
Abstract
The contribution of the Galapagos Islands to increasing our understanding of evolution can hardly be surpassed by any other place of similar size on Earth. Evolutionary biology research on the Galapagos spans studies on the origins of life to molecular phylogeny and evolutionary developmental biology. Most pioneering research conducted on the Galapagos was basic descriptive natural history that focused on the evolutionary ecology of endemic species, with an emphasis on the patterns and mechanisms of adaptation, geographic distribution, and taxonomic affinities, based mostly on appearance and behavior. Modern research, however, such as that conducted by Peter and Rosemary Grant and several other scientists, has made the transition from descriptive patterns to a more quantitative and scientific orientation that features the testing of hypotheses about the evolution and conservation of Galapagos organisms and ecosystems. Now, the study of speciation and adaptive radiation, two common evolutionary processes found on oceanic archipelagos like the Galapagos, can be tracked in time and space, owing to the development of modern molecular genetic techniques and its application to the study of phylogenies (molecular phylogeny) and biography (phylogeography). Island ecosystems and organisms offer a unique opportunity for the study of the fundamental principles of evolution, including natural selection, adaptation, and speciation. Both the Galapagos National Park, as the authority, and scientists deserve much recognition for maintaining the manipulation of organisms and ecosystems at a minimal level to preserve the systems under study. Here, the most relevant subjects on the principles and mechanisms of evolutionary biology are succinctly addressed by research studies conducted in the Galapagos Islands. By no means does the review include all of the fascinating studies that have been conducted since Darwin’s memorable visit to the Galapagos 174 years ago on 16 September 1835.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
Darwin’s Theory of Evolution, more than any other scientific theory, has changed our fundamental understanding of the living world and our own conception of human nature. It is no surprise that it has been considered the most revolutionary idea in modern science (Mayr 1982). The Galapagos Islands played a central role in the development of science, particularly for the field modernly known as evolutionary biology. It was mainly his observations on the species of this isolated group of islands in the Pacific that later led Darwin to doubt the chief concept of the time on the immutability of species and to reflect and develop his ideas about the origin of species and, subsequently, his theory of evolution by natural selection (Sulloway 1982). Ever since Darwin’s memorable trip to the Galapagos Islands 175 years ago in September 1835, and the publication of his theory in 1859, a large number of naturalists and scientists have visited the Galapagos eager to test Darwin’s observations and theory. The Swiss-American naturalist, Louis Agassiz of Harvard University, who visited the Galapagos in 1872; the German geologist Theodor Wolf, who visited twice, in 1875 and 1878; and George Bauer, who traveled there in 1891, are among the most renowned scientists in Darwin’s time to visit the Galapagos (Larson 2001). During the twentieth century, two of the best known expeditions were the ones led by William Beebe, in 1923 and 1925, and the California Academy of Sciences expedition in 1905–1906 led by Rollo Beck (Larson 2001; Quiroga 2009).
Scientific research in the Galapagos was pioneered by Lack (1947), Bowman (1963, 1979) and Eibl-Eibesfeldt (1958). Thereafter, several dozen naturalists and scientists from around the world have visited and conducted research on Galapagos on a wide variety of subjects. Some came to fulfill an academic requirement like a dissertation, while a few others made the Galapagos their veritable home (e.g., see Quiroga 2009). Among the latter, there are celebrities like Peter and Rosemary Grant, now at Princeton University, who have visited the Galapagos for several months every single year for nearly 40 years since 1973 when their study of Darwin’s finches began. Others include Tjitte de Vries, of the Universidad Católica del Ecuador, who pioneered research on the endemic Galapagos hawk (Buteo galapagoensis), now, in collaboration with Patty Parker of the University of Saint Louis; Fritz Trillmich, of the University of Bielefeld, Germany, who researched the Galapagos fur seal (Arctocephalus galapagoensis) and the Galapagos sea lion (Zalophus wollebaeki); Andrew Laurie, whose work on the marine iguana (Amblyrhynchus cristatus) was continued and expanded by Martin Wikelski of the University of Konstanz; Tom Fritts, who researched the giant tortoises; Howard and Heidi Snell, who worked on land iguanas (Conolophus subcristatus and C. pallidus) and lava lizards (Microlophus spp.); and David Anderson, who carried out research on siblicide of the Nazca Booby (Sula granti) and the ecology of the Galapagos boobies (Sula spp.). Lately, a new generation of studies that takes advantage of modern molecular techniques has led to an explosion of phylogenetic and phylogeographic studies that are clarifying our understanding of ecological and evolutionary processes, particularly, the processes and mechanisms of speciation, both in time and space.
The Galapagos Archipelago is one of the most studied places on Earth, even though much of its species’ ecological and evolutionary processes still remain unknown. In the marine realm, very little is known about the intertidal, subtidal, and pelagic species and populations, as well as their communities’ and ecosystems’ ecology and evolutionary processes. The terrestrial biota has been better studied, but, even so, the knowledge base is strongly biased toward the most conspicuous and charismatic species, such as reptiles and some groups of birds. Among plants, only a few flowering plants have been relatively well studied, while very little is known about cryptic species such as cryptogamic plants (especially mosses and lichens).
Here, I briefly summarize some of the most relevant aspects of the natural history of the Galapagos by pointing out a few selected case studies that describe the most important research findings in different fields of the natural sciences studies conducted there. This mini review also addresses the conservation status and threats as well as the conservation achievements in the islands.
Geological Research
Darwin can be considered the first geologist to observe and describe the geology of the Galapagos and to correctly affirm their geological youth and volcanic origin (emerging from the ocean) as oceanic islands that had not been in contact with continental landmasses. Referring to the geology of the Galapagos, he once wrote, “We are led to believe that within a period, geologically recent, the unbroken ocean was here spread out.” Modern geological research—aside from petrology and geochemical studies of basalts (e.g., Williams 1966; Williams and McBirney 1979; McBirney and Williams 1969; Swanson et al. 1974) and volcanology (Simkin and Howard 1970; Simkin 1984)—has focused mainly on the origin and age of the islands using several dating techniques including paleomagnetic polarities (geomagnetic reversals), geomorphology analyses, and radioactive potassium–argon dating (Cox 1971, 1983; Geist 1996). Modern radiometric work estimated that Española Island is about 3.3 million years old (Bailey 1976) and that the oldest islands of the archipelago range from about 2 to 3 million years old (Geist 2009).
Estimates of the ages of individual islands vary a lot, but the latest studies estimate the age of the oldest current extant islands (i.e., San Cristobal or Española) at about 3.5 million years and only about 60,000–300,000 years for Fernandina Island (Bailey 1976; Geist 1996). Once again, Darwin was correct when he inferred that the islands emerged from the unbroken ocean, as their origin is currently explained by hot spot and plate tectonic theories. Plate tectonics—the theory that continents move—was first proposed by de Candolle, a French biogeographer, who was originally discredited for his idea. However, it was formally proposed by Alfred Wegener in 1912 and was only gradually accepted after nearly 50 years, mostly due to paleomagnetic evidence. The hot spot theory, on the other hand, is a recent one and refers to a huge and extremely hot solid, but plastic, column of rock that probably rises from the deep mantle due to radioactive enrichment (and radioactive heat). The column (mantle plume) rises due to thermal buoyancy, melts near the surface owing to decompression, and breaks the Earth’s crust from beneath, giving rise to a shield volcano like those found in the Galapagos, Hawaii, and other oceanic archipelagos.
The Galapagos settles over the Nazca plate that moves eastward to South America and runs under the South American continental plate along the so-called subduction zone. The islands travel from their center of origin (the Galapagos hot spot under Fernandina Island) at a variable rate between 2 and 7 cm per year. That has created a rough age gradient with a cluster of oldest islands in the east, a cluster of middle-aged islands in the center, and still another cluster of the youngest islands in the west of the archipelago (Simkin 1984). Although most of the Galapagos Islands owe their origin to the mantle hot spot, the origin of the two northernmost islands (Darwin and Wolf) and the three northwestern islands (Pinta, Marchena, and Genovesa) is likely related to the Galapagos ridge, the Nazca-Cocos plates spreading zone, which is located north of the Galapagos Islands (Geist 2009). Current marine geological exploration (Christie et al. 1992) has corroborated the hypotheses, first advanced by biologists (Wyles and Sarich 1983), that the older sunken Galapagos Islands may have been in existence for at least 10 million years. By now, most geologists and biologists working in the Galapagos readily accept that as the Nazca plate moves eastward and the islands travel away from the hot spot, they decrease in altitude—apparently owing to the cooling and contraction of the crust—and eventually subside (Geist 2009). Recent evidence about the genetic distance between the marine and land iguanas, and especially the new species of land iguana (Gentile et al. 2009), provides further support for the currently drawn Galapagos Islands and an older origin of the archipelago.
