
Abstract
To date, the emotion regulation literature has focused primarily on the down-regulation of negative emotion, with far fewer studies interrogating the mechanisms at work in positive emotion regulation. This body of work has suggested that nonaffective mechanisms, such as cognitive load have a role to play in reducing emotional response. For example, the late positive potential (LPP), which tracks attention to salient stimuli, is reduced when task-irrelevant negative and neutral stimuli are presented under high compared with low working memory load. Using positive stimuli, working memory load has been shown to reduce the LPP elicited by positive words and faces but has not previously been shown to modulate the LPP elicited by positive scenes. Emotional scenes are the predominant type of stimuli used in the broader emotion regulation literature, are more arousing than faces, and have been shown to more strongly modulate the LPP. Here, 41 participants performed a working memory task interspersed with the presentation of positive and neutral scenes, while electroencephalography was recorded. Results showed that the LPP was increased for positive compared with neutral pictures and reduced on high-load compared to low-load trials. Working memory performance was worse on high-load compared with low-load trials, although it was not significantly correlated with the LPP, and picture type did not affect working memory performance. Results bridge to the willful emotion regulation literature to increase understanding of the mechanisms underlying positive emotion regulation, which has been relatively unexamined.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Reducing or limiting positive emotion may be beneficial in certain contexts (Kalokerinos et al., 2014), such as when positive emotion motivates actions that are not in line with a person’s goals. Nonetheless, most emotion regulation work has focused on the downregulation of negative emotion. This work has shown that it is possible to reduce attention toward negative stimuli through a variety of willful emotion regulation techniques (Buhle et al., 2014), but also through nonaffective tasks that are cognitively demanding, such as those that engage working memory (MacNamara, Ferri, et al., 2011a). As such, this work has provided insight into the mechanism(s) that may underlie the downregulation of emotional response and has demonstrated that reducing emotional response is possible even in the absence of a willful attempt. By comparison, only a handful of studies have examined how cognitively demanding tasks affect the processing of positive stimuli (Kopf et al., 2013; Van Dillen & Derks, 2012) and gaps in the literature (e.g., no investigation of load-related modulation of electrocortical response using positive scenes) limit comparison with other work. Filling these gaps would facilitate a more thorough understanding of cognitive-load-related regulation of response to positive stimuli and would help to address the relative neglect of positive emotion regulation in the field.
Event-related potentials, such as the late positive potential (LPP), provide a useful means of assessing the effects of emotion regulation and cognitive load on the processing of emotional stimuli. The LPP begins approximately 300 ms after stimulus onset, is thought to measure stimulus salience, and is larger for positive and negative compared with neutral stimuli (Cuthbert et al., 2000; Hajcak, MacNamara, et al., 2010b). The LPP is sensitive to both intrinsic (e.g., personal relevance; Tacikowski & Nowicka, 2010) and extrinsic (Schindler & Straube, 2020; Schupp et al., 2007; Weinberg et al., 2012) modulations of stimulus significance and is believed to measure the motivational salience of stimuli. Its neural generators are widespread and include the amygdala, visual cortex, insula, and temporal cortex (Liu et al., 2012).
Willful emotion regulation techniques, such as cognitive reappraisal have been found to reduce the LPP to negative pictures (Hajcak & Nieuwenhuis, 2006; Moran et al., 2013; Moser et al., 2006; Parvaz et al., 2012). The down-regulation of negative emotion (e.g., via reappraisal) has been shown to engage the dorsolateral prefrontal cortex (dlPFC, Buhle et al., 2014), a brain region that shows a reciprocal relationship with activation in regions involved in the generation of emotional response, such as the amygdala (Banks et al., 2007). Structurally, the dlPFC has few direct connections to brain regions involve in emotion generation, such as the amygdala, but may interact with these regions via indirect connection (e.g., orbitofrontal cortex, Amaral & Price, 1984; Cavada, 2000; Ray & Zald, 2012). Therefore, down-regulation of emotional response via reappraisal might be attributed, in part, to an obligatory influence of the lateral prefrontal cortex on brain regions involved in the generation of emotional response (see also Hajcak, Anderson, et al., 2010a). In our prior work, we tested the potential for a task known to activate the dlPFC (Smith et al., 1998)—i.e., a working memory task—to reduce the processing of emotional pictures. Participants performed a letter recall task interspersed with the presentation of task-irrelevant negative and neutral pictures. Results showed that pictures that were presented on high-load (6 letters) compared with low-load (2 letters) trials elicited smaller LPPs (MacNamara, Ferri, et al., 2011a), suggesting that participants allocate less attention to task irrelevant negative and neutral pictures as cognitive load increases. This effect has been replicated several times for negative pictorial scenes (MacNamara et al., 2019; MacNamara & Proudfit, 2014) and has been shown for faces (MacNamara et al., 2012).
