
Abstract
Visual search benefits from advance knowledge of nontarget features. However, it is unknown whether these negatively cued features are suppressed in advance (proactively) or during search (reactively). To test this, we presented color cues varying from trial-to-trial that predicted target or nontarget colors. Experiment 1 (N = 96) showed that both target and nontarget cues speeded search. To test whether attention proactively modified cued feature representations, in Experiment 2 (N = 200), we interleaved color probe and search trials and had participants detect the color of a briefly presented ring that could either match the cued color or not. People detected positively cued colors better than other colors, whereas negatively cued colors were detected no better or worse than other colors. These results demonstrate that nontarget features are not suppressed proactively, and instead suggest that anticipated nontarget features are ignored via reactive mechanisms.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Perception frequently involves selecting some stimuli while ignoring others, and major theories of visual attention propose that this selection process comprises two distinct mechanisms of enhancement and suppression. Indeed, in tasks like visual search, people are faster when either told which feature to select (e.g., by ‘positive cues’ indicating an upcoming target color) or which feature to ignore (e.g., by ‘negative cues’ indicating an upcoming nontarget color; Arita et al., 2012; Chang & Egeth, 2019); however, it is unclear how exactly these cues, especially negative cues, aid target selection. Several studies show that, when participants attend a particular feature (e.g., the color red), processing of that feature is enhanced, both globally across the visual field (Chapman & Stoermer, 2021; Ho et al., 2012; Martinez-Trujillo & Treue, 2004) and in some cases even before search stimuli appear (Giesbrecht et al., 2006). In contrast, relatively little is known about how people use negative cues to ignore distractors. While some studies suggest that distractor ignoring is supported by proactive suppression of visual features, others suggest that ignoring is reactive and only occurs during search itself.
Analogous to anticipatory enhancement (Giesbrecht et al., 2006; Kastner et al., 1999), proactive suppression accounts propose that negative cues attenuate processing of specific nontarget features prior to the onset of the task display (for reviews, see Geng, 2014; Geng et al., 2019). The most direct evidence for proactive suppression is reports of trial-by-trial correlations between the magnitude of prestimulus alpha power (an electrophysiological measure that has been linked to attentional suppression) and the degree of attentional capture (Mazaheri et al., 2011). Similarly, both EEG and functional MRI have shown preparatory suppression of neural activity in visual areas in response to negative feature cues (de Vries et al., 2019; Reeder et al., 2017). While these findings have been interpreted as evidence for proactive suppression, these neural effects do not necessarily reflect suppression of a specific feature value, as they might instead index more general attentional control processes (Reeder et al., 2018). Thus, it remains unknown whether specific feature values can be voluntarily suppressed in advance.
Other studies have proposed an alternative to proactive suppression in which distractor ignoring is not anticipatory but follows display onset. For example, one study cued participants about to-be-attended or to-be-ignored colors for a search task and measured attentional allocation to the locations of these colors on separate probe trials, in which participants recalled the identities of letters overlaid on search items. Negative cues had no effect on letter recall when presented shortly after the search array (25-ms delay); with a 100-ms delay, however, participants recalled more probes on target-colored items following both positive and negative cues, though these effects were smaller for negative cues, mirroring the pattern found during search itself (Zhang et al., 2020). These results suggest that people may reactively suppress cued distractors shortly after search display onset, and that this is less effective than enhancement. Another proposed process of reactive ignoring is ‘search-and-destroy,’ in which negatively cued features are initially attended and then ignored (Beck et al., 2017; Gaspelin et al., 2019; Moher & Egeth, 2012). Search-and-destroy assumes that both to-be-attended and to-be-ignored feature representations are enhanced but then used differently during the search process itself: items with to-be-attended features are marked for selection, while items with to-be-ignored features are marked to be discarded, perhaps via spatial attention.
In sum, studies have provided mixed evidence regarding how advance knowledge of to-be-ignored features benefits visual search. One shortcoming of past studies, in our view, is that processing of irrelevant items is often measured after a multi-item search display appears. These paradigms cannot directly assess proactive attention to specific features, because as soon as a multi-item displays appears, both feature-based and space-based attention contribute to selection. Because spatial and featural attention may involve different mechanisms of enhancement and suppression, assessing these processes during multi-item tasks tells us little about whether and how precues proactively affect feature representations themselves.
