deficiencies will result and, if left uncontrolled long enough, will lead to death.
The hypermetabolic state of the stressed host is attenuated by providing energy substrates. After an insult however, a compensatory anti-inflammatory response syndrome (CARS) has been described following periods of metabolic stress [105]. If CARS persists following injury or illness, the patient is metabolically deranged, immunocompromised, and susceptible to infectious complications. This is a major contributor to late deaths after trauma and injury and puts patients who have survived the acute illness at high risk for other or repeated infections.
Metabolic Response in Obesity
There are divergent outcomes about the effects of obesity in critical illness and injury. There are a number of studies that suggest that obesity confers a survival advantage to patients, or at minimum has no increased risk of mortality. However, there is also a growing body of literature demonstrating obesity is an independent predictor of mortality and significantly increases the risk of morbidity in this patient population. Studies that demonstrate obesity does not increase morbidity or mortality surmise the higher levels of anti-inflammatory adipokines such as IL-10 and leptin positively modulate surgical stress [106]. These studies have demonstrated that there is a decreased incidence of ARDS in the severely obese, a trend toward reduced length of stay, and despite a higher incidence of organ failure, no difference in mortality [107–110].
These results must be considered with the knowledge that extremely obese patients have a higher number of comorbid conditions, and that those conditions contribute to ICU-related mortality and morbidity [111, 112]. Studies dealing directly with the severely obese or those with centripetal obesity (the so-called “metabolic syndrome”), demonstrate increased risk of mortality, pneumonia, organ failure, and other complications [113–116]. These studies argue that the circulating levels of adipokines that are anti-inflammatory, such as IL-10, are actually decreased in the morbidly obese patient compared to their lean counterparts. In addition, circulating levels of proinflammatory adipokines, such as resistin and visfatin, are significantly increased in obesity, contributing to increased complications and death [117]. As the number of patients with obesity continues to grow worldwide, the mechanisms behind these findings are increasingly important to understand.
Metabolism in Malnutrition/Starvation
Starvation has a linear and temporally progressive negative impact on the patient. In early starvation, blood glucose concentrations decrease and serum insulin levels decrease in response to this change. Glycogen stores within the liver are depleted within 12–24 h. Soon after glycogen stores are depleted, the body induces a catabolic response, turning over protein stores for gluconeogenesis by the liver. Lipolysis increases as well, providing substrate for hepatic ketogenesis that peaks after 2–3 days of starvation [118].
After the early starvation phase peaks and wanes, the body enters an adapted starvation response. This host metabolism is designed to minimize protein breakdown by reducing energy expenditure. Keto-acids become the primary metabolic fuel for the brain and erythrocytes, replacing glucose. Ever decreasing circulating insulin levels allow for increased lipolysis. After all lipid stores are depleted, protein is the only source available for cellular metabolism. While the relationship of decay is linear and the resultant complications pattern or weakness, bedridden, skin breakdown, pneumonia follows, most deaths ensue from cardiac arrhythmias or arrest due to electrolyte abnormalities [119].
Areas of Future Research
Genomic Considerations
After injury or illness ensues, the inflammatory response is regulated by a release of cytokines, often termed a “cytokine storm.” These cytokines and the subsequent inflammatory cascade are implicated in the pathophysiology of organ failure after injury or illness [120]. Completely obliterating this inflammatory cascade has detrimental effects, often contributing to significant morbidity and early mortality. Conversely, a brisk and pronounced inflammatory response early after an insult has also been demonstrated to be disadvantageous [121].
It follows from the patient unique cytokine response to injury and illness that varies in time from insult and in degree to type of insult, that genetic encoding is responsible for the metabolic response to stress. The systemic inflammatory response syndrome (SIRS) is associated with activation of innate immunity. With severe SIRS, increased levels of TNF-α and toll-like receptor 4 (TLR4) circulate, and decreased circulating levels of IL-10 and transforming growth factor β1 (TGFβ1) are present [122–124]. A balance between pro and anti-inflammatory cytokines is crucial to prevention of SIRS-related complications and survival [125]. To date we have not been able to achieve this likely because the triggers for synthesis is unique to the DNA. Altered responses to stress such as trauma survival and ARDS frequency seem to be sex linked.