In contrast to the negative emotion regulation literature, there have been few investigations of positive emotion regulation or its underlying mechanisms. Reappraisal, the “gold standard” for regulation of negative emotion, has been shown to be effective for positive pictures (Kim & Hamann, 2007), although depending on stimulus intensity, it may be less effective than other strategies (Shafir et al., 2018). To date, three studies have examined the effect of working memory load on the processing of positive stimuli. Using fMRI, Erk et al. (2007) found that working memory load reduced activation in the right amygdala for negative scenes and in the left ventral striatum for positive scenes (Erk et al., 2007). These results suggest that working memory load attenuates the processing of both positive and negative emotional scenes, but it may do so via different neurocircuit mechanisms. In addition to this fMRI work, two studies investigated the effect of working memory load on positive stimuli using the LPP. In one study, Kopf et al. (2013) used an affective n-back task to assess the effect of working memory load on the LPP elicited by positive, negative, and neutral words. They found that some levels of working memory load reduced the LPP elicited by positive words, but not others, with more consistent effects observed for working memory modulation of the LPP elicited by negative words. In another study, Van Dillen and Derks (2012) found that working memory load reduced the LPP to angry and happy faces, but this effect was stronger for angry faces. However, because this study lacked a neutral “control” condition, it is difficult to know whether the observed results reflect reduced emotional differentiation under high working memory load or a stronger effect of working memory load on negative versus positive stimuli.
Together, this prior work suggests that working memory load modulates the LPP to positive words and faces but raises some questions about the robustness of this effect, and leaves untested whether working memory load modulates the LPP elicited by positive scenes (e.g., IAPS; Lang et al., 2008). This omission limits comparison with a broader body of work, particularly the emotion regulation literature, which has primarily used emotional scenes (Buhle et al., 2014; Picó-Pérez et al., 2017). Moreover, prior work indicates that emotional scenes are more arousing (Britton et al., 2006) and may modulate the LPP more robustly than emotional faces (Thom et al., 2014), suggesting that they might be harder to ignore/less affected by working memory load. Therefore, although we suspected that working memory load would reduce the LPP elicited by positive scenes, we set out to test this question directly.
We used a modified version of the working memory paradigm used in our prior work (MacNamara et al., 2012, 2019; MacNamara, Ferri, et al., 2011a; MacNamara & Proudfit, 2014) to determine whether working memory load would reduce the LPP elicited by positive scenes. We focused our analyses on the LPP because, as in our prior work using this task, we were interested in the effects of working memory load on the sustained processing and motivational significance of positive pictures. In addition, we wanted to be able to relate our work to prior ERP work on emotion regulation, which has primarily examined the LPP (Hajcak & Nieuwenhuis, 2006; MacNamara et al., 2009; MacNamara, Ochsner, et al., 2011b; Moran et al., 2013; Moser et al., 2006).
Based on prior work that used positive words and faces, we predicted that working memory load would reduce the positive picture-elicited LPP (Kopf et al., 2013; Van Dillen & Derks, 2012). Also, based on prior work that found that positive scenes were associated with improved working memory performance (compared to neutral scenes; Erk et al., 2007), as well as other work suggesting that positive mood might facilitate performance on working memory tasks (Storbeck & Maswood, 2016; Yang et al., 2013), we expected that positive pictures would improve letter recall on high-load trials. This prediction was in contrast to our prior work, in which negative pictures were found to interfere with working memory performance (MacNamara, Ferri, et al., 2011a).
Method
Participants
Participants were 41 undergraduates who completed the experiment for course credit (27 females; age M = 19.82 years, SD = 1.94). Sample size was decided according to the a priori decision to run the study for one semester. Study procedures were in compliance with the Helsinki Declaration of 1975 (as revised in 1983) and were approved by the Texas A&M University institutional review board.
Stimulus materials
Sixty positive pictures (e.g., cute animals, erotic scenes) and 60 neutral pictures (e.g., household objects, neutral faces) were selected from the International Affective Picture System (IAPS: Lang et al., 2008) and Emotional Picture Set (Wessa et al., 2010).Footnote 1 Compared with neutral pictures, normative ratings for positive pictures were more pleasant, t(106.49) = 13.45, p < 0.001 (positive: M = 6.70, SD = 0.74; neutral, M = 5.12, SD = 0.53) and more arousing, t(93.87) = 13.62, p < 0.001 (positive: M = 4.77, SD = 0.87; neutral, M = 3.01, SD = 0.50).
Letter strings were the same as in the original version of the task that used negative and neutral pictures (MacNamara, Ferri, et al., 2011a). In brief, letter strings were created using a random number generator (Reed, 2002). Vowels were not included in the strings; there were 60 two-consonant strings and 60 six-consonant strings (Ashcraft & Kirk, 2001).
