
Abstract
In eastern Central Europe the abandonment of traditional land use represents a major threat for biodiversity. Evidence on species loss and shifts in assemblages is often based on butterfly surveys since these are known as sensitive indicators of habitat changes. Butterfly assemblages were studied in meadows of the Transcarpathian lowland in three consecutive years (2012–2014) with standard transect walks in six different sites (two transects/site). More than 6500 individuals of 66 species were recorded. The less disturbed habitats surrounded by natural forests have shown the highest diversity (Shannon-Wiener, dominance profiles). In faunal types the widely distributed, generalist Euro-Siberian species predominated with significant presence of Holo-Mediterranean and southern Continental elements. Three main types of habitats were separated and characterised by indicator species, i.e. we hierarchically classified the species according to their fidelity by the IndVal method. The dry sites were characterised by a few generalist species only, while the humid ones and mostly the transitional sites were inhabited by numerous habitat and/or food plant specialists. The assemblages were compared with multivariate analysis and the concordance of inter-annual changes of the assemblages were surveyed. The concordance profiles of the less diverse dry habitats were clearly separated from others while other sites with dominance profiles with longer sequence of scarce species have also shown similar concordance profiles. The importance of nature-like forest fringe structures was pointed out for both habitat and species conservation. Conservation efforts should be focused to sustain the general level of biodiversity by the preservation of nature-like habitats and the possible re-establishment of some kinds of traditional use.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The relevant references generally agree that we are confronting with a large-scale loss of biodiversity (e.g. Hambler and Speight 2004; Thomas et al. 2004; Thomas 2005). In Central and Western Europe the sinking of species richness is mostly attributed to the loss and fragmentation of nature-like and semi-natural habitats (Harrison and Bruna 1999; Wenzel et al. 2006; Millennium Ecosystem Assessment 2005). Surrounding intensive agriculture also negatively affects the remaining habitats due to increasing load of pesticides and fertilisers (Huemer and Tarmann 2001; Lethmate 2005). In eastern Central Europe, however, the abandonment of traditional land use connected with the decrease of rural population, and the intensification of farming represent the major threats for biodiversity (Cremene et al. 2005; Baur et al. 2006; Schmitt and Rákosy 2007; Csergő et al. 2013; Bubová et al. 2015).
Evidence on species loss and changes in assemblages is often based on butterfly surveys since these are known as sensitive indicators of habitat changes (Erhardt and Thomas 1991; New et al. 1995; MacNally and Fleishman 2004). Butterflies are often food plant and/or habitat specialists, therefore they respond quickly to the changes of vegetation structure and composition (Thomas et al. 2004; Cremene et al. 2005; Rákosy and Schmitt 2011). They are taxonomically and ecologically generally well described, most species can be identified during the field work, they are sufficiently abundant and diverse in most grassland habitats, thus they are relatively easy to sample, and also the monitoring protocols are well elaborated (Scoble 1992; Caro and O’Doherty 1999).
As many butterflies are host-specific, e.g. the myrmecophilous species (e.g. Thomas and Settele 2004; Thomas 2005), they are often considered as umbrella species (Andelman and Fagan 2000; Settele et al. 2005) and used as biodiversity indicators both of plants and other phytophagous insects. Furthermore, specialist (oligo- and/or monophagous) butterfly species are considered to be more affected by environmental changes than generalist ones (Erhardt and Thomas 1991; Steffan-Dewenter and Tscharntke 2002; Tscharntke et al. 2002).
The survey of the inter-annual variation of insect assemblages has a growing importance in the age of human disturbance and climate change (Strange et al. 2011). Recent surveys have shown that those communities display the lowest aggregate variability which include communities with limited fluctuations of species or those in which the compositional variability is sufficient to compensate for fluctuations of individual species (Cottingham et al. 2001; Downing et al. 2008). Conversely, assemblages with the greatest aggregate variability will be those in which individual species’ population growth rates are positively correlated, leading to synchronized species fluctuations or which have limited compositional variability (Houlahan et al. 2007; Valone and Barber 2008). It was recently recognised that communities can display extreme temporal variability in the absence of concordant dynamics. Rank-abundance curves for some trophically similar communities illustrate that a few species may be very abundant, while most of these are comparatively rare (Tokeshi 1993; Magurran 2005). Thus, high community aggregate variability can also arise from high temporal variability of a small number of relatively abundant species even if their population dynamics are not correlated with the rest of the community. Therefore, it is necessary to test whether the inter-annual variations of abundance arise from positively correlated population dynamics of multiple species, against the alternative hypothesis that community abundance patterns are the result of unassociated fluctuations of a small number of abundant species. Additionally, these results should be compared with the rank-abundance curves of the surveyed communities.
We tested these hypotheses in the butterfly assemblages of different meadow types (xeric vs. wet) in a relatively understudied pre-Carpathian region. Our surveys were carried out close to the village Velyka Dobron’, partly in the Velyka Dobron’ Game Reserve but also in some adjacent territories. We were mostly interested in the connections between the composition, seasonal and annual changes of butterfly assemblages. We evaluated our data in connection to the character of the vegetation and tried to forecast the possible changes, formulating some suggestions for the conservation management of the Game Reserve.
We collected data on the frequency of butterfly species in three subsequent years at different sampling sites and analysed the data to test the following hypotheses:
-
i.
We hypothesised that the seasonal numbers of observed individuals mostly depend on the abundance of some polycyclic generalist species.
-
ii.
We also hypothesised that the abundances of generalist species should be basically similar in some consecutive years, at least in the more nature-like habitats. It means that we expect basically concordant changes in individual numbers of such species during the succeeding years.
-
iii.
We hypothesised that these more nature-like habitats show a larger number of nearly equally (sub-)dominant species while the assemblages of disturbed habitats show a more abruptly declining profile.
-
iv.
We also hypothesised that the seasonal and yearly numbers of species show a different pattern due to the fluctuating presence and seasonal dynamics of some specialist species.
