
Abstract
Over the last 25 years, with the concerted effort of many researchers, we have learned a lot about the pathophysiology of celiac disease (CD). Now, CD is the best understood autoimmune/immune-mediated illness. This chapter will discuss current understanding of gluten as an antigen, antigen-presentation, and the role of HLA-DQ2/DQ8, how gluten peptides cross the intestinal epithelial cell barrier, the importance of tissue transglutaminase for modifying gluten peptides for presentation, the role of intestinal T cells in coordinating inflammation, how intraepithelial lymphocytes are triggered to attack intestinal epithelial cells, and the role of antibody response.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Celiac Disease
- Lamina Propria
- Intestinal Epithelial Cell
- Intestinal Inflammation
- Refractory Celiac Disease
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
There are more than 80 recognized autoimmune and immune-mediated inflammatory diseases. Celiac disease (CD) is unique because it shares features of an autoimmune disease (production of self-reactive antibodies) and an inappropriate immune response to an external provocateur. Furthermore, the pathogenesis of CD is better understood than that of any other immune-mediated disease. CD results from a series of cause and effect actions, the first of which is ingestion of gluten.
Gluten as an Antigen
The term “gluten” refers to a broad mixture of storage proteins present in wheat that provide elasticity to dough. Although originally referring to wheat, now gluten is used to connote similar proteins from rye and barley that contain protein sequences that elicit an immune response (i.e., antigens). Oats also contain gluten-type proteins (avenins) that have some of the same peptide sequences, but in much lower amounts. Gluten-type prolamin proteins from other grains mostly lack the peptide sequences that serve as antigens triggering inflammation in patients with CD.
Lymphocytes (B and T cells) each go through a process that involves rearranging gene segments to develop a unique receptor that can recognize specific antigens. Because of this feat, they constitute the “adaptive” immune system. Cells that comprise the “innate” immune system must rely on germline encoded receptors to recognize potential pathogens. Receptors on B cells recognize locations (epitopes) on native protein antigens, and when stimulated by that antigen, make antibodies. Receptors on helper T cells recognize epitopes on fragments of a protein that are processed and displayed by an antigen-presenting cell (APC). When stimulated by the antigenic epitope and APC, helper T cells make cytokines that direct the behavior of other cells, adaptive and innate, to mount, maintain, and regulate an immune reaction.
Gluten from wheat, rye, and barley is composed of numerous proteins that contain antigenic epitopes for patients with CD. Gluten proteins were originally separated into groups according to their solubility in water, salt solutions, or alcohol. Gliadins are those components from wheat gluten that are soluble in alcohol. Gliadins are further divided into α, β, γ, and ω subtypes. Most of the research into the T-cell antigenicity of gluten has been focused on α-gliadins [1]. Evaluation of these proteins revealed short sequences rich in proline (P) and glutamine (Q) that elicit responses from T cells isolated from patients with CD. These epitopes have sequences similar to PQPQLPYPQ. A section of α2-gliadin contains a 33 amino acid sequence that is resistant to digestion by human gut and pancreatic enzymes. This sequence contains several epitopes that can stimulate CD patient T cells (Fig. 4.1) [2]. Because this polypeptide is resistant to digestion, it remains available to stimulate the immune system. However, many proteins present in wheat, rye, barley, and oats contain potential CD-stimulating epitopes [3].

Gluten epitopes. Gliadin contains a 33 amino acid section packed with a peptide sequence (PQPQLPYPQ) that can activate T cells from patients with CD. This sequence is also present in other proteins made by wheat, rye, and barley
Gluten Epitopes and Antigen Presentation
Antigen-presenting cells display epitopes to helper T cells on cell surface protein molecules. These antigen-presenting molecules hold epitopes in correct orientation for review by T cells much like picture frames hold paintings for perusal by museum patrons. Any one picture frame can hold different paintings of the right size, but cannot hold paintings of the wrong size. Any one antigen-presenting molecule can hold different epitopes with correctly aligned charge distributions, but cannot hold epitopes with misaligned charges. Therefore, the specific antigen epitopes that can be displayed by a person’s APCs depends on the set of antigen-presenting molecules encoded in that person’s genome within the human leukocyte antigen (HLA) locus on the short arm of chromosome 6. This section of the genome is highly polymorphic, meaning that many different versions of these genes exist. This wide variation permits a range of responses by different members of a population, ensuring that some will be able to react to a new pathogen.