Procedure
Participants performed a letter recall task adapted from prior work (MacNamara, Ferri, et al., 2011a) while continuous EEG was recorded. Each trial began with the presentation of either a two-letter (low-load) or six-letter (high-load) string that was displayed for 5,000 ms. This was followed by a positive or neutral picture that was presented for 2,000 ms. Following picture offset, the text “What were the letters? (then press enter):” was presented in the center of the screen. Participants were instructed to use the keyboard to enter the letters they had viewed at the beginning of the trial and to enter them in the same order they originally appeared. They were also told that they could use the backspace key to erase a letter if they made a mistake. Participants were told to enter letters with only one finger and to keep their hands in their lap when not typing. Once participants pressed the enter key, the trial ended. A white fixation cross was presented in the center of a black background during the intertrial interval, which varied randomly from 2,000 to 2,500 ms.
Participants saw all pictures and all letter strings exactly one time. Picture and letter string pairings were pseudorandom, with 120 trials in total, intermixed and presented randomly over four blocks (30 trials per block). Each trial was comprised of either a two-letter (low-load) or six-letter (high-load) string followed by either a positive or a neutral picture, for a total of four conditions with 30 trials each (low-load neutral, low-load positive, high-load neutral, and high-load positive). Participants completed four practice trials (one for each condition) before beginning the experiment. Each consisted of a low-load or high-load trial followed by a randomized positive or neutral picture that was not presented during the actual experiment.
EEG recording and data reduction
Continuous EEG recordings were collected using an ActiCap and the ActiChamp amplifier system (Brain Products GmbH, Gilching Germany). Thirty-two electrode sites were used based on the 10/20 system. The electrooculogram (EOG) was recorded from four facial electrodes: two that were placed approximately 1 cm above and below the right eye, forming a bipolar channel to measure vertical eye movement and blinks and two that were approximately 1 cm beyond the outer edges of each eye, forming a bipolar channel to measure horizontal eye movements. The EEG data were digitized at 24-bit resolution and a sampling rate of 1,000 Hz.
EEG data were processed offline using BrainVision Analyzer 2 software (Brain Products GmbH). The data were segmented for each trial beginning 200 ms before picture onset and continuing for 2,000 ms (i.e., the entire picture duration). Baseline correction was performed for each trial using the 200 ms before picture onset. The signal from each electrode was re-referenced to the average of the left and right mastoids (TP9/10) and band-pass filtered with high-pass and low-pass filters of 0.01 and 30 Hz, respectively. Eyeblink and ocular corrections used the method developed by Miller, Gratton, and Yee (1988). Artifact analysis was used to identify a voltage step of more than 50.0 μV between sample points, a voltage difference of 300.0 μV within a trial, and a maximum voltage difference of less than 0.50 μV within 100 ms intervals. Trials also were inspected visually for any remaining artifacts, and data from individual channels containing artifacts were rejected on a trial-to-trial basis. The average percentage of trials rejected per participant and condition were as follows: low-load, neutral = 1.64% (SD = 5.18), low-load, positive = 1.90% (SD = 5.59), high-load, neutral = 1.74% (SD = 5.61), high-load, positive = 1.77% (SD = 5.70). The percentage of trials rejected did not vary by working memory load, picture type, or their interaction, all ps > 0.48. In line with our prior work that has used this task with negative pictures (MacNamara et al., 2012, 2019; MacNamara, Ferri, et al., 2011a; MacNamara & Proudfit, 2014), trials were not excluded from LPP analyses based on working memory performance. Based on our prior work using this task and on visual inspection, the LPP was scored by averaging amplitudes at a pooling of five sites: FC1, FC2, Cz, CP1, and CP2 between 400-2,000 ms following picture onsetFootnote 2 (Holmes et al., 2009).
Working memory performance
Responses on the letter recall task were considered correct if they contained the same letters as those presented at the beginning of the trial, entered in the exact same order as originally presented. The percentage of correct responses per condition was calculated as the number of correct trials divided by 30 trials in each condition.
Data analyses
The LPP and accuracy data on the working memory task were analyzed using a 2 (working memory load: low, high) x 2 (picture type: neutral, positive) repeated measures analysis of variance (ANOVA). Statistical analyses were performed using SPSS statistical software version 25.0 (IBM, Armonk, NY).
Results
Table 1 presents means and standard deviations for all dependent variables, shown separately for each condition.
Working memory performance
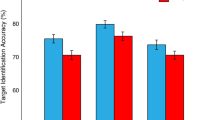
Overall, participants performed well on the letter recall task (M = 84.43% correct, SD = 8.71). Working memory performance was lower on high-load (M = 70.85%, SD = 16.19) compared with low-load trials (M = 98.01%, SD = 2.45), F(1,40) = 129.86, p < 0.001, ηp2 = 0.77. The effect of picture type did not reach significance, and there was no interaction between picture type and working memory load, both ps > 0.15.
