
Abstract
Our search performance is strongly influenced by our past experience. In the lab, this influence has been demonstrated by investigating a variety of phenomena, including intertrial priming, statistical learning, and reward history, and collectively referred to as selection history. The resulting findings have led researchers to claim that selection history guides attention, thereby challenging the prevailing dichotomy, according to which top-down and bottom-up factors alone determine attentional priority. Here, we re-examine this claim with regard to one selection-history phenomenon, feature intertrial priming (aka priming of pop-out). We evaluate the evidence that specifically pertains to the role of feature intertrial priming in attentional guidance, rather than in later selective processes occurring after the target is found. We distinguish between the main experimental rationales, while considering the extent to which feature intertrial priming, as studied through different protocols, shares characteristics of top-down attention. We show that there is strong evidence that feature intertrial priming guides attention when the experimental protocol departs from the canonical paradigm and encourages observers to maintain the critical feature in visual working memory or to form expectations about the upcoming target. By contrast, the current evidence regarding the standard feature intertrial priming phenomenon is inconclusive. We propose directions for future research and suggest that applying the methodology used here in order to re-evaluate of the role of other selection history phenomena in attentional guidance should clarify the mechanisms underlying the strong impact of past experience on visual search performance.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In a very influential study, Maljkovic and Nakayama (1994) reported a counter-intuitive observation. They asked their subjects to perform a simple task: to search for a uniquely colored object (or color singleton) that could be either the only red object among objects that were all green, or the only green object among objects that were all red (see Fig. 1). This task is so easy that the target is said to pop out, that is, it is immediately spotted, no matter how many distractors surround it. In addition, the target’s actual color should not matter, because it suffices to search for the most salient object, the odd-one-out, in order to find it. Yet, the authors found responses to be considerably faster when the target happened to have the same color on consecutive trials than when its color changed.Footnote 1

Sample sequence of trials in a typical feature intertrial priming experiment (Maljkovic & Nakayama, 1994). The target is the diamond with the unique color
This effect, which was called priming of pop-out (or PoP), has been extensively replicated (see, e.g., Kristjánsson & Campana, 2010, and Lamy & Kristjánsson, 2013, for reviews). It was extended to target singletons differing from distractors on a large variety of dimensions such as shape (e.g., Lamy et al., 2006b; Pinto et al., 2005), orientation (e.g., Hillstrom, 2000; Lamy et al., 2013), brightness (e.g., Becker, 2008a, 2008b), size (Huang et al., 2004; Kristjánsson, 2006) and facial expressions of emotion (Amunts et al., 2014; Lamy et al., 2008a), as well as to conjunction search (e.g., Hillstrom, 2000). As this effect is not confined to pop-out tasks, we refer to it as “feature intertrial priming” (henceforth, feature ITP) in the remainder of this paper.
Since then, a myriad of additional effects of memory on visual search have been documented and are typically referred to as “selection history” (e.g., Brascamp et al., 2011) or “attentional priming” effects (e.g., Kristjánsson & Ásgeirsson, 2019). These include other intertrial priming effects, such as dimension priming (e.g., Found & Müller, 1996; Olivers & Humphreys, 2003), the distractor preview effect (e.g., Ariga & Kawahara, 2004; Wan & Lleras, 2010), location priming (Geyer & Müller, 2009; Maljkovic & Nakayama, 1996), and singleton priming (Lamy et al., 2006a; Lamy et al., 2008c; Won et al., 2019), statistical learning effects, such as contextual cueing (Chun & Jiang, 1998; Sisk et al., 2019) and probability cueing (e.g., Ferrante et al., 2018; Geng & Behrmann, 2005), and reward history effects (Della Libera & Chelazzi, 2006; Failing & Theeuwes, 2018).
A challenge to the bottom-up versus top-down dichotomy in attentional guidance
The recent surge of research showing effects of memory on visual search has promoted the view that selection history plays an important role in biasing the competition for attention (e.g., Awh et al., 2012; Belopolsky, 2015; Theeuwes, 2013; see also Anderson, 2016; Chelazzi et al., 2013; Goldstein & Beck, 2018; Jiang, 2018; Luck et al., 2020; Theeuwes, 2018; Todd & Manaligod, 2018; Wolfe, 2019, 2021). In its most extreme form, this view puts forward selection history as an alternative account to most of the effects traditionally attributed to top-down control of attentional priority (e.g., Belopolsky et al., 2010; Theeuwes, 2018). In its milder form, which is the focus of the present review, it proposes that a common spatial map codes attentional priorities and integrates signals related not only to physical salience and current goals, but also to selection history; the competition on this map determines in a winner-take-all fashion which object is selected at any given time. In other words, this view suggest that models of attention should incorporate selection history as an additional source of attentional guidance (e.g., Awh et al., 2012; Luck et al., 2020)– a suggestion that has been widely and quickly embraced by the field, including by visual search models most identified with the notion that attentional priority is entirely determined by stimulus salience and current goals (e.g., Guided Search; Wolfe & Horowitz, 2017; Wolfe, 2021).
However, before we seal the fate of this “failed dichotomy” (Awh et al., 2012), it is important to review the evidence that specifically demonstrates that selection history influences attentional guidance, that is, the relative priority weights assigned to different objects in the visual field. The question arises because, as was the case in Maljkovic and Nakayama’s (1994) seminal experiments, selection history effects often manifest as faster search performance on a given trial when some aspect of the search array repeats from a previous trial or matches a learned regularity (e.g., Chun & Jiang, 2003; Druker & Anderson, 2010; Maljkovic & Nakayama, 1996; Sha & Jiang, 2016). Yet, faster performance does not necessarily indicate that the target is found earlier, and may instead denote that the target is processed faster after it is found.
Objective of the present review
In the present review, we do not question the widely endorsed idea (e.g., Awh et al., 2012; Luck et al., 2020; Wolfe, 2021) that selection history affects selective processes such as processing of the target after it was located (aka attentional engagement, e.g., Folk et al., 2009; Zivony & Lamy, 2016) or reactive attentional control (i.e., rapid disengagement of attention from an irrelevant distractor, e.g., Luck et al., 2020). Instead, we evaluate the evidence that specifically addresses whether selection history modulates attentional priority, that is, whether it biases the competition, the resolution of which determines which item is selected first.
As selection history refers to a heterogenous collection of phenomena, there is no a priori reason to assume that the phenomena currently placed under the selection-history umbrella either all affect attentional guidance or do not. Here, we focus on one selection history phenomenon, feature ITP (aka PoP; Maljkovic & Nakayama, 1994), because it has been the most intensively studied, but the methodology used here can readily be applied to other instances of selection history.
We first summarize the different accounts of the mechanisms underlying feature ITP. We show that while these accounts have mainly contrasted perceptual and post-selective views, there has been a less structured effort to distinguish attentional guidance from later perceptual/selective processes – a state of affairs that prevails also for other selection history phenomena (see, e.g., Sisk et al., 2019, for contextual cueing). Then, we discuss the differences and similarities that exist between feature ITP and top-down attention, and classify the different experimental protocols used to investigate feature ITP according to which characteristics, if any, the critical manipulation shares characteristics of top-down attention. Then, taking this classification into account, we review the relevant evidence, separately for different measures of attentional priority. We conclude by re-evaluating the current consensus on the status of feature ITP as a source of attentional guidance and by suggesting guidelines for further research.
Theoretical accounts of feature intertrial priming (ITP)
Since Maljkovic and Nakayama’s (1994) seminal study, researchers have actively debated the mechanisms underlying feature ITP. The models that have emerged from this debate fall into three broad categories, often referred to as the independent feature-weighting, episodic retrieval, and hybrid accounts (for recent reviews, see Kruijne & Meeter, 2015; Kristjánsson & Ásgeirsson, 2019; Thomson & Milliken, 2013).
The independent feature-weighting account is based on the ideas initially developed by Maljkovic and Nakayama (1994, 1996) to account for feature ITP. During a search event, the features associated with the target become more activated, while the features associated with the distractors are suppressed. These activations persist for some time and then decay. The priority of the locations that share these features on subsequent trials is modified accordingly (e.g. Kristjansson, 2006; Becker & Horstmann, 2009; Chun & Nakayama, 2000; Lee et al., 2009; Maljkovic & Martini, 2005; Martini, 2010; Theeuwes, 2018; Wolfe et al., 2003). Thus, according to the feature-weighting view, an object sharing a previous target’s feature is more likely to be selected, just as if it had become physically more salient, and conversely, an object sharing a previous distractor’s feature is less likely to be selected, just as if it had become less salient.
By contrast, the episodic retrieval model posits that feature ITP reflects mechanisms that occur after the target is found. Multiple aspects of each search trial are stored as bound episodic memory traces. On a new trial, these memory traces are automatically retrieved; they speed performance if they match the features of the current search episodes and impair performance otherwise. One of the main findings supporting this view is that the effects of repeating different features of the target interact. In particular, repeating the response feature speeds performance when the target-defining feature repeats but slows performance when this feature changes from the previous trial (e.g., Hillstrom, 2000; Huang et al., 2004; Lamy et al., 2010; Lamy, Zivony & Yashar, 2011; for additional evidence supporting the episodic retrieval account, see Thomson & Milliken, 2013).
Unlike feature-weighting and episodic-retrieval accounts, hybrid models of feature ITP suggest that it may occur at several stages during search. For instance, Meeter and Olivers (2006; see also Hickey, Olivers, Meeter, & Theeuwes, 2011; Olivers & Meeter, 2006, 2008; Olivers & Hickey, 2010) suggested an interpretation of feature ITP, known as the ambiguity account. They argued that the more ambiguous the task is, the larger the benefit of repetitions from previous trials, and that such ambiguity may arise at different levels. Accordingly, they suggested that “if it is ambiguous what the target is, visual selection will rely relatively more on what was selected in previous trials. If it is ambiguous what the response should be, response selection will rely more heavily on what response was coupled with a stimulus on previous trials” (Meeter & Olivers, 2006). In support for these claims, they showed that increasing perceptual ambiguity (e.g., by introducing a salient distractor in the search displays) and increasing response-related ambiguity (e.g., by alternating the response requirements) both resulted in larger priming effects.
Lamy et al. (2010) suggested a dual-stage account of feature ITP that also offers a compromise between the independent feature-weighting and episodic retrieval accounts. They tracked the time course of the interaction between feature ITP and response repetition that is the hallmark of episodic retrieval accounts. They reported a robust feature ITP effect at early stages of processing, before response-related processes could kick in (within 100 ms from search display onset), and an interaction between pop-out feature repetition and response repetition later on (after 200–400 ms). Moreover, later studies showed that this interaction results from repetition of the motor response, and not from repetition of the response feature (Yashar et al., 2013; Yashar & Lamy, 2011)– a finding that situates retrieval processes at play in feature ITP at a late stage. The authors concluded that feature ITP speeds both a perceptual stage and a response-related stage of visual search.
Processing stages in a typical feature ITP experiment
The brief foregoing review suggests that feature ITP operates at both early and late stages during visual search. However, further scrutiny of the literature reveals that there is quite some variance as to what these stages stand for. By integrating the different suggestions raised with regard to how feature ITP speeds performance in a typical feature ITP experiment, one can distinguish between four candidate stages, as illustrated in Fig. 2: (1) When a search display comes on, the basic features present in the display are analyzed in parallel and attentional priority weights are assigned to each location/object(attentional priority stage, e.g., Maljkovic & Nakayama, 1994). (2) The object with the highest priority weight is selected,Footnote 2 that is, its features are processed (selection stage, e.g., Yashar & Lamy, 2010a, b). (3) Then, prior to responding, the current search episode is compared with previous episodes (episodic retrieval stage, e.g., Hillstrom, 2000; Huang et al., 2004; Thomson & Milliken, 2012). (4) Finally, the appropriate response is selected and executed (response-selection stage; e.g., Yashar & Lamy, 2011).

Schematic illustration of the processing stages that may be modulated by feature intertrial priming (ITP). In the present review, we focus exclusively on the evidence pertaining to the effects of feature ITP on attentional guidance
To determine whether selection history in general, and feature ITP in particular, should be combined with current goals and physical salience to shape an integrated priority map, as suggested by many authors (e.g., Awh et al., 2012; Luck et al., 2020; Wolfe, 2021), it is not enough to evaluate the evidence from studies that tested the independent feature-weighting, episodic retrieval, and hybrid accounts against each other. Instead, a more focused review is necessary, which examines the findings from studies that used experimental strategies specifically suited to investigate whether feature ITP affects the attentional priority stage and guides attention, as do bottom-up and top-down factors. This was the goal of the present paper.
Feature ITP and top-down guidance of attention
The claim that the classical dichotomy between top-down and bottom-up factors should be updated to include selection history critically hinges on the premise that selection history can be distinguished from bottom-up and top-down sources of attentional guidance. Feature ITP is, by definition, not a stimulus-driven bias, as it results from prior experience. The distinction between inter-trial priming and top-down attention is more controversial. Some authors argue that selecting information previously associated with successful search establishes a default top-down bias towards the same information, even if it is currently irrelevant (e.g., Egeth, 2018; Egeth et al., 2010; Wolfe et al., 2003). Others suggest that feature ITP is unrelated to top-down attention because observers do not purposefully attend to the repeated feature (Theeuwes, 2018). Although debating this issue could appear as mere semantic quibbling, it may be informative to examine the extent to which feature ITP, as studied through different experimental protocols, shares characteristics of top-down attention. Such inspection can inform us on whether different feature ITP manipulations have qualitatively different effects on attention, that is, influence different processes. More broadly, it should also help us delineate the boundary conditions of attentional guidance by feature ITP.