To address this, we informed participants about upcoming target or nontarget features for a visual search task and directly measured visual representations of to-be-attended and to-be-ignored features using a single-item color probe task. In Experiment 1, we establish that both positive and negative cues that vary trial by trial improve search performance. In Experiment 2, we directly assess whether these cues modulate feature representations prior to search onset. We randomly intermixed visual search trials with trials in which participants detected the presence or absence of hue in a briefly presented probe that could either match the cued color or not (i.e., reported whether the probe was gray or any other color). Because probes were presented alone, briefly (~30 ms), before search, and at random locations, results cannot be explained by strategic shifts of spatial attention. If participants enhance cued feature representations in advance, this should result in better probe detection for matching colors; conversely, if cued features are suppressed, this should decrease detection performance for the cued color. Overall, we find that positive cues boost color representations prior to search onset, consistent with proactive enhancement, whereas negative cues aid target selection reactively, challenging the proactive suppression account of negative cueing.
Experiment 1
This experiment tested how positive and negative cues affect search response times (RT). Experiment 1a used positive cues that were 75% predictive of the upcoming search target color; Experiment 1b used negative cues that were 75% predictive of the upcoming nontarget color. Based on past results (Arita et al., 2012), we expected both positive and negative cues to speed performance when valid, with larger RT benefits for positive than negative cues.
Method
Participants
A power analysis of an unpublished experiment similar to Experiment 1b (cue color varied block-by-block rather than trial-by-trial) indicated that detecting significant cue validity effects at 80% power and an alpha level of .05 would require 48 participants per experiment (1a and 1b). We collected data until we had 48 participants per experiment that met our inclusion criteria: we excluded 16 participants in Experiment 1a (two for taking longer than 90 minutes to complete the experiment; 14 for accuracy below 80% in any block) and 19 in 1b (three for taking longer than 90 minutes; 15 for accuracy below 80% in any block; one for exceeding 10% of trials with RTs below 200 ms or above 4 seconds in any block). After exclusion, Experiment 1a included 37 women and 11 men with a mean age of 20.1 (range: 18–26 years); Experiment 1b included 36 women, 11 men, and 1 nonbinary person with a mean age of 20.6 (range: 18–24 years).
For all experiments, participants were students from Dartmouth College or the University of California, San Diego, participant pools. Participants volunteered and were compensated with extra course credit. Both experiments were approved by the Dartmouth College and the UCSD Institutional Review Boards and participants provided informed consent prior to participating. Participants completed this experiment in a web browser on devices they provided. We requested that they complete the experiment in full-screen mode on a computer (not a mobile device).
Stimuli
Throughout the experiment (except when the central color cue was present), a black fixation cross was present at the center of the 600 × 600-pixel stimulus display window. Cues were colored rings 90 pixels in diameter (~1° visual angle on a 13-inch MacBook Pro from 60 cm distance) with a line thickness of 15 pixels presented centrally. Search items were identical to the cue, except they had a 5-pixel gap at 0°, 90°, 180°, or 270°. All distractors had gaps at the top or bottom, and the target item had its gap on the left or right. Eight search items were presented on an invisible circle 150 pixels (~1.7°) in radius. For each participant, four colors were used as both cues and search items. Colors were chosen 90° apart from a color circle in CIE L*a*b space (radius 49, centered at L = 54, a = 21.5, b = 11.5; Suchow et al., 2013). Four specific colors were randomly chosen for each participant. Each search display contained four items in each of two colors.
Procedure
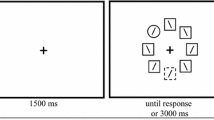
Participants began each trial by pressing the ‘down’ arrow key. This was followed by a search cue for 500 ms, then fixation for 500 ms, after which the search array appeared until response. On each trial, the target and three distractors appeared in one color (the ‘target color’) and four other distractors appeared in another color (the ‘nontarget color’). Locations of colored items were randomly intermixed throughout the search array. Participants searched for the circle with a gap on the left or right side and indicated the gap’s direction using the left and right arrow keys. Instructions emphasized both speed (‘try to respond in under 1.5 s’) and accuracy and told participants to direct attention towards (Exp. 1a) or away from (Exp. 1b) the cue color. Accuracy and RT feedback followed each trial. See Fig. 1 for a trial schematic.

