
Abstract
Pet dogs are known to be responsive to human pointing gestures, but shelter dogs have repeatedly demonstrated poor abilities to follow human pointing, although they can be explicitly trained quickly. This study evaluated the time course in which shelter dogs learn to follow points without explicit training, when given typical interactions with humans. In a longitudinal evaluation, the development of point following was tracked in seven shelter dogs in a training program (enriched human exposure), seven dogs in a traditional shelter (control population), and evaluated once in pet dogs. Twice a week for 6 weeks, shelter dogs’ point-following performance was evaluated in 10 probe trials in which an experimenter pointed to one of two containers equidistant from the dog. To avoid direct training, dogs were given a treat for approaching and touching either container; although correct responses were recorded for touching the pointed-towards container within 30 s. Pet dogs were tested in only one session. All shelter dogs initially showed the expected poor performance. However, enriched shelter dogs receiving enriched human exposure showed significant improvements reaching an identical performance to pet dogs within 7 weeks. In comparison, shelter dogs under standard conditions showed an initial improvement, but performance reached asymptote close to chance levels and lower than that of enriched dogs or pet dogs. Together, these results suggest that enriched experiences with humans, typical of pet dogs, is sufficient for dogs to learn to follow points without explicit training.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Pointing is a form of social referencing and is an important stage in child development (Behne, Carpenter, & Tomasello, 2005; Grassmann & Tomasello, 2010; Hodges, Özçalışkan, & Williamson, 2018; Thoermer & Sodian, 2001). Comparative cognition research over the past 20 years has investigated the development of similar point-following behaviors in humans and nonhuman animals (Call, Hare, & Tomasello, 1998; Hall, Udell, Dorey, Walsh, & Wynne, 2011; Kaminski, Riedel, Call, & Tomasello, 2005; Malassis & Delfour, 2015; Maros, Gácsi, & Miklósi, 2008; Miklósi, Pongrácz, Lakatos, Topál, & Csányi, 2005; Smet & Byrne, 2013; Hall et al., 2011). Such research has focused on domestic dogs, as pet dogs generally perform well on point-following tasks (Lazarowski & Dorman, 2015; Miklósi, Polgárdi, Topál, & Csányi, 1998; Udell, Dorey, & Wynne, 2008). Furthermore, initial studies have suggested that wolves performed poorly at point-following tasks, especially compared with dogs (Hare, Brown, Williamson, & Tomasello, 2002; Virányi et al., 2008). This suggested that perhaps point following was an evolved behavior as a by-product of domestication and cohabitation with humans (Hare et al., 2002; Hare et al., 2010; Hare & Tomasello, 2005; Riedel, Schumann, Kaminski, Call, & Tomasello, 2008; Virányi et al., 2008). This behavior at first seemed to be unique to domestic dogs, as other nonhuman species, even chimpanzees, have been shown to fail to follow human gestures (Kirchhofer, Zimmermann, Kaminski, & Tomasello, 2012). However, subsequent studies have found evidence supporting that chimpanzees and wolves are both capable of using human pointing gestures at the same level or better than dogs with intensive socialization or extensive training (Hopkins, Russell, McIntyre, & Leavens, 2013; Udell et al., 2008; Virányi et al., 2008). Virányi et al. (2008) showed that at the age of 4 months, dog pups outperform wolf pups at a pointing task, however adult wolves are able to perform comparably to adult dogs with extensive training and socialization.
Two primary hypotheses have been established to explain why domesticated dogs understand human social cues—the two-stage hypothesis and the domestication hypothesis. However, mixed support has been found for these hypotheses. The two-stage hypothesis treats the responsiveness of animals to human gestures as dependent on the animal’s acceptance of humans as social companions (Stage 1) and their level of conditioning to follow human limbs (Stage 2; Dorey, Udell, & Wynne, 2010; Udell et al., 2008; Udell, Dorey, & Wynne, 2010a, 2010b; Wynne, Udell, & Lord, 2008). Thus, this hypothesis considers that the amount of exposure to humans is a critical factor that affects point-following performance. In support of this hypothesis, Udell et al. (2010a) found that shelter dogs performed poorly on a point-following task, suggesting dogs with less human exposure do not perform as well. Further, this study showed that the dogs could be trained to follow points within 40 trials, suggesting that given appropriate training history and associations, dogs can learn to follow points, supporting the two-stage hypothesis (Udell et al., 2010a). This study, however, used a training paradigm, rewarding correct responses and not rewarding incorrect response. Under these conditions, theoretically, any animal capable of operant conditioning would be expected to learn the task.
The domestication hypothesis suggests that domesticated dogs perform well at social tasks, such as point following, due to an evolutionary process that selected for this ability (Hare et al., 2005; Hare et al., 2010; Hare & Tomasello, 2005; Riedel et al., 2008). One study found that 6, 8, 16, and 24-week-old puppies all performed well at point-following tasks, indicating that young pups with little human experience were successful, supporting the domestication hypothesis (Riedel et al., 2008). Other studies have further shown little to no age effects among 180 dogs, ages 2 months to 3.88 years in following human cues to find food in two-object choice tasks (Gácsi, McGreevy, Edina, & Miklósi, 2009b), and 32 dogs ages 9–24 weeks (Hare et al., 2002). Further, other domesticated species such as goats (Kaminski et al., 2005), horses (Maros et al., 2008), and cats (Miklósi et al., 2005) have all shown success on point following. Point following, however, does not appear limited to domesticated species, as nondomesticated species such as bats (Hall et al., 2011), elephants (Smet & Byrne, 2013), and sea lions (Malassis & Delfour, 2015) can perform well on point-following tasks.