Characteristics of top-down guidance of attention
When discussing whether selection history differs from top-down attention, three characteristics of top-down attention most often emerge: awareness, flexibility, and goal-directedness. Top-down guidance is thought to be explicit and voluntary because when searching for an object defined by a given property, observers are typically aware of this property and purposefully establish it as their search goal. Top-down guidance is also thought to be flexible: when directing one’s attention to a given feature, one can cease to prioritize that feature if it no longer serves current goals. Finally, and most importantly for the purposes of this review, top-down guidance is thought to be goal-directed: observers use some information because it serves a goal, typically because it is necessary or at least useful for performing an upcoming task. In this case, holding a representation of the task goal in working memory (WM) is thought to be the mechanism that mediates guidance of attention (e.g., Desimone & Duncan, 1995; Duncan & Humphreys, 1989; Oberauer, 2002; Olivers et al., 2011; see also Gao & Theeuwes, 2020). Accordingly, when a feature is held in WM for a memory test, attention is biased towards items matching this feature in an unrelated search task that intervenes during the retention interval, even if it is detrimental to the task at hand (e.g., Downing, 2000; Pashler & Shiu, 1999; Olivers et al., 2006; Soto et al., 2005; see also Li et al., 2020). Likewise, contingent capture (e.g., Folk et al., 1992), which is a hallmark of goal-directed attention, refers to involuntary allocation of attention to a distractor that shares the target-defining feature held in WM.
Feature ITP experimental protocols and top-down guidance
In a typical ITP experiment (e.g., Maljkovic & Nakayama, 1994) a repetition is no more likely to occur than a change, such that searching for the feature that previously characterized the target does not serve any objective goal. In addition, as a feature repetition from as far as five to eight trials back exerts an influence, it is unlikely that the target color on previous trials is maintained in WM or that conscious traces of the target feature, so far back, drive the effect (see Maljkovic & Nakayama, 2000). Whether feature priming is flexible, however, remains controversial, with mixed findings as to whether or not it is influenced by knowledge of the probability of a target feature change (Maljkovic & Nakayama, 1994 vs. Cochrane & Pratt, 2020; see also Shurygina et al., 2019).
Importantly, however, feature ITP has been investigated via additional paradigms, in which it seems reasonable to claim that guidance of attention shares the goal-directed aspect of top-down attention, at least to some extent. Two main departures from the typical paradigm fall into this category.
In a number of studies, the task did not entail monitoring displays for a feature discontinuity (e.g., Bravo & Nakayama, 1992; Maljkovic & Nakayama, 1994) but instead required participants to store a template of the target-defining feature in WM (e.g., Ásgeirsson & Kristjánsson, 2019; Becker & Horstmann, 2009; Eimer & Kiss, 2010, Exp. 2; Folk & Remington, 2008, Exp. 2; Hillstrom, 2000, Exp. 4; Irons et al., 2012, Exp. 2; Koshino, 2001; Kristjánsson et al., 2002; Meeter & Olivers, 2006, Exp. 3; Olivers & Hickey, 2010; Olivers & Meeter, 2012; Schoeberl, Goller, & Ansorge, 2019a, b, Exp. 1; Theeuwes & van der Burg, 2011). For instance, Theeuwes and van der Burg (2011, Exp. 4) used a variant of the additional singleton paradigm (Theeuwes, 1991), in which a precue indicated the color of the upcoming target on each trial. The search display that followed contained two color singletons: one singleton (the target) shared the precue color, while the other singleton (the distractor) had a different color. The salient distractor interfered with search more when the target color changed on successive trials than when it repeated (see Olivers & Meeter, 2006, for related evidence). The authors concluded that repeating the target, which they labeled “automatic bottom-up intertrial priming,” biased the competition in favor of the repeated singleton. However, if one agrees that maintaining a feature in working memory is the mechanism that mediates goal-directed attention (e.g., Gao & Theeuwes, 2020), such findings describe a feature ITP phenomenon that is related to the search goals maintained in WM: they show that using the same feature template on successive trials increases the competitive edge of objects sharing that feature.Footnote 3
In line with our argument, note that this form of feature ITP has been dissociated from the classical feature ITP: Leonard and Egeth (2008) showed that feature ITP was independent of target salience when an informative cue indicated the upcoming target feature (i.e., when the target feature was stored in WM), whereas it was strongly modulated by target salience when the target feature on any given trial was unknown (as in the classical feature ITP paradigm).
The second departure from the typical paradigm worth mentioning is when a feature repetition is more likely than a feature change across the experiment (e.g., Bichot & Schall, 2002; Feldmann-Wüstefeld & Schubö, 2016; Gaspelin et al., 2019, Exp. 3; Shurygina et al., 2019; Sigurdardottir et al., 2008; Westerberg et al., 2020). In that case, prioritizing the previously selected feature is objectively beneficial for performance, and feature ITP therefore shares the goal-related aspect of top-down attention (irrespective of whether observers are aware of the critical contingency, which was never assessed in these studies).
Based on the above considerations, it seems important to examine the possibility that feature ITP may be more likely to guide attention when it shares characteristics of goal-directed attention than when it does not. Therefore, in the review that follows, we single out the studies in which the task required or encouraged participants to maintain in WM the feature for which feature ITP was measured, or in which feature repetitions were more likely than feature changes. For the purpose of the present review, we refer to both these phenomena as “goal-related feature ITP,” which we contrast with “basic feature ITP.”
Does feature ITP affect attentional guidance? Review of the evidence
Several different experimental rationales have been employed to investigate whether feature ITP modulates the attentional priority of a given object in the visual field. For each of these, we first explain why it is thought to provide a diagnostic measure of attentional guidance and then review the studies in which it was used in the context of feature ITP. Note that, as we focus exclusively on whether feature ITP modulates attentional priority, we do not review studies that address whether feature ITP affects perceptual processes (vs. post-perceptual processes) when their design does not allow one to distinguish between attentional guidance and later perceptual mechanisms (i.e., selection). In addition, our inclusion criterion was whether a given experiment included a manipulation that could isolate effects of feature ITP on attentional priority – even if it was not the authors’ objective.
We first review studies that relied only on manual responses and then turn to describe studies that relied on eye movements, event-related potentials, and single-cell recordings,Footnote 4 in separate sections.
Feature ITP and search slopes
A popular method to study attentional guidance during visual search is to vary the number of distractors presented together with the target and measure the time it takes to respond to the target as a function of the number of distractors (e.g., Wolfe, 2016). The slope of the response time (RT) × set size function is used as a measure of the search efficiency. When performance deteriorates as more distractors are added (positive slopes), shallower search slopes are taken to indicate that fewer distractors are attended before the target is found (e.g., Wolfe & Horowitz, 2017). Accordingly, many previous studies have shown that both bottom-up and top-down factors modulate search slopes (e.g., Cave & Wolfe, 1990; Duncan & Humphreys, 1989; Treisman & Sato, 1990; but for criticisms of search slopes as a measure of attention, see Christie et al., 2014 and Kristjánsson, 2015, and for responses to some of these, see Wolfe, 2016), in line with the idea that these factors guide attention. However, performance may also improve as set size increases. This occurs when the target feature is unpredictable and search is guided by the target’s salience (e.g., Bravo & Nakayama, 1992): in that case, adding distractors increases target salience and facilitates search, which manifests in negative search slopes.
Feature PoP and positive search slopes
If feature ITP modulates attentional priority, positive search slopes should be shallower when the target and distractors repeat from the previous trial than when they don’t. However, this finding is open to alternative interpretations: for instance, the target and distractors priorities may not change at all as a function of feature repetition, but instead, flatter search slopes may occur because it is easier to disengage attention from a distractor that has a previously ignored feature. Negative evidence is less ambiguous when considering the influence of feature ITP on search slopes: failure to observe shallower search slopes when features repeat would argue against the priority-based account. As we describe next, most studies showed that feature ITP does not reduce positive search slopes.
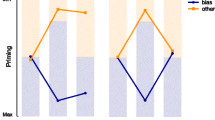
Three studies used tasks in which the target was defined by a unique feature that randomly repeated or changed across trials, as in Maljkovic and Nakayama’s (1994) study. All of them showed a robust feature ITP effect and generated positive search slopes (Amunts et al., 2014; Ásgeirsson & Kristjánsson, 2011; Becker & Ansorge, 2013, Exp.2). However, none of them showed a reliable effect of feature ITP on search slopes (see Figure 3 for an example).

Sample display (left panel) and mean reaction times on target-present trials preceded by a target-present trial in the upright-face condition as a function of set size, target emotion and emotion repetition (right panel) in Amunts et al. (2014, Exp.1). Participants had to detect the presence of a face that differed from all others, unpredictably either a neutral face among angry faces (as depicted here) or an angry face among neutral faces. Repeating the target and distractors emotion speeded performance when the target was angry but not when it was neutral. While search slopes were positive, they were not modulated by emotion repetition.
Other studies used conjunction search tasks (Becker & Horstmann, 2009; Hillstrom, 2000; Exp.4; Geyer et al., 2006; Koshino, 2001; Kristjánsson et al., 2002Footnote 5; Lamy et al., 2008b) and can be divided in three types. One group of studies falls into the goal-related feature ITP category: participants were instructed to search for one of two conjunction targets (e.g., either a red horizontal or a green vertical bar presented among red vertical and green horizontal distractors) across the experiment (Becker & Horstmann, 2009; Hillstrom, 2000; Exp.4) or were instructed as to what target to search for on any given trial (Koshino, 2001). Thus, in order to find the target participants had to maintain the templates of the candidate targets in working memoryFootnote 6. Only one study in this group reported a reduction in search slopes when the target features repeated (Becker & Horstmann, 2009).
One conjunction-search study (Geyer et al., 2006) did not require maintaining in WM the feature for which the repetition effect was computed: one of the target features was constant and the other could vary unpredictably. For instance, the target might be either a red vertical bar among green vertical bars and red horizontal bars or a red horizontal bar among green horizontal bars and red vertical bars. The target was thus always the orientation singleton within the red subset. In this study, search slopes did not interact with feature ITP.
Finally, one study (Lamy et al., 2008b) examined the effect of repeating the target’s color on search slopes when participants searched for a T among Ls in multi-color displays and reported its orientation. Color was thus an irrelevant dimension of the target. Although there was a robust color repetition effect on overall performance, it did not interact with set size.
Taken together, most of the current literature converges to show that across a variety of features (color, orientation, and facial expressions), feature ITP strongly reduces overall RTs but does not increase search efficiency measured by positive search slopes.
Feature priming of pop-out(PoP) and negative search slopes
In their original demonstration of feature ITP, Maljkovic and Nakayama (1994) used a pop-out search in which search slopes were negative. Negative search slopes are taken to indicate that the target becomes more salient the more populated the displays (e.g., Bravo & Nakayama, 1992). Consistent with the idea that top-down and bottom-up factors interact to guide attention (e.g., Geng & Behrmann, 2005; Otten et al., 2016; Proulx & Egeth, 2006; Rangelov et al., 2017), search slopes are less negative when the target’s feature is pre-cued or fixed across a block (e.g., Bravo & Nakayama, 1992; Leonard & Egeth, 2008). Several studies showed that, likewise, search slopes are less negative when the target feature repeats than when it does not: in other words, the repetition benefit is larger for sparse than for dense displays (e.g., Leonard & Egeth, 2008; Meeter & Olivers, 2006; Rangelov et al., 2017). This finding is unlikely to reflect floor effects on dense-display trials: Rangelov et al. (2017) showed that although responses were much slower for orientation than for color targets (and benefitted more strongly from dense displays), feature ITP was significant for sparse displays and disappeared for dense displays, in both color and orientation search. These findings therefore support the feature-weighting account.
To summarize, repeating the target feature typically did not reduce positive search slopes but consistently reduced negative search slopes. Possible interpretations of this discrepancy are considered in the Discussion.
Feature ITP and high-priority distractors
The presence of a distractor that benefits from high attentional priority impairs search performance because this distractor competes with the target for attention. Thus, if feature ITP affects attentional priority, repeating the target feature should increase the target’s relative attentional weight and reduce the distractor’s influence. Conversely, when the high-priority distractor takes on the previous target’s feature, more weight should accrue to the distractor, which should increase its influence. Studies that adopted this logic relied on two main paradigms: the additional singleton paradigm and the spatial cueing paradigm.
Feature ITP in the additional singleton paradigm
In the additional singleton paradigm pioneered by Theeuwes (1991), participants search for a target on a given dimension (e.g., shape). All the items in the display share the same feature on an irrelevant dimension (e.g., color) except for one, the singleton distractor. Distractor interference refers to poorer performance when the singleton distractor is present relative to when it is absent. This cost is generally taken to indicate that the singleton distractor enjoys higher priority than the target, and that as a result, spatial attention is automatically shifted to the distractor’s location before it is redirected to the target (but for alternative interpretations, see Ester & Awh, 2008; Mathôt et al., 2010). Several studies showed that both bottom-up and top-down factors affect singleton-distractor interference. On the one hand, increasing the target’s salience decreases interference, whereas increasing the singleton distractor’s salience increases such interference (e.g., Theeuwes, 1991, 1992). On the other hand, decreasing the match between the singleton distractor and the target-defining feature reduces the interference (e.g., Bacon & Egeth, 1994), thus demonstrating sensitivity also to top-down factors.
If feature ITP affects attentional guidance, repeating the target should reduce the singleton distractor’s interference. Three groups of studies are relevant for testing this prediction. The experimental differences between them are highlighted in Fig. 4.

Schematic displays illustrating the different measures of feature intertrial priming (ITP) using the additional singleton paradigm. Panel A: Participants search for the unique shape and the salient distractor is a color singleton. Feature ITP is measured for repetitions vs. swaps on the shape (task-relevant) dimension. Panel B: The task is the same but feature ITP is measured for repetitions vs. swaps on the color (task-irrelevant) dimension. Panel C: Participants search for the circle among heterogeneous shapes. Feature ITP is measured for repetitions vs. swaps on the color (task-irrelevant) dimension. Panel D: Participants search for a target that is either red or green and the salient distractor is either blue or yellow. Feature ITP is measured for repetitions vs. changes of the target color. The lower panels depict a swap/switch on trial n relative to trial n-1 (upper panels)
Repetition on the target-defining(task-relevant) dimension
One group of studies used Theeuwes’ (1991) original paradigm and examined the effect of repeating versus swapping the target and distractors’ features on the target-defining dimension (Fig. 4A). That is, the target’s salient shape either repeated from the previous trial or took on the previous non-targets’ feature, but while the non-salient shape of the color distractor either repeated or swapped accordingly, its salient color remained the same. Pinto et al. (2005) showed that distractor interference was reduced when the target and nontarget shapes repeated versus swapped, whereas Lamy et al. (2006b) found no such reduction using a similar task. Lamy and Yashar (2008) showed that the critical difference between the two studies was that conditions of distractor-presence were blocked in the former and randomly mixed in the latter – which should not matter if feature repetition strengthened the priority weight of the target relative to the distractor. Moreover, Becker (2008a) showed that repetition of a target defined by its size did not reduce the large interference produced by a color singleton distractor, even though she used a blocked design (Exps. 1 and 3, but see Exp. 4 where repetition of a target defined by its color did reduce the small interference produced by a size singleton distractor).
