
Abstract
The ability of cells to generate, maintain, and repair tissues with complex architecture, in which distinct cells function as coherent units, relies on polarity cues. Polarity can be described as an asymmetry along a defined axis, manifested at the molecular, structural, and functional levels. Several types of cell and tissue polarities were described in the literature, including front-back, apical-basal, anterior–posterior, and left-right polarity. Extensive research provided insights into the specific regulators of each polarization process, as well as into generic elements that affect all types of polarities. The actin cytoskeleton and the associated adhesion structures are major regulators of most, if not all, known forms of polarity. Actin filaments exhibit intrinsic polarity and their ability to bind many proteins including the mechanosensitive adhesion and motor proteins, such as myosins, play key roles in cell polarization. The actin cytoskeleton can generate mechanical forces and together with the associated adhesions, probe the mechanical, structural, and chemical properties of the environment, and transmit signals that impact numerous biological processes, including cell polarity. In this article we highlight novel mechanisms whereby the mechanical forces and actin-adhesion complexes regulate cell and tissue polarity in a variety of natural and experimental systems.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
1.1 The Metazoan Revolution and the Emergence of Multicellular Morphogenesis
The evolution of multicellular forms of life (“metazoa”) was a long and gradual process, which is commonly believed to have taken place between 1,300 and 600 million years ago (Conway Morris 1998). During that period, unicellular organisms started to develop colonial forms of life, which led, over time, to the development of independent, “individualized” metazoan organisms, most likely resembling today’s sponges (Muller and Muller 2003). This transition from relatively simple unicellular organisms (protozoa) to the considerably more complex multicellular life was apparently associated with major changes in the cells’ survival strategy; while in protozoan organisms, all life processes (e.g., reproduction, motility, feeding, and protection) are independently executed by all individual cells. In metazoans these missions are performed by specialized sub-populations of cells. With the advent of metazoan evolution, mainly during the “Cambrian explosion,” an increasing order in body plan was observed (mainly, through detailed fossil records), with a common form, characterized by tissue polarization along a single longitudinal axis [anterior (front)–posterior (back)], which is often associated with the direction of locomotion (Martindale 2005) (Fig. 1). The cellular and molecular processes underlying this tissue polarization will be discussed below.

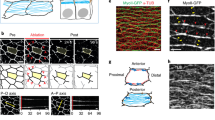
Schematic overview of polarity from the cell and tissue level to the whole organism level. The example shown here is of simple epithelia that exhibit front-back polarity, apico-basal polarity, and anterior–posterior polarity (a). At the whole organism level (bilaterians) three distinct axes can be evident: anterior–posterior, dorsal-ventral, and left-right (b)
Interestingly, many studies, initially based on biochemical information (e.g., Burger and Jumblatt 1977; Muller et al. 1982; Wiens et al. 2003) and later on molecular genetic data (Wiens et al. 2003), revealed diverse adhesion molecules in sponges (“aggregation factors,” Burger and Jumblatt 1977), which promote species-specific clustering of sponge cells. Moreover, the inter-cellular space in sponges contains galectin (Wagner-Hulsmann et al. 1996), as well as additional components related to the extracellular matrix (ECM) of higher metazoans, including collagen, and fibronectin-like protein (Labat-Robert et al. 1981). Naturally, in higher metazoa the repertoire of ECM proteins is considerably larger than that of sponges, reflecting the increasing complexity of tissue and organ architecture.
1.2 Cell-to-Tissue Polarity
Asymmetric distribution of molecules, sub-cellular structures, and organelles is a hallmark of many cell types. These basic asymmetries can lead to local changes in cell shape and/or function, transiently or constitutively, and are largely known as cell polarity. In fact, polarity is a fundamental property of nearly all living cells; it can be detected in vitro and in vivo, in a wide variety of cell types from those forming complex metazoans to simpler unicellular eukaryotes such as amoeba and yeast and also in some bacteria.
Polarity plays a critical role in cells’ life. The fate of a new daughter cell may be determined by polarity cues that orient the mitotic spindle and assign the daughter cells into a specific niche (Williams and Fuchs 2013; Vorhagen and Niessen 2014). Polarity also plays a key role in cells’ ability to execute many biological functions, such as migrating directionally as individuals or collectively (Vicente-Manzanares et al. 2009; Devreotes and Horwitz 2015). It is also essential for cell ability to maintain properly its architecture and function by regulating cytoskeletal organization, trafficking, and signaling events (Guyer and Macara 2015; Narimatsu et al. 2015; Sasaki 2015). Morphogenesis, a process that shapes tissues and organisms, rely on complex polarity cues, and once a tissue takes shape, polarity allows it to maintain homeostasis and function as a coherent and, largely, independent unit (Roignot et al. 2013; Tellkamp et al. 2014). Moreover, upon injury, polarity cues allow cells to repair the tissue effectively and resume normal function.
With so many vital processes that take place throughout the life of an organism, it is not surprising that defects in cell polarity may have dire consequences, such as severe neural tube closure defects (Juriloff and Harris 2012), deafness and kidney diseases, and severe homeostasis defects associated with the development of cancer (Fischer et al. 2006; Martin-Belmonte and Perez-Moreno 2012).
While most cells exhibit some sort of asymmetry, polarity can be quite diverse in space and time (Fig. 1). Some cell types maintain a highly polarized architecture throughout life, while other cells transiently polarize to execute a specific biological function (e.g., immune cells during inflammation).
As briefly mentioned above, polarity can be classified, according to the axis that defines the asymmetry. The most common types of polarity are
-
Front-back polarity, found in many types of migratory cells; it can be recognized by the assembly of motility related actin-rich protrusions at the front of the cell (the leading edge), while at the back of the cell is enriched with contractile structures that facilitate the retraction of the trailing edge of the cell (Devreotes and Horwitz 2015).
-
Apical-basal polarity is commonly found in simple, single-layered epithelia (Figs. 1, 2, and 3) where it relates to the following sub-cellular domains: (a) The apical domain – the cell’s part that faces the lumen of an organ or the environment; (b) The lateral domain – the cell’s part that faces the neighboring cells in the monolayer; and (c) The basal domain – the part of the cell that faces and interacts with the ECM. In some simple epithelia apico-basal polarity can be recognized by the appearance of actin-rich microvilli at the apical region, cell–cell adhesion structures at the lateral domain, and cell–ECM adhesions at the basal aspect (Roignot et al. 2013; Rodriguez-Boulan and Macara 2014).
Fig. 2 
Apico-basal and planar cell polarity in the skin epidermis. The apico-basal polarity protein PAR3 (green) labels the apical domain of basal layer cells; the basement membrane protein Nidogen (red) labels the dermal-epidermis boundary (a). The core PCP protein Celsr1 (green) labels the anterior and posterior faces of basal layer cells (b). Scale bar = 10 μm
Fig. 3 
Organization of inter-cellular tight junctions and cell–ECM adhesions formed by cultured intestinal epithelial (CaCo2) cells. (a) A panel showing immunofluorescently labeled CaCo2 cells visualizing cell adhesions formed at the apical (left) and basal (right) areas of the cells. The cells were triple-labeled for the tight junction component cingulin (top, left), actin, which is associated with both cell–cell adherens junctions and cell–ECM focal adhesions (middle) and paxillin (top, right), associated with cell–ECM FAs. Two-color (pink-green) super-positions of cingulin/actin and paxillin-actin are depicted in the images shown at the bottom (courtesy of Inna Grosheva). (b) Transmission electron microscopy, showing (in a grazing section) the apical tight junction (TJ) and the sub-apical adherens junction (AJ). (c) Transmission electron microscopy, showing (in a cross section) a basal focal adhesion (marked by the two arrows); N nucleus (electron micrographs were provided by Ilana Sabanay)
-
Planar cell polarity (PCP) refers to the collective polarization of cells along the plain of a tissue, orthogonal to apical-basal polarity (Figs. 1 and 2). This polarity allows long-range communication between cells, affecting their internal organization relative to the anterior–posterior/proximal-distal axes and supports their functional coordination as coherent units (McNeill 2010; Devenport 2014; Sebbagh and Borg 2014; Sokol 2015) (for more details, see Chapter 2).