Shelter dogs and pet dogs differ significantly in the amount of human interaction they experience. This presents a unique opportunity to test the effects of experience with humans on point-following abilities. Previous research has found, as predicted by the two-stage hypothesis, that shelter and kennel (research) dogs perform significantly worse on pointing and human-guided tasks than do pet dogs (D’Aniello et al., 2017a, b; Lazarowski & Dorman, 2015; Udell et al., 2008, 2010a; Zaine, Domeniconi, & Wynne, 2015). There have been some exceptions (e.g., Cunningham & Ramos, 2014; Hare et al., 2002), which found no difference between shelter/kennel dogs and pet dogs. One potential difference between these findings is that some studies use dogs with known limited history of human interaction, whereas other studies use shelter dogs with unknown histories, meaning that there might be variability in the background population of dogs.
Although shelter dogs tend to perform poorly on point-following tasks compared with pet dogs, it remains unclear why. Perhaps shelter dogs simply have not had the required exposure to humans and human behavior that is required to develop point following, as suggested by the two-stage hypothesis (Udell et al., 2010b). To address this question, we conducted a longitudinal assessment of shelter dogs as they receive typical experiences with humans, without explicit training on a point-following task. The aim of the present study was to track the development of point-following behavior in a cohort of dogs given an enriched experience around humans, as might be expected in a pet home. The development of point following in this cohort was compared with a control group that did not receive the enrichment (i.e., standard shelter housing). Dogs in the two groups were selected from the same shelter population. One group was randomly assigned to receive the enriched conditions, whereas the other group maintained the standard housing practices. To allow for repeated assessments, without directly training dogs to follow a point, we rewarded dogs for correct and incorrect responses on a standard two-choice point-following paradigm. This allowed us to test dogs twice weekly for 6.5 weeks without explicitly training the correct response (as both would lead to reward: see Supplemental Video 1 for an example demonstration of a dog being rewarded following an incorrect choice). We hypothesized that all dogs would initially perform poorly, but dogs that received enriched human experiences (e.g., going for walks, petting, playtime with human caretakers, obedience training) would develop point-following behavior typical of pet dogs, whereas similar dogs not receiving this enrichment would not develop this behavior.
Demonstration of one pointing trial in which the dog makes an incorrect response. Note both correct and incorrect responses are nondifferentially reinforced to prevent explicit training of point following. Pointing Data. csv shows the performance of each dogs across all session. Ncorrect indicates the number for correct responses. Complete indicates the number of trials to dog responded on (i.e., made a correct or incorrect response). Total refers to the number of trials attempted. PropCorrect refers to the proportion correct (Ncorrect/Total). Age refers to the dog’s estimated age in years. Pointing Analysis. R provides the R code for the analyses conducted in the manuscript. (MP4 1786 kb)
Method
Subjects
Twenty-three dogs were included in this study: seven pet dogs, nine shelter dogs housed in a shelter, and seven shelter dogs under enriched conditions. Two of the shelter dogs housed at the shelter were adopted during the study period (just before Sessions 8 and 10, respectively). Thus, two dogs from the standard shelter were excluded, yielding complete data for 21 dogs. Individual information on ages, source, and experimental condition of each dog is listed in Table 1. Note that ages are general best estimates provided by the shelter, which are generally known for owner surrenders, but are estimated by the shelter from dental evaluation on stray dogs.
Experimental design
To test whether point following is learned by dogs through typical interactions with humans, we compared the performance of three different populations of dogs that varied in the degree of human interactions they received.
Pet dogs
Pet dogs were owned dogs living in a home for at least the past 6 months. Dogs were tested in a room in the owner’s house. Pet dogs were tested in one session, as we did not expect their performance to increase over a period of time due to the consistency of interaction with humans.
Shelter dogs under standard conditions
Dogs were tested in a 3.5-m × 5-m room at a local shelter twice a week for a total of 13 sessions (spanned 6.5 weeks). Shelter dogs were tested longitudinally to match testing of the dogs in the enriched condition. Standard housing conditions for these dogs included kennels with immediate indoor and outdoor access. A caretaker fed them once a day and refilled their water twice a day. Their social interaction was limited to volunteers that are allowed to come twice a week. It is also important to note that one of the standard shelter dogs was taken to a home setting for a period of 3 days twice during this experiment.
Enriched shelter dogs
Enriched shelter dogs were tested in a 3.5-m × 6.5-m room at the housing facility twice a week for 13 sessions (spanned 6.5 weeks). Testing of this cohort began as soon as they arrived at the enriched facility from the same shelter as the dogs in the standard condition. Dogs had kennels with immediate indoor and outdoor access. They received twice daily walks and playtime twice a day along with behavioral training for future adoption (“Sit,” “Down,” “Stay”), and participation in other observational studies. Dogs therefore received direct human interaction typical of a pet home multiple times per day.