Repetition on the salient distractor’s (task-irrelevant) dimension
A second group of studies used a variant of Theeuwes’ (1991) paradigm in which the color singleton distractor had one color and all the remaining items (including the target) another, with the colors either repeating or swapping unpredictably across trials. In some studies, the target was an unpredictable shape singleton (i.e., observers had to use singleton detection mode, Hickey & Theeuwes, 2011; Hickey et al., 2011; Fig. 4B), while in others, it was defined by its known shape and appeared among heterogeneously shaped nontargets (i.e., observers had to use feature search mode, e.g., Gaspelin et al., 2019, Exp. 3Footnote 7; Gaspelin & Luck, 2018, Exps. 1 and 3; Graves & Egeth, 2015, Exp. 1; Fig. 4C). These studies focused on whether or not color repetition (i.e., repetition vs. swap on a task-irrelevant dimension) reduces distractor interference. The critical reduction was reported in all of them.
Repetition of the target-defining feature with a singleton distractor in the same dimension
In a third group of studies, both the target and the critical distractor were singletons on the same dimension (Fig. 4D). In this case, it was necessary to let participants know which one is the target. In some studies, this was achieved by using non-overlapping target and distractor color sets: throughout the experiment, the target might be either red or green and the distractor either yellow or blue, among gray items (Meeter & Olivers, 2006, Exp. 3; Olivers & Hickey, 2010; Olivers & Meeter, 2012). In other studies, the target feature was specified by a precue on a trial-by-trial basis (Theeuwes & van der Burg, 2011). Thus, these studies all fall into the goal-related feature ITP category. In all of them, distractor interference was reduced when the target color was the same on consecutive trials relative to when it changed (but see Ásgeirsson & Kristjánsson, 2019, for a failure to replicate Theeuwes & van der Burg’s, 2011, findings).
To summarize, repeating the target feature clearly reduced distractor interference when a goal-related feature ITP protocol was used, whereas the findings emanating from studies in which feature ITP shared no characteristic with top-down attention were more ambiguous: feature repetition most often did not reduce distractor interference when it pertained to the target-defining feature, but clearly did when the repetition versus swap concerned the distractor’s unique feature on an irrelevant dimension. How can we explain this discrepancy? If feature ITP affects attentional priority, repeating a feature of the target should increase its competitive advantage relative to the salient distractor irrespective of whether repetition occurs on the relevant or on an irrelevant dimension. One possibility is that rather than affecting attentional priority, feature ITP modulates how fast observers are able to disengage their attention from the salient distractor (see Hickey & Theeuwes, 2011, for a similar suggestion). According to this account, the critical difference between the two groups of studies is that on swap trials, the irrelevant singleton is the only object that takes on the previous target’s feature when the feature repetition versus swap occurs on a task-irrelevant dimension. By contrast, when feature repetition versus swap occurs on the target-defining dimension, the previous target’s feature is shared both by the non-targets and the irrelevant singleton. Thus, it should take more time to disengage attention from the irrelevant singleton on swapped than on repeated-feature trials only in the former case – hence the consistent finding that in that case only, feature ITP modulates distractor interference. More evidence is clearly needed to test this conclusion.
Feature ITP in the spatial cueing paradigm
The other paradigm used to study the effect of feature ITP on the impact of salient distractors during search is the spatial cueing paradigm (e.g., Folk et al., 1992). In a typical experiment, observers search for a target among distractors and shortly prior to the search display, a spatially uninformative cue appears at one of the potential target locations. Faster search performance when the target appears at the same location as the cue (valid-cue trials) than at another location (invalid-cue trials) is taken to indicate that attention was shifted to the cue (e.g., Folk et al., 1992; Gaspelin et al., 2016 or that the cue was effective in later biasing attention in favor of its location (Gabbay et al., 2019; Lamy et al., 2018; Yaron & Lamy, 2021).
Early studies using this paradigm demonstrated that top-down factors strongly modulate the cue validity effect (e.g., Folk et al., 1992). They showed that a cue produces a validity effect only when it matches the target-defining feature, for example, in search for a red target, when the cue is red but not when it is green (e.g., Folk & Remington, 1998; for a review, see Büsel et al., 2020). Moreover, only slightly larger effects were reported when the matching cues were made more salient (e.g., Lamy, 2005; Lamy et al., 2004; Schoeberl et al., 2019b). However, recent studies revealed that bottom-up guidance also plays an important role in the spatial cueing paradigm: cue validity effects from cues not matching the search set emerge during difficult search (Gaspelin et al., 2016; Lamy et al., 2018) but only when these cues are abrupt onsets and not when they are static color cues (Ruthruff et al., 2020).
The effect of feature ITP on attentional guidance can be probed using the spatial cueing paradigm by examining whether a cue produces larger validity effects when it shares the feature of the target on the previous trial relative to when it does not (see Fig. 5). When the target feature is constant, effects of feature ITP are confounded with top-down guidance (see Folk & Remington, 2008, for the first formulation of this idea): a cue sharing the previous target’s color always matches the target template, whereas a cue that does not share the previous target’s color never matches the target template. This problem is avoided in two types of studies.

Sample sequence of events in a typical spatial cueing paradigm adapted to examine the effect of feature intertrial priming on the cue validity effect. In this example, the target is either the red or green singleton in the search display (here, it is green) and the cue is either the red or green singleton in the cue display (here, it is red). The comparison of interest is whether the cue validity effect is larger when the cue on trial n shares the color of the target on trial n-1 than when it has the alternative color. ISI interstimulus interval
Two-feature search tasks
In some studies, participants searched for two possible targets, for example, either a green or a red object. The critical question was whether a red cue would produce larger validity effects when the target on the previous trial had been red than when it had been green. The evidence is mixed: some studies reported the critical interaction (Belopolsky et al., 2010, Exp. 2; Folk & Remington, 2008, Exp. 1; Schoeberl, Goller, & Ansorge, 2019a, b, Exp. 1), whereas other studies did not (Biderman et al., 2017; Eimer & Kiss, 2010, Exp. 1; Irons et al., 2012; Schoeberl et al., 2019a, Exp. 3 – see also Yashar et al., 2017). In some studies, participants maintained the possible target features in WM and used feature-search mode (Irons et al., 2012, Exp. 2; Schoeberl et al., 2019a, b, Exp. 1), in others, they searched for the object with the unique feature in the task-relevant dimension (Eimer & Kiss, 2010, Exp. 1; Irons et al., 2012, Exp. 1; Yashar et al., 2017), while in the remaining studies, what strategy was used cannot be unequivocally established (Belopolsky et al., 2010, Exp. 2; Biderman et al., 2017; Folk & Remington, 2008, Exp. 1; Schoeberl et al., 2019a, b, Exp. 3).
Go/no-go tasks
The other group of studies used go/no-go tasks. Displays contained one of two possible targets (say, either a red or a green color singleton), but participants had to respond if the target had one feature and refrain from responding if it had the other. The question of main interest was whether a cue sharing the go feature would produce a larger validity effect when the target on the previous trial had been a go (same feature) versus a no-go (different feature) target. The findings showed that it did not (Eimer & Kiss, 2010, Exp. 2; Folk & Remington, 2008, Exp. 2).
To conclude, there is mixed evidence from spatial cueing studies as to whether feature ITP guides attention – although there is overall more evidence against this possibility. What factors might account for these inconsistent findings remains unclear. Note that target-cue feature repetition never increased the cue validity effect when observers unambiguously used singleton-detection mode and therefore did not store the potentially repeating features in WM. However, there is no clear evidence that goal-related feature ITP affected the cue validity effect.
Prior-entry effects
According to the law of prior entry (Titchener, 1908) “the objects of attention come more quickly to consciousness than the objects which we are not attending to”. Empirical evidence for this phenomenon comes from studies using temporal order judgement (TOJ; e.g., Stelmach & Herdman, 1991) and simultaneity judgement (SJ, e.g., Carver & Brown, 1997) tasks. In both tasks, two objects appear at variable stimulus onset asynchronies (SOAs) from one another. In the TOJ task, participants report which of the objects appeared first; the prior-entry effect is quantified by the point of subjective simultaneity (PSS), that is, the average SOA at which participants are equally likely to report that the object that appeared first was the attended or the unattended object. In the SJ task, participants report whether or not the two objects appeared simultaneously. The typical finding in both tasks is that the attended object must be presented later for participants to judge the two objects as simultaneous (e.g., Schneider & Bavelier, 2003; Stelmach & Herdman, 1991). Accordingly, several authors have relied on prior entry effects to investigate which objects enjoy higher priority (e.g., Born et al., 2015; Donk & Soesman, 2011).
Two studies used this rationale to examine the role of feature ITP. Theeuwes and Van der Burg (2013) used both TOJ and SJ tasks and reported that an object primed by a same-color cue was attended first. However, unlike in classical feature ITP studies, where a benefit is observed for the target color although both the target and distractor colors repeat, here, only the cued color repeated. Thus, the findings may reflect simple perceptual priming.
Burnham (2015) used visual arrays more similar to the classical feature ITP search displays and therefore did not suffer from this problem. He relied on a ToJ task: on each trial, four task-irrelevant squares appeared, either one red among green ones or one green among red ones, randomly. Two probes, flashed one after the other at various SOAs, were superimposed on two of these squares and participants had to determine which appeared first. Probes appearing on the color singleton needed to be presented earlier on swapped than on repeated-color trials in order to be perceived simultaneously with a probe appearing on a nonsingleton. It is noteworthy, however, that PSS calculated from TOJ tasks have been criticized because they might reflect decision biases that emerge more strongly the more ambiguous temporal order is (Shore et al., 2001; Spence & Parise, 2010). In Burnham’s (2015) study, such a decision bias may have manifested in participants’ selecting the same object color on successive trials (see also Brascamp et al., 2011). Further research is therefore needed to ensure that his findings are replicated when response biases are prevented (see Spence & Parise, 2010, for procedures allowing such prevention).
To conclude, while the two studies relying on prior-entry suggest that feature ITP affects attentional priority, their findings are open to alternative accounts. Therefore, further research is required in order to establish this conclusion.
Feature ITP and search accuracy with brief displays
A small group of studies investigated whether feature ITP affects perceptual processing by measuring search performance accuracy in the typical feature ITP task with briefly presented, masked search displays. They relied on the idea that RTs with extended viewing times index both perceptual and post-perceptual stages, whereas accuracy under data-limited conditions indexes only perceptual stages. However, such perceptual processes may be enhanced both if the target is found faster (attentional guidance) and if its features are processed faster after it is found (selection). Yashar and Lamy et al. (2010) suggested that any factor that increases the attentional priority accruing to the target relative to the distractors should improve search accuracy both when the task requires attention to be focused on the target and when it requires only detecting the target. By contrast, a factor that expedites selection should improve accuracy only for focused-attention tasks.
The findings of several studies confirmed the latter prediction over the former. Repetition of the target and distractors features (feature ITP) improved accuracy when the task involved fine discrimination of the response feature (e.g., Kristjánsson, 2016; Pascucci et al., 2012; Sigurdardottir et al., 2008, Exp. 2; Yashar & Lamy, 2010a; see also Ásgeirsson et al., 2014), but not when the task was a left/right hemifield localization of the target feature (Huang et al., 2004; Yashar & Lamy, 2010a). Note that Sigurdardottir et al. (2008, Exp. 1) reported higher accuracy for repeated targets in a detection task. However, in that study, the target was defined as the conjunction of color and orientation, a difficult task that may have encouraged participants to focus their attention on the target. The authors did not report whether accuracy also improved when location repeated, which would be diagnostic of whether or not attention was focused at the target location. Note also that these authors investigated goal-related feature ITP, since the probability of a repeated target color was very high (it was 75% or higher until after the seventh color repetition).
Taken together, these findings suggest that feature ITP affects processes that occur after attention is focused on the target and therefore does not modulate attentional priority. However, two observations invite caution before adopting this conclusion. First, this conclusion relies on the assumption that higher attentional priority should be associated with better detection or coarse localization performance. To instantiate this claim, it would be important to examine whether bottom-up and top-down factors affect accuracy in both the discrimination and the left/right hemifield localization tasks, as predicted if both tasks are indeed sensitive to attentional guidance. Second, findings from a study where accuracy for briefly presented displays was compared for single- and multiple-object displays are at odds with the conclusion that feature ITP affects processing after the target is found. Specifically, Ásgeirsson et al. (2015) reasoned that if feature ITP biases attentional priority, it should improve identification accuracy only when the target competes with non-targets and not when it is presented alone. Their findings confirmed this prediction. Possible avenues for resolving the apparent discrepancies between brief-display studies are considered in the Discussion.
Feature ITP and eye movements
Eye movements (or saccades) that bring an object in the fovea are often interpreted as an overt marker of attentional shifting (Deubel & Schneider, 1996; Hoffman & Subramaniam, 1995). As a result, many visual search studies have used eye-tracking methodology to study the factors that contribute to attentional priority, relying on three main measures.
Measures of attentional priority in eye-movement studies
The proportion of first saccades is thought to be the most straightforward measure, based on the premise that the first object to which the gaze is shifted is the object with the highest priority. Accordingly, the proportion of first saccades to an object was found to increase the more salient this object is (e.g., Irwin et al., 2000; Theeuwes et al., 1998, 1999; van Zoest & Donk, 2005) and when it matches the search template relative to when it does not (e.g., Ludwig & Gilchrist, 2002, 2003a, 2003b; Wu & Remington, 2003).