As we further elucidate the role of genetic encoding and its role in the development of the metabolic response to stress, potential targets for inflammatory modulation could be identified. If these targets could be manipulated early enough in the stress response and cytokine storm cascade, perhaps survival rates could be impacted dramatically.
References
“metabolism.” Merriam-Webster.com. 2011. http://www.merriam-webster.com (8 May 2011).
Dunn PM. Aristotle (384–322 bc): philosopher and scientist of ancient Greece. Arch Dis Child Fetal Neonatal Ed. 2006;91(1):F75–7.
Eknoyan G. Santorio Sanctorius (1561-1636) - founding father of metabolic balance studies. Am J Nephrol. 1999;19(2):226–33.
Karamanou M, Androutsos G. Antoine-Laurent de Lavoisier (1743-1794) and the birth of respiratory physiology. Thorax. 2013. doi:10.1136/thoraxjnl-2013-203840.
Fleck A. Obituary Notice: Sir David P. Cuthbertson. Br J Nutr. 1990;63:1–4.
Ferrannini E. The theoretical bases of indirect calorimetry: a review. Metabolism. 1988;37(3):287–301.
Saffle JR. The 1942 fire at Boston’s Cocoanut Grove nightclub. Am J Surg. 1993;166(6):581–91.
Dudrick SJ, Wilmore DW, Vars HM, Rhoads JE. Long-term total parenteral nutrition with growth, development, and positive nitrogen balance. Nutr Hosp. 2001;16(6):287–92.
Kudsk KA, Croce MA, Fabian TC, Minard G, Tolley EA, Poret HA, Kuhl MR, Brown RO. Enteral versus parenteral feeding. Effects on septic morbidity after blunt and penetrating abdominal trauma. Ann Surg. 1992;215(5):503–11. discussion 511–3.
The Veterans Affairs Total Parenteral Nutrition Cooperative Study Group. Perioperative total parenteral nutrition in surgical patients. N Eng J Med. 1991;325(8):525–32.
Bistrian B, Blackburn J, Vitale J, Cochran D, Naylor J. Prevalence of malnutrition in general medical patients. JAMA. 1976;235:1567–70.
Rivers E, Nguyen B, Ressler J, Muzzin A, Knoblich B, Peterson E, et al. Early goal directed therapy in the treatment of severe sepsis and septic shock. NEJM. 2001;345:1368–77.
Van den Berghe G, Wouters P, Weekers F, Verwaest C, Bruyninckx F, Schetz E, et al. Intensive insulin in critically ill patients. NEJM. 2001;345:1359–67.
Villet S, Chiolero RL, Bollmann MD, Revelly JP, Cayeux RNMC, Delarue J, Merger MM. Negative impact of hypocaloric feeding and energy balance on critical outcome in ICU patients. Clin Nutr. 2005;24:502–9.
Lin E, Calvano SE, Lowry SF. Systemic response to injury and metabolic support. In: Brunicardi FC, Anderson DK, et al., editors. Schwartz’s principles of surgery. New York: McGraw-Hill; 2005. p. 3–41.
Van Ooijen AM, Lichtenbelt VM, van Steenhoven AA, Westerterp KR. Seasonal changes in metabolic responses to cold in humans. Physiol Behav. 2004;82:545–53.
Compher CW, Frankenfield DC, Roth-Yousey L, Evidence Analysis Working Group. Best practice methods to apply to measurement of resting metabolic rate in adults: a systematic review. J Am Diet Assoc. 2006;106:881–903.