-
Left-right polarity (also known as “chirality” or “handedness”) refers to bilateral asymmetry that can be detected in many organisms. It can be evident at the cell level as many cultured cell types establish asymmetry with consistent handedness. At the tissue/organ level left-right polarity cues shape our organs, such as heart and viscera that exhibit clear asymmetry (Coutelis et al. 2014; Hamada and Tam 2014; Yoshiba and Hamada 2014) (for more details, see Chapter 3).


While every type of polarity has unique regulators, some proteins and cellular structures are essential for most, if not all, types of polarity. The actin cytoskeleton and its associated adhesion structures are the primary drivers of polarization at the cell’s level (see Figs. 3 and 4 for actin/adhesion complexes in epithelial cells and fibroblasts). Below, we discuss the roles of the actin cytoskeleton and its adhesions in cell and tissue polarity. Given that cellular and trans-cellular mechanical forces are major regulators of both the actin cytoskeleton and its associated adhesions, we will also briefly address here classical examples (Chapter 1) and novel mechanisms that play a role in the bio-mechanical regulation of planar, and left-right polarities (Chapters 2 and 3).

Effect of matrix rigidity on cell polarization, focal adhesion formation, and actin organization. Fibroblasts, stably expressing YFP-paxillin (green), were plated on elastomeric PDMS substrates with different stiffness (“rigid” = 2 MPa (A); “soft” = 5kPa (B)), incubated for 6 h, and then fixed-permeabilized and labeled with phalloidin to visualize actin (red), and DAPI (blue), to visualize the nucleus. Notice the polarization of the cell adhering to the rigid surface, and the radial spreading of the cell adhering to compliant surface (courtesy of Alexandra Lichtenstein)
1.3 The Roles of the Actin Cytoskeleton in the Acquisition and Maintenance of Cell Polarity
The cytoskeleton is made of three major types of filaments: actin filaments (F-actin), microtubule, and intermediate filaments. The first two exhibit an intrinsic asymmetry and play a major role in polarity (Li and Gundersen 2008; Mullins 2010). The actin filament is a polymer composed of globular subunits made of the monomeric G-actin, which polymerizes in a uniform orientation, giving rise to a polarized linear polymer with two distinct ends that exhibit different structural and dynamic properties. The two ends of F-actin are the “barbed” (+) and “pointed” (−) ends. At steady state, G-actin associates with the polymer primarily at the barbed end and dissociates at the pointed end, a process that leads to a “treadmilling” process that is characteristic of the internal dynamics of the actin filament in cells. These dynamic properties also involve chemical energy: new G-actin that undergoes polymerization binds ATP (G-actin-ATP) that will be hydrolyzed soon after polymerization (G-actin-ADP-Pi) followed by a slower dissociation of the phosphate (G-actin-ADP) eventually leading to the dissociation of G-actin from the pointed end of the filament. ATP hydrolysis alters the local conformation of the polymer and therefore allows actin binding proteins such as cofilin to distinguish between “new” and “old” parts of the actin filaments. Beyond cofilin, more than 100 different proteins bind actin filaments. Some of these can differentiate between the barbed and the pointed ends (e.g., capZ and tropomodulin, respectively), others can alter the stability of the filament and modify its dynamic properties (e.g., tropomyosins), create bundles with different orientation/polarity (e.g., fimbrin that forms parallel bundles, α-actinin that is associated with anti-parallel bundles, and filamins that support the formation of F-actin networks). This extraordinary variety of actin modulators allows the actin cytoskeleton to function as a highly versatile platform that regulates many processes including cell polarity (Pollard 2007; Pollard and Cooper 2009; De La Cruz and Gardel 2015).
Special players in actin-based mechanics are myosins that are actin binding molecular motors that translate chemical energy (ATP) into mechanical work (transport or contraction). Myosins’ ability to “walk” unidirectionally along actin fibers and rotate the actin filament with constant handedness (Sase et al. 1997; Beausang et al. 2008) renders them key players in the generation and maintenance of cell polarity. The myosin superfamily is large and versatile; however, it can be divided into two groups: conventional and unconventional myosins. Conventional myosin, also known as myosin II, assembles into bipolar filaments that together with actin filaments generate tension forces. Myosin II plays a key role in many processes that require the generation of tension within and between cells, such as muscle contraction, cell migration, cytokinesis, and cell shape regulation (Zaidel-Bar et al. 2015). Unconventional myosins that do not form filaments play a role in many processes including transport of cargo and functional assembly of actin-rich structures such as stereocilia in the inner ear, epithelial microvilli, and filopodia (Hartman and Spudich 2012; Vicente-Manzanares et al. 2009). Although the two groups of myosins have distinct structures, enzymatic properties, and functions, members of both groups play key roles in the establishment of polarity.
In the C. elegans zygote myosin II activity plays a key role in the polarization of PAR (“partitioning defective”) proteins and the establishment of anterior/posterior polarity (Munro et al. 2004). Sperm entry into C. elegans egg alters cortical actomyosin activity (“actomyosin” refers to a contractile complex of F-actin and myosin II). Specifically, it induces flow of actin and myosin II from the sperm entry site that will become the posterior of the embryo, to the opposite pole, that will become the embryo’s head.
A role for non-conventional myosin was demonstrated in the budding yeast S. cerevisiae. Polarization of non-polar yeast (G1 arrested) can be induced by expressing of active Cdc42, a Rho family small GTPase, and a key regulator of the actin cytoskeleton and polarity (Wedlich-Soldner et al. 2003). This process involves amplification of stochastic Cdc42 activity to cross a certain threshold and establish polarity. In this system Cdc42 mediates the polymerization of actin cables on which the unconventional myosin V transports more Cdc42 towards the membrane and amplifies random signals into a stable site of active Cdc42 that interacts with actin cables (Wedlich-Soldner et al. 2003; Li and Gundersen 2008).
1.4 Thoughts About the Roles of Actin Associated Adhesion Structures in Acquisition of Polarity
How does the assembly of the actin cytoskeleton contribute to embryonic morphogenesis and acquisition of polarity? In multicellular organisms, cytoskeleton-anchored adhesions of cells to the ECM or to their neighbors play a key morphogenetic role. Consequently, the characterization of cell adhesions, and their structural and functional diversity was extensively pursued in recent years (Wolfenson et al. 2013; Winograd-Katz et al. 2014; Lecuit and Yap 2015; Priya and Yap 2015).
Adhesive interactions, as part of the cells’ communication and interactions network, are highly complex processes that enable the sensing of the chemical and physical properties of external surfaces. These environmental cues can be further integrated by the cells, and activate signaling processes that regulate cell proliferation, survival, differentiation, and migration. Furthermore, specific classes of cell adhesions, mainly integrin mediated cell–ECM adhesions (Meldolesi 2016), and cadherin-mediated cell–cell adherens junctions (AJ) (Harris and Tepass 2010) have an inherent association with the actin cytoskeleton, and consequently, play a key role in cellular mechanics, including cell polarization processes.