Another popular measure is saccade latency to the target: the higher the attentional priority of the target, the faster one should be to select it. Saccade latencies to a target are typically faster the more salient it is (e.g., Theeuwes et al., 2003) and when it matches the search template compared to when it does not (e.g., Al-Aidroos & Pratt, 2010). However, saccade latencies may be susceptible to post-guidance processes, such as the decision to launch a saccade after the target is located (Findlay, 1997). As such, they are less unequivocally diagnostic of attentional guidance than proportion of first saccades.
Finally, deviations of saccade trajectory from a straight line to the target can also be used to assess the target’s attentional priority relative to other objects in the visual field (van der Stigchel, Meeter, & Theewues, 2006). Accordingly, the presence of a distractor leads to a more substantial deviation in the saccade path the more salient this distractor is (e.g., Theeuwes et al., 1998) and when this distractor matches the search-relevant feature relative to when it does not (e.g., Ludwig & Gilchrist, 2003a). Thus, saccade trajectories may reveal changes in the relative attentional priority accruing to a target when a distractor competes with it for attention, changes to which the proportion of first saccades may not be sensitive.
Feature ITP in saccade tasks and free-viewing manual response tasks
The studies that relied on eye movements to examine the role of feature ITP on attentional guidance can be divided into two broad categories. In one group of studies, participants were required to make a saccade towards the location of the target. There were fewer erroneous saccades when the target repeated from the previous trial than when it swapped, and correct saccades (to the target) were faster (Becker, 2008b; McPeek, Maljkovic, & Nakayama, 1999). In addition, on correct-saccade swap trials, saccades were more likely than on correct-saccade repeat trials to deviate in the direction of a distractor before being corrected “in-flight” and landing at the actual target’s location (McPeek et al., 2000; for more frequent in-flight corrections of manual reaching responses on swap trials, see also Song & Nakayama, 2006, 2008). Similar findings were reported when monkeys served as participants (e.g., McPeek & Keller, 2001).
In the other group of studies, participants were required to make a manual response and allowed to move their gaze freely while their eye movements were recorded. Therefore, moving one’s eyes to a non-target location did not constitute an error. These studies relied on a variety of search tasks similar to those used in the studies reviewed in the foregoing sections, in which only manual response performance was measured.
Pop-out search
With the standard pop-out task, target fixation latencies (i.e., the time that elapses from the onset of the search display until the target is fixated) were found to be faster when the target repeated than when it swapped (Becker, 2008c; Becker & Ansorge, 2013). Importantly, in the latter study, both target repetition and target salience increased the proportion of first fixations to the target, irrespective of whether the target was defined on the color dimension or on the shape dimension. The attention-guidance account predicts that the effects of feature ITP and target salience should interact, because if feature ITP increases the target’s competitive advantage, its influence should be larger the less salient the target. Yet, while this prediction was confirmed with color-defined targets, the opposite pattern was observed with size-defined targets. Finally, Shurygina et al. (2019) used Maljkovic and Nakayama’s (1994) displays, and in the condition that is most relevant for the present purposes, they randomly mixed sequences of one, two, five, or seven trials of the same color. Therefore, the probability of a color repetition was .73. They showed that first saccades deviated in the direction of distractors sharing the previous target’s color. This finding indicates that goal-related feature ITP guides attention.
Conjunction search
One study (Becker & Horstmann, 2009) examined feature ITP in a conjunction search (see the section Feature ITP and search slopes for more details). Mirroring their RT data, feature ITP reduced search slopes measured on target fixation latencies. In addition, the first fixation was more likely to land on the target on repeated- than on swapped-feature trials. However, although increasing the number of distractors decreased the proportion of first saccades to the target, priming did not reduce this set size effect, as would be expected if repeating the target feature increased its attentional priority. Moreover, the priming effect on the proportion of first saccades to the target interacted with response-feature repetition. First saccades landed less often on the target when one of its features (either its response or defining feature) repeated and the other swapped. Interpreted within the retrieval-based account of feature ITP, the latter finding raises the intriguing possibility that first saccades may not be driven by attentional priority alone, but also by post-selection processes.
Additional singleton search
Becker (2008a, 2010) used the additional singleton paradigm while measuring eye movements. When feature repetition was measured on the target-defining dimension (Becker, 2008a), first saccades were more likely to land on a non-target on swap than on repetition trials, and on distractor-present than on distractor-absent trials, in line with the attentional guidance account of feature ITP. Again, the attention-guidance account predicts that the two effects should interact, because if feature ITP increases the target’s competitive advantage, its influence should be larger when the target competes with a salient distractor than when it does not. Yet, the expected interaction was not observed. To explain this result, Becker (2008a) suggested that the effects of priming may have been too weak to override the large competitive advantage of the salient distractor over the target (p. 338). While this argument is valid, it cannot be easily applied to Becker’s (2008a) results: in Experiment 1, for instance, an interaction would be expected because the magnitude of the priming effect was similar to that of the distractor-presence effect: the average number of non-target fixations per trial rose from .57 to .86 on repetition versus swap trials, respectively, and from .53 to .81 on distractor-absent versus distractor-present trials.
In a later study, Becker (2010) measured effects of feature repetition on the salient distractor’s (irrelevant) dimension but relied on a different index to gauge attentional priority. The salient distractor was present on all trials, and allocation of attention to this distractor was measured as a larger proportion of first saccades to its location relative to other nontargets. This effect was larger when the target (and salient distractor’s) features swapped than when they repeated, suggesting that repeating the target and distractor’s features decreased the salient distractor’s attentional priority relative to that of the nontargets. Gaspelin and colleagues reported compatible findings using a task where participants had to use feature-search mode (i.e., the target was not a singleton and participants had to search for a specific shape). First saccades to the distractor were less likely when its color repeated relative to when it swapped, both when repetitions and swaps occurred randomly (Gaspelin & Luck, 2018, Exp. 3) and when they occurred in predictable five-trial streaks (Gaspelin et al., 2019, Exp. 3).
To conclude, most of the evidence relying on eye movements shows that repeating the target feature increases the probability that the first fixation should land on the target and reduces target fixation latencies, both in saccade tasks and in a variety of manual response tasks where eye movements were monitored. In addition, the probability of making the first saccade to a salient distractor decreases when the target-defining feature repeats, or when the irrelevant feature on which the distractor is salient repeats. These findings support the view that feature ITP guides attention.
Two findings, however, do not fully align with this conclusion. If first saccades are a proxy for priority in the competition for overt attention, and feature ITP modulates first saccades, one expects other manipulations of competition – namely, search slopes and distractor interference – to interact with feature ITP in determining where the first saccade lands – yet such interactions did not emerge (Becker & Horstmann, 2009, and Becker, 2008a, respectively). They are expected because competition for attention is thought to occur on a common map that integrates bottom-up, top-down, and selection history activations (e.g., Awh et al., 2012; Luck et al., 2020; Wolfe, 2021). Therefore, if feature ITP, for instance, increases the target’s priority, it should also reduce the interference from a competing salient distractor (see Becker, 2008a, for a similar argument). Further research is needed to clarify these discrepant findings. In addition, it would be useful to conduct manual-response studies probing the effect of basic feature ITP on saccade deviations, as none was published to date.
Feature ITP and event-related potentials
Event-related potentials (ERPs) are ongoing electroencephalogram (EEG) waveforms that are temporally locked to a specific event and averaged across many trials. Unlike behavioral studies, in which the end outcome of the processes that precede the response is measured, ERP studies provide a direct window into how these processes unfold in time, with high temporal precision.
The N2pc has been the most often studied component with regard to feature ITP. The N2pc is an enhanced negativity over the posterior electrodes contralateral to the presentation of the critical stimulus, and is typically observed between 180 and 330 ms after the visual array is presented. The precise mechanisms associated with the N2pc have been debated. Luck and colleagues initially proposed that the N2pc indexes the suppression of distractor stimuli that interfere with target identification (Luck & Hillyard, 1994; Luck et al., 1997; see also Hopf et al., 2006), Eimer (1996) suggested that “the N2pc primarily reflects the selection of target stimuli …and not a process by which irrelevant stimulation is attenuated or filtered out” (p.233), while Mazza and Caramazza (2011) linked the N2pc with target individuation. Despite these divergences, most accounts of the N2pc converge to suggest that this component reflects processes related to spatial selection. Consequently, the N2pc has become the gold standard ERP marker of attentional allocation in visual search tasks (e.g., Eimer & Kiss, 2010; Hickey et al., 2006; but see McDonald et al., 2013; Lien et al., 2010).
Studies that used ERPs to clarify the mechanisms underlying feature ITP have mainly investigated whether feature ITP affects perceptual or response-related processes (e.g., Becker et al., 2014; Eimer et al., 2010; Olivers & Hickey, 2010; but see Feldmann-Wüstefeld & Schubö, 2016), They focused on the N2pc to answer this question and reasoned that if feature ITP modulates the N2pc, one may conclude that it affects processes that occur prior to response selection. In line with behavioral evidence for a perceptual component of feature ITP (e.g., Ásgeirsson & Kristjánsson, 2011; Lamy et al., 2010), most ERP studies reported that a target elicited an N2pc with an earlier onset (Becker et al., 2014;Christie et al., 2014 ; Tay et al., 2019) sometimes accompanied by higher amplitude (Eimer et al., 2010; Olivers & Hickey, 2010) when this target repeated relative to when it did not.Footnote 8 These findings support the idea that both the basic feature ITP (Becker et al., 2014; Eimer et al., 2010) and goal-related feature ITP (Feldmann-Wüstefeld & Schubö, 2016; Olivers & Hickey, 2010) affect attentional selection.
However, moving beyond the consensus that broadly links the N2pc to attentional selection, several findings converge to show that the N2pc more specifically indexes processes that occur after the competition for attentional priority is resolved. For instance, Tay et al. (2019) suggested an ERP chronometry of visual singleton detection, according to which the N2pc reflects the allocation of attention to the singleton’s location “after the relevance and localization of the singleton are established and encoded” (p.12). Likewise, Kiss et al. (2008) proposed that “N2pc triggered in response to pop-out visual search targets does not reflect processes involved in covert shifts of spatial attention, but is instead linked to spatially selective attentional mechanisms that occur after such shifts are completed” (p.248). This characterization of the N2pc implies that the N2pc reflect processes that follow attentional guidance – namely, “attentional selection” as described in the section titled Processing stages in a typical feature ITP experiment(see Zivony et al., 2018, for related evidence). In other words, while finding a target-locked N2pc is good evidence that the target was attended, finding that feature ITP modulates the N2pc component does not necessarily entail that such modulation reflects a change in attentional priority: it may instead indicate that feature ITP affects processes that occurred after attention was shifted to the target. Accordingly, in order to defend the idea that feature ITP affects attentional priority, one has to show that feature ITP also affects ERP waveforms prior to the N2pc (see Olivers & Hickey, 2010, for a similar argument).
Only three studies examined effects of feature ITP on components that precede the N2pc, and focused on the P1 and N1 components. Two of them (Christie et al., 2014; Tay et al., 2019) reported no modulation of either P1- or N1-related components. Olivers and Hickey (2010) reported an earlier P1 on repeated- relative to switched-target color trials, in a study in which participants had to keep the possible target features in WM in order to perform the task and therefore examined goal-related ITP. What ERP components specifically index attentional priority, however, is not yet clear. As P1 and N1 have been associated with early attentional gating (e.g., Heinze et al., 1994), they may be good candidates (see, e.g., Luck & Kappenman, 2011; Tay et al., 2019; Zhang & Luck, 2009).
To conclude, the extant evidence from ERP studies strongly indicates that feature ITP affects attentional processes. However, most studies relied on the N2pc, which is thought to reflect selective processes that occur after the target is located (e.g., Kiss et al., 2008; Tay et al., 2019; Wascher & Beste, 2010; Zivony et al., 2018). Further research is needed to determine (a) what components might unambiguously index attentional priority and (b) whether these are modulated by feature ITP.
Feature ITP and single-cell recordings
The theoretical concept of “priority map” is invoked by many models of attention to designate a representation of space that codes the relative priority of objects present in the visual field (e.g., Itti & Koch, 2001; Treisman, 1988; Wolfe, 1994; Zelinsky & Bisley, 2015). These objects compete for attentional selection and the most highly prioritized object is chosen for enhanced processing in a winner-take-all fashion. Several brain regions have been singled out as potential priority maps, among them the superior colliculus (SC), the lateral intraparietal area (LIP), and the frontal eye fields (FEFs) and, recently, the posterior inferotemporal cortex (PITd)(for reviews, see Bisley & Goldberg, 2010; Krauzlis et al., 2013; Stemmann & Freiwald, 2019; Thompson & Bichot, 2005).
Finding that feature ITP modulates the activity of neurons in priority maps would provide direct evidence that feature ITP affects attentional guidance (see Westerberg & Schall, 2021, for a review). Bichot and Schall (2002) published the first study that examined the effects of feature ITP on single-cell activity (see also Thompson et al., 2005), and focused on FEF neurons. The task was similar to Maljkovic and Nakayama’s (1994) classical pop-out search task. When the target and distractor colors repeated over a sequence of trials, FEF cell activity increased when the target fell in its receptive field and decreased when a distractor was in its receptive field (see also Westerberg et al., 2020, for similar findings with single-cell recordings from area V4, and Muggleton et al., 2009, for increased RTs on switch trial when left FEF activity was disrupted using transcranial magnetic stimulation (TMS)).