To test these hypotheses, in each year (i) we compared the species numbers of butterflies at the different sites, (ii) the proportions of the different faunal elements and ecological components, of the specialist vs. generalist species, and the oligo- vs. polyphagous ones, according to the relevant references (Varga 2011; Varga et al. 2004). Based on the individual numbers we calculated the relative frequencies of species and the Shannon-Wiener diversity indices of assemblages. We characterised the assemblages according to their indicator species by IndVal method and also analysed the dominance profiles of species in the different habitat types. Finally, we surveyed the concordance of the changes of species/individual numbers of species in the succeeding years.
Materials and methods
Study sites
The sites of our surveys are located on the north-eastern edge of the Pannonian lowland (Szatmár-Bereg plain) near to the river Tisza and tributaries which is a traditional, low-input agricultural region with several remnants of the natural and semi-natural vegetation, i.e. lowland oak-hornbeam forests, hard- and softwood gallery forests, humid meadows and forest clearings (Simon 1952), but recently also with abandoned pastures and fallow lands. A considerable part of the area is dissected by drainage canals of the former extensive peatland which existed here until the last decades of the nineteenth century (Boros 1964). The Szatmár-Bereg lowland is – with the exception of some few small volcanic hills – covered by fine-grained fluviatile sediments and deep, clayey alluvial soils. The climate is slightly cooler (annual mean 8.9 °C) and more continental than the average of the Great Hungarian Plain (January −3.4 °C) (Kormány 1976, 2006). The yearly sum of precipitation is also higher, approximately amounting more than 600 mm in average (610–640 mm). The area biogeographically belongs to the Pannonian region, however, with significant Carpathian influences in some groups of terrestrial invertebrates (land gastropods, ground beetles, orthopterans see: Deli et al. 1997; Ködöböcz and Magura 1999; Gálik et al. 2001; Szanyi et al. 2015a).
We characterised the sites in a preliminary publication (Szanyi et al. 2015b and Table 1) as xeric ([i], [ii]), transitional ([iii], [iv]) and wet ([v], [vi]), see Results (Online Resource 1, Table 2 and Figs. 1, 2, 3, 4, 5 and 6).

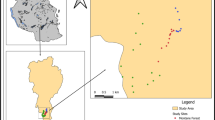
Map of the sampling area with the location of six sampling sites studied between 2012 and 2014 in surroundings of Velyka Dobron’ (West Ukraine, Transcarpathia). Codes, and characterisation of sites are detailed in the text. Dark grey: forests; black: sampling sites

Scatterplot of the diversity (Shannon-Wiener H) and the total species number of the six studied sites by studied years (2013–2015). Squares: wet habitats, circles: transitional habitats, triangles: xeric habitats

Cluster analysis of the butterfly samples taken on the six sites between 2012 and 2014 using Bray-Curtis MISSQ on the relative frequency data of species. Squares: wet habitats, circles: transitional habitats, triangles: xeric habitats

Principal Component (PCoA) based on Bray-Curtis distances of the Lepidoptera samples taken between 2012 and 2014. The symbols are the same as on Fig. 2. Information content: 1st: 42.59%, 2nd: 14.14%, cum: 56.73%

Dominance profiles of studied sampling sites on the mean relative frequencies of species between 2012 and 2014. Squares: wet habitats; circles: transitional habitats, triangles: xeric habitats

Kendall’s concordance profiles of Lepidoptera assemblages of six studied sites based on the data collected between 2012 and 2014. The symbols are the same as in the Fig. 5
Sampling and identification of material
The surveys were carried out by standardized transect walks in 2012, 2013 and 2014 in four, five and eight repetitions, respectively (2012: 16.-18.06; 11.-14.07; 04.-06.08; 06.-08.09; 2013: 05.-06.06; 05.-08.07; 21–23.07; 13–15.08; 30.08.-01.09; 2014: 23.-24.04; 21.-23.05; 09.-10.06; 11.-13.07; 22.-24.07; 14.-16.08; 27.-29.08; 08.-10.09). For quantitative analyses the data of those four-four transects walks were only considered which were phenologically overlapping (i.e. early summer, mid-summer, late summer, early autumn, figures in italics) in each years.
We designated two transects at each of the 6 sites (with lengths of 50 m) and recorded all butterflies in a distance range of 2.5 m to their right, 2.5 m to their left, 5 m ahead of them and 5 m above them. Visits were conducted when the temperature was above 20 °C in sunny weather, without strong wind and rain (see: Van Swaay 2002). Most butterflies were readily identified by observation. Individuals with doubtful species identity were captured by net and they were either immediately released after identification or preserved as voucher specimen for exact determination (e.g. some Lycaenidae or Melitaea species).
For identification, we used the recent taxonomic atlas of Macrolepidoptera of Hungary (Varga 2011). The biogeographical categorisation of species followed the principle elaborated by Varga (1977) and modified by Varga et al. (2004). The classification of faunal components was based on the perception that the bionomy of the lepidopterans is primarily influenced by the larval hostplants, and therefore it can be characterized by a certain type of habitats, like softwood vs. hardwood forests, humid vs. dry swards, humid tall forb formations, etc. (Varga et al. 2004).
Statistics
The composition of butterfly assemblages was compared by Principal Coordinate (PCoA) and cluster analysis in which Bray-Curtis distance was used, in clustering the MISSQ (incremental sum of squares method of Ward and Orlóci) was chosen (Podani 1997a). The assemblages were compared using their yearly data sets containing relative abundance of species (abbreviated: RF%) in the samples. The analyses were carried out with the SynTax 2000 programme package (Podani 1997b).
Studied sites were a priori categorized to xeric, transitional and wet types on the basis of our former results (Szanyi et al. 2015b). The correspondence between the a priori groups and groups formed was evaluated with multivariate analysis. Assemblage types identified using multivariate analysis were characterised and compared on the basis of their species richness, mean number of species and individuals, relative frequencies of biogeographical groups and feeding types of species (Online Resource 1 Table S2).
The quantitative character species (indicator species) of the assemblages were classified by the IndVal method, using the programme package IndVal (Dufrêne and Legendre 1997). We hierarchically classified the species according to their fidelity (constancy within group). Value IV of species is the highest (100) if the given species is present in all samples of the given group and is also exclusive for this group of samples. The program calculates the values IV of each species for each hierarchic level of clustering and the maximum value will be considered as indicator value of the given species. The significance of values IV was calculated by randomisation (1000 iterations).