It is noteworthy that in addition to integrin- and cadherin-mediated adhesions, discussed here, cells contain a wide variety of additional adhesive mechanisms, which affect cell behavior and fate. To mention just a few – cells can bind to the connective tissue and basement membrane via a variety of lectins and proteoglycan receptors, as well as intermediate filament-anchored hemidesmosomes (McDonald and Mecham 1991). Similarly, the canonical inter-cellular junctional complex of epithelial cells consists of apical tight junctions, which seal epithelial sheets, and sets the apical-basolateral polarity. Along their lateral membranes, epithelial cells communicate via gap junctions and form robust cytokeratin adhesions, namely desmosomes (Alberts 2015). In this article we will focus only on the actin-associated, integrin- and cadherin-mediated junctions.
Cellular characterization of integrin-mediated ECM adhesion was (and still is) strongly based on the investigation of focal adhesions (FA) and related structures. FAs are specialized and defined regions (commonly measuring 0.25–2 by 2–10 μm) along the ventral plasma membrane, which are directly interacting with the substrate. Moreover, FAs are associated via their cytoplasmic faces with the termini of bundles of actin filaments, known as stress fibers. The molecular composition of FAs was addressed by a wide variety of experimental approaches, ranging from immunofluorescence microscopy to biochemistry to advanced genomics and proteomics, yielding a long list of proteins that belong to the so-called integrin adhesome (Table 1; Zaidel-Bar et al. 2007; Zaidel-Bar and Geiger 2010; Horton et al. 2016). Among them are “scaffolding proteins” (e.g., talin, vinculin, paxillin, and zyxin) that physically bridge, directly or indirectly, between the cytoplasmic domains of integrins and the actin cytoskeleton, and “signaling proteins” (e.g., a variety of kinases, phosphatases, and regulators of GTPases), which can affect cell behavior, as well as the fate of the adhesion site itself (Zaidel-Bar and Geiger 2010). In recent years much information has accumulated not only on the physiological roles of the integrin adhesome, but also on its involvement in human diseases (Winograd-Katz et al. 2014).
FAs, and additional forms of integrin-mediated contacts with the ECM such as focal complexes, fibrillar adhesions, podosomes, and invadopodia (Geiger and Yamada 2011), play key roles in the induction of cell spreading in a wide variety of tissue cells (e.g., mesenchyme-derived fibroblasts, epithelial cells, and endothelial cells), followed by anterior–posterior polarization. The mechanism underlying this process involves a combination of integrin-mediated signaling and scaffolding. Interaction with the ECM was shown to activate the small GTPases Rac1 and CDC42, which trigger cell spreading by activating lamellipodial and filopodial protrusion (Price et al. 1998; Lawson and Burridge 2014). Nascent adhesions, which form under the expanding lamellae, are consequently exposed to mechanical stress, which promotes their growth and maturation. This is followed by the nucleation and assembly of stress fibers that run between FAs, and apply contractile forces to the two FAs associated with its ends. This Rho A-activated actomyosin contractility applies a stable tension, via the stress fibers to FAs (~5.5 nN/μm2; see Balaban et al. 2001), which was shown to be crucial for maintaining the integrity of the FA-stress fiber complex.
How are integrin-mediated FAs involved in regulating cell polarization? Examination of cells, plated on a rigid surface, indicated that cell spreading on the substrate is, initially, a radial process, leading to the development of circular (“fried-egg”) morphology, with peripheral, mostly radially oriented FAs. The rate and extent of spreading and FA formation vary between cells, and are usually affected by ligand density and other surface features (e.g., Reinhart-King et al. 2005). Upon longer incubation, the radial symmetry is usually broken (most prominently in fibroblasts), and the cells tend to polarize, and assume an elongated morphology. In cells with migratory properties, the two ends of the major axis of the cells are distinct; one end developing a protrusive lamellipodium (“leading edge”), while the opposite end is dominated by focal adhesions (“trailing edge”). The acquisition of such anterior–posterior polarity plays a crucial role in physiological processes such as embryonic development and wound healing, as well as pathological states like invasive and metastatic migration of cancer cells. This transition from radial to axial symmetry is regulated by both the rigidity of the underlying substrate and the cell’s adhesion-signaling system. Specifically, it was shown that cell polarization/ elongation depends on substrate rigidity, and is usually preceded by polarization of the peripheral FAs (Fig. 4). Furthermore, it was shown, by siRNA screening, that the polarization process is tightly regulated by tyrosine kinases within the adhering cells (for details, see: Prager-Khoutorsky et al. 2011).
Actin associated cell–cell AJs also play key roles in tissue coherence, primarily in epithelial, endothelial, and cardiac muscle cells (Geiger et al. 1983; Volberg et al. 1986). In simple epithelial cells (e.g., intestinal or renal epithelia) they are usually organized as sub-apical rings, located just below the apical tight junctions (Farquhar and Palade 1963), while in other cell types they form patchy adhesions of variable sizes and distributions (Fig. 3). The adhesion receptors of AJs are different cadherins, which commonly mediate calcium-dependent homotypic interactions (between similar cells) and act in a homophilic manner (interacting with the same cadherin type on the neighboring cell). Just like integrins, in ECM adhesions, cadherins interact with actin via a network of scaffolding and signaling molecules, referred to, collectively, as the “cadherin adhesome” or “cadhesome” (Table 1; Guo et al. 2014; Zaidel-Bar et al. 2015). These components, collectively, contribute to the mechanical stability of the inter-cellular junction, and to its capacity to generate and transmit adhesion signals. Moreover, some of these molecules (e.g., vinculin, α-actinin, and VASP) are present in both types of adhesions, while others are associated only with focal adhesions (e.g., paxillin, ILK) or only with AJs (e.g., α and β catenin).
The regulation of AJ formation and stability is based on the same general principles which operate in integrin adhesions, including mechanical stimulation and integration (Lecuit and Yap 2015), coordinated activation of stimulatory GEFs and inhibitory GAPs (Braga and Yap 2005), and the recruitment of different kinases and phosphatases (Bertocchi et al. 2012). While the regulatory mechanisms underlying cell–cell and cell–ECM adhesion appear to be quite similar (Table 1), the two types of adhesions are far from being up- and down-regulated simultaneously. On the contrary; commonly, the state of organization of the two types of adhesions is reciprocally related. Whether the mechanism whereby these adhesion systems are coordinated and/or differentially regulated is not clear. Different possibilities are considered, including the activation of specific signaling pathways, stimulation of a mesenchymal-epithelial transition-like process, and direct mechanical cross-talk between the two structures. Whichever the underlying mechanism is, the transition from epithelial to mesenchymal characteristics leads to the loss of apical-basal polarity and (in some cases), to acquisition of a “migratory” phenotype, based on anterior–posterior polarity.
1.5 The Roles of Mechanical Forces in Acquisition of Polarity
Mechanical forces are major regulators of biological processes. These forces can be divided into two types according to their source: intrinsic forces, developed inside the cell, and extrinsic, “environmental” forces, generated by whole body movements, muscle activity, blood flow, and the like. Mechanical forces affect multiple cellular processes, ranging from cell architecture to signal transduction to cell fate determination (Iskratsch et al. 2014; Zaidel-Bar et al. 2015).
