
Abstract
To understand how different management interventions influence the forest structure and biodiversity, various components of vascular plant communities were studied and compared in differently managed forest stands and clearings. The number of species, the percentage of the overall cover of alien species, different Raunkier’s life forms’, CRS strategists, the amount of litter and canopy openness were monitored and used for assessment of forest stands condition. We discovered that the species richness and composition of intensively managed forests significantly differ from those with extensive management. Intensive management interventions in commercial plantations, such as mechanical site preparation and the application of herbicides significantly contribute to the decrease of diversity and the spread of alien and ruderal plant species at the expense of native species. On the other hand, when new stands are founded and cared for sensitively, common types of floodplain forests show good regeneration abilities and their species diversity is preserved or quickly renewed. Thus, extensive floodplain forest management results in higher biodiversity and better conditions for the fulfillment of both wood-producing and non-wood-producing functions as well as for sustainable management. We further propose that mechanical site preparation with soil milling and application of herbicides may have a very strong negative impact on the overall biodiversity in forest plantations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Lowland floodplain forests dominated by oaks cover approximately 7% of the total forest area in the Czech Republic. These are important land-use types due to their wood production (Wenger et al. 1990), but mainly non-productive functions (Ward 1998). The largest complex of floodplain forest in the Czech Republic is still preserved in South Moravia (Miklín and Čížek 2014) and it was this area of forest habitats that was surveyed for this study.
The floodplain forest ecosystem was formed by a specific water regime in the given localities (Piegay 2003), such as the undulating groundwater level and floods (Gurnell and Petts 2002). The characteristic vegetation composition and structure in native stands changes, especially, in connection with the frequency of floods (Ward 1998). The differences in the occurrence of plant species in floodplain forests are often also linked to diaspores being washed down from the higher parts of the river basin. In recent decades, this phenomenon has been particularly important for the frequent occurrence of alien species (Řepka et al. 2015). Another common trait is the high concentration of nutrients and the related high production of plant biomass (Maděra and Kovářová 2004). The present structure of the vegetation and floristic composition also reflect the development in the Holocene - human population and coppice management from as early as the Neolithic period (Řepka et al. 2015). Many associated organisms, including the thermophilous species of numerous plants, fungi, and animals have colonized these managed, heterogeneous and often bright habitats (Hédl et al. 2010). Many of those organisms (mainly invertebrates) are specialized for microhabitats and thus sensitive to the changes in the structure of forest stands (Košulič et al. 2016).
It is widely accepted that the current condition of floodplain forest plantations in South Moravia is mainly the result of changes in the hydrological conditions caused by water management adjustments. These have been carried out since the 1970s in the study area in the lower reach of the River Svratka. The steady decline of Quercus robur L. in the tree layers, together with the declining level of groundwater, is particularly shown by Vrška et al. (2006) and Maděra (2001a). Another important point in the changes of floodplain forest ecosystems is the intensification of forest management practices since the second half of the nineteenth century (Janík et al. 2008). Floodplain forests are also strongly invaded by alien species, which significantly influence the size and spatial layout of the autochthonous species (Chytrý et al. 2009).
Despite all these changes, floodplain forests are endangered yet still species-rich communities, in comparison to other types of Central European forest vegetation, and still strongly react to the changes in the environment (Maděra 2001a, b; Chytrý et al. 2009). At the present time, commercial plantations are planted in stands of different ages (forest cycling; e.g. Purchart et al. 2013), maintaining the heterogenous mosaic of spatially divided stands, which create a number of different habitats – from open xerothermic sites, over more shaded shrubland, to strongly shaded stands with closed herb and tree layers (Mullen et al. 2008). Thus, it may be logically assumed that, under certain conditions, the site heterogeneity can substantially increase landscape biodiversity and create suitable conditions for both heliophytes and typical sciophytes of lowland forests. However, the management of such stands (e.g. timber logging, mechanical site preparation) may have an impact on biotic and abiotic conditions which, in turn, affect their plant and animal communities (Bengtsson et al. 2000).
The aim of this study is to assess the influence of management intensity on the vegetation of clearings and the latter stages of forest stands. We also investigate the changes in species and functional diversity during the successional development of commercial lowland floodplain forests, and protected non-regulated forest stands based on the number of species, the percentage cover of alien species (Pyšek et al. 2002), different Raunkier’s life forms (Raunkiaer 1934), and CRS strategists (Grime 1979).
Material and methods
Characteristics of the studied territory
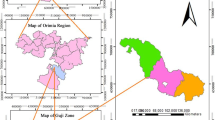
The model territory is situated in the South Moravian Region of the Czech Republic (Fig. 1), in the alluvium of the lower reach of the River Svratka in the vicinity of the villages of Vranovice, Pouzdřany and Ivaň. The Dolní Mušovský luh Natural Monument and the Plačkův les and říčka Šatava Natural Reserve, which were established in 1990, are situated in this territory. According Mucina et al. (2016) is the largest part of the monitored area covered by hardwood floodplain forest vegetation, Fraxino-Quercion roboris, often with altered tree layers.