Antigen presenting molecules that display epitopes to helper T cells are made of an α and β chain, each chain encoded by a different gene. Three sets of these “Class II” antigen-presenting genes are encoded on chromosome 6. The sets are named DP, DQ, and DR. However, the α chain from one chromosome (e.g., paternal) can bind with the β chain of the same set from the other chromosome (e.g., maternal), so four different DQ molecules can be produced. The α and β chains that bind gluten epitopes and contribute to development of CD are encoded by the DQA1*0501 and DQB1*0201 genes [4]. People with this combination have the DQ2.5 molecule. If the DQ2.5 molecule is encoded on one chromosome, the person carries a DR3-DQ2 haplotype. If the DQ2.5 molecule is produced by combining an α and β chain from different chromosomes, the person carries the DR5-DQ7/DR7-DQ2 haplotypes.
As further elaborated in Chap. 5 by de Haas et al., about 95 % of patients with CD express DQ2.5. People who inherit a copy of DQB1*0201 from each parent are at higher risk of developing CD than are those with one copy of DQB1*0201 [5]. Thus, people who are homozygous for DQ2.5 have higher risk than those who are heterozygotes. A very similar set of α and β chains that can bind gluten epitopes are DQA1*0201 and DQB1*0202, which compose the DQ2.2 molecule. Major gluten epitope presentation is not as sustained on DQ2.2 as it is with DQ2.5 [6]. However, other similar gluten epitopes can be stably displayed on DQ2.2 molecules [7]. Moreover, DQ2.5/DQ2.2 heterozygotes have a risk that is higher than a regular DQ2.5 heterozygote. About 5 % of patients with CD lack a DQ2.5 haplotype. Most of these express DQ8, which is composed of the DQB1*0302 and DQA1*0301 chains [8]. There are rare patients with biopsy proven CD that does not express either DQ2 or DQ8 [9]. About one-third of the Caucasian population expresses either DQ2 or DQ8 antigen-presenting molecules. Therefore, the vast majority (97 %) of people with DQ2 or DQ8 antigen presenting molecules will not develop celiac disease.
Intestinal Barrier and Antigen Access
In order to induce inflammation, gluten antigens need to cross from the intestinal lumen into the lamina propria where antigen-presenting cells and lymphocytes reside. Intestinal epithelial cells (IEC) form a polarized sheet that acts as a barrier to luminal contents. Macromolecules like gluten epitopes can cross this barrier through four pathways: (1) by transport through the epithelial cell (transcellular passage), (2) by transport between epithelial cells (paracellular passage), (3) by direct antigen-presenting dendritic cell sampling of luminal contents, and (4) through a break in the epithelium due to some sort of injury (Fig. 4.2).

Routes of passage across the epithelial barrier. To initiate inflammation, gluten peptides need to move from the intestinal lumen to the lamina propria, crossing the epithelial cell barrier. Pathways include (1) transcellular passage, (2) paracellular passage, (3) direct sampling by dendritic cells, and (4) passage through an injured area
Intestinal epithelial cells can transport material through the cell from the apical surface to the basal surface. Microfold (M) cells are specialized epithelial cells that transport macromolecules and particulate matter (e.g., bacteria) across the cell to underlaying lymphoid follicles for immunologic evaluation. This pathway permits routine sampling of luminal contents and is upregulated in rodents with nonsteroidal anti-inflammatory drug-induced intestinal inflammation [10]. Dietary antigens also are able to pass through normal columnar intestinal epithelial cells. This pathway likely dominates in active CD [11]. One important mechanism is mediated by gluten binding to IgA antibodies in the lumen that then associate with cell surface transferrin receptor (CD71), triggering endocytosis and passage of gluten through the epithelial cell [12]. Apical expression of CD71 increases in active CD as does production of anti-gliadin IgA, creating a potential for progressively worsening inflammation. Other pathways of gluten antigen transport through epithelial cells also exist [13, 14].
Intestinal epithelial cells are bound to each other by a cellular organelle called the “tight junction” or “zonula occludens,” which controls passage of ions and macromolecules between cells (paracellular pathway). Tight junction transmembrane, structural, and regulatory proteins form a web that holds adjacent cell membranes in close opposition. The complex is composed of about 30 proteins, including junction adhesion molecule (JAM), claudins, VAP-33, zonula occludens proteins (ZO-1, ZO-2, ZO-3), cingulin, occludin, and regulatory proteins [15]. Tight junction function is regulated by cytokines like interferon-γ (IFN-γ) and tumor necrosis factor-α (TNF-α) in a process that requires occludin [16]. Inflammatory cytokines cause the tight junctions to loosen permitting paracellular transport of macromolecules to the lamina propria for immunologic evaluation and, conversely, rapid egress of fluid and bactericidal molecules to “wash out” offending agents. Zonulin (prehaptoglobin-2) is another human protein that opens tight junctions [17]. Zonulin production is upregulated in active CD, and gliadin stimulates zonulin release from epithelial cells. Thus, regulation of tight junctions provides another circuit for progressively worsening inflammation. Larazotide acetate (AT-1001) prevents opening of tight channels in response to cytokines or zonulin and was recently tested in clinical trials to determine if the compound would prevent changes in intestinal permeability upon gluten challenge in patients with celiac disease. Larazotide acetate did not appear to be effective in controlling permeability, but the study may have been complicated by high variability in how individual patients respond to gluten challenge [18].