Conclusions from the composition of assemblages would be consistent if assemblages under consideration are stable over time. Temporal constancy of assemblages was measured using Kendall’s Coefficient of Concordance (W), a non-parametric test based on the abundance ranks of species among years (Legendre 2005). If the structure of assemblages persist, abundance rank of coexisting species should be similar leading to a high and significant W. If the structure is not constant, species ranks greatly vary among years and W values decrease. To get concordance profiles of assemblages, W values were calculated for a sequence of assemblages from the three most abundant species (based on the average of three studied years) until all species were included. In this way we examined the sensitivity of the analysis to inclusion of species in an assemblage (Zar 1984; Joern and Pruess 1986; Podani 1997a).
Results
Species and individual numbers, diversity of assemblages
The sampling sites proved to be moderately rich in species and individuals: 66 species of butterflies in 6556 individuals were registered during the transect walks (Online Resource 1 Table S1.). The highest species numbers were recorded at the transitional site [iii] (“Felső-erdő”), and the wet site [v] “Reserve 2”). Oppositely, the xeric sites [i] and [ii] (“Szapat”, “Körerdő”) were found rather poor, especially the earlier overgrazed and thus, degraded site [i]. These conditions were essentially similar in each year. However, the observed species numbers were somewhat higher in each site in 2014 but the number of individuals were found below the values of the preceding years.
The species composition of the xeric sites was the poorest as nearly only generalist species were recorded. Eight generalist species were observed at all sites: Pyrgus malvae, Thymelicus silvestris*, Pieris rapae, P. napi, Cupido argiades, Polyommatus icarus, Coenonympha pamphilus*, Maniola jurtina*. These species are polyphagous, three of them are connected to grasses* (Poaceae), while the others are feeding on herbaceous plants. Two further species, feeding on Fabaceae (Lotus, Lathyrus, Medicago, etc.) occur everywhere with the exception of the degraded xeric site: L. sinapis/juvernica, Cupido alcetas.
The repartition of individual numbers was rather similar. The highest individual numbers were recorded in the transitional [iii] and the wet site [v] while the xeric sites [i]-[ii] showed the lowest values. Some species reached considerable abundance. The aggregate number of the most common ten species exceeded 60% of the total number of observed butterflies: Coenonympha pamphilus (846), Polyommatus icarus (691), Argynnis paphia (552, only in transitional and wet sites), Maniola jurtina (454), Pieris rapae (344), Leptidea sinapis/ juvernica (286) Minois dryas (273, in a single transitional site [iii] only!), Pieris napi (268), Aphantopus hyperanthus (255), Pyrgus malvae (182) (Table 2). Other species showed also a rather similar distribution: absence or rather low presence in xeric sites and high or moderate number of individuals in transitional and/or wet sites, e.g. the Nymphalid species feeding on common nettle (Urtica dioica), with highest frequency of Araschnia levana. High numbers of some fritillary species were also registered in transitional and wet sites, such as Argynnis paphia, Brenthis daphne, Boloria selene, B. dia, Melitaea phoebe and M. athalia.
Although most sites were surrounded by hardwood forests with nature-like edges, the typical species of this habitat, e.g. the hairstreak (Satyrium) species, were scarce. The large, protected Nymphalidae species connected to woody vegetation (Apatura ilia, Nymphalis antiopa, N. polychloros, N. xanthomelas) were also recorded in low numbers. Two grass-feeding, more or less generalist species (Minois dryas, Melanargia galathea) were observed on a single site [iii] only. The former species was common at one of the wet sites, only during the first two years, however.
The values of Shannon-Wiener diversity showed an increasing gradient from the disturbed (drained and formerly overgrazed) pasture ([i]” Szapat”) towards the mosaic-like habitats with scrubby patches as well as natural-like forest edges ([iv] Reserve 1 and [vi]” Kismakkos”). The species diversities of the sites surrounded by forests and/or natural-like forest edges reached about the same values (Fig. 2, see sites 3–6). This trend was observed during our whole sampling period (2012–2014). Therefore, it seems to be a general tendency that the nature-like forest edges support a higher diversity of species, faunal elements and ecological types.
Biogeographical and ecological composition of butterfly assemblages
For the further quantitative analyses only the results of samplings were included which were made in phenologically identical periods of year. Thus, the following analyses are based on 4846 individuals of 57 species.
The sites of our surveys are mostly surrounded by anthropogenic habitats. Thus, the majority of species belong to widely distributed Euro-Siberian faunal elements with broad ecological tolerance (see: hyper-euryoecious and euryoecious species in Online Resource 1 Table S2.). They generally do not have any food plant specialisation and thus, occur both in disturbed or transformed secondary habitats (see: high frequency of polyphagous species in xeric sites, Online Resource 1 Table S2.). Special biogeographic elements, such as Holo-Mediterranean (incl. -West Asiatic), Ponto-Mediterranean, Southern Continental or Boreo-Continental species are much less represented. The number of „extra-Palaearctic” migrant species is relatively high. However, they are represented only by lower numbers of individuals (see: RF% of migrant species, Online Resource 1 Table S2.). The most widely distributed and common migrant species was Cupido argiades was often observed during the last years, also outside of our sampling areas.
The repartition of faunal components (RF%, Table S1) also proved to be rather similar to the faunal elements since the species with wide tolerance predominated. Highest proportion of polyphagous species was observed at the xeric site [i]. The RF% of the oligophagous species was between 40 and 50% at all sites. The specialists are mostly connected either to humid habitats, e.g. Lycaena dispar rutila, Boloria selene, to light-penetrated forests as Neptis sappho, Lopinga achine or forest fringe formations, as some Satyrium species, Brenthis daphne, etc. In these connections we could not find any differences among the three consecutive years.
Dominance structure and similarity of butterfly assemblages
Steepest and shortest log-series dominance profiles of the less diverse xeric sites, especially the site [i], showed a less balanced structure of these species poor assemblages (Whitakker 1965). The profiles of the other sites were more similar with the exception of the species with lowest frequency forming the “tail” of the curves (see: sites [ii]-[v]). The most even, nearly log-normal course was shown in case of the species with medium frequency (ranks 10–23) at the wet site [vi]. The compositional similarity of the assemblages was compared by Cluster Analysis and Principal Component (PCoA) based on Bray-Curtis distances (programme package SynTax 2000) (Figs. 3 and 4). The analyses were carried out separately for each year.