The position of the studied territory located in southern Moravia of the Czech Republic and layout of the study plots. The second number represents categorization according to Table 1; the first number represents the number of study plots in a category. The map background was downloaded from free maps platform system (https://app.gisonline.cz/chytre-mapy/) and modified in Adobe Photoshop CS4
The floodplain forests in the alluvium of the lower reach of the River Svratka are important floodplain forest complexes in the Czech Republic. The more frequent occurrence of thermophilous species and the potential high species richness, which make it different from other types of floodplain forests in Northern and Central Moravia and Bohemia (Řepka et al. 2015), provide a sufficient species pool for the assessment of different influences on species and functional diversity.
Study plots
The research was conducted during the vegetation periods of 2015 and 2016. All sites studied were afforested with at least 90% representation of Quercus robur. All the study plots were divided into 10 categories according to the age and management type (Table 1). Two types of management were recognized, intensive/commercial and extensive/natural. There was herbicide application and wide area mechanical soil preparation on intensively managed clearings and thinning in commercial stands. All commercial forests evolved from intensively managed clearings. Extensive managed/natural clearings or stands were without soil preparation, mowing of unwanted vegetation and thinning. All native stands evolved from extensively managed clearings or natural regeneration. The plots classified in categories 1 to 4 (extensive clearings and natural stands) are located in the Dolní Mušovský Luh Natural Monument and the Plačkův les and říčka Šatava Natural Reserve and are all managed extensively. The plots in categories 5 to 10 are commercial forests used for timber harvesting and are managed intensively by mechanical soil preparation and herbicide application. The layout of the plots studied is shown in Fig. 1.
In total, 26 study plots were established in 10 categories and 2 phytosociological relevés were made on each study plot, which means that 52 phytosociological relevés were made. All the categories and numbers of plots and relevés per category are recorded in Table 1. The phytosociological relevés on the plots studied were 10 × 20 m in size. The transformed nine-point Braun-Blanquet scale was used to evaluate the abundance and dominance of plants (Westhoff and Van der Maarel 1978). Canopy openness and overall layer of litter were determined for each plot.
Environmental variables
Canopy openness evaluation
The light volume gradient was calculated using imaging software (GAP Light Analyzer, version 2.0) to extract canopy structural parameters and gap light transmission indices from true-color fisheye photographs (Frazer et al. 1999). All the hemispherical fisheye photographs were taken at ground level from the center of the plots within a single day in order to rule out seasonal effects (July 14, 2015 and 2016). This date was selected because the canopy was fully developed during this period.
Litter
Amount of litter on each plot was measured from random samples, always taken within a study plot of phytosociological relevés, with five repetitions per plot. Litter depth of each plot was determined as the mean of these five samples.
Evaluation of phytosociological data
All the phytosociological relevés, including the environmental characteristics measured, were entered in the Turboveg for Windows database program (Hennekens and Schaminee 2001) and then transported into the JUICE 6.0 program (Tichý 2002). In this program, the nomenclature of the species found was unified, based on Kubát et al. (2002). The species number per releve, percentage of the cover of archaeophytes, neophytes and native species based on Pyšek et al. (2002), the percentage of cover of plant life forms (Raunkiaer 1934), and the percentage of the cover of CRS strategists (Grime 1979) out of the overall cover were calculated for each phytosociological relevé. For the calculation of the percentage of cover, the formula \( \frac{Tc}{Sc}\ast 100= Pc \) was used in which Tc is the summation of the cover of all the species recorded in the phytosociological relevé, Sc is the summation of the cover of all the species in the relevé categorised in one group (e.g. all archeophytes) and Pc is the percentage of the cover of the given group.
All the phytosociological relevés were assigned to plant associations, using the expert system implemented in the Juice software (Tichý 2002) for determination of associations and then manually assigned to plant alliances. The names of syntaxons were adapted according to Mucina et al. (2016). The analyses were performed within the JUICE 6.0 program (Tichý 2002).
Statistical analyses
Univariate analyses were performed using R (R Development Core Team 2018), and multivariate analyses were evaluated using CANOCO 5 (ter Braak and Šmilauer 2012). Abovementioned characteristics were consequently used to express the differences between intensive and extensive clearings (0 to 5 years old) and between natural and commercial adult forest stands (50 to 120 + years old). Comparison of species richness among the four forest types was analyzed using generalized linear models (GLMs). This method is intended for analysis of non-normally distributed data with counts (Lepš and Šmilauer 2016). Due to the presence of overdispersion, GLM with negative binomial distribution (GLM-nb) within the “MASS” package was used. This model offers an alternative to the Poisson model with overdispersion (Pekár and Brabec 2009). Dependent variable was species richness per plot and the explanatory variables were the forest types. Differences in percentage cover of species origins, life forms and CRS strategies among four forest types were tested by one-way analysis of variance (ANOVA). This model is used for data with a normal distribution (Lepš and Šmilauer 2016). The percentage covers were logit-transformed to obtain normality. Logit transformation was performed within the “car” package with using adjustment factor to avoid proportions of 0 or 1. The logit transformation is proposed as suitable method when analyzing proportional data in ecology (Warton and Hui 2011). Linear models with logit-transformed percentage cover of species origins, life forms and CRS strategies per plot as the dependent variable were used with forest type as explanatory variables. Post-hoc comparisons among management methods were conducted through treatment contrasts (Pekár and Brabec 2009).