While these findings align with behavioral reports of target facilitation and distractor suppression (Lamy et al., 2008; Kristjánsson & Driver, 2008; Maljkovic & Nakayama, 1994), it should be noted that they all pertain to goal-related ITP. In Bichot and Schall’s (2002) study, the modulation of cell activity by feature repetitions and swaps was reported across a heterogenous combination of short search blocks, randomly intermixed: the color of the target and distractors swapped across trials with a probability of 50% or 33%, or in blocks of ten trials. Thus, overall, a color repetition was considerably more probable than a color swap. This situation prevailed in all the other single-cell studies of feature ITP (Muggleton et al., 2009; Thompson et al., 2005; Westerberg et al., 2020). Although Maljkovic and Nakayama (1994) initially claimed that expectations with regard to repetition and swap probabilities do not affect feature ITP, later studies provided evidence to the contrary (e.g., Cochrane & Pratt, 2020; Shurygina et al., 2019).
To conclude, the evidence from single-cell recording studies strongly suggests that goal-related ITP modulates the activity of neurons in brain areas held to serve as priority maps. Further research is needed to determine whether these findings can be replicated with the standard ITP protocol. In addition, since previous studies all used saccade tasks, it would be useful to examine whether similar findings are observed when participants are required to make manual responses.
Discussion
The idea that selection history guides attentional guidance on par with bottom-up and top-down factors enjoys a wide consensus in the current literature (e.g., Awh et al., 2012; Luck et al., 2020; Wolfe, 2021). Here, we reviewed the empirical studies that tested this claim, while focusing on one selection history phenomenon, feature ITP (aka PoP). Our review suggests that the consensus may be premature, at least with regard to feature ITP.
Goal-related versus basic ITP
The first conclusion that arises from this review is that in order to evaluate the status of feature ITP as a separate source of attentional guidance, it is important to clarify the boundaries of this phenomenon. In Maljkovic and Nakayama’s (1994) initial demonstration, observers searched for a target unique on a salient dimension, and the target feature was equally likely to repeat or swap, unpredictably. As many later studies moved away from the original paradigm, broader definitions of the phenomenon have surfaced. For instance, Ásgeirsson and Kristjánsson (2019) recently defined attentional priming as “… the finding that as observers repeatedly search for the same target, the search becomes faster” – a definition that is inclusive of feature repetitions that occur in streaks (e.g., Bichot & Schall, 2002), manifest as the repetition of one of the two possible target features or feature combinations held in WM (e.g., Olivers & Hickey, 2010), or result from the consecutive presentation of the same informative precue indicating the upcoming target’s feature (e.g., Theeuwes & van der Burg, 2011). In all these cases, the task requires or encourages participants to maintain in WM the feature for which feature ITP is measured and feature ITP therefore shares a characteristic of top-down control: goal-directedness (see the section titled Feature ITP experimental protocols and top-down guidance for more ample justifications). Accordingly, we referred to this phenomenon as goal-related feature ITP and contrasted it with basic feature ITP (for which the task does not require maintaining the critical features in WM).
The findings of our review underscore the importance of this distinction: evidence that goal-related feature ITP guides attention was stronger and more consistent than evidence pertaining to basic feature ITP. This observation is most informative for paradigms in which both types of feature ITP were investigated: in particular, only for goal-related ITP was distractor interference consistently reduced when the target-defining feature repeated. Likewise, the only study that reported an effect of feature repetition on ERP components earlier than the N2pc investigated goal-related feature ITP (Olivers & Hickey, 2010). Single-cell studies also provided very strong support for the idea that goal-related feature ITP modulates attentional priority, but there is to date no single-cell study exploring basic feature ITP. Conjunction search and spatial cueing studies, however, yielded inconsistent findings. While all the studies investigating basic feature ITP most often failed to show any reduction of either search slopes or spatial cueing effects, studies investigating goal-related ITP provided mixed evidence on both measures.
To summarize, the current literature provides fairly strong support for the notion that goal-related ITP modulates attentional priority, and the benefits of additional research on this issue would be therefore limited.
Measures of attentional priority
With regard to basic feature ITP, the different measures of attentional priority reviewed here yielded conflicting results (see Table 1). On the one hand, research relying on positive search slopes and distractor interference with manual performance measures provided little support overall for the priority view. On the other hand, research relying on negative search slopes, prior entry, eye movements (especially on the proportion of first saccades), and ERPs strongly supported the priority view. Studies relying on accuracy with brief masked displays were relatively scarce and yielded mixed findings.
These inconsistencies are most likely to arise from the fact that every measure taken in isolation is open to alternative interpretations. We already underscored this possibility with regard to the N2pc, which many authors take to reflect processes that occur after the candidate target is located, rather than indexing the locus of maximum priority; with regard to distractor interference and positive search slopes, which may not only gauge the relative priorities of the target and distractors but also how long it takes to disengage one’s attention from distractors; and with regard to temporal order judgments, which may reflect decision biases instead of prior entry. In other words, while a given measure may index attentional priority, it may also index additional processes.
To address this difficulty, a potentially useful solution is to jointly manipulate a variable known to affect attentional priority (e.g., physical stimulus salience) together with feature ITP, and observe their effects on each of the candidate measures of attentional priority reviewed here. The critical prediction is that the effects of stimulus salience and feature ITP should interact on the measure of interest (see Sternberg’s (1998) additive factors method). Let us consider, for instance, the finding that feature repetition on a salient distractor’s task-irrelevant dimension consistently reduces distractor interference in the additional singleton paradigm (e.g., Graves & Egeth, 2015). If this effect indicates that feature ITP modulates attentional priority, it should be smaller the more salient the target is. Failure to observe this interaction would indicate that feature ITP affects processes that may be related to selection but are independent of attentional priority. The few studies that provided evidence relevant to this issue yielded mixed findings (Becker & Ansorge, 2013; Becker & Horstmann, 2009). A more systematic application of this rationale is therefore needed.
A role for uncertainty?
Beyond the discrepancies that exist between different measures of attentional priority, unresolved issues also remain within each paradigm. For instance, if feature ITP affects processes that occur after the competition for attention is resolved, how can we explain that it improves search accuracy for masked displays only when the target competes with distractors and not when it appears alone (Ásgeirsson et al., 2015)? Likewise, if adding distractors to the search display increases the competition incurred by the target in tasks yielding positive search slopes and decreases such competition in tasks yielding negative search slopes, how can we explain that feature ITP reliably modulates negative search slopes (e.g., Leonard & Egeth, 2008) but not positive search slopes (e.g., Geyer et al., 2006)?
The idea, suggested by Meeter and Olivers (2006; see also Olivers & Meeter, 2006, 2008), that feature ITP plays a role in resolving ambiguity may help us address at least some of these issues. The foregoing review suggests that several manipulations of competition (e.g., the number of distractors in difficult search) did not modulate feature ITP effects. Therefore, we do not construe ambiguity as being synonymous with competition, and instead define it as the level of uncertainty that prevails after a candidate target is located. When uncertainty is high, repeating the target feature may speed the decision that the candidate target is indeed the target, and therefore how fast attention is engaged in the target.Footnote 9 Uncertainty should be higher when the display contains distractors than when the target appears alone, which explains why effects of feature ITP were absent in the latter condition (Ásgeirsson et al., 2015). Likewise, when the feature that defines the target is its salience, uncertainty should be higher when the display is sparse (and the target may be only marginally more salient than the distractors) than when it is dense (and the target pops out). This explains why feature ITP modulates negative (e.g., Leonard & Egeth, 2008) but not positive search slopes (e.g., Geyer et al., 2006).
Previous work has typically manipulated competition and uncertainty simultaneously and therefore did not allow one to disentangle their respective roles. For instance, Olivers and his colleagues sought to measure the role of uncertainty by comparing trials in which an irrelevant singleton was present or absent (resulting in high vs. low competition, respectively, e.g., Meeter & Olivers, 2006, Exp. 3) or by increasing display size during salience-based search (and thereby reducing competition, e.g., Meeter & Olivers, 2006, Exp. 1). In order to test whether, as we suggest, differences in uncertainty explain inconsistencies in the reviewed literature, further research is needed, in which uncertainty is manipulated independently of competition. For instance, one might vary the proportion of trials in which the target is present across blocks in go/no-go discrimination tasks requiring focused attention. Our tentative uncertainty account predicts that for similar trial sequences, feature ITP should be larger in low-presence (high uncertainty) than on high-presence(low-uncertainty) blocks, irrespective of competition level.
Conclusion
Research on selection history has exploded in the last decade. It has shown that recent experience during search has remarkably large and replicable effects on performance, and has prompted far-reaching changes in how leading figures in the field describe the architecture of attentional control (e.g., Awh et al., 2012; Jiang, 2018; Luck et al., 2020; Wolfe, 2021). It is therefore important to understand what processes are influenced by selection history.
Here, we reviewed whether feature ITP (aka priming of pop-out (PoP)) – one of the most intensively investigated selection history phenomena – guides attention, as do stimulus salience and search goals. We found that it most often does when the task encourages observers to maintain the critical feature in WM – that is, when feature ITP shares the goal-directedness feature of top-down attention. The picture, however, is less clear with regard to the basic feature ITP phenomenon, in which the specific features that the target can take on need not be kept in WM and are equally likely to repeat or switch. Thus, the jury is still out on whether feature ITP is a source of attentional guidance when it shares no characteristic of top-down attention.
To explain many of the inconsistencies that remain in this literature, we speculated that feature ITP may affect processes that occur after the competition for attention is resolved, namely, how fast the winner of the competition is rejected (disengagement) or selected (engagement). We further proposed that uncertainty may strongly modulate this effect. These suggestions yield clear predictions that could be tested in future research.
Beyond the insights it provides about feature ITP, the present review also makes a useful methodological contribution. The role of other selection history phenomena (e.g., location priming, dimension priming, target/distractor location/feature statistical learning, and reward learning) can be clarified by applying the methodology used here, that is: (a) by classifying the different experimental protocols used to test the effects of a selection history phenomenon on attentional priority according to what characteristics associated with top-down attention this phenomenon shares (awareness, flexibility, goal-directedness, etc…), within each protocol; and (b) by reviewing the relevant evidence separately for each of the most widely used measures of attentional priority.
Notes
In the remainder of this review, a feature swap refers to a transition in which the target and distractors exchange their features from one trial to the next (e.g., a red target among green distractors following a green target among red distractors), whereas a feature switch refers to a transition in which the target (or distractors) takes on a feature that was not present on the previous trial (e.g., a red target among blue distractors following a green target among blue distractors for a target-feature switch, and a red target among blue distractors following a red target among green distractors for a distractor-feature switch).
The attentional selection stage is sometimes subdivided into shifting and engagement (e.g., Zivony & Lamy, 2018), but this distinction is not relevant for the current purposes.
In apparent contrast with this claim, Gunseli et al. (2016) presented findings suggesting that repeating the target of a search task reduces the strength of the representation of that target in WM, because this representation is transferred to long-term memory. However, in their study (1) participants had to maintain an additional template in WM for a subsequent memory task and (2) they were informed that the search target would repeat on 20 consecutive trials (in Experiment 1) or on six consecutive trials (in Experiment 2). These characteristics created a strong incentive to transfer the search template to long-term memory. It is unlikely that such transfer should occur when the search template changes randomly from trial to trial and in the absence of the competition of a concurrent template, as was the case in Theeuwes and van der Burg’s (2011) study.
Several studies (e.g., Brinkhuis et al., 2020; Kristjánsson et al., 2007; Rorden et al., 2011) convincingly demonstrated that regions in the fronto-parietal attentional network are involved in feature ITP, in line with the view that feature ITP affects attentional selection. However, in these studies, it was not possible to disentangle activity modulations related to the target, to distractors, or to the display as a whole – which is necessary in order to address the more specific question asked in the present review: does feature ITP bias the relative priority weights of the target versus distractors?
Wang et al. (2005), Exp.6) also used a conjunction search and found no interaction with set size but their finding is not informative for the present purposes because search slopes in their conjunction search were flat (4.5ms / item).
As suggested by Hillstrom (2000), although this task is indeed likely to involve top-down guidance, it remains possible that participants may selectively search through one subset to find a discrepant item, the target (e.g., search through the green bars to find the only vertical bar among horizontal ones), and if unsuccessful, then search through the red bars to find the horizontal only bar among vertical ones.
In that experiment, the colors remained fixed for alternating streaks of five trials (a letter cue also indicated the upcoming target color). In addition, participants were strongly encouraged to use this regularity. Therefore, the phenomenon investigated in that study falls into the category of goal-related feature ITP.
Another study using a variant of the standard PoP paradigm (Burra & Kerzel, 2013) reported no effect of shape repetition on the N2pc but did not report whether shape repetition had a behavioral effect. Hickey et al. (2011) also measured the N2pc elicited by the target and salient distractor in the additional singleton paradigm, but the large variability of the temporal windows used for the various comparisons does not allow one to draw strong conclusions from the findings (see Hickey & Theeuwes, 2011, for additional criticisms of this study).
Note that unlike the retrieval account, which stipulates that feature ITP speeds response-related processes (e.g., Huang et al., 2004), the proposed uncertainty account suggests that feature ITP speeds attentional engagement (as defined by, e.g., Folk et al., 2009; Nieuwenstein et al., 2005; Zivony & Lamy, 2016). This account therefore explains the perceptual rather than the response-related component of feature ITP.
References
Al-Aidroos, N., & Pratt, J. (2010). Top-down control in time and space: Evidence from saccadic latencies and trajectories. Visual Cognition, 18(1), 26–49. https://doi.org/10.1080/13506280802456939
Amunts, L., Yashar, A., & Lamy, D. (2014). Inter-trial priming does not affect attentional priority in asymmetric visual search. Frontiers in Psychology, 5. https://doi.org/10.3389/fpsyg.2014.00957
Anderson, B. A. (2016). The attention habit: How reward learning shapes attentional selection. Annals of the New York Academy of Sciences, 1369(1), 24-39.
Ariga, A., & Kawahara, J. I. (2004). The perceptual and cognitive distractor-previewing effect. Journal of Vision, 4(10), 5-5.