The actin cytoskeleton is not only force generator, but is also sensitive to mechanical perturbations, which affect its polymerization (Goeckeler and Wysolmerski 1995; Hirata et al. 2008; Jegou et al. 2013), turnover (Wilson et al. 2010), and the assembly of actin-rich structures such as stress fibers and lamellipodia (Giuliano et al. 1992; Svitkina et al. 1997; Hotulainen and Lappalainen 2006; Goeckeler et al. 2008; Naumanen et al. 2008; Senju and Miyata 2009). As mentioned above, actin associated adhesion structures, both cell–cell and cell–ECM, are also sensitive to mechanical cues that alter their adhesive properties, their molecular composition, and signal transduction (reviewed in Geiger et al. 2001; Schiller and Fassler 2013; Wolfenson et al. 2013; Ladoux et al. 2010; Sumida et al. 2011; Priya and Yap 2015).
With such profound impact of cellular mechanics on the cytoskeleton and associated adhesion structures, it is not surprising that mechanical forces are major regulators of polarity, including front-back polarity (Vicente-Manzanares et al. 2009), apical-basal polarity (Yu et al. 2008), PCP (Lee et al. 2012; Walters et al. 2006), and left-right polarity (Naganathan et al. 2014; Tee et al. 2015).
To illustrate the mechanisms, we would like to describe here the “nodal model.” Ciliated cells at the node of the mouse embryo (the node is a transient structure, a cavity at the posterior part of the notochord) rotate clockwise and generate a directional flow that can be detected by sensory structures and translated into expression of specific genes and establishment of left-right polarity. Defects in the assembly (Nonaka et al. 1998; Takeda et al. 1999) or motility (Supp et al. 1997) of cilia give rise to left-right polarity defects. Moreover, an artificial flow system was able to rescue motile cilia mutants and flip left-right polarity, providing direct evidence for the role of mechanical forces in the establishment of left-right symmetry (Nonaka et al. 2002).
2 Novel Mechanisms in the Establishment of Planar Cell Polarity
2.1 Some Basic Facts
PCP is a tissue-level phenomenon, which plays a fundamental role in tissue morphogenesis. For instance, PCP regulates collective cell migration and convergent extension (a process whereby tissue extends on one axis and narrows down in the orthogonal direction) (Wallingford et al. 2000; Darken et al. 2002; Goto and Keller 2002), as well as mitotic spindle orientation (Lake and Sokol 2009; Segalen and Bellaiche 2009) and cilia positioning and function (Park et al. 2006; Park et al. 2008; Tarchini et al. 2013). Studies in vertebrate model organisms showed that mutations in PCP genes may give rise to many developmental defects and diseases including neural tube closure defects, inner ear and heart development defects, wound healing defects, kidney disease, and cancer (Munoz-Soriano et al. 2012). In humans PCP defects are associated with neural tube closure defects such as spina bifida and craniorachischisis (Lei et al. 2010; Seo et al. 2011; Robinson et al. 2012), and were also reported in several models of human cancers (Hatakeyama et al. 2014).
At the molecular level, two groups of genes control PCP: the “core-PCP” and the “Fat-Dachsaus” groups, yet the mode of molecular interaction between the two pathways is not well understood. Frizzled and disheveled are core-PCP proteins; however, they are also important regulators of the Wnt signaling pathway. Therefore the core-PCP pathway is often referred to as non-canonical Wnt pathway. An important hallmark of these proteins is their asymmetric localization within the cells. Only Celsr (also known as flamingo), a transmembranal atypical cadherin core-PCP protein, is localized on both the anterior and posterior sides of the cell where it creates homodimers that connect neighboring cells (Fig. 2b), all the other PCP proteins in the two groups exhibit asymmetric distribution (reviewed in McNeill 2010; Devenport 2014; Sebbagh and Borg 2014; Sokol 2015).
2.2 Involvement of External Mechanical Forces in Planar Cell Polarity
One of the major challenges in the PCP studies is to identify the upstream cue(s) that provide directionality and determine the global axis of PCP. Recently it was shown that wingless (Wg) and Wnt4 function redundantly to orient PCP in the Drosophila wing disc and affect the axis of polarity by regulating the interaction between two core-PCP proteins [frizzeled and Vangl (Van Gogh)] (Wu et al. 2013). An important work on the same tissue suggested that mechanical forces also affect the orientation of the PCP axis and couple PCP establishment with wing morphogenesis (Aigouy et al. 2010).
During the development of the drosophila wing, the tissue PCP changes its orientation. In early stage PCP is oriented towards the edge of the wing, later on it reorients along the proximal-distal axis (Classen et al. 2005). Live imaging experiments conducted by Aigout and coworkers (Aigouy et al. 2010) demonstrated that the PCP reorientation is temporally correlated with cell and tissue shape changes. At the tissue level the shape dynamics involve the contraction of the wing hinge that loses health of its size while the blade becomes more elongated and narrows down. At the cellular level, oriented cell divisions, changes in cell shape, and establishment of new contacts can be detected. By severing the contracting hinge the authors demonstrated that PCP reorientation and tissue/cell dynamics are coupled. Laser ablation experiments showed that hinge contraction alters force distribution in the tissue from isotropic to anisotropic (Aigouy et al. 2010). Together, these data suggest a key role for mechanical forces in PCP orientation in the fly.
During gastrulation mechanical forces strain the developing skin in Xenopus. At the same time skin planar axis is established. These PCP cues will then guide the development and coherent function of multiciliated epithelial cells at the surface of the tadpole. Chien et al. (2015) noted that ventralized Xenopus embryos that exhibit many patterning defects manage to establish PCP. Since these abnormal embryos do undergo gastrulation that strains the developing skin, the authors asked whether PCP establishment is regulated by mechanical cues. To check this hypothesis, the authors exposed isolated ectoderm that normally does not establish PCP to external forces. By doing so Chien et al. nicely demonstrated that mechanical forces are very effective in establishing the axis of PCP in the developing Xenopus skin (Chien et al. 2015).
2.3 Involvement of Internal Mechanical Forces in Planar Cell Polarity
While external forces (hinge contraction) play a key role in wing development, wing cells are not indifferent bystanders in the process. Wing cells actively generate internal forces that respond to the external forces and together affect wing morphogenesis (Etournay et al. 2015). Moreover, recently, a tissue-specific knockdown of Wdr1 (restricted to the mouse epidermis) demonstrated a key role for internal, actomyosin-generated forces in PCP establishment in a mammalian system (Luxenburg et al. 2015).
During the development of the mouse skin, epidermal cells of the basal layer establish PCP that orients the growth of hair follicles towards the anterior of the embryo (Fig. 2). This process results in a highly ordered pattern of hairs throughout the mouse fur. Defects in the core PCP gene Frizzled6 give rise to an abnormal hair pattern (Guo et al. 2004; Wang et al. 2006). Early in epidermal development [embryonic day (E)12.5] core PCP proteins can be detected throughout the cortex of the cell, a day later early signs of anterior–posterior enrichment can be detected and 2 days later (E14.5) a polarized pattern is readily evident (Devenport and Fuchs 2008). E14.5 is also the first time point in development in which hair follicles begin their development.