Detrended correspondence analysis (DCA) was used to visualize the heterogeneity in the species composition of vascular plants in differently managed forests stands and clearings (extensive, intensive).
Variation in species composition in relation to management intensity (extensive, intensive), canopy openness and litter amount was examined by canonical correspondence analysis (CCA). The data from the first and second age stages were also analyzed separately in order to clarify the direct effect of management on vegetation of clearings in real time. For this analysis, the canopy openness was excluded. Canonical correspondence analysis (CCA) was used because an initial analysis of the data using detrended correspondence analysis suggested a unimodal response by the species (Šmilauer and Lepš 2014). Gradient length for the first DCA axis was 3.52 (stands and clearings) and 4.49 (clearings). Before analysis, logarithmic transformation of species cover values was performed. The significance of the effect of management intensity and environmental variables was tested using Monte Carlo permutation tests with 999 permutations. First, a model with all selected variables was performed to obtain the total explained variability. In order to detect marginal effects of selected variables, their significance was tested separately. Conditional effects of individual variables were assessed by successively entering competing variables as a covariate (Šmilauer and Lepš 2014). The results are presented in Table 2.
Results
Species number and their origin
Out of all 181 species – out of which 129 are native to the Czech territory – 31 archaeophytes and 21 neophytes were recorded in the researched plots (Fig. 2). The most common neophyte and archaeophytes were Erigeron annuus and Solidago gigantea, and Capsella bursa-pastoris, Carduus crispus and Cirsium arvense, respectivelly.

Species richness in phytosociological relevés. Letters in x-axis are as follow: A – Extensive clearings, B – Extensive stands, C – Intensive clearings, D – Intensive stands. Horizontal lines on bars indicate median values, box boundaries show quartiles, whiskers denote 1.5 times the interquartile range. Different letters indicate statistically significant differences (P < 0.050)
The total number of species differed significantly among forest types (χ23 = 60.65, P < 0.001; Fig. 2). The total number of species in adult intensive stands was significantly lower than in the other monitored types (contrasts, P < 0.001). Conversely, the numbers of species in native stands, extensive and intensive clearings showed no significant differences (contrasts, P > 0.050).
The percentage cover of archaeophytes differed significantly among monitored forest types (F3,34 = 28.14, P < 0.001; Fig. 3a). The percentage cover of archaeophytes in intensive clearings ranged from 4 to 72% and as expected, was highest there (contrasts, P < 0.001). There was significantly higher percentage cover of archaeophytes in extensive clearings than in extensive stands (contrasts, P = 0.017) but its percentage cover did not differ significantly from this of intensive stands (contrasts, P = 0.051). Stands had similar percentage cover of archaeophytes (contrasts, P = 0.453).

The percentage cover of non-native and native vascular plants: (a) archeophyte, (b) neophyte, (c) natural species in different stand categories. Letters in x-axis are as follow: A – Extensive clearings, B – Extensive stands, C – Intensive clearings, D – Intensive stands. Combination of letters show non-significant difference between each of the categories. Horizontal lines on bars indicate median values, box boundaries show quartiles, whiskers denote 1.5 times the interquartile range. Different letters indicate statistically significant differences (P < 0.050)
There was a significant difference in the percentage cover of neophytes among forest types (F3,34 = 4.04, P = 0.014; Fig. 3b). The percentage cover of neophytes in intensive clearings was significantly higher than in stands (contrasts, P < 0.050), but it did not differ from that in extensive clearings (contrasts, P = 0.124). The other monitored forest types had similar percentage cover of neophytes (contrasts, P > 0.050).
The percentage cover of native species differed significantly by forest types (F3,34 = 27.33, P < 0.001; Fig. 3c). Native species had significantly lowest percentage cover in intensive clearings in comparison with extensive clearings and stands (contrasts, P < 0.001). There was no significant difference in the percentage cover of native species among stands (contrasts, P = 0.606) but there was higher percentage cover in comparison to extensive clearings (contrasts, P < 0.050). The most common native species were Galium aparine, Glechoma hederacea, Lamium maculatum and Urtica dioica.
Life forms of plants
The percentage cover of life-forms shows the apparent changes in the dominance of individual groups in native and commercial stands and clearings (Fig. 4).

