
Abstract
The emotional attentional blink (EAB) refers to a temporary impairment in the ability to identify a target when it is preceded by an emotional distractor. It is thought to occur because the emotional salience of the distractor exogenously captures attention for a brief duration, rendering the target unattended and preventing it from reaching awareness. Here we tested the extent to which the EAB can be attenuated by inducing a diffuse top-down attentional state, which has been shown to improve target identification in an analogous attentional phenomenon, the attentional blink. Rapid sequences of landscape images were presented centrally, and participants reported the orientation of a ± 90° rotation of a landscape target. To induce a diffuse state of attention, participants were given a secondary task of monitoring for the appearance of a colored dot in the periphery. We found that emotional distractors impaired target recognition performance to comparable extents, regardless of whether or not participants concurrently performed the peripheral-monitoring task. Moreover, we found that performance of the secondary task led to an impaired ability to ignore neutral distractors. Subjective ratings of target vividness mirrored the behavioral accuracy, with frequent reports of intermediate levels of vividness suggesting that the EAB might impair target visibility in a graded manner. Our results demonstrate that the EAB is robust to manipulations of top-down attention, suggesting that the temporary capture of attention by emotionally salient stimuli involves processes that are distinct from those that produce the attentional blink.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
To cope with the constant bombardment of excess visual information from our environment, we must attend to behaviorally relevant information and ignore information that is irrelevant. Selective attention to information usually leads to a behavioral advantage for perception, but it also has a measurable cost, as evidenced by a phenomenon called the “attentional blink.”
The attentional blink (AB) refers to transient impairment in the ability to perceive the second of two masked targets when the second target appears approximately 200–500 ms after the attended first target (Chun & Potter, 1995; Giesbrecht & Di Lollo, 1998; Raymond, Shapiro, & Arnell, 1992). This momentary impairment in the ability to detect or identify a second target has been suggested to occur because of an overinvestment of limited-capacity resources to processing of the first target, rendering the second target unattended and preventing its perception (Olivers & Nieuwenhuis, 2006). Research on the AB has suggested that the overinvestment of attentional resources to the first target can be mitigated by the addition of task-irrelevant visual motion (Arend, Johnston, & Shapiro, 2006) or by requiring participants to perform a secondary task involving concurrent monitoring of stimuli in the periphery (Wierda et al., 2010). Performance can also be facilitated by providing a purely cognitive instruction to focus less attention on the first target (Olivers & Nieuwenhuis, 2006). Such seemingly paradoxical attenuation of the AB is theorized to result from the participant’s adoption of a more diffuse top-down attentional state, which prevents the participant from allocating too much attentional resource to the first target (Olivers & Nieuwenhuis, 2006).
Other research has revealed a compelling analogue to the AB that can be elicited by emotionally salient stimuli. The emotional attentional blink (EAB) refers to a temporary impairment in the ability to identify a single target when that target is preceded by an emotionally salient distractor, such as a disgusting, violent, or erotic image (Ciesielski, Armstrong, Zald, & Olatunji, 2010; McHugo, Olatunji, & Zald, 2013; Most, Smith, Cooter, Levy, & Zald, 2007). In the typical EAB paradigm, the participant must discriminate a single rotated target image of a building or a landscape that is embedded in a rapid serial visual presentation (RSVP) of nontarget images, such as upright landscape images (Most, Chun, Widders, & Zald, 2005; Olatunji, Ciesielski, Armstrong, Zhao, & Zald, 2011; Piech et al., 2011; Piech, Pastorino, & Zald, 2010). Participants exhibit a significant decrement in their ability to discriminate the target’s orientation if an emotional distractor appears 100–600 ms prior to the target, but they show minimal impairment for neutral distractor images (i.e., Fig. 1a; Ciesielski et al., 2010; Most & Jungé, 2008). Although the time course of impairment in the EAB resembles the traditional AB, a key difference lies in the fact that the EAB arises from the presentation of a task-irrelevant distractor that nevertheless captures the participant’s attention. In this respect, the EAB paradigm provides a powerful approach for characterizing how an emotionally salient stimulus can exogenously capture attention in a stimulus-driven manner. Identifying similarities and dissimilarities between the AB and EAB could therefore help reveal the shared or distinct mechanisms involved in goal-directed and stimulus-driven attention.

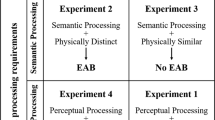
An outline of anticipated results. In the typical EAB paradigm, an impairment in target performance is observed when an emotional distractor is presented close in time to the target (A). If this impairment is the result of an overinvestment of limited attentional resources toward the emotional distractor, similar to that observed in the AB, then the induction of a diffuse attentional state via the introduction of a peripheral monitoring task should alleviate the EAB (B). However, given that the EAB is the result of involuntary attentional capture prior to a target rather than of voluntary attention allocation, as in the AB, is also possible that the introduction of a concurrent task would have no effect (C) or might even worsen task performance by increasing demands (D)
Although we have some understanding of how a diffuse state of attention, induced by a concurrent visual or cognitive task, can affect the traditional AB (Arend et al., 2006; Olivers & Nieuwenhuis, 2006; Wierda, van Rijn, Taatgen, & Martens, 2010), researchers have yet to investigate its potential impact on the EAB. We sought to investigate this question by developing a dual-task paradigm that required participants to monitor a central rapid serial visual presentation (RSVP) sequence of images while concurrently monitoring the periphery to perform another task (Olivers & Nieuwenhuis, 2006; Wierda et al., 2010). In Fig. 1 we present three potential competing hypotheses regarding how the EAB might be modulated by the introduction of a peripheral monitoring task to induce a diffuse attentional state.