Climate and Oceanographic Research
The Galapagos Islands are located where several marine currents converge, modifying what should be basically a tropical climate into a predominantly dry region during most of the year (Palmer and Pyle 1966). Such a geographical and oceanographic setting makes these islands attractive for oceanographers and climate scientists interested in the study of Earth’s paleoclimate and for those attempting to understand and predict current climatic phenomena with a global impact, such as the El Niño–Southern Oscillation (ENSO) (e.g., Wyrtki 1975; Houvenaghel 1984; Cane 1983). The El Niño event, and its usual counterpart La Niña, affect the terrestrial and marine Galapagos biota in particular ways. During the conditions associated with El Niño (high sea surface temperature and a heavy rainy season), terrestrial organisms feast, breed freely, improve survival, and increase population sizes, while marine organisms fast and stop breeding, die, and decline in population. However, the reverse is true during the drop in sea surface temperature and drought conditions associated with La Niña (e.g., see Robinson and del Pino 1985).
Global-scale research has tremendously advanced our understanding of how the Earth’s climate has changed over millions of years. Astronomic evidence and glacial–interglacial cycles, as well as local changes in landmass connectivity associated with plate tectonics, attest to the major climatic changes that the Earth has undergone on a global scale (e.g., Zachos et al. 2001; Ferodov et al. 2006). Thus, the opening and closing of the Isthmus of Panama had major consequences on the patterns of atmospheric and ocean circulation in the eastern Pacific (Cronin and Dowsett 1996). From these studies, it has been inferred how the climate of the Galapagos has changed over the last 10 million years from when the islands had a warmer and more humid tropical climate as compared to today.
Furthermore, studies conducted on the Galapagos Islands themselves have also improved our understanding of the climate both in these islands and at larger geographic scales, including the eastern Pacific. Most paleoclimate research on the Galapagos is based on analyses of sediment cores from a number of lakes but mainly at Junco Lake on San Cristobal Island, Lake Arcturus on Genovesa Island, and the lake on Bainbridge Rock islet near Santiago Island. Pollen stratigraphy and geochemical and mineralogical analyses of sediment cores and C14 dating (Colinvaux 1968, 1972; Colinvaux and Schofield 1976a, b), carbon/nitrogen ratios, and isotopic hydrogen and oxygen analyses (Riedinger et al. 2002) have opened a window that has allowed us to picture the history of climatic changes in the Galapagos and the whole eastern Pacific Ocean. These studies provide evidence that during the last 50,000 years, the climate of the Galapagos has undergone profound changes in temperature and precipitation, in association with the Northern Hemisphere ice-age cycles. The islands were dry during glaciations and humid and rainy during interglacial periods, such as the present (Colinvaux 1984), and the frequency of El Niño events started increasing during the last 2,500 years, particularly in the last 1,000 years (Riedinger et al. 2002; Conroy et al. 2008). Research on lake sediments continues today with modern and improved devices and methods; research on El Niño and the climate dynamics of the Galapagos has also been tracked by studies of marine organisms including coral (Shen et al. 1992; Urban et al. 2000) and foraminifera (Lea et al. 2006). One of the major effects of climate change associated with glacial and interglacial cycles in the Galapagos relates to changes in sea levels. Sea level rises (interglacial) and falls (glacial) of over 100 m (Lambeck and Chappell 2001) are presumed to have affected levels of isolation, as well as the size and shape of the islands (Grant and Grant 2008); changes, that in turn, are expected to have affected evolutionary patterns and processes. In summary, global local-scale climate studies have shown that climate of the Galapagos has been highly dynamic and has changed dramatically over geological and evolutionary time scales.
Research on Ecology and Evolutionary Biology
Research on Ecology
Ecological research in the Galapagos is mainly of autoecological character and has focused on geographic distribution, demography, and the behavioral ecology of the most conspicuous species of terrestrial vertebrates (Bowman 1984; Clark 1984; Eibl-Eibesfeldt 1984a, b; de Vries 1984; Grant 1984, 1999; Grant and Grant 2008; Harris 1984; Trillmich 1984; de Roy 2009) and a few species of vascular plants (Eliasson 1984; Porter 1984; Tye 2007, 2008). Even so, several gaps remain regarding the ecology of these relatively well-studied groups, and even less is known about most species of terrestrial invertebrates and most vascular and nonvascular plants, as well as most marine organisms. Furthermore, except for a few marine studies (Withman and Smith 2003; Edgar et al. 2004) and a handful of terrestrial organisms (Abbott and Abbott 1978; Schluter 1986; Schluter and Grant 1984; Schluter et al. 1985; Grant and Grant 2006), ecological research in the Galapagos has severely neglected community ecology at the ecosystem level, with almost a complete lack of in-depth studies about interspecific interaction at the community level and its role on the structuring of natural communities. Thus, we know very little about ecological processes such as pollination, seed dispersal, symbiotic interactions (e.g., parasitism, commensalism), competition, predation, decomposition, predator-/herbivore-mediated coexistence, and the occurrence and role of ecological guilds.
Research on Evolutionary Biology
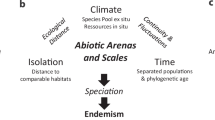
A volcanic, highly isolated, oceanic archipelago like the Galapagos that was never connected to the mainland is expected to have been devoid of terrestrial life when it emerged from the ocean. The terrestrial life now inhabiting these islands, as well as those organisms now extinct, got there from somewhere else and became rapidly and effectively isolated from their parental populations. These two aspects of colonization and further isolation have profoundly shaped the ecological and evolutionary history of the Galapagos organisms through a three-step process involving (1) organisms’ arrival, (2) establishment (colonization) in their new habitat, and (3) in situ evolution (i.e., adaptive and nonadaptive genetic and phenotypic divergence, speciation, evolutionary radiation).
Arrival of Organisms to the Archipelago
Arrival requires transportation to the islands from somewhere else, and 1,000 km of ocean represents a major ecological barrier for a vast number of organisms. A key factor for dispersal and arrival was the organisms’ vagility (dispersal ability), which is dependent upon their intrinsic dispersal abilities (i.e., production of small propagules that disperse easily or are easy to transport by winds or by animals) and their ability to survive a long journey through the ocean. Organisms reached the islands by three means of dispersal: through the ocean, either by floating and drifting with the ocean currents (passive dispersal) or swimming (active dispersal); through the air, taking advantage of the trade winds blowing from southeast and northwest; or by attaching themselves to other organisms (e.g., birds). Many organisms, including several types of invertebrates (snails, arthropods) and vertebrates (lizards, snakes, and even poor-flying birds like rails), may have reached the islands on those masses of vegetation that usually get to the ocean when continental rivers overflow during the rainy season and then drift into the open ocean. Duncan Porter (1976) inferred the mean of transportation for the 378 indigenous ancestral taxa that colonized the Galapagos and suggests that birds may have been the vector for about 60% (of these, 64% in the digestive tract, 21% attached to feathers, and 15% attached to mud on the legs), while 32% were transported by the wind and 9% by the sea.
Establishment of Organisms in the Archipelago
Getting to the Galapagos was only part of the process; colonization required the establishment of a viable population. It can be inferred that many successful arrivals did not lead to a successful colonization due to a number of factors, including (1) the ecological successional stage of the island, (2) reproductive viability, and (3) demographic (population) viability. Many organisms may have been prevented from settling after not finding a suitable habitat upon arrival. For example, as we can see today on a barren lava flow, only a few pioneer species (e.g., lichens and few other plants) would be expected to have established themselves first, while the more habitat-demanding species would have had to wait for conditions that were more suitable. For organisms with strict sexual reproduction (most animals and monoecious plants), an unavoidable condition for establishment was arriving with a mate, or in a small flock with individuals of both sexes (as may be the case of the birds), or a fertilized female. Populations that are too small and isolated, such as those that just have colonized a remote archipelago from one single flock, a fertilized female, or a single asexual individual, run a high risk of extinction due to stochastic factors (including genetic, demographic, and environmental), as well as the occurrence of natural catastrophes (Lande 1980; Lande and Barrowclough 1987).
Evolution in the Galapagos Archipelago
Perhaps the most prevalent biological feature of isolated oceanic archipelagos is the high level of endemism among every native taxonomic group. Insular endemics evolve from a mainland-colonizing ancestor either through linear evolution, when a colonizing ancestor transforms into a new insular species, or through an evolutionary radiation, when either an ancestral colonizer or an insular endemic splits further into several new species.