LPP
Figure 1 presents grand-averaged waveforms at the pooling of FC1, FC2, Cz, CP1, and CP2 for each of the four conditions, as well as headmaps depicting the voltage differences between positive minus neutral (left) and low-load minus high-load (right) trials in the 400-2,000 ms window. A main effect of picture type indicated that positive pictures elicited larger LPP amplitudes than neutral pictures, F(1,40) = 71.47, p < 0.001, ηp2 = 0.64. In addition, a main effect of working memory load indicated that pictures presented on high-load trials elicited smaller LPPs compared to pictures presented on low-load trials, F(1,40) = 55.32, p < 0.001, ηp2 = 0.58. There was no interaction between picture type and working memory load, p > 0.67.

Grand-averaged waveforms depict amplitudes elicited by pictures in each of the four conditions at a pooling of FC1, FC2, Cz, CP1, and CP2, where the LPP was scored. Headmaps illustrate the difference between positive minus neutral pictures (left) and pictures presented on low-load minus high-load (right) trials, between 400-2,000 ms after picture onset
Correlations
Correlations were performed to determine whether the LPP was associated with performance on the working memory task. Difference scores reflecting the effect of working memory load were calculated separately for each of the LPP and working memory performance, i.e., low-load minus high-load positive scenes, low-load minus high-load neutral scenes. Correlations between the corresponding difference scores for the LPP and performance failed to reach significance, all rs > −0.23 and < 0.04; ps > 0.16.
Discussion
The current study examined the effect of working memory load on the processing of positive and neutral pictures. A main effect of picture type on the LPP indicated that despite the task-irrelevant nature of pictures, positive pictures received increased processing resources compared with neutral pictures. In addition, working memory load reduced the LPP elicited by positive and neutral pictures, in line with prior results observed using the same paradigm with negative pictures (MacNamara et al., 2012, 2019; MacNamara, Ferri, et al., 2011a; MacNamara & Proudfit, 2014). As expected, participants recalled more letters on low- compared to high-load trials, but positive pictures neither facilitated nor interfered with working memory performance, and no significant associations between the LPP and performance on the working memory task were observed.
Our results build on prior work, which found that working memory load reduced the LPP to positive words (Kopf et al., 2013) and faces (Van Dillen & Derks, 2012). Extension of effects to emotional scenes is important, because emotional scenes may be more arousing than other types of stimuli (Britton et al., 2006) and appear to modulate the LPP more robustly than emotional faces (Thom et al., 2014). Moreover, our results facilitate comparison with a broader body of work on emotion regulation, which has primarily used standardized emotional scenes (e.g., the IAPS; Lang et al., 2008).
Our results also fit with work that suggests that cognitive effort and/or nonaffective engagement of the prefrontal cortex may account for some of the downregulation observed in willful emotion regulation (Wyczesany & Ligeza, 2017). That is, in the absence of willful attempts at emotion regulation, cognitively demanding tasks may provide a means of reducing attention to positive stimuli that are not in line with an individual’s goals (e.g., high calorie food; van Dillen & van Steenbergen, 2018; see also van Dillen & Andrade, 2016). While this could be beneficial in contexts where reducing attention to positive stimuli might aid in adopting healthier behaviors (e.g., dieting), high levels of cognitive load might also play a role in positive affect impairments, such as those observed in depression (Vanderlind et al., 2020). This seems especially plausible given that states, such as rumination and worry, have been associated with increased working memory load and engagement of the dlPFC (Hayes et al., 2008; Steinfurth et al., 2017).
While prior work by Kopf et al. (2013) used an emotional n-back task and positive, negative and neutral words, Van Dillen and Derks (2012) used a paradigm that more closely resembled the one used here (i.e., emotional stimuli were task-irrelevant). Results showed a main effect of working memory load on the LPP elicited by angry and happy faces (neutral faces were not shown). In addition, Van Dillen and Derks (2012) observed an interaction between working memory load and face type, such that working memory load reduced the LPP more for angry compared with happy faces. This interaction was attributed to a negativity bias for angry compared with happy faces (i.e., larger LPPs to angry faces) under low-load that was no longer evident when faces were presented under high-load. Here, working memory load had a similar effect for both positive and neutral pictures (i.e., there was no interaction between working memory load and picture type). Therefore, when using emotional scenes (not faces), working memory load may be insufficient to attenuate affective potentiation of the LPP; rather, overall reductions in stimulus salience are observed. Nonetheless, future work may wish to include positive, negative, and neutral pictures to compare the relative effect of working memory load on the LPP elicited by each picture type.
One reason that working memory load might reduce the processing of task-irrelevant stimuli could be to attenuate interference. We failed to observe an adverse effect of positive pictures on working memory performance, even though we had previously found that negative pictures potentiated the adverse effect of working memory load on performance (MacNamara, Ferri, et al., 2011a). The current results are in line with prior work suggesting that positive stimuli may be less likely to interfere with performance than negative stimuli (Grissmann et al., 2017) but differ somewhat from those of Erk et al. (2007), who, in the context of an fMRI study, found that positive pictures facilitated working memory performance. A comparison of accuracy rates from the current study and this prior work suggests that the working memory task used here may have been more difficult than that used by Erk et al. (2007), in which participants did not need to recall a string of letters but only recognize a probe letter. Therefore, one possibility is that positive stimuli might facilitate performance on tasks with relatively low, but not high working memory load. Nonetheless, positive and negative pictures can differ in terms of arousal, not just valence. Therefore, our failure to observe an effect of positive stimuli on working memory performance cannot be definitively attributed to stimulus valence, and future work might delve further into this issue by using pictures that are matched on arousal but differ on valence.
