
Abstract
Neuropeptide Y (NPY) plays an important role in maintaining energy homeostasis. Within the hypothalamus, Npy is primarily expressed in the arcuate nucleus (Arc) and the dorsomedial hypothalamus (DMH). ARC NPY acts as an orexigenic neuromodulator integrating energy-related systemic signals (such as the adiposity signal leptin) to modulate food intake and energy balance. In contrast, DMH NPY acts independently of leptin. In addition to its feeding effects, DMH NPY has specific actions on brown adipocyte formation and thermoregulation. Knockdown of NPY in the DMH promotes development of brown adipocytes in white adipose tissue and increases brown adipocyte activity, leading to increased energy expenditure and lowered fat contents. DMH NPY knockdown prevents high-fat diet-induced obesity. Peripheral NPY also directly acts on adipose tissue and promotes white adipogenesis. Overall, NPY plays critical roles in the control of energy balance through affecting food intake, body adiposity, and energy expenditure. These actions provide evidence for the potential target(s) for combatting obesity and diabetes.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
- Dorsomedial hypothalamus
- Neuropeptide Y
- Brown adipose tissue
- Uncoupling protein 1
- Adipogenesis
- Adipocyte transformation
- Thermogenesis
- Food intake
- Energy expenditure
- Obesity
1 Introduction
Two types of fat, white adipose tissue (WAT) and brown adipose tissue (BAT), exist in mammals, including in adult humans as recently reported (Cypess et al. 2009; van Marken Lichtenbelt et al. 2009; Virtanen et al. 2009). WAT consists of unilocular adipocytes that contain a large lipid droplet. WAT acts as an energy reservoir to store excess calories and supply the stored lipid once a body is starved or increases energy needs. An excessive accumulation of WAT over time due to constantly increased energy intake and/or decreased energy expenditure causes overweight and obesity, a disorder that has been linked to various life-threatening diseases such as cardiovascular diseases, type II diabetes, and some cancers. Since the discovery of leptin in 1994 (Zhang et al. 1994), a hormone produced by adipocytes, the understanding of the adipocyte has significantly increased. In addition to storing energy, the adipocyte acts as a key endocrine gland secreting various hormones and/or cytokines such as leptin (Ahima et al. 1996; Zhang et al. 1994), adiponectin (Yamauchi et al. 2001), resistin (Steppan et al. 2001), tumor necrosis factor-alpha (TNF-α) (Uysal et al. 1997), and interleukin 6 (IL-6) (Wallenius et al. 2002) and playing a fundamental role in regulating energy balance and glucose homeostasis. Dysfunctions of adipocytes have been demonstrated to cause disordered energy balance and impaired glucose homeostasis. For instance, deficits in leptin signaling derived from either leptin deficiency in ob/ob mice (Ingalls et al. 1950) or its receptor mutations in db/db mice (Hummel et al. 1966) lead to obesity and diabetes syndromes.
BAT is comprised of multilocular and mitochondrial-rich adipocytes that contain multiple small lipid droplets. BAT dissipates lipid energy to produce heat via mitochondrial uncoupling protein 1 (UCP1)-mediated nonshivering thermogenesis as a defense against cold and the potential for combatting obesity/diabetes. BAT is traditionally considered as a thermogenic organ and primarily present in small mammals and human infants for protection against cold environment. Recently, using a combination of 18fluoro-labeled 2-deoxyglucose (18FDG) positron emission tomography and computed tomography (PET-CT) technology, active BAT has been detected in adult humans (Cypess et al. 2009; van Marken Lichtenbelt et al. 2009; Virtanen et al. 2009). Adult humans have increased BAT activity during cold exposure, and lowered BAT activity when they are overweight or obese (van Marken Lichtenbelt et al. 2009; Virtanen et al. 2009), although the origin of these brown adipocytes remains to be determined. Nevertheless, the findings of BAT in adult humans have led to a great interest in its potential for fighting against obesity/diabetes, i.e., searching for ways to elevate BAT activity or to turn WAT into BAT that burns calories instead of storing them.
Both types of fat are innervated by the sympathetic nervous system (SNS) (Bartness and Bamshad 1998; Cannon and Nedergaard 2004). Activation of the sympathetic innervation induces lipolysis in WAT (Fredholm and Karlsson 1970; Weiss and Maickel 1968) and produces thermogenesis through mitochondrial UCP1 in BAT (Cannon and Nedergaard 2004). Intriguingly, the sympathetic activation via treatment of β3-adrenergic receptor (β3-AR) agonists or cold stress also causes development of brown adipocytes in white fat cells (Himms-Hagen et al. 1994; Jimenez et al. 2003; Nagase et al. 1996). Follow up studies demonstrate the transdifferentiation of white to brown adipocytes as multilocular fat cells (brown-like adipocytes) in WAT induced by β3-AR agonist derive from unilocular white adipocytes (Himms-Hagen et al. 2000). Consistent with this finding, the emergence of cold-induced brown adipocytes in white fat depots is determined predominantly by white to brown adipocyte transdifferentiation with evidence of transdifferentiating paucilocular adipocytes (Barbatelli et al. 2010). In contrast, Petrovic et al. (2010) argued against this idea and proposed two different progenitor cells in BAT and WAT, respectively. The classical brown adipocytes such as interscapular BAT are derived from “adipomyocytes” that share their origin with myocytes (Seale et al. 2008). The other is atypical brown adipocytes called “brite” adipocytes (brown-in-white, i.e., the brown adipocyte-like adipocytes induced in white adipocytes, or “beige” cells) developing from an origin that shares a common precursor with white adipocytes (Petrovic et al. 2010). In support of this view, Wu and colleagues (2012) have identified these beige cells as a distinct type of thermogenic fat cell in both mouse and human. Using viral-mediated RNA interference (RNAi) for specifically knocking down neuropeptide Y (NPY) in the dorsomedial hypothalamus (DMH), Chao et al. (2011) reported that DMH NPY knockdown promotes development of brown adipocytes in WAT, elevates interscapular BAT activity, and prevents high-fat diet-induced obesity and impaired glucose homeostasis. This chapter will focus on the role of NPY in brown adipocyte formation and actions on obesity.