All dogs were required to be over 2 months and less than 14 years of age. This was done as Gácsi, Kara, Belényi, Topál, and Miklósi (2009a) has previously shown no effect of age in point-following performance of dogs older than 2 months. Older dogs were not selected to avoid potential health effects. Dogs in both the standard conditions and enriched conditions originated from the same population of dogs at the start of the study and were randomly assigned to one of the two conditions. Dogs were randomly assigned without respect to age given that previous research has found no effect (Gácsi et al., 2009a; Hare et al., 2002; Riedel et al., 2008). Pet dogs were recruited by word of mouth.
Procedure
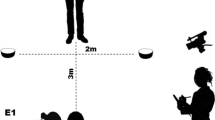
Dogs were tested on a two-object choice task in one brief session composed of two motivation trials, 10 testing trials, and two control trials (D’Aniello et al., 2017; Scandurra et al., 2019; Udell et al., 2008, 2010a; Udell, Ewald, Dorey, & Wynne, 2014; Udell, Hall, Morrison, Dorey, & Wynne, 2013; Zaine et al., 2015). Motivation trials were used to confirm the dog’s food motivation and that the dog was comfortable responding to the response objects (upside down dog bowls). Dog bowls were chosen as the response object on the basis that the participants were already familiar with this object. For motivation trials, the primary experimenter (E1) placed freely available food on top of the response object. The dog was given 30 s to approach and consume the food. One trial was run for each response side (D’Aniello et al., 2017; Udell et al., 2008, 2010a; Udell et al., 2014; Udell et al., 2013; Zaine et al., 2015). All dogs accepted food from the experimenter before testing started and approached the response objects to consume the food, ensuring that they were food motivated and did not display any signs of fear that would interfere with testing.
Experimental trials
E1 stood between two upside-down bowls, each 0.5 meters away from her, and there was a distance of at least 40 cm from the tip of the finger used to point and the respective bowl being pointed at. Experimenter 2 (E2) stood approximately 2.5 meters away from E1 with the dog equidistant from both bowls. To start a trial, E2 would face the dog toward E1. E1 would then call the dog by name to gain the dog’s attention, which consisted of waiting for eye contact before using a dynamic distal point to point to one of the two response objects. A dynamic distal point is accomplished from the standing position, extending the ipsilateral arm and hand into a traditional point in the direction of the food bowl, and held until the end of the trial. The tip of the finger of the experimenter was approximately 50 cm from the closest edge of the food bowl. After making eye contact with the participant, E1 would keep looking directly straight ahead. E2 would then release the participant. E2 was naïve to which bowl was the correct choice, as the dog was rewarded for either response. If the participant approached within 10 cm or touched either bowl within 30 s, E2 would call out the number ascribed to the bowl the dog chose, and E1 would place a palatable commercial dog treat on top of the respective bowl for the dog to eat (the same treat used in the motivation assessment). To prevent explicit training of point following, dogs were rewarded for responding to either bowl regardless of whether E1 pointed to it. Thus, nondifferential reinforcement was used to probe changes in the spontaneous response of the participants without explicit training of the task. If dogs were tested such that food was only available for point following, we would expect all animals capable of operant conditioning to learn this task, preventing a longitudinal analysis of the effects of human experience. If a dog did not respond to either bowl within 30s, a “no choice” was scored and counted as an incorrect response. “No choices” were counted as incorrect as it indicated that the dog did not respond appropriately to the point. This analysis falls in line with the procedures used by D’Aniello et al. (2017), Udell et al. (2008, 2010a), Udell et al. (2014), Udell et al. (2013), and Zaine et al. (2015), but differs from that of other authors (e.g., Colbert-White, Tullis, Andresen, Parker, & Patterson, 2018; Hare et al., 2002; Lazarowski & Dorman, 2015; Miklósi et al., 1998; Oliva, Rault, Appleton, & Lill, 2015).
The position E1 pointed toward was pseudorandomly determined such that points were made equally often to the left and right side. At the end of pointing trials, two control trials in which the experimenter did not point, but the participant was still released by E2 to approach either bowl, were conducted. The correct bowl was predetermined, but similar to experimental trials, a response to either bowl was rewarded. Control trials evaluated whether E1 was giving unintentional cues. This is the same procedure used by past research (D’Aniello et al., 2017; Udell et al., 2008, 2010a; Udell et al., 2014; Udell et al., 2013; Zaine et al., 2015). If dogs were using unintentional cues, we expected dogs to have an above-chance performance on control trials. Otherwise, we expected performance to be at or below chance (due to dogs not responding if no cue is given).
Statistical analysis
To evaluate differences between enriched and standard shelter dogs, we used a generalized linear mixed-effects model from the lme4 package in R (R Core Team, 2019). The dependent variable was the number correct (out of 10) assuming a binomial distribution and a logistic link. We included fixed effects of enriched versus nonenriched, session number (treated as a continuous variable), their interaction, and age. A random intercept was included for each dog. To test for significance, a model was fit with and without the term of interest and compared. False discovery rate adjusted post hoc tests were computed using the lsmeans package (Length, 2016) to compare the enriched and standard dogs’ performance during the first and last sessions. Although session number was treated as a continuous variable, post hoc comparisons were made with reference to the first and last session between enriched and standard shelter dogs using the lsmeans package, as this provided more relevant comparisons in the change in performance than simply comparing the two groups at the mean session value. Raw data and R code are provided as supplementary files.