In the Galapagos Islands, most species that colonized the archipelago have evolved at least into single endemic taxa including subspecies, species, and genera (see Baert 2000; Peck 1996; Tye et al. 2002). Endemics at the species level included mammals (∼88%), birds (52%), reptiles (100%), fishes and algae (∼20–30%) insects (47%), other terrestrial invertebrates (53%) and vascular plants (32%), bryophytes (mosses, liverworts; ∼10%), lichens (∼7%), and pteridophytes (ferns; ∼4%). Examples of a single colonizing ancestor evolving into a new species (an insular endemic) abound, including the two species of sea lions, the Galapagos penguin (Spheniscus mendiculus), the flightless cormorant (Phalacrocorax harrisi), and several others. A lower number of colonizers evolved even further and underwent evolutionary radiation when a single colonizing species evolved into several new endemic species. The best examples of evolutionary radiation, most of them derived from a single colonizing event (i.e., monophyletic groups), include land snails of the genus Bulimulus (71 species, all endemic, perhaps the most spectacular example of adaptive radiation), Darwin’s finches (15 species), and giant tortoises (originally about 15 species). Among plants, the genus Scalesia (Asteraceae; 15 species, 19 taxa including subspecies and varieties) and Alternanthera (Amaranthaceae; 14 species and 20 taxa), Opuntia (Cactaceae; six species and 14 varieties, from two independent colonizations).
The geological youth of the Galapagos Archipelago, together with the high level of endemism among all taxonomic groups that colonized it, implies that evolution in the Galapagos has generally proceeded rapidly particularly among those species that have evolved into different species through the process of linear evolution. The rate of evolution (speciation) has been even more dramatic among those groups that have radiated into many species from a single ancestor that colonized the islands.
The process of colonization and evolution of all organisms in the Galapagos fits nicely into the founder effect speciation model first suggested by Huxley (1938) and formally proposed by Mayr (1954). That is, a new species arises when a new population is founded in a remote isolated place, usually by a small group on immigrants. Under such a model, theoretically, there are a number of ecological and genetic factors and processes driving rapid evolution in an isolated archipelago like the Galapagos. These factors and processes include founder effect, genetic drift, divergent selection, interrupted or restricted gene flow, multiple isolation and ecological opportunity.
Founder Effect: The role of the founder effect in immediately driving genetic divergence and subsequent phenotypic divergence between a founding (island) and a parental (continental) population is expected to be high; few individuals will bring, at best, only a small fraction of the whole genetic variation from a large parental population simply due to random genetic sampling error. Furthermore, a very small number of founding individuals may not necessarily be a representative sample of the whole parental population (e.g., Huxley 1938). A genetically nonrepresentative sample means that the island’s founding population will start with a biased sample of alleles relative to the continental population. All of this suggests that a new insular founding population is likely to diverge from its continental parental population from the very beginning. Founder effects are also expected to lead to inbreeding, due to the small number of colonists. Inbreeding is predicted to lead to increased homozygosis in the population, which would potentially lead to inbreeding depression, thus increasing the probability that the founding population could become extinct. Only those populations that managed to go through such a demographic bottleneck and survive were the ones that become established (colonized) in the Galapagos. Such populations, at least on theoretical grounds, may be able to survive, evolve, and remain well adapted through a coadapted set of genes; for them, outbreeding could be deleterious in the sense that it would break the coadapted set of genes thus reducing or wiping out their adaptive value. Over time, however, most populations are expected to increase in number and in genetic variation.
Owing to the development of molecular genetics, studies only recently started to address this issue, as well as that of the role of genetic drift in the process of evolution in the Galapagos. The few studies addressing this issue seem to indicate that some species may in fact have started with extremely low numbers (i.e., a founder effect genetic signature has been found). On Daphne Major Island, Grant and Grant (2008) observed and neatly tracked a founder event of a population of the large ground finch (G. magnirostris) that had colonized the islet during the last 25 years.
Genetic Drift: Genetic drift, a random fluctuation of allele frequencies leading to an eventual random loss/fixation of alleles in a population, is expected to play an important role in the evolution of small and isolated populations (Wright 1931, 1932). The Galapagos’ recently founded populations that have experienced a demographic bottleneck due to a founder effect would constitute an ideal scenario for drift to occur. Thus, colonizing alleles would become lost/fixed at a faster rate, leading to rapid further genetic divergence between the island and the continental population. Such a stochastic evolutionary process would result in nonadaptive evolution, which is theoretically expected to be a common pattern at the onset of colonization in a place like the Galapagos. Genetic erosion or loss of alleles (a consequence of drift) and inbreeding (a consequence of a founder effect) are expected to lead to increased homozygosis in the population, perhaps contributing in turn to inbreeding depression and increasing chance of extinction.
Divergent Selection: Isolated populations are expected to diverge rapidly as a result of divergent natural selection for local adaptation. Selection pressures usually vary geographically and doubtless are expected to differ greatly between the continent and a new colonized insular habitat, as well as from island to island. Selection, acting upon a biased and impoverished genetic variation (due to founder effect and genetic drift), as well as upon new alleles (i.e., originated by independent mutation) and new genotypes (due to recombination), will rapidly or gradually lead to genetic and phenotypic divergence between insular and continental populations. Such divergence is expected to happen rapidly among small and isolated populations or rather gradually among much larger populations.
Interrupted or Restricted Gene Flow: Gene flow is a homogenizing evolutionary force that prevents conspecific populations from diverging due to local adaptation. Therefore, if gene flow became effectively interrupted or severely restricted due to geographical isolation, genetic divergence between source (continental) and founder (island) population would be rapid. Recent molecular genetics and ecological inferences suggest that, in this sense, the almost 1,000 km of open ocean have acted as an effective ecological barrier preventing gene flow for most of the terrestrial organisms that colonized the Galapagos. Founder effect, genetic drift, and divergent selection, aided with lack of gene follow, will result in genetic and phenotypic divergence between conspecific populations, eventually leading to speciation and the origin of endemic taxa (species or subspecies).
Multiple Isolation: Archipelagos, as opposed to single islands, present opportunities for multiple and repeated events of dispersal and colonization followed by further isolation (an archipelago effect). This means that within archipelagos, a natural evolutionary experiment of colonization of one island from the mainland, as described above, is replicated every time that a species already present in the archipelago disperses and colonizes a new island. Such a combination of ecological and genetic factors and processes repeated several times on different islands would be another ingredient for rapid evolutionary diversification of a lineage into several endemic taxa (genera, species, subspecies). The Galapagos, where single colonization events have led to evolutionary radiation in a number of endemic organisms (e.g., Darwin’s finches, bulimulid land snails, Scalesia plants, and Opuntia cacti), fits these theoretical expectations very closely.
Ecological Opportunity: Oceanic archipelagos that were originally devoid of terrestrial life, like the Galapagos, will only gradually fill with species that arrive and establish on the islands. Insular ecological communities, therefore, are expected to be less packed than continental ones, especially at the early and middle stages of their ecological succession. These more relaxed ecological communities will have a number of empty ecological niches that will not only favor continuous colonization of new species (McArthur and Wilson 1967) but will also allow already established species the opportunity to explore and, in some cases, eventually shift into a new ecological niche. Species that show large genetic variation for morphological and behavioral adaptive traits, such as Darwin’s finches (e.g., Grant and Grant 1989, 2008), will be more likely to undertake such an evolutionary path and thus radiate into several new species, each one adapted to a particular ecological niche (Lack 1947; Grant 1999; Grant and Grant 2008).
On theoretical grounds, in isolated archipelagos like the Galapagos, genetic drift has the potential to be a relevant factor in evolution, particularly following a founder event. Thereafter, because the population increases in size or becomes less isolated within the archipelago, the effect of drift as an evolutionary force is expected to decline, while natural selection is likely to catch up and become the chief mechanism of further divergence. This does not imply that natural selection may not be an important evolutionary force from the very beginning following a founder event.
In the past, evolutionary ecologists usually assumed that all or most speciation resulted from Darwinian (adaptive) evolution and quickly accepted most evolutionary radiations as putative examples of adaptive radiation. Most modern-day evolutionary biologists would accept that a combination of stochastic processes (i.e., drift and founder effect) and adaptive (Darwinian selection) mechanisms are usually involved in evolution. The relative importance of natural selection and stochastic processes, however, is still a matter of debate on both theoretical and empirical grounds. Whether evolutionary radiations are mainly adaptive or the product of stochastic process still deserves in-depth research in the Galapagos and elsewhere.