The percentage cover of life forms of plants in different stand categories: (a) nano phanerophytes, (b) phanerophytes, (c) geophytes, (d) therophytes, (e) hydrophytes. Letters in x-axis are as follow: A – Extensive clearings, B – Extensive stands, C – Intensive clearings, D – Intensive stands. Horizontal lines on bars indicate median values, box boundaries show quartiles, whiskers denote 1.5 times the interquartile range. Different letters indicate statistically significant differences (P < 0.050)
Percentage cover of nano phanerophytes differ significantly among monitored types of stands and clearings (F3,36 = 9.39, P < 0.001; Fig. 4a). The percentage cover of nano phanerophytes was the highest in native stands (contrasts, P < 0.05). The difference in the percentage cover of nano phanerophytes between intensive clearings and intensive stands was not significant (contrasts, P = 0.738), however it was significantly lower than in extensive clearings (contrasts, P = 0.111). The most dominant nano phanerophytes were Cornus sanguinea and Viburnum opulus.
There was a significant difference in percentage cover of phanerophytes among monitored forest types (F3,36 = 15.64, P < 0.001; Fig. 4b). The percentage cover of phanerophytes was highest in stands (contrasts, P < 0.001). Percentage cover of phanerophytes in intensive clearings was comparable to that in extensive clearings but was lower than in extensive and intensive stand (contrasts, P = 0.195).
Percentage cover of hemicryptophytes was not significantly different (F3,36 = 2.006, P = 0.130) in the monitored types of plots as it ranges from 12 to 86% in extensive clearings, from 5 to 59% in intensive clearings, from 9 to 58% in natural forests, and from 21 to 70% in commercial forests. The most dominant hemicryptophytes were Brachypodium sylvaticum, Glechoma hederacea, Urtica dioica.
Percentage cover of geophytes varied significantly among forest types (F3,36 = 12.85, P < 0.001; Fig. 4c). The percentage cover of geophytes was highest in intensive clearings (contrasts, P < 0.050) where their cover reached 3 to 64%. In extensive clearings (2 to 16%), there was higher percentage cover of geophytes than in intensive stands (0 to 6%) (contrasts, P = 0.048), but it did not differ significantly from that in extensive stands (contrasts, P = 0.438). There was no significant difference in the percentage cover of geophytes among stands (contrasts, P = 0.242). Most dominant geophyte was Cirsium arvense and Ficaria verna subsp. bulbifera.
Percentage cover of therophytes differ significantly among monitored forest types (F3,36 = 8.71, P < 0.001; Fig. 4d). The highest percentage cover of therophytes (10 to 33%) was again observed in intensive clearings (contrasts, P < 0.050), whereas it did not differ significantly among other monitored forest types (contrasts, P > 0.050). Most dominant therophytes were Amaranthus retroflexus, Capsella bursa-pastoris and Chenopodium album.
Percentage cover of hydrophytes differed significantly among monitored forest types (F3,36 = 5.66, P = 0.002; Fig. 4e). The percentage cover of hydrophytes was highest in extensive clearings (contrasts, P < 0.050). There was no significant difference in percentage cover of hydrophytes among other monitored forest types (contrasts, P > 0.050). Most dominant hydrophytes were Glyceria fluitans and Phragmites australis.
Representation of the CRS strategist in stands
There was a significant difference in percentage cover of competitors (C) among monitored forest types (F3,34 = 3.96, P = 0.015; Fig. 5a). The lowest percentage cover of competitors was observed in extensive clearings (contrasts, P < 0.050), whereas there was no significant difference among other monitored types of forests (contrasts, P > 0.050). Most dominant competitors in herb layer were Deschampsia caespitosa, Rubus caesius and Urtica dioica.

The percentage cover of CRS plant life strategies in different stand categories: (a) C strategist, (b) R strategist, (c) S strategist. Letters in x-axis are as follow: A – Extensive clearings, B – Extensive stands, C – Intensive clearings, D – Intensive stands. Horizontal lines on bars indicate median values, box boundaries show quartiles, whiskers denote 1.5 times the interquartile range. Different letters indicate statistically significant differences (P < 0.050)
The percentage cover of ruderal species (R) varied significantly among forest types (F3,34 = 6.45, P = 0.001; Fig. 5b). The percentage cover of ruderal species was significantly lowest in extensive stand (contrasts, P < 0.050). The difference in the percentage cover of ruderal species among other forest types was not significant (contrasts, P > 0.050). Most common ruderal species were Capsella bursa – pastoris, Polygonum arenastrum and Veronica hederifolia.
The percentage cover of S strategists differed significantly among forest types (F3,34 = 5.47, P = 0.003; Fig. 5c).The percentage cover of S strategists was significantly lowest in intensive clearings (contrasts, P < 0.050). Extensive clearings had significantly higher percentage cover than intensive stands (contrasts, P = 0.037) and similar percentage cover like extensive stands (contrasts, P = 0.076). Stands had comparable percentage cover of S strategists (contrasts, P = 0.836). The typical S strategists were quite rare; the mixed C-S strategists like Brachypodium sylvaticum, Circaea lutetiana and Crex riparia were more common.
Plant aliances
The classification of relevés to alliances is depicted in Table 1. In particular, it is worth noting that most of phytosociological relevés taken in native forests were assigned to the Fraxino-Quercion roboris alliance, regardless of stand age. Phytosociological relevés from freshly planted and very young stands correspond with the Fraxino-Quercion roboris alliance only due to their species composition, not the stand structure.