The differentiation of keratinocytes involves dramatic changes in cell shape as cells of the basal layer alter their cuboidal architecture and turn flat. Recently, Luxenburg et al. (2015) demonstrated that cytoskeleton-driven cell shape changes take place within the basal layer at the same time that the tissue establishes PCP. During this process the basal layer of epidermal cells becomes more compact and changes their orientation in the tissue. The knockdown of Wdr1, a scaffolding protein that enhances cofilin mediated actin severing activity, negatively affects both cell shape dynamics and the establishment of PCP. In line with these PCP defects, hair follicle orientation and molecular asymmetry are abnormal without Wdr1. Laser ablation experiments showed that boundary tension is significantly reduced without Wdr1, highlighting the involvement of this gene in the ability to generate and maintain cytoskeleton-derived forces and cortical tension in the cell.
In the mouse epidermis core PCP proteins undergo endocytosis on mitosis and will be recycled to the cell surface of the daughter cells (Devenport et al. 2011). When core-PCP mitotic internalization is blocked, cell shape changes that normally accompany mitosis (e.g., mitotic rounding (Luxenburg et al. 2011) abolish the localization of core PCP proteins and planar polarity (Shrestha et al. 2015). This observation further supports a key role for cell shape dynamics in the PCP.
3 Cytoskeletal Involvement in the Acquisition of Left-Right Polarity
3.1 Some Basic Facts
The establishment of left-right symmetry takes place early in development. In C. elegans and Xenopus left-right cues are generated by the actin cytoskeleton and can be detected at the single cell embryo stage (Danilchik et al. 2006; Naganathan et al. 2014). In most vertebrates left-right polarity cues affect the development of many organs, including the heart, lungs, stomach, liver, gallbladder, pancreas, spleen, and the brain. Moreover, many of the large arteries and veins also establish left-right asymmetry during development (Casey and Hackett 2000; Levin 2005). In cultured cells left-right polarity is evident by consistent handedness of cytoskeletal structures (Tee et al. 2015) and consistent directional migration under both confined and isotropic conditions (Nonaka et al. 2002; Xu et al. 2007; Tamada et al. 2010; Yamanaka and Kondo 2015).
A range of left-right polarity defects was described in humans, from complete reversal of all organs known as “situs inversus” to a range of partial defects known as “partial situs inversus,” “situs ambiguus,” “heterotaxy,” or “heterotaxia.” Some of these conditions may be fetal or negatively affect the health of the affected individual (Casey and Hackett 2000).
While defects in left-right polarity were described in snails more than a century ago, the first molecular components that mediate it were described only 20 years ago (Levin et al. 1995). Since then studies in several model organisms and in cultured cells identified a variety of molecular mechanisms that establish, maintain, and amplify left-right polarity. These mechanisms involve both the actin and the microtubule cytoskeleton and their motor proteins, as well as external forces and ion fluxes (for recent reviews, see Coutelis et al. 2014; Hamada and Tam 2014; Yoshiba and Hamada 2014).
3.2 Key Roles for the Actin Cytoskeleton in Left-Right Polarity: Insights from Cultured Cells
The notion that the actin cytoskeleton plays a role in left-right polarity is well established. Wan et al. (2011) demonstrated this phenomenon in many different cell types. In this experimental system cells were cultured in a confined environment and cell alignment and migration were analyzed to determine left-right polarity. Strikingly, all types of cells exhibit handedness. Interestingly, normal and transformed skin fibroblasts exhibit opposite handedness, suggesting that chirality might be altered in specific pathological states, such as cancer. Treatment with drugs that modulate actin dynamics alters handedness, emphasizing the key role of the actin cytoskeleton in this type of polarity (Wan et al. 2011).
Recently Tee and coworkers (2015) provided intriguing insights into the roles of the actin cytoskeleton in left-right polarity. These authors cultured fibroblasts on circular adhesive islands and tracked the actin cytoskeleton by video microscopy. A sequence of five distinct patterns was detected that included a shift from isotropic to chiral pattern with defined handedness. The isotropic pattern was made of radial actin fibers growing from peripheral focal adhesions towards the cell center and transverse fibers that move centripetally. The shift to a chiral pattern takes place at the level of the radial fibers, which uniformly tilt to the same direction, inducing the swirling of the transverse fibers.
To gain mechanistic insights into the establishment of left-right polarity in this system the authors studied the structural, molecular, and dynamic properties of the chiral actin pattern. They demonstrated that while the radial and traverse actin fibers interact physically, their molecular composition is different: the actin bundling protein α-actenin is enriched in the radial fibers while the traverse fibers are enriched in myosin II. Moreover, myosin II motor activity is essential for the assembly of both radial and traverse fibers and its activity within the traverse fibers propel their movement along the radial fibers that rely on formin-mediated actin polymerization for their assembly. Overexpressing of α-actinin flips the handedness of the actin network, suggesting a key role for actin bundling in the development of cell chirality.
3.3 Key Roles for the Actin Cytoskeleton in Left-Right Polarity: Physiological insights
The aforementioned studies demonstrate that the actin cytoskeleton has an intrinsic ability to establish handedness under experimental conditions. A recent study by Naganathan et al. (2014) demonstrated that the actin cytoskeleton plays a key role in establishing left-right polarity also in a physiologically relevant system, the C. elegans embryo. More than a decade ago Munro et al. (2004) showed that sperm entry into C. elegans egg induces flow of actin and myosin that polarizes PAR proteins and establishes anterior–posterior polarity. More recently Naganathan et al. (2014) revisited these early events in C. elegans development, and found that while cortical flow is readily detectable along the anterior–posterior axis, an orthogonal flow can also be detected, leading the anterior and posterior parts of the embryo to counter-rotate, relative to each other. Apparently a significant portion of myosin II activity is dedicated to the generation of torque that propels both the establishment of chirality and anterior–posterior flow. This process is regulated by Rho signaling, and subtle changes in this pathway can alter torque without affecting tension, affecting left-right but not anterior–posterior polarity. This mechanism is relevant also at the 4-cell stage at which C. elegans handedness is established. Interestingly, manipulating Wnt signaling genes that are known to play a role in left-right symmetry altered chiral actin flow. This observation further emphasizes the link between left-right polarity and actomyosin activity.
4 Conclusion
Cells rely on internal and/or external cues to establish asymmetry. In recent years it became evident that mechanical forces applied by the pericellular environment or generated inside cells, by the cytoskeleton, function as important upstream signals that trigger the establishment of cell and tissue polarity.
In the last decade advances in high-resolution microscopy, ECM nano- and micro-fabrication, as well as computational analysis shed new light on the molecular mechanisms underlying the mechanobiology of cell polarity. That said, many key questions are still poorly understood. For instance, it will be important to understand the cross-talk between chemical and mechanical cues in these processes and the identity of the diverse proteins that mediate the acquisition of polarity. These studies will, hopefully, advance our knowledge of cell polarity, with relevance to diverse physiological and pathological processes, including embryonic development, stem cell differentiation, and cancer biology.