NPY is a 36-amino acid neuropeptide that was discovered by Tatemoto and colleagues (1982) and belongs to the pancreatic polypeptide family that includes peptide tyrosine-tyrosine (PYY) and pancreatic polypeptide (PP) (Tatemoto 1982; Tatemoto et al. 1982). NPY is ubiquitously distributed in both central and peripheral nervous systems. It is prevalent in central cortical, limbic, and hypothalamic regions (Adrian et al. 1983; Allen et al. 1983) and in the peripheral SNS (Lundberg et al. 1982). NPY exhibits a variety of biological and physiological actions including feeding, thermoregulation, locomotor activity, cardiovascular function, cognition and memory, and stress-related behaviors (Bi 2007; Colmers and Wahlestedt 1993; Gray and Morley 1986). Hypothalamic NPY plays a pivotal role in the control of food intake and body weight. Central administration of NPY via intracerebroventricular (icv) (Clark et al. 1984; Levine and Morley 1984) or intrahypothalamic injection (Stanley et al. 1986; Stanley and Leibowitz 1985) causes robust increases in food intake and body weight and, with chronic administration, can eventually produce obesity (Zarjevski et al. 1993). In addition to its orexigenic effects, icv administration of NPY promotes WAT lipid storage and decreases BAT thermogenesis (Billington et al. 1991). Central administration of NPY suppresses sympathetic activity in interscapular BAT in rats (Egawa et al. 1991). Thus, hypothalamic NPY not only acts as an orexigenic peptide, but also affects adipocyte lipid mobilization and BAT functions.
2 Role of Arcuate NPY in Adiposity
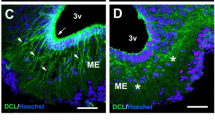
Within the hypothalamus, NPY-expressing neurons have been identified in the arcuate nucleus (ARC) and the DMH (Bi et al. 2003; White and Kershaw 1990). As shown in Fig. 9.1, Npy gene expression is found in both the ARC and the DMH of rat (Fig. 9.1a) and nonhuman primate brains (Fig. 9.1b), although the expression level of DMH NPY is much lower than that within the ARC. The actions of ARC NPY in the control of food intake and energy balance have been documented. The ARC contains two distinct populations of neurons: orexigenic neuropeptide NPY/agouti-related protein (AgRP) neurons and anorexigenic proopiomelanocortin (POMC) neurons. Both types of neurons contain leptin receptors (LepRbs). Leptin acts on these neurons to down-regulate Npy/Agrp gene expression and upregulate Pomc gene expression (Schwartz et al. 2000). Collectively, these two neural systems integrate adiposity signals (such as leptin) and nutrient signals as well as other hormonal signals (such as ghrelin) to modulate food intake and energy balance (Cone 2006; Elmquist et al. 1999; Flier 2004; Friedman and Halaas 1998; Nakazato et al. 2001; Schwartz et al. 2000). Consistent with this view, while ARC Npy mRNA or peptide levels are elevated in genetic obesity animals with leptin signaling deficits including fatty Zucker (fa/fa) rats, obese (cp/cp) JCR:LA corpulent (Koletsky) rat, ob/ob mice, and db/db mice (Beck 2006; Sanacora et al. 1990; Wilding et al. 1993), such elevations are not evident in other obesity models such as those resulting from hyperphagia (Bi et al. 2001), overfeeding (McMinn et al. 1998), or access to a high-fat diet (Giraudo et al. 1994; Stricker-Krongrad et al. 1998) in which levels of ARC Npy expression are actually decreased, likely as a response to increased levels of leptin and body weight. Although data from mouse models with targeted disruption of NPY [either knockout (Erickson et al. 1996a) or transcriptional alterations through doxycycline-regulated system (Ste Marie et al. 2005)] have failed to demonstrate significant effects on food intake or body weight, the deletion of NPY does attenuate a hyperphagic and obese phenotype of leptin-deficient ob/ob mice (Erickson et al. 1996b) and genetic ablation of neurons expressing NPY/AgRP in adult mice results in a hypophagic and lean phenotype (Bewick et al. 2005; Gropp et al. 2005). Moreover, consistent with ARC NPY mediation of food deprivation-induced feeding, specific knockdown of NPY in the ARC via adeno-associated virus (AAV)-mediated RNAi attenuates the feeding response to food deprivation (Yang et al. 2009). AAV-mediated expression of antisense Npy cRNA in the ARC of adult rats decreases NPY expression and results in decreased food intake and body weight (Gardiner et al. 2005), whereas AAV-mediated overexpression of NPY in the ARC causes overeating, sustained body weight gain, and severe obesity in adult rats (Sousa-Ferreira et al. 2011). Together, these data demonstrate that ARC NPY acts as an orexigenic peptide to modulate food intake and energy balance to affect adiposity, but whether ARC NPY has adipocyte-specific effects, in addition to feeding effects, has yet to be reported.