Some divergent traits among closely related species [e.g., morphological and behavioral traits such as song and beak size and shape in Darwin finches (Lack 1947; Bowman 1979; Grant 1999; Grant and Grant 2008)] show a clear adaptive function and evolutionary divergence. Speciation among those lineages can safely be attributed to the role of selection although that does not imply that the role of drift at some point on their evolutionary history should be disregarded. The adaptive value of other traits is less obvious since there is not an apparent function for survival and their evolution may be either the result of sexual selection or genetic drift. Geographical and interspecific divergence at the molecular level (i.e., DNA sequences) is regarded to be mainly the result of genetic drift, and, although the issue is still controversial, new molecular techniques and statistical methods have started to reveal evidence for selection at this level.
Most biological research on the Galapagos has been centered on the evolutionary ecology of vertebrates and a few invertebrates and plants. The evolutionary biology of marine organisms largely remains an unexplored field. Modern molecular genetic techniques have prompted a new generation of evolutionary studies in the Galapagos, including fields such as population genetics, phylogenetics, phylogeography, and evolutionary developmental biology. These new and recent studies are shedding light on the patterns of genetic variation within and between populations, information that is important for conservation. These studies are also clarifying the taxonomic status of many different taxa, such as the taxonomic position of each species (previously considered as subspecies) and the finding of cryptic taxa of giant tortoises. Just to list a few, the taxonomic positions of species whose Galapagos populations are now accepted as full new and endemic species include the Nazca booby (Sula granti), the Galapagos petrel (Pterodroma phaeopygia), the Galapagos shearwater (Puffinus subalaris), and the green heron (Butorides striatus). More than that, these studies are allowing the exploration of the evolutionary history of Galapagos organisms, both in space and time. These studies have begun to clarify more accurately several issues that were previously difficult to track, such as the ancestral species and geographical origins of the different native species, especially of those that have radiated. We have still a long way to go, but thanks to molecular genetic studies, at least we have gained a better understanding of the historical evolution of Darwin’s finches (Petren et al. 1999, 2005; Sato et al. 2001a, b), Galapagos mockingbirds (Arbogast et al. 2006; Hoeck et al. 2010), Galapagos hawk (Bollmer et al. 2005, 2006, 2007; Whiteman et al. 2007; Hull et al. 2008; Parker 2009a, b), Galapagos cormorant (Kennedy et al. 2009; Duffie et al. 2009), giant tortoises (Caccone et al. 1999, 2002), land and marine iguanas (Wyles and Sarich 1983; Rassmann 1997; Gentile et al. 2009), lava lizards (Lopez et al. 1992; Jordan et al. 2002; Kizirian et al. 2004; Jordan and Snell 2008), bulimulid land snails (Parent and Crespi 2006; Parent et al. 2008), and a few others (Sequeira et al. 2000; Schmitz et al. 2007).
Biological Conservation
Oceanic archipelagos and island ecosystems, in general, are highly vulnerable to disturbance, especially to invasion by exotic (introduced) organisms (Crawley 1987). Such a high level of vulnerability is likely explained by a history of evolution in isolation from the mainland (i.e., insular organisms evolved free from major and diversified competition, predation, and disease) that has resulted in their ecosystems’ low resistance and low resilience (Carlquist 1965; Connell and Sousa 1983). Other causes for their ecosystems’ fragility may be explained by a pattern of low species diversity, low complexity and demographic factors, including small population size and restricted distribution range for a large number of island species and the existence of vacant niches (Connell and Sousa 1983; Herbold and Moyle 1986; Mace and Lande 1991).
The Galapagos Archipelago fully fits these generalizations of being ecologically fragile. The islands’ ecosystems are species-poor and simple (Snell et al. 2002); however, relative to its size, the Galapagos’ contribution to global biodiversity is high, due to a high endemism among all taxa. Also, the Galapagos biota evolved largely in isolation from mainland South America with very low presumed rates of natural arrival and colonization of species over their geological history. On the demographic side, a large number of native species of plants and animals have an extremely small population size and/or distributions restricted to a single island or even to only a small area within an island. The population size of several of these species, as well as their distribution areas, has been further reduced due to current threats.
Conservation Threats
The Galapagos’ ecosystems are under pressure on two fronts: terrestrial and marine, each one having its own peculiarities.
Terrestrial Ecosystems
The main threat to terrestrial ecosystems is the large and diversified number of exotic organisms (plant, animals, bacterial, and viral diseases), many of which have become invasive. Most of the introduced organisms are competitors, predators, parasites for native species, vectors, or reservoirs of diseases that later spread to organisms. The ecological impact of exotic organisms is worsened, because native species have not usually evolved immunological defenses against recently introduced diseases or developed behaviors and other life-history strategies to counteract the effects of exotic predators and competitors. The problems of exotic species in the Galapagos are further worsened because, while roads are the main means of dispersing exotic plants and animals, current development trends aim to build new roads across each inhabited island.
Exotic organisms have been introduced to the islands voluntarily or involuntarily by humans for 400 years, since pirates started using the islands regularly. The rate of introductions has worsened during last two decades, mainly due to a dramatic increase in the resident human population and the increased rate of transportation both within the archipelago and between the archipelago and the continent (Tye et al. 2008; Causton and Sevilla 2008). Contrasting with 112 introduced species recorded by 1900, the Charles Darwin Research Station and the Galapagos National Park confirmed the establishment of 748 species of vascular plants (cf. ∼500 native), 543 invertebrates (cf. ∼3,000 native), mostly insects (490 introduced cf. 1,555 native), and 30 vertebrates (Tapia et al. 2000; Roque-Albelo 2008; Causton and Sevilla 2008; Tye 2007, 2008; Tye et al. 2008) by 2007. Most remain in the place they were introduced on the five human-inhabited islands and within the colonized zones of these islands, but they are gradually spreading over much of the archipelago. By now, none of the 19 larger islands of the archipelago are free from introduced organisms.
The worst introduced species are the most invasive ones that rapidly spread from their center on introduction. Among these are hill blackberry (Rubus niveus) and guava (Psidium guajava), which are invading extensive areas of the humid zone on San Cristobal, Santa Cruz, Floreana, and Isabela Islands, and quinine (Cinchona pubescens), which is widespread on Santa Cruz (Tye et al. 2008). Their negative impact on animals is widely recognized (Causton and Sevilla 2008; Jiménez-Uzcátegui et al. 2008a, b). Goats (Capra hircus) compete for food with giant tortoises, causing the decline of a number of rare plants and provoking the decline of otherwise rather common plants, such as the cactus trees (Opuntia sp.) in islands like Santiago which, in turn, endangers the cactus finch (Geospiza scandens), which is highly dependent on the cactus for survival. Rats (black Rattus rattus; Norwegian, Rattus norvegicus), dogs (Canis familiaris), cats (Felis catus), and pigs (Sus scrofa) are very active and voracious predators, attacking insects (e.g., native beetles), land and marine birds and even nests and hatchlings of birds, and all reptiles, including giant tortoise hatchlings, lava lizards, land and marine iguanas, snakes, and sea turtles. The recently introduced ani (Crotophaga ani) spread through the archipelago during the El Niño of 1982–1983 (Valle, personal observations), likely causing the population to decline and perhaps even causing future extinctions of several land birds (e.g., the vermilion flycatcher), although this issue needs further investigation.
Introduced invertebrates seem to be the worst among the animals, because of their rapid spread and their apparent impact on the invertebrate and vertebrate community (Causton and Sevilla 2008; Roque-Albelo 2008; Fessl et al. 2006). Of primary concerns are two fire ants (Wasmannia auropunctata and Solenopsis geminata) and two wasps (Brachygastra lechiguana, Polistes versicolor). The cottony cushion scale (Icerya purchasi) spread rapidly along the coastal zone in Santa Cruz and caused the death of mangroves and several other plant species. The fly (Philornis downsi) infests birds in their nests. The potential for further introduction is remarkably high due to the high volume of organic products (mostly fruits and vegetables) brought to the islands every week as food supply for the local population and tourists.
Diseases pose a major threat for oceanic fauna, as exemplified in Hawaii, where more than one-half of native birds became extinct from avian malaria (Van Riper et al. 1988). In the Galapagos, introduced vertebrates, particularly birds and mammals, are known vectors or reservoirs of several viral, bacterial, and protozoan diseases (Vargas and Snell 1997; Miller et al. 2001; Wikelski et al. 2004). A viral disease carried by black rats was the most likely reason for the extinction of most Galapagos native rice rats. Viral and protozoan diseases are the subject of an investigation started by the Charles Darwin Foundation (Vargas and Snell 1997) and now continued by Patricia Parker’s team (Gottdenker et al. 2005; Whiteman et al. 2005; Parker et al. 2006, 2009a, b; Duffie et al. 2008; Santiago-Alarcon et al. 2008; Levin et al. 2009).