On the other hand, the phytosociological relevés of the intensive clearings and young commercial forest stands up to 20 years aged, where mechanical site preparation and herbicides were used on the plantation, were assigned to ruderal vegetation. Young stands showed the highest similarity to the Atriplicion alliance, while the older stands were similar to the Aegopodion podagrariae alliance. Both of these phyto-sociological alliances contain mainly ruderal vegetation typical for highly disturbed and unstable habitats under strong anthropogenic influence.
Multivariate analysis
Figure 6a shows the results of DCA that revealed a major difference between communities of vascular plants in extensively and intensively managed plots of old forest stands. The first axis explained 8.9% of the variability, while the second axis explained 5% of the variability in community composition. Tight clustering of intensive plots reflects greater homogeneity than in extensive plots.

(a) DCA diagrams summarazing differences within vascular plant communities of individual relevés according to different management intensity in the stands and clearings: squares – intensive, cirles – extensive; (b) CCA diagrams summarizing differences within vascular plant communities in relation to canopy opennes, litter amount and management intensity; (c) DCA diagrams summarazing differences within vascular plant communities of individual relevés according to different management intensity in the clearings: squares – intensive, cirles – extensive; (d) CCA diagrams summarizing differences within vascular plant communities in relation to litter amount and management intensity. Fifty best fitted plant species are plotted. Species names are abbreviated by the first four letters of the genus and species names in graph
Canonical correspondence analysis (CCA) revealed a significant effect of canopy openness, litter amount and management intensity on community composition of vascular plants in forest stands and clearings (CCA, pseudo-F = 2.3, P = 0.001; Fig. 6b). The first three (constrained) axes explained 15.8% of the variability and the remaining (unconstrained) axes explained 7.7% of the variability. Canopy opennes had the largest marginal effect (6.9%), followed by amount of litter (5.5%) and management (5.2%; Table 2). The second ordination axis distinguished between extensively and intensively managed forests. Extensive forests were characterized by high amount of litter and low canopy opennes, while intensive forests were characterized by high canopy opennes and low amount of litter. On the right side, the ordination biplot showed species related to extensive forests such as Cornus sanguinea, Fraxinus excelsior or Viburnum opulus which rejuvenates naturally in native (especially older) stands, together with Circaea lutetiana and Rubus caesius. Species related to higher intensity in forest stands under increasing level of canopy openness are typical ruderal open habitat species such as Amaranthus retroflexus and Capsella bursa – pastoris.
The DCA revealed difference between communities of vascular plants in the extensively and intensively managed plots of forest clearings (Fig. 6c). The first axis explained 12.2% of the variability and the second axis explained 8.2% of the variability in the community composition. The tight clustering of intensive plots confirmed that extensive plots are more heterogeneous than intensive one.
Canonical correspondence analysis (CCA) showed a significant effect of litter amount and management intensity on community composition of vascular plants in clearings (CCA, pseudo-F = 1.8, P = 0.001; Fig. 6d). The first two (constrained) axes explained 17.2% of the variability, while the other two (unconstrained) axes explained 11.2% and 8.4% of the variability in the community composition. Amount of litter had larger marginal effect (9.4%) than management (8.3%; Table 2). The second ordination axis distinguished between extensively and intensively managed clearings. Extensive clearings were characterized by high amount of litter, on the contrary intensive clearings were characterized by low amount of litter. The ordination biplot showed species related to extensive clearings on the right side, while species related to intensive clearings showed on the left side. The high intensity of management is associated with the occurrence of Capsella bursa-pastoris, Circaea lutetiana and Cirsium arvense which are typical for degraded habitats under strong level of anthropogenic pressure. On the other hand, the high amount of litter in extensive plots is associated with species of humid and mesophytic forests that are able to survive the disturbance of wood harvesting and the founding of clearings without mechanical site preparation (e.g. soil milling, herbicide applications) like Carex riparia, Festuca gigantea and Scirpus sylvaticus.
Discussion
The results of this study show that native floodplain forests are an interesting and species-rich forest community, where 181 species – out of which 129 are native to the Czech territory – 31 archaeophytes and 21 neophytes (Pyšek et al. 2002) were recorded in the researched plots.
Previous studies have shown that floodplain forests react sensitively to changes in environmental conditions and management (Buček and Lacina 1995; Piegay 2003) and that occasional strong disturbance is a natural part of their ecosystems (Bengtsson et al. 2000). The current biodiversity is the result of a long-term natural development (Thomas et al. 1979) as well as human influence (Košulič et al. 2014, 2016). In the past decades, especially after the influence of floods had been minimized by regulations of river flow, the effect of natural disturbance has declined. At the same time, the intensity of anthropogenic disturbance has increased and its character has changed (Piegay 2003).