The position of the eighteen samples clearly reflected the ‘a priori’ categorisation of the habitats. The basic split was shown between the ‘xeric‘ and ‘not xerix’ sites. Additionally, both the xeric and the wet sites were clustered to one group. In all sites the samples of all years clustered together. The differences among the different sites were usually larger than the differences between the samples of different years within each site. The most diverse ‘Felső-erdő’ samples were clearly differentiated both form xeric and transitional und humid sites, while the transitional Reserve 1 was clustered near to the humid sites (Fig. 3).
According to the Principal Component (PCoA) analysis (Fig. 4) the xeric and not xeric sites were clearly differentiated on the Axis 1 explaining 42.59% of the variance. However, the transitional and wet sites were only differentiated according to the second axis (with lower information content of 14.14%) and the less differentiated transitional site (Reserve 1) was nested into the humid sites while the most diverse other transitional site was clearly differentiated.
The ordination of the sites was completed with an Indicator Species Analysis (IndVal) (Table 3). Thirteen generalist species were associated with all sampling sites, while the disturbed xeric sites were characterised by only five species, and also five species seemed to be typical for the wet sites, but only three species showed significant IV values. All ‘non-xeric’ sites were characterised by nine species, including rather frequent ones (e.g. Argynnis paphia, Leptidia sinapis/ juvernica). The distinction of the two transitional groups was supported by seven-seven significantly characteristic species. The relatedness of the transitional [iv] and we t sites was demonstrated by three symmetric character species (IV > 55). Species of conservation importance (see: Discussion) are to be found in the species groups typical for mesic and/or scrubby, semi-dry sites. We could not differentiate further sub-groups since the indicator species of the lowest hierarchic level would be very weakly supported.
According to the dominance rank structure the xeric sites were clearly separated from the othersboth by the highest frequency of the one or two most dominant species but also by the shortest “tail” due to the low species numbers, especially in site I (Fig. 5). The curves of all other sites were less steep but only in one case (VI) approaching the log-normal course.
The concordance profiles showed nearly the same regularities as the dominance profiles (Fig. 6). The course of the concordance profiles of the xeric sites was strikingly different from the other sites (Fig. 6) since the dominant species were nearly identical in all years while it was much more variable in all other sites.
In the xeric sites the most dominant species did not change from year to year and they showed high initial W values (0.8 to 1). These profiles also indicated the shortest “tail” due to the lack of most specialist species (see: Results on ecological composition, Table 3). In other sites, the most dominant species significantly changed from year to year. Therefore, these profiles started with much lower W values (0.1 to 0.4). The most diverse site (III) showed already more balanced W values between the 5–9 species, while in other cases only the ranks 13–35 proved to be more balanced. The [vi] wet site with the least variable frequencies of species (with highest Shannon-Wiener diversity) showed a gradual course of the profile with less “tail” species.
It means that the results of the Principal Coordinate (PCoA) and cluster analysis, the frequency profiles and the results of the concordance analysis strongly support each other despite of the fact that the survey period was rather short (three years only).
Discussion
Several studies have shown that in Europe the diversity of the insect assemblages is highly threatened by fragmentation and degradation of nature-like and semi-natural habitats (Wilcox and Murphy 1985; Steffan-Dewenter and Tscharntke 2002; Tscharntke et al. 2002). Despite of these alarming facts, previous surveys have also shown that the traditionally used cultural landscapes of Eastern Central Europe, incl. The Carpathian basin, are housing a significant level of biodiversity even in recent times. Especially the semi-dry grasslands were shown as hotspots of diversity, floristically and also considering the butterfly assemblages (e.g. Schmitt and Rákosy 2007; Dengler et al. 2012; Roleček et al. 2014).
The agrarian mosaic landscapes are suitable to demonstrate the effects of fragmentation of grasslands on the diversity of insect assemblages (e.g. Bergman et al. 2004; Öckinger and Van Dyck 2012; Filz et al. 2013). The humid, riverine landscapes, as the Bereg lowland with remnants of forests, wetland habitats and traditionally managed grasslands offer optimal conditions for such studies (Magura et al. 1997) despite the fact that due to drainage and abandonment of the land use habitat degradation and fragmentation have started (Ködöböcz and Magura 1999; Kormány 2006; Szanyi et al. 2015b). These changes make possible to test our working hypotheses but also some general hypotheses of the community ecology.
According to the generally accepted hypothesis, strong disturbance can result in the increase of frequency of some generalist species connected with the decrease and/or extinction of specialists, and in homogenisation in the composition of assemblages (Devictor et al. 2008; Brückmann et al. 2010; Ekroos et al. 2010). Our results show, however, that the two processes should be disentangled. Although we confirmed the high frequency of widely distributed generalist species, the number of species and the diversity values did not decrease compared with other regions within the Carpathian basin (66 vs. 68 species, see: Baur et al. 2006), and was even slightly higher than in some cultural landscapes of Western Europe (Öckinger and Smith 2006), of southern Poland (Skórka et al. 2007; Rosin et al. 2012), or of Toscana (Maccherini et al. 2009) but the species numbers were somewhat lower than in the intensively surveyed Central and Southern European calcareous grasslands (Marini et al. 2009; Kramer et al. 2012; Filz et al. 2013). Thus, the predicted homogenisation of assemblages was also not observed. However, our working hypotheses on the frequency and concordance of polycyclic generalist species were much more supported.
Furthermore, we could also confirm our hypothesis on restricted localisation and fluctuating numbers of some specialists, as species connected with natur-like forest fringes, e.g. some Theclini (Satyrium pruni, S. ilicis (Esper, 1779)), with mesic habitats (Cupido alcetas, Polyommatus semiargus (Rottemburg, 1775)) or to light-penetrated forests (Lopinga achine, Euphydryas maturna (Linnaeus, 1758), latter only outside of sampling sites!). These latter species belong to the mostly decreasing and threatened ones at European level due to the abandoning of some traditional methods of forest management as coppicing, threadwise cutting, etc. (Freese et al. 2006; Van Swaay et al. 2006).