Additionally, we found that the LPP and working memory performance were not correlated. This could be due in part to the temporal separation between measurement of the LPP and behavioral response (i.e., participants viewed a picture and then entered the letters they recalled from the beginning of the trial). Nonetheless, even when neural and behavioral response are measured at the same point in time/to the same stimuli, behavior is more “downstream” than neural response—i.e., many intervening variables may affect behavioral response beyond attention to task-irrelevant stimuli (measured by the LPP), including motivation, decision making, motor speed, etc. (MacNamara et al., 2013). Relatedly, compensatory effects (e.g., trying harder, recruiting additional brain regions) might help to offset any potential decrements in behavioral performance among individuals who allocate more attention to task-irrelevant pictures.
From a limited capacity perspective, demanding cognitive tasks, such as the working memory task used here, may exert their effects on task-irrelevant stimuli via distraction (Van Dillen & Koole, 2007). That is, when a person is engaged with a task that is cognitively demanding, s/he may have fewer processing resources available for the processing of task-irrelevant stimuli that would normally capture attention. Distraction has been associated with reduced memory for emotional events (Richards & Gross, 2006; Sheppes & Meiran, 2008) and stimuli that elicit smaller LPPs are remembered poorly at subsequent encounter compared with pictures that elicit larger LPPs (Dolcos & Cabeza, 2002). Therefore, smaller LPPs to pictures presented on high- compared with low-load trials suggest that these pictures may not have been encoded as well as the pictures presented in the low-load condition. Future work may wish to test this possibility directly by testing the effect of working memory load on the LPP and memory for pictures at subsequent encounter.
Conclusions
In sum, results observed here reveal that working memory load reduces the LPP to positive and neutral scenes and may be useful in understanding both adaptive and nonadaptive downregulation of attention to positive stimuli. For instance, the current paradigm could be used to examine how incentives might be used to bolster modulation of attention to positive distracters (Jones et al., 2020) or to assess the role of working memory load in disorders of reduced positive affect, such as depression.
Notes
The numbers of the IAPS images used were the following: pleasant (1440, 1460, 1601, 1604, 1721, 2034, 2060, 2160, 2332, 4006, 4233, 4311, 4240, 4525, 4530, 4550, 4561, 4574, 4611, 4624, 4625, 4641, 4647, 4669, 4672, 4698, 5010, 5480, 5661, 5825, 7220, 7283, 7325, 7340, 7482, 7489, 7508, 7515, 7580, 8193, 8205, 8208, 8460, 8461, 8540) and neutral (2036, 2280, 2515, 2840, 5000, 5532, 5534, 5635, 6150, 7001, 7012, 7014, 7020, 7021, 7025, 7031, 7033, 7036, 7052, 7205, 7217, 7224, 7236, 7705, 9360). The numbers of the EmoPicS images used were the following: pleasant (001, 005, 012, 014, 019, 022, 026, 029, 033, 039, 061, 067, 073, 332, 333) and neutral (083, 092, 110, 116, 119, 120, 122, 124, 128, 131, 138, 160, 161, 168, 169, 173, 175, 178, 181, 185, 189, 191, 200, 201, 276, 281, 284, 298, 336, 339, 340, 341, 352, 354, 366).
The LPP was also scored at early and late windows by averaging amplitudes at a pooling of FC1, FC2, Cz, CP1, and CP2. For the early LPP window (400-1,000 ms), there was a main effect of picture type, F(1,40) = 75.28, p < .001, ηp2 = .65 (positive > neutral). There was also a main effect of working memory load, F(1,40) = 33.98, p < .001, ηp2 = .46 (low-load > high-load). There was no interaction between picture type and working memory load, p > .66. For the late LPP window (1,000-2,000 ms), there was a main effect of picture type, F(1,40) = 57.04, p < .001, ηp2 = .59 (positive > neutral). There was also a main effect of working memory load, F(1,40) = 62.00, p < .001, ηp2 = .61 (low-load > high-load). There was no interaction between picture type and working memory load, p > .69.
References
Amaral, D. G., & Price, J. L. (1984). Amygdalo-cortical projections in the monkey (Macaca fascicularis). The Journal of Comparative Neurology, 230, 465–496.