References
Aigouy B, Farhadifar R, Staple DB, Sagner A, Roper JC, Julicher F, Eaton S (2010) Cell flow reorients the axis of planar polarity in the wing epithelium of Drosophila. Cell 142(5):773–786
Alberts B (2015) Molecular biology of the cell. Garland Science, Taylor and Francis Group, New York
Balaban NQ, Schwarz US, Riveline D, Goichberg P, Tzur G, Sabanay I, Mahalu D, Safran S, Bershadsky A, Addadi L, Geiger B (2001) Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat Cell Biol 3(5):466–472
Beausang JF, Schroeder HW 3rd, Nelson PC, Goldman YE (2008) Twirling of actin by myosins II and V observed via polarized TIRF in a modified gliding assay. Biophys J 95(12):5820–5831
Bertocchi C, Vaman Rao M, Zaidel-Bar R (2012) Regulation of adherens junction dynamics by phosphorylation switches. J Signal Transduct 2012:125295
Braga VM, Yap AS (2005) The challenges of abundance: epithelial junctions and small GTPase signalling. Curr Opin Cell Biol 17(5):466–474
Burger MM, Jumblatt J (1977) Membrane involvement in cell-cell interactions: a two-component model system for cellular recognition that does not require live cells. Soc Gen Physiol Ser 32:155–172
Casey B, Hackett BP (2000) Left-right axis malformations in man and mouse. Curr Opin Genet Dev 10(3):257–261
Chien YH, Keller R, Kintner C, Shook DR (2015) Mechanical strain determines the axis of planar polarity in ciliated epithelia. Curr Biol 25(21):2774–2784
Classen AK, Anderson KI, Marois E, Eaton S (2005) Hexagonal packing of Drosophila wing epithelial cells by the planar cell polarity pathway. Dev Cell 9(6):805–817
Conway Morris S (1998) The question of metazoan monophyly and the fossil record. Prog Mol Subcell Biol 21:1–19
Coutelis JB, Gonzalez-Morales N, Geminard C, Noselli S (2014) Diversity and convergence in the mechanisms establishing L/R asymmetry in metazoa. EMBO Rep 15(9):926–937
Danilchik MV, Brown EE, Riegert K (2006) Intrinsic chiral properties of the Xenopus egg cortex: an early indicator of left-right asymmetry? Development 133(22):4517–4526
Darken RS, Scola AM, Rakeman AS, Das G, Mlodzik M, Wilson PA (2002) The planar polarity gene strabismus regulates convergent extension movements in Xenopus. EMBO J 21(5):976–985
De La Cruz EM, Gardel ML (2015) Actin mechanics and fragmentation. J Biol Chem 290(28):17137–17144
Devenport D (2014) The cell biology of planar cell polarity. J Cell Biol 207(2):171–179
Devenport D, Fuchs E (2008) Planar polarization in embryonic epidermis orchestrates global asymmetric morphogenesis of hair follicles. Nat Cell Biol 10(11):1257–1268
Devenport D, Oristian D, Heller E, Fuchs E (2011) Mitotic internalization of planar cell polarity proteins preserves tissue polarity. Nat Cell Biol 13(8):893–902
Devreotes P, Horwitz AR (2015) Signaling networks that regulate cell migration. Cold Spring Harb Perspect Biol 7(8)
Etournay R, Popovic M, Merkel M, Nandi A, Blasse C, Aigouy B, Brandl H, Myers G, Salbreux G, Julicher F, Eaton S (2015) Interplay of cell dynamics and epithelial tension during morphogenesis of the pupal wing. Elife 4:e07090
Farquhar MG, Palade GE (1963) Junctional complexes in various epithelia. J Cell Biol 17:375–412
Fischer E, Legue E, Doyen A, Nato F, Nicolas JF, Torres V, Yaniv M, Pontoglio M (2006) Defective planar cell polarity in polycystic kidney disease. Nat Genet 38(1):21–23
Geiger B, Yamada KM (2011) Molecular architecture and function of matrix adhesions. Cold Spring Harb Perspect Biol 3(5)
Geiger B, Schmid E, Franke WW (1983) Spatial distribution of proteins specific for desmosomes and adhaerens junctions in epithelial cells demonstrated by double immunofluorescence microscopy. Differentiation 23(3):189–205
Geiger B, Bershadsky A, Pankov R, Yamada KM (2001) Transmembrane crosstalk between the extracellular matrix – cytoskeleton crosstalk. Nat Rev Mol Cell Biol 2(11):793–805
Giuliano KA, Kolega J, DeBiasio RL, Taylor DL (1992) Myosin II phosphorylation and the dynamics of stress fibers in serum-deprived and stimulated fibroblasts. Mol Biol Cell 3(9):1037–1048
Goeckeler ZM, Wysolmerski RB (1995) Myosin light chain kinase-regulated endothelial cell contraction: the relationship between isometric tension, actin polymerization, and myosin phosphorylation. J Cell Biol 130(3):613–627
Goeckeler ZM, Bridgman PC, Wysolmerski RB (2008) Nonmuscle myosin II is responsible for maintaining endothelial cell basal tone and stress fiber integrity. Am J Physiol Cell Physiol 295(4):C994–C1006
Goto T, Keller R (2002) The planar cell polarity gene strabismus regulates convergence and extension and neural fold closure in Xenopus. Dev Biol 247(1):165–181
Guo N, Hawkins C, Nathans J (2004) Frizzled6 controls hair patterning in mice. Proc Natl Acad Sci U S A 101(25):9277–9281
Guo Z, Neilson LJ, Zhong H, Murray PS, Zanivan S, Zaidel-Bar R (2014) E-cadherin interactome complexity and robustness resolved by quantitative proteomics. Sci Signal 7(354):rs7
Guyer RA, Macara IG (2015) Loss of the polarity protein PAR3 activates STAT3 signaling via an atypical protein kinase C (aPKC)/NF-kappaB/interleukin-6 (IL-6) axis in mouse mammary cells. J Biol Chem 290(13):8457–8468
Hamada H, Tam PP (2014) Mechanisms of left-right asymmetry and patterning: driver, mediator and responder. F1000Prime Rep 6:110
Harris TJ, Tepass U (2010) Adherens junctions: from molecules to morphogenesis. Nat Rev Mol Cell Biol 11(7):502–514
Hartman MA, Spudich JA (2012) The myosin superfamily at a glance. J Cell Sci 125(Pt 7):1627–1632
Hatakeyama J, Wald JH, Printsev I, Ho HY, Carraway KL 3rd (2014) Vangl1 and Vangl2: planar cell polarity components with a developing role in cancer. Endocr Relat Cancer 21(5):R345–R356
Hirata H, Tatsumi H, Sokabe M (2008) Mechanical forces facilitate actin polymerization at focal adhesions in a zyxin-dependent manner. J Cell Sci 121(Pt 17):2795–2804
Horton ER, Astudillo P, Humphries MJ, Humphries JD (2016) Mechanosensitivity of integrin adhesion complexes: role of the consensus adhesome. Exp Cell Res 343(1):7–13
Hotulainen P, Lappalainen P (2006) Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J Cell Biol 173(3):383–394
Iskratsch T, Wolfenson H, Sheetz MP (2014) Appreciating force and shape-the rise of mechanotransduction in cell biology. Nat Rev Mol Cell Biol 15(12):825–833
Jegou A, Carlier MF, Romet-Lemonne G (2013) Formin mDia1 senses and generates mechanical forces on actin filaments. Nat Commun 4:1883
Juriloff DM, Harris MJ (2012) A consideration of the evidence that genetic defects in planar cell polarity contribute to the etiology of human neural tube defects. Birth Defects Res A Clin Mol Teratol 94(10):824–840