Results
Figure 1 shows the performance of the enriched and standard shelter dogs across all sessions. Both populations showed identical performance in the first session, which was very low, largely due to a failure to respond and approach the containers. This is not unexpected given that dogs were randomly assigned from the same population. Both groups quickly showed increases in performance to chance performance levels, as they rapidly started responding to the containers; however, only enriched dogs continued to increase, with performances significantly exceeding chance. Comparing a generalized linear mixed-effect model with and without an interaction term indicates a significant Group × Session interaction (χ2 = 6.57, p = .01). Comparing a model with and without an age term did not significantly improve model fit (χ2 = 1.36, p = .24), indicating no significant effect of age differences. When controlling for age, post hoc test for Session 1 indicated that enriched and standard housed dogs do not differ (z = 1.33, p = .18); however, enriched dogs significantly outperformed standard housed dogs in Session 13 (z = 3.52, p < .001). Thus, when dogs are initially randomly assigned into our two populations, performance is similar; however, enriched dogs show significant gains in performance compared with dogs remaining in standard conditions.

Performance across sessions. Solid line indicates mean performance of enriched dogs. Dashed line shows performance of nonenriched dogs. Error bars show the 95% confidence interval. Horizontal line shows chance performance
To evaluate how standard housed dogs and enriched dogs compare with pet dogs, we conducted a generalized linear model. We evaluated performance of enriched and standard dogs in the last session and performance to that of pet dogs in the single session tested (see Fig. 2). There was a significant effect of enrichment type (χ2 = 11.44, p < .01), but no effect of age (χ2 = 0.87, p = .35). When accounting for age in the model, post hoc tests indicate that enriched dogs and pet dogs outperformed standard housed shelter dogs (enriched vs. standard: z = 3.24, p < .01; pet vs. standard: z = 2.70, p = .01). Importantly, pet dogs and enriched dogs performed similarly (z = 0.65, p = .52).

Performance of enriched shelter dogs, nonenriched shelter dogs, and pet dogs. Dashed line indicates chance performance. Error bars show the 95% confidence interval. Beginning enriched shows the first session for enriched dogs. Beginning standard shows the first session for standard house dogs. End enriched shows the last session for enriched dogs. End standard shows the last session for standard housed dogs
One important consideration is whether differences in the number of “no choice” responses across groups strongly influenced performance. Figure 3 shows the data from the final session for the standard housed dogs, enriched dogs, and pet dogs when scoring no choices as incorrect (left graph) and when removing no choices from the data (right graph) and only accounting for trials in which a response was made. This improved the performance of nonenriched dogs to slightly above chance, but the conclusions of our group comparisons remain unchanged. Overall, there was a main effect of group (χ2 = 10.97, p < .01), and no effect of age (χ2 = 1.17, p < .28) on point-following accuracy. Post hoc tests again confirm that enriched dogs and pet dogs outperform the standard shelter dogs (enriched vs. standard: z = 2.15, p = .047; pet vs. standard: z = 3.03, p < .01). Lastly, enriched dogs performed at the same level as pet dogs (z = 1.08, p = .28). Thus, regardless of whether a lack of response was scored as incorrect or missing, the conclusions remain unchanged and suggest the group differences are not related to a difference to no choice responses alone.

Performance on the last session for enriched and standard housed dogs with performance of pet dogs. Bars show the mean, and error bars show the 95% confidence interval. Dashed line show chance performance. Left graph shows the proportion correct counting no choices as incorrect responses (all 10 trials scored). Right graph removes all no choice responses and calculates proportion correct out of total trials completed
Controls
Performance did not appear to be controlled by unintentional cuing that may have arisen in the experiment. If dogs were using unintentional cues, we expected dogs to perform at above chance levels (>50%). If dogs were not using other cues, we expected performance to be at or below chance, as many dogs would simply not respond if the cue were not given. Performance on controls averaged less than 50% for all groups.
Discussion
The results show that when controlled for age, both standard and enriched dogs were at similar point-following performance levels at the start of the experiment. This was expected, given that both groups of dogs were randomly assigned from the same source, and that minor differences in the shelter estimated age would not likely lead to significant differences alone, given that several studies have tested dogs over the range of 2 months to 3.88 years (adult group mean = 3.88 years; Gácsi et al., 2009a), 9–24 weeks (Hare et al., 2002), and 6–24 weeks (Riedel et al., 2008) and have not previously shown significant age effects. In fact, the only age differences found in pointing studies have been restricted between 8 and 24 weeks of age. One study found that young pups outperformed older pups between ages of 8 and 12 weeks (Zaine et al., 2015) and the other found that older pups outperformed younger pups between 9 and 24 weeks of age (Dorey et al., 2010). Further, we included age as a covariate in all of our analyses and found no statistically significant effect.