Dendritic cells are the most prodigious antigen-presenting cells and are scattered in the lamina propria beneath the basement membrane [19]. Dendritic cells can send processes up between and even through epithelial cells to sample luminal contents [20]. Dendritic cell sampling can be upregulated by epithelial cell exposure to potentially pathogenic bacteria like salmonella [21]. Small intestinal dysbiosis also may increase antigen presentation by this mechanism [22]. Furthermore, gliadin peptide fragments alone induce maturation of dendritic cells to augment APC function [23]. Regulation of lamina propria dendritic cell function provides another circuit for progressively worsening intestinal inflammation.
Severe injury or inflammation can kill an area of epithelial cells causing an ulcer. Enteric virus infection or medications like aspirin, ibuprofen, and naproxen frequently cause small bowel ulceration. In young children, active CD can present with duodenal ulceration [24]. In adults, ulceration is rare in active CD and suggests ulcerative jejunitis, refractory CD type 2, or enteropathy-associated T-cell lymphoma, which is on the spectrum of aberrant T-cell neoplasms [25]. However the ulcer forms, the break in the epithelial cell lining permits direct access of luminal contents to lamina propria and submucosal APC. Thus, an ulcer due to injury or inflammation can permit ingested gluten peptides to bathe the lamina propria, prompting a reaction that can worsen the ulcer in patients with CD.
It is not surprising that large gluten peptides can cross the epithelial cell barrier and initiate inflammation. Each of the four pathways (see Fig. 4.2) could cause transient or low-grade display of gluten epitopes to T cells by APC. Each of the four pathways is likely active at any given time. Each of the four pathways will increase with worsening inflammation due to active CD, which can cause increase in the other pathways. It is surprising that food allergies are not more common.
Tissue Transglutaminase: A Matchmaker
Tissue transglutaminase (TG2, TTG) is a ubiquitously expressed cellular enzyme that crosslinks proteins through a lysine-glutamine bridge. It can also serve as a deamidase, removing the amide group on the side chain of glutamine and converting it to glutamate. Tissue transglutaminase can deamidate gluten peptides [26]. For example, PQPQLPYPQ is the sequence derived from native 33mer (see Fig. 4.1). If the residue has been deamidated by TTG, the sequence PQPELPYPQ is produced through degradation [3]. This change alters the charge on the polypeptide, which increases its ability to bind in the antigen-presenting groove of DQ2.5. Gluten serves as an ideal substrate for TTG and is quickly and specifically deamidated by the enzyme. The preferred (Q, underlined) glutamine target for TTG is in the sequence QXPF (Y, W, M, L, I, or V) [27], where X is any amino acid. This sequence set is rare or absent in oat avenins [27]. Glutamine can deamidate to glutamate nonenzymatically, but the conversion is slow. It is likely that TTG is central to the process of rendering gluten epitopes highly antigenic by increasing their binding strength (avidity) to CD-associated antigen-presenting molecules. In addition, TTG may also assist in shuttling gluten epitopes through epithelial cells [14].
Lamina Propria T Cells: Stokers of Intestinal Inflammation
We have now set most of the stage for intestinal inflammation. There is an antigen (gluten) that has crossed the epithelial cell barrier and has been acted on by tissue TTG to increase binding to DQ2.5 or DQ8 antigen-presenting molecules on antigen-presenting cells in the lamina propria and draining lymph node. For inflammation to occur, several cell types need to work in concert, communicating by cell surface receptor display and elaboration of signaling molecules called cytokines or interleukins (IL). Appropriate inflammatory reactions are highly regulated and tightly focused. Inappropriate inflammation, like that which occurs in CD, is a result of either excessive pro-inflammatory or ineffective anti-inflammatory (regulatory) communication. At the center of this communication are the T helper lymphocytes.