In situ hybridization with [35S]-labeled antisense riboprobe of rodent or human Npy shows Npy gene expression in the arcuate nucleus (ARC) and the dorsomedial hypothalamus (DMH) in both rats (a) and adult rhesus monkey brain (b)
3 Role of DMH NPY in Adiposity
Elevation or induction of Npy gene expression in the DMH in specific animal models with increased energy demands (Bi et al. 2003; Kawaguchi et al. 2005; Smith 1993) and several rodent models of obesity (Bi et al. 2001; Guan et al. 1998a, 1998b; Kesterson et al. 1997; Tritos et al. 1998) implicates DMH NPY in the control of energy balance. In support of this view, recent evidence has demonstrated a role for DMH NPY in modulating food intake and energy balance (Yang et al. 2009) and intriguingly in affecting brown adipocyte formation and thermoregulation (Chao et al. 2011).
3.1 Leptin-Independent Control of DMH NPY
While ARN NPY serves as one of the downstream mediators of leptin’s actions or is under the control of the adiposity signal leptin, DMH NPY acts independently of leptin. DMH NPY neurons do not contain LepRbs, although abundant LepRbs are present in the DMH (Bi et al. 2003). Dual in situ hybridization histochemistry revealed that while Npy and LepRbs are co-expressed in ARC neurons, DMH Npy-expressing neurons do not co-express LepRbs (Bi et al. 2003). Npy gene expression is increased in the ARC in response to acute food deprivation, a time when circulating leptin levels are low, whereas DMH Npy expression is only significantly increased in rats with chronic food restriction (Bi et al. 2003). Moreover, Npy gene expression is elevated or induced in the DMH of specific rodent models of obesity including the lethal yellow agouti (A y) (Kesterson et al. 1997), MC4R knockout (Kesterson et al. 1997), diet-induced obese (Guan et al. 1998a), tubby (Guan et al. 1998b), and brown adipose tissue-deficient obese mice (Tritos et al. 1998), and Otsuka Long-Evans Tokushima Fatty (OLETF) rats (Bi et al. 2001), but such elevation or induction is not evident in leptin-deficient ob/ob mice (Kesterson et al. 1997). In fact, while Npy gene expression is significantly increased in the ARC of obese animals with leptin signaling deficiency (Beck 2006; Sanacora et al. 1990; Wilding et al. 1993), obese animals with DMH NPY overexpression generally have significantly decreased expression of Npy in the ARC (Bi et al. 2001; Guan et al. 1998a; Kesterson et al. 1997). Thus, ARC and DMH NPY signals are differentially regulated and they each likely elicit distinct actions in the control of energy balance.
3.2 DMH NPY in Brown Adipocyte Development
Previous studies have shown that in addition to its orexigenic effects, icv administration of NPY increases WAT lipoprotein lipase activity (suggesting increased lipid storage) and decreases BAT GDP binding activity (indicating decreased thermogenic activity) (Billington et al. 1991) and that central administration of NPY suppresses sympathetic activity in interscapular BAT in rats (Egawa et al. 1991). These data suggest a role for NPY in lipid mobilization and thermoregulation, but the source of central NPY contributing to these actions has not been determined. Manipulation of Npy gene expression in the DMH using AAV-mediated RNAi revealed that knockdown of NPY in the DMH produces not only a nocturnal and meal size-specific reduction in feeding (Yang et al. 2009), but also site-specific effects on adiposity (Chao et al. 2011). DMH NPY knockdown decreases subcutaneous inguinal fat mass in rats on regular chow and ameliorates high-fat diet-induced increases in fat accumulations including inguinal and epididymal fat and interscapular BAT mass (Chao et al. 2011). Intriguingly, DMH NPY knockdown promotes development of brown adipocytes in inguinal WAT. As shown in Fig. 9.2, while inguinal adipocytes of control rats contain unilocular adipocytes, typical white adipocytes (Fig. 9.2a), inguinal adipose tissue of NPY knockdown rats contains large clusters of multilocular adipocytes (brown-like adipocytes) (Fig. 9.2b). The BAT-specific marker UCP1 is highly detected in these new formed cells and also in a number of unilocular adipocytes around these clusters (Chao et al. 2011). UCP1 expression in this inguinal fat is further confirmed by using both real-time RT-PCR (reverse transcriptase-polymerase chain reaction) and Western blot analyses (Chao et al. 2011). But, whether these browning adipocytes in WAT are directly derived from specific precursor cells such as beige cells, or transdifferentiated from unilocular white adipocytes, or both, remains to be determined.

Representative H&E (hematoxylin and eosin) stain shows unilocular adipocytes in inguinal white adipose tissue (WAT) of rats receiving bilateral DMH injections of control adeno-associated viral vector containing scrambled shRNA (AAVshCTL) (a) and multilocular adipocytes (brown-like adipocytes) in inguinal fat of rats receiving bilateral DMH injections of AAV-mediated RNAi vector (AAVshNPY) (b). Inguinal fat temperature over the 24-h period (c) and mean ± SEM temperature during the dark and the light (d) in rats at an ambient room temperature condition. Black bar indicates the dark cycle. *P < 0.05 compared to AAVshCTL rats
Elevation of UCP1 contributes to increased thermogenesis of BAT (Cannon and Nedergaard 2004). Consistent with this view, we have demonstrated increased thermogenesis in inguinal browning fat of NPY knockdown rats. In the study, the inguinal fat temperature was directly examined using a radio transmitter device (E-mitter, Mini-Mitter) that was buried under the inguinal fat. At an ambient room temperature condition (23 ± 1 °C), the inguinal fat temperature is significantly increased during the dark period in rats with DMH NPY knockdown compared to control animals (Fig. 9.2c, d).
