
Abstract
Plant hormones cytokinins stimulate cell division; regulate shoot and root development; promote leaf growth and flower, fruit, and seed formation; stabilize photosynthetic machinery; suppress senescence; and enhance sink strength and nitrogen acquisition. Cytokinin signaling is mediated by multistep phosphorelay. Binding of the cytokinin molecule to CHASE domain of the histidine kinase receptors triggers an autophosphorylation of the histidine domain and subsequent intramolecular transfer to receiver domain. Phosphoryl group is then transmitted to histidine phosphotransfer proteins and subsequently to type B response regulators (transcription factors) in the nucleus. Phosphotransfer proteins interact also with transcription factors CRFs (cytokinin response factors) that represent signaling side branch. The signal strength is regulated by cytokinin metabolism, which controls levels of active cytokinins, through feedback inhibition of signal transduction via type A response regulators (primary response genes), S-nitrosylation of phosphotransfer proteins, and/or proteasome degradation of type B and type A response regulators. Practical applications of cytokinins include their use in in vitro micropropagation, stimulation of flower branching, crop tillering or berry formation, and prolongation of fruit or tuber shelf life. Targeted elevation of cytokinin levels was found to increase the tolerance of plants to abiotic stresses, at least partially by diminishing the negative stress effects on photosynthesis. Recently, function of cytokinins in biotic stress responses has been also recognized. Full utilization of cytokinin potential to improve plant productivity by regulation of plant development has been until now limited by the necessity of targeting modulation of their levels or signal transduction in a time- and tissue-specific manner.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
In plants, the cytokinins (CKs) were defined as substances stimulating cell division (cytokinesis) in tissue cultures (Miller et al. 1955). Apart of this effect, CKs exhibit a wide range of physiological functions, including regulation of shoot and root apical meristems, stimulation of branching, vascular development, chloroplast differentiation, stabilization of the structure and function of the photosynthetic machinery, delay of senescence, stomata opening, and elevation of the sink strength and nutritional signaling (Mok and Mok 2001; Spichal 2012; Chernyadev 2009; Kiba et al. 2011; Ruffel et al. 2011). Naturally occurring CKs are N6-substituted adenine derivatives with either isoprenoid or aromatic side chain. Due to their predominance in plant tissues, attention has been focused mainly on isoprenoid CK research. The first identified natural CK, trans-zeatin (tZ), was named according to the species from which it was isolated (Zea mays, Letham 1963). Later on, other physiologically active CKs were identified: isopentenyladenine (iP), cis-zeatin (cZ), and dihydrozeatin (DHZ) as well as their ribosides (Sakakibara 2006). According to the activity in bioassays as well as the affinity to CK receptors, the most active CK is tZ, followed by iP (Spichal et al. 2004). Considerably less active cZ also widely occurs, being highly abundant in some species, especially in the monocots (Gajdosova et al. 2011). Apart from the species specificity, cZ seems to play a role in the stress responses (Dobra et al. 2010). DHZ was found predominantly in dormant seeds and apical buds, where it may serve as a source of active CKs before acceleration of de novo biosynthesis after germination (Frebort et al. 2011). CKs were detected not only in land plants but also in algae (Ordog et al. 2004), mosses (von Schwartzenberg et al. 2007), cyanobacteria, or symbiotic bacteria (Droog et al. 1997). Production of CKs by biotroph pathogens, e.g., Agrobacterium tumefaciens (Akiyoshi et al. 1984) or Rhodococcus fascians (Crespi et al. 1994), is part of their plant invading strategy. Detailed phylogenetic analysis of the occurrence of CKs and components of their signaling pathway was described by Spichal (2012).
Biosynthesis and Metabolism of CKs
The rate-limiting step of CK biosynthesis is transfer of isopentenyl moiety from dimethylallyl diphosphate (DMAPP) or its hydroxylated derivative (HMBDP) to adenosine 5′-phosphate (ATP and ADP in plants or AMP in bacteria). This reaction is catalyzed by isopentenyltransferase (IPT). In fact, CK biosynthetic genes were first characterized in A. tumefaciens. Plant IPTs were identified in Arabidopsis thaliana based on the homology with bacterial genes (Kakimoto 2001; Takei et al. 2001a). Plant IPTs strongly prefer ATP and ADP to AMP as well as DMAPP. The CK side chain may originate either from the plastid methylerythritol phosphate (MEP) pathway or from the cytoplasmic mevalonate (MEV) pathway. The prevalence of plastid pathway, in case of tZ and iP, was reported by Kasahara et al. (2004). The scheme of CK metabolism is shown in Fig. 1.

The scheme of cytokinin metabolism. Donor of isoprenoid side chain can originate either from plastid methylerythritol phosphate (MEP) or from the cytoplasmic mevalonate (MEV) pathway. IPT isopentenyltransferase, CYP450 cytochrome P450 monooxygenase, LOG cytokinin phosphoribohydrolase (LONELY GUY), CKX cytokinin oxidase/dehydrogenase, ZOG zeatin O-glucosyltransferase, N-GT cytokinin N-glucosyltransferase, GLU β-glucosidase, IS zeatin cis-trans isomerase, ZR zeatin reductase (adapted from Frebort et al. 2011, J Exp Bot 62: 2431–2452)
Some IPTs (AtIPT2 and AtIPT9) are tRNA-IPTs that prenylate adenine moieties adjacent to the 3′-end of the anticodon of specific tRNAs. This modification supports codon–anticodon interaction, as indicated by the disturbed interactions in bacterial tRNA mutants lacking CK moiety at A37 (Einset et al. 1976). Highly prevailing CK moiety in tRNAs is cZ. The tRNA degradation has been considered as an important source of cZ (Kamada-Nobusada and Sakakibara 2009).
In plants, the IPT-catalyzed reaction results in iP nucleotides that may be hydroxylated with cytochrome P450 monooxygenases (CYP735A1 and CYP735A2, Takei et al. 2004). As CK nucleotides are not active, gradual cleavage to nucleosides (i.e., ribosides) and bases by nucleotidase and nucleosidase was anticipated. Later on, CK-activating enzyme phosphoribohydrolase LOG (LONELY GUY), which converts all CK mononucleotides directly to the CK bases, was found (Kurakawa et al. 2007).
Due to the high physiological activity of CKs, the levels of the active forms need to be strictly regulated with respect to the plant developmental stage as well as environmental conditions. The key enzymes downregulating CK levels are cytokinin oxidases/dehydrogenases (CKX), which cleave the N6 side chain. Oxidative cleavage of CKs was first described by Paces et al. (1971). Later on, the enzyme was identified as cytokinin oxidase (Whitty and Hall 1974). The structure of CKX was first reported by Houba-Hérin et al. (1999). Galuszka et al. (2001) reclassified the enzyme as a dehydrogenase possessing slight oxidase activity. The substrates for CKXs are CKs with unsaturated side chain (iP, tZ, and cZ), while DHZ, containing a saturated side chain, and CKs with aromatic ring at N6 position are resistant to these enzymes.
Another important mechanism of suppression of CK activity is their glycosylation. Irreversible glucosylation occurs at the adenine ring in N7 or N9 position. CK N-glucosyltransferase was first isolated from radish (Entsch and Letham 1979). O-glucosylation (or much less frequent O-xylosylation) of the side chains containing a hydroxyl group or quite rarely observed N3-glucosylation is reversible (Mok and Mok 2001). The enzymes trans-zeatin O-glucosyltransferase, trans-zeatin O-xylosyltransferase, and cis-zeatin O-glucosyltransferase were identified by Martin et al. (1999a, b, 2001, respectively). CK O- and N3-glucosides can be hydrolyzed back by glucosidases (Brzobohaty et al. 1993). Very rare CK modification is the binding of l-alanine to N9 position of purine ring reported in lupine (Entsch et al. 1983) or the formation of methylthioderivatives in R. fascians (Pertry et al. 2009). Recently, in vitro interaction between CKs and NO (or more precisely with the product of NO reaction with superoxide–peroxynitrite) was reported (Liu et al. 2013). The authors identified several nitro- and nitroso-trans-zeatin and iP derivatives and suggested that reaction of CKs with NO can control levels of reactive nitrogen species in vivo.
Signal Perception and Execution of CK-Induced Responses
The CK signaling pathway is mediated by the multistep phosphorelay, similar to a two-component system found in bacteria for perception of extracellular stimuli (Argueso et al. 2009).
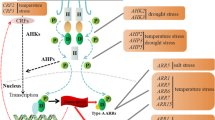
The scheme of cytokinin signaling pathway mediated by phosphorelay is shown in Fig. 2.

The scheme of cytokinin signaling pathway mediated by phosphorelay. The binding of cytokinin molecule by a CHASE-domain histidine kinase receptor (CHK) at the endoplasmic reticulum or plasma membrane results in autophosphorylation of the histidine kinase domain and transfer of the phosphoryl group to the receiver domain. Phosphoryl group is then transferred to histidine phosphotransfer proteins (HPts) in the cytoplasm, and the phosphorylated HPts cycle between the cytoplasm and nucleus and pass the phosphoryl group to type B response regulators (type B RRs). The activated type B RRs stimulate transcription of cytokinin primary response genes, including those encoding type A response regulators (type A RRs), which negatively affect signal transduction, at least partially by competing with type B RRs for the phosphate transferred by HPts. The signal from HPts may also be transferred to other transcription factors, the cytokinin response factors (CRFs)
Cytokinin Receptors
CK molecule is bound by CHASE-domain-containing histidine kinase receptors (CHKs, Heyl et al. 2013). The receptors contain sensor CHASE domain, which serves for interaction with CKs (Heyl et al. 2007), histidine kinase (HK) domain, and receiver domain (West and Stock 2001; Ueguchi et al. 2001). The binding of CKs to CHASE domain triggers an autophosphorylation of the receptor and subsequent intramolecular transfer of phosphoryl group (Gruhn and Heyl 2013). The phosphoryl residue is transferred to histidine phosphotransfer proteins (HPts) and subsequently to type B response regulators (RRs) (Shi and Rashotte 2012). The first CK receptor was described in Arabidopsis as a two-component hybrid molecule that regulates vascular morphogenesis WOL (WOODEN LEG, Mahonen et al. 2000). This CK receptor kinase was named also as CRE1 (cytokinin response 1, Inoue et al. 2001) or AHK4 (Arabidopsis histidine kinase 4, Suzuki et al. 2001; Yamada et al. 2001). Ueguchi et al. (2001) reported AHK4 together with two other CK receptors, the AHK2 and AHK3. The CK receptors exhibit partially overlapping activity, especially in roots, but play specific roles, given by their expression pattern and ligand specificity (Stolz et al. 2011). Recently, receptors were reported to be localized to the endoplasmic reticulum membrane (Caesar et al. 2011; Lomin et al. 2011; Wulfetange et al. 2011). However, their localization to plasma membrane can still be anticipated, too.