James WPT. From SDA to DIT to TEF. In: Kinney MM, Tucker HN, editors. Energy metabolism tissue determinants and cellular corollaries. New York, NY: Raven; 1992. p. 163–86.
Weissman C, Kemper M, Elwyn DH, Askanazi J, Hyman AI, Kinney JM. The energy patient: an analysis. Chest. 1986;89:254–9.
Wooley JA, Sax HC. Indirect calorimetry: applications to practice. Nutr Clin Pract. 2003;18:434–9.
Brandi LS, Bertonlini R, Calafà M. Indirect calorimetry in critically ill patients: clinical applications and practical advice. Nutrition. 1997;13:349–58.
Frankenfield DC, Sarson GY, Blosser SA, Cooney RN, Smith JS. Validation of a 5-minute steady state indirect calorimetry protocol for resting energy expenditure in critically ill patients. J Am Coll Nutr. 1996;15:397–402.
Ogawa AM, Shikora SA, Burke LM, Heetderks-Cox JE, Bergren CT, Muskat PC. The thermodilution technique for measuring resting energy expenditure does not agree with indirect calorimetry for the critically ill patient. JPEN J Parenter Enteral Nutr. 1998;22:347–51.
Frankenfield DC, Muth E, Rowe WA. The Harris-Benedict studies of human basal metabolism: history and limitations. J Am Diet Assoc. 1998;98:439–45.
Ireton-Jones CS, Jones JD. Why use predictive equations for energy expenditure assessment? Am Diet Assoc. 1997;97:A-44.
Frankenfield DC, Smith JS, Cooney RN. Validation of two approaches to predicting resting metabolic rate in critically ill patients. JPEN J Parenter Enteral Nutr. 2004;28:259–64.
Cerra FB, Benitez, Blackburn GL, Irwin RS, Jeejeebhoy K, Katz DP, et al. Applied nutrition in ICU patients: a concensus statement of the American College of Chest Physicians. Chest. 1997;111:769–78.
Wilmore DW. Metabolic response to severe surgical illness: overview. World J Surg. 2000;24:705–11.
Chandramouli V, Ekberg K, Schumann WC, Kalhan SC, Wahren J, Landau BR. Quantifying gluconeogenesis during fasting. Am J Phsiol. 1997;273:E1209–15.
Landau BR, Wahren J, Chandramouli V, Schumann WC, Ekberg K, Kalhan SC. Contributions of gluconeogenesis to glucose production in the fasted state. J Clin Invest. 1996;53(suppl):S101–6.
McCowen KC, Malhotra A, Bistrian B. Stress-induced hyperglycemia. Crit Care Clin. 2001;17:107–24.
Montori VM, Bistrian BR, McMahon MM. Hyperglycemia in acutely ill patients. JAMA. 2002;288:2167–9.
Young LS, Kearns LR, Schoepfel SL. Protein in the chamber. In: Gottschlich MM, editor. The A.S.P.E.N. Nutrition Support Curriculum. Cincinatti: A.S.P.E.N., 2007; p. 71–87.
Jabbar A, Wei-Kuo C, McClave SA, Dryden GW, McClave SA. Gut immunology and the differential response to feeding and starvation. Nutr Clin Pract. 2003;18:461–82.
Shaw JH, Wildborne M, Wolfe RR. Whole body protein kinetics in severely septic patients. The response to glucose infusion and total parenteral nutrition. Ann Surg. 1987;205:288–94.
Reeds PJ. Dispensible and indispensible amino acids for humans. J Nutr. 2000;130:1835S–40.
Griffiths RD, Allen KD, Andrews FJ, Allen KD, Andrews FJ, Jones C. Infection, multiple organ failure, and survival in the intensive care unit: influence of glutamine-supplemented parenteral nutrition on acquired infection. Nutrition. 2002;18:546–52.
Alpers DH. Glutamine: do the data support the cause for glutamine supplementation in humans? Gastroenterology. 2006;130:S106–16.