Experiment 1’s trial structure. On each trial, a colored ring indicated the upcoming target (in Exp. 1a) or nontarget (Exp. 1b) color with 75% validity. Following a blank display, a circular display of eight search stimuli was presented until the end of the trial. Participants judged the gap location of the single target Landolt C with its gap on the left or right, with distractors containing gaps on the top or bottom. The depicted trial could be either an invalid positive cue trial (Exp. 1a) or a valid negative cue trial (Exp. 1b.)
Participants completed five practice trials followed by 648 trials separated into six experimental blocks. Cues appeared equally often in each color in each block and were 75% predictive of either the target color (Exp. 1a) or the nontarget color (Exp. 1b). There were three cue conditions: valid, when the cue is accurate (i.e., congruent with either the target color in Exp. 1a or the nontarget color in Exp. 1b); invalid, when the cue is inaccurate (i.e., congruent with the nontarget color in Exp. 1a or the target color in Exp. 1b); and neutral, when the cue color is absent from the search array. We counterbalanced the frequency of these conditions within each block according to frequencies expected by randomly selecting colors with the restriction that cues were 75% predictive. This resulted in 81 valid trials, 18 neutral trials, and 9 invalid trials per block.
Analysis
All exclusion criteria were determined a priori and were the same across all experiments. Data from participants were excluded if mean accuracy was below 80% in any experimental block; if a participant took more than 90 minutes to complete the experiment; or if more than 10% of trials were had outlier RTs (see Participants section for exclusion rates). We excluded individual trials with inaccurate responses or outlier RTs (below 200 ms or above 4 s). We then calculated mean RT for correct trials for each participant in each cue condition (valid, invalid, and neutral), and conducted a one-way analysis of variance (ANOVA) on these data, separately for Experiment 1a and 1b. We conducted planned pairwise comparisons on ANOVA results significant at a .05 alpha level. As expected, accuracy was near ceiling in all cue conditions (minimum condition accuracy was 95.7%), so we did not conduct analyses on accuracy data. Analyses were conducted in R using the rstatix package (Kassambara, 2020).
Results
Experiment 1a: Positive cues
Positive cues sped responses to congruent targets and slowed responses when the cue color was present as the nontarget color. A one-way ANOVA showed strong evidence for an effect of cue condition (valid, invalid, neutral) on RT, F(2, 94) = 25.588, p < .001, ηp2 = .35 (see Fig. 2a). Follow-up dependent-samples t tests compared each pair of cue conditions, finding evidence for reliable differences between all condition pairs. Valid cue trials were on average 60 ms faster than neutral trials (95% CI [32, 87]), t(47) = 4.36, p < .001, d = 0.63, and 159 ms faster than invalid trials (95% CI [99, 219]), t(47) = 5.36, p < .001, d = 0.77, and neutral trials were 100 ms faster than invalid trials (95% CI [57, 142]), t(47) = 4.70, p < .001, d = 0.68.

Results from all experiments. a Mean search RT as a function of cue condition in Experiment 1a, which used 75% predictive positive cues. Valid cues matched the target color; neutral cues were in a color not present in the trial; invalid cues matched the nontarget color. b Mean search response time as a function of cue condition in Experiment 1b, which used 75% predictive negative cues. Valid cues matched the nontarget color; neutral cues were in a color that was absent from the trial; invalid cues matched the target color. c Mean color detection accuracy as a function of congruency between cue and probe color in Experiment 2a, which used positive cues that indicated the target color in search trials. Congruent probes matched the cue color; incongruent cues matched another possible search color. d Mean color detection accuracy as a function of congruency between cue and probe color in Experiment 2b, which used negative cues that indicated the nontarget color in search trials. Congruent probes matched the cue color; incongruent cues were another possible search color. Error bars reflect 95% confidence intervals corrected for within-subjects designs as recommended in Morey (2008)
Experiment 1b: Negative cues
Like positive cues, negative cues sped responses on valid trials.