The ligand specificity of CK receptors is given by their CHASE domain, which was originally described as “Cyclase/Histidine kinase-Associated Sensing Extracellular” domain (Anantharaman and Aravind 2001; Mougel and Zhulin 2001). This full name was recently suggested to be changed to “Cyclase/Histidine kinase-Associated SEnsing” (Steklov et al. 2013). The crystal structure of the AHK4 CHASE domain was determined by Hothorn et al. (2011), who described also the AHK4 interactions with iP, tZ, benzyladenine, kinetin, and thidiazuron. The N-terminus of the AHK4 molecule folds into a long stalk α-helix, followed by the CHASE domain, which consists of two PAS-like (Per–Arnt–Sim-like) domains connected by a helical linker. The last β-strand of the membrane-proximal PAS domain (proximal to the C-terminus) is covalently linked to the N-terminus of the stalk helix by a disulfide bridge, which brings the flanking membrane helices into close proximity. The membrane-distal PAS domain forms binding cavity for CK molecule. In the lower part of the ligand-binding pocket, the central β-sheet of the PAS subdomain is lined by small hydrophobic residues, such as Ala and Gly. The hydrophobic upper part of the binding site is formed by two β-strands. The purine ring of CK molecule is oriented in the binding cavity by hydrogen bonds with Asp262 and Leu284. Approximately 20 amino acid residues are in contact with tZ. Three water molecules in the cavity mediate the additional interactions, including the hydrogen bond between Thr294 and side-chain hydroxyl group of tZ, which is the reason for much higher affinity of AHK4 to tZ than cZ (Spichal et al. 2004).
Phylogenetic analysis of ca. 100 receptors (Steklov et al. 2013) indicated that CHASE domain (ca. 220 amino acids) together with adjacent domains (totally about 280 amino acid residues) is enclosed at both sides with transmembrane helices. These hydrophobic regions seem to play a role in the correct subcellular localization as well as in intramolecular signaling. One transmembrane region occurs between CHASE and downstream kinase domains. This means that CHASE domain and catalytic part of the protein are always located at different sides of the membrane. Specific mutations in this downstream transmembrane helix render receptors constitutively active regardless of the CK presence. The number of upstream transmembrane helices may vary among the receptor orthologous groups from 1 to 4. The CRE1/AHK4 orthologs possess only one upstream transmembrane region, whereas AHK2 orthologs have three or four transmembrane helices. In AHK3 orthologs, the number of upstream transmembrane regions may vary. Steklov et al. (2013) suggested that upstream transmembrane helices are predominantly responsible for receptor subcellular localization, while downstream helices are involved in the signal transduction. The conserved N-terminal helix α1, upstream of the CHASE domain, may fix the appropriate conformation of the distal PAS domain and may regulate its movement upon CK binding. The substrate high-affinity binding results in specific conformational rearrangements of the PAS region in the sensory module. The signaling mechanisms of PAS domains were reviewed by Moglich et al. (2009). Signals originated within the conserved core generate structural and dynamic changes, which are propagated via amphipathic α-helical and coiled-coil linkers at the N- or C-termini of the core to the covalently attached effector domain. Many CK receptors were found to have a short coiled-coil motif that connects transmembrane helix with histidine kinase domain. Steklov et al. (2013) suggested that CK binding can affect the mode of interaction between ligand-binding PAS subdomains in the receptor dimer(s). Such change in the interaction mode of PAS subdomains might induce a mutual rotation of sensory modules relative to each other. The twist of sensory modules can in turn change the mutual position of transmembrane helices and cytoplasmic parts of receptors in dimer. As the histidine phosphorylation obviously occurs in trans by the parallel receptor, the change in relative position of receptors in dimer can switch on or off their kinase activity. Thus, formation of CK receptor complex results in HK activation and autophosphorylation of the conserved histidine in the catalytic module (West and Stock 2001). The phosphoryl group is then transferred intramolecularly to the conserved aspartate in the receiver domain. The HK domains of all Arabidopsis receptors have conserved histidine residue and five consensus motifs (H, N, G1, F, and G2). The receiver domain has conserved aspartate residue and three regions containing the conserved D, D, and K amino acid residues (Ueguchi et al. 2001). The subsequent transfer of CK signal is based on His–Asp phosphorelay (Grefen and Harter 2004; Muller and Sheen 2007; To and Kieber 2008; Schaller et al. 2011; Gupta and Rashotte 2012).
When ligand specificity of CK receptors was tested, AHK4 showed very high preference for tZ, followed by iP, and very low to cZ (Spichal et al. 2004). AHK3 showed only slight preference for tZ in comparison with tZR, iP, cZ, and DHZ. AHK2 has similar ligand specificity as AHK4 (Stolz et al. 2011). CK receptors differ also in their localization. AHK4 is expressed predominantly in roots, especially in vascular cylinder and pericycle of primary roots. AHK3 is expressed in rosette leaves, roots, stems, and flowers (Ueguchi et al. 2001; Higuchi et al. 2004). AHK2 is expressed in leaves, roots, and flowers (Ueguchi et al. 2001). In accordance with their expression pattern, CK receptors were reported to be involved in the regulation of root vascular morphogenesis (Mahonen et al. 2000) and shoot vascular development (Hejatko et al. 2009), control of root meristem (Dello Ioio et al. 2007) and shoot apical meristem size and activity (Higuchi et al. 2004; Skylar et al. 2010), as well as retardation of leaf senescence (Kim et al. 2006) and abiotic stress responses (Tran et al. 2007; Jeon et al. 2010).
Since its identification, AHK4 role in regulation of root vascular development has been recognized. AHK4 is the main CK receptor involved in the control of root vascular tissues (Mahonen et al. 2000, 2006b). AHK2 and AHK3, together with CKI1 (CYTOKININ-INDEPENDENT1) HK, are important regulators of shoot vascular tissue development. Their mutation results in defects in procambium proliferation and absence of secondary growth (Hejatko et al. 2009). The size of the root meristem was found to be negatively affected by AHK3 signal transduction (Dello Ioio et al. 2007). AHK3, but not the other CK receptors, plays a major role in CK-mediated chlorophyll retention and leaf longevity (Riefler et al. 2006; Kim et al. 2006). Homologues of AHK4 were reported to be indispensable for root nodulation in Medicago truncatula (Gonzalez-Rizzo et al. 2006) and Lotus japonicus (Tirichine et al. 2007). Cold-induced expression of a subset of type A Arabidopsis RR (ARR) genes, including ARR5, ARR6, ARR7, and ARR15, was shown to be mediated by the receptors AHK2 and AHK3 (Jeon et al. 2010).
It is interesting that AHK4 may function in the absence of CKs as a phosphatase, which dephosphorylates HPts and further suppresses CK signaling (Mahonen et al. 2006b).
Cytokinin Phosphotransfer Proteins
The components of CK signaling cascade downstream of receptors are the HPts that function as intermediate proteins to transfer the phosphoryl group from hybrid kinase receptors to downstream RRs (West and Stock 2001). In Arabidopsis, there are five authentic HPts (AHP1–5), which carry the conserved phospho-accepting His residue (Heyl and Schmulling 2003; Hutchison et al. 2006), and a pseudo-HPt (AHP6), which does not contain the conserved His residue necessary for phosphotransfer activity (Suzuki et al. 2000; Mahonen et al. 2006a). The AHPs have approximately 150 amino acids (Suzuki et al. 2000), except AHP4, which may occur in longer (145 aa) and shorter (127 aa) versions, and AHP5, which exhibits alternative splicing (Hradilova and Brzobohaty 2007). The authentic AHPs are positive regulators of CK signal transduction, which function to transfer phosphoryl group, obtained from AHKs, from the cytoplasm into the nucleus. Their continuous shuttling between the cytoplasm and nucleus was reported to be independent of their phosphorylation status (Punwani et al. 2010). The function of AHP4 is not clear; AHP4 had a slight positive effect in hypocotyl elongation assay, while in lateral root (LR) formation assay it acted as a negative regulator of CK response (Hutchison et al. 2006). However, it needs to be taken into account that AHP4 transcription levels are very low in most tissues. AHP4 may play a role in specific developmental processes (e.g., anther endothecium formation, Jung et al. 2008). The pseudo-HPt AHP6 is a negative regulator of CK signaling. Its transcription is downregulated by CKs (Mahonen et al. 2006a). Recently, S-nitrosylation of AHP1 by NO at Cys115 was reported, which suppressed AHP1 phosphorylation and subsequent transfer of phosphoryl group to ARR1 (Feng et al. 2013). This finding indicates an important mechanism for regulation of CK-induced phosphorelay activity in plants.
The crystal structure of one HPt protein from maize, the ZmHP2, was determined several years ago (Sugawara et al. 2005). ZmHP2 contains four C-terminal helices that form an antiparallel bundle connected to two N-terminal helices by a β-turn. The phospho-accepting residue is His80. The conserved residues surrounding His80 possibly act as a docking interface for receiver domains, while the non-conserved residues seem to be responsible for specific activities of different HPt proteins. More recently, the crystal structure of MtHPt1, an HPt from Medicago truncatula (MtHPt1), was reported. The MtHPt1, with His79 as its phosphorylation site, consists of six α-helices, four of which form a C-terminal helix bundle. The coiled-coil structure of the bundle is stabilized by a network of S-aromatic interactions involving highly conserved sulfur-containing residues (Ruszkowski et al. 2013).
The Arabidopsis AHP1 is expressed mainly in the roots; AHP2, AHP3, and AHP5 transcripts are widely spread in plants (in roots, stems, leaves, flowers, and siliques). The highest AHP2 expression is in roots and flowers, while AHP3 is predominantly expressed in roots and leaves (Suzuki et al. 1998; Hradilova and Brzobohaty 2007). The Arabidopsis AHPs, especially AHP2, AHP3, and AHP5, were found to be negative regulators of the drought response (Nishiyama et al. 2013). The loss-of-function mutants of these three AHP genes exhibited strong drought tolerance, improved cell membrane integrity under stress conditions, and increased sensitivity to abscisic acid.