Habitat degradation and loss is probably the second most important threat for terrestrial biodiversity in the Galapagos and the most likely direct cause of recent extinctions among some native endemic invertebrates and vertebrates (especially land birds). The humid zones of the four major islands (which also happen to be the islands inhabited by humans) are probably the most important habitats for most exclusive terrestrial Galapagos organisms, both vertebrates and invertebrates. The vegetation zone that remained largely unaltered up to the 1970s has now almost been completely replaced by introduced pastures and other invasive exotic plant species, especially on Santa Cruz and San Cristobal and to a lesser extent on Isabela and Floreana where large areas have been invaded also. The removal and alteration of natural habitats in these areas have severe implications for the conservation of a large number of native species. This alone, or in combination with other factors, may be the cause of the extinction of two species of endemic snails and the virtual disappearance of the vermilion flycatcher on San Cristobal and its apparent decline in Santa Cruz.
Marine Ecosystems
The Galapagos marine ecosystems remained largely pristine until very recently, in spite of heavy exploitation during the nineteenth century by whalers, fur sea lion hunters and, since the early 1900s, tuna fishers (Larson 2001). Besides the direct and severe effect on the exploited populations (i.e., whales and the Galapagos fur sea lion Arctocephalus galapagoensis), the ecosystem as a whole was not largely impacted, and apparently, at least for fur seals, these exploited populations recovered in full (Trillmich, personal communication). Recent research suggests that the Galapagos sea lion (Zalophus wollebaeki) population may be declining; however, preliminary results await further confirmation since methodologies between past and recent censuses differed substantially (Bustamante et al. 2002). Marine predators and grazers, both among vertebrates and invertebrates are, however, affected negatively by recurrent El Niños. The most severe effects of El Niño in the Galapagos were recorded during 1982–1983. In that year, most sea birds ceased reproduction, experienced an unprecedented high mortality, and some species including the Galapagos penguin (Spheniscus mendiculus) and the Galapagos cormorant (Phalacrocorax harrisi) suffered the most severe population declines ever recorded (Valle 1985; Valle and Coulter 1987; Valle et al. 1987). A similar pattern, although without the dramatic decline of penguins and cormorants, was also recorded for the Galapagos fur seal and the Galapagos sea lion (Limberger 1985; Trillmich and Limberger 1985; Trillmich 1985), marine iguana (Amblyrhynchus cristatus) (Laurie 1985), fishes (Grove 1985), corals that almost disappeared (Glynn 1986, 1994), and several other organisms (see Robinson and del Pino 1985). Another exceptionally strong El Niño took place in 1997–1998 that also led to interruptions and mortality and population declines of a large number of marine organisms (Vargas et al. 2006). Marine biologists suggest that the effects of the El Niño and anthropogenic factors combined to threaten several marine organisms, which in turn are severely altering the community composition of Galapagos marine ecosystems (Branch et al. 2002). Although it is still debatable, some researchers think that the frequency and intensity of El Niños may be strengthening due to anthropogenic factors at the global scale.
The main current threat to marine ecosystems is overfishing. The targets of traditional small-scale fisheries—the grouper bacalao (Mycteroperca olfax) and three species of lobster (Panulirus penicillatus, P. gracilis, Scyllarides astori)—are already overexploited. Sea cucumber (Stichopus fuscus) fishing began as a high income opportunity in the early 1990s, but became overexploited in less than a decade, as some predicted (Valle 1994). By now it has become economically unprofitable. Although baseline data are still scanty, circumstantial evidence strongly suggests that several marine invertebrates of the intertidal and subtidal zones (e.g., snails, crabs, chitons, octopi) around local communities are heavily impacted by on-foot fishing. A preliminary assessment failed to detect any impact from tourism on the marine visitor sites (Bustamante et al. 2002). However, there is growing concern that marine subtidal zones at visitor sites may become affected especially by the dramatic increase in tourism over the last 10 years. All ecosystems, but particularly the marine ones, face the potential impact from accidental fuel spills from the nearly 100 tourist boats, half-dozen cargo boats, and tankers that provide fuel to the islands for a remarkably fast-growing number of automobiles. The most striking fuel spill accident was that of the Jessica tanker in January 2001, when the tanker runs aground on Wreck Bay on San Cristobal and more than 240,000 gallons of fuel spilled into the ocean.
Conservation Achievements
Much effort has been devoted to the conservation of the Galapagos by both Ecuador and the world. Ecuador pioneered conservation efforts in South America when it declared a number of island protected areas in the 1930s and, subsequently, when it declared all terrestrial land that was not yet colonized in the Galapagos by 1959 (97.3% of the land area) as a national park. Another conservation landmark was an agreement signed by Ecuador and the Charles Darwin Foundation that allowed the creation of the Charles Darwin Research Station, which began its operation in 1964. Since 1964, both the Galapagos National Park and the Charles Darwin Foundation (through its operative arm, the Charles Darwin Research Station) have exerted tremendous effort to fulfill their mission of conserving the Galapagos Islands. Their main conservation efforts focused on (1) the protection and restoration of native endangered species and habitats; (2) the control and eradication of exotic species, focusing particularly on the most invasive flora and fauna; and (3) the environmental education of a rapidly growing resident population.
Achievements in protecting and restoring native endangered species has mainly taken place via the captive breeding program for land iguanas and giant tortoises. Their subsequent repatriation into their natural habitat qualifies as an unprecedented conservation success. The most celebrated case is that of the Espanola giant tortoise (Milinkovitch et al. 2004) that was at the brink of extinction in 1964 with only 14 individuals in the wild (12 females and two males) and a third male at the San Diego Zoo, all of whom were brought to the Darwin Station for breeding in captivity; by 2010, nearly 2,000 young tortoises had been repatriated and a few of the first repatriated young already had started breeding in the wild (Milinkovitch et al. 2004). There is, however, still room for concern about the long-term viability of this population, due both to the small number of parents, the consequent low levels of heterozygosis, and a small effective population size.
Eradication of feral introduced vertebrates, including goats (Capra hircus), pigs (Sus scrofa), dogs (Canis familiaris), cats (Felis catus), donkeys (Equus asinus), and pigeons (Columbia livia) has been successful on a number of small islands and at particular locales on the larger islands (Tapia et al. 2000; Jiménez-Uzcátegui et al. 2008a, b). Another notable conservation achievement was the Isabela Project, a well-planned intensive eradication project that led to the eradication of goats on northern Isabela Island, a vast area of difficult terrain, and the virtual eradication of goats on Pinta Island and goats and pigs on Santiago Island (Campbell et al. 2004). The local control of introduced species, particularly mammals, has proved effective in particular locations at decreasing predation and increasing in situ reproductive success and population numbers for a large number of native species including the Galapagos Petrel (Pterodroma phaeopygia) on Floreana and Santa Cruz Islands, land iguanas on Santa Cruz and Isabela Islands, and giant tortoises on several islands. Along the same lines, the first time biological control systems was applied in the Galapagos to control the introduced cottony cushion scale (Icerya purchasi) that started devastating native vegetation with the introduction and controlled release of the ladybug beetle (Rodolia cardinalis). The Galapagos National Park and the Charles Darwin Research Station have several eradication projects underway aiming to eradicate or control at least the most invasive introduced plants, vertebrates, and invertebrates. Enormous further investment is needed to succeed, especially with the need to fight not only invasive species already on the islands but also new ones that are being introduced every year, due to an ineffective and poorly implemented quarantine control.
On the marine side, the creation in 1998 of the Galapagos Marine Reserve (GMR), one of the world’s largest marine reserves backed by an organic special law, was, without a doubt, the greatest marine conservation achievement. The GMR which embraces 140,000 km2 of marine waters, both within and around the islands, is a multiple-use reserve (protection, small-scale extractive and non-extractive activities are allowed) that excludes industrial fishing and confers a high level of protection to marine ecosystems and species within 40 miles surrounding the Galapagos. The GMR is under the administration of the Galapagos National Park directorate but has several bodies that allow a process of participatory management where local stakeholders are part of the decision-making process. To implement the management of the reserve, a management plan was developed and an on-the-ground zoning system of the coastal waters was put in place and is being implemented.