T helper (Th) cells respond to antigen epitopes displayed on MHC class II (DP,DQ,DR) molecules by a high affinity engagement of a clonally unique T cell receptor complexed to a co-receptor named CD4. The terms “Th cells” and “CD4+ T cells” are functionally synonymous. There are several types of Th identified by the kind of cytokines they make [28]. Major types include Th1 (makers of IFN-γ), Th2 (makers of IL-4), Th17 (makers of IL-17), and T regulatory cells (makers of IL-10, TGF-β, and/or inhibitory cell surface molecules). Each of these Th cell types makes cytokines that amplify development of that type while inhibiting development of the other types. This causes chronic reactions to polarize into a discrete Th type cytokine profile. The cytokines made by Th cells instruct other cells how to respond within an inflammation. Therefore, the mix of T cell types generated in response to an epitope displayed by an APC will determine whether or not an inflammation results and if so, what type of inflammation occurs. This initial decision likely takes place in the mesenteric draining lymph nodes where intestinal dendritic cells have migrated to display captured antigens [29]. Most of the time, the decision produces tolerance (regulatory reaction) toward food antigens and no inflammation occurs. Instead, in CD, the decision is to react. It is likely that several factors conspire to produce a decision to react, such as the strength of epitope/antigen-presenting molecule interaction on an APC [6, 7], the affinity of a randomly generated T cell receptor for that specific gluten epitope, and the coincident mix of cytokines and co-stimulatory molecules present when that Th cell engages its cognate antigen [28, 30]. In patients with active CD, pro-inflammatory IFN-γ and IL-17 producing (Th1/Th17) cells control the response [31]. These cells leave the draining lymph node and migrate (traffic) back to the intestinal mucosa where, in response to locally displayed gluten epitopes, they help drive the inflammation.
Within the mucosa, several different cell types work together to cause an inflammatory response (Fig. 4.3). Inflammatory Th1/Th17 cells respond to gluten epitopes displayed by lamina propria macrophages and dendritic cells by producing IFN-γ and IL-17. Each of these cytokines has multiple effects on surrounding cells, and both are often found at sites of autoimmune inflammation [32]. IFN-γ feeds back on APC to change their cytokine and cell surface receptor display, enhancing development of more pro-inflammatory T cells. IFN-γ also instructs macrophages to upregulate the ability to kill ingested bacteria. Importantly, IFN-γ instructs epithelial cells to display HLA-E [33], potentially targeting them for injury by intraepithelial lymphocytes (IEL). Th17 cells make IL-17, which increases neutrophil activity and other cytokines like IL-6 and TNF-α, which drive tissue inflammation.

Inflammatory circuits in CD. Multiple cell types are involved in the intestinal inflammation in CD. Anti-deamidated gliadin is taken up by an APC (macrophage or dendritic cell) and is presented to a T cells using a DQ2 or DQ8 antigen-presenting molecule. The activated T cells make IFN-γ and IL-17. The IFN-γ instructs epithelial cells to display HLA-E. The APC also makes IL-15 that causes IEL to proliferate and display NK receptors that recognize HLA and MICA on epithelial cells. The activated IEL then kills the targeted epithelial cells. B cells in the lamina propria also can present antigen to T cells that help the B cells mature into plasma cells that make anti-TTG or anti-deamidated gliadin antibodies
Lamina propria T cell IFN-γ also likely signals APC to make IL-15 [34]. In addition, a specific gliadin peptide (p31-43) can trigger IL-15 production by lamina propria APC [35]. IL-15 is a central cytokine in CD and other immune-mediated inflammatory diseases [36]. Many cell types, including intestinal epithelial cells, can make IL15. Mice engineered to overproduce IL-15 by their intestinal epithelial cells develop inflammation that recapitulates CD [37, 38]. IL-15 has many effects; it augments inflammatory IFN-γ and TNF-α production, suppresses T regulatory cell function [39], and can feedback to promote its own production [34]. IL-15 may also instruct epithelial cells to display greater levels of “MHC class I chain-related gene A” (MICA) [40]. Most importantly, IL-15 promotes the proliferation of cytolytic CD8+ IEL [36, 41] and causes them to express receptors that target killing of epithelial cells.
Intraepithelial Lymphocytes: Agents of Destruction
Small numbers of T cells normally reside above the basement membrane and between epithelial cells. These are called intraepithelial lymphocytes (IEL). IELs likely aid in the identification, removal, and replacement of damaged epithelial cells. There are few different types of IEL. The majority are CD8+ T cells that utilize a T cell receptor (TCR) generated by recombining gene segments in the α and β TCR loci (CD8+α/β T cells). These cells recognize antigens displayed on Class I MHC molecules. Class I MHC molecules mostly display pieces of proteins that were made by the cell as part of its normal functioning. Cytolytic CD8+ T cells survey this display to find evidence for production of unusual proteins suggesting presence of a mutated or virally infected cell. When discovered, they kill the abnormal cell. Another important group of IEL is CD8+ T cells that utilize a TCR generated by recombining gene segments in the γ and δ TCR loci (CD8+γ/δ T cells). These cells often recognize antigens that can be displayed on “nonclassical” antigen-presenting molecules like CD1 and MICA, which are both expressed by epithelial cells.