AHP6 is expressed in developing protoxylem and pericycle cells, shoot apices, and young leaves (Mahonen et al. 2006a). It promotes protoxylem formation by counteracting CK signaling (Mahonen et al. 2006a). AHP6 also functions as a CK repressor during early stages of lateral root (LR) development. AHP6 is expressed at different developmental stages during LR formation. It is required for the correct orientation of cell divisions at the onset of LR development. Recently, AHP6 was found to influence localization of the auxin efflux carrier PIN1 that is necessary for patterning the LR primordia (Moreira et al. 2013).
Cytokinin Response Regulators
As mentioned above, the HPts transport the phosphate signal, received from receptor AHKs, from the cytoplasm to the nucleus, and transfer the phosphoryl groups to response regulators (RRs) (Gupta and Rashotte 2012). In Arabidopsis, there are two main classes of RRs, type A and type B ARRs. Type B ARRs are transcription factors (Sakai et al. 2000) which upon phosphorylation of a conserved Asp residue activate transcription of CK response genes (including type A ARRs). The type B ARRs possess an N-terminal phospho-accepting receiver domain and a C-terminal output domain containing a GARP (GOLGI-ASSOCIATED RETROGRADE PROTEIN) family Myb-like DNA-binding and transactivating region. In addition, there is a conserved nuclear targeting sequence located in the Myb-like/B motif of the type B RRs (Imamura et al. 2001; Hosoda et al. 2002). Three subfamilies of type B ARRs may be distinguished. Subfamily I includes ARR1, ARR2, ARR10, ARR11, ARR12, ARR14, and ARR18. This subfamily is the most important in mediation of CK responses (Hwang and Sheen 2001; Sakai et al. 2001; Argyros et al. 2008). Subfamily II consists of ARR13 and ARR21, and subfamily III of ARR19 and ARR20. ARR1, ARR2, ARR10, and ARR12 are expressed in young leaves. Their expression is restricted to the vascular tissues and hydathodes during the leaf maturation. ARR1, ARR2, ARR10, ARR11, and ARR12 are expressed in the roots, especially in root apical meristem and elongation zone (Birnbaum et al. 2003; Imamura et al. 2003; Mason et al. 2004; Tajima et al. 2004). The ARR1 is expressed at similar level throughout the stele, endodermis, cortex, and epidermis, but ARR10 is expressed at higher level in epidermis than in the other tissues (Mason et al. 2004; Birnbaum et al. 2003; Argyros et al. 2008).
The role of type B ARRs as positive regulators of CK signaling was demonstrated using ARR2-overexpressing plants which proved to be able to stimulate cell proliferation and shoot formation in the absence of exogenous CKs (Hwang and Sheen 2001). The fact that ARR1, ARR2, ARR10, and ARR12 (but not ARR11, ARR14, ARR18, ARR13, ARR19, and ARR20) were able to complement arr1arr12 mutant indicates functional diversities among the type B ARRs (Hill et al. 2013). The role of ARR1 and ARR12 in the control of cell division in shoot apical meristem seems to be mediated by transcriptional control of SHY2 (SHORT HYPOCOTYL 2), a suppressor of the auxin response (Dello Ioio et al. 2008). ARR2 was found to be activated downstream of AHK3 in the delay of leaf senescence (Kim et al. 2006). ARR1 and ARR12 were reported to suppress the expression of AtHKT1;1 (Arabidopsis thaliana high-affinity K+ transporter 1;1) that functions to remove sodium ions from the root xylem (Mason et al. 2010). ARR1 and ARR12 were thus suggested to delay the response to salinity stress.
The effects of the individual type B RRs on meristem size are generally consistent with their absolute transcript abundance, as well as with temporal changes in the expression (Hill et al. 2013). However, the ability of the type B RRs to stimulate transcription of CK response genes may be affected not only by the affinity or specificity to the target but also by potential interactions with HPts or transcriptional coregulators (Dortay et al. 2006; Kim et al. 2006). Promoter deletion analysis of the primary CK response gene ARR6 showed that a combination of two extended motifs within the promoter is required to mediate the full transcriptional activation by ARR1 and other type B ARRs. The identification of a novel enhancer, which is not bound by the DNA-binding domain of ARR1, indicates that apart from type B RRs additional proteins might be involved in mediating the transcriptional CK response (Ramireddy et al. 2013).
The function of the type B RRs may be also affected by the protein stability (Kim et al. 2012). Recently, specific degradation of type B ARRs upon binding to a family of F-box proteins KMD (KISS ME DEADLY) was reported (Kim et al. 2013). KMD proteins form an S-PHASE KINASE-ASSOCIATED PROTEIN1 (SKP1)/cullin/F-box protein (SCF) E3 ubiquitin ligase complex and directly interact with type B ARR proteins. The KMD family members are broadly expressed, predominantly in shoot apical meristem (especially KMD1 and KMD2) and in root tip (especially KMD2 and KMD3) (Kim et al. 2006). They are localized both in the nucleus and in the cytoplasm. KMD proteins interact with ARR1, ARR12, and ARR20, less with ARR2 and ARR10. ARR1 and ARR12 were found unstable, readily to be degraded by proteasome, independently of CK presence. In contrast, degradation of ARR2 by proteasome requires CK-induced phosphorylation (Kim et al. 2012). KMD proteins seem to be key players of an important mechanism that is responsible for reducing the levels of activated type B RRs, thereby preventing continued transcriptional activation by CKs (Kim et al. 2013). The representative members of type A ARRs, ARR4 and ARR7, were found not to be the substrates of KMD proteins.
Plant hormones regulate most physiological processes in an intensive cross talk. ARR2 seems to represent a link between CK and ethylene signaling pathways (Hass et al. 2004). ARR2 also makes a complex with TGA3 (TGACG-motif-binding transcription factor 3), a salicylic acid response factor. Salicylic acid signaling via NPR1 (NON-EXPRESSOR OF PATHOGENESIS-RELATED GENE1) enhanced binding of ARR2/TGA3 to the PR1 (PATHOGENESIS-RELATED PROTEIN1) promoter. CKs were thus found to promote resistance against Pseudomonas syringae in Arabidopsis (Choi et al. 2010).
Type B ARRs stimulate the expression of the type A ARRs (Hwang and Sheen 2001; Sakai et al. 2001), which are negative regulators of CK signaling and represent a negative feedback loop (CK signal switch-off). In Arabidopsis, ten type A ARRs were identified: ARR3, ARR4, ARR5, ARR6, ARR7, ARR8, ARR9, ARR15, ARR16, and ARR17 (To et al. 2004). These ARRs contain a phospho-accepting receiver domain, but no DNA-binding domain as do the type B ARRs. Phosphorylation of type A ARRs, for example, ARR5 and ARR7, at an aspartate of the phosphate receiver domain is a necessary prerequisite of their action as negative regulators (Lee et al. 2007; To et al. 2007). The mode of action of type A RRs seems to include competitive binding of the phosphoryl group from HPts at the expense of type B RRs. ARR5, ARR6, ARR7, and ARR15 were detected only in the nucleus; ARR4 and ARR16 were found both in the cytoplasm and in the nucleus (Hwang and Sheen 2001; Imamura et al. 2001).
Expression of several type A ARRs is rapidly induced by CKs, even after the inhibition of de novo protein synthesis (Brandstatter and Kieber 1998; Sakakibara et al. 1999; D’Agostino et al. 2000), suggesting that type A ARRs are CK primary response genes. To et al. (2007) specified a subset of type A ARRs stabilized by CKs, in part via phosphorylation (ARR5, ARR6, and ARR7), while ARR4 and ARR9 were not stabilized. The function of CKs as well as of proteasome in regulation of type A RR stability was studied by Ren et al. (2009). They found regulatory effect of CKs in case of ARR5, ARR6, ARR7, ARR8, ARR15, ARR16, and ARR17. Proteasome affected stability of ARR3, ARR5, ARR7, ARR8, ARR15, ARR16, and ARR17 (Ren et al. 2009).
Comparative analysis of Arabidopsis plants over-expressing individual members of type A ARRs showed their differential roles (Ren et al. 2009). The inhibitory effect of CKs on the primary root elongation was suppressed predominantly by ARR3 and ARR5 over-expression, followed by that of ARR4, ARR16, and ARR17. Inhibition of lateral root initiation was affected by most type A RRs, with the exception of ARR4, ARR5, and ARR7. Most type A RRs speeded up flowering, while only ARR16 was active in regulation of dark-induced leaf senescence. The strongest inhibition of CK-induced shoot formation was exhibited by ARR3, ARR5, ARR6, ARR16, and ARR17 (Ren et al. 2009). The expression of ARR5, ARR6, ARR7, and ARR15 is repressed by transcription factor WUS (WUSCHEL, “wuscheligen habitus”), in order to maintain optimal CK levels in shoot apical meristem (Leibfried et al. 2005). In root apical meristem, ARR7 and ARR15 transcription is positively regulated by auxin to maintain a balance between auxin and CK levels (Muller and Sheen 2008). A subset of type A ARRs, especially ARR5, ARR6, ARR7, and ARR15, are induced by cold (Jeon et al. 2010). Upregulation of type A RRs at the early phase of cold stress response is in accordance with transient downregulation of the active CK levels observed in winter wheat after exposure to cold (Kosova et al. 2012). ARR4 was found to represent a link between CKs and light signaling, interacting with phytochrome B (Fankhauser 2002; Sweere et al. 2001). ARR3 and ARR4 are involved in regulation of circadian rhythms (Salome et al. 2006).
Similar to type A RRs are the type C RRs, which are sometimes included into the type A RR group. Type C RRs have also only receiver domain. This domain is more related to the receiver domain of the hybrid histidine kinase receptors (Kiba et al. 2004; To and Kieber 2008). Additionally, type C RRs are not induced by CKs. This ARR group includes ARR22 and ARR24 (Gattolin et al. 2006). ARR22 is expressed in flowers and developing pods, where it undergoes alternative splicing. Expression of ARR24 was found restricted to pollen grains (Gattolin et al. 2006). Transcription of ARR22 is induced by wounding, which may indicate a possible role of type C ARRs in response to biotic stresses (Gattolin et al. 2006). ARR22 interacts with AHP2, AHP3, and AHP5, acting as phosphohistidine phosphatase (Horak et al. 2008).