Labat-Robert J, Robert L, Auger C, Lethias C, Garrone R (1981) Fibronectin-like protein in Porifera: its role in cell aggregation. Proc Natl Acad Sci U S A 78(10):6261–6265
Ladoux B, Anon E, Lambert M, Rabodzey A, Hersen P, Buguin A, Silberzan P, Mege RM (2010) Strength dependence of cadherin-mediated adhesions. Biophys J 98(4):534–542
Lake BB, Sokol SY (2009) Strabismus regulates asymmetric cell divisions and cell fate determination in the mouse brain. J Cell Biol 185(1):59–66
Lawson CD, Burridge K (2014) The on-off relationship of Rho and Rac during integrin-mediated adhesion and cell migration. Small GTPases 5, e27958
Lecuit T, Yap AS (2015) E-cadherin junctions as active mechanical integrators in tissue dynamics. Nat Cell Biol 17(5):533–539
Lee J, Andreeva A, Sipe CW, Liu L, Cheng A, Lu X (2012) PTK7 regulates myosin II activity to orient planar polarity in the mammalian auditory epithelium. Curr Biol 22(11):956–966
Lei YP, Zhang T, Li H, Wu BL, Jin L, Wang HY (2010) VANGL2 mutations in human cranial neural-tube defects. N Engl J Med 362(23):2232–2235
Levin M (2005) Left-right asymmetry in embryonic development: a comprehensive review. Mech Dev 122(1):3–25
Levin M, Johnson RL, Stern CD, Kuehn M, Tabin C (1995) A molecular pathway determining left-right asymmetry in chick embryogenesis. Cell 82(5):803–814
Li R, Gundersen GG (2008) Beyond polymer polarity: how the cytoskeleton builds a polarized cell. Nat Rev Mol Cell Biol 9(11):860–873
Luxenburg C, Pasolli HA, Williams SE, Fuchs E (2011) Developmental roles for Srf, cortical cytoskeleton and cell shape in epidermal spindle orientation. Nat Cell Biol 13(3):203–214
Luxenburg C, Heller E, Pasolli HA, Chai S, Nikolova M, Stokes N, Fuchs E (2015) Wdr1-mediated cell shape dynamics and cortical tension are essential for epidermal planar cell polarity. Nat Cell Biol 17(5):592–604
Martin-Belmonte F, Perez-Moreno M (2012) Epithelial cell polarity, stem cells and cancer. Nat Rev Cancer 12(1):23–38
Martindale MQ (2005) The evolution of metazoan axial properties. Nat Rev Genet 6(12):917–927
McDonald JA, Mecham RP (1991) Receptors for extracellular matrix. Academic, San Diego
McNeill H (2010) Planar cell polarity: keeping hairs straight is not so simple. Cold Spring Harb Perspect Biol 2(2):a003376
Meldolesi J (2016) Pharmacology of the cell/matrix form of adhesion. Pharmacol Res 107:430–436
Muller WE, Muller IM (2003) Origin of the metazoan immune system: identification of the molecules and their functions in sponges. Integr Comp Biol 43(2):281–292
Muller WE, Zahn RK, Conrad J, Kurelec B, Uhlenbruck G (1982) Aggregation of sponge cells: stage dependent, distinct adhesion mechanisms in Cliona celata. Eur J Cell Biol 28(2):243–250
Mullins RD (2010) Cytoskeletal mechanisms for breaking cellular symmetry. Cold Spring Harb Perspect Biol 2(1):a003392
Munoz-Soriano V, Belacortu Y, Paricio N (2012) Planar cell polarity signaling in collective cell movements during morphogenesis and disease. Curr Genomics 13(8):609–622
Munro E, Nance J, Priess JR (2004) Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo. Dev Cell 7(3):413–424
Naganathan SR, Furthauer S, Nishikawa M, Julicher F, Grill SW (2014) Active torque generation by the actomyosin cell cortex drives left-right symmetry breaking. Elife 3, e04165
Narimatsu M, Samavarchi-Tehrani P, Varelas X, Wrana JL (2015) Distinct polarity cues direct Taz/Yap and TGFbeta receptor localization to differentially control TGFbeta-induced Smad signaling. Dev Cell 32(5):652–656
Naumanen P, Lappalainen P, Hotulainen P (2008) Mechanisms of actin stress fibre assembly. J Microsc 231(3):446–454
Nonaka S, Tanaka Y, Okada Y, Takeda S, Harada A, Kanai Y, Kido M, Hirokawa N (1998) Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 95(6):829–837
Nonaka S, Shiratori H, Saijoh Y, Hamada H (2002) Determination of left-right patterning of the mouse embryo by artificial nodal flow. Nature 418(6893):96–99
Park TJ, Haigo SL, Wallingford JB (2006) Ciliogenesis defects in embryos lacking inturned or fuzzy function are associated with failure of planar cell polarity and Hedgehog signaling. Nat Genet 38(3):303–311
Park TJ, Mitchell BJ, Abitua PB, Kintner C, Wallingford JB (2008) Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nat Genet 40(7):871–879
Pollard TD (2007) Regulation of actin filament assembly by Arp2/3 complex and formins. Annu Rev Biophys Biomol Struct 36:451–477
Pollard TD, Cooper JA (2009) Actin, a central player in cell shape and movement. Science 326(5957):1208–1212
Prager-Khoutorsky M, Lichtenstein A, Krishnan R, Rajendran K, Mayo A, Kam Z, Geiger B, Bershadsky AD (2011) Fibroblast polarization is a matrix-rigidity-dependent process controlled by focal adhesion mechanosensing. Nat Cell Biol 13(12):1457–1465
Price LS, Leng J, Schwartz MA, Bokoch GM (1998) Activation of Rac and Cdc42 by integrins mediates cell spreading. Mol Biol Cell 9(7):1863–1871
Priya R, Yap AS (2015) Active tension: the role of cadherin adhesion and signaling in generating junctional contractility. Curr Top Dev Biol 112:65–102
Reinhart-King CA, Dembo M, Hammer DA (2005) The dynamics and mechanics of endothelial cell spreading. Biophys J 89(1):676–689
Robinson A, Escuin S, Doudney K, Vekemans M, Stevenson RE, Greene ND, Copp AJ, Stanier P (2012) Mutations in the planar cell polarity genes CELSR1 and SCRIB are associated with the severe neural tube defect craniorachischisis. Hum Mutat 33(2):440–447
Rodriguez-Boulan E, Macara IG (2014) Organization and execution of the epithelial polarity programme. Nat Rev Mol Cell Biol 15(4):225–242
Roignot J, Peng X, Mostov K (2013) Polarity in mammalian epithelial morphogenesis. Cold Spring Harb Perspect Biol 5(2)
Sasaki H (2015) Position- and polarity-dependent Hippo signaling regulates cell fates in preimplantation mouse embryos. Semin Cell Dev Biol 47–48:80–7
Sase I, Miyata H, Ishiwata S, Kinosita K Jr (1997) Axial rotation of sliding actin filaments revealed by single-fluorophore imaging. Proc Natl Acad Sci U S A 94(11):5646–5650
Schiller HB, Fassler R (2013) Mechanosensitivity and compositional dynamics of cell-matrix adhesions. EMBO Rep 14(6):509–519
Sebbagh M, Borg JP (2014) Insight into planar cell polarity. Exp Cell Res 328(2):284–295
Segalen M, Bellaiche Y (2009) Cell division orientation and planar cell polarity pathways. Semin Cell Dev Biol 20(8):972–977
Senju Y, Miyata H (2009) The role of actomyosin contractility in the formation and dynamics of actin bundles during fibroblast spreading. J Biochem 145(2):137–150