If similar cognitive mechanisms underlie the AB and EAB, then performance of a concurrent spatial monitoring task of the periphery should lead to more diffuse allocation of limited attentional resources, and might thereby mitigate the overinvestment of limited resources to the emotional RSVP distractor (i.e., Fig. 1b; Olivers & Nieuwenhuis, 2006). An improvement in target discrimination at lag 2 would demonstrate that the involuntary capture of limited resources by an emotional distractor can be modulated by manipulations of voluntary spatial attention. Such a finding would suggest that the deficits in target identification observed in the EAB and AB both result from an excessive allocation of limited attentional resource.
Unlike the AB, however, the EAB involves the exogenous capture of attention by irrelevant emotional information. Thus, one might alternatively expect that the allocation of attention in the EAB would involve mechanisms that are distinct from those that mediate the AB (Most & Jungé, 2008; Most & Wang, 2011; Wang, Kennedy, & Most, 2012). Should this be the case, then the EAB might be robust to manipulations of central attention, such that adding a secondary task in order to distribute attention more diffusely across space might have no effect on dampening the EAB, as is illustrated in Fig. 1c.
A final possibility to consider is that the added task might incur a greater cost to target processing by further taxing attentional control mechanisms (i.e., Fig. 1d). Previous work has demonstrated that participants exhibit poorer attentional filtering and greater interference from task-irrelevant stimuli when they must perform a secondary cognitive task, particularly when that task places greater demands on top-down control and working memory (de Fockert & Bremner, 2011; de Fockert, Rees, Frith, & Lavie, 2001; Kelley & Lavie, 2010; Lavie, Hirst, de Fockert, & Viding, 2004). This increase in distractability is posited to occur because the attentional control mechanisms needed to maintain goal-directed priorities are taxed, thereby impairing the ability to select and filter the appropriate stimuli. As such, the concurrent task could increase the degree to which the distractors in the EAB paradigm capture limited attentional resources and thereby incur a greater cost to target processing.
Another goal of this study was to provide an exploratory investigation of whether the EAB modifies awareness of the target stimulus in a discrete or graded manner. Prior research using neutrally valenced stimuli has suggested that the AB may disrupt processing of the second target in a discrete, all-or-none manner, as evidenced by a bimodal distribution of subjective ratings of the target’s visibility (Sergent & Dehaene, 2004) and by estimated rates of random guessing in studies that allow the target’s precise identity to be reported in a continuous feature space (Asplund, Fougnie, Zughni, Martin, & Marois, 2014). However, others have reported evidence to suggest that graded levels of perception occur during the AB (Nieuwenhuis & de Kleijn, 2011; Overgaard, Rote, Mouridsen, & Ramsøy, 2006; Sy, Marois, & Tong, 2018). To test whether conscious perception during the EAB reflects a discrete or graded loss of information about the target, we required participants to rate the vividness of their experience of the target on each trial, using a 4-point rating scale (Overgaard et al., 2006). If the EAB impedes conscious perception of the target in a discrete, all-or-none manner, then reported vividness should reveal a dichotomous pattern of vividness responses, indicating either no experience or a clear experience of the target. In contrast, reports of intermediate levels of vividness would imply a graded representation of the target.
Our dual-task paradigm required participants to monitor a central RSVP of upright landscape images, as is typical of the EAB, while concurrently monitoring the color of dots that briefly flashed at random peripheral locations (Olivers & Nieuwenhuis, 2006; Wierda et al., 2010). Specifically, participants had to discriminate whether a clockwise- or counterclockwise-rotated landscape target occurred within each RSVP sequence of upright landscape images. A distractor image was presented on every trial, preceding the target by either two (187 ms) or eight (747 ms) images, and consisted of either an emotionally erotic image or a neutral image portraying people engaged in everyday activities. We investigated how different manipulations of voluntary spatial attention affected performance by evaluating three experimental tasks. In the single-task control, participants simply had to focus on the central RSVP stream and report their perceptions of the landscape target. In two different dual-task versions of this experiment, participants were additionally asked to detect the infrequent appearance of a peripheral red dot target (20% of trials) among more frequent presentations of a black dot. The dual tasks differed in the spatial extent of possible target locations. In the diffuse-attention condition, the target dot could appear at any randomly chosen peripheral location along a radius of 9° visual angle from the center of the screen. In the cued version of the dual task, spatial attention was narrowed because the potential location of the peripheral target was cued before the start of the trial. The addition of a concurrent task, along with these manipulations of the spatial spread of attention, allowed us to investigate the effect of diffuse attention on the EAB. Finally, we investigated whether the EAB affected target perception in a graded or all-or-none fashion by asking participants to rate the vividness of their target experience on each trial.
Method
Participants
Nineteen participants (ages 20–30; 11 female, 8 male) from the Vanderbilt community participated in the hour-long study and received monetary compensation for their efforts. Each participant provided informed written consent prior to the study’s start, as per Vanderbilt University Institutional Review Board guidelines. Participants were informed of the nature of the visual stimuli they would be viewing. One participant was excluded from the analyses because he reported ignoring the spatial cue in the cued-attention task. This sample size was chosen on the basis of previous studies probing similar effects of diffuse-attention manipulations (Arend et al., 2006; Olivers & Nieuwenhuis, 2006; Wierda et al., 2010) and previous studies of the EAB (Kennedy, Rawding, Most, & Hoffman, 2014; Most et al., 2005; Most et al., 2007; Smith, Most, Newsome, & Zald, 2006), ranging from 11 to 24 participants.
Apparatus
Stimuli were presented on a 17-in. (43.18-cm), 75-Hz CRT monitor via the Psychophysics Toolbox in MATLAB (Brainard, 1997; Pelli, 1997).