CRFs represent a side branch of CK signaling pathway. These proteins can interact directly with HPt proteins. The Arabidopsis CRFs are induced by CKs and belong to AP2-/ERF-like (APETALA2/ethylene-responsive factor) transcription factor family, distinct from type B RRs (GARP-/Myb-related family). CRFs share some targets with type B RRs but also activate some other genes (Rashotte et al. 2003, 2006). CRFs occur broadly in land plants (Rashotte and Goertzen 2010) and are involved in the normal development of embryos, cotyledons, and leaves (Rashotte et al. 2006).
Improvement of Plant Productivity with Biotechnological Manipulation of Cytokinin Biosynthesis and Signaling
The multiple physiological functions of CKs, which include regulation of germination, shoot and root development, leaf growth, flower and fruit formation, suppression of leaf senescence, enhancement of sink strength as well as uptake of nitrogen (Mok and Mok 2001), make this hormone class very perspective for practical applications. Unfortunately, effective regulation of particular physiological processes requires very precise time- and site-specific targeting of modulation of CK levels or signaling.
Until now, exogenous applications of CKs have been predominantly used in practice, to enhance shoot formation, branching, and tillering, to improve nitrogen acquisition, or to delay senescence (e.g., see Mala et al. 2013; Malabug et al. 2010; Gapper et al. 2005). Aromatic CKs are preferentially used, as these types of CKs are not subjected to fast degradation by CKXs. N6-(3-hydroxybenzyl)adenine (meta-topolin) became a very promising alternative for widely used N6-benzyladenine (Strnad 1997). meta-Topoline has a high biological activity, which is in accordance with its relatively high affinity to AHK4 (Mok et al. 2005). As a good substrate of trans-zeatin O-glucosyltransferase (Mok et al. 2005), meta-topoline is metabolized to a storage O-glucoside, which can be gradually converted back to the active compound, in contrast to N6-benzyladenine that is quickly N-glucosylated, resulting in a stable metabolite accumulated in basal parts of plants (Werbrouck et al. 1996). Recently, an alternative approach—suppression of degradation of endogenous CKs by inhibition of CKXs—has been tested (Zatloukal et al. 2008; Motte et al. 2013).
Since their discovery in 1955, CKs are routinely used in in vitro cultures for stimulation of shoot differentiation and propagation. Micropropagation techniques are used, for example, for cultivation of ornamental plants (orchids, chrysanthemums or carnations, e.g., see Ferreira et al. 2006), for multiplication of elite clones of forest trees (pine, elm, poplar, eucalyptus, and teak, e.g., see Mala et al. 2013), or for propagation of potato (Baroja-Fernandez et al. 2002). Exogenous CKs are used in classical horticulture to increase branching and thus the amount stem cuttings and flowers (Kaminek et al. 1987). Exogenous CKs have been also used for prolongation of the flower vase life, e.g., of gerberas (Danaee et al. 2011). Benzyladenine together with gibberellin was shown to promote plant growth and yield in three strawberry cultivars (Momenpour et al. 2011). When synthetic CK CPPU [N-(2-chloro-4-pyridyl)-N-phenylurea] and gibberellic acid were applied to various grapevine varieties at the fruit setting stage, these hormones increased berry size in Perlette, Superior, and Thompson Seedless cultivars. Gibberellin was found to enhance cell expansion, while CKs to increase cell number and density (Ben-Arie et al. 1997). CPPU was also found to increase berry mass and firmness, as well as cluster mass and compactness in Vitis labrusca and V. labrusca × V. vinifera in field trials (Zabadal and Bukovac 2006). Souza et al. (2010) reported positive effect of benzyladenine application on the quality of clusters of cv. Superior Seedless grapes. CKs were tested to increase a wheat grain yield by promotion of tillering. Their effect was significantly positive when the original plant density was low. In case of high plant density, the amount of seeds was also increased, but their size was reduced; thus, the yield was not enhanced.
CK application may also allow reduction of nitrogen fertilization, as CKs promote nitrogen acquisition (Takei et al. 2001b; Sykorova et al. 2008; Kiba et al. 2011; Pavlikova et al. 2012). This may solve the problems associated with high nitrogen levels in the field soils and underground waters which often result from heavy fertilization used to maintain high grain yields.
Surprisingly, CKs have been commercially used also in “non-plant” areas. Their antiaging effects were proved also for human skin, as evidenced by the fact that several types of cosmetics contain CKs (e.g., Pyratine-6 antiaging cream). Some CK analogues were found to block cell cycle progression not only in plant cells (Vesely et al. 1994) but also in humans (Vermeulen et al. 2002). These CK analogues were successfully tested as anticancer agents (Casati et al. 2011; Molinsky et al. 2013).
When genetic approach to elevation of CK levels is applied, it is necessary to prevent too strong IPT over-expression. High CK levels cause morphological abnormalities and very high levels may induce cell apoptosis (Mlejnek and Prochazka 2002). This problem was solved by Gan and Amasino (1995), who expressed IPT gene under the control of senescence-inducible promoter (SAG12) in tobacco. Stimulation of senescence program in plants resulted in an enhanced activity of SAG12 promoter, leading to an increase of CK biosynthesis. Increased CK levels in turn suppressed the promoter activity, which prevented their overproduction. Repetition of these cycles allowed prolongation of the plant life-span. The SAG:IPT construct was successfully used in Lactuca sativa to delay developmental and postharvest leaf senescence in mature lettuce heads (McCabe et al. 2001). No significant effect of transformation on the head diameter or fresh weight of leaves or roots was observed. Postponed plant senescence, accompanied by the delay in the loss of photosynthetic activity, was observed in maize expressing ipt under the control of a senescence-enhanced maize promoter (Robson et al. 2004). The elevation of CK content by expression of SAG:ipt in cassava plants delayed substantially the post-harvest senescence of cassava tuberous roots (Zhang et al. 2010). Prolongation of the shelf life may be very important in some developing regions, where cassava represents substantial part of the diet. When promotion of photosynthetic activity and delay of senescence are desirable, stimulation of CK biosynthesis is advantageous (Gregersen et al. 2013). In some cases, however, delay of leaf senescence may interfere with the developmental program. The over-expression of SAG:ipt in wheat prolonged substantially the vegetative period, increasing the sink strength of leaves, which interfered with grain filling (Sykorova et al. 2008). Due to the smaller seed size, no yield improvement was observed, in spite of their increased number. Apart from the changed sink/source relations, increase in seed number may result in plant exhaustion that limits the seed growth. A similar situation was observed after stimulation of flower branching in chrysanthemum plants by over-expression of ipt under LEACO1 promoter, which resulted in substantial increase in flower number, but their diameter was smaller than in wild type (Khodakovskaya et al. 2009).
Recent climate changes have strengthened the demands for crops with improved stress tolerance, as unfavorable environmental conditions, including various abiotic and biotic stresses, may cause more than 50 % loss of the crop yield, especially in developing countries. As the response to the stress conditions requires vast re-programming of the metabolism to reallocate the energy supplies from the developmental programs to fast and effective stimulation of defense pathways, downregulation of CK levels, associated with low growth rate, was tested. CK deficiency achieved by over-expression of CKX or downregulation of CK biosynthesis was found to increase substantially drought tolerance (Werner et al. 2001; Mytinova et al. 2010; Nishiyama et al. 2011). The same phenomenon has been described with knock-out mutants of CK receptor (Tran et al. 2010) and of AHP-encoding genes (Nishiyama et al. 2013). Apart from drought, also salinity or heat stress tolerance was improved (Nishiyama et al. 2011; Mackova et al. 2013). Constitutive CKX expression promotes growth of the root system, a trait that positively correlates with tolerance to water deficit (Tuberosa et al. 2002). However, it has strong negative effect on the shoot growth. This drawback can be avoided by utilization of root-specific promoters. Targeting of CKX expression only to roots (Werner et al. 2010) resulted in plants which maintained enhanced root system, but their shoot phenotype was similar to wild type. Their stress tolerance was lower in comparison with 35S:CKX plants but still significantly higher than that of wild type (Mackova et al. 2013).
Interestingly, opposite strategy—elevation of CK levels by expression of CK biosynthetic gene (IPT)—also resulted in strong elevation of abiotic stress tolerance. In contrast to constitutive elevation of CK levels, which was associated with high sensitivity to drought (pssu:ipt, Synkova et al. 1999), IPT over-expression under the senescence- or stress-inducible promoters (SAG12, SARK, or rd29A) enhanced tolerance to drought (Rivero et al. 2007, 2009, 2010; Merewitz et al. 2010, 2012; Peleg et al. 2011; Qin et al. 2011; Qiu et al. 2012; Kuppu et al. 2013), heat (Xing et al. 2009), salinity (Ghanem et al. 2011), cold (Hu et al. 2005; Belintani et al. 2012), or flooding (Huynh et al. 2005). The underlying mechanism seems to be diminishing of the stress-induced suppression of photosynthetic activity and stabilization of photosynthetic machinery (Rivero et al. 2009), which improves the energy supply. Moreover, transcription of many stress-inducible genes could be stimulated by CKs (Hare et al. 1997). Recent reports indicate intensive cross talk of CKs with salicylic acid and jasmonic acid and potential positive effect of CKs in biotic stress responses (Choi et al. 2010; Synkova et al. 2006). CK functions in stress responses were recently reviewed by Argueso et al. (2009) or Ha et al. (2012).
The CK functions in regulation of plant development offer unique opportunities to target different processes using suitable promoters. The over-expression of ipt under cysteine protease promoter resulted in transgenic rice plants with early flowering and higher number of emerged panicles (Liu et al. 2010). Expression of ipt under the control of seed-specific lectin promoter in tobacco promoted cell division in the embryo, resulting in an increase in the number of plerome cell layers and cell number in cotyledons (Ma et al. 2008). Dry weight of seeds was higher and transgenic seedlings grew faster.