Monk D, Plank LD, Franch-Arcas G, Finn PJ, Streat SJ, Hill GL. Sequential changes in the metabolic response in critically injured patients during the first 25 days after blunt trauma. Ann Surg. 2001;234(2):245–55.
Martindale RG, Zhou M. Nutrition and metabolism. In: O’Leary JP, Tabuenca A, editors. Physiologic basis of surgery. 4th ed. Philadelphia: Lippincott Williams & Wilkins; 2008. p. 112–49.
Wilmore DW. Metabolic management of the critically ill. New York: Plenum; 1977, p. 193.
Stroud M. Protein and the critically ill; do we know what to give? Proc Nutr Soc. 2007;66:378–83.
Kreymann KG, Berger MM, Deutz NE, Hiesmayr M, Jolliet P, Kazandjiev G, Nitenberg G, et al. ESPEN (European Society for Parenteral and Enteral Nutrition): ESPEN guidelines on enteral nutrition: intensive care. Clin Nutr. 2006;25:210–33.
Jacobs DG, Jacobs DO, Kudsk KA, Moore FA, Oswanski MF, Poole GV, et al. Practice management guidelines for nutritional support of the trauma patient. J Trauma. 2004;57:660–79.
McClave SA, Martindale RG, Vanek VW, et al. A.S.P.E.N. Board of Directors; American College of Critical Care Medicine; Society of Critical Care Medicine: Guidelines for the provision and assessment of nutrition support therapy in the adult critically ill patient: Society of Critical Care Medicine (SCCM) and American Society for Parenteral and Enteral Nutrition (ASPEN). JPEN J Parenter Enteral Nutr. 2009; 33:277–316.
Chadalavada R, Sappati BRS, Maxwell J, Sappati Biyyani RS, Maxwell J, Mullen K. Nutrition in hepatic encephalopathy. Nutr Clin Pract. 2010;25(3):257–64.
Schulz GJ, Campos AC, Coelho JC. The role of nutrition in hepatic encephalopathy. Curr Opin Clin Nutr Metab Care. 2008;11(3):275–80.
Garneata L, Mircescu G. Nutritional intervention in uremia – myth or reality? J Ren Nutr. 2010;20(5 Suppl):S31–4.
Masson S, Latini R, Tacconi M, Bernasconi R. Incorporation and washout of n-3 polyunsaturated fatty acids after diet supplementation in clinical studies. J Cardiovasc Med (Hagerstown) 2007;8 Suppl 1:S4.
Kudsk KA. Immunonutrition in surgery and critical care. Annu Rev Nutr. 2006;26:463–79.
Calder PC. n-3 fatty acids, inflammation, and immunity – relevance to postsurgical and critically ill patients. Lipids. 2004;39(12):1147–61.
Kumar KV, Rao SM, Gayani R, Mohan IK, Naudu MU. Oxidant stress and essential fatty acids in patients with risk and established ARDS. Clin Chim Acta. 2000;298(1–2):111–20.
Gadek JE, et al. Enteral nutrition in ARDS study group. Effect of enteral feeding with eicosapentaenoic acid, gamma-linolenic acid, and antioxidants in patients with acute respiratory distress syndrome. Crit Care Med. 1999;27(8):1409–20.
Singer P, DeMichele SJ, Karlstad MD, Pacht ER, et al. Benefit of an enteral diet enriched with eicosapentaenoic acid and gamma-linolenic acid in ventilated patients with acute lung injury. Crit Care Med. 2006;34(4):1033–8.
Pontes-Arruda A, Aragão AM, Albuquerque JD. Effects of enteral feeding with eicosapentaenoic acid, gamma-linolenic acid, and antioxidants in mechanically ventilated patients with severe sepsis and septic shock. Crit Care Med. 2006;34(9):2325–33.