Stimuli
A black fixation cross, 0.7° × 0.7° in size, appeared in the center of the screen against a uniform gray background prior to and during each trial. The stimuli consisted of color photographs sampled from 56 erotic images, 56 neutral pictures, 256 upright landscape/architectural scenes, and 244 target images (122 landscape/architectural photos rotated 90° to the left and right). The landscape images were drawn from the set used by Most et al. (2005), and the neutral images were drawn mostly from the International Affective Picture System (IAPS; Lang, Bradley, & Cuthbert, 2008). The emotional distractors consisted of erotic images drawn partly from the IAPS and supplemented with images drawn from the Internet (Most et al., 2007). The erotic pictures included images of mixed-sex nude couples engaging in sexual acts, whereas the neutral distractor images consisted of portraits depicting one person or several people in everyday settings and were rated as eliciting no emotional reaction. The stimuli measured 11.6° × 9.3° of visual angle in size. In addition to the central photograph, a black or red dot (0.43° × 0.43°) was presented in one of 39 possible positions along a virtual circle at 9° eccentricity from the central fixation point. The images used in this study are available upon request from either David Zald or Steven Most.
Procedures
Each participant completed three different tasks over the course of an hour. Across tasks, every trial involved the presentation of both the RSVP images and the peripheral dot stimulus. Participants were instructed to keep their eyes fixed on a fixation cross at the center of the screen throughout each experimental trial. A spacebar press initiated an RSVP of 17 images presented at fixation for 93.33 ms each; an example is shown in Fig. 2. This rate of stimulus presentation was dictated by the refresh rate of the monitor used in the study. All images on a given trial consisted of different upright landscape/architectural photographs, except for two: the critical distractor and the target stimulus. The critical distractor was displayed as either the fourth or sixth stimulus in the stream. The distractor was either a neutral-valence image of everyday interactions of people or an emotionally salient image of a nude couple engaged in erotic acts, presented with equal probabilities within each experimental block. The target was presented at a lag of either two or eight images after the critical distractor (186.67 or 746.67 ms) and consisted of a landscape/architectural scene rotated 90° clockwise or counterclockwise from vertical. Concurrent with each image presentation, a dot was presented at 9° eccentricity from fixation. The location of the dot changed with each new RSVP image to one of 39 locations with equal probabilities, excluding the preceding location. In 80% of the trials, all dots in that trial were black. In 20% of the trials, one of the dots appeared red rather than black for one 93.33-ms image frame. The red dot could appear at any time during the RSVP stream, excluding the time of the first, second, or final image.

Participants discriminated the clockwise or counterclockwise rotation of the target landscape image, presented in a rapid serial visual presentation (RSVP) of upright landscape images. An irrelevant emotional (erotic) or neutral distractor preceded the target by either two or eight frames. In separate experimental blocks, attention to the RSVP stream was manipulated on the basis of task instructions to ignore or also attend to the peripherally presented dots
The three tasks differed in how attention was directed to the peripheral dots. In the single task, participants were asked to ignore all dots and focus attention on the RSVP stream. In the distributed-attention task, participants were instructed to detect the presence or absence of a red dot that could potentially occur in any of the 39 possible locations. In this distributed-attention task variant, attention was likely diffusely distributed across the two tasks and across a wide spatial extent. In the spatially cued attention task, a white dot 0.43° in diameter was presented for 250 ms, 1 s prior to the start of the RSVP stream, indicating with 100% validity the spatial position at which the red dot would appear if one were presented on that trial. The 1-s interval between the cue onset and the start of the RSVP stream provided enough time so that any bottom-up attentional effects of the cue should have been negligible (Theeuwes, Atchley, & Kramer, 2000), but ample time for exerting top-down spatial focusing. In this cued-attention paradigm, attention was likely distributed across discriminating the orientation of the landscape target and detecting a red dot at the cued location, but spatial attention could be restricted to those two spatial locations.
It is important to note that all trials in which a red dot appeared were excluded from the analysis of the EAB in order to avoid effects due to having to process the red dot. For each task, participants performed a 20-trial practice block with positive auditory feedback for correct responses. Three consecutive experimental blocks of the same experimental task followed each practice block. Each experimental block consisted of 40 total trials, eight of which included the red dot and were thus excluded from analysis of the EAB. The order of the three tasks was counterbalanced across participants, and manipulations of distractor valence and lag were randomized and counterbalanced within each experimental block.
In all three tasks, participants were required to report the direction in which the target image was rotated and how vivid the target appeared by using keyboard button-presses after each trial. In both dual-target tasks, participants were additionally asked whether a red dot had appeared during that trial, which they reported using a keyboard button-press following their reports of target rotation and vividness. Participants reported target vividness using the Perceptual Awareness Scale (PAS; Ramsøy & Overgaard, 2004), which allows participants to report one of four levels of awareness:
-
No experience. No impression of the stimulus is experienced. All answers are experienced as mere guessing.
-
Brief glimpse. A feeling that something was present, even though the content cannot be specified any further.
-
Almost clear experience. Feeling of having seen the stimulus, but only somewhat sure about it.
-
Clear experience. Nonambiguous experience of the stimulus.
Results
Peripheral target detection performance
Participants’ accuracy in red dot detection in both the distributed-attention task (mean = .89, SEM = .02) and the cued-attention task (mean = .84, SEM = .03) confirmed that some attention was allocated to the peripheral locations in both tasks. Indicative of equivalent effort across both peripheral tasks, detection of the red dot did not differ significantly between the distributed- and cued-attention tasks, t(17) = 1.49, p = .16.
Central target discrimination performance
Emotional distractors had a clear impact on the participants’ ability to discriminate the orientation of the rotated landscape target (90° clockwise or counterclockwise), especially when the target appeared at lag 2 as compared to lag 8 following the task-irrelevant emotional distractor. This impact was evident in each of the attention conditions (Fig. 3, top). By lag 8, performance returned to equally good levels for the neutral and emotional distractors, indicating the temporary nature of the attentional disruption of the EAB (Fig. 3, top).