The alternative approach to over-expression of biosynthetic gene is silencing of the expression of deactivating gene (CKX). Bartrina et al. (2011) reported that Arabidopsis ckx3ckx5 double mutant formed larger inflorescence and floral meristems. Cellular differentiation was also retarded in this mutant, leading to higher cell number and larger flowers. Silencing of HvCKX1 expression in barley and wheat resulted in higher grain yield (Zalewski et al. 2010). As HvCKX1 exhibits high activity in the regulatory aleurone layer of the seeds, the positive effect of downregulation of CKX activity seems to be based on the increase of CK concentration in this layer with positive effect on sink strength and starch accumulation during grain filling (Zalabak et al. 2013).
The perspectives of genetic engineering of CK metabolism for the improvement of agricultural traits of crop plants were discussed by Zalabak et al. (2013), who also provided a comprehensive list of transgenic plants with altered expression of CK-related genes and their traits. The abovementioned data indicate that modulation of CK metabolism and/or signaling may represent a promising strategy for improvement of plant productivity, especially in combination with suitable tissue- and time-specific promoters that allow direct control of grain filling or stimulation of inflorescence meristems.
References
Akiyoshi DE, Klee H, Amasino RM, Nester EW, Gordon MP (1984) T-DNA of Agrobacterium tumefaciens encodes an enzyme of cytokinin biosynthesis. Proc Natl Acad Sci U S A 81:5994–5998
Anantharaman V, Aravind L (2001) The CHASE domain: a predicted ligand-binding module in plant cytokinin receptors and other eukaryotic and bacterial receptors. Trends Biochem Sci 26:579–582
Argueso CT, Ferreira FJ, Kieber JJ (2009) Environmental perception avenues: the interaction of cytokinin and environmental response pathways. Plant Cell Environ 32:1147–1160
Argyros RD, Mathews DE, Chiang YH, Palmer CM, Thibault DM, Etheridge N, Argyros DA, Mason MG, Kieber JJ, Schaller GE (2008) Type B response regulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell 20:2102–2116
Baroja-Fernandez E, Aguirreolea J, Martinkova H, Hanus J, Strnad M (2002) Aromatic cytokinins in micropropagated potato plants. Plant Physiol Biochem 40:217–224
Bartrina I, Otto E, Strnad M, Werner T, Schmulling T (2011) Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 23:69–80
Belintani NG, Guerzoni JTS, Moreira RMP, Vieira LGE (2012) Improving low-temperature tolerance in sugarcane by expressing the ipt gene under a cold inducible promoter. Biol Plant 56:71–77
Ben-Arie R, Sarig P, Cohen-Ahdut Y, Zutkhi Y, Sonego L, Kapulonov T, Lisker N (1997) CPPU and GA3 effects on pre- and post-harvest quality of seedless and seeded grapes. In: 8th International symposium on plant bioregulators in fruit production. Acta Hort 463:349–357
Birnbaum K, Shasha DE, Wang JY, Jung JW, Lambert GM, Galbraith DW, Benfey PN (2003) A gene expression map of the Arabidopsis root. Science 302:1956–1960
Brandstatter I, Kieber JJ (1998) Two genes with similarity to bacterial response regulators are rapidly and specifically induced by cytokinin in Arabidopsis. Plant Cell 10:1009–1020
Brzobohaty B, Moore I, Kristoffersen P, Bako L, Campos N, Schell J, Palme K (1993) Release of active cytokinin by a beta-glucosidase localized to the maize root-meristem. Science 262:1051–1054
Caesar K, Thamm AMK, Witthoft J, Elgass K, Huppenberger P, Grefen C, Horak J, Harte K (2011) Evidence for the localization of the Arabidopsis cytokinin receptors AHK3 and AHK4 in the endoplasmic reticulum. J Exp Bot 62:5571–5580
Casati S, Ottria R, Baldoli E, Lopez E, Maier JAM, Ciuffreda P (2011) Effects of cytokinins, cytokinin ribosides and their analogs on the viability of normal and neoplastic human cells. Anticancer Res 31:3401–3406
Chernyadev II (2009) The protective action of cytokinins on the photosynthetic machinery and productivity of plants under stress. Appl Biochem Microbiol 45:351–362
Choi J, Huh SU, Kojima M, Sakakibara H, Paek KH, Hwang I (2010) The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev Cell 19:284–295
Crespi M, Vereecke D, Temmerman W, Van Montagu M, Desomer J (1994) The fas operon of Rhodococcus fascians encodes new genes required for efficient fasciation of host plants. J Bacteriol 176:2492–2501
D’Agostino IB, Deruere J, Kieber JJ (2000) Characterization of the response of the Arabidopsis response regulator gene family to cytokinin. Plant Physiol 124:1706–1717
Danaee E, Mostofi Y, Moradi P (2011) Effect of GA(3) and BA on postharvest quality and vase life of Gerbera (Gerbera jamesonii. cv. Good Timing) cut flowers. Hort Environ Biotechnol 52:140–144
Dello Ioio R, Linhares FS, Scacchi E, Casamitjana-Martinez E, Heidstra R, Costantino P, Sabatini S (2007) Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation. Curr Biol 17:678–682
Dello Ioio R, Nakamura K, Moubayidin L, Perilli S, Taniguchi M, Morita MT, Aoyama T, Costantino P, Sabatini S (2008) A genetic framework for the control of cell division and differentiation in the root meristem. Science 322:1380–1384
Dobra J, Motyka V, Dobrev P, Malbeck J, Prasil IT, Haisel D, Gaudinova A, Havlova M, Gubis J, Vankova R (2010) Comparison of hormonal responses to heat, drought and combined stress in tobacco plants with elevated proline content. J Plant Physiol 167:1360–1370
Dortay H, Mehnert N, Burkle L, Schmulling T, Heyl A (2006) Analysis of protein interactions within the cytokinin-signaling pathway of Arabidopsis thaliana. FEBS J 273:4631–4644
Droog FN, Taller BJ, Stevens SE (1997) Isolation of cytokinin biosynthesis genes from cyanobacteria. Plant Physiol 114:791
Einset JW, Swaminathan S, Skoog F (1976) Ribosyl-cis-zeatin in a leucyl transfer-RNA species from peas. Plant Physiol 58:140–142
Entsch B, Letham DS (1979) Enzymic glycosylation of the cytokinin, 6-benzylaminopurine. Plant Sci Lett 14:205–212
Entsch B, Parker CW, Letham DS (1983) An enzyme from lupin seeds forming alanine derivatives of cytokinins. Phytochemistry 22:375–381
Fankhauser C (2002) Light perception in plants: cytokinins and red light join forces to keep phytochrome B active. Trends Plant Sci 7:143–145
Feng J, Wang C, Chen QG, Chen H, Ren B, Li XM, Zuo JR (2013) S-nitrosylation of phosphotransfer proteins represses cytokinin signaling. Nat Commun 4:1529
Ferreira WD, Kerbauy GB, Costa APP (2006) Micropropagation and genetic stability of a Dendrobium hybrid (Orchidaceae). In Vitro Cell Dev Plant 42:568–571
Frebort I, Kowalska M, Hluska T, Frebortova J, Galuszka P (2011) Evolution of cytokinin biosynthesis and degradation. J Exp Bot 62:2431–2452
Gajdosova S, Spichal L, Kaminek M, Hoyerova K, Novak O, Dobrev PI, Galuszka P, Klima P, Gaudinova A, Zizkova E, Hanus J, Dancak M, Travnicek B, Pesek B, Krupicka M, Vankova R, Strnad M, Motyka V (2011) Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J Exp Bot 62:2827–2840
Galuszka P, Frebort I, Sebela M, Sauer P, Jacobsen S, Pec P (2001) Cytokinin oxidase or dehydrogenase? Mechanism of cytokinin degradation in cereals. Eur J Biochem 268:450–461
Gan S, Amasino RM (1995) Inhibition of leaf senescence by autoregulated production of cytokinin. Science 270:1986–1988
Gapper NE, Coupe SA, McKenzie MJ, Sinclair BK, Lill RE, Jameson PE (2005) Regulation of harvest-induced senescence in broccoli (Brassica oleracea var. italica) by cytokinin, ethylene, and sucrose. J Plant Growth Regul 24:153–165
Gattolin S, Alandete-Saez M, Elliott K, Gonzalez-Carranza Z, Naomab E, Powell C, Roberts JA (2006) Spatial and temporal expression of the response regulators ARR22 and ARR24 in Arabidopsis thaliana. J Exp Bot 57:4225–4233
Ghanem ME, Albacete A, Smigocki AC, Frebort I, Pospisilová H, Martinez-Andujar C, Acosta M, Sanchez-Bravo J, Lutts S, Dodd IC, Perez-Alfocea F (2011) Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L.) plants. J Exp Bot 62:125–140
Gonzalez-Rizzo S, Crespi M, Frugier F (2006) The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 18:2680–2693
Grefen C, Harter K (2004) Plant two-component systems: principles, functions, complexity and cross talk. Planta 219:733–742
Gregersen PP, Culetic A, Boschian L, Krupinska K (2013) Plant senescence and crop productivity. Plant Mol Biol 82:603–622
Gruhn N, Heyl A (2013) Updates on the model and the evolution of cytokinin signaling. Curr Opin Plant Biol 16:1–6
Gupta S, Rashotte AM (2012) Down-stream components of cytokinin signaling and the role of cytokinin throughout the plant. Plant Cell Rep 31:801–812
Ha S, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Tran LSP (2012) Cytokinins: metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci 17:172–179
Hare PD, Cress WA, van Staden J (1997) The involvement of cytokinins in plant responses to environmental stress. Plant Growth Regul 23:79–103
Hass C, Lohrmann J, Albrecht V, Sweere U, Hummel F, Yoo SD, Hwang I, Zhu T, Schafer E, Kudla J, Harter K (2004) The response regulator 2 mediates ethylene signalling and hormone signal integration in Arabidopsis. EMBO J 23:3290–3302