Ashcraft, M. H., & Kirk, E. P. (2001). The relationships among working memory, math anxiety, and performance. Journal of Experimental Psychology: General, 130(2), 224–237. https://doi.org/10.1037/0096-3445.130.2.224
Banks, S. J., Eddy, K. T., Angstadt, M., Nathan, P. J., & Phan, K. L. (2007). Amygdala–frontal connectivity during emotion regulation. Social Cognitive and Affective Neuroscience, 2(4), 303–312. https://doi.org/10.1093/scan/nsm029
Britton, J. C., Taylor, S. F., Sudheimer, K. D., & Liberzon, I. (2006). Facial expressions and complex IAPS pictures: Common and differential networks. NeuroImage, 31(2), 906–919. https://doi.org/10.1016/j.neuroimage.2005.12.050
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., Weber, J., & Ochsner, K. N. (2014). Cognitive Reappraisal of Emotion: A Meta-Analysis of Human Neuroimaging Studies. Cerebral Cortex, 24(11), 2981–2990. https://doi.org/10.1093/cercor/bht154
Cavada, C. (2000). The anatomical connections of the macaque monkey orbitofrontal cortex. A review. Cerebral Cortex, 10, 220–242.
Cuthbert, B. N., Schupp, H. T., Bradley, M. M., Birbaumer, N., & Lang, P. J. (2000). Brain potentials in affective picture processing: Covariation with autonomic arousal and affective report. Biological Psychology, 52, 95–111. https://doi.org/10.1016/S0301-0511(99)00044-7
Dolcos, F., & Cabeza, R. (2002). Event-related potentials of emotional memory: Encoding pleasant, unpleasant, and neutral pictures. Cognitive, Affective, & Behavioral Neuroscience, 2(3), 252–263. https://doi.org/10.3758/CABN.2.3.252
Erk, S., Kleczar, A., & Walter, H. (2007). Valence-specific regulation effects in a working memory task with emotional context. NeuroImage, 37(2), 623–632. https://doi.org/10.1016/j.neuroimage.2007.05.006
Grissmann, S., Faller, J., Scharinger, C., Spüler, M., & Gerjets, P. (2017). Electroencephalography Based Analysis of Working Memory Load and Affective Valence in an N-back Task with Emotional Stimuli. Frontiers in Human Neuroscience, 11. https://doi.org/10.3389/fnhum.2017.00616
Hajcak, G., Anderson, B. S., Arana, A., Borckardt, J., Takacs, I., George, M. S., & Nahas, Z. (2010a). Dorsolateral prefrontal cortex stimulation modulates electrocortical measures of visual attention: Evidence from direct bilateral epidural cortical stimulation in treatment-resistant mood disorder. Neuroscience, 170, 281–288. https://doi.org/10.1016/j.neuroscience.2010.04.069
Hajcak, G., MacNamara, A., & Olvet, D. M. (2010b). Event-related potentials, emotion, and emotion regulation: An Integrative Review. Developmental Neuropsychology, 35, 129–155. https://doi.org/10.1080/87565640903526504
Hajcak, G., & Nieuwenhuis, S. (2006). Reappraisal modulates the electrocortical response to unpleasant pictures. Cognitive, Affective & Behavioral Neuroscience, 6, 291–297. https://doi.org/10.3758/CABN.6.4.291
Hayes, S., Hirsch, C., & Mathews, A. (2008). Restriction of working memory capacity during worry. Journal of Abnormal Psychology, 117(3), 712–717. https://doi.org/10.1037/a0012908
Holmes, A., Nielsen, M. K., Tipper, S., & Green, S. (2009). An electrophysiological investigation into the automaticity of emotional face processing in high versus low trait anxious individuals. Cognitive, Affective, & Behavioral Neuroscience, 9(3), 323–334. https://doi.org/10.3758/CABN.9.3.323
Jones, N. P., Schlund, M., Kerestes, R., & Ladouceur, C. D. (2020). Emotional Interference in Early Adolescence: Positive Reinforcement Modulates the Behavioral and Neural Effects of Negative Emotional Distracters. Cerebral Cortex, 30(4), 2642–2657. https://doi.org/10.1093/cercor/bhz266
Kalokerinos, E. K., Greenaway, K. H., Pedder, D. J., & Margetts, E. A. (2014). Don’t grin when you win: The social costs of positive emotion expression in performance situations. Emotion, 14(1), 180–186. https://doi.org/10.1037/a0034442
Kim, S. H., & Hamann, S. (2007). Neural Correlates of Positive and Negative Emotion Regulation. Journal of Cognitive Neuroscience, 19(5), 776–798. https://doi.org/10.1162/jocn.2007.19.5.776
Kopf, J., Dresler, T., Reicherts, P., Herrmann, M. J., & Reif, A. (2013). The Effect of Emotional Content on Brain Activation and the Late Positive Potential in a Word n-back Task. PLoS ONE, 8(9). https://doi.org/10.1371/journal.pone.0075598
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2008). International affective picture system (IAPS): Affective ratings of pictures and instruction manual. University of Florida, Gainesville. Tech Rep A-8.