Ásgeirsson, Á. G., & Kristjánsson, Á. (2011). Episodic retrieval and feature facilitation in intertrial priming of visual search. Attention, Perception, & Psychophysics, 73(5), 1350. https://doi.org/10.3758/s13414-011-0119-5
Ásgeirsson, Á. G., & Kristjánsson, Á. (2019). Attentional priming does not enable observers to ignore salient distractors. Visual Cognition, 27(5–8), 595–608. https://doi.org/10.1080/13506285.2019.1657215
Ásgeirsson, Á.G., Kristjánsson, Á. & Bundesen, C. (2014) Independent priming of location and color in identification of briefly presented letters. Attention, Perception and Psychophysics,76, 40–48. https://doi.org/10.3758/s13414-013-0546-6
Ásgeirsson, Á. G., Kristjánsson, Á., & Bundesen, C. (2015). Repetition priming in selective attention: A TVA analysis. Acta Psychologica, 160, 35-42. https://doi.org/10.1016/j.actpsy.2015.06.008
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16(8), 437–443. https://doi.org/10.1016/j.tics.2012.06.010
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. https://doi.org/10.3758/BF03205306
Becker, S. I. (2008a). The mechanism of priming: Episodic retrieval or priming of pop-out? Acta Psychologica, 127(2), 324–339. https://doi.org/10.1016/j.actpsy.2007.07.005
Becker, S. I. (2008b). The stage of priming: Are intertrial repetition effects attentional or decisional? Vision Research, 48(5), 664–684. https://doi.org/10.1016/j.visres.2007.10.025
Becker, S. I. (2008c). Can intertrial effects of features and dimensions be explained by a single theory? Journal of Experimental Psychology. Human Perception and Performance, 34(6), 1417–1440. https://doi.org/10.1037/a0011386
Becker, S. I. (2010). Oculomotor capture by colour singletons depends on intertrial priming. Vision Research, 50(21), 2116–2126. https://doi.org/10.1016/j.visres.2010.08.001
Becker, S. I., & Ansorge, U. (2013). Higher set sizes in pop-out search displays do not eliminate priming or enhance target selection. Vision Research, 81, 18–28. https://doi.org/10.1016/j.visres.2013.01.009
Becker, S. I., & Horstmann, G. (2009). A feature-weighting account of priming in conjunction search. Attention, Perception, & Psychophysics, 71(2), 258–272. https://doi.org/10.3758/APP.71.2.258
Becker, S. I., Grubert, A., & Dux, P. E. (2014). Distinct neural networks for target feature versus dimension changes in visual search, as revealed by EEG and fMRI. NeuroImage, 102, 798–808. https://doi.org/10.1016/j.neuroimage.2014.08.058
Belopolsky, A. V. (2015). Common priority map for selection history, reward and emotion in the oculomotor system. Perception, 44(8-9), 920-933.
Belopolsky, A. V., Schreij, D., & Theeuwes, J. (2010). What is top-down about contingent capture? Attention, Perception, & Psychophysics, 72(2), 326–341. https://doi.org/10.3758/APP.72.2.326
Bichot, N. P., & Schall, J. D. (2002). Priming in Macaque Frontal Cortex during Popout Visual Search: Feature-Based Facilitation and Location-Based Inhibition of Return. Journal of Neuroscience, 22(11), 4675–4685. https://doi.org/10.1523/JNEUROSCI.22-11-04675.2002
Biderman, D., Biderman, N., Zivony, A., & Lamy, D. (2017). Contingent capture is weakened in search for multiple features from different dimensions. Journal of Experimental Psychology. Human Perception and Performance, 43(12), 1974–1992. https://doi.org/10.1037/xhp0000422
Bisley, J. W., & Goldberg, M. E. (2010). Attention, Intention, and Priority in the Parietal Lobe. Annual Review of Neuroscience, 33(1), 1–21. https://doi.org/10.1146/annurev-neuro-060909-152823
Born, S., Kerzel, D., & Pratt, J. (2015). Contingent capture effects in temporal order judgments. Journal of Experimental Psychology: Human Perception and Performance, 41(4), 995.
Brascamp, J. W., Blake, R., & Kristjánsson, Á. (2011). Deciding where to attend: Priming of pop-out drives target selection. Journal of Experimental Psychology: Human Perception and Performance, 37(6), 1700–1707. https://doi.org/10.1037/a0025636
Bravo, M. J., & Nakayama, K. (1992). The role of attention in different visual-search tasks. Perception & Psychophysics, 51(5), 465–472. https://doi.org/10.3758/BF03211642
Brinkhuis, M., Kristjánsson, Á., Harvey, B. M., & Brascamp, J. W. (2020). Temporal Characteristics of Priming of Attention Shifts Are Mirrored by BOLD Response Patterns in the Frontoparietal Attention Network. Cerebral Cortex, 30(4), 2267–2280. https://doi.org/10.1093/cercor/bhz238
Burnham, B. R. (2015). Intertrial priming of popout search on visual prior entry. Journal of Vision, 15(14), 8–8. https://doi.org/10.1167/15.14.8
Burra, N., & Kerzel, D. (2013). Attentional capture during visual search is attenuated by target predictability: Evidence from the N2pc, Pd, and topographic segmentation. Psychophysiology, 50(5), 422–430. https://doi.org/10.1111/psyp.12019
Büsel, C., Voracek, M., & Ansorge, U. (2020). A meta-analysis of contingent-capture effects. Psychological Research, 84(3), 784-809.
Carver, R. A., & Brown, V. (1997). Effects of amount of attention allocated to the location of visual stimulus pairs on perception of simultaneity. Perception & Psychophysics, 59(4), 534-542.
Cave, K. R., & Wolfe, J. M. (1990). Modeling the role of parallel processing in visual search. Cognitive Psychology, 22(2), 225-271.
Chelazzi, L., Perlato, A., Santandrea, E., & Della Libera, C. (2013). Rewards teach visual selective attention. Vision Research, 85, 58-72.
Christie, G. J., Livingstone, A. C., & McDonald, J. J. (2014). Searching for Inefficiency in Visual Search. Journal of Cognitive Neuroscience, 27(1), 46–56. https://doi.org/10.1162/jocn_a_00716
Chun, M. M., & Jiang, Y. (1998). Contextual Cueing: Implicit Learning and Memory of Visual Context Guides Spatial Attention. Cognitive Psychology, 36(1), 28–71. https://doi.org/10.1006/cogp.1998.0681
Chun, M. M., & Jiang, Y. (2003). Implicit, long-term spatial contextual memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29(2), 224–234. https://doi.org/10.1037/0278-7393.29.2.224
Chun, M. M., & Nakayama, K. (2000). On the functional role of implicit visual memory for the adaptive deployment of attention across scenes. Visual Cognition, 7(1-3), 65-81.
Cochrane, B. A., & Pratt, J. (2020). Re-examining Maljkovic and Nakayama (1994): Conscious expectancy does affect the Priming of Pop-out effect. Attention, Perception, & Psychophysics. https://doi.org/10.3758/s13414-020-02034-x
Desimone, R., & Duncan, J. (1995). Neural Mechanisms of Selective Visual Attention. Annual Review of Neuroscience, 18(1), 193–222. https://doi.org/10.1146/annurev.ne.18.030195.001205
Deubel, H., & Schneider, W. X. (1996). Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research, 36(12), 1827-1838.
Donk, M., & Soesman, L. (2011). Object salience is transiently represented whereas object presence is not: Evidence from temporal order judgment. Perception, 40(1), 63-73.
Downing, P. E. (2000). Interactions between visual working memory and selective attention. Psychological Science, 11(6), 467-473.
Druker, M., & Anderson, B. (2010). Spatial Probability Aids Visual Stimulus Discrimination. Frontiers in Human Neuroscience, 4. https://doi.org/10.3389/fnhum.2010.00063
Duncan, J., & Humphreys, G. W. (1989). Visual search and stimulus similarity. Psychological Review, 96(3), 433.
Egeth, H. (2018). Comment on Theeuwes’s characterization of visual selection. Journal of Cognition, 1(1).
Egeth, H. E., Leonard, C. J., & Leber, A. B. (2010). Why salience is not enough: reflections on top-down selection in vision. Acta Psychologica, 135(2), 130–139. https://doi.org/10.1016/j.actpsy.2010.05.012
Eimer, M. (1996). The N2pc component as an indicator of attentional selectivity. Electroencephalography & Clinical Neurophysiology, 99(3), 225–234. https://doi.org/10.1016/0013-4694(96)95711-9
Eimer, M., & Kiss, M. (2010). Top-down search strategies determine attentional capture in visual search: Behavioral and electrophysiological evidence. Attention, Perception, & Psychophysics, 72(4), 951–962. https://doi.org/10.3758/APP.72.4.951
Eimer, M., Kiss, M., & Cheung, T. (2010). Priming of pop-out modulates attentional target selection in visual search: Behavioural and electrophysiological evidence. Vision Research, 50(14), 1353–1361. https://doi.org/10.1016/j.visres.2009.11.001
Ester, E. F., & Awh, E. (2008). The processing locus of interference from salient singleton distractors. Visual Cognition, 16(2–3), 166–181. https://doi.org/10.1080/13506280701489510
Failing, M., & Theeuwes, J. (2018). Selection history: How reward modulates selectivity of visual attention. Psychonomic Bulletin & Review, 25(2), 514–538. https://doi.org/10.3758/s13423-017-1380-y
Feldmann-Wüstefeld, T., & Schubö, A. (2016). Intertrial priming due to distractor repetition is eliminated in homogeneous contexts. Attention, Perception, & Psychophysics, 78(7), 1935–1947.
Ferrante, O., Patacca, A., Di Caro, V., Della Libera, C., Santandrea, E., & Chelazzi, L. (2018). Altering spatial priority maps via statistical learning of target selection and distractor filtering. Cortex, 102, 67–95. https://doi.org/10.1016/j.cortex.2017.09.027
Findlay, J. M. (1997). Saccade Target Selection During Visual Search. Vision Research, 37(5), 617–631. https://doi.org/10.1016/S0042-6989(96)00218-0
Folk, C. L., Ester, E. F., & Troemel, K. (2009). How to keep attention from straying: Get engaged! Psychonomic Bulletin & Review, 16(1), 127–132. https://doi.org/10.3758/PBR.16.1.127
Folk, C. L., & Remington, R. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 847–858. https://doi.org/10.1037/0096-1523.24.3.847
Folk, C. L., & Remington, R. W. (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16(2–3), 215–231. https://doi.org/10.1080/13506280701458804
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology. Human Perception and Performance, 18(4), 1030–1044.
Found, A., & Müller, H. J. (1996). Searching for unknown feature targets on more than one dimension: Investigating a “dimension-weighting” account. Perception & Psychophysics, 58(1), 88-101.
Gabbay, C., Zivony, A., & Lamy, D. (2019). Splitting the attentional spotlight? Evidence from attentional capture by successive events. Visual Cognition, 0(0), 1–19. https://doi.org/10.1080/13506285.2019.1617377
Gao, Y., & Theeuwes, J. (2020). Learning to suppress a distractor is not affected by working memory load. Psychonomic Bulletin & Review, 27(1), 96–104. https://doi.org/10.3758/s13423-019-01679-6
Gaspelin, N., Gaspar, J. M., & Luck, S. J. (2019). Oculomotor inhibition of salient distractors: Voluntary inhibition cannot override selection history. Visual Cognition, 27(3–4), 227–246.
Gaspelin, N., & Luck, S. J. (2018). Distinguishing among potential mechanisms of singleton suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626. https://doi.org/10.1037/xhp0000484
Gaspelin, N., Ruthruff, E., & Lien, M.-C. (2016). The problem of latent attentional capture: Easy visual search conceals capture by task-irrelevant abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 42(8), 1104–1120. https://doi.org/10.1037/xhp0000214
Geng, J. J., & Behrmann, M. (2005). Spatial probability as an attentional cue in visual search. Perception & Psychophysics, 67(7), 1252–1268. https://doi.org/10.3758/BF03193557
Geyer, T., & Müller, H. J. (2009). Distinct, but top-down modulable color and positional priming mechanisms in visual pop-out search. Psychological Research, 73(2), 167-176.
Geyer, T., Müller, H. J., & Krummenacher, J. (2006). Cross-trial priming in visual search for singleton conjunction targets: Role of repeated target and distractor features. Perception & Psychophysics, 68(5), 736–749. https://doi.org/10.3758/BF03193697
Goldstein, R. R., & Beck, M. R. (2018). Visual search with varying versus consistent attentional templates: Effects on target template establishment, comparison, and guidance. Journal of Experimental Psychology: Human Perception and Performance, 44(7), 1086-1102. https://doi.org/10.1037/xhp0000520
Graves, T., & Egeth, H. E. (2015). When does feature search fail to protect against attentional capture? Visual Cognition, 23(9–10), 1098–1123. https://doi.org/10.1080/13506285.2016.1145159
Gunseli, E., Olivers, C. N. L., & Meeter, M. (2016). Task-irrelevant memories rapidly gain attentional control with learning. Journal of Experimental Psychology: Human Perception and Performance, 42(3), 354–362. https://doi.org/10.1037/xhp0000134
Heinze, H. J., Mangun, G. R., Burchert, W., Hinrichs, H., Scholz, M., Münte, T. F., Gös, A., Scherg, M., Johannes, S., & Hundeshagen, H. (1994). Combined spatial and temporal imaging of brain activity during visual selective attention in humans. Nature, 372(6506), 543–546. https://doi.org/10.1038/372543a0
Hickey, C., & Theeuwes, J. (2011). Context and competition in the capture of visual attention. Attention, Perception, & Psychophysics, 73(7), 2053. https://doi.org/10.3758/s13414-011-0168-9
Hickey, C., McDonald, J. J., & Theeuwes, J. (2006). Electrophysiological evidence of the capture of visual attention. Journal of Cognitive Neuroscience, 18(4), 604–613. https://doi.org/10.1162/jocn.2006.18.4.604
Hickey, C., Olivers, C., Meeter, M., & Theeuwes, J. (2011). Feature priming and the capture of visual attention: Linking two ambiguity resolution hypotheses. Brain Research, 1370, 175–184. https://doi.org/10.1016/j.brainres.2010.11.025
Hillstrom, A. P. (2000). Repetition effects in visual search. Perception & Psychophysics, 62(4), 800–817. https://doi.org/10.3758/BF03206924
Hoffman, J. E., & Subramaniam, B. (1995). The role of visual attention in saccadic eye movements. Perception & psychophysics, 57(6), 787-795.