Besides a good level of scientific knowledge relevant to conservation, the Galapagos has the legal framework and elements to achieve its conservation goals in the long run. In Ecuador, Galapagos is the only province with an organic special law that declares and promotes conservation and sustainable development as its fundamental principles. However, due to a general lack of law enforcement due to limited resources and limited willingness, the long-term conservation of the Galapagos cannot yet be guaranteed. The islands were included in the 2007 list of World Heritage sites in peril. There is an urgent need for an Ecuadorean state policy for the conservation of the islands. Although there is some disagreement about how much of the Galapagos native biota is still in place, nobody doubts that the Galapagos remains one of the most pristine places on Earth, something that humanity cannot afford to lose.
References
Abbott I, Abbott LK (1978) Multivariate study of morphological variation in Galapagos and Ecuadorian mockingbirds. Condor 80:302–308
Arbogast BS, Drovetski SV, Curry LR, Boag PT, Seutin G, Grant PR, Grant BR, Anderson DJ (2006) The origin and diversification of Galapagos mockingbirds. Evolution 60:370–382
Baert L (2000) Invertebrate research overview: 1. Terrestrial arthropods. In: Sitwell N, Baert L, Coppois G (eds) Science for conservation in Galapagos. Bulletin van het Koninklijk Belgisch Instituut voor Natuurwetenschappen, Suplement 70:23–25
Bailey K (1976) Potassium–argon ages from the Galapagos Islands. Science 192:465–466
Bollmer JL, Whiteman NK, Cannon MD, Bednarz JC, de Vries T, Parker PG (2005) Population genetics of the Galapagos Hawk (Buteo galapagoensis): genetic monomorphism within isolated populations. Auk 122:1210–1224
Bollmer JL, Kimball RT, Whiteman NK, Sarasola JH, Parker PG (2006) Phylogeography of the Galapagos hawk (Buteo galapagoensis): a recent arrival to the Galapagos Islands. Mol Phylogenet Evol 39:237–247
Bollmer JL, Vargas FH, Parker PG (2007) Low MHC variation in the endangered Galapagos penguin (Spheniscus mendiculus). Immunogenetics 59:593–602
Bowman RI (1963) Evolutionary patterns in Darwin’s finches. Occas Pap Calif Acad Sci 44:107–140
Bowman RI (1979) Adaptive morphology of song dialects in Darwin’s finches. J Ornithol 120:353–389
Bowman RI (1984) Contributions to science from the Galapagos. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 277–311
Branch GM, Whitman JD, Bensted-Smith R, Bustamante RH, Wellington GM, Smith F, Edgar GJ (2002) Conservation criteria for the marine biome. In: Benson-Smith R (ed) A biodiversity vision for the Galapagos Islands. Charles Darwin Foundation and World Wildlife Fund, Puerto Ayora, Galapagos, pp 80–95
Bustamante RH, Branch GM, Bensted-Smith R, Edgar GJ (2002) In: Benson-Smith R (ed) A biodiversity vision for the Galapagos Islands. Charles Darwin Foundation and World Wildlife Fund, Puerto Ayora, Galapagos, pp 80–95
Caccone A, Gibbs JP, Ketmaier V, Suatoni E, Powell JR (1999) Origin and evolutionary relationships of giant Galapagos tortoises. Proc Natl Acad Sci USA 96:13223–13228
Caccone A, Gentile G, Gibbs JP, Fritts TH, Snell HL, Betts J, Powell JR (2002) Phylogeography and history of giant Galapagos tortoises. Evolution 56:2052–2066
Campbell K, Donlan CJ, Cruz F, Carrion V (2004) Eradication of feral goats Capra hircus from Pinta Island, Galapagos, Ecuador. Oryx 38:1–6
Cane MA (1983) Oceanographic events during El Niño. Science 222:1189–1194
Carlquist S (1965) Island life. Natural History Press, Garden City, New York
Causton C, Sevilla C (2008) Latest records of introduced invertebrates in Galapagos and measures to control them. Galapagos report 2006–2007. CDF, GNP and INGALA, Puerto Ayora, Galapagos, Ecuador, pp 142–145
Christie DM, Duncan RA, McBirney AR, Richards MA, White WM, Harpp KS, Fox CG (1992) Drowned islands downstream from the Galapagos hotspot imply extended speciation times Nature 355:246–248
Clark D (1984) Native land mammals. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 225–231
Colinvaux PA (1968) Reconnaissance and chemistry of the lakes and bogs of the Galapagos Islands. Nature 219:590–594
Colinvaux PA (1972) Climate and the Galapagos Islands. Nature 240:17–20
Colinvaux PA (1984) The Galapagos climate: present and past. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 55–69
Colinvaux PA, Schofield EK (1976a) Historical ecology in the Galapagos Islands. I. A Holocene pollen record from Isla San Cristobal. J Ecol 64:989–1012
Colinvaux PA, Schofield EK (1976b) Historical ecology in the Galapagos Islands. II. A Holocene spore record from Isla San Cristobal. J Ecol 64:1013–1026
Connell JH, Sousa WP (1983) On the evidence needed to judge ecological stability or persistence. Am Nat 121:789–824
Conroy JL, Overpeck JT, Cole JE, Shanahan TM, Steinitz-Kannan M (2008) Holocene changes in eastern tropical Pacific climate inferred from a Galapagos lake sediment record. Quaternary Sci Rev 27:1166–1180
Cox A (1971) Paleomagnetism of San Cristobal Island, Galapagos. Earth Planet Sci Lett 11:152–160
Cox A (1983) Ages of the Galapagos Islands. In: Bowman RI, Berson M, Leviton AE (eds) Patterns of evolution in Galapagos organisms. American Association for the Advancement of Science, Pacific Division, San Francisco, CA, pp 123–155
Crawley MJ (1987) What makes a community invasible? In: Gray AJ, Crawley MJ, Edwards PJ (eds) Colonization, succession and stability. Blackwell Scientific, Oxford, pp 429–453
Cronin TM, Dowsett HJ (1996) Biotic and oceanographic response to the Pliocene closing of the Central American Isthmus. In: Jackson JBC, Budd AF, Coates AG (eds) Evolution and environment in tropical America. University of Chicago Press, Chicago, pp 76–104
De Roy T (ed) (2009) Galapagos: preserving Darwin’s legacy. David Bateman Ltd., New Zealand
De Vries T (1984) The giant tortoises: a natural history disturbed by man. In: Perry R (ed) Key environments. Oxford Pergamon Press, Galapagos, pp 145–156
Duffie CV, Glenn TC, Hagen C, Parker PG (2008) Microsatellite markers isolated from the flightless cormorant (Phalacrocorax harrisi). Mol Ecol Resour 8:625–627
Duffie CV, Glenn TC, Vargas FH, Parker PG (2009) Genetic structure within and between island populations of the flightless cormorant (Phalacrocorax harrisi). Mol Ecol 18:2103–2111
Edgar GJ, Banks S, Fariña M, Calvopiña M, Martínez C (2004) Regional biogeography of shallow reef fish and macro-invertebrate communities in the Galapagos archipelago. J Biogeogr 31:1107–1124
Eibl-Eibesfeldt I (1958) The Galapagos Islands: a laboratory of evolution. New Sci 4:250–253
Eibl-Eibesfeldt I (1984a) The large iguanas of the Galapagos Islands. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 157–173
Eibl-Eibesfeldt I (1984b) The Galapagos seals. Part 1. Natural history of the Galapagos sea lion (Zalophus californianus wollebaeki, Sivertsen). In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 207–214
Eliasson U (1984) Native climax forest. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 101–114
Ferodov AV, Dekens PS, McCarthy M, Revelo AC, de Menocal PB, Barreiro M, Pacanowski RC, Philander SG (2006) The Pliocene paradox (mechanisms for a permanent El Niño). Science 312:1485–1489
Fessl B, Kleindorfer S, Tebbich S (2006) An experimental study on the effects of an introduced parasite in Darwin’s finches. Biol Conserv 127:55–61
Geist D (1996) On the emergence and submergence of the Galapagos Islands. Not Galapagos 56:5–9
Geist D (2009) Islands on the move: significance of hotspot volcanoes. In: de Roy T (ed) Galapagos: preserving Darwin’s legacy. David Bateman Ltd, New Zealand, pp 28–35
Gentile G, Fabiani A, Marquez C, Snell HL, Snell HM, Tapia W, Sbordoni V (2009) An overlooked pink species of land iguana in the Galapagos. Proc Natl Acad Sci 106:507–511