Proportions of correct responses (top row) and average vividness ratings (bottom row) for the target are plotted as a function of distractor–target lag, distractor valence, and task. Ratings were assigned ordinal values: 1 = no experience (no exp), 2 = brief glimpse (brief), 3 = almost clear experience (almost), and 4 = clear experience (clear exp). Error bars represent the standard errors of the means (SEMs)
To confirm the presence of the EAB, we compared the proportions of correct target responses in each attention condition by performing a three-factor repeated measures analysis of variance (ANOVA) using lag (second or eighth item following distractor), distractor valence (neutral or emotional), and attention task (single, distributed, or cued) as within-subjects factors. As expected, the ANOVA revealed a statistically significant main effect of lag, F(1, 17) = 37.38, p < .001, ηp2 = .69, demonstrating impaired target performance at lag 2 as compared to lag 8 trials. There was also a main effect of distractor valence, F(1, 17) = 6.85, p = .018, ηp2 = .29, indicating poorer performance when the distractor was emotional than when it was neutral. Characteristic of the EAB, there was a statistically significant interaction between lag and distractor valence, F(1, 17) = 18.48, p < .001, ηp2 = 0.52, indicating that the effect of distractor valence was proportionally greater at lag 2 than at lag 8. Separate analyses for each lag comparing distractor valence (neutral or emotional) and attention task (single, distributed, cued) revealed that target performance was significantly worse following emotional than following neutral distractors at lag 2, F(1, 17) = 12.30, p = .003, ηp2 = .42, but not significantly different at lag 8, F(1, 17) = 2.19, p = .158, ηp2 = .11. These results provide a clear demonstration of the EAB.
All subsequent analyses were focused on lag 2, in order to determine the modulatory effects of distributed attention and distractor valence on target discrimination. If similar mechanisms underlie the AB and EAB, manipulations meant to diffuse top-down attention across tasks and/or space should similarly attenuate attentional capture by the emotional critical distractor and subsequently improve target discrimination. Conversely, if distinct mechanisms underlie the AB and the EAB, then manipulations of top-down attention might not affect or might even exacerbate the magnitude of the EAB. Critically, we found no main effect of attention task, F(2, 34) = 1.85, p = .173, ηp2 = .10, or interaction between attention task and valence, F(2, 34) = 1.97, p = .16, ηp2 = .10, at lag 2. These findings demonstrate that the EAB is unaffected by the additional task and the spatial diffusion of attention.
To further elucidate the effects of attentional state on performance in the neutral and emotional conditions, we performed two separate repeated measures analyses for neutral and emotional distractors at lag 2, using attention task as a factor. When the distractors were neutral, there was a significant, albeit modest, main effect of attention task, F(2, 34) = 5.05, p = .01, ηp2 = .23. Post-hoc pairwise t tests comparing performance across attention tasks with neutral distractors at lag 2, corrected for multiple comparisons using a Holm–Bonferroni correction (Holm, 1979), indicated that the main effect of attention was due to significantly worse performance when attention was distributed, t(17) = 3.07, p = .03, d = 0.72, or cued to a single peripheral location, t(17) = 2.50, p = .04, d = 0.59, when compared to the single-task control condition. However, there was no significant difference in performance between the distributed- and cued-attention tasks, t(17) = 0.16, p = .87, d = 0.04. As can be seen in Fig. 3, both peripheral-attention tasks, regardless of their spatial distribution, reduced participants’ ability to prevent a categorically salient but neutrally valenced distractor from capturing attention at lag 2, in comparison to target performance in the single-task condition. In contrast, we found no effect of attention task when the distractors were emotional, F(2, 34) = 0.14, p = .87, ηp2 = .01. These data support the robust ability of emotionally salient stimuli to capture attention and impair target performance at short temporal lags and indicate that, unlike the traditional AB, the bottom-up capture of attention by emotional stimuli is not impacted substantially by the concurrent distribution of attention.
Vividness ratings
As can be seen in Fig. 3 (bottom row), the average ratings of target vividness qualitatively mirrored the accuracy of participants’ performance in the target discrimination tasks. Targets appearing at lag 2 were rated as less vivid when they followed an emotional than when they followed a neutral distractor, whereas the vividness ratings were higher and more comparable across the two valence conditions by lag 8.
To quantitatively confirm this pattern, we compared the average ratings of target vividness in each attention condition across participants by performing a three-factor repeated measures ANOVA with lag (two or eight items), distractor valence (neutral or emotional), and attention task (single, distributed, or cued) as within-subjects factors. The ANOVA revealed significant main effects of lag, F(1, 17) = 75.20, p < .001, ηp2 = .82, and distractor valence, F(1, 17) = 57.91, p < .001, ηp2 = .77, reflecting the fact that targets were reported as being less vivid at short than at long lags and when they were preceded by an emotional rather than a neutral distractor. Consistent with the target discrimination data, we also found a significant interaction between lag and distractor valence, F(1, 17) = 43.02, p < .001, ηp2 = .72, revealing that the difference in vividness ratings between distractor valence conditions was larger at lag 2 than at lag 8. Finally, there was a significant interaction between attention task and distractor valence, F(2, 34) = 5.27, p = .01, ηp2 = .24. This interaction demonstrated that the addition of a concurrent task, irrespective of the extent of spatial attention, reduced the difference between valence conditions as compared to the single task.
We performed two separate analyses at lags 2 and 8, using distractor valence and attention task as within-subjects factors, to evaluate the differences in average vividness ratings at each lag. Consistent with an EAB, there was a significant main effect of distractor valence in the reported vividness of the target at lag 2, F(1, 17) = 62.98, p < .001, ηp2 = .79, which was not present at lag 8, F(1, 17) = 3.09, p = .097, ηp2 = 0.15. At lag 2, we also observed a significant interaction between distractor valence and attention task, F(2, 34) = 6.19, p = .005, ηp2 = .27, indicating a smaller valence effect when either diffuse- or cued-attention tasks were compared to the single attention task. In contrast, there was only a modest main effect of attention task at lag 8, F(2, 34) = 3.29, p = .049, ηp2 = .16. A post-hoc pairwise t test indicated that the main effect of attention task at lag 8 was driven by a significant difference between the single and cued-attention tasks, t(17) = 3.02, p = .008, d = 0.72.