Munroe C, Frantz D, Martindale RG, McClave SA. The optimal lipid formulation in enteral feeding in critical illness: clinical update and review of the literature. Curr Gastroenterol Rep. 2011;13:368–75.
Calder PC, Dangour AD, Diekman C, Eilander A, Koletzko B, Meijer GW, et al. Essential fats for future health. Proceedings of the 9th Unilever nutrition symposium, 26–27 May 2010. Eur J Clin Nutr. 2010;64 Suppl 4:S1–13.
Vanek VW, Borum P, Buchman A, et al. A.S.P.E.N. Position paper: Recommendations for changes in commercially available parenteral multivitamin and multi-trace element products. Nutr Clin Pract. 2012;27(4):440–91.
Hou CT, Wu YH, Huang PN, Cheng CH, Huang YC. Higher plasma pyridoxal 5′-phosphate is associated with better blood glucose responses in critically ill surgical patients with inadequate vitamin B-6 status. Clin Nutr. 2011;30(4):478–83.
Hou CT, Wu YH, Cheng C, Huang PN, Huang YC. Higher plasma homocysteine is associated with lower vitamin B6 status in critically ill surgical patients. Nutr Clin Pract. 2012;27(5):695–700.
Cheng CH, Chang SJ, Lee BJ, Chang SJ, Lee BJ, Lin KL, Huang YC. Vitamin B6 supplementation increases immune responses in critically ill patients. Eur J Clin Nutr. 2006;60(10):1207–13.
Manzanares W, Hardy G. Thiamine supplementation in the critically ill. Curr Opin Clin Nutr Metab Care. 2011;14(6):610–7.
Vasilaki AT, McMillan DC, Kinsella J, Duncan A, O’Reilly DS, Talwar D. Relation between riboflavin, flavin mononucleotide and flavin adenine dinucleotide concentrations in plasma and red cells in patients with critical illness. Clin Chim Acta. 2010;411(21–22):1750–5.
Gariballa S, Forster S, Powers H. Riboflavin status in acutely ill patients and response to dietary supplements. JPEN J Parenter Enteral Nutr. 2009;33(6):656–61.
Roy CC, Bouthillier L, Seidman E, Levy E. New lipids in enteral feeding. Curr Opin Clin Nutr Metab Care. 2004;7(2):117–22.
Calder PC, Deckelbaum RJ. Harmful, harmless or helpful? The n-6 fatty acid debate goes on. Curr Opin Clin Nutr Metab Care. 2011;14(2):113–4.
Simopoulos AP. Essential fatty acids in health and chronic disease. Am J Clin Nutr. 1999;70(3Suppl):560S–9S.
Calder PC. The 2008 ESPEN Sir David Cuthbertson Lecture: Fatty acids and inflammation – from the membrane to the nucleus and from the laboratory bench to the clinic. Clin Nutr. 2010;29(1):5–12.
Fan YY, Chapkin RS. Importance of dietary gamma-linolenic acid in human health and nutrition. J Nutr. 1998;128(9):1411–4.
Singer P, Shapiro H, Theilla M, Anbar R, Singer J, Cohen J. Anti-inflammatory properties of omega-3 fatty acids in critical illness: novel mechanisms and an integrative perspective. Intensive Care Med. 2008;34(9):1580–92.
Supinski GS, Vanags J, Callahan LA. Eicosapentaenoic acid preserves diaphragm force generation following endotoxin administration. Crit Care. 2010;14(2):R35.
Tiesset H, Pierre M, Desseyn JL, Guéry B, Beermann C, Galabert C, Gottrand F, Husson MO. Dietary (n-3) polyunsaturated fatty acids affect the kinetics of pro- and anti-inflammatory responses in mice with Pseudomonas aeruginosa lung infection. J Nutr. 2009;139(1):82–9.