Hejatko J, Ryu H, Kim GT, Dobesova R, Choi S, Choi SM, Soucek P, Horak J, Pekarova B, Palme K, Brzobohaty B, Hwang I (2009) The histidine kinases cytokinin-independent1 and Arabidopsis histidine kinase 2 and 3 regulate vascular tissue development in Arabidopsis shoots. Plant Cell 21:2008–2021
Heyl A, Schmulling T (2003) Cytokinin signal perception and transduction. Curr Opin Plant Biol 6:480–488
Heyl A, Wulfetange K, Pils B, Nielsen N, Romanov GA, Schmulling T (2007) Evolutionary proteomics identifies amino acids essential for ligand-binding of the cytokinin receptor CHASE domain. BMC Evol Biol 7:62
Heyl A, Brault M, Frugier F, Kuderova A, Lindner AC, Motyka V, von Schwartzenberg K, Vankova R, Schaller GE (2013) Nomenclature for members of the two-component signaling pathway of plants. Plant Physiol 161:1063–1065
Higuchi M, Pischke MS, Mahonen AP, Miyawaki K, Hashimoto Y, Seki M, Kobayashi M, Shinozaki K, Kato T, Tabata S, Helariutta Y, Sussman MR, Kakimoto T (2004) In planta functions of the Arabidopsis cytokinin receptor family. Proc Natl Acad Sci U S A 101:8821–8826
Hill K, Mathews DE, Kim HJ, Street IH, Wildes SL, Chiang YH, Mason MG, Alonso JM, Ecker JR, Kieber JJ, Schaller GE (2013) Functional characterization of type-B response regulators in the Arabidopsis cytokinin response. Plant Physiol 162:212–224
Horak J, Grefen C, Berendzen KW, Hahn A, Stierhof YD, Stadelhofer B, Stahl M, Koncz C, Harter K (2008) The Arabidopsis thaliana response regulator ARR22 is a putative AHP phospho-histidine phosphatase expressed in the chalaza of developing seeds. BMC Plant Biol 8:77
Hosoda K, Imamura A, Katoh E, Hatta T, Tachiki M, Yamada H, Mizuno T, Yamazaki T (2002) Molecular structure of the GARP family of plant Myb-related DNA binding motifs of the Arabidopsis response regulators. Plant Cell 14:2015–2029
Hothorn M, Dabi T, Chory J (2011) Structural basis for cytokinin recognition by Arabidopsis thaliana histidine kinase 4. Nat Chem Biol 7:766–768
Houba-Hérin N, Pethe C, d’Alayer J, Laloue M (1999) Cytokinin oxidase from Zea mays: purification, cDNA cloning and expression in moss protoplasts. Plant J 17:615–626
Hradilova J, Brzobohaty B (2007) Expression pattern of the AHP gene family from Arabidopsis thaliana and organ specific alternative splicing in the AHP5 gene. Biol Plant 51:257–267
Hu YL, Jia WL, Wang JD, Zhang YQ, Yang LL, Lin ZP (2005) Transgenic tall fescue containing the Agrobacterium tumefaciens ipt gene shows enhanced cold tolerance. Plant Cell Rep 23:705–709
Hutchison CE, Li J, Argueso C, Gonzalez M, Lee E, Lewis MW, Maxwell BB, Perdue TD, Schaller GE, Alonso JM, Ecker JR, Kieber JJ (2006) The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell 18:3073–3087
Huynh LN, VanToai T, Streeter J, Banowetz G (2005) Regulation of flooding tolerance of SAG12: ipt Arabidopsis plants by cytokinin. J Exp Bot 56:1397–1407
Hwang I, Sheen J (2001) Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 413:383–389
Imamura A, Yoshino Y, Mizuno T (2001) Cellular localization of the signaling components of Arabidopsis His-to-Asp phosphorelay. Biosci Biotechnol Biochem 65:2113–2117
Imamura A, Kiba T, Tajima Y, Yamashino T, Mizuno T (2003) In vivo and in vitro characterization of the ARR11 response regulator implicated in the His-to-Asp phosphorelay signal transduction in Arabidopsis thaliana. Plant Cell Physiol 44:122–131
Inoue T, Higuchi M, Seki M, Hashimoto Y, Kobayashi M, Kato T, Tabata S, Shinozaki K, Kakimoto T (2001) Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 409:1060–1063
Jeon J, Kim NY, Kim S, Kang NY, Novak O, Ku SJ, Cho C, Lee DJ, Lee EJ, Strnad M, Kim J (2010) A subset of cytokinin two component signalling system plays a role in cold temperature stress response in Arabidopsis. J Biol Chem 285:23371–23386
Jung KW, Oh SI, Kim YY, Yoo KS, Cui MH, Shin JS (2008) Arabidopsis histidine-containing phosphotransfer factor 4 (AHP4) negatively regulates secondary wall thickening of the anther endothecium during flowering. Mol Cells 25:294–300
Kakimoto T (2001) Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases. Plant Cell Physiol 42:677–685
Kamada-Nobusada T, Sakakibara H (2009) Molecular basis for cytokinin synthesis. Phytochemistry 70:444–449
Kaminek M, Vanek T, Kalendova-Kulasova A, Pilar J (1987) The effect of two cytokinins on production of stem cuttings by stock plants of Euphorbia pulcherrima Willd. and Gerbera jamesonii Hook. Sci Hortic-Amsterdam 33:281–289
Kasahara H, Takei K, Ueda N, Hishiyama S, Yamaya T, Kamiya Y, Yamaguchi S, Sakakibara H (2004) Distinct isoprenoid origins of cis- and trans-zeatin biosyntheses in Arabidopsis. J Biol Chem 279:14049–14054
Khodakovskaya M, Vankova R, Malbeck J, Li A, Li Y, McAvoy R (2009) Enhancement of flowering and branching phenotype in chrysanthemum by expression of ipt under the control of a 0.821 kb fragment of the LEACO1 gene promoter. Plant Cell Rep 28:1351–1362
Kiba T, Aoki K, Sakakibara H, Mizuno T (2004) Arabidopsis response regulator, ARR22, ectopic expression of which results in phenotypes similar to the wol cytokinin-receptor mutant. Plant Cell Physiol 45:1063–1077
Kiba T, Kudo T, Kojima M, Sakakibara H (2011) Hormonal control of nitrogen acquisition: roles of auxin, abscisic acid, and cytokinin. J Exp Bot 62:1399–1409
Kim HJ, Ryu H, Hong SH, Woo HR, Lim PO, Lee IC, Sheen J, Nam HG, Hwang I (2006) Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis. Proc Natl Acad Sci U S A 103:814–819
Kim K, Ryu H, Cho YH, Scacchi E, Sabatini S, Hwang I (2012) Cytokinin facilitated proteolysis of Arabidopsis response regulator 2 attenuates signaling output in two-component circuitry. Plant J 69:934–945
Kim HJ, Chianga YH, Joseph J, Kieber JJ, Schaller GE (2013) SCFKMD controls cytokinin signaling by regulating the degradation of type-B response regulators. Proc Natl Acad Sci U S A 110:10028–10033
Kosova K, Prasil IT, Vitamvas P, Dobrev P, Motyka V, Flokova K, Novak O, Tureckova V, Rolcik J, Pesek B, Travnickova A, Gaudinova A, Galiba G, Janda T, Vlasakova E, Prasilová P, Vankova R (2012) Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J Plant Physiol 169:567–576
Kuppu S, Mishra N, Hu RB, Sun L, Zhu XL, Shen GX, Blumwald E, Payton P, Zhang H (2013) Water-deficit inducible expression of a cytokinin biosynthetic gene IPT improves drought tolerance in cotton. PLoS One 8:e64190
Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, Nagato Y, Sakakibara H, Kyozuka J (2007) Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature 445:652–655
Lee DJ, Park JY, Ku SJ, Ha YM, Kim S, Kim MD, Oh MH, Kim J (2007) Genome-wide expression profiling of Arabidopsis response regulator (ARR7) overexpression in cytokinin response. Mol Genet Genomics 277:115–137
Leibfried A, To JP, Busch W (2005) WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 438:1172–1175
Letham DS (1963) Zeatin, a factor inducing cell division isolated from Zea mays. Life Sci 8:569–573
Liu L, Zhou Y, Szczerba MW, Xianghua L, Yongjun L (2010) Identification and application of a rice senescence-associated promoter. Plant Physiol 153:1239–1249
Liu WZ, Kong DD, Gu XX, Gao HB, Wang JZ, Xia M, Gao Q, Tian LL, Xu ZH, Bao F, Hu Y, Ye NS, Pei ZM, He YK (2013) Cytokinins can act as suppressors of nitric oxide in Arabidopsis. Proc Natl Acad Sci U S A 110:1548–1553
Lomin SN, Yonekura-Sakakibara K, Romanov GA, Sakakibara H (2011) Ligand-binding properties and subcellular localization of maize cytokinin receptors. J Exp Bot 62:5149
Ma QH, Wang XM, Wang ZM (2008) Expression of isopentenyl transferase gene controlled by seed-specific lectin promoter in transgenic tobacco influences seed development. J Plant Growth Regul 27:68–76
Mackova H, Hronkova M, Dobra J, Tureckova V, Novak O, Lubovska Z, Motyka V, Haisel D, Hajek T, Prasil IT, Gaudinova A, Storchova H, Werner T, Schmulling T, Vankova R (2013) Enhanced drought and heat stress tolerance of tobacco plants with ectopically enhanced cytokinin oxidase/dehydrogenase gene expression. J Exp Bot 54:2805–2815
Mahonen AP, Bonke M, Kauppinen L, Marjukka R, Benfey PN, Helariutta Y (2000) A novel two-component hybrid molecule regulates vascular morphogenesis of the Arabidopsis root. Genes Dev 14:2938–2943
Mahonen AP, Bishopp A, Higuchi M, Nieminen KM, Kinoshita K, Tormakangas K, Ikeda Y, Oka A, Kakimoto T, Helariutta Y (2006a) Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311:94–98
Mahonen AP, Higuchi M, Tormakangas K, Miyawaki K, Pischke MS, Sussman MR, Helariutta Y, Kakimoto T (2006b) Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis. Curr Biol 16:1116–1122
Mala J, Machova P, Cvrckova H, Karady M, Novak O, Mikulik J, Dostal J, Strnad M, Dolezal K (2013) The role of cytokinins during micropropagation of wych elm. Biol Plant 57:174–178
Malabug LU, Cruz PCS, Banayo NPMC, Aguilar EA, Hernandez JE (2010) Improving the grain filling and yield of Indica rice through kinetin (N-6-furfuryl adenine) application at flowering stage. Philipp J Crop Sci 35:22–35
Martin RC, Mok MC, Mok DWS (1999a) Isolation of a cytokinin gene, ZOG1, encoding zeatin O-glucosyltransferase from Phaseolus lunatus. Proc Natl Acad Sci U S A 96:284–289