In active CD, the number of IELs greatly expands. Indeed, this expansion is a distinctive feature of gluten-induced intestinal inflammation [42]. This expansion is driven by IL1-5 made by lamina propria APC and by stressed epithelial cells [36, 41]. IL-15 also instructs IELs to express a set of receptors normally displayed by “natural killer” (NK) cells. NK cells are similar to CD8+ lymphocytes, but they do not utilize a TCR. Instead, they have a set of germ-line receptors that probe for the absence or presence of abnormal antigen-presenting molecules on a cell’s surface [43]. NK cells dispose of virally infected or mutant cells that may not be recognized by CD8+ T cells. IL-15 induces expression of NKG2D [44], which interacts with MICA (and MICB) displayed on epithelial cells. Unlike IEL in normal intestine, IELs in patients with CD also express another NK receptor called CD94/NKG2C [45]. CD94/NKG2C recognizes HLA-E, which is upregulated in epithelial cells in response to IFN-γ [33]. Engagement of NKG2D with MICA/B and CD94/NKG2C with HLA-E activates the IELs without requiring CD8/TCR recognition of an abnormal protein. Acquisition of NKG2D and CD94/NKG2C receptors by IEL and display of MICA/B and HLA-E by epithelial cells target the epithelial cells for destruction.
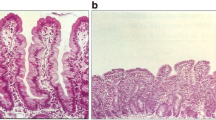
Thus, the hallmark injury in CD, villous atrophy, results from destruction of intestinal epithelial cells by IEL in response to IL-15 made by intestinal epithelial cells, lamina propria macrophages, and dendritic cells. Epithelial cells are targeted for destruction by cell surface display of MICA/B, induced by cell stress and possibly IL-15, and HLA-E, induced by IFN-γ made by gluten-responsive lamina propria T cells. The cells causing injury are the more numerous CD8+α/β IELs. The CD8+ γ/δ IEL (particularly those expressing inhibitory receptor CD94/NKG2A) may be uninvolved or even protective in CD [42, 46, 47], though γ/δ IEL may also be the cell type that gives rise to enteropathy associated T cell lymphoma (EATL) in refractory CD.
Refractory Celiac Disease: Antigen-Independent Inflammation
The vast majority of patients respond to a gluten-free diet with resolution of their intestinal inflammation and normalization of their serum anti-TTG and anti-deamidated gliadin levels. Rarely, intestinal inflammation remains or returns even though the patient is maintaining a gluten-free lifestyle. Refractory celiac disease (RCD) is defined as persistent or recurrent malabsorptive symptoms and signs, with villous atrophy despite a strict gluten-free diet for more than 12 months [48]. RCD is further broken down into two types based on the absence or presence of abnormal intestinal T cells. Normal T cells express the marker CD3, a component of the TCR complex, on their surface. In RCD I, lamina propria T cells and IEL are surface CD3+ and appear identical to those in untreated active celiac disease. In RCD II, there is development of an aberrant CD4-CD8- γδ T cell clone that expresses intracellular but not surface CD3 and makes up more than 50 % of IEL by immunohistochemistry. These cells also express the integrin CD103+ (αE) common to IEL suggesting they arise from the IEL population [49]. Approximately, 50 % of patients with RCD II develop EATL within 5 years of diagnosis, which is why RCDII has a poor prognosis.
In RCD, intestinal inflammation continues in the absence of known gluten exposure. In RCD I, it is possible that unidentified gluten-like epitopes present in other food are sufficient to maintain the inflammation. In a study of ten patients with RCD I placed on a non-immunogenic elemental diet, eight of the nine patients that completed the study had histologic improvement and seven had a decrease in mucosal IFNγ RNA expression [50]. This suggests that RCD I reflects an ongoing response to an unidentified antigen.
On the other hand, it is possible that RCD results from a self-sustaining inflammation in the absence of Th direction. Epithelial cells are capable of making IL15 [51], and this production is upregulated in active celiac disease [41]. IELs can make IFN-γ, and this production is upregulated in active CD [41, 52]. Thus, epithelial cells can make IL-15 to drive proliferation of IEL that display NKG2D and CD94/NKG2C. These IELs in turn produce IFN-γ that upregulates HLA-E expression by epithelial cells that also display MICA/B. This could create a positive-feedback loop, creating IL-15-dependent inflammation in the absence of specific antigen exposure. Moreover, IL-15 supports the survival proliferation of aberrant T cells in RCD II [53]. Thus, it is easy to hypothesize self-amplifying circuits in RCD. The centrality of IL-15 in these circuits fosters interest in therapeutic trials involving IL-15 blockade [37, 54].