Glynn PW (1986) Ecological effects of the 1982/83 El Niño Associate Disturbance to Eastern Pacific coral reefs. Progress Report 1986. CDRS Library, pp 1–13
Glynn PW (1994) State of coral reefs in the Galapagos Islands: natural vs. anthropogenic impacts. Mar Pollut Bull 29:131–140
Gottdenker N, Walsh T, Vargas H, Duncan M, Merkel J, Jimenez G, Miller RE, Dailey M, Parker PG (2005) Assessing the risks of introduced chickens and their pathogens to native birds in the Galapagos Archipelago. Biol Conserv 126:429–439
Grant PR (1984) The endemic land birds. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 175–189
Grant PR (1999) Ecology and evolution of Darwin’s finches, 2nd edn. Princeton University Press, Princeton, NJ
Grant BR, Grant PR (1989) Evolutionary dynamics of a natural population: the large cactus finch of the Galapagos. University of Chicago Press, Chicago
Grant PR, Grant BR (2006) Evolution of character displacement in Darwin’s finches. Science 313:224–226
Grant PR, Grant BR (2008) How and why species multiply: the radiation of Darwin’s finches. Princeton University Press, Princeton, NJ
Grove J (1985) Influence of the 1982–1983 El Niño event upon the icthyofauna of the Galapagos archipelago. In: Robinson G, Del Pino E (eds) El Niño en las Islas Galapagos: Evento 1982–1993. Charles Darwin Foundation, Galapagos, Ecuador, pp 245–258
Harris MP (1984) Seabirds. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 191–206
Herbold R, Moyle PB (1986) Introduced species and vacant niches. Am Nat 128:751–760
Hoeck PEA, Bollmer JL, Parker PG, Keller LF (2010) Differentiation with drift: a spatio-temporal genetic analysis of Galapagos mockingbird populations (Mimus spp.). Phil Trans R Soc B 365:1127–1138
Houvenaghel GT (1984) Oceanographic setting of the Galapagos Islands. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 225–231
Hull JM, Savage WK, Bollmer JL, Kimball RT, Parker PG, Whiteman NK, Ernest HB (2008) On the origin of the Galapagos hawk: an examination of phenotypic differentiation and mitochondrial paraphyly. Biol J Linn Soc 95:779–789
Huxley JS (1938) Species formation and geographic isolation. Proc Linn Soc Lond 150:253–264
Jiménez-Uzcátegui G, Carrión V, Zabala J, Buitrón P, Milstead B (2008) Status of introduced vertebrates in Galapagos. Galapagos report 2006–2007. Charles Darwin Foundation, Puerto Ayora, pp 136–141
Jiménez-Uzcátegui G, Carrión V, Zabala J, Buitrón P, Milstead B (2008) Status of introduced vertebrates in Galapagos. Galapagos report 2007–2008. Charles Darwin Foundation, Puerto Ayora, pp 97–102
Jordan MA, Snell HL (2008) Historical fragmentation of islands and genetic drift in populations of Galapagos lava lizards (Microlophus albemarlensis complex). Mol Ecol 17:1224–1237
Jordan MA, Hammond RL, Snell HL, Jordan WC (2002) Isolation and characterization of microsatellite loci from Galapagos lava lizards (Microlophus spp.). Mol Ecol Notes 2:349–351
Kennedy M, Valle CA, Spencer HG (2009) The phylogenetic position of the Galápagos Cormorant. Mol Phylogenet Evol 53:94–98
Kizirian D, Trager A, Donnelly MA, Wright JW (2004) Evolution of Galapagos Island lava lizards (Iguania: Tropiduridae: Microlophus). Mol Phylogenet Evol 32:761–769
Lack D (1947) Darwin’s finches. Cambridge University Press, Cambridge
Lambeck K, Chappell J (2001) Sea level change through the last glacial cycle. Science 292:679–686
Lande R (1980) Genetic variation and phenotypic evolution during allopatric speciation. Am Nat 116:463–479
Lande R, Barrowclough GF (1987) Effective population size, genetic variation, and their use in population management. In: Soule ME (ed) Viable populations for conservation. Cambridge University Press, New York, pp 87–123
Larson EJ (2001) Evolution’s workshop: God and science on the Galapagos Islands. Basic Books, New York
Laurie A (1985) Santa Fe news letter. Noticias de Galapagos 41:20–21
Lea DW, Pak DK, Belanger CL, Spero HJ, Hall MA, Shackleton NJ (2006) Paleoclimate history of Galapagos surface waters over the last 135,000 yr. Quaternary Sci Rev 25:1152–1167
Levin I, Outlaw DC, Vargas FH, Parker PG (2009) Plasmodium blood parasite found in endangered Galapagos penguins (Spheniscus mendiculus). Biol Conserv 142:3191–3195
Limberger D (1985) El Niño on Fernandina. In: Robinson G, Del Pino E (eds) El Niño en las Islas Galapagos: Evento 1982–1993. Charles Darwin Foundation, Galapagos, Ecuador, pp 245–258
Lopez TJ, Hauselman ED, Maxson LR, Wright JW (1992) Preliminary analysis of phylogenetic relationships among Galapagos Island lizards of the genus Tropidurus. Amphibia-Reptilia 13:327–339
Mace GM, Lande R (1991) Assessing extinction threats: toward a reevaluation of IUCN threatened species categories. Conserv Biol 5:148–157
Mayr E (1954) Change in genetic environment and evolution. In: Huxley J, Hardy AC, Ford EB (eds) Evolution as a process. Allen & Unwin, London, pp 157–180
Mayr E (1982) The growth of biological thought: diversity, evolution, and inheritance. Harvard University Press, Cambridge
McArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton, NJ
McBirney AR, Williams H (1969) Geology and petrology of the Galapagos Islands. In: Bowman RI (ed) The Galapagos, Proceedings of the Symposia G.I.S.P. University of California Press, Berkeley, pp 65–70
Milinkovitch MC, Monteyne D, Gibbs JP, Fritts TH, Tapia W, Snell HL, Tiedemann R, Caccone A, Powell JR (2004) Genetic analysis of a successful repatriation program: giant Galapagos tortoises. Proc R Soc Lond B 271:341–345
Miller GD, Hofkin BV, Snell H, Hahn A, Miller RD (2001) Avian malaria and Marek’s disease: potential threats to Galapagos penguin Spheniscus mendiculus. Mar Ornithol 29:43–46
Palmer CE, Pyle RL (1966) The climatological setting of the Galapagos. In: Bowman RI (ed) The Galapagos, Proceedings of the Symposia G.I.S.P. University of California Press, Berkeley, pp 65–70
Parent CE, Crespi BJ (2006) Sequential colonization and diversification of Galapagos endemic land snail genus Bulimulus (Gastropoda, Stylommatophora). Evolution 60:2311–2328
Parent CE, Caccone A, Petren K (2008) Colonization and diversification of Galapagos terrestrial fauna: a phylogenetic and biogeographical synthesis. Phil Trans R Soc B 363:3347–3361
Parker P (2009a) A most unusual hawk: one mother and several fathers. In: de Roy T (ed) Galapagos: preserving Darwin’s legacy. David Bateman Ltd., New Zealand, pp 130–137
Parker PG (2009b) Parasites and pathogens: threats to native birds. In: de Roy T (ed) Galapagos: preserving Darwin’s legacy. New Zealand, David Bateman Ltd, pp 177–183
Parker PG, Whiteman NK, Miller RE (2006) Perspectives in ornithology: conservation medicine in the Galapagos Islands: partnerships among behavioral, population and veterinary scientists. Auk 123:625–638
Peck SB (1996) Origin and development of an insect fauna on a remote archipelago: the Galapagos Islands, Ecuador. In: Keast A, Miller SE (eds) The origin and evolution of Pacific Island biotas, New Guinea to Eastern Polynesia: patterns and processes. Academic Publishing, Amsterdam, pp 91–122
Petren K, Grant BR, Grant PR (1999) A phylogeny of Darwin’s finches based on microsatellite DNA length variation. Phil Trans R Soc B 266:321–329
Petren K, Grant PR, Grant BR, Keller LF (2005) Comparative landscape genetics and the adaptive radiation of Darwin’s finches: the role of peripheral isolation. Mol Ecol 14:2943–2957
Porter DM (1976) Geography and dispersal of Galapagos Islands vascular plants. Nature 264:745–746
Porter D (1984) Endemism and evolution in terrestrial plants. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 85–99
Quiroga D (2009) Galapagos, laboratorio natural de la evolucion: una aproximacion historica. In: Tapia W, Ospina P, Quiroga D, Gonzales JA, Montes C (eds) Ciencia para la sostenibilidad en Galapagos. Parque Nacional Galapagos, Ecuador