Despite starting at identical performance levels, over the course of the 6 weeks, the standard housed dogs remained at chance levels to perhaps slightly above chance (when excluding no-choice responses), whereas the enriched dogs’ performance improved to the point that it was comparable to the pet dogs and exceeded that of the standard shelter dogs. The improvement in performance of the enriched dogs is consistent with our prediction of an increase in correctly followed points among the enriched dogs as a result of enriched conditions that include more exposure to humans. This adds an important finding to the literature, as this is the first time in which the development of point-following behavior has been tracked as dogs gain more experience with humans. Our results suggest that within 7 weeks, shelter dogs that previously performed poorly on a point-following task perform the same as pet dogs. This highlights an important finding that over a relatively short period of time, dogs can learn to follow points through typical “everyday” interactions with a human.
These results suggest that learning is an important component to point following in canines and that typical experiences with humans can lead to the development of the behavior, as suggested by the two-stage hypothesis. Further, this runs counter to the domestication hypothesis given the increase in point-following performance with more human experience. This will, however, require further testing in which the specific gestures and mannerisms used by caretakers is strictly controlled and manipulated to provide a more robust analysis of what specific experiences with humans control changes in performance.
We modeled our test procedures after the two-object choice task done in previous experiments (D’Aniello et al., 2017; Scandurra et al., 2019; Udell et al., 2008, 2010a; Udell et al., 2014; Udell et al., 2013; Zaine et al., 2015). Recently, discussions have been raised concerning how the food rewards are specifically delivered (whether hidden or delivered after a choice; see Hare et al., 2010; Udell et al., 2008; Udell & Wynne, 2010). We selected the procedure that inherently controls for odor stimuli and potential food baiting cues, and is consistent with a wide body of the literature (D’Aniello et al., 2017; Scandurra et al., 2019; Udell et al., 2008, 2010a; Udell et al., 2014; Udell et al., 2013; Zaine et al., 2015), though it differs from other authors (e.g., Colbert-White et al., 2018; Hare et al., 2002; Hare et al., 2005; Hare & Tomasello, 2005; Miklósi et al., 1998, 2003, 2005).
One consideration is whether the enriched dogs showed greater motivation to respond to the experimenter than the standard shelter dogs did. We view this potential difference as unlikely, given that all dogs completed all pretraining trials (i.e., motivation assessment) trials indicating they would readily approach the experimenter and containers when the dog could see food available. Further, our coding of no choices as incorrect responses is consistent with a large portion of the literature (D’Aniello et al., 2017; Udell et al., 2008, 2010a; Udell et al., 2014; Udell et al., 2013; Zaine et al., 2015), as a no-choice score can reflect a dog failing to respond to the point appropriately. We choose this scoring scheme because we wanted strictest criterion for whether a dog was or was not responding to the point. Importantly, however, this study is unable to directly resolve the mechanism of the enriched dogs’ improvement. Although dogs may have learned to follow limbs through increased interaction with humans, it is also possible that dogs became more “comfortable” with responding to a point with increased human interaction. We find the latter explanation less likely, however, because all dogs could obtain reward equally by approaching the pointed toward, or nonpointed toward, container, so it is unclear why specifically correct choices would increase only for enriched dogs. Further, when we remove all no-choice trials in the final session, we find the same group differences, suggesting that enriched dogs showed increased propensity for correct responses, not just simply increases in response rates to the human experimenter.
In addition, we need to consider the importance of ostensive cues before pointing, such as eye contact and addressing the dog’ name, as it has been shown that ostensive cues can modify a dog’s behavior (Duranton, Range, & Virányi, 2017; Kaminski, Schulz, & Tomasello, 2012; Tauzin, Csík, Kis, Kovács, & Topál, 2015a; Tauzin, Csík, Kis, & Topál, 2015b; Téglás, Gergely, Kupán, Miklósi, & Topál, 2012; Wallis et al., 2015). In particular, previous research has shown that the dog better attends to the experimenter when more ostensive cues are used. In this study, eye contact was established to ensure the dog was attending to E1, then E1 continued to stare straight ahead, not looking at the dog or bowls. Thus, the present study did include some ostensive cues, which has been previously found to increase dog performance (Duranton et al., 2017; Kaminski et al., 2012; Tauzin et al., 2015b; Téglás et al., 2012; Wallis et al., 2015). Future research could evaluate if differences in the experience with humans influences the effect of ostensive cues on performance.
There are several further important limitations to the present study. First, given that we used shelter populations, it is unclear whether dogs in both the enriched and standard shelter dog cohorts received a comparable amount of exposure to humans before the experiment. Given, however, that both groups responded identically in the first sessions and that dogs came from the same population suggests this was not a large limiting factor. Another weakness to this study is that one of the dogs was taken home to be fostered for two nights in the middle of our study. We cannot know for certain if, or how much, this affected the subject’s performance; however, we would like to note that the subject’s performance did not change noticeably as a result of being in a human’s home for a short period of time. Given that our results from the enriched conditions suggests that 6–7 weeks of experiences with humans was necessary to reach pet dogs levels. Further, we reanalyzed our data excluding this one participant and found that this had no effect on any of our statistical conclusions (i.e., p values originally <.05 remained <.05 after exclusion).