Anti-TTG and Anti-Deamidated Gliadin Antibodies
An important screening tool for CD is testing for high titer (elevated concentration) of antibodies against TTG and deamidated gliadin. However, their role in the pathogenesis of CD is unclear. Anti-gliadin antibodies probably help shuttle gluten peptides transcellularly across the epithelial layer [12]. Antibodies are made by B lymphocytes. B cells differentiate into plasma cells, which secrete copious amounts of antibody. Both cell types are numerous in the intestinal lamina propria. B cells also function as “nonprofessional” APC and can present antigens to T cells. B cells express their clonally unique antibody on their cell surface. Here, the antibody can capture its cognate antigen (gluten or TTG) and cause the B cell to ingest the molecule and process it for presentation to T cells in conjunction with MHC Class 2 molecules like DQ2. T cells that recognize the antigen can secrete growth factors for the B cells to ramp up production of the antibody. In addition, B cells may stimulate T cells to drive inflammation. However, identification of a direct role for celiac-associated antibodies in the pathogenesis of the intestinal inflammation remains elusive [55]. Mice that are engineered to over-express IL15 from intestinal epithelial cells develop celiac-like inflammation with numerous lamina propria B cells and plasma cells and produce elevated serum anti-TTG antibody [38]. This suggests that anti-TTG antibody develops in response to IL-15-driven inflammation rather than causing that inflammation.
Conclusion
CD begins with ingestion of gluten, which finds its way across the epithelial cell barrier to APC. If those APCs utilize DQ2 or DQ8 antigen-presenting molecules, they can display epitopes to Th cells that start to produce IFN-γ and stimulate APC to make IL-15. These cytokines induce IEL to kill epithelial cells and cause the damage that results in celiac disease. Central to the inflammation is IL-15. Antibodies to TTG and anti-deamidated gliadin may increase sensitivity to gluten, but are probably more important as clinical measures of gluten exposure. Although a lot is known, much remains to be discovered.
References
Ciccocioppo R, Di Sabatino A, Corazza GR. The immune recognition of gluten in coeliac disease. Clin Exp Immunol. 2005;140(3):408–16.
Shan L, Molberg Ø, Parrot I, Hausch F, Filiz F, Gray GM, et al. Structural basis for gluten intolerance in celiac sprue. Science. 2002;297(5590):2275–9.
Sollid LM, Qiao SW, Anderson RP, Gianfrani C, Koning F. Nomenclature and listing of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics. 2012;64(6):455–60.
Sollid LM, Markussen G, Ek J, Gjerde H, Vartdal F, Thorsby E. Evidence for a primary association of celiac disease to a particular HLA-DQ alpha/beta heterodimer. J Exp Med. 1989;169(1):345–50.
Murray JA, Moore SB, Van Dyke CT, Lahr BD, Dierkhising RA, Zinsmeister AR, et al. HLA DQ gene dosage and risk and severity of celiac disease. Clin Gastroenterol Hepatol. 2007;5(12):1406–12.
Fallang LE, Bergseng E, Hotta K, Berg-Larsen A, Kim CY, Sollid LM. Differences in the risk of celiac disease associated with HLA-DQ2.5 or HLA-DQ2.2 are related to sustained gluten antigen presentation. Nat Immunol. 2009;10(10):1096–101.
Bodd M, Kim CY, Lundin KE, Sollid LM. T-cell response to gluten in patients with HLA-DQ2.2 reveals requirement of peptide-MHC stability in celiac disease. Gastroenterology. 2012;142(3):552–61.
Lundin KE, Gjertsen HA, Scott H, Sollid LM, Thorsby E. Function of DQ2 and DQ8 as HLA susceptibility molecules in celiac disease. Hum Immunol. 1994;41(1):24–7.
Karell K, Louka AS, Moodie SJ, Ascher H, Clot F, Greco L, et al. HLA types in celiac disease patients not carrying the DQA1*05-DQB1*02 (DQ2) heterodimer: results from the European Genetics Cluster on Celiac Disease. Hum Immunol. 2003;64(4):469–77.
Lugering A, Floer M, Lügering N, Cichon C, Schmidt MA, Domschke W, et al. Characterization of M cell formation and associated mononuclear cells during indomethacin-induced intestinal inflammation. Clin Exp Immunol. 2004;136(2):232–8.
Heyman M, Abed J, Lebreton C, Cerf-Bensussan N. Intestinal permeability in coeliac disease: insight into mechanisms and relevance to pathogenesis. Gut. 2012;61(9):1355–64.
Matysiak-Budnik T, Moura IC, Arcos-Fajardo M, Lebreton C, Ménard S, Candalh C, et al. Secretory IgA mediates retrotranscytosis of intact gliadin peptides via the transferrin receptor in celiac disease. J Exp Med. 2008;205(1):143–54.
Berin MC. Mucosal antibodies in the regulation of tolerance and allergy to foods. Semin Immunopathol. 2012;34(5):633–42.