Martin RC, Mok MC, Mok DWS (1999b) A gene encoding the cytokinin enzyme zeatin O-xylosyltransferase of Phaseolus vulgaris. Plant Physiol 120:553–558
Martin RC, Mok MC, Habben JE, Mok DWS (2001) A maize cytokinin gene encoding an O-glucosyltransferase specific to cis-zeatin. Proc Natl Acad Sci U S A 98:5922–5926
Mason MG, Li J, Mathews DE, Kieber JJ, Schaller GE (2004) Type-B response regulators display overlapping expression patterns in Arabidopsis. Plant Physiol 135:927–937
Mason MG, Jha D, Salt DE, Tester M, Hill K, Kieber JJ, Schaller GE (2010) Type-B response regulators ARR1 and ARR12 regulate expression of AtHKT1;1 and accumulation of sodium in Arabidopsis shoots. Plant J 64:753–763
McCabe MS, Garratt LC, Schepers F, Jordi WJRM, Stoopen GM, Davelaar E, van Rhijn JHA, Power JB, Davey MR (2001) Effects of PSAG12-IPT gene expression on development and senescence in transgenic lettuce. Plant Physiol 127:505–516
Merewitz E, Gianfagna T, Huang B (2010) Effects of SAG12-ipt and HSP18.2-ipt expression on cytokinin production, root growth and leaf senescence in creeping bentgrass exposed to drought stress. J Am Soc Hortic Sci 135:230–239
Merewitz EB, Du HM, Yu WJ, Liu YM, Gianfagna T, Huang BR (2012) Elevated cytokinin content in ipt transgenic creeping bentgrass promotes drought tolerance through regulating metabolite accumulation. J Exp Bot 63:1315–1328
Miller CO, Skoog F, von Saltza MH, Strong FM (1955) Kinetin, a cell division factor from deoxyribonucleic acid. J Am Chem Soc 77:1392
Mlejnek P, Prochazka S (2002) Activation of caspase-like proteases and induction of apoptosis by isopentenyladenosine in tobacco BY-2 cells. Planta 215:158–166
Moglich A, Ayers RA, Moffat K (2009) Structure and signaling mechanism of Per-ARNT-Sim domains. Structure 17:1282–1294
Mok DWS, Mok MC (2001) Cytokinin metabolism and action. Annu Rev Plant Physiol Plant Mol Biol 89:89–118
Mok MC, Martin RC, Dobrev PI, Vankova R, Shing Ho P, Yonekura-Sakakibara K, Sakakibara H, Mok DWS (2005) Topolins and hydroxylated thidiazuron derivatives are substrates of cytokinin O-glucosyltransferase with position specificity related to receptor recognition. Plant Physiol 137:1057–1066
Molinsky J, Klanova M, Koc M, Beranova L, Andera L, Ludvikova Z, Bohmova M, Gasova Z, Strnad M, Ivanek R, Trneny M, Necas E, Zivny J, Klener P (2013) Roscovitine sensitizes leukemia and lymphoma cells to tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis. Leuk Lymphoma 54:372–380
Momenpour A, Taghavi TS, Manochehr S (2011) Effects of benzyladenine and gibberellin on runner production and some vegetative traits of three strawberry cultivars. Afr J Agric Res 6:4357–4361
Moreira S, Bishopp A, Carvalho H, Campilho A (2013) AHP6 inhibits cytokinin signaling to regulate the orientation of pericycle cell division during lateral root initiation. PLoS One 8:e56370
Motte H, Galuszka P, Spichal L, Tarkowski P, Plihal O, Smehilova M, Jaworek P, Vereecke D, Werbrouck S, Geelen D (2013) Phenyl-adenine, identified in a LIGHT-DEPENDENT SHORT HYPOCOTYLS4-assisted chemical screen, is a potent compound for shoot regeneration through the inhibition of cytokinin oxidase dehydrogenase activity. Plant Physiol 161:1229–1241
Mougel C, Zhulin IB (2001) CHASE: an extracellular sensing domain common to transmembrane receptors from prokaryotes, lower eukaryotes and plants. Trends Biochem Sci 26:582–584
Muller B, Sheen J (2007) Advances in cytokinin signaling. Science 318:68–69
Muller B, Sheen J (2008) Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature 453:1094–1097
Mytinova Z, Motyka V, Haisel D, Gaudinova A, Lubovska Z, Wilhelmova N (2010) Effect of abiotic stresses on the activity of antioxidative enzymes and contents of phytohormones in wild type and AtCKX2 transgenic tobacco plants. Biol Plant 54:461–470
Nishiyama R, Watanabe Y, Fujita Y, Le DT, Kojima M, Werner T, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Kakimoto T, Sakakibara H, Schmulling T, Tran LSP (2011) Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 23:2169–2183
Nishiyama R, Watanabe Y, Leyva-Gonzalez MA, Van Ha C, Fujita Y, Tanaka M, Seki M, Yamaguchi-Shinozaki K, Shinozaki K, Herrera-Estrella L, Tran LSP (2013) Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. Proc Natl Acad Sci U S A 110:4840–4845
Ordog V, Stirk WA, van Staden J, Novak O, Strnad M (2004) Endogenous cytokinins in three genera of microalgae from the Chlorophyta. J Phycol 40:88–95
Paces V, Werstiuk E, Hall RH (1971) Conversion of N6-(∆2-isopentenyl)adenosine to adenosine by enzyme activity in tobacco tissue. Plant Physiol 48:775–778
Pavlikova D, Neuberg M, Zizkova E, Motyka V, Pavlik M (2012) Interactions between nitrogen nutrition and phytohormone levels in Festulolium plants. Plant Soil Environ 58:367–372
Peleg Z, Reguera M, Tumimbang E, Walia H, Blumwald E (2011) Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol J 9:747–758
Pertry I, Vaclavikova K, Depuydt S, Galuszka P, Spichal L, Temmerman W, Stes E, Schmulling T, Kakimoto T, Van Montagu MCE, Strnad M, Holsters M, Tarkowski P, Vereecke D (2009) Identification of Rhodococcus fascians cytokinins and their modus operandi to reshape the plant. Proc Natl Acad Sci U S A 106:929–934
Punwani JA, Hutchison CE, Schaller GE, Kieber JJ (2010) The subcellular distribution of the Arabidopsis histidine phosphotransfer proteins is independent of cytokinin signaling. Plant J 62:473–482
Qin H, Gu Q, Zhang JL, Sun L, Kuppu S, Zhang YZ, Burow M, Payton P, Blumwald E, Zhang H (2011) Regulated expression of an isopentenyltransferase gene (IPT) in peanut significantly improves drought tolerance and increases yield under field conditions. Plant Cell Physiol 52:1904–1914
Qiu WM, Liu MY, Qiao GR, Jiang J, Xie LH, Zhuo RY (2012) An isopentyl transferase gene driven by the stress-inducible rd29A promoter improves salinity stress tolerance in transgenic tobacco. Plant Mol Biol Rep 30:519–528
Ramireddy E, Brenner WG, Pfeifer A, Heyl A, Schmulling T (2013) In planta analysis of a cis-regulatory cytokinin response motif in Arabidopsis and identification of a novel enhancer sequence. Plant Cell Physiol 54:1079–1092
Rashotte AM, Goertzen LR (2010) The CRF domain defines cytokinin response factor proteins in plants. BMC Plant Biol 10:74
Rashotte AM, Carson SD, To JPC, Kieber JJ (2003) Expression profiling of cytokinin action in Arabidopsis. Plant Physiol 132:1998–2011
Rashotte AM, Mason MG, Hutchison CE, Ferreira FJ, Schaller GE, Kieber JJ (2006) A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two component pathway. Proc Natl Acad Sci U S A 103:11081–11085
Ren B, Liang Y, Deng Y, Chen Q, Zhang J, Yang X, Zuo J (2009) Genome-wide comparative analysis of type-A Arabidopsis response regulator genes by overexpression studies reveals their diverse roles and regulatory mechanisms in cytokinin signaling. Cell Res 19:1178–1190
Riefler M, Novak O, Strnad M, Schmulling T (2006) Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 18:40–54
Rivero RM, Kojima M, Gepstein A, Sakakibara H, Mittler R, Gepstein S, Blumwald E (2007) Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc Natl Acad Sci U S A 104:19631–19636
Rivero RM, Shulaev V, Blumwald E (2009) Cytokinin-dependent photorespiration and the protection of photosynthesis during water deficit. Plant Physiol 150:1530–1540
Rivero RM, Gimeno J, Van Deynze A, Walia H, Blumwald E (2010) Enhanced cytokinin synthesis in tobacco plants expressing P-SARK::IPT prevents the degradation of photosynthetic protein complexes during drought. Plant Cell Physiol 51:1929–1941
Robson PRH, Donnison IS, Wang K, Frame B, Pegg SE, Thomas A, Thomas H (2004) Leaf senescence is delayed in maize expressing the Agrobacterium IPT gene under the control of a novel maize senescence-enhanced promoter. Plant Biotechnol J 2:101–112
Ruffel S, Krouk RD, Shasha D, Birnbaum KD, Coruzzi GM (2011) Nitrogen economics of root foraging: transitive closure of the nitrate-cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc Natl Acad Sci U S A 108:18524–18529
Ruszkowski M, Brzezinski K, Jedrzejczak R, Dauter M, Dauter Z, Sikorski M, Jaskolski M (2013) Medicago truncatula histidine-containing phosphotransfer protein: structural and biochemical insights into the cytokinin transduction pathway in plants. FEBS J 280:3709–3720
Sakai H, Aoyama T, Oka A (2000) Arabidopsis ARR1 and ARR2 response regulators operate as transcriptional activators. Plant J 24:703–711
Sakai H, Honma T, Aoyama T, Sato S, Kato T, Tabata S, Oka A (2001) ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 294:1519–1521
Sakakibara H (2006) Cytokinins: activity, biosynthesis, and translocation. Annu Rev Plant Biol 57:431–449
Sakakibara H, Hayakawa A, Deji A, Gawronski SW, Sugiyama T (1999) His-Asp phosphotransfer possibly involved in the nitrogen signal transduction mediated by cytokinin in maize: molecular cloning of cDNAs for two-component regulatory factors and demonstration of phosphotransfer activity in vitro. Plant Mol Biol 41:563–573