Consistent with the SNS mediation of WAT into BAT transformation (Himms-Hagen et al. 1994; Jimenez et al. 2003; Nagase et al. 1996), the sympathetic innervation contributes DMH NPY knockdown-induced transformation of white to brown adipocytes in inguinal WAT. The study of sympathetic denervation with a regional injection of the neurotoxin 6-hydroxydopamine (6-OHDA) revealed that in the rats with DMH NPY knockdown, unilateral injection of 6-OHDA into the inguinal fat area results in significantly less browning on the side of 6-OHDA injection, whereas the site of inguinal fat receiving saline injection becomes brown-like fat (Chao et al. 2011). This observation was further confirmed by the findings that (1) numerous clusters of brown-like adipocytes (multilocular adipocytes) remain on the side of saline-treated inguinal fat, whereas brown-like adipocytes are dramatically reduced by 6-OHDA treatment and (2) 6-OHDA treatment also prevents UCPl expression in inguinal adipocytes at both the protein and mRNA levels (Chao et al. 2011).
Although the specific neural signaling pathway underlying the effects of DMH NPY on inguinal adiposity remains to be determined, evidence from viral transsynaptic retrograde tracing studies provides support for the idea that DMH NPY modulates SNS innervation on inguinal WAT. Less viral tracing is detected in the DMH in animals receiving epididymal viral injection than those receiving inguinal injection (Bamshad et al. 1998), implying that the central nervous control of inguinal WAT is more DMH related than that of epididymal WAT, i.e., DMH NPY may serve as a central modulator influencing SNS outflow to inguinal WAT.
On the molecular level, a number of transcription factors regulate development of brown adipocytes. Peroxisome proliferator-activated receptor-γ (PPAR-γ) is an essential factor for the development of both white and brown fat cells (Rosen et al. 1999). Chronic treatment with the PPAR-γ agonist rosiglitazone promotes brown adipogenesis in WAT characterized with distinct adipocytes, namely, “brite” adipocytes (Petrovic et al. 2010). The rosiglitazone treatment causes not only the expression of the PPAR-γ coactivator-1α (PGC-1α) and mitochondriogenesis, but also norepinephrine (NE)-augmentable expression of UCP1 in these brite cells (Petrovic et al. 2010). PGC-1a is another key factor involved in brown adipocyte development and thermogenesis (Handschin and Spiegelman 2006). Ectopic expression of PGC-1α in white fat cells induces a number of mitochondrial genes and thermogenic genes, such as Ucp1, whereas genetic ablation of PGC-1α results in reduced capacity for cold-induced thermogenesis in vivo (Puigserver et al. 1998). DMH NPY knockdown promotes both Ppar-γ and Pgc-1α gene expression in inguinal fat (Chao et al. 2011), suggesting that the browning effects of DMH NPY knockdown on inguinal WAT are likely mediated through affecting these key transcription factors, but the detailed molecular control of this white to brown adipocyte transformation merits further investigation.
3.3 DMH NPY Affects Inguinal Lipid Mobilization and Blood Fat Levels
Fatty acid synthase (FAS) and carnitine palmitoyltransferase 1 (CPT1) are two important enzymes involved in fat metabolism. FAS plays a key role in fatty acid synthesis, whereas CPT1 is the rate-limiting enzyme controlling fatty acid oxidation. Compared to control rats, Cpt1 gene expression is significantly increased in the inguinal fat of NPY knockdown rats with a trend toward a decrease in Fas gene expression (Chao et al. 2011), indicating a shift from lipogenesis to fatty acid oxidation in this tissue. Consistent with this view, the weight of inguinal fat mass is significantly decreased in NPY knockdown rats as compared with their control counterparts (Chao et al. 2011). Importantly, DMH NPY knockdown results in decreased levels of both blood triglyceride (Fig. 9.3a) and free fatty acid (Fig. 9.3b), whereas NPY overexpression in the DMH causes increased levels of blood triglyceride (Fig. 9.3a). These data indicate that DMH NPY is an important neuromodulator in modulation of fat utilization. Specifically, the browning effects of NPY knockdown on fat may contribute to these reductions of blood triglyceride and free fatty acid levels since BAT activity has been demonstrated to control triglyceride clearance (Bartelt et al. 2011).

Effects of manipulation of NPY expression in the DMH on blood triglyceride (a) and free fatty acid levels (b). AAVGFP, rats receiving bilateral DMH injections of control AAV vector (AAVGFP); AAVNPY, rats receiving bilateral DMH injections of AAV-mediated NPY expression vector (AAVNPY)
3.4 DMH NPY Affects Interscapular BAT Activity
Evidence has indicated the importance of the DMH in regulating sympathetic nerve activity to interscapular BAT and thermogenesis. Stimulation or disinhibition of neurons in the DMH by parenchymal microinjection of glutamate or γ-aminobutyric acid (GABA)-A receptor antagonist results in great increases in sympathetic nerve activity to interscapular BAT and increases in BAT and core body temperature (Dimicco and Zaretsky 2007; Morrison and Nakamura 2011). Collectively, the DMH has been proposed as an intermediate relay receiving the inputs from the hypothalamic preoptic area (POA), a center of integrating centrally and peripherally thermal signals, and sending the outputs to the rostral raphe pallidus (rRP) in the medulla, the area containing premotor neurons that innervate SNS to interscapular BAT (Dimicco and Zaretsky 2007; Morrison and Nakamura 2011). Although the nature of DMH neurons mediating these effects has not yet been fully characterized, DMH NPY may serve as one of the mediators. DMH NPY knockdown results in increased Ucp1 gene expression in the interscapular BAT, whereas NPY overexpression in the DMH causes decreased expression of Ucp1 in this fat tissue (Fig. 9.4a). This suggests that DMH NPY affects interscapular BAT activity or thermogenesis. In support of this view, DMH NPY knockdown results in increased interscapular BAT thermogenesis as directly determined by increased BAT temperature. At an ambient room temperature condition (23 ± 1 °C), interscapular BAT temperature is significantly increased during the dark with a trend for increases during the light compared to control animals (Fig. 9.4b, c). Thus, DMH NPY also modulates interscapular BAT thermogenesis or activity to affect energy expenditure.