Lastly, the rooms used to test the enriched, nonenriched, and pet dogs were different given that the tests occurred in different facilities and owner’s homes. The primary concern and reason for choosing the used spaces was to ensure the dogs would become familiar with the space by always having access to the same space, and that there was limited extraneous noise and distractions. Despite the unavoidable limitation of different testing environments, such as the 3.5-m × 6.5-m room for the enriched shelter dogs and 3.5-m × 5-m room for the standard shelter dogs, all of the dogs successfully completed pretraining and started at an identical point of performance. It is therefore unlikely that the present results are related to the testing room.
Further research is needed to determine whether additional social behaviors change with human enrichment, such as gazing. We did not evaluate gazing in the present study, but rather had the experimenter look straight ahead. This was to present a single cue to the dog rather than a compound cue, but would be an interesting future direction, because canine gazing has previously been shown to be amenable ontogenetic experiences (Barrera, Mustaca, & Bentosela, 2011; D’Aniello & Scandurra, 2016; D’Aniello, Scandurra, Prato-Previde, & Valsecchi, 2015; Scandurra, Prato-Previde, Valsecchi, Aria, & D’Aniello, 2015; Topál, Miklósi, & Csányi, 1997).
Together, these results show that social enrichment has a significant impact on point-following behavior in shelter dogs. Enriched conditions with more human–animal interaction led to marked changes in point-following performance; whereas, dogs that started at a comparable level showed no such changes under standard housing conditions. Further, enriched conditions led to dogs showing equivalent levels of performance as pet dogs within 7 weeks.
References
Barrera, G., Mustaca, A., & Bentosela, M. (2011). Communication between domestic dogs and humans: Effects of shelter housing upon the gaze to the human. Animal Cognition, 14(5), 727–734. https://doi.org/10.1007/s10071-011-0407-4
Behne, T., Carpenter, M., & Tomasello, M. (2005). One-year-olds comprehend the communicative intentions behind gestures in a hiding game. Developmental Science, 8(6), 492–499. https://doi.org/10.1111/j.1467-7687.2005.00440.x
Call, J., Hare, B., & Tomasello, M. (1998). Chimpanzee gaze following in an object-choice task. Animal Cognition, 1(2), 89–99. https://doi.org/10.1007/s100710050013
Colbert-White, E. N., Tullis, A., Andresen, D. R., Parker, K. M., & Patterson, K. E. (2018). Can dogs use vocal intonation as a social referencing cue in an object choice task? Animal Cognition, 21(2), 253–265. https://doi.org/10.1007/s10071-018-1163-5
Cunningham, C. L., & Ramos, M. F. (2014). Effect of training and familiarity on responsiveness to human cues in domestic dogs (Canis familiaris). Animal Cognition, 17(3), 805–814. https://doi.org/10.1007/s10071-013-0714-z
D’Aniello, B., & Scandurra, A. (2016). Ontogenetic effects on gazing behaviour: A case study of kennel dogs (Labrador Retrievers) in the impossible task paradigm. Animal Cognition, 19(3), 565–570. https://doi.org/10.1007/s10071-016-0958-5
D’Aniello, B., Scandurra, A., Prato-Previde, E., & Valsecchi, P. (2015). Gazing toward humans: A study on water rescue dogs using the impossible task paradigm. Behavioural Processes, 110, 68–73. https://doi.org/10.1016/j.beproc.2014.09.022
D’Aniello, B., Alterisio, A., Scandurra, A., Petremolo, E., Iommelli, M. R., & Aria, M. (2017). What’s the point? Golden and Labrador retrievers living in kennels do not understand human pointing gestures. Animal Cognition, 20(4), 777–787. https://doi.org/10.1007/s10071-017-1098-2
Dorey, N. R., Udell, M. A. R., & Wynne, C. D. L. (2010). When do domestic dogs, Canis familiaris, start to understand human pointing? The role of ontogeny in the development of interspecies communication. Animal Behaviour, 79(1), 37–41. https://doi.org/10.1016/j.anbehav.2009.09.032
Duranton, C., Range, F., & Virányi, Z. (2017). Do pet dogs (Canis familiaris) follow ostensive and non-ostensive human gaze to distant space and to objects? Royal Society Open Science, 4(7). https://doi.org/10.1098/rsos.170349
Gácsi, M., Kara, E., Belényi, B., Topál, J., & Miklósi, Á. (2009a). The effect of development and individual differences in pointing comprehension of dogs. Animal Cognition, 12(3), 471–479. https://doi.org/10.1007/s10071-008-0208-6
Gácsi, M., McGreevy, P., Edina, K., & Miklósi, Á. (2009b). Effects of selection for cooperation and attention in dogs. Behavioral and Brain Functions, 5(31). Retrieved from https://behavioralandbrainfunctions.biomedcentral.com/articles/10.1186/1744-9081-5-31
Grassmann, S., & Tomasello, M. (2010). Young children follow pointing over words in interpreting acts of reference. Developmental Science, 13(1), 252–263. https://doi.org/10.1111/j.1467-7687.2009.00871.x
Hall, N. J., Udell, M. A. R., Dorey, N. R., Walsh, A. L., & Wynne, C. D. L. (2011). Megachiropteran bats (Pteropus) utilize human referential stimuli to locate hidden food. Journal of Comparative Psychology, 125(3), 341–346. https://doi.org/10.1037/a0023680
Hare, B., & Tomasello, M. (2005). Human-like social skills in dogs? Trends in Cognitive Sciences, 9(9), 439–444. https://doi.org/10.1016/j.tics.2005.07.003
Hare, B., Brown, M., Williamson, C., & Tomasello, M. (2002). The domestication of social cognition in dogs. Science, 298(5598), 1634–1636.