Rassmann K (1997) Evolutionary age of the Galapagos iguanas predates the age of the present Galapagos Islands. Mol Phylogenet Evol 7:158–172
Riedinger M, Steinitz Kannan M, Last W, Brenner M (2002) A 6100 14-C record of El Niño activity from the Galapagos Islands. J Paleolimnol 27:1–7
Robinson G, del Pino EM (1985) El Niño in the Galapagos Islands. Charles Darwin Foundation, Quito
Roque-Albelo L (2008) Evaluating land invertebrate species: prioritizing endangered species. Galapagos report 2006–2007. Charles Darwin Foundation, Puerto Ayora, 111–117
Santiago-Alarcon D, Whiteman NK, Ricklefs RE, Parker PG, Valkiunas G (2008) Patterns of parasite abundance and distribution in island populations of Galapagos endemic birds. J Parasitol 94:584–590
Sato A, Tichy H, O’Huigin C, Grant PR, Grant BR, Klein J (2001a) On the origin of Darwin’s finches. Mol Biol Evol 18:299–311
Sato A, Mayer WE, Tichy H, Grant PR, Grant BR, Klein J (2001b) Evolution of Mhc class II B genes in Darwin’s finches and their closest relatives: birth of a new gene. Immunogenetics 53:792–801
Schluter D (1986) Character displacement between distantly related taxa? Finches and the bees in the Galapagos. Am Nat 127:95–102
Schluter D, Grant PR (1984) Determinants of morphological patterns in communities of Darwin’s finches. Am Nat 123:175–196
Schluter D, Price TD, Grant PR (1985) Ecological character displacement in Darwin’s finches. Science 227:1056–1059
Schmitz P, Cibois A, Landry B (2007) Molecular phylogeny and dating of an insular endemic moth radiation inferred from mitochondrial and nuclear genes: the genus Galagete (Lepidoptera: Autostichidae) of the Galapagos Islands. Mol Phylogenet Evol 45:180–192
Sequeira AS, Lanteri AA, Scataglini MA, Confalonieri VA, Farrell BD (2000) Are flightless Galapaganus weevils older than the Galapagos Islands they inhabit? Heredity 85:20–29
Shen GT, Cole JE, Lea DW, Linn LJ, McConnaughey TA, Fairbanks RG (1992) Surface ocean variability at Galapagos from 1936–1982: calibration of geochemical tracers in corals. Paleoceanography 7:563–588
Simkin T (1984) Geology of Galapagos Islands. In: Perry R (ed) Key environments: Galapagos. Pergamon Press, New York, pp 15–41
Simkin T, Howard KA (1970) Caldera collapse in the Galapagos Islands, 1968. Science 169:429–437
Snell HL, Tye A, Causton CE, Bensted-Smith R (2002) Current status and threats to the terrestrial biodiversity of Galapagos. A biodiversity vision for the Galapagos Islands. Charles Darwin Foundation and World Wildlife Fund, Puerto Ayora, Galapagos, pp 30–47
Sulloway FJ (1982) Darwin and his finches: the evolution of a legend. J Hist Biol 15:1–53
Swanson FJ, Baitis HW, Lexa J, Dymond J (1974) Geology of Santiago, Rabida and Pinzon Islands, Galapagos. Geol Soc Am Bull 85:1803–1810
Tapia W, Patry M, Snell H, Carrión V (2000) Estado actual de los vertebrados introducidos a las islas Galapagos. Fundación Natura: Informe Galapagos 1999–2000, Quito, Ecuador
Trillmich F (1984) Part 2. Natural history of the Galapagos fur seal (Arctocephalus galapagoensis, Heller). In: Perry R (ed) Key environments: Galapagos. Pergamon Press, Oxford, pp 85–99
Trillmich F (1985) Effects of the 1982/83 El Niño on Galapagos fur seals and sea lions. Not Galapagos 42:22–23
Trillmich F, Limberger D (1985) Drastic effect of El Niño on Galapagos, Ecuador pinnipeds. Oecologia 67:19–22
Tye A (2007) The status of the endemic flora of Galapagos: the number of threatened species is increasing. Galapagos report 2006–2007. Charles Darwin Foundation, Puerto Ayora, pp 97–103
Tye A (2008) The status of the endemic flora of Galapagos: the number of threatened species is increasing. Galapagos report 2007–2008. Charles Darwin Foundation, Puerto Ayora, pp 97–102
Tye A, Snell HL, Peck SB, Andersen H (2002) Outstanding terrestrial features of the Galapagos Archipelago. A biodiversity vision for the Galapagos Islands. Charles Darwin Foundation and World Wildlife Fund, Puerto Ayora, Galapagos, pp 12–23
Tye A, Atkinson R, Carrión V (2008) Increase in the number of introduced plant species in Galapagos. Galapagos report 2006–2007. Charles Darwin Foundation, Puerto Ayora, pp 133–135
Urban FE, Cole JE, Overpeck JT (2000) Influence of mean climate change on climate variability from a 155-year tropical Pacific coral record. Nature 407(6807):989–993
Valle CA (1985) Alteración de las poblaciones del cormorán no volador, el pingüino y otras aves marinas en Galapagos por efecto de El Niño 1982–83 y su subsecuente recuperación. In: Robinson G, Del Pino E (eds) El Niño en las Islas Galapagos: Evento 1982–1993. Charles Darwin Foundation, Galapagos, Ecuador, pp 245–258
Valle CA (1994) Pepino war, 1992—is conservation just a matter for the elite? Not Galapagos 53:2
Valle CA, Coulter MC (1987) Present status of the flightless cormorant, Galapagos penguin, and greater flamingo populations in the Galapagos Islands, Ecuador after the 1982–83 el Niño. Condor 89:276–281
Valle CA, Cruz F, Cruz JB, Merlen G, Coulter MC (1987) The impact of the 1982–1983 El Niño Southern oscillation on seabirds in the Galapagos Islands, Ecuador. J Geophys Res 92:14437–14443
Van Riper C, Van Riper SG, Goff ML, Laird M (1988) The epizootiology and ecological significance of malaria in Hawaiian land birds. Ecol Monogr 58:111–127
Vargas H, Snell HM (1997) The arrival of Marek’s disease to Galapagos. Not Galapagos 58:4–5
Vargas FH, Harrison S, Rea S, Macdonald DW (2006) Biological effects of El Niño on the Galapagos penguin. Biol Conserv 127:107–114
Whiteman NK, Goodman SJ, Sinclair BJ, Walsh T, Cunningham AA, Kramer LD, Parker PG (2005) Establishment of the avian disease vector Culex quinquefasciatus Say, 1823 (Diptera: Culicidae) on the Galapagos Islands, Ecuador. Ibis 147:844–847
Whiteman NK, Kimball RT, Parker PG (2007) Co-phylogeography and comparative population genetics of the Galapagos Hawk and three co-occurring ectoparasite species: natural history shapes population histories within a parasite community. Mol Ecol 16:4759–4773
Wikelski M, Foufopoulos J, Vargas H, Snell H (2004) Galapagos birds and diseases: invasive pathogens as threats for island species. Ecol Soc 9:5
Williams H (1966) Geology of the Galapagos Islands. In: Bowman RI (ed) The Galapagos, Proceedings of the Symposia G.I.S.P. University of California Press, Berkeley, pp 65–70
Williams H, McBirney AR (1979) Volcanology. Freeman, Cooper, San Francisco
Withman JD, Smith F (2003) Rapid community change at a site in the Galapagos marine reserve. Biodivers Conserv 12:25–45
Wright S (1931) Evolution in Mendelian populations. Genetics 16:97–159
Wright S (1932) The roles of mutation, inbreeding, crossbreeding and selection in evolution. Proc 6th Int Cong Genet 1:356–366
Wyles JS, Sarich VM (1983) Are the Galapagos iguanas older than the Galapagos? Molecular evolution and colonization models for the archipelago. In: Bowman RI, Berson M, Levinton AE (eds) Patterns of evolution in Galapagos organisms. Amer. Assoc. Advanc. Sc., Pacific Div, San Francisco, pp 177–185
Wyrtki K (1975) ‘El Niño’: the dynamic response of the Equatorial Pacific Ocean to atmospheric forcing. J Phys Oceanogr 5:572–584
Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292:686–693
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Valle, C.A. (2013). Science and Conservation in the Galapagos Islands. In: Walsh, S., Mena, C. (eds) Science and Conservation in the Galapagos Islands. Social and Ecological Interactions in the Galapagos Islands, vol 1. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-5794-7_1
Download citation
DOI: https://doi.org/10.1007/978-1-4614-5794-7_1
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-5793-0
Online ISBN: 978-1-4614-5794-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)