Liu, Y., Huang, H., McGinnis-Deweese, M., Keil, A., & Ding, M. (2012). Neural Substrate of the Late Positive Potential in Emotional Processing. Journal of Neuroscience, 32(42), 14563–14572. https://doi.org/10.1523/JNEUROSCI.3109-12.2012
MacNamara, A., Ferri, J., & Hajcak, G. (2011a). Working memory load reduces the late positive potential and this effect is attenuated with increasing anxiety. Cognitive, Affective & Behavioral Neuroscience, 11(3), 321–331. https://doi.org/10.3758/s13415-011-0036-z
MacNamara, A., Foti, D., & Hajcak, G. (2009). Tell me about it: Neural activity elicited by emotional stimuli and preceding descriptions. Emotion, 9, 531–543.
MacNamara, A., Jackson, T. B., Fitzgerald, J. M., Hajcak, G., & Phan, K. L. (2019). Working Memory Load and Negative Picture Processing: Neural and Behavioral Associations With Panic, Social Anxiety, and Positive Affect. Biological Psychiatry. Cognitive Neuroscience and Neuroimaging, 4(2), 151–159. https://doi.org/10.1016/j.bpsc.2018.04.005
MacNamara, A., Kappenman, E. S., Black, S. R., Bress, J. N., & Hajcak, G. (2013). Integrating Behavioral and Electrocortical Measures of Attentional Bias Toward Threat. In K. C. Barrett, N. A. Fox, G. A. Morgan, D. J. Fidler, & L. A. Daunhauer (Eds.), Handbook of Self-Regulatory Processes in Development: New Directions and International Perspectives (pp. 215–246). Psychology Press.
MacNamara, A., Ochsner, K. N., & Hajcak, G. (2011b). Previously reappraised: The lasting effect of description type on picture-elicited electrocortical activity. Social Cognitive and Affective Neuroscience, 6(3), 348–358. https://doi.org/10.1093/scan/nsq053
MacNamara, A., & Proudfit, G. H. (2014). Cognitive load and emotional processing in generalized anxiety disorder: Electrocortical evidence for increased distractibility. Journal of Abnormal Psychology, 123(3), 557–565. https://doi.org/10.1037/a0036997
MacNamara, A., Schmidt, J., Zelinsky, G. J., & Hajcak, G. (2012). Electrocortical and ocular indices of attention to fearful and neutral faces presented under high and low working memory load. Biological Psychology, 91, 349–356. https://doi.org/10.1016/j.biopsycho.2012.08.005
Miller, G. A., Gratton, G., & Yee, C. M. (1988). Generalized implementation of an eye movement correction procedure. Psychophysiology, 25(2), 241–243. https://doi.org/10.1111/j.1469-8986.1988.tb00999.x.
Moran, T. P., Jendrusina, A. A., & Moser, J. S. (2013). The psychometric properties of the late positive potential during emotion processing and regulation. Brain Research, 1516, 66–75. https://doi.org/10.1016/j.brainres.2013.04.018
Moser, J. S., Hajcak, G., Bukay, E., & Simons, R. F. (2006). Intentional modulation of emotional responding to unpleasant pictures: An ERP study. Psychophysiology, 43, 292–296. https://doi.org/10.1111/j.1469-8986.2006.00402.x
Parvaz, M. A., MacNamara, A., Goldstein, R. Z., & Hajcak, G. (2012). Event-related induced frontal alpha as a marker of lateral prefrontal cortex activation during cognitive reappraisal. Cognitive, Affective, & Behavioral Neuroscience, 12, 730–740. https://doi.org/10.3758/s13415-012-0107-9
Picó-Pérez, M., Radua, J., Steward, T., Menchón, J. M., & Soriano-Mas, C. (2017). Emotion regulation in mood and anxiety disorders: A meta-analysis of fMRI cognitive reappraisal studies. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 79(Pt B), 96–104. https://doi.org/10.1016/j.pnpbp.2017.06.001
Ray, R. D., & Zald, D. H. (2012). Anatomical insights into the interaction of emotion and cognition in the prefrontal cortex. Neuroscience & Biobehavioral Reviews, 36(1), 479–501. https://doi.org/10.1016/j.neubiorev.2011.08.005
Reed, D. (2002). Random letter sequence generator. http://www.dave-reed.com/Nifty/randSeq.html
Richards, J. M., & Gross, J. J. (2006). Personality and emotional memory: How regulating emotion impairs memory for emotional events. Journal of Research in Personality, 40(5), 631–651. https://doi.org/10.1016/j.jrp.2005.07.002
Schindler, S., & Straube, T. (2020). Selective visual attention to emotional pictures: Interactions of task-relevance and emotion are restricted to the late positive potential. Psychophysiology, n/a(n/a), e13585. https://doi.org/10.1111/psyp.13585
Schupp, H. T., Stockburger, J., Codispoti, M., Junghöfer, M., Weike, A. I., & Hamm, A. O. (2007). Selective Visual Attention to Emotion. Journal of Neuroscience, 27(5), 1082–1089. https://doi.org/10.1523/JNEUROSCI.3223-06.2007
Shafir, R., Zucker, L., & Sheppes, G. (2018). Turning off hot feelings: Down-regulation of sexual desire using distraction and situation-focused reappraisal. Biological Psychology, 137, 116–124. https://doi.org/10.1016/j.biopsycho.2018.07.007
Sheppes, G., & Meiran, N. (2008). Divergent cognitive costs for online forms of reappraisal and distraction. Emotion, 8(6), 870–874. https://doi.org/10.1037/a0013711
Smith, E. E., Jonides, J., Marshuetz, C., & Koeppe, R. A. (1998). Components of verbal working memory: Evidence from neuroimaging. Proceedings of the National Academy of Sciences of the United States of America, 95(3), 876–882.