Gadek JE, DeMichele SJ, Karlstad MD, Pacht ER, Donahoe M, Albertson TE, et al. Effect of enteral feeding with Eicosapentaenoic acid, gamma-linolenic acid and antioxidants in patients with acute respiratory distress syndrome. Enteral Nutrition in ARDS Study Group. Crit Care Med. 1999;27(8):1409–20.
Pacht ER, DeMichele SJ, Nelson JL, Hart J, Wennberg AK, Gadek JE. Enteral nutrition with Eicosapentaenoic acid, gamma-linolenic acid, and antioxidants reduces alveolar inflammatory mediators and protein influx in patients with acute respiratory distress syndrome. Crit Care Med. 2003;31(2):491–500.
Chang CS, Sun HL, Lii CK, Chen HW, Chen PY, Liu KL. Gamma-linolenic acid inhibits inflammatory responses by regulating NF-kappaB and AP-1 activation in lipopolysaccharide-induced RAW 264.7 macrophages. Inflammation. 2010;33(1):46–57.
Mancuso P, Whelan J, DeMichele SJ, Snider CC, Guszcza JA, Karlstad MD. Dietary fish oil and fish and borage oil suppress intrapulmonary proinflammatory eicosanoid biosynthesis and attenuate pulmonary neutrophil accumulation in endotoxic rats. Crit Care Med. 1997;25(7):1198–206.
Murray MJ, Kumar M, Gregory TJ, Banks PL, Tazelaar HD, DeMichele SJ. Select dietary fatty acids attenuate cardiopulmonary dysfunction during acute lung injury in pigs. Am J Physiol. 1995;269(6 Pt 2):H2090–9.
Calder PC. Satellite symposium: throw another fish on the fire: the role of n-3 in inflammation. Rationale and use of n-3 fatty acids in artificial nutrition. Proc Nutr Soc. 2010;36(2):289–95.
Martindale RG, McClave SA, Vanek VW, McCarthy M, Roberts P, Taylor B, et al. Guidelines for the provision and assessment of nutrition support therapy in the adult critically ill patient: Society of Critical Care Medicine and American Society for Parenteral and Enteral Nutrition: Executive Summary. Crit Care Med. 2009;37(5):1757–61.
Oudemans-van Straaten HM, Bosman RJ, Treskes M, van der Spoel HJ, Zandstra DF. Plasma glutamine depletion and patient outcome in acute ICU admissions. Intensive Care Med. 2001;27:84–90.
Wischmeyer P. Glutamine: role in gut protection in critical illness. Curr Opin Clin Nutr. 2006;9:607–12.
Novak F, Heyland DK, Avenell A, Heyland DK, Avenell A, Drover JW, Su X. Glutamine supplementation in serious illness: a systematic review of the evidence. Crit Care Med. 2002;30:2022–9.
Macario AJL, Conway de Macario E. Sick chaperones, cellular stress and disease. NEJM. 2005;353:1489–1501.
Oliveira GP, Dias CM, Rocco PR. Understanding the mechanisms of glutamine action in critically ill patients. Ann Acad Bras Cienc. 2010;82(2):417–30.
Kelly D, Wischmeyer PE. Role of L-glutamine in critical illness: new insights. Curr Opin Clin Nutr Metab Care. 2003;6(2):217–22.
Singleton KD, Beckey VE, Wischmeyer PE. Glutamine prevents activation of nfkappab and stress kinase pathways, attenuates inflammatory cytokine release, and prevents acute respiratory distress syndrome (ARDS) following sepsis. Shock. 2005;24(6):583–9.
Zhou M, Martindale RG. Arginine in the critical care setting. J Nutr. 2007;137:1687S–92.
Albina JE, Mills CD, Henry Jr WL, Caldwell MD. Regulation of macrophage physiology by L-arginine: role of the oxidative L-arginine deaminase pathway. J Immunol. 1989;143:3641–6.
Morris SM. Recent advances in arginine metabolism. Curr Opin Clin Nutr Metab Care. 2004;7:45–51.