Hopf, J. M., Luck, S. J., Boelmans, K., Schoenfeld, M. A., Boehler, C. N., Rieger, J., & Heinze, H. J. (2006). The neural site of attention matches the spatial scale of perception. Journal of Neuroscience, 26(13), 3532-3540. https://doi.org/10.1523/JNEUROSCI.4510-05.2006
Huang, L., & Pashler, H. (2005). Expectation and repetition effects in searching for featural singletons in very brief displays. Perception & Psychophysics, 67(1), 150–157. https://doi.org/10.3758/BF03195018
Huang, L., Holcombe, A. O., & Pashler, H. (2004). Repetition priming in visual search: Episodic retrieval, not feature priming. Memory & Cognition, 32(1), 12–20. https://doi.org/10.3758/BF03195816
Irons, J. L., Folk, C. L., & Remington, R. W. (2012). All set! Evidence of simultaneous attentional control settings for multiple target colors. Journal of Experimental Psychology: Human Perception and Performance, 38(3), 758–775. https://doi.org/10.1037/a0026578
Irwin, D. E., Colcombe, A. M., Kramer, A. F., & Hahn, S. (2000). Attentional and oculomotor capture by onset, luminance and color singletons. Vision Research, 40(10), 1443–1458. https://doi.org/10.1016/S0042-6989(00)00030-4
Itti L, Koch C. Computational modelling of visual attention. Nat Rev Neurosci. 2001;2(3):194-203. doi:https://doi.org/10.1038/35058500
Jiang, Y. V. (2018). Habitual versus goal-driven attention. Cortex, 102, 107–120. https://doi.org/10.1016/j.cortex.2017.06.018
Kiss, M., Van Velzen, J., & Eimer, M. (2008). The N2pc component and its links to attention shifts and spatially selective visual processing. Psychophysiology, 45(2), 240-249.
Koshino, H. (2001). Activation and inhibition of stimulus features in conjunction search. Psychonomic Bulletin & Review, 8(2), 294–300. https://doi.org/10.3758/BF03196164
Krauzlis, R. J., Lovejoy, L. P., & Zénon, A. (2013). Superior Colliculus and Visual Spatial Attention. Annual Review of Neuroscience, 36(1), 165–182. https://doi.org/10.1146/annurev-neuro-062012-170249
Kristjánsson, A. (2006). Simultaneous priming along multiple feature dimensions in a visual search task. Vision research, 46(16), 2554-2570.
Kristjánsson, Á. (2015). Reconsidering Visual Search. I-Perception. https://doi.org/10.1177/2041669515614670
Kristjánsson, Á. (2016). Priming of Visual Search Facilitates Attention Shifts: Evidence From Object-Substitution Masking. Perception, 45(3), 255–264. https://doi.org/10.1177/0301006615607121
Kristjánsson, Á., & Ásgeirsson, Á. G. (2019). Attentional priming: recent insights and current controversies. Current Opinion in Psychology, 29, 71-75.
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72(1), 5-18.
Kristjánsson, Á., & Driver, J. (2008). Priming in visual search: Separating the effects of target repetition, distractor repetition and role-reversal. Vision Research, 48(10), 1217-1232.
Kristjánsson, A., Vuilleumier, P., Schwartz, S., Macaluso, E., & Driver, J. (2007). Neural basis for priming of pop-out during visual search revealed with fMRI. Cerebral cortex, 17(7), 1612–1624. https://doi.org/10.1093/cercor/bhl072
Kristjánsson, A., Wang, D., & Nakayama, K. (2002). The role of priming in conjunctive visual search. Cognition, 85(1), 37-52.
Kruijne, W., & Meeter, M. (2015). The long and the short of priming in visual search. Attention, Perception, & Psychophysics, 77(5), 1558-1573.
Lamy, D. (2005). Temporal expectations modulate attentional capture. Psychonomic Bulletin & Review, 12(6), 1112-1119.
Lamy, D. F., & Kristjánsson, Á. (2013). Is goal-directed attentional guidance just intertrial priming? A review. Journal of Vision, 13(3), 14–14. https://doi.org/10.1167/13.3.14
Lamy, D., & Yashar, A. (2008). Intertrial target-feature changes do not lead to more distraction by singletons: Target uncertainty does. Vision Research, 48(10), 1274–1279. https://doi.org/10.1016/j.visres.2008.02.021
Lamy, D., Amunts, L., & Bar-Haim, Y. (2008a). Emotional priming of pop-out in visual search. Emotion, 8(2), 151–161. https://doi.org/10.1037/1528-3542.8.2.151
Lamy, D., Antebi, C., Aviani, N., & Carmel, T. (2008b). Priming of pop-out provides reliable measures of target activation and distractor inhibition in selective attention. Vision Research, 48(1), 30-41.
Lamy, D., Bar-Anan, Y., & Egeth, H. E. (2008c). The role of within-dimension singleton priming in visual search. Journal of Experimental Psychology: Human Perception and Performance, 34(2), 268.
Lamy, D., Bar-Anan, Y., Egeth, H. E., & Carmel, T. (2006a). Effects of top-down guidance and singleton priming on visual search. Psychonomic Bulletin & Review, 13(2), 287-293.
Lamy, D., Carmel, T., Egeth, H. E., & Leber, A. B. (2006b). Effects of search mode and intertrial priming on singleton search. Perception & Psychophysics, 68(6), 919–932. https://doi.org/10.3758/BF03193355
Lamy, D., Darnell, M., Levi, A., & Bublil, C. (2018). Testing the Attentional Dwelling Hypothesis of Attentional Capture. Journal of Cognition, 1(1), 43. https://doi.org/10.5334/joc.48
Lamy, D., Goshen-Kosover, A., Aviani, N., Harari, H., & Levkovitz, H. (2008d). Implicit memory for spatial context in depression and schizophrenia. Journal of Abnormal Psychology, 117(4), 954–961. https://doi.org/10.1037/a0013867
Lamy, D., Leber, A., & Egeth, H. E. (2004). Effects of Task Relevance and Stimulus-Driven Salience in Feature-Search Mode. Journal of Experimental Psychology: Human Perception and Performance, 30(6), 1019–1031. https://doi.org/10.1037/0096-1523.30.6.1019
Lamy, D., Yashar, A. & Ruderman, L. (2010). A dual-stage account of inter-trial priming effects. Vision Research, 48, 1274-1279.
Lamy, D., Yashar, A., & Ruderman, L. (2013). Orientation search is mediated by distractor suppression: Evidence from priming of pop-out. Vision Research, 81, 29-35.
Lamy, D., Zivony, A. & Yashar, A. (2011). The role of search difficulty in intertrial feature priming. Vision Research, 51, 2099-2109.
Lee, H., Mozer, M. C., & Vecera, S. P. (2009). Mechanisms of priming of pop-out: Stored representations or feature-gain modulations? Attention, Perception, & Psychophysics, 71(5), 1059–1071. https://doi.org/10.3758/APP.71.5.1059
Leonard, C. J., & Egeth, H. E. (2008). Attentional guidance in singleton search: An examination of top-down, bottom-up, and intertrial factors. Visual Cognition, 16(8), 1078–1091. https://doi.org/10.1080/13506280701580698
Li, A., Wolfe, J. M., & Chen, Z. (2020). Implicitly and explicitly encoded features can guide attention in free viewing. Journal of Vision, 20(6), 8-8.
Libera, C. D., & Chelazzi, L. (2006). Visual selective attention and the effects of monetary rewards. Psychological Science, 17(3), 222-227.
Lien, M. C., Ruthruff, E., & Cornett, L. (2010). Attentional capture by singletons is contingent on top-down control settings: Evidence from electrophysiological measures. Visual Cognition, 18(5), 682-727.
Luck, S. J., Chelazzi, L., Hillyard, S. A., & Desimone, R. (1997). Neural mechanisms of spatial selective attention in areas V1, V2, and V4 of macaque visual cortex. Journal of Neurophysiology, 77(1), 24-42. https://doi.org/10.1152/jn.1997.77.1.24
Luck, S. J., & Kappenman, E. S. (Eds.). (2011). The Oxford handbook of event-related potential components. Oxford university press.
Luck, S. J., Gaspelin, N., Folk, C. L., Remington, R. W., & Theeuwes, J. (2020). Progress Toward Resolving the Attentional Capture Debate. Visual cognition, 29(1), 1–21. https://doi.org/10.1080/13506285.2020.1848949
Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during visual search: Evidence from human electrophysiology. Journal of Experimental Psychology: Human Perception and Performance, 20(5), 1000–1014. https://doi.org/10.1037/0096-1523.20.5.1000
Ludwig, C. J. H., & Gilchrist, I. D. (2002). Stimulus-driven and goal-driven control over visual selection. Journal of Experimental Psychology: Human Perception and Performance, 28(4), 902–912. https://doi.org/10.1037/0096-1523.28.4.902
Ludwig, C. J. H., & Gilchrist, I. D. (2003a). Target similarity affects saccade curvature away from irrelevant onsets. Experimental Brain Research, 152(1), 60–69. https://doi.org/10.1007/s00221-003-1520-7
Ludwig, C. J. H., & Gilchrist, I. D. (2003b). Goal-driven modulation of oculomotor capture. Perception & Psychophysics, 65(8), 1243–1251. https://doi.org/10.3758/BF03194849
Maljkovic, V., & Martini, P. (2005). Implicit short-term memory and event frequency effects in visual search. Vision Research, 45(21), 2831-2846.
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22(6), 657–672. https://doi.org/10.3758/BF03209251
Maljkovic, V., & Nakayama, K. (1996). Priming of pop-out: II. The role of position. Perception & Psychophysics, 58(7), 977–991. https://doi.org/10.3758/BF03206826
Maljkovic, V., & Nakayama, K. (2000). Priming of popout: III. A short-term implicit memory system beneficial for rapid target selection. Visual cognition, 7(5), 571-595.
Martini, P. (2010). System identification in priming of pop-out. Vision research, 50(21), 2110-2115.
Mathôt, S., Hickey, C., & Theeuwes, J. (2010). From reorienting of attention to biased competition: Evidence from hemifield effects. Attention, Perception, & Psychophysics, 72(3), 651-657.
Mazza, V., & Caramazza, A. (2011). Temporal brain dynamics of multiple object processing: the flexibility of individuation. PloS one, 6(2), e17453. https://doi.org/10.1371/journal.pone.0017453
McDonald, J. J., Green, J. J., Jannati, A., & Di Lollo, V. (2013). On the electrophysiological evidence for the capture of visual attention. Journal of Experimental Psychology: Human Perception and Performance, 39(3), 849–860.
McPeek, R. M., & Keller, E. L. (2001). Short-term priming, concurrent processing, and saccade curvature during a target selection task in the monkey. Vision Research, 41(6), 785-800.
McPeek, R. M., Maljkovic, V., & Nakayama, K. (1999). Saccades require focal attention and are facilitated by a short-term memory system. Vision Research, 39(8), 1555–1566. https://doi.org/10.1016/S0042-6989(98)00228-4
McPeek, R. M., Skavenski, A. A., & Nakayama, K. (2000). Concurrent processing of saccades in visual search. Vision Research, 40(18), 2499–2516. https://doi.org/10.1016/S0042-6989(00)00102-4
Meeter, M., & Olivers, C. N. L. (2006). Intertrial priming stemming from ambiguity: A new account of priming in visual search. Visual Cognition, 13(2), 202–222. https://doi.org/10.1080/13506280500277488
Mounts, J. R. (2000). Attentional capture by abrupt onsets and feature singletons produces inhibitory surrounds. Perception & Psychophysics, 62(7), 1485-1493.
Muggleton, Neil & Juan, Chi-Hung, Cowey, Alan & Walsh, Vincent & O'Breathnach, Uinsionn. (2009). Human frontal eye fields and target switching. Cortex; A Journal Devoted to the Study of the Nervous System and Behavior. 46. 178-84. https://doi.org/10.1016/j.cortex.2009.01.011.
Nieuwenstein, M. R., Chun, M. M., van der Lubbe, R. H. J., & Hooge, I. T. C. (2005). Delayed Attentional Engagement in the Attentional Blink. Journal of Experimental Psychology: Human Perception and Performance, 31(6), 1463–1475. https://doi.org/10.1037/0096-1523.31.6.1463
Oberauer, K. (2002). Access to information in working memory: Exploring the focus of attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 28(3), 411–421. https://doi.org/10.1037/0278-7393.28.3.411
Olivers, C. N. L., & Hickey, C. (2010). Priming resolves perceptual ambiguity in visual search: Evidence from behaviour and electrophysiology. Vision Research, 50(14), 1362–1371. https://doi.org/10.1016/j.visres.2009.11.022
Olivers, C. N. L., & Meeter, M. (2006). On the dissociation between compound and present/absent tasks in visual search: Intertrial priming is ambiguity driven. Visual Cognition, 13(1), 1–28. https://doi.org/10.1080/13506280500308101
Olivers, C. N. L., & Meeter, M. (2008). Feature priming in visual search does not depend on the dimensional context. Visual Cognition, 16(6), 785-803.
Olivers, C. N. L., & Meeter, M. (2012). Current versus past ambiguity in intertrial priming. Visual cognition, 20(6), 627-646.
Olivers, C. N. L., Meijer, F., & Theeuwes, J. (2006). Feature-based memory-driven attentional capture: visual working memory content affects visual attention. Journal of Experimental Psychology: Human Perception and Performance, 32(5), 1243.
Olivers, C. N. L., Peters, J., Houtkamp, R., & Roelfsema, P. R. (2011). Different states in visual working memory: When it guides attention and when it does not. Trends in cognitive sciences, 15(7), 327-334.
Olivers, C. N. L & Humphreys, G. W. (2003). Visual marking inhibits singleton capture. Cognitive Psychology, 47(1), 1-42.