To further understand the effects of attention for each distractor valence at lag 2, we performed separate ANOVAs for the neutral and emotional conditions at lag 2, using attention task as a within-subjects factor. We found no effect of attention task in the emotional condition at lag 2, F(2, 34) = 1.19, p = .317, ηp2 = .07, but did find an effect of attention in the neutral condition, F(2, 34) = 4.65, p = .016, ηp2 = .22. Consistent with our analysis of target discrimination accuracy, these results indicated that the introduction of a concurrent task specifically affected the reported vividness of the target in the neutral-distractor condition and had no reliable effect in the emotional-distractor condition.
We created an additional figure in order to display the relationship between subjective ratings of vividness and objective performance at the target discrimination task. Specifically, we were interested in how the accuracy of target responses might have varied according to which of the four possible levels of rated vividness the participant reported. As can be seen in Fig. 4a, participants’ average target accuracy increased monotonically as a function of vividness rating, with performance only somewhat greater than the chance level of .50 for ratings of no experience, ranging upward to greater than .95 accuracy for targets judged as clear experiences. These results indicate that rated vividness was likely not a result of response biases or arbitrary reports; instead, participants have excellent metacognitive awareness of the reliability of their target percept.

a Average proportions of correct target responses for each confidence rating for each distractor valence and lag. Error bars represent the standard errors of the means (SEMs). b Proportions of responses for each vividness rating are plotted as a function of attention task for each distractor valence (neutral, top row; emotional, bottom row) and lag (columns)
The behavioral cost in target performance in the EAB could arise from one of two possible perceptual experiences. First, target performance could be impaired because attentional capture by an emotionally salient distractor might probabilistically prevent the target from reaching conscious awareness in an all-or-none fashion. In this case, vividness ratings should reflect a dichotomous distribution, with most reports indicating either complete perception or a total lack of perceptual awareness of the target. Alternatively, the capture of attention by the emotional distractor might impair the clarity of the target percept in a graded manner. In this case, the vividness ratings would also reflect a graded distribution of vividness responses, in which some proportion of responses should be characterized by intermediate ratings of vividness. Figure 4 clearly indicates that target accuracy improved monotonically with each increase in rated vividness, supporting a continuity of responses across the vividness scale that was not restricted to all-or-none perceptual experiences of the target.
Discussion
Motivated by previous research on the AB, we evaluated whether the performance of a peripheral-monitoring task would improve or impair target discrimination performance in the EAB paradigm. In the single-task RSVP design, we replicated previous work showing that discrimination performance was significantly impaired for targets that followed an emotionally salient distractor at lag 2. Importantly, the EAB remained robust when participants had to perform a concurrent peripheral-monitoring task, since we observed a similar transient impairment in discrimination performance for targets preceded by emotional distractors at short lags. In this respect, our results run contrary to the predictions of the overinvestment hypothesis (Olivers & Nieuwenhuis, 2006). Instead, our findings suggest that the processing of an emotional distractor robustly outweighs the top-down prioritization of task-relevant stimuli.
Interestingly, we found that the performance of a concurrent dot-monitoring task led to impaired target discrimination for targets that followed the visually distinctive neutral distractor. The disruptive effect of the neutral distractor resembled the pattern of disruption caused by emotional distractors, insofar as it was specific to lag 2. However, unlike emotional distractors, neutral distractors led to a temporary cost in target processing only in the dual-task conditions. This finding was somewhat unexpected, since we did not anticipate that performance of a secondary task would lead to a selective disruption of target processing following neutral distractors without a corresponding decrement following emotional distractors in the dual-task paradigm. Nevertheless, the decrement following neutral distractors is broadly consistent with previous research on attentional and cognitive load; both attentional selection and distractor filtering tend to become less effective under conditions of high load (Akyürek & Hommel, 2005; Burnham, 2010; Kim, Kim, & Chun, 2005; Lavie & de Fockert, 2005; Park, Kim, & Chun, 2007). In the present study, it appears that performance of the secondary dot-monitoring task led to poorer top-down filtering of the neutral distractors, such that target discrimination was impaired at short lags. These neutral distractors were visually complex and categorically distinct from the landscape distractors, consisting of a large set of photos of people engaged in various real-world activities and social interactions. It should be noted that images of faces and people tend to attract greater attentional processing than do other categorically salient real-world stimuli (Evans & Treisman, 2005; Langton, Law, Burton, & Schweinberger, 2008; Theeuwes & Van der Stigchel, 2006), which may have boosted the disruptive impact of the neutral distractors under dual-task conditions even though they lacked clear emotional salience. However, categorical differences alone would not be expected to cause sustained attentional capture. Similar single-task RSVP studies have indicated that attention is captured only by the first few occurrences of a new but otherwise neutral stimulus category, suggesting that participants are able to rapidly learn to filter out categorically unique stimuli (Asplund, Todd, Snyder, Gilbert, & Marois, 2010). Within the context of our dual-task EAB paradigm, we found that categorically distinct neutral distractors are filtered out less effectively when attentional control mechanisms are taxed.