It was also hypothesised that size and quality of habitat patches play a decisive role in the sustaining of a high species diversity (Báldi 2007; Kramer et al. 2012). During our surveys we separated (see: ‘a priori’ categorisation, Szanyi et al. 2015b) three different types of habitats, also with different extension. Our results confirm that the extended but drained and heavily disturbed habitats (I-II) could not support any high species diversity. Oppositely, the restricted site V, surrounded by nature-like forest skirt, has nearly the same species number as the more extended transitional site IV.
These data cannot be evaluated without the knowledge of the former land use of the sites, however. Considering the military maps from the end of the 18th century, all these sites appear to be forested with the exception of the site III which was used as hayfield during the whole 19th and 20th centuries. Furthermore, the sites I-II. were clearcut after the World War I., probably during the economical crisis (the same proceeded also in the Hungarian side of the Bereg plain) and later drained an used as pasture. This scenario is strongly supported by the data of the proportion species number vs. Shannon-Wiener diversity which shows in sites I-II a nearly linear connection while this connection is more diffuse in other cases where the highest figures were shown at the transitional site III.
In this connection experience of some Authors (e.g. Weidemann 1988; Tews et al. 2004; Halder et al. 2015) was also confirmed that the nature-like forest fringes can support a higher species diversity due to the presence of specialists. We have observed in such sites some species which are connected by the larval food plants to edge structures, as Satyrium spp. (Prunus, Rhamnus spp.), Brenthis daphne (Rubus spp.), Melitaea athalia (Veronica, Melampyrum spp.) or to open forests as Lopinga achine (Brachypodium sylvaticum, Carex brizoides; Bergman 1999, 2001). One could accapt that the presence of other, e.g. mesophilous species (Boloria selene, B. dia) may be favoured by more balanced micro-climatic conditions but we could not confirm these statements since practically all climatic niche models are based on large-scale macro-climatic data (e.g. Parmesan 2006; Phillips et al. 2006; Habel et al. 2011).
Large-scale ‘a priori’ subdivisions (e.g. on physiognomy or composition of the vegetation) are often used as first step in community ecological surveys. If so, it must be tested whether these subdivisions could be confirmed by ‘a posteriori’ analyses. In our case, we experienced a good agreement both in species composition and diversity. In the next step we considered the results of the ordinations (cluster analysis and Principal Component Analysis) and of IndVal analysis. The step-by-step subdivision of the indicator species (Table 3) can be completely parallelized with the results of the PCoA and cluster analysis (Figs. 2 and 3) where the basal split was found between the xeric and all other types of sites. In this context we should compare which species have shown the highest IV-values (IV ≥ 95) in transitional and wet habitats combined with high number of individuals.
These species are: Aphantopus hyperanthus (255), Argynnis paphia (552), Melitaea athalia (113), Minois dryas (273). However, A. paphia and M. dryas suffered a strong decline in the third year, probably due to the extreme drought of the spring and early summer periods. As a consequence of this dry period the number of nectar sources was strongly reduced, although it was shown (Kalarus and Nowicki 2015) that the number of nectar sources may be critical for the M. dryas poulation. The same negative tendency was also observed in two nymphalid species (Araschnia levana, Inachis io) feeding on nettle (Urtica dioica) which also need diverse nectar sources. These species may compete according to a recent survey (Audusseau et al. 2016) in which was shown that the population growth of the northwards expanding A. levana was connected with the decrease of the I. io population. More even numbers were registered in some fritillary species in transitional and wet sites, such as Brenthis daphne, Boloria selene, B. dia, Melitaea phoebe and M. athalia, although the latter two species belong to the r-strategists according to the oviposition behaviour and social early larval period (Warren 1987a, b; Weidemann 1995; Wahlberg 2000; Tóth et al. 2015).
The basic split between xeric and “non-xeric” sites became mostly evident, however, from the dominance and concordance profiles. These profiles showed both sides of the same regularity: (i) few but highly dominant species in xeric sites with hardly any chance of change during the consecutive years, and (ii) fluctuating change of the dominant/subdominant species followed by a long sequence of medium frequent or scarce, partly characteristic, i.e. indicator species (see: Table 3). The presence of a relatively high number of such species had the consequence that we have chosen the IV ≥ 40 as criteria for quantitative character species in consense with the references (Legendre 2005). Oppositely, the constant rank positions of the dominant species in xeric habitats was interpreted as a consequence of the extreme conditions as drainage and partial overgrazing and trampling. This degradation effect is clearly reflected by the rather steep course of the dominance profile of these sites. The non-xeric sites were forming much more a “continuum” according to the earlier hypothesis of Whitakker (1965) in which, although the bulk of species fluctuate from year to year, but the assemblage displays a more or less even dominance hierarchy as e.g. shown by Root and Cappuccino (1992). This evenness is mostly expressed at the site [vi] with smaller number of “tail” species while all other transitional and wet sites have shown a higher number of less frequent species which obviously do not compete with each other.
Based on our analyses, we concluded that the drainage of habitats represents the most important threat for the studied butterfly assemblages. It was demonstrated not only by the strong separation of the xeric habitats in PCoA and cluster analyses, but also the reduced species number and the steep dominance structure show the marks of degradation. However, despite of the change of land use, fragmentation and partial drainage of habitats, the other sites support a diverse assemblage of butterflies characterised by a log-normal dominance structure and the presence of numerous species with intermediate or fluctuating frequency. These habitats could preserve also some protected species (e.g. the large copper – Lycaena dispar, protected by the Habitats Directive, and several large Nymphalids). Compared with the composition and modest diversity of butterfly assemblages in other regions (e. g. Schneider and Fry 2001; Skórka et al. 2007; Rosin et al. 2012) we expect that the conservation status of these sites could be improved by some soft management. If so, the conservation efforts should be mostly focused to sustain the generally high level of biodiversity by the conservation of the nature-like forest edge structures but also by the possible re-establishment of some kinds of traditional use such as mowing or extensive grazing by cattle.