Seo JH, Zilber Y, Babayeva S, Liu J, Kyriakopoulos P, De Marco P, Merello E, Capra V, Gros P, Torban E (2011) Mutations in the planar cell polarity gene, Fuzzy, are associated with neural tube defects in humans. Hum Mol Genet 20(22):4324–4333
Shrestha R, Little KA, Tamayo JV, Li W, Perlman DH, Devenport D (2015) Mitotic control of planar cell polarity by polo-like kinase 1. Dev Cell 33(5):522–534
Sokol SY (2015) Spatial and temporal aspects of Wnt signaling and planar cell polarity during vertebrate embryonic development. Semin Cell Dev Biol 42:78–85
Sumida GM, Tomita TM, Shih W, Yamada S (2011) Myosin II activity dependent and independent vinculin recruitment to the sites of E-cadherin-mediated cell-cell adhesion. BMC Cell Biol 12:48
Supp DM, Witte DP, Potter SS, Brueckner M (1997) Mutation of an axonemal dynein affects left-right asymmetry in inversus viscerum mice. Nature 389(6654):963–966
Svitkina TM, Verkhovsky AB, McQuade KM, Borisy GG (1997) Analysis of the actin-myosin II system in fish epidermal keratocytes: mechanism of cell body translocation. J Cell Biol 139(2):397–415
Takeda S, Yonekawa Y, Tanaka Y, Okada Y, Nonaka S, Hirokawa N (1999) Left-right asymmetry and kinesin superfamily protein KIF3A: new insights in determination of laterality and mesoderm induction by kif3A−/− mice analysis. J Cell Biol 145(4):825–836
Tamada A, Kawase S, Murakami F, Kamiguchi H (2010) Autonomous right-screw rotation of growth cone filopodia drives neurite turning. J Cell Biol 188(3):429–441
Tarchini B, Jolicoeur C, Cayouette M (2013) A molecular blueprint at the apical surface establishes planar asymmetry in cochlear hair cells. Dev Cell 27(1):88–102
Tee YH, Shemesh T, Thiagarajan V, Hariadi RF, Anderson KL, Page C, Volkmann N, Hanein D, Sivaramakrishnan S, Kozlov MM, Bershadsky AD (2015) Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat Cell Biol 17(4):445–457
Tellkamp F, Vorhagen S, Niessen CM (2014) Epidermal polarity genes in health and disease. Cold Spring Harb Perspect Med 4(12):a015255
Vicente-Manzanares M, Ma X, Adelstein RS, Horwitz AR (2009) Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat Rev Mol Cell Biol 10(11):778–790
Volberg T, Geiger B, Kartenbeck J, Franke WW (1986) Changes in membrane-microfilament interaction in intercellular adherens junctions upon removal of extracellular Ca2+ ions. J Cell Biol 102(5):1832–1842
Vorhagen S, Niessen CM (2014) Mammalian aPKC/Par polarity complex mediated regulation of epithelial division orientation and cell fate. Exp Cell Res 328(2):296–302
Wagner-Hulsmann C, Bachinski N, Diehl-Seifert B, Blumbach B, Steffen R, Pancer Z, Muller WE (1996) A galectin links the aggregation factor to cells in the sponge (Geodia cydonium) system. Glycobiology 6(8):785–793
Wallingford JB, Rowning BA, Vogeli KM, Rothbacher U, Fraser SE, Harland RM (2000) Dishevelled controls cell polarity during Xenopus gastrulation. Nature 405(6782):81–85
Walters JW, Dilks SA, DiNardo S (2006) Planar polarization of the denticle field in the Drosophila embryo: roles for Myosin II (zipper) and fringe. Dev Biol 297(2):323–339
Wan LQ, Ronaldson K, Park M, Taylor G, Zhang Y, Gimble JM, Vunjak-Novakovic G (2011) Micropatterned mammalian cells exhibit phenotype-specific left-right asymmetry. Proc Natl Acad Sci U S A 108(30):12295–12300
Wang Y, Badea T, Nathans J (2006) Order from disorder: self-organization in mammalian hair patterning. Proc Natl Acad Sci U S A 103(52):19800–19805
Wedlich-Soldner R, Altschuler S, Wu L, Li R (2003) Spontaneous cell polarization through actomyosin-based delivery of the Cdc42 GTPase. Science 299(5610):1231–1235
Wiens M, Mangoni A, D’Esposito M, Fattorusso E, Korchagina N, Schroder HC, Grebenjuk VA, Krasko A, Batel R, Muller IM, Muller WE (2003) The molecular basis for the evolution of the metazoan bodyplan: extracellular matrix-mediated morphogenesis in marine demosponges. J Mol Evol 57(Suppl 1):S60–S75
Williams SE, Fuchs E (2013) Oriented divisions, fate decisions. Curr Opin Cell Biol 25(6):749–758
Wilson CA, Tsuchida MA, Allen GM, Barnhart EL, Applegate KT, Yam PT, Ji L, Keren K, Danuser G, Theriot JA (2010) Myosin II contributes to cell-scale actin network treadmilling through network disassembly. Nature 465(7296):373–377
Winograd-Katz SE, Fassler R, Geiger B, Legate KR (2014) The integrin adhesome: from genes and proteins to human disease. Nat Rev Mol Cell Biol 15(4):273–288
Wolfenson H, Lavelin I, Geiger B (2013) Dynamic regulation of the structure and functions of integrin adhesions. Dev Cell 24(5):447–458
Wu J, Roman AC, Carvajal-Gonzalez JM, Mlodzik M (2013) Wg and Wnt4 provide long-range directional input to planar cell polarity orientation in Drosophila. Nat Cell Biol 15(9):1045–1055
Xu J, Van Keymeulen A, Wakida NM, Carlton P, Berns MW, Bourne HR (2007) Polarity reveals intrinsic cell chirality. Proc Natl Acad Sci U S A 104(22):9296–9300
Yamanaka H, Kondo S (2015) Rotating pigment cells exhibit an intrinsic chirality. Genes Cells 20(1):29–35
Yoshiba S, Hamada H (2014) Roles of cilia, fluid flow, and Ca2+ signaling in breaking of left-right symmetry. Trends Genet 30(1):10–17
Yu W, Shewan AM, Brakeman P, Eastburn DJ, Datta A, Bryant DM, Fan QW, Weiss WA, Zegers MM, Mostov KE (2008) Involvement of RhoA, ROCK I and myosin II in inverted orientation of epithelial polarity. EMBO Rep 9(9):923–929
Zaidel-Bar R, Geiger B (2010) The switchable integrin adhesome. J Cell Sci 123(Pt 9):1385–1388
Zaidel-Bar R, Itzkovitz S, Ma’ayan A, Iyengar R, Geiger B (2007) Functional atlas of the integrin adhesome. Nat Cell Biol 9(8):858–867
Zaidel-Bar R, Zhenhuan G, Luxenburg C (2015) The contractome – a systems view of actomyosin contractility in non-muscle cells. J Cell Sci 128(12):2209–2217
Acknowledgments
The authors are thankful to E. Fuchs for providing Celsr1 antibody. C.L. is supported by a grant from the I-CORE Gene Regulation in Complex Human Disease, Center number 41/11, and the Israel Science Foundation grant number 1113/15. BG is the incumbent of the Erwin Netter Chair in cell and tumor biology and recipient of grants from the Leona M. and Harry B. Helmsley Charitable Trust and the ERC SynAd project (Grant agreement No.: 294852)
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Luxenburg, C., Geiger, B. (2016). Multiscale View of Cytoskeletal Mechanoregulation of Cell and Tissue Polarity. In: Jockusch, B. (eds) The Actin Cytoskeleton. Handbook of Experimental Pharmacology, vol 235. Springer, Cham. https://doi.org/10.1007/164_2016_34
Download citation
DOI: https://doi.org/10.1007/164_2016_34
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-46369-8
Online ISBN: 978-3-319-46371-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)