In contrast, the EAB did not become stronger under dual-task conditions. This finding is consistent with the notion that participants found it very difficult to filter out or disengage from processing these emotionally evocative stimuli, even in the single-task condition. A key aspect of the EAB paradigm is that all stimuli are presented foveally, including the sought-after target. As a consequence, participants must spatially attend to the location of the distractor. The visual complexity of the stimuli, as well as the fact that a different distractor image is shown on every trial, renders it difficult for participants to filter out the distractors on the basis of their low-level visual properties. Once the emotional distractor appears and is initially processed, the visual–attentional system appears to have difficulty disengaging from this stimulus, leading to a temporary blink effect in target discrimination. Taken together, our dual-task results suggest that attentional control is critical for top-down filtering of visually salient distractors but that this control, although highly effective for neutral stimuli, is severely limited in its ability to filter out emotional distractors regardless of the state of other attentional demands.
One might ask whether performance of the secondary dot-monitoring task might have had a greater impact on the EAB if it were performed at the same central location as the emotional distractor rather than in the periphery. Although this condition was not tested directly in our paradigm, we consider such an outcome to be unlikely. First, our analyses were focused exclusively on trials in which the target dot probe was withheld and therefore avoided direct competition with the emotional distractor. This approach was taken because the goal of the present experimental design was to induce a diffuse attentional state, not to investigate the impact of direct competition between the dot target and the emotional distractor. Second, participants already had to allocate some attentional resources to the central visual field to monitor for the rotated target landscape image; as a consequence, shifting the dot task to central vision would have simply led to a top-down spatial profile of attention that resembled the single-task condition. Although concurrent presentation of the dot target and emotional distractor might have led to attenuated processing of the emotional distractor, previous research has provided inconclusive evidence as to whether this would depend on their spatial proximity or overlap. Indeed, the spatial dynamics of emotional attentional capture warrant further exploration, given the apparent heterogeneity of current findings on the topic. Consistent with the present study, prior research has demonstrated that attentional capture by emotional distractors does not appear to depend upon the initial distribution of top-down spatial attention. A wealth of data indicate that emotional stimuli are capable of attracting attention when presented outside of the focus of top-down spatial attention (Erthal et al., 2005; Müller, Andersen, & Keil, 2008; Schönwald & Müller, 2014) and that emotional stimuli can modulate cortical and behavioral responses even when they are presented in the impaired hemifield of patients with visual extinction and neglect (Vuilleumier, Armony, Driver, & Dolan, 2001; Vuilleumier & Schwartz, 2001). In general, our results are in line with research suggesting that the ability of emotional distractors to capture attention is nonspatial, but that the effects of emotional stimuli are spatially selective once this attentional capture has occurred. In a pair of experiments by Most and Wang (2011), participants were asked to monitor two parallel RSVP streams for the occurrence of a rotated target image. In the first experiment, the targets were always followed by trailing nontarget landscape images, as in the present study. The researchers found that the introduction of an emotional distractor two stimuli before the target substantially impaired target discrimination when the emotional distractor and the subsequent target appeared in the same RSVP location, but only weakly and nonsignificantly affected target discrimination when the two stimuli occurred in separate locations. This suggests a spatial specificity of the disruptive effect of the emotional distractor. In the second experiment, target images were sometimes not followed by trailing landscape distractors. In these trials without landscape images trailing the target, performance was modestly impaired by emotional distractors that occurred at a different location and was unaffected by those that occurred at the target location. Presumably, spatial attention tended to be captured by the emotional distractor and could linger to process further information about the target if there was a persisting iconic trace of the target at that location. Consequently, the authors argued that the EAB is the result of spatially specific competition for resources at an early stage of perceptual processing rather than competition at a later, or central, stage (Wang et al., 2012). Convergent evidence for the spatial specificity of emotional effects has come from attentional-cueing studies demonstrating that the presentation of a task-irrelevant emotional face can enhance the perception of subsequent targets that appear at a common location (Phelps, Ling, & Carrasco, 2006; Pourtois, Grandjean, Sander, & Vuilleumier, 2004). In sum, the spatial characteristics of our dual-task paradigm should not have been a factor in the ability of the concurrent task to create a diffuse attentional state. That said, future studies should investigate the impact of requiring participants to attend to emotional and neutral stimuli concurrently, and should determine how the spatial relationship between these stimuli may impact the effects of either stimulus.
Additionally, we found that participants’ subjective ratings of their experience of the central target mirrored the pattern of target discrimination performance. Participants reported a weaker perceptual experience of the target when it shortly followed an emotional distractor, irrespective of the attentional task, as well as when the target soon followed a neutral distractor when participants were engaged in either dual-task paradigm. Critically, we found that participants rated their experience of the central target along a graded range of vividness, suggestive of graded effects of the EAB on conscious perception. These subjective ratings clearly deviate from a bimodal distribution of having either no experience or a clear experience of the target. Interestingly, the addition of a concurrent task attenuated the effects of distractor valence on vividness ratings at the shortest lag. Like target accuracy, this attenuation was due to a reduction in target vividness ratings for the neutral-distractor trials, with no evidence of a change in ratings on the emotional-distractor trials.
The frequent reports of intermediate levels of target vividness in the present study run contrary to previous studies that suggested all-or-none losses of target awareness during the traditional AB (Asplund et al., 2014; Sergent & Dehaene, 2004). However, it has previously been argued that the bimodal responses in confidence or vividness ratings in the AB can at least partially be attributed to potential biases that arise with the users’ interpretations of less descriptive rating scales (Nieuwenhuis & de Kleijn, 2011; Overgaard et al., 2006; Sy et al., 2018). Here we found graded levels of target vividness in the EAB using a rating scale that provides clear verbal descriptions to describe different levels of awareness; the use of this scale has led to positive reports of graded percepts in other visual tasks, such as the AB (Overgaard et al., 2006). The distribution of vividness levels reported in the present experiment cannot be readily attributed to participants’ failure to use the ratings consistently, because the average ratings of stimulus vividness mirrored the qualitative pattern observed in behavioral accuracy, and higher ratings of vividness corresponded to monotonic increases in target discrimination accuracy.