References
Andelman SJ, Fagan WF (2000) Umbrellas and flagships: efficient conservation surrogates or expensive mistakes? Proc Natl Acad Sci U S A 9797:5954–5959. https://doi.org/10.1073/pnas.100126797
Audusseau H, Vaillant ML, Janz N, Nylin S, Karlsson B, Schmucki R (2016) Species range expansion constrains the ecological niches of resident butterflies. J Biogeogr 44(1):28–38. https://doi.org/10.1111/jbi.12787
Báldi A (2007) Habitat heterogeneity overrides the species–area relationship. J Biogeogr 35(4):675–681. https://doi.org/10.1111/j.1365-2699.2007.01825.x
Baur B, Cremene C, Groza G, Rákosy L, Schileyko AA, Baur A, Stoll P, Erhardt A (2006) Effects of abandonment of subalpine hay meadows on plant and invertebrate diversity in Transylvania, Romania. Biol Conserv 132:261–273. https://doi.org/10.1016/j.biocon.2006.04.018
Bergman K (1999) Habitat utilization by Lopinga achine (Nymphalidae: Satyrinae) larvae and ovipositing females: implications for conservation. Biol Conserv 88:69–74. https://doi.org/10.1016/S0006-3207(98)00088-3
Bergman KO (2001) Population dynamics and the importance of habitat management for the conservation of the butterfly Lopinga achine. J Appl Ecol 38:1303–1313. https://doi.org/10.1046/j.0021-8901.2001.00672.x
Bergman KO, Askling J, Ekberg O, Ignell H, Wahlman H, Milberg P (2004) Landscape effects on butterfly assemblages in an agricultural region. Ecography 27:619–628. https://doi.org/10.1111/j.0906-7590.2004.03906.x
Boros Á (1964) A tőzegmoha és a tőzegmohás lápok Magyarországon. [Sphagnum and peat bogs in Hungary.]. Vasi Szemle 18:53–68 (In Hungarian)
Brückmann SV, Krauss J, Steffan-Dewenter I (2010) Butterfly and plant specialists suffer from reduced connectivity in fragmented landscapes. J Appl Ecol 47(4):799–809. https://doi.org/10.1111/j.1365-2664.2010.01828.x
Bubová T, Vrabec V, Kulma M, Nowicki P (2015) Land management impacts on Europeanbutterflies of conservation concern: a review. J Insect Conserv 19:805–821. https://doi.org/10.1007/s10841-015-9819-9
Caro TM, O’Doherty G (1999) On the use of surrogate species in conservation biology. Conserv Biol 13:805–814. https://doi.org/10.1046/j.1523-1739.1999.98338.x
Cottingham KL, Brown BL, Lennon JT (2001) Biodiversity may regulate the temporal variability of ecological systems. Ecol Lett 4:72–85. https://doi.org/10.1046/j.1461-0248.2001.00189.x
Cremene C, Groza G, Rakosy L, Schileyko AA, Baur A et al (2005) Alterations of steppe-like grasslands in Eastern Europe: a threat to regional biodiversity hotspots. Conserv Biol 19:1606–1618. https://doi.org/10.1111/j.1523-1739.2005.00084.x
Csergő AM, Demeter L, Turkington R (2013) Declining diversity in abandoned grasslands of the Carpathian mountains: do dominant species matter? PLoS One 8:e73533. https://doi.org/10.1371/journal.pone.0073533
Deli T, Sümegi P, Kiss J (1997) Biogeographical characterisation of the mollusc fauna on Szatmár-Bereg plain. In: Tóth E, Horváth R (eds) ANP Füzetek Aggtelek Vol I. Aggtelek, Hungary, pp 123–129
Dengler J, Becker T, Ruprecht E, Szabó A, Becker U, Beldean M, Bita-Nicolae C, Dolnik C, Goia I, Peyrat J, Sutcliffe LME, Turtureanu PD, Uğurlu E (2012) Festuco-Brometea communities of the Transylvanian plateau (Romania) – a preliminary overview on syntaxonomy, ecology, and biodiversity. Tuexenia 32:319–359
Devictor V, Julliard R, Jiguet F (2008) Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117:507–514. https://doi.org/10.1111/j.2008.0030-1299.16215.x
Downing AL, Brown BL, Perrin EM, Keitt TH, Leibold M (2008) Environmental fluctuations induce scale-dependent compensation and increase stability in plankton ecosystems. Ecology 89:3204–3214. https://doi.org/10.1890/07-1652.1
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for flexible asymmetrical approach. Ecol Monogr 67:345–366. https://doi.org/10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Ekroos J, Heliöla J, Kuussaari M (2010) Homogenization of lepidopteran communities in intensively cultivated agricultural landscapes. J Appl Ecol 47:459–467. https://doi.org/10.1111/j.1365-2664.2009.01767.x
Erhardt A, Thomas JA (1991) Lepidoptera as indicators of change in the semi-natural grasslands of lowland and upland Europe. In: Collins NM, Thomas JA (eds) The conservation of insects and their habitats. Academic Press, London, pp 213–236
Filz KJ, Engler JO, Stoffels J, Weitzel M, Schmitt T (2013) Missing the target? A critical view on butterfly conservation efforts on calcareous grasslands in South-Western Germany. Biodivers Conserv 22(10):2223–2241. https://doi.org/10.1007/s10531-012-0413-0
Freese A, Benes J, Bolz R et al (2006) Habitat use of the endangered butterfly Euphydryas maturna and forestry in Central Europe. Anim Conserv 9:388–397. https://doi.org/10.1111/j.1469-1795.2006.00045.x
Gálik K, Deli T, Sólymos P (2001) Comparative malacological investigations on the Kaszonyi Hill (NE Hungary). Malac News 19:81–88
Habel JC, Rödder D, Schmitt T, Nèves G (2011) Global warming will affect the genetic diversity and uniqueness of Lycaena helle populations. Glob Chang Biol 17:194–205. https://doi.org/10.1111/j.1365-2486.2010.02233.x
Halder I, Barnagaud JY, Jactel H, Barbaro L (2015) Woodland habitat quality prevails over fragmentation for shaping butterfly diversity in deciduous forest remnants. For Ecol Manag 357:171–180. https://doi.org/10.1016/j.foreco.2015.08.025
Hambler C, Speight MR (2004) Extinction rates and butterflies. Science 305:1563. https://doi.org/10.1126/science.305.5690.1563b