A one-way ANOVA showed evidence for an effect of cue condition (valid, invalid, neutral) on RT, F(2, 94) = 4.772, p = .011, ηp2 = .092 (see Fig. 2b). Dependent-samples t tests compared each pair of cue conditions, finding significant differences between valid cues and both invalid cues, t(47) = 2.46, p = .018, d = 0.35, and neutral cues, t(47) = 3.27, p = .002, d = 0.47. In contrast, there was no reliable difference between invalid and neutral cue trials, t(47) = 1.21, p = 0.231, d = 0.18. Numerically, response times in each condition were consistent with expected effects of negative cueing: 40-ms faster on valid trials than invalid trials (95% CI [7, 73]), 23 ms faster on valid than neutral trials (95% CI [8, 37]), and 17 ms faster on neutral than invalid trials (95% CI [−11, 45]).
Comparing positive and negative cues
We also compared the strength of positive and negative cueing by calculating the average RT difference between each pair of cue validity conditions (valid–neutral, valid–invalid, and neutral–invalid). We then used independent-means t tests for each of these cue validity pairs across cue types. For all three tests, positive cues yielded significantly larger RT effects than did negative cues. Positive cues yielded a 36-ms larger benefit than negative cues on valid relative to neutral trials (95% CI [5, 67]), t(94) = 2.37, p = .020, d = 0.36, a 119-ms larger advantage on valid relative to invalid trials (95% CI [52, 186]), t(94) = 3.52, p < .001, d = 0.54, and an 82-ms larger advantage on neutral trials relative to invalid trials (95% CI [32, 133]), t(94) = 3.27, p = .001, d = 0.51. Thus, positive cues elicited larger search benefits than negative cues: boosting performance more than negative cues when valid and impairing performance more than negative cues when invalid.
Experiment 2
Experiment 1 showed clear benefits of positive and negative cues during visual search, even when cued features varied from trial to trial (in contrast to some studies; e.g., Cunningham & Egeth, 2016). We also showed that cues providing target information yield stronger effects than equally informative cues providing distractor information, consistent with other studies that have reported smaller search benefits for negative relative to positive cues (e.g., Arita et al., 2012). However, search performance alone cannot identify what mechanisms underlie these effects or why negative cueing speeds search less than positive cueing. Thus, Experiment 2 directly assesses whether attention proactively enhances or suppresses color representations in anticipation of search. We intermixed visual search trials with infrequent probe trials that allowed us to measure perceptual sensitivity to—and thus the activation level of—the cued color. Specifically, on a small number of trials, we briefly presented a probe stimulus that could either match the cued color or not and asked participants to discriminate whether the probe was colorful or gray; in other words, participants had to detect any hue in the probe. If attending towards positive cues enhances processing of the cued color, as expected based on results from feature-based attentional enhancement (Martinez-Trujillo & Treue, 2004; Störmer & Alvarez, 2014), the detection of color probes matching the cue should be higher relative to nonmatching colors. If cues suppress feature representations, as may be the case for negative cues, and assuming this suppression is reflected in the attenuation of specific color representations (Störmer & Alvarez, 2014), detection of the cued color should be worse relative to other colors. Thus, by measuring the perceptual sensitivity to cued colors, this experiment directly tested whether the representations of to-be-selected and to-be-ignored features are boosted or attenuated.
Method
Participants
A power analysis on a pilot version of Experiment 2a with positive cues indicated that detecting a significant effect with 80% power at an alpha level of .05 would require collecting data from 55 participants. To evaluate the strength of evidence for a small or potentially null effect of negative cues on color detection, we preregistered recruitment of 100 participants who passed our exclusion criteria in each of Experiments 2a and 2b. We excluded data from 72 participants in Experiment 2a (16 for taking longer than 90 minutes to complete the experiment; 53 for search accuracy below 80% in any block; three for overall detection sensitivity below 0.5 d′). We excluded data from 99 participants in Experiment 2b (15 for taking longer than 90 minutes to complete the experiment; 84 for search accuracy below 80% in any block). After exclusion, Experiment 2a included 66 women, 28 men, five nonbinary people and one participant who declined to report their gender, with a mean age of 20.4 (range: 18–34 age); Experiment 2b included 71 women, 26 men, and one nonbinary person and two people who declined to report their genders, with a mean age of 20.2 years (range: 18–36 years).
Stimuli
Stimuli were like those in Experiment 1, with the following exceptions. The fixation point was a black dot rather than a cross. To ensure the consistent use of color cues, each search trial contained two potential search targets with rightward or leftward gaps (one matching the cue color; gap locations of the two potential targets were random and determined independently of each other), and six distractors with gaps on their top or bottom, with four items in each of two colors.