We discovered that a significant number of native species in floodplain forests is linked to extensively managed clearings and native stands and we recorded a sudden rise in the cover of hydrophytes and S-strategists. The differences in occurrence of hydrophytes in natural and commercial localities are probably related to the micro-topography (Simmons et al. 2011). In the studied locations, the terrain in natural forests and especially in extensive clearings appears to be much more ragged and therefore hydrophytes can find a suitable niche in terrain depressions (e.g. holes caused by windthrows). Moreover, we observed that the surface in commercial stands is much more flat, probably as a result of milling and other terrain adjustments carried out by silviculture activities during stand establishment. We suggest that these activities maintain various level of micro-topography, thus significantly affect species diversity and compositional variation in hard-wood floodplain forest as shown also by other studies (Janík et al. 2008; Miklín and Čížek 2014).
On the other hand, the cover of ruderal species is significantly lower in native stands than in commercial stands and intensive clearings. It can be explained by a higher representation of nanophanerophytes in natural stands, since nanophanerophytes, which are not classified as ruderal species, competitively suppress short-lived ruderal species (Maděra 2001b).
According to our results, the stress tolerant herb layer species manage to survive the disturbance connected with the creation of extensive forest clearings, and are consequently able to regenerate in the cleared space (for example Carex riparia, Lythrum salicaria or Scirpus sylvaticus). However, they have low occurrence on intensive clearings where the dominant life form are geophytes. It is obvious, that wood harvesting, unlike mechanical site preparation and herbicide application, is apparently not a limiting disturbance to these stress tolerant species.
The dominance of geophytes on intensive clearings is likely caused by milling which leads to fragmentation of underground storage organs and the consequent regeneration of the seperated pieces as has also been shown by other authors (Lososová et al. 2003). The fragments of storage organs, deprived of leaves and stems during mulching, are also less exposed to the effects of herbicides, which leads to the advantegement of geophytes at the expense of other life forms.
In the field, we observed that litter is cut into tiny pieces during milling which quickly decompose or become buried under the ground which makes it impossible to measure. The presence of litter can be therefore partly understood as an expression of absent soil milling and mechanical site preparation in the forest undergrowth (Newmaster et al. 2007). For this reason, we assume that the amount of litter is not as important for the distribution of S-strategists and hydrophytes as the absence of milling. On the other hand, this assertion should be more tested in future studies focused also in other forest habitats affected by soil milling (e.g. Löf et al. 2012).
We suggest that non-forest species (mostly archaeophytes or neophytes) disappear during succesion development but in native stands they are replaced with forest species of more advanced succession stages, therefore the species diversity will not significantly decrease. According to the study by Řepka et al. (2015), we can expect a decrease in plant species diversity in native stands as late in age 140 years. A number of researchers have explained this decrease by a long-term lack of disturbances, leading to the displacement of the competitively weaker species (Denslow 1980; Huston 1979), and a considerable uniformity of older stands. We have also noted a significant rise of competitive strong species in adult natural stands.
The results of the classification of relevés to alliances show that there are no areas covered with a ruderal community in extensively managed clearings and the forest ecosystem operates continuously. We suggest that mechanical soil preparation and herbicide application cause replacing of forest vegetation by non-forest ruderal communities with a minimum number of floodplain-forest species, but with a strong presence of certain alien species. The ruderal communities persist for at least 15 years. Similar conclusions were also reached by Miklín and Čížek (2014).
In adult commercial stands, as well as in natural stands, a number of species which dominated in intensive clearings disappear. However, they are not replaced by the same number of forest species, only the cover of some surviving tolerant species rises. Brunet and Von Oheimb (1998) states that the number and percentage cover of woodland species in recent woods (founded on agricultural soil) decreased with increasing distance to ancient woods, which happens even with stands 30 to 75 years old. Brunet and Von Oheimb (1998) also states that median migration rates calculated on the occurrence of the farthest individual is only 0.53 m−1 /year. We can therefore assume that also in the territory monitored by us, many forest species do not manage to migrate between adult stands and replace the populations destroyed by milling and herbiciding, especially if the individual adult stands are separated by migration barriers in the form of intensive clearings.
It follows from our study that anthropogenic disturbance can have a positive, but also a very negative impact on vegetation. The study also shows that maintaining proper (environmentally-friendly) methods of restoring alluvial forests has a remarkable regenerative capacity (Janík et al. 2008). Many plant species in floodplain forests have adapted to the occasional strong disturbance of removing (harvesting) the tree layer (Wenger et al. 1990; Ward 1998). However, our results clearly confirmed that mechanical site preparation presented by soil milling and the subsequent application of herbicides during the establishment of new stands, together with planting large areas of homogeneous stands, are the greatest obstacles to maintaining high species diversity in floodplain forests (Miklín and Čížek 2014).
Conclusion
Our study showed that intensive management interventions in commercial plantations significantly contribute to the decrease of diversity in forest stands and the spread of alien and ruderal plant species at the expense of native species of vascular plants. On the contrary, when new forest stands were founded and cared for sensitively, common types of floodplain forests showed good regeneration properties and their species diversity was preserved or quickly renewed. We therefore recommend that mechanical site preparation and herbicide application on freshly cutted stands should be avoided as it showed the most negative effect on diversity of vascular plants. To conclude, according to our results and the data from other authors (e.g. Ward 1998; Janík et al. 2008; Řepka et al. 2015), high species and functional diversity and high structural heterogeneity are key conditions for the development of resilient stands.