Harrison S, Bruna E (1999) Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography 22:225–232. https://doi.org/10.1111/j.1600-0587.1999.tb00496.x
Houlahan JE, Currie DJ, Cottenie K, Cumming GS, Ernest SKM, Findlay CS, Fuhlendorf SD (2007) Compensatory dynamics are rare in natural ecological communities. Proc Natl Acad Sci U S A 104:3273–3277. https://doi.org/10.1073/pnas.0603798104
Huemer P, Tarmann G (2001) Artenvielfalt und Bewirtschaftungsintensität: Problemanalyse am Beispiel der Schmetterlinge auf Wiesen und Weiden Südtirols. Gredleriana 1:331–418
Joern A, Pruess KT (1986) Temporal constancy in grasshopper assemblies (Orthoptera: Acrididae). Ecol Entom 11:379–385. https://doi.org/10.1111/j.1365-2311.1986.tb00316.x
Kalarus K, Nowicki P (2015) How do landscape structure, management and habitat quality drive the colonization of habitat patches by the dryad butterfly (Lepidoptera: Satyrinae) in fragmented grassland? PLoS One 10:e0138557. https://doi.org/10.1371/journal.pone.0138557
Ködöböcz V, Magura T (1999) Biogeographical connections of the carabid fauna (Coleoptera) of the Beregi-síkság to the Carpathians. Folia Ent Hung 60:195–203
Kormány G (1976) Szabolcs-Szatmár megye éghajlata. [The climate of the County Szabolcs-Szatmár.]. Szab-Szat Szem 1:32–40 (In Hungarian)
Kormány, Gy (2006) A Szatmár-Beregi síkság klimatikus és domborzati viszonyai. [The climate and relief of the Szatmár-Beregi plain.] Research report of OTKA T/F 046821. (In Hungarian)
Kramer B, Poniatowski D, Fartmann T (2012) Effects of landscape and habitat quality on butterfly communities in pre-alpine calcareous grasslands. Biol Conserv 152:253–261. https://doi.org/10.1016/j.biocon.2012.03.038
Legendre P (2005) Species associations: the Kendall coefficient of concordance revisited. J Agric Biol Environ Stat 10:226–245
Lethmate J (2005) Stickstoff-Regen: Ein globales Eutrophierungsexperiment. Biologie in unserer Zeit 35:118–135
Maccherini S, Bacaro G, Favilli L, Piazzinia S, Santi E, Marignani M (2009) Congruence among vascular plants and butterflies in the evaluation of grassland restoration success. Acta Oecol 35:11–17. https://doi.org/10.1016/j.actao.2008.12.002
MacNally R, Fleishman E (2004) A successful predictive model of species richness based on indicator species. Conserv Biol 18:646–654. https://doi.org/10.1111/j.1523-1739.2004.00328_18_3.x
Magura T, Ködöböcz V, Tóthmérész B, Molnár T, Elek Z, Szilágyi G, Hegyessy G (1997) Carabid fauna of the Beregi-síkság and its biogeographical relations (Coleoptera Carabidae). Folia Ent Hung 58:73–82
Magurran AE (2005) Species abundance distributions: pattern or process? Funct Ecol 19:177–181. https://doi.org/10.1111/j.0269-8463.2005.00930.x
Marini L, Fontana P, Klimek S, Battisti A, Gaston KJ (2009) Impact of farm size and topography on plant and insect diversity of managed grasslands in the alps. Biol Conserv 142:394–403. https://doi.org/10.1016/j.biocon.2008.10.034
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: biodiversity synthesis. World Resources Institute, Washington
New TR, Pyle RM, Thomas JA, Thomas CD, Hammond PC (1995) Butterfly conservation management. Annu Rev Entomol 40:57–83. https://doi.org/10.1146/annurev.en.40.010195.000421
Öckinger E, Smith HG (2006) Semi-natural grasslands as population sources for pollinating insects in agricultural landscapes. J Appl Ecol 44(1):50–59. https://doi.org/10.1111/j.1365-2664.2006.01250.x
Öckinger E, Van Dyck H (2012) Landscape structure shapes habitat finding ability in a butterfly. PLoS One 7(8):e41517. https://doi.org/10.1371/journal.pone.0041517
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37:637–669. https://doi.org/10.1146/annurev.ecolsys.37.091305.110100
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Podani J (1997a) Bevezetés a többváltozós biológiai adatfeltárás rejtelmeibe. [Introduction to the multivariate statistics of biological data] Scientia, Budapest 412 p. (In Hungarian)
Podani J (1997b) SYNTAX 5.1.: a new version of PC and Macinthosh computers. Coenoses 12:149–152
Rákosy L, Schmitt T (2011) Are butterflies and moths suitable ecological indicator systems for restoration measures of semi-natural calcareous grassland habitats? Ecol Indic 11:1040–1045. https://doi.org/10.1016/j.ecolind.2010.10.010
Roleček J, Čornej II, Tokarjuk AI (2014) Understanding the extreme species richness of semi-dry grasslands in east-Central Europe: a comparative approach. Preslia 86:13–34
Root R, Cappuccino N (1992) Patterns in population change and the organization of the insect community associated with goldenrod. Ecol Monogr 62:393–420. https://doi.org/10.2307/2937117
Rosin ZM, Myczko L, Skórka P, Lenda M, Moroń D, Sparks TH, Tryjanowski P (2012) Butterfly responses to environmental factors in fragmented calcareous grasslands. J Insect Conserv 16(3):321–329. https://doi.org/10.1007/s10841-011-9416-5
Schmitt T, Rákosy L (2007) Changes of traditional agrarian landscapes and their conservation implications: a case study of butterflies in Romania. Divers Distrib 13:855–862. https://doi.org/10.1111/j.1472-4642.2007.00347.x
Schneider C, Fry GLA (2001) The influence of landscape grain size on butterfly diversity in grasslands. J Insect Conserv 5(3):163–171. https://doi.org/10.1023/A:1017932931192
Scoble MJ (1992) The Lepidoptera: form, function and diversity. Oxford University Press, New York
Settele J, Kühn E, Thomas JA (eds) (2005) Studies on the ecology and conservation of butterflies in Europe. Vol. 2. Species ecology along a European Gradient: Maculinea butterflies as a model. Pensoft Publishers, Sofia-Moscow