Color probes were rings 90 pixels in diameter and an eight-pixel line thickness. Probes could be any of the four search item colors or a gray approximately equiluminant to the other colors. The response screen on color detection trials asked participants, “Was the probe a non-gray color?” with the options “Press ‘left’ for yes (it flashed a color besides gray)” and “Press ‘right’ for no (it flashed gray).” Note that we never asked participants which color they saw, but simply whether they detected a color at all, avoiding non-perceptual biases to report the cued color.
Procedure
Participants completed intermixed visual search and color detection trials. First, participants completed thirty practice trials. They then completed six 88-trial experimental blocks, each of which randomly intermixed 72 search trials and 16 detection trials. Participants did not know the upcoming trial type.
On visual search trials, the cue was presented for 600 ms, followed by a 663 ms blank screen, then the search array until response. The cue color indicated the target color with 100% accuracy (in Exp. 2a; nontarget color in Exp. 2b), and two potential targets (with rightward or leftward gaps) were present on each trial. One of these potential targets was in the cued color and one was in a random one of the other three colors. Rather than simply responding with the orientation of a target as in Experiment 1, participants were now asked to respond with the target orientation of either the potential target in the cue color (Exp. 2a) or not in the cue color (Exp. 2b). Thus, accurate search performance required use of the cue. In other aspects the visual search task was identical to that of Experiment 1.
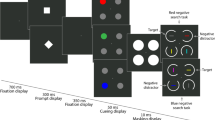
On color detection trials, the cue was presented for 600 ms, followed by a blank screen for a random duration between 300 and 400 ms, followed by a color detection probe that appeared randomly at one of the search item locations. This probe was presented for only about 30 ms in the main experiment, but for between 50 and 80 ms during practice to familiarize participants with the task. This was followed by a blank screen for between 233 and 333 ms, with its duration selected based on the cue-to-probe interval duration for a total time between cue offset and response screen of 663 ms. Probes were gray on half of trials and a random one of the four search colors on the other half of trials. Counterbalancing ensured that the cue color did not predict probe color, resulting in eight gray probes, two probes in the cue color (congruent probes), and six probes in uncued colors in each block (incongruent probes). Participants were instructed to respond whether the probe was gray (right arrow press) or any color (left arrow press). Participants were told to emphasize accuracy during color detection trials and were given accuracy feedback after each trial. See Fig. 3 for a trial schematic.

Experiment 2’s trial structure. On visual search trials, a brief cue indicated the upcoming target (in Exp. 2a) or nontarget (Exp. 2b) color. Each search array displayed 8 items in two colors, with two potential targets, one in each color. Participants judged the gap location of either the potential target that matched the cue color (Exp. 2a) or the one that did not match the cue color (Exp. 2b). On color detection trials, the same cue was presented, followed by a blank display and then a color detection probe at a random one of the search item locations. Half of the probes were gray, while the other half were equally likely to occur in any of the four search item colors. This yielded three probe conditions: congruent, in which probes matched the search cue color; incongruent, in which probes matched an uncued potential search color; and neutral gray probes. Participants judged whether probes were gray or any other color
Analysis
Analysis focused on the color detection task. We used dependent-samples t-tests to compare the accuracy for hue detection on congruent and incongruent trials, separately for Experiments 2a and 2b. We also compared the difference between congruent and incongruent accuracy across experiments using an independent-samples t-test and a two-sample Bayesian test. Exclusion criteria were the same as in Experiment 1; we also excluded participants with overall sensitivity (d′) lower than 0.5 in the probe task, indicating near or below chance performance.
Results
Experiment 2a: Positive cues
Positive cues induced greater sensitivity for probes in the cued color than uncued colors, consistent with evidence for proactive enhancement. In particular, color detection accuracy was 4.5% higher for probes congruent with the to-be-attended cue color than for incongruent probes (95% CI [2.3%, 6.7%]), t(99) = 3.97, p < .001, d = 0.39. Overall search accuracy (M = 96.0%, 95% CI [95.5, 96.5]) and RT (M = 815 ms, 95% CI [797, 834]) suggested that participants who passed the exclusion criteria performed well in the search task.