Salome PA, To JPC, Kieber JJ, McClung CR (2006) Arabidopsis response regulators ARR3 and ARR4 play cytokinin-independent roles in the control of circadian period. Plant Cell 18:55–69
Schaller GE, Shiu SH, Armitage JP (2011) Two-component systems and their co-option for eukaryotic signal transduction. Curr Biol 21:R320–R330
Shi X, Rashotte AM (2012) Advances in upstream players of cytokinin phosphorelay: receptors and histidine phosphotransfer proteins. Plant Cell Rep 31:789–799
Skylar A, Hong FX, Chory J, Weigel D, Wu XL (2010) STIMPY mediates cytokinin signaling during shoot meristem establishment in Arabidopsis seedlings. Development 137:541–549
Souza ER, Pereira MD, Santos LD, Ribeiro VG, Pionorio JAD, De Araujo EA (2010) Quality of grapes “Superior Seedless” with benzyladenine combined with applications or not of gibberellic acid. Rev Caatinga 23:144–148
Spichal L (2012) Cytokinins—recent news and views of evolutionally old molecules. Funct Plant Biol 39:267–284
Spichal L, Rakova NY, Riefler M, Mizuno T, Romanov GA, Strnad M, Schmulling T (2004) Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol 45:1299–1305
Steklov MY, Lomin SN, Osolodkin DI, Romanov GA (2013) Structural basis for cytokinin receptor signaling: an evolutionary approach. Plant Cell Rep 32:781–793
Stolz A, Riefler M, Lomin SN, Achazi K, Romanov GA, Schmulling T (2011) The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J 67:157–168
Strnad M (1997) The aromatic cytokinins. Physiol Plant 101:674–688
Sugawara H, Kawano Y, Hatakeyama T, Yamaya T, Kamiya N, Sakakibara H (2005) Crystal structure of the histidine-containing phosphotransfer protein ZmHP2 from maize. Protein Sci 14:202–208
Suzuki T, Imamura A, Ueguchi C, Mizuno T (1998) Histidine containing phosphotransfer (HPt) signal transducers implicated in His-to-Asp phosphorelay in Arabidopsis. Plant Cell Physiol 39:1258–1268
Suzuki T, Sakurai K, Imamura A, Nakamura A, Ueguchi C, Mizuno T (2000) Compilation and characterization of histidine-containing phosphotransmitters implicated in His-to-Asp phosphorelay in plants: AHP signal transducers of Arabidopsis thaliana. Biosci Biotechnol Biochem 6:2486–2489
Suzuki T, Miwa K, Ishikawa K, Yamada H, Aiba H, Mizuno T (2001) The Arabidopsis sensor His-kinase, AHK4, can respond to cytokinins. Plant Cell Physiol 42:107–113
Sweere U, Eichenberg K, Lohrmann J, Mira-Rodado V, Baurle I, Kudla J, Nagy F, Schafer E, Harter K (2001) Interaction of the response regulator ARR4 with phytochrome B in modulating red light signaling. Science 294:1108–1111
Sykorova B, Kuresova G, Daskalova S, Trckova M, Hoyerova K, Raimanova I, Motyka V, Travnickova A, Elliott MC, Kaminek M (2008) Senescence-induced ectopic expression of the A. tumefaciens ipt gene in wheat delays leaf senescence, increases cytokinin content, nitrate influx, and nitrate reductase activity, but does not affect grain yield. J Exp Bot 59:377–387
Synkova H, Van Loven K, Pospisilova J, Valcke R (1999) Photosynthesis of transgenic pssu-ipt tobacco. J Plant Physiol 155:173–182
Synkova H, Semoradova S, Schnablova R, Muller K, Pospisilova J, Ryslava H, Malbeck J, Cerovska N (2006) Effects of biotic stress caused by Potato virus Y on photosynthesis in ipt transgenic and control Nicotiana tabacum L. Plant Sci 171:607–616
Tajima Y, Imamura A, Kiba T, Amano Y, Yamashino T, Mizuno T (2004) Comparative studies on the type-B response regulators revealing their distinctive properties in the His-to-Asp phosphorelay signal transduction of Arabidopsis thaliana. Plant Cell Physiol 45:28–39
Takei K, Sakakibara H, Sugiyama T (2001a) Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J Biol Chem 276:26405–26410
Takei K, Sakakibara H, Taniguchi M, Sugiyama T (2001b) Nitrogen-dependent accumulation of cytokinins in root and the translocation to leaf: implication of cytokinin species that induces gene expression of maize response regulator. Plant Cell Physiol 42:85–93
Takei K, Yamaya T, Sakakibara H (2004) Arabidopsis CYP735A1 and CYP735A2 encode cytokinin hydroxylases that catalyze the biosynthesis of trans-zeatin. J Biol Chem 279:41866–41872
Tirichine L, Sandal N, Madsen LH, Radutoiu S, Albrektsen AS, Sato S, Asamizu E, Tabata S, Stougaard J (2007) A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 315:104–107
To JP, Kieber JJ (2008) Cytokinin signaling: two-component and more. Trends Plant Sci 13:85–92
To JPC, Haberer G, Ferreira FJ, Deruere J, Mason MG, Schaller GE, Alonso JM, Ecker JR, Kieber JJ (2004) Type-A Arabidopsis response regulators are partially redundant negative regulators of cytokinin signaling. Plant Cell 16:658–671
To JPC, Deruere J, Maxwell BB, Morris VF, Hutchison CE, Ferreira FJ, Schaller GE, Kieber JJ (2007) Cytokinin regulates type-A Arabidopsis response regulator activity and protein stability via two-component phosphorelay. Plant Cell 19:3901–3914
Tran LSP, Urao T, Qin F, Maruyama K, Kakimoto T, Shinozaki K, Yamaguchi-Shinozaki K (2007) Functional analysis of AHK1/AtHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought and salt stress in Arabidopsis. Proc Natl Acad Sci U S A 104:20623–20628
Tran LS, Shinozaki K, Yamaguchi-Shinozaki K (2010) Role of cytokinin responsive two-component system in ABA and osmotic stress signalings. Plant Signal Behav 5:148–150
Tuberosa R, Sanguineti MC, Landi P, Giuliani M, Salvi S, Conti S (2002) Identification of QTLs for root characteristics in maize grown in hydroponics and analysis of their overlap with QTLs for grain yield in the field at two water regimes. Plant Mol Biol 48:697–712
Ueguchi C, Koizumi H, Suzuki T, Mizuno T (2001) Novel family of sensor histidine kinase genes in Arabidopsis thaliana. Plant Cell Physiol 42:231–235
Vermeulen K, Strnad M, Krystof V, Havlicek L, Van der Aa A, Lenjou M, Nijs G, Rodrigus I, Stockman B, van Onckelen H, Van Bockstaele DR, Berneman ZN (2002) Antiproliferative effect of plant cytokinin analogues with an inhibitory activity on cyclin-dependent kinases. Leukemia 16:299–305
Vesely J, Havlicek L, Strnad M, Blow JJ, Donelladeana A, Pinna L, Letham DS, Kato J, Detivaud L, Leclerc S, Meijer L (1994) Inhibition of cyclin-dependent kinases by purine analogs. Eur J Biochem 224:771–786
von Schwartzenberg K, Nunez MF, Blaschke H, Dobrev PI, Novak O, Motyka V, Strnad M (2007) Cytokinins in the bryophyte Physcomitrella patens: analyses of activity, distribution, and cytokinin oxidase/dehydrogenase overexpression reveal the role of extracellular cytokinins. Plant Physiol 145:786–800
Werbrouck SPO, Strnad M, VanOnckelen HA, Debergh PC (1996) meta-Topolin, an alternative to benzyladenine in tissue culture? Physiol Plant 98:291–297
Werner T, Motyka V, Strnad M, Schmulling T (2001) Regulation of plant growth by cytokinin. Proc Natl Acad Sci U S A 98:10487–10492
Werner T, Nehnevajova E, Kollmer I, Novak O, Strnad M, Kramer U, Schmulling T (2010) Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 22:3905–3920
West AH, Stock AM (2001) Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem Sci 26:369–376
Whitty CD, Hall RH (1974) A cytokinin oxidase in Zea mays. Can J Biochem 52:787–799
Wulfetange K, Lomin SN, Romanov GA, Stolz A, Heyl A, Schmulling T (2011) The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum. Plant Physiol 156:1808–1818
Xing JP, Xu Y, Tian JA, Gianfanga T, Huang BR (2009) Suppression of shade- or heat-induced leaf senescence in creeping bentgrass through transformation with the ipt gene for cytokinin synthesis. J Am Soc Hortic Sci 134:602–609
Yamada H, Suzuki T, Terada K, Takei K, Ishikawa K, Miwa K, Yamashino T, Mizuno T (2001) The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane. Plant Cell Physiol 42:1017–1023
Zabadal TJ, Bukovac MJ (2006) Effect of CPPU on fruit development of selected seedless and seeded grape cultivars. HortScience 41:154–157
Zalabak D, Pospisilova H, Smehilova M, Mrizova K, Frebort I, Galuszka P (2013) Genetic engineering of cytokinin metabolism: prospective way to improve agricultural traits of crop plants. Biotechnol Adv 31:97–117
Zalewski W, Galuszka P, Gasparis S, Orczyk W, Nadolska-Orczyk A (2010) Silencing of the HvCKX1 gene decreases the cytokinin oxidase/dehydrogenase level in barley and leads to higher plant productivity. J Exp Bot 61:1839–1851
Zatloukal M, Gemrotová M, Dolezal K, Havlicek L, Spichal L, Strnad M (2008) Novel potent inhibitors of A. thaliana cytokinin oxidase/dehydrogenase. Bioorg Med Chem 16:9268–9275
Zhang P, Wang WQ, Zhang GL, Kaminek M, Dobrev P, Xu J, Gruissem W (2010) Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J Integr Plant Biol 52:653–669
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Vankova, R. (2014). Cytokinin Regulation of Plant Growth and Stress Responses. In: Tran, LS., Pal, S. (eds) Phytohormones: A Window to Metabolism, Signaling and Biotechnological Applications. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-0491-4_3
Download citation
DOI: https://doi.org/10.1007/978-1-4939-0491-4_3
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-0490-7
Online ISBN: 978-1-4939-0491-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)