Hare, B., Plyusnina, I., Ignacio, N., Schepina, O., Stepika, A., Wrangham, R., & Trut, L. (2005). Social cognitive evolution in captive foxes is a correlated by-product of experimental domestication. Current Biology, 15(3), 226–230. https://doi.org/10.1016/j.cub.2005.01.040
Hare, B., Rosati, A., Kaminski, J., Bräuer, J., Call, J., & Tomasello, M. (2010). The domestication hypothesis for dogs’ skills with human communication: A response to Udell et al. (2008) and Wynne et al. (2008). Animal Behaviour, 79(2), e1–e6. https://doi.org/10.1016/j.anbehav.2009.06.031
Hodges, L. E., Özçalışkan, Ş., & Williamson, R. (2018). Type of iconicity influences children’s comprehension of gesture. Journal of Experimental Child Psychology, 166, 327–339. https://doi.org/10.1016/j.jecp.2017.08.009
Hopkins, W. D., Russell, J., McIntyre, J., & Leavens, D. A. (2013). Are chimpanzees really so poor at understanding imperative pointing? Some new data and an alternative view of canine and ape social cognition. PLOS ONE, 8(11), e79338. https://doi.org/10.1371/journal.pone.0079338
Kaminski, J., Riedel, J., Call, J., & Tomasello, M. (2005). Domestic goats, Capra hircus, follow gaze direction and use social cues in an object choice task. Animal Behaviour, 69(1), 11–18. https://doi.org/10.1016/j.anbehav.2004.05.008
Kaminski, J., Schulz, L., & Tomasello, M. (2012). How dogs know when communication is intended for them. Developmental Science, 15(2), 222–232. https://doi.org/10.1111/j.1467-7687.2011.01120.x
Kirchhofer, K. C., Zimmermann, F., Kaminski, J., & Tomasello, M. (2012). Dogs (Canis familiaris), but not chimpanzees (Pan troglodytes), understand imperative pointing. PLOS ONE, 7(2), e30913–e30913. https://doi.org/10.1371/journal.pone.0030913
Lazarowski, L., & Dorman, D. C. (2015). A comparison of pet and purpose-bred research dog (Canis familiaris) performance on human-guided object-choice tasks. Behavioural Processes, 110, 60–67. https://doi.org/10.1016/j.beproc.2014.09.021
Length, R. (2016). Least-Squares Means: The R Package lsmeans. Journal of Statistical Software, 69(1), 1-33. https://doi.org/10.18637/jss.v069.i01
Malassis, R., & Delfour, F. (2015). Sea lions’ (Zalophus californianus ) use of human pointing gestures as referential cues. Learning & Behavior, 43(2), 101–112. https://doi.org/10.3758/s13420-014-0165-7
Maros, K., Gácsi, M., & Miklósi, A. (2008). Comprehension of human pointing gestures in horses (Equus caballus). Animal Cognition, 11(3), 457–466. https://doi.org/10.1007/s10071-008-0136-5
Miklósi, Á., Polgárdi, R., Topál, J., & Csányi, V. (1998). Use of experimenter-given cues in dogs. Animal Cognition, 1(2), 113–121. https://doi.org/10.1007/s100710050016
Miklósi, Á., Kubinyi, E., Topál, J., Gácsi, M., Virányi, Z., & Csányi, V. (2003). A Simple Reason for a Big Difference: Wolves Do Not Look Back at Humans, but Dogs Do. Current Biology, 13(9), 763–766. https://doi.org/10.1016/S0960-9822(03)00263-X
Miklósi, Á., Pongrácz, P., Lakatos, G., Topál, J., & Csányi, V. (2005). A comparative study of the use of visual communicative signals in interactions between dogs (Canis familiaris) and humans and cats (Felis catus) and humans. Journal of Comparative Psychology, 119(2), 179–186. https://doi.org/10.1037/0735-7036.119.2.179
Oliva, J. L., Rault, J.-L., Appleton, B., & Lill, A. (2015). Oxytocin enhances the appropriate use of human social cues by the domestic dog (Canis familiaris) in an object choice task. Animal Cognition, 18(3), 767–775. https://doi.org/10.1007/s10071-015-0843-7
R Core Team (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Riedel, J., Schumann, K., Kaminski, J., Call, J., & Tomasello, M. (2008). The early ontogeny of human–dog communication. Animal Behaviour, 75(3), 1003–1014. https://doi.org/10.1016/j.anbehav.2007.08.010
Scandurra, A., Prato-Previde, E., Valsecchi, P., Aria, M., & D’Aniello, B. (2015). Guide dogs as a model for investigating the effect of life experience and training on gazing behaviour. Animal Cognition, 18(4), 937–944. https://doi.org/10.1007/s10071-015-0864-2
Scandurra, A., Alterisio, A., Di Cosmo, A., D'Ambrosio, A., & D'Aniello, B. (2019). Ovariectomy impairs socio-cognitive functions in dogs. Animals: An Open Access Journal from MDPI, 9(2) https://doi.org/10.3390/ani9020058
Smet, A. F., & Byrne, R. W. (2013). African elephants can use human pointing cues to find hidden food. Current Biology, 23(20), 2033–2037. https://doi.org/10.1016/j.cub.2013.08.037