Experiment 2b: Negative cues
Unlike like positive cues, negative cues did not significantly change sensitivity for the cued color. Color detection accuracy was 2.1% higher for congruent probes (those matching the to-be-ignored cue color) than for incongruent probes (95% CI [0.0, 4.6]), but this difference was not statistically reliable, t(99) = 1.64, p = .104, d = 0.16. Overall search accuracy (M = 94.7%, 95% CI [94.0, 95.3]) and RT (M = 985 ms, 95% CI [959, 1011]) were again indicative of good performance in the main task.
Comparing positive and negative cues
Cross-experiment analyses did not provide conclusive evidence on whether the increased detection rate for the cued color in Experiments 2a and 2b were statistically comparable. An independent-samples t test showed that the mean accuracy advantage for congruent probes was not significantly higher for positive cues than negative cues (mean accuracy advantage was 2.4% greater for positive cues, 95% CI [−1.0, 5.8]), t(198) = 1.42, p = 0.160, d = 0.17. To assess evidence for the null effect, we ran an unplanned post hoc Bayesian two-sample test using a default prior of .707 (Morey et al., 2015; Rouder et al., 2009). This test yielded a small Bayes factor of 2.53 in favor of the null hypothesis that congruency effects on color detection were equivalent for positive and negative cues.
Discussion
Overall, Experiment 2 suggests that cues providing information about upcoming target colors yield proactive enhancement of the cued colors. In contrast, cues about upcoming nontarget colors yield no proactive suppression of cued colors, instead yielding small to negligible effects of proactive enhancement. Given the inconclusive data regarding whether negative cues elicit no proactive effects or proactive enhancement of the cued color, we conducted a near replication of Experiment 2 (see Supplemental Materials), using colors more similar to each other in CIElab space. Data from this replication again showed no proactive suppression of negatively cued colors, in fact showing significant proactive enhancement. Taken together, data from Experiment 2 and our supplemental replication provide strong evidence for a lack of proactive suppression, and some evidence for proactive enhancement, of negatively cued colors.
It is worth noting that considerably more participants were excluded for low search accuracy in the negative cueing (84) compared with the positive cueing experiment (53). It is likely that this reflects overall greater difficulty in using negative than positive cues. While the general difficulty of using negative cues is itself interesting, this is not a novel observation (e.g., Cunningham & Egeth, 2016). As our research question was not about whether negative cueing could benefit search, but how this occurs when it does, our strict exclusion criteria is crucial for ensuring that our included participants are using cues as intended.
General discussion
Across two experiments, we demonstrate that search benefits from cueing upcoming nontarget features does not occur due to proactive suppression of the cued feature. In Experiment 1, both 75% valid positive and negative cues benefited search RT, replicating past reports that positive cues speed search more than negative cues (e.g., Arita et al., 2012). Experiment 2 (and a near replication; see Supplemental Materials) demonstrated that positive cues increased sensitivity to the cued color relative to uncued colors, whereas negative cues elicited no proactive suppression and perhaps small effects of proactive enhancement. These results indicate that participants did not proactively suppress a color to subsequently ignore it, at least under accounts in which suppression operates analogously to attentional enhancement (for instance, by decreasing gain of suppressed feature channels; Störmer & Alvarez, 2014). Rather, these results are consistent with reactive accounts of distractor ignoring, including purely reactive accounts claiming that suppression begins after display onset (see Geng, 2014; Geng et al., 2019, for reviews), the search-and-destroy hypothesis (Moher & Egeth, 2012), and translation accounts in which knowledge about nontarget features is used to inform which target feature should be enhanced during search (Becker et al., 2015).
This study provides clear evidence against proactive suppression of negatively cued features. In our view, the reactive processes involved in ignoring features during search may underlie the smaller RT benefits from negative compared with positive cues, as the time taken to implement feature-based attention occurs during search itself, reducing the potential RT benefit relative to if these processes occur proactively. Furthermore, because our results diverge from other research in which data from similar search tasks have been taken as evidence for active suppression (Arita et al., 2012; Chang & Egeth, 2019; Zhang et al., 2020), they suggest that data from search tasks themselves—including ones probing perceptual sensitivity shortly after search array onset—should not be taken as evidence for proactive suppression. Rather, proactive suppression should be measured only prior to the onset of search stimuli using single-item displays that do not confound spatial and feature-based attention.