In summary, the present study demonstrated the robustness of the EAB to manipulations of voluntary spatial attention. We found that the addition of a concurrent peripheral-monitoring task can increase attentional capture by distractors, due to increases in top-down task demands; however, this dual-task cost to target performance only occurred in the presence of neutral distractors. This finding suggests that the dissociation between the processing of emotional- and neutral-valence distractor stimuli lies within attentional control mechanisms necessary to prevent attentional capture by distractors. Altogether, the present study may indicate that the AB and EAB result from limitations at different stages of information processing. Where the overinvestment hypothesis of the AB suggests that the AB results from temporary overinvestment of limited attentional resources, our work suggests that the EAB reflects limitations in the ability to appropriately select relevant stimuli to gain access to central attentional resources.
Author note
A National Science Foundation grant BCS-1228526 to F.T. and a National Institutes of Health Research Training Grant 5T32EY007135-18 awarded to the Vanderbilt Vision Research Center supported this work. Research was facilitated by a National Institutes of Health P30-EY-008126 center grant to the Vanderbilt Vision Research Center.
Change history
21 February 2019
In this issue, there is an error in the citation information on the opening page of each article HTML. The year of publication should be 2019 instead of 2001. The Publisher regrets this error.
References
Akyürek, E. G., & Hommel, B. (2005). Short-term memory and the attentional blink: Capacity versus content. Memory & Cognition, 33, 654–663. https://doi.org/10.3758/BF03195332
Arend, I., Johnston, S., & Shapiro, K. (2006). Task-irrelevant visual motion and flicker attenuate the attentional blink. Psychonomic Bulletin & Review, 13, 600–607.
Asplund, C. L., Fougnie, D., Zughni, S., Martin, J. W., & Marois, R. (2014). The attentional blink reveals the probabilistic nature of discrete conscious perception. Psychological Science, 25, 824–831.
Asplund, C. L., Todd, J. J., Snyder A. P., Gilbert, C. M., & Marois, R. (2010). Surprise-induced blindness: A stimulus-driven attentional limit to conscious perception. Journal of Experimental Psychology: Human Perception and Performance, 36, 1372–1381.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial Vision 10, 433–436. https://doi.org/10.1163/156856897X00357
Burnham, B. R. (2010). Cognitive load modulates attentional capture by color singletons during effortful visual search. Acta Psychologica, 135, 50–58.
Chun, M. M., & Potter, M. C. (1995). A two-stage model for multiple target detection in rapid serial visual presentation. Journal of Experimental Psychology: Human Perception and Performance, 21, 109–127. https://doi.org/10.1037/0096-1523.21.1.109
Ciesielski, B. G., Armstrong, T., Zald, D. H., & Olatunji, B. O. (2010). Emotion modulation of visual attention: Categorical and temporal characteristics. PLoS ONE, 5, e13860. https://doi.org/10.1371/journal.pone.0013860
de Fockert, J. W., & Bremner, A. J. (2011). Release of inattentional blindness by high working memory load: Elucidating the relationship between working memory and selective attention. Cognition, 121, 400–408. https://doi.org/10.1016/j.cognition.2011.08.016
de Fockert, J. W., Rees, G., Frith, C. D., & Lavie, N. (2001). The role of working memory in visual selective attention. Science, 291, 1803–1806. https://doi.org/10.1126/science.1056496
Erthal, F. S., De Oliveira, L., Mocaiber, I., Pereira, M. G., Machado-Pinheiro, W., Volchan, E., & Pessoa, L. (2005). Load-dependent modulation of affective picture processing. Cognitive, Affective, & Behavioral Neuroscience, 5, 388–395. https://doi.org/10.3758/CABN.5.4.388
Evans, K. K., & Treisman, A. (2005) Natural cross-modal mappings between visual and auditory features. Journal of Experimental Psychology: Human Perception and Performance, 31, 1476–1492. https://doi.org/10.1037/0096-1523.31.6.1476
Giesbrecht, B., & Di Lollo, V. (1998). Beyond the attentional blink: Visual masking by object substitution. Journal of Experimental Psychology: Human Perception and Performance, 24, 1454–1466. https://doi.org/10.1037/0096-1523.24.5.1454
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 6, 65–70. Retrieved from www.jstor.org/stable/4615733
Kelley, T. A., & Lavie, N. (2010). Working memory load modulates distractor competition in primary visual cortex. Cerebral Cortex, 21, 659–665.
Kennedy, B. L., Rawding, J., Most, S. B., & Hoffman, J. E. (2014). Emotion-induced blindness reflects competition at early and late processing stages: An ERP study. Cognitive, Affective, & Behavioral Neuroscience, 14, 1485–1498.
Kim, S.-Y., Kim, M.-S., & Chun, M. M. (2005). Concurrent working memory load can reduce distraction. Proceedings of the National Academy of Sciences, 102, 16524–16529. https://doi.org/10.1073/pnas.0505454102
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2008). International Affective Picture System (IAPS): Affective ratings of pictures and instruction manual (Technical Report No. A-8). Gainesville, FL: University of Florida, NIMH Center for Research in Psychophysiology.
Langton, S. R. H., Law, A. S., Burton, A. M., & Schweinberger, S. R. (2008). Attention capture by faces. Cognition, 107, 330–342. https://doi.org/10.1016/j.cognition.2007.07.012
Lavie, N., & de Fockert, J. (2005). The role of working memory in attentional capture. Psychonomic Bulletin & Review, 12, 669–674. https://doi.org/10.3758/BF03196756
Lavie, N., Hirst, A., de Fockert, J. W., & Viding, E. (2004). Load theory of selective attention and cognitive control. Journal of Experimental Psychology: General, 133, 339–354. https://doi.org/10.1037/0096-3445.133.3.339
McHugo, M., Olatunji, B. O., & Zald, D. H. (2013). The emotional attentional blink: What we know so far. Frontiers in Human Neuroscience, 7, 151. https://doi.org/10.3389/fnhum.2013.00151
Most, S. B., Chun, M. M., Widders, D. M., & Zald, D. H. (2005). Attentional rubbernecking: Cognitive control and personality in emotion-induced blindness. Psychonomic Bulletin & Review, 12, 654–661. https://doi.org/10.3758/BF03196754
Most, S. B., & Jungé, J. A. (2008). Don’t look back: Retroactive, dynamic costs and benefits of emotional capture. Visual Cognition, 16, 262–278.