Otten, M., Schreij, D., & Los, S. A. (2016). The interplay of goal-driven and stimulus-driven influences on spatial orienting. Attention, Perception, & Psychophysics, 78(6), 1642–1654. https://doi.org/10.3758/s13414-016-1121-8
Pascucci, D., Mastropasqua, T., & Turatto, M. (2012). Permeability of priming of pop out to expectations. Journal of Vision, 12(10), 21-21. https://doi.org/10.1167/12.10.21
Pashler, H., & Shiu, L. P. (1999). Do images involuntarily trigger search? A test of Pillsbury’s hypothesis. Psychonomic Bulletin & Review, 6(3), 445-448.
Pinto, Y., Olivers, C. L., & Theeuwes, J. (2005). Target uncertainty does not lead to more distraction by singletons: Intertrial priming does. Perception & Psychophysics, 67(8), 1354–1361. https://doi.org/10.3758/BF03193640
Proulx, M. J., & Egeth, H. E. (2006). Target-nontarget similarity modulates stimulus-driven control in visual search. Psychonomic Bulletin & Review, 13(3), 524–529. https://doi.org/10.3758/bf03193880
Rangelov, D., Müller, H. J., & Zehetleitner, M. (2017). Failure to pop out: Feature singletons do not capture attention under low signal-to-noise ratio conditions. Journal of Experimental Psychology: General, 146(5), 651–671. https://doi.org/10.1037/xge0000284
Rorden, C., Kristjansson, A., Revill, K. P., & Saevarsson, S. (2011). Neural correlates of inter-trial priming and role-reversal in visual search. Frontiers in human neuroscience, 5, 151. https://doi.org/10.3389/fnhum.2011.00151
Ruthruff, E., Faulks, M., Maxwell, J.W., & Gaspelin, N. (2020). Attentional dwelling and capture by color singletons. Attention, Perception, & Psychophysics.
Schneider, K. A., & Bavelier, D. (2003). Components of visual prior entry. Cognitive psychology, 47(4), 333-366.
Schoeberl, T., Goller, F., & Ansorge, U. (2019a). Testing a priming account of the contingent-capture effect. Attention, Perception, & Psychophysics, 81(5), 1262–1282. https://doi.org/10.3758/s13414-019-01672-0
Schoeberl, T., Goller, F., & Ansorge, U. (2019b). Top-down matching singleton cues have no edge over top-down matching nonsingletons in spatial cueing. Psychonomic Bulletin & Review, 26(1), 241–249. https://doi.org/10.3758/s13423-018-1499-5
Sha, L. Z., & Jiang, Y. V. (2016). Components of reward-driven attentional capture. Attention, Perception, & Psychophysics, 78(2), 403–414. https://doi.org/10.3758/s13414-015-1038-7
Shore, D. I., Spence, C., & Klein, R. M. (2001). Visual Prior Entry. Psychological Sciencehttps://doi.org/10.1111/1467-9280.00337
Shurygina, O., Kristjánsson, Á., Tudge, L., & Chetverikov, A. (2019). Expectations and perceptual priming in a visual search task: Evidence from eye movements and behavior. Journal of Experimental Psychology: Human Perception and Performance, 45(4), 489-499. https://doi.org/10.1037/xhp0000618
Sigurdardottir, H. M., Kristjánsson, A., & Driver, J. (2008). Repetition streaks increase perceptual sensitivity in visual search of brief displays. Visual cognition, 16(5), 643–658. https://doi.org/10.1080/13506280701218364
Sisk, C. A., Remington, R. W., & Jiang, Y. V. (2019). Mechanisms of contextual cueing: A tutorial review. Attention, Perception, & Psychophysics, 81(8), 2571–2589. https://doi.org/10.3758/s13414-019-01832-2
Song, J. H., & Nakayama, K. (2006). Role of focal attention on latencies and trajectories of visually guided manual pointing. Journal of vision, 6(9), 11-11.
Song, J.-H., & Nakayama, K. (2008). Target selection in visual search as revealed by movement trajectories. Vision Research, 48(7), 853–861. https://doi.org/10.1016/j.visres.2007.12.015
Soto, D., Heinke, D., Humphreys, G. W., & Blanco, M. J. (2005). Early, involuntary top-down guidance of attention from working memory. Journal of Experimental Psychology: Human Perception and Performance, 31(2), 248.
Spence, C., & Parise, C. (2010). Prior-entry: a review. Consciousness and cognition, 19(1), 364–379. https://doi.org/10.1016/j.concog.2009.12.001
Stelmach, L. B., & Herdman, C. M. (1991). Directed attention and perception of temporal order. Journal of Experimental Psychology: Human Perception and Performance, 17(2), 539.
Stemmann, H., & Freiwald, W. A. (2019). Evidence for an attentional priority map in inferotemporal cortex. Proceedings of the National Academy of Sciences, 116(47), 23797–23805. https://doi.org/10.1073/pnas.1821866116
Sternberg, S. (1998). Discovering mental processing stages: The method of additive factors. In D. Scarborough & S. Sternberg (Eds.), An invitation to cognitive science. Methods, models, and conceptual issues: An invitation to cognitive science, Vol. 4 (p. 703–863). The MIT Press.
Tay, D., Harms, V., Hillyard, S. A., & McDonald, J. J. (2019). Electrophysiological correlates of visual singleton detection. Psychophysiology, 56(8), e13375. https://doi.org/10.1111/psyp.13375
Theeuwes, J. (1991). Cross-dimensional perceptual selectivity. Perception & Psychophysics, 50(2), 184–193. https://doi.org/10.3758/BF03212219
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J. (2013). Feature-based attention: It is all bottom-up priming. Philosophical Transactions of the Royal Society B: Biological Sciences, 368(1628), 20130055.
Theeuwes, J. (2018). Visual Selection: Usually Fast and Automatic; Seldom Slow and Volitional. Journal of Cognition, 1(1), 29. https://doi.org/10.5334/joc.13
Theeuwes, J., & Van der Burg, E. (2011). On the limits of top-down control of visual selection. Attention, Perception, & Psychophysics, 73(7), 2092. https://doi.org/10.3758/s13414-011-0176-9
Theeuwes, J., & Van der Burg, E. (2013). Priming makes a stimulus more salient. Journal of Vision, 13(3), 21–21.
Theeuwes, J., de Vries, G.-J., & Godijn, R. (2003). Attentional and oculomotor capture with static singletons. Perception & Psychophysics, 65(5), 735–746. https://doi.org/10.3758/BF03194810
Theeuwes, J., Kramer, A. F., Hahn, S., & Irwin, D. E. (1998). Our eyes do not always go where we want them to go: Capture of the eyes by new objects. Psychological Science, 9(5), 379-385.
Theeuwes, J., Kramer, A. F., Hahn, S., Irwin, D. E., & Zelinsky, G. J. (1999). Influence of attentional capture on oculomotor control. Journal of experimental psychology: human perception and performance, 25(6), 1595.
Thompson, K. G., & Bichot, N. P. (2005). A visual salience map in the primate frontal eye field. In Progress in Brain Research (Vol. 147, pp. 249–262). Elsevier. https://doi.org/10.1016/S0079-6123(04)47019-8
Thompson, K. G., Bichot, N. P., & Sato, T. R. (2005). Frontal eye field activity before visual search errors reveals the integration of bottom-up and top-down salience. Journal of neurophysiology, 93(1), 337–351. https://doi.org/10.1152/jn.00330.2004
Thomson, D. R., & Milliken, B. (2012). Perceptual distinctiveness produces long-lasting priming of pop-out. Psychonomic Bulletin & Review, 19(2), 170–176. https://doi.org/10.3758/s13423-011-0199-1
Thomson, D. R., & Milliken, B. (2013). Contextual distinctiveness produces long-lasting priming of pop-out. Journal of Experimental Psychology: Human Perception and Performance, 39(1), 202.
Titchener, E. B. (1908). Lectures on the elementary psychology of feeling and attention. Macmillan.
Todd, R. M., & Manaligod, M. G. (2018). Implicit guidance of attention: The priority state space framework. Cortex, 102, 121-138.
Treisman, A. (1988). Features and Objects: The Fourteenth Bartlett Memorial Lecture. The Quarterly Journal of Experimental Psychology Section A, 40(2), 201–237. https://doi.org/10.1080/02724988843000104
Treisman, A., & Sato, S. (1990). Conjunction search revisited. Journal of experimental psychology: human perception and performance, 16(3), 459.
Van der Stigchel, S., Meeter, M., & Theeuwes, J. (2006). Eye movement trajectories and what they tell us. Neuroscience & Biobehavioral Reviews, 30(5), 666–679. https://doi.org/10.1016/j.neubiorev.2005.12.001
van Zoest, W., & Donk, M. (2005). The effects of salience on saccadic target selection. Visual Cognition, 12(2), 353–375. https://doi.org/10.1080/13506280444000229
Westerberg, J. A., Maier, A., Woodman, G. F., & Schall, J. D. (2020). Performance Monitoring during Visual Priming. Journal of cognitive neuroscience, 32(3), 515–526. https://doi.org/10.1162/jocn_a_01499
Westerberg, J. A., & Schall, J. D. (2021). Neural mechanism of priming in visual search. Attention, perception & psychophysics, 83(2), 587–602. https://doi.org/10.3758/s13414-020-02118-8
Wan, X., & Lleras, A. (2010). The effect of feature discriminability on the intertrial inhibition of focused attention. Visual Cognition, 18(6), 920-944.
Wang, D., Kristjansson, A., & Nakayama, K. (2005). Efficient visual search without top-down or bottom-up guidance. Perception & Psychophysics, 67(2), 239-253.
Wascher, E., & Beste, C. (2010). Tuning Perceptual Competition. Journal of Neurophysiology, 103(2), 1057–1065. https://doi.org/10.1152/jn.00376.2009
Wolfe, J. M. (1994). Guided Search 2.0 A revised model of visual search. Psychonomic Bulletin & Review, 1(2), 202–238. https://doi.org/10.3758/BF03200774
Wolfe, J. M. (2016). Visual Search Revived: The Slopes Are Not That Slippery: A comment on Kristjansson (2015). i-Perception, 1–6.
Wolfe, J. M. (2019). Visual attention: The multiple ways in which history shapes selection. Current Biology, 29(5), R155-R156.
Wolfe J. M. (2021). Guided Search 6.0: An updated model of visual search. Psychonomic bulletin & review. Advance online publication. https://doi.org/10.3758/s13423-020-01859-9
Wolfe, J. M., & Horowitz, T. S. (2017). Five factors that guide attention in visual search. Nature Human Behaviour, 1(3), 1-8.
Wolfe, J. M., Butcher, S. J., Lee, C., & Hyle, M. (2003). Changing your mind: On the contributions of top-down and bottom-up guidance in visual search for feature singletons. Journal of Experimental Psychology: Human Perception and Performance, 29(2), 483–502. https://doi.org/10.1037/0096-1523.29.2.483
Won, B. Y., Kosoyan, M., & Geng, J. J. (2019). Evidence for second-order singleton suppression based on probabilistic expectations. Journal of Experimental Psychology: Human Perception and Performance, 45(1), 125.
Wu, S.-C., & Remington, R. W. (2003). Characteristics of covert and overt visual orienting: Evidence from attentional and oculomotor capture. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1050–1067. https://doi.org/10.1037/0096-1523.29.5.1050
Yaron, I., & Lamy, D. (2021). Spatial cueing effects are not what we thought: On the timing of attentional deployment. Journal of Experimental Psychology: Human Perception and Performance, 47(7), 946–962. https://doi.org/10.1037/xhp0000918
Yashar, A., & Lamy, D. (2010a). Intertrial repetition affects perception: The role of focused attention. Journal of vision, 10(14), 3-3.
Yashar, A., & Lamy, D. (2010b). Intertrial repetition facilitates selection in time: common mechanisms underlie spatial and temporal search. Psychological Science, 21(2), 243–251. https://doi.org/10.1177/0956797609357928
Yashar, A., & Lamy, D. (2011). Refining the dual-stage account of intertrial feature priming: Does motor response or response feature matter? Attention, Perception, & Psychophysics, 73(7), 2160.
Yashar, A., Makovski, T., & Lamy, D. (2013). The role of motor response in implicit encoding: Evidence from intertrial priming in pop-out search. Vision research, 93, 80-87.
Yashar, A., White, A. L., Fang, W., & Carrasco, M. (2017). Feature singletons attract spatial attention independently of feature priming. Journal of Vision, 17(9), 7–7. https://doi.org/10.1167/17.9.7
Zelinsky, G. J., & Bisley, J. W. (2015). The what, where, and why of priority maps and their interactions with visual working memory. Annals of the New York Academy of Sciences, 1339(1), 154–164. https://doi.org/10.1111/nyas.12606
Zhang, W., & Luck, S. J. (2009). Feature-based attention modulates feedforward visual processing. Nature Neuroscience, 12(1), 24–25. https://doi.org/10.1038/nn.2223
Zivony, A., Allon, A. S., Luria, R., & Lamy, D. (2018). Dissociating between the N2pc and attentional shifting: An attentional blink study. Neuropsychologia, 121, 153–163. https://doi.org/10.1016/j.neuropsychologia.2018.11.003
Zivony, A., & Lamy, D. (2016). Attentional capture and engagement during the attentional blink: A “camera” metaphor of attention. Journal of Experimental Psychology: Human Perception and Performance, 42(11), 1886–1902. https://doi.org/10.1037/xhp0000286
Zivony, A., & Lamy, D. (2018). Contingent Attentional Engagement: Stimulus- and Goal-Driven Capture Have Qualitatively Different Consequences. Psychological Science, 29(12), 1930–1941. https://doi.org/10.1177/0956797618799302
Author’s note
Support was provided by the Israel Science Foundation (ISF) grant no. 1286/16 to Dominique Lamy.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ramgir, A., Lamy, D. Does feature intertrial priming guide attention? The jury is still out. Psychon Bull Rev 29, 369–393 (2022). https://doi.org/10.3758/s13423-021-01997-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-021-01997-8