Our results do not imply that proactive suppression cannot occur. People may strategically suppress features in other contexts. However, we did use several strategies to maximize the likelihood of participants using proactive suppression. First, the task necessitated using the cue to identify which of two potential targets to identify, meaning participants could not simply ignore the cue if they found it difficult to use. Second, we used a relatively long delay between cue and probe onset (900–1,000 ms) to allow proactive suppression to emerge. Thus, if voluntary proactive suppression is possible, its absence here likely means it is used in only a narrow range of circumstances.
Broadly, the present results suggest that ignoring cued visual features depends on reactive mechanisms, which contrasts with the proactive effects of attending to upcoming target features. Our results (especially the Supplemental Experiment) also provide some preliminary evidence that these reactive effects may sometimes rely on proactive enhancement of the negatively cued feature, suggesting that reactive ignoring may rely on some of the same feature-based attention mechanisms used in attending to target features—only deployed in strategic ways to ignore the negatively cued item during search. While the extent of this proactive enhancement of negatively cued features is not yet clear, our results provide a clear lack of evidence for proactive suppression. This may explain why ignoring distractors may generally be less efficient than selecting targets.
References
Arita, J. T., Carlisle, N. B., & Woodman, G. F. (2012). Templates for rejection: Configuring attention to ignore task-irrelevant features. Journal of Experimental Psychology. Human Perception and Performance, 38(3), 580–584. https://doi.org/10.1037/a0027885
Beck, V. M., Luck, S. J., & Hollingworth, A. (2017). Whatever you do, don’t look at the . . .: Evaluating guidance by an exclusionary attentional template. Journal of Experimental Psychology. Human Perception and Performance, 44(4), 645–662. https://doi.org/10.1037/xhp0000485
Becker, M. W., Hemsteger, S., & Peltier, C. (2015). No templates for rejection: A failure to configure attention to ignore task-irrelevant features. Visual Cognition, 23(9/10), 1150–1167. https://doi.org/10.1080/13506285.2016.1149532
Chang, S., & Egeth, H. E. (2019). Enhancement and suppression flexibly guide attention. Psychological Science, 30(12), 1724–1732. https://doi.org/10.1177/0956797619878813
Chapman, A. F., & Störmer, V. S. (2021). Feature-based attention is not confined by object boundaries: Spatially global enhancement of irrelevant features. Psychonomic Bulletin & Review. https://doi.org/10.3758/s13423-021-01897-x
Cunningham, C. A., & Egeth, H. E. (2016). Taming the white bear: Initial costs and eventual benefits of distractor inhibition. Psychological Science, 27(4), 476–485. https://doi.org/10.1177/0956797615626564
de Vries, I. E. J., Savran, E., van Driel, J., & Olivers, C. N. L. (2019). Oscillatory mechanisms of preparing for visual distraction. Journal of Cognitive Neuroscience, 31(12), 1873–1894. https://doi.org/10.1162/jocn_a_01460
Gaspelin, N., Gaspar, J. M., & Luck, S. J. (2019). Oculomotor inhibition of salient distractors: Voluntary inhibition cannot override selection history. Visual Cognition, 1–27. https://doi.org/10.1080/13506285.2019.1600090
Geng, J. J. (2014). Attentional mechanisms of distractor suppression. Current Directions in Psychological Science, 23(2), 147–153. https://doi.org/10.1177/0963721414525780
Geng, J. J., Won, B.-Y., & Carlisle, N. B. (2019). Distractor ignoring: Strategies, learning, and passive filtering. Current Directions in Psychological Science, 28(6), 600–606. https://doi.org/10.1177/0963721419867099
Giesbrecht, B., Weissman, D. H., Woldorff, M. G., & Mangun, G. R. (2006). Pre-target activity in visual cortex predicts behavioral performance on spatial and feature attention tasks. Brain Research, 1080(1), 63–72. https://doi.org/10.1016/j.brainres.2005.09.068
Ho, T. C., Brown, S., Abuyo, N. A., Ku, E.-H. J., & Serences, J. T. (2012). Perceptual consequences of feature-based attentional enhancement and suppression. Journal of Vision, 12(8), 15. https://doi.org/10.1167/12.8.15
Kassambara, A. (2020). Rstatix: Pipe-friendly framework for basic statistical tests (R Package Version 0.6.0) [Computer software]. https://cran.r-project.org/web/packages/rstatix/index.html. Accessed 6 Jan 2022.