Most, S. B., Smith, S. D., Cooter, A. B., Levy, B. N., & Zald, D. H. (2007). The naked truth: Positive, arousing distractors impair rapid target perception. Cognition and Emotion, 21, 964–981.
Most, S. B., & Wang, L. (2011). Dissociating spatial attention and awareness in emotion-induced blindness. Psychological Science, 22, 300–305.
Müller, M. M., Andersen, S. K., & Keil, A. (2008). Time course of competition for visual processing resources between emotional pictures and foreground task. Cerebral Cortex, 18, 1892–1899. https://doi.org/10.1093/cercor/bhm215
Nieuwenhuis, S., & de Kleijn, R. (2011). Consciousness of targets during the attentional blink: A gradual or all-or-none dimension? Attention, Perception, & Psychophysics, 73, 364–373. https://doi.org/10.3758/s13414-010-0026-1.
Olatunji, B. O., Ciesielski, B. G., Armstrong, T., Zhao, M., & Zald, D. H. (2011). Making something out of nothing: Neutral content modulates attention in generalized anxiety disorder. Depression and Anxiety, 28, 427–434.
Olivers, C. N. L., & Nieuwenhuis, S. (2006). The beneficial effects of additional task load, positive affect, and instruction on the attentional blink. Journal of Experimental Psychology: Human Perception and Performance, 32, 364–379. https://doi.org/10.1037/0096-1523.32.2.364
Overgaard, M., Rote, J., Mouridsen, K., & Ramsøy, T. Z. (2006). Is conscious perception gradual or dichotomous? A comparison of report methodologies during a visual task. Consciousness and Cognition, 15, 700–708.
Park, S., Kim, M.-S., & Chun, M. M. (2007). Concurrent working memory load can facilitate selective attention: Evidence for specialized load. Journal of Experimental Psychology: Human Perception and Performance, 33, 1062–1075. https://doi.org/10.1037/0096-1523.20.5.1062
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10, 437–442. https://doi.org/10.1163/156856897X00366
Phelps, E. A., Ling, S., & Carrasco, M. (2006). Emotion facilitates perception and potentiates the perceptual benefits of attention. Psychological Science, 17, 292–299. https://doi.org/10.1111/j.1467-9280.2006.01701.x
Piech, R., McHugo, M., Smith, S., Dukic, M., Meer, J., Abou-Khalil, B., ... Zald, D. (2011). Attentional capture by emotional stimuli is preserved in patients with amygdala lesions. Neuropsychologia, 49, 3314–3319.
Piech, R., Pastorino, M., & Zald, D. (2010). All I saw was the cake: Hunger effects on attentional capture by visual food cues. Appetite, 54, 579–582.
Pourtois, G., Grandjean, D., Sander, D., & Vuilleumier, P. (2004). Electrophysiological correlates of rapid spatial orienting toward fearful faces. Cerebral Cortex, 14, 619–633.
Ramsøy, T. Z., & Overgaard, M. (2004). Introspection and subliminal perception. Phenomenology and the Cognitive Sciences, 3, 1–23.
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992). Temporary suppression of visual processing in an RSVP task: An attentional blink? Journal of Experimental Psychology: Human Perception and Performance, 18, 849–860. https://doi.org/10.1037/0096-1523.18.3.849
Schönwald, L. I., & Müller, M. M. (2014). Slow biasing of processing resources in early visual cortex is preceded by emotional cue extraction in emotion–attention competition. Human Brain Mapping, 35, 1477–1490.
Sergent, C., & Dehaene, S. (2004). Is consciousness a gradual phenomenon? Evidence for an all-or-none bifurcation during the attentional blink. Psychological Science, 15, 720–728.
Smith, S. D., Most, S. B., Newsome, L. A., & Zald, D. H. (2006). An emotion-induced attentional blink elicited by aversively conditioned stimuli. Emotion, 6, 523–527.
Sy, J. L., Marois, R., & Tong, F. (2018). Conscious perception can be both graded and discrete. Manuscript submitted for publication.
Theeuwes, J., Atchley, P., & Kramer, A. F. (2000). On the time course of top-down and bottom-up control of visual attention. In S. Monsell & J. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 105–124). Cambridge, MA: MIT Press.
Theeuwes, J., & Van der Stigchel, S. (2006). Faces capture attention: Evidence from inhibition of return. Visual Cognition, 13, 657–665.
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2001). Effects of attention and emotion on face processing in the human brain: An event-related fMRI study. Neuron, 30, 829–841. https://doi.org/10.1016/S0896-6273(01)00328-2
Vuilleumier, P., & Schwartz, S. (2001). Emotional facial expressions capture attention. Neurology, 56, 153–158.
Wang, L., Kennedy, B. L., & Most, S. B. (2012). When emotion blinds: A spatiotemporal competition account of emotion-induced blindness. Frontiers in Psychology, 3, 438. https://doi.org/10.3389/fpsyg.2012.00438
Wierda, S. M., van Rijn, H., Taatgen, N. A., & Martens, S. (2010). Distracting the mind improves performance: An ERP study. PLoS ONE, 5, 15024. https://doi.org/10.1371/journal.pone.0015024
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Keefe, J.M., Sy, J.L., Tong, F. et al. The emotional attentional blink is robust to divided attention. Atten Percept Psychophys 81, 205–216 (2019). https://doi.org/10.3758/s13414-018-1601-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1601-0