Effects of manipulation of DMH NPY expression on the expression of uncoupling protein 1 (UCP1) in interscapular brown adipose tissue (BAT) (a), and effects of NPY knockdown in the DMH on interscapular BAT temperature over the 24-h period (b) and mean ± SEM BAT temperature during the dark and light period (c) in rats at an ambient room temperature condition. *P < 0.05 compared to AAVshCTL rats
3.5 Role of DMH NPY in Energy Expenditure
Consistent with the actions of DMH NPY on activity of both inguinal fat and interscapular BAT, manipulation of DMH NPY expression affects energy expenditure and thermogenesis. Using an oxymax equal flow indirect calorimetric system, we determined that DMH NPY knockdown results in increased oxygen consumption during both dark and light phases of the circadian cycle at an ambient room temperature condition (23 ± 1 °C) (Fig. 9.5a), indicating that rats with DMH NPY knockdown have increased daily energy metabolism. NPY knockdown rats also have decreased respiratory exchange rate (RER) during the dark and across 24 hours (Fig. 9.5b), indicating increased lipid oxidation. As a result, energy expenditure (EE) is significantly increased over the 24 h period (Fig. 9.5c). Furthermore, although core body temperature does not differ between NPY knockdown and control rats at room temperature (23 ± 1 °C), NPY knockdown rats have a greater increase in thermogenic response to 6 h of cold exposure (6 °C) compared to their control counterparts (Chao et al. 2011). In response to 6 h of cold exposure, Ucp1 gene expression is significantly elevated in the interscapular BAT of control rats (Fig. 9.6a) and DMH NPY knockdown enhances this elevation (Fig. 9.6a). Six hours of cold exposure does not induce Ucp1 gene expression in inguinal WAT of control rats, but Ucp1 gene expression is largely induced in inguinal WAT of rats with DMH NPY knockdown (Fig. 9.6b). Overall, DMH NPY plays a functional role in the regulation of thermogenesis and energy expenditure likely through affecting the activity of both inguinal fat and interscapular BAT.

Effects of DMH NPY knockdown on oxygen consumption (a), respiratory exchange rate (RER) (b), and energy expenditure (EE) (c) in rats at room temperature. Black bar indicates the dark cycle. *P < 0.05 compared to AAVshCTL rats

Effects of DMH NPY knockdown on Ucp1 gene expression in interscapular BAT (a) and inguinal fat (b) in response to 6 h of cold exposure (6 °C). AAVshCTL RT, AAVshCTL rats at room temperature; AAVshCTL Cold, AAVshCTL rats at 6 °C; and AAVshNPY Cold, AAVshNPY rats at 6 °C. *P < 0.05 compared to AAVshCTL rats. #P < 0.05 compared to AAVshCTL Cold
4 Role of Peripheral NPY in Adiposity
Peripheral NPY is primarily produced in the SNS and co-operates with NE to affect sympathetic control of various tissues (Lundberg et al. 1982) including adipose tissue (Bartness and Bamshad 1998). Adipose tissue also contains NPY and its Y1 and Y2 receptors (Kuo et al. 2007; Yang et al. 2008). Peripheral NPY acts directly on adipose tissue and mediates stress-induced obesity and metabolic syndrome through its interacting with Y2 receptors (Kuo et al. 2007). While activation of the sympathetic innervation and the resultant release of NE induces lipolysis in WAT (Fredholm and Karlsson 1970; Weiss and Maickel 1968), the release of NPY from sympathetic nerves upregulates NPY and Y2 receptors in adipose tissue, leading to adipogenesis directly by stimulating proliferation and differentiation of preadipocytes and indirectly by angiogenesis (Kuo et al. 2007). Consistent with this view, pharmacological antagonism or genetic knockdown of peripheral Y2 receptors prevents high-fat diet-induced obesity (Kuo et al. 2007; Shi et al. 2011). Adipose Y1 receptors have also been shown to contribute to the effects of peripheral NPY on adipogenesis. Adipose NPY promotes proliferation of adipocyte precursor cells via the Y1 receptor (Yang et al. 2008) and adipose Y1 receptors regulate lipid oxidation and fat accretion (Zhang et al. 2010). Thus, while central NPY-Y1/2 systems have specific effects on feeding to modulate energy balance (Lin et al. 2004), adipose Y1/2 receptors mediate the actions of peripheral NPY on adipogenesis and fat accumulation, eventually leading to development of obesity.
5 Conclusions
NPY plays an important role in maintaining energy homeostasis. In response to energy needs or a change in energy status, ARC NPY acts as an orexigenic peptide to modulate food intake and energy balance. Particularly, ARC NPY serves as one of the downstream mediators of leptin’s actions. In contrast, DMH NPY acts independently of leptin. In addition to its feeding effect, DMH NPY has specific actions on brown adipocyte formation and thermoregulation. Knockdown of NPY in the DMH promotes development of brown adipocytes in WAT and elevates BAT activity, leading to increased energy expenditure and lowered fat contents. DMH NPY knockdown prevents high-fat diet-induced obesity. Peripheral NPY also directly acts on adipose tissue and promotes white adipogenesis. Overall, NPY plays critical roles in the control of energy balance through affecting food intake, body adiposity, and energy expenditure. These actions provide potential target(s) for combatting obesity and diabetes.