References
Bengtsson J, Nilsson SG, Franc A, Menozzi P (2000) Biodiversity, disturbances, ecosystem function and management of European forests. For Ecol Manag 132:39–50. https://doi.org/10.1016/S0378-1127(00)00378-9
Brunet J, Von Oheimb G (1998) Migration of vascular plants to secondary woodlands in southern Sweden. J Ecol 86:429–438. https://doi.org/10.1046/j.1365-2745.1998.00269.x
Buček A, Lacina J (1995) Diferenciace krajiny v geobiocenologickém pojetí a její aplikace v krajinném plánování při navrhování územních systémů ekologické stability. Zpr Čes Bot Společ 12:92–102 [in Czech]
Chytrý M, Wild J, Pyšek P, Tichý L, Danihelka J, Knollová I (2009) Maps of the level of invasion of the Czech Republic by alien plants. Preslia 81:187–207
Denslow JS (1980) Patterns of plant species diversity during succession under different disturbance regimes. Oecologia 46:18–21
Frazer GW, Canham CD, Lertzman KP (1999) Gap Light Analyzer (GLA), version 2.0: imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, user’s manual and program documentation. Simon Fraser University, British Columbia and the Institute of Ecosystem Studies, New York
Grime JP (1979) Plant strategies and vegetation processes. Wiley, Chichester, New York, Brisbane, Toronto
Gurnell AM, Petts GE (2002) Island-dominated landscapes of large floodplain rivers, a European perspective. Freshw Biol 47:581–600. https://doi.org/10.1046/j.1365-2427.2002.00923.x
Hédl R, Kopecký M, Komárek J (2010) Half a century of succession in a temperate Oakwood: from species-rich community to Mesic forest. Divers Distrib 16:267–276. https://doi.org/10.1111/j.1472-4642.2010.00637.x
Hennekens SM, Schaminee JH (2001) Turboveg, a comprehensive database management system for vegetation data. J Veg Sci 12:589–591. https://doi.org/10.2307/3237010
Huston M (1979) A general hypothesis of species diversity. Am Nat 113:81–101
Janík D, Adam D, Vrška T, Hort L, Unar P, Král K, Šamonil P, Horal D (2008) Tree layer dynamic of the Cahnov – Soutok, near natural floodplain forest after 33 years (1973 – 2006). Eur J For Sci 127:337–345. https://doi.org/10.1007/s10342-008-0210-1
Košulič O, Michalko R, Hula V (2014) Recent artificial vineyard terraces as a refuge for rare and endangered spiders in modern agricultural landscape. Ecol Eng 68:137–154. https://doi.org/10.1016/j.ecoleng.2014.03.030
Košulič O, Michalko R, Hula V (2016) Impact of canopy openness on spider communities: implications for conservation management of formerly coppiced oak forests. PLoS One. https://doi.org/10.1371/journal.pone.0148585
Kubát K, Hrouda L, Chrtek J, Kaplan Z, Kirschner J, Štěpánek J (2002) Klíč ke květeně České Republiky. Academia, Praha [in Czech]
Lepš J, Šmilauer P (2016) Biostatistika. Episteme, Jihočeská univerzita, České Budějovice [in Czech]
Löf M, Dey DC, Navarro RM, Douglass FJ (2012) Mechanical site preparation for forest restoration. New For 43:825–848. https://doi.org/10.1007/s11056-012-9332-x
Lososová Z, Danihelka J, Chytrý M (2003) Seasonal dynamics and diversity of weed vegetation in tilled and mulched vineyards. Biológia 58:49–57
Maděra P (2001a) Effect of water regime changes on the diversity of plant communities in floodplain forests. Ekológia 20:116–129. https://doi.org/10.1111/j.1654-1103.2009.05282.x
Maděra P (2001b) Response of the floodplain forest communities herb layer to changes in the water regime. Biológia 56:63–72
Maděra P, Kovářová P (2004) Primary succession of white willow communities in the supraregional biocorridor in the middle water reservoir of Nové Mlýny. Ekológia 23:191–204
Miklín J, Čížek L (2014) Erasing a European biodiversity hot-spot: open woodlands, veteran trees and mature forests succumb to forestry intensification, succession, and logging in a UNESCO biosphere reserve. J Nat Conserv 22:35–41. https://doi.org/10.1016/j.jnc.2013.08.002
Mucina L, Bültmann H, Dierßen K, Theurillat JP, Raus T, Čarni A, Šumberová K, Willner W, Dengler J, García RG, Chytrý M, Hájek M, Pietro R, Lakushenko D, Pallas J, Daniëls AJF, Bergmeier E, Guerra AS, Ermakov NG, Valachovič M, Schaminée JHJ, Lysenko T, Didukh PY, Pignatti S, Rodwell SJ, Capelo J, Weber EH, Solomeshch A, Dimopoulos P, Aguiar C, Hennekens MS, Tichý L (2016) Vegetation of Europe: hierarchical floristic classification system of vascular plant, lichen, and algal communities. Appl Veg Sci 19:1–264. https://doi.org/10.1111/avsc.12257
Mullen K, O’Halloran J, Breen J, Giller P, Pithon J, Kelly T (2008) Distribution and composition of carabid beetle (Coleoptera, Carabidae) communities across the plantation forest cycle implications for management. For Ecol Manag 256:624–632. https://doi.org/10.1016/j.foreco.2008.05.005