Chiarla C, Giovannini I, Siegel JH. Plasma arginine correlations in trauma and sepsis. Amino Acids. 2006;30:81–6.
Arrigoni F, Ahmetaj B, Leiper J. The biology and therapeutic potential of the DDAH/AMDA pathway. Curr Pharm Des. 2010;16(37):4089–102.
Pope AJ, Karrupiah K, Xia Y, Karrupiah K, Kearns PN, Xia Y, Cardounel AJ. Role of dimethylarginine dimethylaminohydrolases in the regulation of nitric oxide production. J Biol Chem. 2009;284(51):36338–47.
Fortin CF, McDonald PP, Fülöp T, Lesur O. Sepsis, leukocytosis, and nitric oxide (NO): an intricate affair. Shock. 2010;33(4):344–52.
Suchner U, Heyland DK, Peter K. Immune-modulatory actions of arginine in the critically ill. Br J Nutr. 2002;87 Suppl 1:S121–32.
Kao CC, Bandi V, Guntupalli KK, Wu M, Castillo L, Jahoor F. Arginine, citrulline and nitric oxide metabolism in sepsis. Clin Sci (Lond). 2009;117(1):23–30.
Luiking YC, Poeze M, Ramsay G, Deutz NE. Reduced citrulline production in sepsis is related to diminished de novo arginine and nitric oxide production. Am J Clin Nutr. 2009;89(1):142–52.
Pan M, Choudry HA, Epler MJ, Meng Q, Karinch A, Lin C, Souba W. Arginine transport in catabolic disease states. J Nutr. 2004;134:2826S–9S; discussion 2853S.
Piton G, Belon F, Cypriani B, Regnard J, Puyraveau M, Manzon C, et al. Enterocyte damage in critically ill patients is associated with shock condition and 28-day mortality. Crit Care Med. 2013;41:2169–76.
Seigel JH, Cerra FB, Coleman B, Giovannini I, Shetye M, Border JR, McMenamy RH. Physiological and metabolic correlations in human sepsis. Surgery. 1979;86:163–93.
Cuthbertson DP. Surgical metabolism: historical and evolutionary aspects. In: Wilkinson AW, Cuthbertson DP, editors. Metabolism and the response to injury. Chicago: Year Book Medical Publishers; 1977. p. 1.
Wilmore DW, Goodwin CW, Aulick LH, Powanda MC, Mason Jr AD, Pruitt Jr BA. Effect of injury and infection on visceral metabolism and circulation. Ann Surg. 1980;192:491.
Wilmore DW. Metabolic management of the critically ill. New York: Plenum; 1980.
Hwang TL, Huang SL, Chen MF. The use of indirect calorimetry in critically ill patients-the relationship of measured energy expenditure to Injury Severity Score, Septic Severity Score, and APACHE II Score. J Trauma. 1993;34(2):247–51.
Lawrence PF. Essentials of general surgery. 3rd ed. Philadelphia, PA: Lippincott, Williams & Wilkins; 1997.
Gentile LF, Cuenca AG, Efron PA, Ang D, Bihorac A, et al. Persistent inflammation and immunosuppression: a common syndrome and new horizon for surgical intensive care. J Trauma Acute Care Surg. 2012;72(6):1491–501.
Wu B, Fukuo K, Suzuki K, Yoshino G, Kazumi T. Relationships of systemic oxidative stress to body fat distribution, adipokines, and inflammatory markers in healthy middle-aged women. Endocr J. 2009;56(6):773–82.
Dossett LA, Heffernan D, Lightfoot M, Collier B, Diaz JJ, Sawyer RG, May AK. Obesity and pulmonary complications in critically injured adults. Chest. 2008;134(5):974–80.
Winfield RD, Delano MJ, Dixon DJ, Schierding WS, Cendan JC, Lottenberg L, et al. Differences in outcome between obese and nonobese patients following severe blunt trauma are not consistent with an early inflammatory genomic response. Crit Care Med. 2010;38(1):51–8.