Tauzin, T., Csík, A., Kis, A., Kovács, K., & Topál, J. (2015a). The order of ostensive and referential signals affects dogs’ responsiveness when interacting with a human. Animal Cognition, 18(4), 975–979. https://doi.org/10.1007/s10071-015-0857-1
Tauzin, T., Csík, A., Kis, A., & Topál, J. (2015b). What or where? The meaning of referential human pointing for dogs (Canis familiaris). Journal of Comparative Psychology, 129(4), 334–338. https://doi.org/10.1037/a0039462
Téglás, E., Gergely, A., Kupán, K., Miklósi, Á., & Topál, J. (2012). Dogs’ gaze following is tuned to human communicative signals. Current Biology, 22(3), 209–212. https://doi.org/10.1016/j.cub.2011.12.018
Thoermer, C., & Sodian, B. (2001). Preverbal infants’ understanding of referential gestures. First Language, 21(63), 245–264. https://doi.org/10.1177/014272370102106303
Topál, J., Miklósi, Á., & Csányi, V. (1997). Dog–human relationship affects problem solving behavior in the dog. Anthrozoös, 10(4), 214–224. https://doi.org/10.2752/089279397787000987
Udell, M. A. R., & Wynne, C. D. L. (2010). Ontogeny and phylogeny: Both are essential to human-sensitive behaviour in the genus Canis. Animal Behaviour, 79(2), e9–e14. https://doi.org/10.1016/j.anbehav.2009.11.033
Udell, M. A. R., Dorey, N. R., & Wynne, C. D. L. (2008). Wolves outperform dogs in following human social cues. Animal Behaviour, 76(6), 1767–1773. https://doi.org/10.1016/j.anbehav.2008.07.028
Udell, M. A. R., Dorey, N. R., & Wynne, C. D. L. (2010a). The performance of stray dogs (Canis familiaris) living in a shelter on human-guided object-choice tasks. Animal Behaviour, 79(3), 717–725. https://doi.org/10.1016/j.anbehav.2009.12.027
Udell, M. A. R., Dorey, N. R., & Wynne, C. D. L. (2010b). What did domestication do to dogs? A new account of dogs’ sensitivity to human actions. Biological Reviews of the Cambridge Philosophical Society, 85(2), 327–345. https://doi.org/10.1111/j.1469-185X.2009.00104.x
Udell, M. A. R., Hall, N. J., Morrison, J., Dorey, N. R., & Wynne, C. D. L. (2013). Point Topography and Within-Session Learning Are Important Predictors of Pet Dogs’ (Canis lupus familiaris) Performance on Human Guided Tasks. Revista Argentina de Ciencias del Comportamiento ( RACC ), 5(2), 1.
Udell, M. A. R., Ewald, M., Dorey, N. R., & Wynne, C. D. L. (2014). Exploring breed differences in dogs (Canis familiaris): Does exaggeration or inhibition of predatory response predict performance on human-guided tasks? Animal Behaviour, 89, 99–105. https://doi.org/10.1016/j.anbehav.2013.12.012
Virányi, Z., Gácsi, M., Kubinyi, E., Topál, J., Belényi, B., Ujfalussy, D., & Miklósi, Á. (2008). Comprehension of human pointing gestures in young human-reared wolves (Canis lupus) and dogs (Canis familiaris). Animal Cognition, 11(3), 373. https://doi.org/10.1007/s10071-007-0127-y
Wallis, L. J., Range, F., Müller, C. A., Serisier, S., Huber, L., & Virányi, Z. (2015). Training for eye contact modulates gaze following in dogs. Animal Behaviour, 106, 27–35. https://doi.org/10.1016/j.anbehav.2015.04.020
Wynne, C. D. L., Udell, M. A. R., & Lord, K. A. (2008). Ontogeny’s impacts on human–dog communication. Animal Behaviour, 76(4), e1–e4. https://doi.org/10.1016/j.anbehav.2008.03.010
Zaine, I., Domeniconi, C., & Wynne, C. D. L. (2015). The ontogeny of human point following in dogs: When younger dogs outperform older. Behavioural Processes, 119, 76–85. https://doi.org/10.1016/j.beproc.2015.07.004
Acknowledgements
We would like to thank Annie Steffy and Cristina Lawniczak for their work on this project. We also would like to thank the members of the Texas Tech University Canine Olfaction Lab for their help with the dogs used in this study as well as the Texas Tech University Undergraduate Research Scholars Program supported by the CH and Helen Jones Foundations. Lastly, we would like to thank our shelter partners for their support and assistance.
Open practices statements
Study data and R code for analysis is available as a supplementary material. This study was not preregistered.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical statement
All procedures with animals were approved by the Texas Tech University Animal Care and Use Committee (Protocol # 16111-12).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Jarvis, T., Hall, N.J. Development of point following behaviors in shelter dogs. Learn Behav 48, 335–343 (2020). https://doi.org/10.3758/s13420-020-00415-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-020-00415-8