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., & Ungerleider, L. G. (1999). Increased activity in human visual cortex during directed attention in the absence of visual stimulation. Neuron, 22(4), 751–761. https://doi.org/10.1016/S0896-6273(00)80734-5
Martinez-Trujillo, J. C., & Treue, S. (2004). Feature-based attention increases the selectivity of population responses in primate visual cortex. Current Biology, 14(9), 744–751. https://doi.org/10.1016/j.cub.2004.04.028
Mazaheri, A., DiQuattro, N. E., Bengson, J., & Geng, J. J. (2011). Pre-stimulus activity predicts the winner of top-down vs. bottom-up attentional selection. PLOS ONE, 6(2), Article e16243. https://doi.org/10.1371/journal.pone.0016243
Moher, J., & Egeth, H. E. (2012). The ignoring paradox: Cueing distractor features leads first to selection, then to inhibition of to-be-ignored items. Attention, Perception, & Psychophysics, 74(8), 1590–1605. https://doi.org/10.3758/s13414-012-0358-0
Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 1(1), 42–45. https://doi.org/10.20982/tqmp.04.2.p061
Morey, R., Rouder, J. N., Jamil, T., Urbanek, S., Forner, K., & Ly, A. (2015). Package “BayesFactor” (Version 0.9.12-4.2) [Computer software]. https://cran.r-project.org/web/packages/BayesFactor/index.html. Accessed 6 Jan 2022.
Reeder, R. R., Olivers, C. N. L., & Pollmann, S. (2017). Cortical evidence for negative search templates. Visual Cognition, 25(1/3), 278–290. https://doi.org/10.1080/13506285.2017.1339755
Reeder, R. R., Olivers, C. N. L., Hanke, M., & Pollmann, S. (2018). No evidence for enhanced distractor template representation in early visual cortex. Cortex, 108, 279–282. https://doi.org/10.1016/j.cortex.2018.08.005
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson, G. (2009). Bayesian t tests for accepting and rejecting the null hypothesis. Psychonomic Bulletin & Review, 16(2), 225–237. https://doi.org/10.3758/PBR.16.2.225
Störmer, V. S., & Alvarez, G. A. (2014). Feature-based attention elicits surround suppression in feature space. Current Biology, 24(17), 1985–1988. https://doi.org/10.1016/j.cub.2014.07.030
Suchow, J. W., Brady, T. F., Fougnie, D., & Alvarez, G. A. (2013). Modeling visual working memory with the MemToolbox. Journal of Vision, 13(10). https://doi.org/10.1167/13.10.9
Zhang, Z., Gapelin, N., & Carlisle, N. B. (2020). Probing early attention following negative and positive templates. Attention, Perception, & Psychophysics, 82, 1166–1175. https://doi.org/10.3758/s13414-019-01864-8
Acknowledgements
This research was supported in part by National Science Foundation Grant BCS-1850738 to V.S.S. Thanks to Tim Brady for discussions leading to Experiment 2 and to Angus Chapman for helpful comments.
Author information
Authors and Affiliations
Contributions
D.A.A. and V.S.S. developed the study concept and design. D.A.A. programmed the experiments and was responsible for data collection, analysis, and visualization. D.A.A. drafted the manuscript and V.S.S. provided critical revisions. All authors approved the final version of the manuscript for submission.
Corresponding author
Additional information
Open practices statement
Experiments 1 and 2 were not preregistered but followed the same preregistered exclusion criteria as Supplemental Experiment 1, which was formally preregistered at AsPredicted prior to data collection and can be viewed (https://aspredicted.org/mm4u4.pdf). Raw data, experimental code, and analysis code for all experiments are available on the Open Science Framework (https://osf.io/mb683/).
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 105 kb)
Rights and permissions
About this article

Cite this article
Addleman, D.A., Störmer, V.S. No evidence for proactive suppression of explicitly cued distractor features. Psychon Bull Rev 29, 1338–1346 (2022). https://doi.org/10.3758/s13423-022-02071-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-022-02071-7