Simon T (1952) Montán elemek az Észak-Alföld flórájában és növénytakarójában. [Montane elements in the flora of the Northern Plain.]. Annual Bio Univ Debr 1:146–174 (In Hungarian)
Skórka P, Settele J, Woyciechowski M (2007) Effects of management cessation on grassland butterflies in southern Poland. Agric Ecosyst Environ 121:319–324. https://doi.org/10.1016/j.agee.2006.11.001
Steffan-Dewenter I, Tscharntke T (2002) Insect communities and biotic interactions on fragmented calcareous grasslands – a mini review. Biol Conserv 104:275–284. https://doi.org/10.1016/S0006-3207(01)00192-6
Strange N, Thorsen BJ, Bladt JS, Wilson KA, Rahbek C (2011) Conservation policies and planning under climate change. Biol Conserv 144:2968–2977. https://doi.org/10.1016/j.biocon.2011.08.022
Szanyi S, Katona K, Bernát N, Tamási K, Molnár A (2015a) The flora of the grasslands of the Velyka Dobron’ wildlife reserve (Transcarpathia, West Ukraine). Hung J Land Ecol 13:1–8 (In Hungarian)
Szanyi S, Katona K, Rácz I, Varga Z, Nagy A (2015b) Orthoptera fauna of the Ukrainian part of the Bereg plain (Transcarpathia, western Ukraine). Articulata 30(1):91–104
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92. https://doi.org/10.1046/j.0305-0270.2003.00994.x
Thomas JA (2005) Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philos Trans R Soc Lond Ser B Biol Sci 360:339–357. https://doi.org/10.1098/rstb.2004.1585
Thomas JA, Settele J (2004) Butterfly mimics of ants. Nature 432:283–284. https://doi.org/10.1038/432283a
Thomas JA, Telfer MG, Roy DB, Preston CD, Greenwood JJD, Asher J, Fox R, Clarke RT, Lawton JH (2004) Comparative losses of British butterflies, birds and plants and the global extinction crisis. Science 303:1879–1881. https://doi.org/10.1126/science.1095046
Tokeshi M (1993) Species abundance patterns and community structure. Adv Ecol Res 24:111–186. https://doi.org/10.1016/S0065-2504(08)60042-2
Tóth JP, Bereczki J, Végvári Z, Juhász E, Varga Z (2015) Different host plant utilization ability of two closely related Melitaea species (Lepidoptera: Nymphalidae). Eur J Entomol 112:120–125. https://doi.org/10.14411/eje.2015.001
Tscharntke T, Steffan-Dewenter I, Kruess A, Thies C (2002) Characteristics of insect populations on habitat fragments: a mini review. Ecol Res 7:229–239. https://doi.org/10.1046/j.1440-1703.2002.00482.x
Valone TJ, Barber NA (2008) An empirical evaluation of the insurance hypothesis in diversity-stability models. Ecology 89:522–531. https://doi.org/10.1890/07-0153.1
van Swaay CAM (2002) The importance of calcareous grasslands for butterflies in Europe. Biol Conserv 104:315–318. https://doi.org/10.1016/S0006-3207(01)00196-3
van Swaay CM, Warren S, Löis G (2006) Biotope use and trends of European butterflies. J Insect Conserv 10:189–209. https://doi.org/10.1007/s10841-006-6293-4
Varga Z (1977) Das Prinzip der areal-analytischen Methode in der Zoogeographie und die Faunenelemente-Einteilung der europäischen Tagschmetterlinge (Lep.: Diurna). Acta Biol Debr 14:223–285
Varga Z (ed) (2011) Macrolepidoptera of Hungary. Heterocera Press, Budapest
Varga Z, Ronkay L, Bálint Zs, Gyula LM, Peregovits L (2004) Checklist of the fauna of Hungary. Volume 3. Macrolepidoptera. Hungarian Natural History Museum, Budapest
Wahlberg N (2000) Comparative descriptions of the immature stages and ecology of five Finnish Melitaeine butterfly species (Lepidoptera: Nymphalidae). Ento Fenn 11:167–174
Warren MS (1987a) The ecology and conservation of the heath fritillary butterfly, Mellicta athalia. I. Host selection and phenology. J Appl Ecol 24:467–482. https://doi.org/10.2307/2403887
Warren MS (1987b) The ecology and conservation of the heath fritillary butterfly, Mellicta athalia. II. Adult population structure and mobility. J Appl Ecol 24:483–498. https://doi.org/10.2307/2403888
Weidemann HJ (1988) Tagfalter. Bd. 2. Biologie, Ökologie, Biotopschutz. Verlag J. Neumann-Neudamm, Melsungen, Germany
Weidemann HJ (1995) Tagfalter – beobachten, bestimmen. Naturbuch-Verlag, Augsburg
Wenzel M, Schmitt T, Weitzel M, Seitz A (2006) The severe decline of butterflies on western calcareous grasslands during the last 30 years: a conservation problem. Biol Conserv 28:542–552. https://doi.org/10.1016/j.biocon.2005.10.022
Whitakker RH (1965) Dominance and diversity in land plant communities. Science 147:250–260. https://doi.org/10.1126/science.147.3655.250
Wilcox BA, Murphy DD (1985) Conservation strategy: the effects of fragmentation on extinction. Am Nat 125:879–887
Zar JH (1984) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgments
The authors are highly grateful to Dr. Zs. Végvári for critical comments on the earlier version of the manuscript. Szabolcs Szanyi was supported by the European Union and the State of Hungary, co-financed by the European Social Fund in the framework of TÁMOP-4.2.4.A/ 2-11/1-2012-0001 ‘National Excellence Program’ (2013). The study was supported by the Collegium Talentum Program (2014-2015).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Electronic supplementary material
ESM 1
(DOC 135 kb)
Rights and permissions
About this article

Cite this article
Szanyi, S., Nagy, A. & Varga, Z. Diversity and concordance in the composition of butterfly assemblages of the Transcarpathian (Bereg) plain (SW Ukraine). Biologia 73, 951–964 (2018). https://doi.org/10.2478/s11756-018-0102-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2478/s11756-018-0102-x