References
Adrian TE, Allen JM, Bloom SR, Ghatei MA, Rossor MN, Roberts GW, Crow TJ, Tatemoto K, Polak JM. Neuropeptide Y distribution in human brain. Nature. 1983;306:584–6.
Ahima RS, Prabakaran D, Mantzoros C, Qu D, Lowell B, Maratos-Flier E, Flier JS. Role of leptin in the neuroendocrine response to fasting. Nature. 1996;382:250–2.
Allen YS, Adrian TE, Allen JM, Tatemoto K, Crow TJ, Bloom SR, Polak JM. Neuropeptide Y distribution in the rat brain. Science. 1983;221:877–9.
Bamshad M, Aoki VT, Adkison MG, Warren WS, Bartness TJ. Central nervous system origins of the sympathetic nervous system outflow to white adipose tissue. Am J Physiol. 1998;275:R291–9.
Barbatelli G, Murano I, Madsen L, Hao Q, Jimenez M, Kristiansen K, Giacobino JP, De Matteis R, Cinti S. The emergence of cold-induced brown adipocytes in mouse white fat depots is determined predominantly by white to brown adipocyte transdifferentiation. Am J Physiol Endocrinol Metab. 2010;298:E1244–53.
Bartelt A, Bruns OT, Reimer R, Hohenberg H, Ittrich H, Peldschus K, Kaul MG, Tromsdorf UI, Weller H, Waurisch C, et al. Brown adipose tissue activity controls triglyceride clearance. Nat Med. 2011;17:200–5.
Bartness TJ, Bamshad M. Innervation of mammalian white adipose tissue: implications for the regulation of total body fat. Am J Physiol. 1998;275:R1399–411.
Beck B. Neuropeptide Y in normal eating and in genetic and dietary-induced obesity. Phil Trans Roy Soc London B Biol Sci. 2006;361:1159–85.
Bewick GA, Gardiner JV, Dhillo WS, Kent AS, White NE, Webster Z, Ghatei MA, Bloom SR. Post-embryonic ablation of AgRP neurons in mice leads to a lean, hypophagic phenotype. FASEB J. 2005;19:1680–2.
Bi S. Role of dorsomedial hypothalamic neuropeptide Y in energy homeostasis. Peptides. 2007;28:352–6.
Bi S, Ladenheim EE, Schwartz GJ, Moran TH. A role for NPY overexpression in the dorsomedial hypothalamus in hyperphagia and obesity of OLETF rats. Am J Physiol Regul Integr Comp Physiol. 2001;281:R254–60.
Bi S, Robinson BM, Moran TH. Acute food deprivation and chronic food restriction differentially affect hypothalamic NPY mRNA expression. Am J Physiol Regul Integr Comp Physiol. 2003;285:R1030–6.
Billington CJ, Briggs JE, Grace M, Levine AS. Effects of intracerebroventricular injection of neuropeptide Y on energy metabolism. Am J Physiol. 1991;260:R321–7.
Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004;84:277–359.
Chao PT, Yang L, Aja S, Moran TH, Bi S. Knockdown of NPY expression in the dorsomedial hypothalamus promotes development of brown adipocytes and prevents diet-induced obesity. Cell Metab. 2011;13:573–83.
Clark JT, Kalra PS, Crowley WR, Kalra SP. Neuropeptide Y and human pancreatic polypeptide stimulate feeding behavior in rats. Endocrinology. 1984;115:427–9.
Colmers WF, Wahlestedt C. The biology of neuropeptide Y and related peptides. New Jersey: Humana; 1993.
Cone RD. Studies on the physiological functions of the melanocortin system. Endocr Rev. 2006;27:736–49.
Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng YH, Doria A, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med. 2009;360:1509–17.
Dimicco JA, Zaretsky DV. The dorsomedial hypothalamus: a new player in thermoregulation. Am J Physiol Regul Integr Comp Physiol. 2007;292:R47–63.
Egawa M, Yoshimatsu H, Bray GA. Neuropeptide Y suppresses sympathetic activity to interscapular brown adipose tissue in rats. Am J Physiol. 1991;260:R328–34.
Elmquist JK, Elias CF, Saper CB. From lesions to leptin: hypothalamic control of food intake and body weight. Neuron. 1999;22:221–32.
Erickson JC, Clegg KE, Palmiter RD. Sensitivity to leptin and susceptibility to seizures of mice lacking neuropeptide Y. Nature. 1996a;381:415–21.
Erickson JC, Hollopeter G, Palmiter RD. Attenuation of the obesity syndrome of ob/ob mice by the loss of neuropeptide Y. Science. 1996b;274:1704–7.
Flier JS. Obesity wars: molecular progress confronts an expanding epidemic. Cell. 2004;116:337–50.
Fredholm BB, Karlsson J. Metabolic effects of prolonged sympathetic nerve stimulation in canine subcutaneous adipose tissue. Acta physiologica Scandinavica. 1970;80:567–76.
Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature. 1998;395:763–70.
Gardiner JV, Kong WM, Ward H, Murphy KG, Dhillo WS, Bloom SR. AAV mediated expression of anti-sense neuropeptide Y cRNA in the arcuate nucleus of rats results in decreased weight gain and food intake. Biochem Biophys Res Commun. 2005;327:1088–93.