Caputo I, Barone MV, Lepretti M, Martucciello S, Nista I, Troncone R, et al. Celiac anti-tissue transglutaminase antibodies interfere with the uptake of alpha gliadin peptide 31-43 but not of peptide 57-68 by epithelial cells. Biochim Biophys Acta. 2010;1802(9):717–27.
Anderson JM, Van Itallie CM. Physiology and function of the tight junction. Cold Spring Harb Perspect Biol. 2009;1(2):a002584.
Van Itallie CM, Fanning AS, Holmes J, Anderson JM. Occludin is required for cytokine-induced regulation of tight junction barriers. J Cell Sci. 2010;123(Pt 16):2844–52.
Fasano A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann N Y Acad Sci. 2012;1258:25–33.
Leffler DA, Kelly CP, Abdallah HZ, Colatrella AM, Harris LA, Leon F, et al. A randomized, double-blind study of larazotide acetate to prevent the activation of celiac disease during gluten challenge. Am J Gastroenterol. 2012;107(10):1554–62.
Niess JH, Reinecker HC. Dendritic cells: the commanders-in-chief of mucosal immune defenses. Curr Opin Gastroenterol. 2006;22(4):354–60.
Lelouard H, Fallet M, de Bovis B, Méresse S, Gorvel JP et al. Peyer's patch dendritic cells sample antigens by extending dendrites through M cell-specific transcellular pores. Gastroenterology, 2012; 142(3): 592-601 e3.
Chieppa M, Rescigno M, Huang AY, Germain RN. Dynamic imaging of dendritic cell extension into the small bowel lumen in response to epithelial cell TLR engagement. J Exp Med. 2006;203(13):2841–52.
Wells JM, Rossi O, Meijerink M, van Baarlen P. Epithelial crosstalk at the microbiota-mucosal interface. Proc Natl Acad Sci USA. 2011;108 Suppl 1:4607–14.
Palova-Jelinkova L, Rozková D, Pecharová B, Bártová J, Sedivá A, Tlaskalová-Hogenová H, et al. Gliadin fragments induce phenotypic and functional maturation of human dendritic cells. J Immunol. 2005;175(10):7038–45.
Mones RL, Mercer GO. Ulcerative duodenitis in a child with celiac disease. J Pediatr. 2011;158(5):857.
Rubio-Tapia A, Murray JA. Classification and management of refractory coeliac disease. Gut. 2010;59(4):547–57.
Klock C, Diraimondo TR. C. Khosla C. Role of transglutaminase 2 in celiac disease pathogenesis. Semin Immunopathol. 2012;34(4):513–22.
Vader LW, de Ru A, van der Wal Y, Kooy YM, Benckhuijsen W, Mearin ML, et al. Specificity of tissue transglutaminase explains cereal toxicity in celiac disease. J Exp Med. 2002;195(5):643–9.
Zygmunt B, Veldhoen M. T helper cell differentiation more than just cytokines. Adv Immunol. 2011;109:159–96.
Pabst O, Mowat AM. Oral tolerance to food protein. Mucosal Immunol. 2012;5(3):232–9.
Vahedi G, C Poholek A, Hand TW, Laurence A, Kanno Y, O’Shea JJ, et al. Helper T-cell identity and evolution of differential transcriptomes and epigenomes. Immunol Rev. 2013;252(1):24–40.
Monteleone I, Sarra M, Del Vecchio BG, Paoluzi OA, Franzè E, Fina D, et al. Characterization of IL-17A-producing cells in celiac disease mucosa. J Immunol. 2010;184(4):2211–8.
Moudgil KD, Choubey D. Cytokines in autoimmunity: role in induction, regulation, and treatment. J Interferon Cytokine Res. 2011;31(10):695–703.
Perera L, Shao L, Patel A, Evans K, Meresse B, Blumberg R, et al. Expression of nonclassical class I molecules by intestinal epithelial cells. Inflamm Bowel Dis. 2007;13(3):298–307.
Waldmann TA, Tagaya Y. The multifaceted regulation of interleukin-15 expression and the role of this cytokine in NK cell differentiation and host response to intracellular pathogens. Annu Rev Immunol. 1999;17:19–49.
Maiuri L, Ciacci C, Ricciardelli I, Vacca L, Raia V, Auricchio S, et al. Association between innate response to gliadin and activation of pathogenic T cells in coeliac disease. Lancet. 2003;362(9377):30–7.
Di Sabatino A, Calarota SA, Vidali F, Macdonald TT, Corazza GR. Role of IL-15 in immune-mediated and infectious diseases. Cytokine Growth Factor Rev. 2011;22(1):19–33.
Yokoyama S, Watanabe N, Sato N, Perera PY, Filkoski L, Tanaka T, et al. Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes. Proc Natl Acad Sci USA. 2009;106(37):15849–54.