Newmaster SG, Parker WC, Bell FW, Paterson JM (2007) Effects of forest floor disturbances by mechanical site preparation on floristic diversity in a Central Ontario clearcut. For Ecol Manag 246:196–207. https://doi.org/10.1016/j.foreco.2007.03.058
Pekár S, Brabec M (2009) Moderní analýza biologických dat. 1. vydání. Zobecněné lineární modely v prostředí R. Scientia, Praha [in Czech]
Piegay H (2003) Dynamics of wood in large rivers. In: Gregory SV, Boyer KL, Gurnell AM (eds) The ecology and management of wood in world rivers. American Fisheries Society, Bethesda
Purchart L, Tuf IH, Hula V, Suchomel J (2013) Arthropod assemblages in Norway spruce monocultures during a forest cycle - a multi-taxa approach. For Ecol Manag 306:42–51. https://doi.org/10.1016/j.foreco.2013.06.012
Pyšek P, Sádlo J, Mandák B (2002) Catalogue of alien plants of the Czech Republic. Preslia 74:97–186
R Development Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.Rproject.org/. Accessed 17 Apr 2020
Raunkiaer C (1934) The life forms of plants and statistical plant geography. Clarendon Press, Oxford
Řepka R, Šebesta J, Maděra P, Vahalík P (2015) Comparison of the floodplain forest floristic composition of two riparian corridors: species richness, alien species and the effect of water regime changes. Biologia 70:208–217. https://doi.org/10.1515/biolog-2015-0021
Simmons ME, Ben Wu X, Whisenant SG (2011) Plant and soil responses to created microtopography and soil treatments in bottomland hardwood Forest restoration. Restor Ecol 19:136–146. https://doi.org/10.1111/j.1526-100X.2009.00524.x
Šmilauer P, Lepš J (2014) Multivariete analysis of ecological data using Canoco 5. Cambridge University Press, Cambridge
ter Braak CJF, Šmilauer P (2012) Cannoco 5. Software for multivariate data exploration, testing, and summarization. Microcomputer Power, Ithaca
Thomas JW, Maser C, Rodiek JE (1979) Wildlife habitats in managed forests: the Blue Mountains of Oregon and Washington. Forest Service and Agriculture, Washington DC
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–459. https://doi.org/10.1111/j.1654-1103.2002.tb02069.x
Vrška T, Adam D, Hort L, Odehnálková P, Horal D, Král K (2006) Dynamika vývoje pralesovitých rezervací v České republice. II. Lužní lesy – Cahnov-Soutok, Ranšpurk, Jiřina. Academia, Praha [in Czech]
Ward JV (1998) Riverine landscapes: biodiversity patterns, disturbance regimes and aquatic conservation. Biol Conserv 83:269–278. https://doi.org/10.1016/S0006-3207(97)00083-9
Warton DI, Hui FK (2011) The arcsine is asinine: the analysis of proportions in ecology. Ecology 92:3–10. https://doi.org/10.1890/10-0340
Wenger EL, Zinke A, Gutzweiler KA (1990) Present situation of the European floodplain forests. For Ecol Manag 33:5–12. https://doi.org/10.1016/0378-1127(90)90180
Westhoff V, van der Maarel E (1978) The Braun-Blanquet approach. In: Whittaker RH (ed) Classification of plant communities, 2nd edn. Junk, The Hague, pp 287–399
Acknowledgements
We thank the Židlochovice Enterprise, a division of Czech National Forests, Inc., for access to the forest stands around the villages of Vranovice, Ivaň and Pouzdřany, and for their cooperation within the implementation of this research. We are indebted to the following people who assisted in various aspects of this work: Šárka Mašová, Radek Michalko, Radomír Řepka, Kamila Surovcová and Luboš Purchart. We would like to thank the editor and anonymous reviewers for their valuable comments and suggestions on the manuscript. The study was financially supported by the Specific University Research Fund of the FFWT Mendel University in Brno (Reg. numbers: LDF_VT_2016002/2016 and LDF_PSV_2017004/2017).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
In this manuscript, all authors have no actual or potential conflict of interest including financial, personal or other relationships with journal editors. This submitted work has not been inappropriately influenced.
Submission declaration
This manuscript and its contents have not previously been published nor are under active consideration for publication with any journal or publisher except with Biologia.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Košulič, O., Hamřík, T. & Lvončík, S. Patterns of change in the species composition of vascular plants during different succession stages and management intensity of a lowland floodplain forest. Biologia 75, 1801–1813 (2020). https://doi.org/10.2478/s11756-020-00536-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2478/s11756-020-00536-5