Giraudo SQ, Kotz CM, Grace MK, Levine AS, Billington CJ. Rat hypothalamic NPY mRNA and brown fat uncoupling protein mRNA after high-carbohydrate or high-fat diets. Am J Physiol. 1994;266:R1578–83.
Gray TS, Morley JE. Neuropeptide Y: anatomical distribution and possible function in mammalian nervous system. Life Sci. 1986;38:389–401.
Gropp E, Shanabrough M, Borok E, Xu AW, Janoschek R, Buch T, Plum L, Balthasar N, Hampel B, Waisman A, et al. Agouti-related peptide-expressing neurons are mandatory for feeding. Nat Neurosci. 2005;8:1289–91.
Guan XM, Yu H, Trumbauer M, Frazier E, Van der Ploeg LH, Chen H. Induction of neuropeptide Y expression in dorsomedial hypothalamus of diet-induced obese mice. Neuroreport. 1998a;9:3415–9.
Guan XM, Yu H, Van der Ploeg LH. Evidence of altered hypothalamic pro-opiomelanocortin/ neuropeptide Y mRNA expression in tubby mice. Brain Res Mol Brain Res. 1998b;59:273–9.
Handschin C, Spiegelman BM. Peroxisome proliferator-activated receptor gamma coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr Rev. 2006;27:728–35.
Himms-Hagen J, Cui J, Danforth Jr E, Taatjes DJ, Lang SS, Waters BL, Claus TH. Effect of CL-316,243, a thermogenic beta 3-agonist, on energy balance and brown and white adipose tissues in rats. Am J Physiol. 1994;266:R1371–82.
Himms-Hagen J, Melnyk A, Zingaretti MC, Ceresi E, Barbatelli G, Cinti S. Multilocular fat cells in WAT of CL-316243-treated rats derive directly from white adipocytes. Am J Physiol Cell Physiol. 2000;279:C670–81.
Hummel KP, Dickie MM, Coleman DL. Diabetes, a new mutation in the mouse. Science. 1966;153:1127–8.
Ingalls AM, Dickie MM, Snell GD. Obese, a new mutation in the house mouse. J Hered. 1950;41:317–8.
Jimenez M, Barbatelli G, Allevi R, Cinti S, Seydoux J, Giacobino JP, Muzzin P, Preitner F. Beta 3-adrenoceptor knockout in C57BL/6J mice depresses the occurrence of brown adipocytes in white fat. Eur J Biochem. 2003;270:699–705.
Kawaguchi M, Scott KA, Moran TH, Bi S. Dorsomedial hypothalamic corticotropin-releasing factor mediation of exercise-induced anorexia. American journal of physiology Regulatory, integrative and comparative physiology. 2005;288:R1800–1805.
Kesterson RA, Huszar D, Lynch CA, Simerly RB, Cone RD. Induction of neuropeptide Y gene expression in the dorsal medial hypothalamic nucleus in two models of the agouti obesity syndrome. Mol Endocrinol. 1997;11:630–7.
Kuo LE, Kitlinska JB, Tilan JU, Li L, Baker SB, Johnson MD, Lee EW, Burnett MS, Fricke ST, Kvetnansky R, et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nat Med. 2007;13:803–11.
Levine AS, Morley JE. Neuropeptide Y: a potent inducer of consummatory behavior in rats. Peptides. 1984;5:1025–9.
Lin S, Boey D, Herzog H. NPY and Y receptors: lessons from transgenic and knockout models. Neuropeptides. 2004;38:189–200.
Lundberg JM, Terenius L, Hokfelt T, Martling CR, Tatemoto K, Mutt V, Polak J, Bloom S, Goldstein M. Neuropeptide Y (NPY)-like immunoreactivity in peripheral noradrenergic neurons and effects of NPY on sympathetic function. Acta physiologica Scandinavica. 1982;116:477–80.
McMinn JE, Seeley RJ, Wilkinson CW, Havel PJ, Woods SC, Schwartz MW. NPY-induced overfeeding suppresses hypothalamic NPY mRNA expression: potential roles of plasma insulin and leptin. Regul Pept. 1998;75–76:425–31.
Morrison SF, Nakamura K. Central neural pathways for thermoregulation. Front Biosci. 2011;16:74–104.
Nagase I, Yoshida T, Kumamoto K, Umekawa T, Sakane N, Nikami H, Kawada T, Saito M. Expression of uncoupling protein in skeletal muscle and white fat of obese mice treated with thermogenic beta 3-adrenergic agonist. J Clin Investig. 1996;97:2898–904.
Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, Matsukura S. A role for ghrelin in the central regulation of feeding. Nature. 2001;409:194–8.
Petrovic N, Walden TB, Shabalina IG, Timmons JA, Cannon B, Nedergaard J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J Biol Chem. 2010;285:7153–64.
Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell. 1998;92:829–39.
Rosen ED, Sarraf P, Troy AE, Bradwin G, Moore K, Milstone DS, Spiegelman BM, Mortensen RM. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol cell. 1999;4:611–7.
Sanacora G, Kershaw M, Finkelstein JA, White JD. Increased hypothalamic content of preproneuropeptide Y messenger ribonucleic acid in genetically obese Zucker rats and its regulation by food deprivation. Endocrinology. 1990;127:730–7.
Schwartz MW, Woods SC, Porte Jr D, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature. 2000;404:661–71.
Seale P, Bjork B, Yang W, Kajimura S, Chin S, Kuang S, Scime A, Devarakonda S, Conroe HM, Erdjument-Bromage H, et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature. 2008;454:961–7.