Yokoyama S, Takada K, Hirasawa M, Perera LP, Hiroi T. Transgenic mice that overexpress human IL-15 in enterocytes recapitulate both B and T cell-mediated pathologic manifestations of celiac disease. J Clin Immunol. 2011;31(6):1038–44.
Zanzi D, Stefanile R, Santagata S, Iaffaldano L, Iaquinto G, Giardullo N, et al. IL-15 interferes with suppressive activity of intestinal regulatory T cells expanded in Celiac disease. Am J Gastroenterol. 2011;106(7):1308–17.
Zwirner NW, Fuertes MB, Girart MV, Domaica CI, Rossi LE. Immunobiology of the human MHC class I chain-related gene A (MICA): from transplantation immunology to tumor immune escape. Immunologia. 2006;25(1):25–38.
Di Sabatino A, Ciccocioppo R, Cupelli F, Cinque B, Millimaggi D, Clarkson MM, et al. Epithelium derived interleukin 15 regulates intraepithelial lymphocyte Th1 cytokine production, cytotoxicity, and survival in coeliac disease. Gut. 2006;55(4):469–77.
Abadie V, Discepolo V, Jabri B. Intraepithelial lymphocytes in celiac disease immunopathology. Semin Immunopathol. 2012;34(4):551–66.
Rajalingam R. Overview of the killer cell immunoglobulin-like receptor system. Methods Mol Biol. 2012;882:391–414.
Meresse B, Chen Z, Ciszewski C, Tretiakova M, Bhagat G, Krausz TN, et al. Coordinated induction by IL15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine-activated killer cells in celiac disease. Immunity. 2004;21(3):357–66.
Meresse B, Curran SA, Ciszewski C, Orbelyan G, Setty M, Bhagat G, et al. Reprogramming of CTLs into natural killer-like cells in celiac disease. J Exp Med. 2006;203(5):1343–55.
Verbeek WH, von Blomberg BM, Scholten PE, Kuik DJ, Mulder CJ, Schreurs MW. The presence of small intestinal intraepithelial gamma/delta T-lymphocytes is inversely correlated with lymphoma development in refractory celiac disease. Am J Gastroenterol. 2008;103(12):3152–8.
Bhagat G, Naiyer AJ, Shah JG, Harper J, Jabri B, Wang TC, et al. Small intestinal CD8+TCRgammadelta+NKG2A+ intraepithelial lymphocytes have attributes of regulatory cells in patients with celiac disease. J Clin Invest. 2008;118(1):281–93.
Ludvigsson JF, Leffler DA, Bai JC, Biagi F, Fasano A, Green PH, et al. The Oslo definitions for coeliac disease and related terms. Gut. 2013;62(1):43–52.
Malamut G, Meresse B, Cellier C, Cerf-Bensussan N. Refractory celiac disease: from bench to bedside. Semin Immunopathol. 2012;34(4):601–13.
Olaussen RW, Løvik A, Tollefsen S, Andresen PA, Vatn MH, De Lange T, et al. Effect of elemental diet on mucosal immunopathology and clinical symptoms in type 1 refractory celiac disease. Clin Gastroenterol Hepatol. 2005;3(9):875–85.
Reinecker HC, MacDermott RP, Mirau S, Dignass A, Podolsky DK. Intestinal epithelial cells both express and respond to interleukin 15. Gastroenterology. 1996;111(6):1706–13.
Leon F, Sánchez L, Camarero C, Roy G. Cytokine production by intestinal intraepithelial lymphocyte subsets in celiac disease. Dig Dis Sci. 2005;50(3):593–600.
Malamut G, El Machhour R, Montcuquet N, Martin-Lannerée S, Dusanter-Fourt I, Verkarre V, et al. IL-15 triggers an antiapoptotic pathway in human intraepithelial lymphocytes that is a potential new target in celiac disease-associated inflammation and lymphomagenesis. J Clin Invest. 2010;120(6):2131–43.
Sollid LM, Khosla C. Novel therapies for coeliac disease. J Intern Med. 2011;269(6):604–13.
Mesin L, Sollid LM, Di Niro R. The intestinal B-cell response in celiac disease. Front Immunol. 2012;3:313.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Elliott, D.E. (2014). The Pathophysiology of Celiac Disease. In: Rampertab, S., Mullin, G. (eds) Celiac Disease. Clinical Gastroenterology. Humana Press, New York, NY. https://doi.org/10.1007/978-1-4614-8560-5_4
Download citation
DOI: https://doi.org/10.1007/978-1-4614-8560-5_4
Published:
Publisher Name: Humana Press, New York, NY
Print ISBN: 978-1-4614-8559-9
Online ISBN: 978-1-4614-8560-5
eBook Packages: MedicineMedicine (R0)