Steinfurth, E. C. K., Alius, M. G., Wendt, J., & Hamm, A. O. (2017). Physiological and neural correlates of worry and rumination: Support for the contrast avoidance model of worry. Psychophysiology, 54(2), 161–171. https://doi.org/10.1111/psyp.12767
Storbeck, J., & Maswood, R. (2016). Happiness increases verbal and spatial working memory capacity where sadness does not: Emotion, working memory and executive control. Cognition & Emotion, 30(5), 925–938. https://doi.org/10.1080/02699931.2015.1034091
Tacikowski, P., & Nowicka, A. (2010). Allocation of attention to self-name and self-face: An ERP study. Biological Psychology, 84(2), 318–324. https://doi.org/10.1016/j.biopsycho.2010.03.009
Thom, N., Knight, J., Dishman, R., Sabatinelli, D., Johnson, D. C., & Clementz, B. (2014). Emotional scenes elicit more pronounced self-reported emotional experience and greater EPN and LPP modulation when compared to emotional faces. Cognitive, Affective & Behavioral Neuroscience, 14(2), 849–860. https://doi.org/10.3758/s13415-013-0225-z
van Dillen, L. F., & Andrade, J. (2016). Derailing the streetcar named desire. Cognitive distractions reduce individual differences in cravings and unhealthy snacking in response to palatable food. Appetite, 96, 102–110. https://doi.org/10.1016/j.appet.2015.09.013
Van Dillen, L. F., & Derks, B. (2012). Working memory load reduces facilitated processing of threatening faces: An ERP study. Emotion, 12(6), 1340–1349. https://doi.org/10.1037/a0028624
Van Dillen, L. F., & Koole, S. L. (2007). Clearing the mind: A working memory model of distraction from negative mood. Emotion, 7(4), 715–723. https://doi.org/10.1037/1528-3542.7.4.715
van Dillen, L. F., & van Steenbergen, H. (2018). Tuning down the hedonic brain: Cognitive load reduces neural responses to high-calorie food pictures in the nucleus accumbens. Cognitive, Affective, & Behavioral Neuroscience, 18(3), 447–459. https://doi.org/10.3758/s13415-018-0579-3
Vanderlind, W. M., Millgram, Y., Baskin-Sommers, A. R., Clark, M. S., & Joormann, J. (2020). Understanding positive emotion deficits in depression: From emotion preferences to emotion regulation. Clinical Psychology Review, 76, 101826. https://doi.org/10.1016/j.cpr.2020.101826
Weinberg, A., Hilgard, J., Bartholow, B. D., & Hajcak, G. (2012). Emotional targets: Evaluative categorization as a function of context and content. International Journal of Psychophysiology, 84(2), 149–154. https://doi.org/10.1016/j.ijpsycho.2012.01.023
Wessa, M., Kanske, P., Neumeister, P., Bode, K., Heissler, J., & Schönfelder, S. (2010). EmoPicS: subjective and psychophysiological evaluation of new imagery for clinical biopsychological research. Z. Klin. Psychol. Psychother. Suppl, 1, 11–77.
Wyczesany, M., & Ligeza, T. S. (2017). Is elaboration of affective content sufficient to attenuate emotional responses during reappraisal procedures? Biological Psychology, 127, 99–107. https://doi.org/10.1016/j.biopsycho.2017.05.009
Yang, H., Yang, S., & Isen, A. M. (2013). Positive affect improves working memory: Implications for controlled cognitive processing. Cognition & Emotion, 27(3), 474–482. https://doi.org/10.1080/02699931.2012.713325
Acknowledgements
AM was supported by National Institute of Mental Health grant, K23MH105553 during the design of this study and data collection. AM is a consultant for Aptinyx Inc.
Open practices statement
The study was not pre-registered. However, all data are open and available on the Open Science Framework (https://osf.io/w9tjz/), and we report all conditions, measures, manipulations, and data exclusions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Barley, B., Bauer, E.A., Wilson, K.A. et al. Working memory load reduces the electrocortical processing of positive pictures. Cogn Affect Behav Neurosci 21, 347–354 (2021). https://doi.org/10.3758/s13415-021-00875-z
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-021-00875-z