Shi YC, Lin S, Castillo L, Aljanova A, Enriquez RF, Nguyen AD, Baldock PA, Zhang L, Bijker MS, Macia L, et al. Peripheral-specific y2 receptor knockdown protects mice from high-fat diet-induced obesity. Obesity. 2011;19:2137–48.
Smith MS. Lactation alters neuropeptide-Y and proopiomelanocortin gene expression in the arcuate nucleus of the rat. Endocrinology. 1993;133:1258–1265.
Sousa-Ferreira L, Garrido M, Nascimento-Ferreira I, Nobrega C, Santos-Carvalho A, Alvaro AR, Rosmaninho-Salgado J, Kaster M, Kugler S, de Almeida LP, et al. Moderate long-term modulation of neuropeptide Y in hypothalamic arcuate nucleus induces energy balance alterations in adult rats. PLoS One. 2011;6:e22333.
Stanley BG, Leibowitz SF. Neuropeptide Y injected in the paraventricular hypothalamus: a powerful stimulant of feeding behavior. Proc Natl Acad Sci U S A. 1985;82:3940–3.
Stanley BG, Kyrkouli SE, Lampert S, Leibowitz SF. Neuropeptide Y chronically injected into the hypothalamus: a powerful neurochemical inducer of hyperphagia and obesity. Peptides. 1986;7:1189–92.
Ste Marie L, Luquet S, Cole TB, Palmiter RD. Modulation of neuropeptide Y expression in adult mice does not affect feeding. Proc Natl Acad Sci U S A. 2005;102:18632–7.
Steppan CM, Bailey ST, Bhat S, Brown EJ, Banerjee RR, Wright CM, Patel HR, Ahima RS, Lazar MA. The hormone resistin links obesity to diabetes. Nature. 2001;409:307–12.
Stricker-Krongrad A, Cumin F, Burlet C, Beck B. Hypothalamic neuropeptide Y and plasma leptin after long-term high-fat feeding in the rat. Neurosci Lett. 1998;254:157–60.
Tatemoto K. Neuropeptide Y: complete amino acid sequence of the brain peptide. Proc Natl Acad Sci U S A. 1982;79:5485–9.
Tatemoto K, Carlquist M, Mutt V. Neuropeptide Y–a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature. 1982;296:659–60.
Tritos NA, Elmquist JK, Mastaitis JW, Flier JS, Maratos-Flier E. Characterization of expression of hypothalamic appetite-regulating peptides in obese hyperleptinemic brown adipose tissue-deficient (uncoupling protein-promoter-driven diphtheria toxin A) mice. Endocrinology. 1998;139:4634–41.
Uysal KT, Wiesbrock SM, Marino MW, Hotamisligil GS. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature. 1997;389:610–4.
van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, Schrauwen P, Teule GJ. Cold-activated brown adipose tissue in healthy men. N Engl J Med. 2009;360:1500–8.
Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, Taittonen M, Laine J, Savisto NJ, Enerback S, et al. Functional brown adipose tissue in healthy adults. N Engl J Med. 2009;360:1518–25.
Wallenius V, Wallenius K, Ahren B, Rudling M, Carlsten H, Dickson SL, Ohlsson C, Jansson JO. Interleukin-6-deficient mice develop mature-onset obesity. Nat Med. 2002;8:75–9.
Weiss B, Maickel RP. Sympathetic nervous control of adipose tissue lipolysis. Int J Neuropharmacol. 1968;7:395–403.
White JD, Kershaw M. Increased hypothalamic neuropeptide Y expression following food deprivation. Mol Cell Neurosci. 1990;1:41–8.
Wilding JP, Gilbey SG, Bailey CJ, Batt RA, Williams G, Ghatei MA, Bloom SR. Increased neuropeptide-Y messenger ribonucleic acid (mRNA) and decreased neurotensin mRNA in the hypothalamus of the obese (ob/ob) mouse. Endocrinology. 1993;132:1939–44.
Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, Khandekar M, Virtanen KA, Nuutila P, Schaart G, et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell. 2012;150:366–76.
Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y, Ide T, Murakami K, Tsuboyama-Kasaoka N, et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med. 2001;7:941–6.
Yang K, Guan H, Arany E, Hill DJ, Cao X. Neuropeptide Y is produced in visceral adipose tissue and promotes proliferation of adipocyte precursor cells via the Y1 receptor. FASEB J. 2008;22:2452–64.
Yang L, Scott KA, Hyun J, Tamashiro KL, Tray N, Moran TH, Bi S. Role of dorsomedial hypothalamic neuropeptide Y in modulating food intake and energy balance. J Neurosci. 2009;29:179–90.
Zarjevski N, Cusin I, Vettor R, Rohner-Jeanrenaud F, Jeanrenaud B. Chronic intracerebroventricular neuropeptide-Y administration to normal rats mimics hormonal and metabolic changes of obesity. Endocrinology. 1993;133:1753–8.
Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372:425–32.
Zhang L, Macia L, Turner N, Enriquez RF, Riepler SJ, Nguyen AD, Lin S, Lee NJ, Shi YC, Yulyaningsih E, et al. Peripheral neuropeptide Y Y1 receptors regulate lipid oxidation and fat accretion. Int J Obes. 2010;34:357–73.
Acknowledgments
This work was supported by the US National Institute of Diabetes and Digestive and Kidney Diseases Grants DK074269 and DK087888.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Bi, S. (2013). Role of NPY in Brown Adipocytes and Obesity. In: Cao, Y. (eds) Angiogenesis in Adipose Tissue. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-8069-3_9
Download citation
DOI: https://doi.org/10.1007/978-1-4614-8069-3_9
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-8068-6
Online ISBN: 978-1-4614-8069-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)