Alban RF, Lyass S, Bargulies DR, Shabot MM. Obesity does not affect mortality after trauma. Ann Surg. 2006;72(10):966–9.
Dossett LA, Dageforde LA, Swenson BR, Metzger R, Bonatti H, Sawyer RG, May AK. Obesity and site specific nosocomial infection risk in the intensive care unit. Surg Infect. 2009;10(2):137–48.
Khaodhiar L, McCowen KC, Blackburn GL. Obesity and its comorbid conditions. Clin Cornerstone. 1999;2(3):17–31.
Muñoz E, Rosner F, Friedman R, Sterman H, Goldstein J, Wise L. Financial risk, hospital cost, and complications and comorbidities in medical non-complications and comorbidity-stratified diagnosis-related groups. Am J Med. 1988;84(5):933–9.
Neville AL, Brown CV, Weng J, Demetriades D, Velmahos GC. Obesity is an independent risk factor of mortality in severely injured blunt trauma patients. Arch Surg. 2004;139(9):983–7.
Brown CV, Neville AL, Rhee P, Salim A, Velmahos GC, Demetriades D. The impact of obesity on the outcomes of 1,153 critically injured blunt trauma patients. J Trauma. 2005;59(5):1048–51.
Choban PS, Weireter Jr LJ, Maynes C. Obesity and increased mortality in blunt trauma. J Trauma. 1991;31(9):1253–57.
Bochicchio GV, Joshi M, Bochicchio K, Nehman S, Tracy JK, Scalea TM. Impact of obesity in the critically ill trauma patient: a prospective study. J Am Coll Surg. 2006;203(4):533–8.
Lau DC, Dhillon B, Yan H, Szmitko PE, Verma S. Adipokines: molecular links between obesity and atherosclerosis. Am J Physiol Heart Circ Physiol. 2005;288(5):H2031–41.
Berg JM, Tymoczko JL, Starvation SL. Biochemistry. 5th ed. New York: W. H. Freeman; 2002.
Felig P, Marliss EB, Cahill Jr GF. Metabolic response to human growth hormone during prolonged starvation. J Clin Invest. 1971;50(2):411–21.
Dietch EA. Multiple organ failure. Pathophysiology and potential future therapy. Ann Surg. 1992;216(2):117–34.
Rivers EP, McIntyre L, Rivers KK. Early and innovative interventions for severe sepsis and septic shock: taking advantage of a window of opportunity. CMAJ. 2005;173(9):1054–65.
Song Z, Song Y, Yin J, Shen Y, Yao C, Sun Z, et al. Genetic variation in the TNF gene is associated with susceptibility to severe sepsis, but not with mortality. PLoS One. 2012;7(9):e46113.
Song Z, Yin J, Yao C, Sun Z, Shao M, Zhang Y, et al. Variants in the Toll interacting protein gene are associated with susceptibility to sepsis. Crit Care. 2011;15(1):R12.
Song Z, Yao C, Yin J, Tong C, Zhu D, Sun Z, Jiang J, et al. Genetic variation in the TNF receptor associated factor 6 gene is associated with susceptibility to sepsis induced acute lung injury. J Transl Med. 2012;10:166.
Reid CL, Perrey C, Pravica V, Hutchinson IV, Campbell IT. Genetic variation in proinflammatory and anti-inflammatory cytokine production in multiple organ dysfunction syndrome. Crit Care Med. 2002;30(10):2216–21.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Lawson, C.M., Long, C.A., Bollig, R., Daley, B.J. (2014). Introduction to Metabolism. In: Davis, K., Rosenbaum, S. (eds) Surgical Metabolism. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-1121-9_1
Download citation
DOI: https://doi.org/10.1007/978-1-4939-1121-9_1
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-1120-2
Online ISBN: 978-1-4939-1121-9
eBook Packages: MedicineMedicine (R0)