
Abstract
Two species of oribatid mites of the genus Nothrus (Oribatida, Nothridae) were collected during an expedition to Taiwan in 2017. The species Nothrus anauniensis Canestrini and Fanzago, 1876 is redescribed and illustrated in detail; the main morphological traits for this species are summarized. A new species Nothrus xuejianensis Ermilov sp. n. is described; it differs from the closest species, Nothrus meakanensis Fujikawa, 1999 by the morphology and length of notogastral setae h2 and number of subcapitular setae m.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The oribatid mite genus Nothrus (Acari, Oribatida, Nothridae) was proposed by Koch (1835) with Nothrus palustris Koch, 1839 as type species. Currently, it comprises 87 species, which collectively have a cosmopolitan distribution (Subías 2004, updated 2018). The selective generic characters are summarized by Sellnick and Forsslund (1955), Sitnikova (1975), Olszanowski (1996) and Fujikawa (1999). Identification keys to some Nothrus species groups have been presented in several papers: e.g., Sellnick and Forsslund (1955), Beck (1962), Sitnikova (1975), Mahunka (1978), Olszanowski (1996), Fujikawa (1999), Balogh and Balogh (2002), Weigmann (2006), Lochynska (2008), Bayartogtokh (2010), Ermilov and Hugo-Coetzee (2012).
During taxonomic identification of oribatid mites from Taiwan, we found two species of Nothrus, one is known Nothrus anauniensis Canestrini and Fanzago, 1876 (Canestrini and Fazango 1876), and the other species is new to science. The main goal of the paper is to present a supplementary description of N. anauniensis, on the basis of specimens from Taiwan, and to describe the new species under the name Nothrus xuejianensis Ermilov sp. n.
At present, the oribatid mite fauna of Taiwan is poorly investigated (e.g., Tseng 1982, 1984; Aoki 1991, 1995; Ohkubo 1995; Chu and Aoki 1997; Bayartogtokh et al. 2009; Ermilov and Liao 2017a, b, c), and only two records of Nothrus are known: N. anauniensis (see Chu and Aoki 1997); N. praeoccupatus Subías, 2004 (see Tseng 1982; as Vigilomicrozetes pulchellus Tseng, 1982).
Material and methods
Samples were collected by hand method and extracted into 96% ethanol using Berlese’s funnels without use of an electric lamp during 7 days in the laboratory. Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the ventral plate. Notogastral width refers to the maximum width of the notogaster in dorsal view. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence trochanter–femur–genu–tibia–tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence genu–tibia–tarsus.
Drawings were made with a camera lucida using a Leica transmission light microscope “Leica DM 2500”.
Morphological terminology used in this paper follows that of F. Grandjean: see Travé and Vachon (1975) for references, Norton (1977) for leg setal nomenclature, and Norton and Behan-Pelletier (2009) for overview.
The following abbreviations are used: dep – depression; br – bothridial ridge; ro, le, in, bs, ex – rostral, lamellar, interlamellar, bothridial and exobothridial setae, respectively; cf. –circummarginal furrows; cr – caudal ridge; c, d, e, f, h, p – notogastral setae; ia, im, ip, ih, ips – notogastral lyrifissures; gla – opisthonotal gland opening; h, m, a – subcapitular setae; or – adoral seta; l, d, cm, acm, ul, sul, vt, lt – palp setae; ω – palp and leg solenidion; ep – postpalpal seta; cha, chb – cheliceral setae; Tg – Trägårdh’s organ; g, an, ad – genital, anal and adanal setae, respectively; ian – anal lyrifissure; iad – adanal lyrifissure; σ, φ – solenidia; ɛ – leg famulus; v, ev, bv, l, d, ft., tc, p, u, a, s, pv, pl – leg setae.
The following abbreviations of collections are used: NTU – National Taiwan University, Taipei, Taiwan; TSUMZ – Tyumen State University Museum of Zoology, Tyumen, Russia.
Systematics
Nothrus anauniensis Canestrini and Fanzago, 1876 (Figs 1–20)
Supplementary description
Measurements. Body length: 697–747 (five specimens, all females); notogaster width: 348–381 (five specimens).
Integument (Figs 1, 12–14). Body color yellow-brownish, covered with thin layer of gel-like cerotegument. Surface densely microfoveolate (visible under high magnification, ×1000). Prodorsum, notogaster and genital plates with rounded or elongate oval alveoles (their diameter or length up to 14).
Prodorsum (Figs 1–4). One unpaired mediobasal (in interbothridial region) and one pair of laterobasal (anteriorly to bothridia) depressions clearly visible. Rostrum broadly rounded, with distinct medial indentation. Rostral (20–24), lamellar (36–41) and interlamellar (45–49) setae covered with broad, phylliform cerotegument, heavily barbed, inserted on tubercles, ro–ro and le–le connected by thin ridges. Bothridial setae long (213–217), rod-like, slightly dilated distally, sparsely barbed. Bothridial ridges distinct. Exobothridial setae (8–10) covered with narrow, phylliform cerotegument, smooth.
Notogaster (Figs 1, 5–8, 12, 13). Anterior notogastral margin straight. Posterior part evenly rounded (not dilated). Weakly convex in dorsocentral and lateral parts and with circummarginal furrows between them. Caudal ridges strong. Sixteen pairs of notogastral setae covered with broad, phylliform cerotegument, inserted on small tubercles, c1–c3, d1, d2, cp, e1, e2 heavily barbed, h2 smooth, other setae sparsely barbed; h2 (90–98) longer than c1, d1, d2, e1 (49–57), c3, cp, e2, f1, f2, h1, h3, p1–p3 (41–49) and c2 (28–32). Setae c1 d1, d2 almost reaching the insertions of d1, d2, e1, respectively. Distance between setae c1–c2 shorter than between c2–c3. Lyrifissures ia, im, ip, ih, ips and opisthonotal gland openings well visible, located in typical positions.
Gnathosoma (Figs 14–16). Subcapitulum longer than wide (151–157 × 131–135). Subcapitular setae h (20–22) and m2 (8) covered with narrow phylliform cerotegument, smooth, m1 (20–22) and a (26–28) setiform, barbed. Two pairs of adoral setae (24–28) present; or1 setifrom, smooth, or2 dilated distally, truncate, smooth. Palps (82–86) with setation 0–1–1–3–9(+1ω). Postpalpal setae (12–14) simple, erect. Chelicerae (151–157) with two setiform, barbed setae; cha (49–53) longer than chb (32–36). Trägårdh’s organ elongate triangular.
Epimeral and lateral podosomal regions (Fig. 12). Anterior tectum of epimere I distinctly incised medially. Epimeral setal formula 7–4–6–8. Setae short (12–16), covered with narrow phylliform cerotegument, smooth.
Anogenital region (Figs 9–13). Nine pairs of genital (16–20), two pairs of anal (16–20) and 3 pairs of adanal (ad1, 30–32; ad2, ad3, 16–20) setae covered with narrow phylliform cerotegument, smooth. Lyrifissures ian and iad clearly visible.
Legs (Figs 17–20). Tarsi with 3 smooth claws, median claw distinctly thicker than laterals. Formulas of leg setation and solenidia: I (1–9–5–6–27) [1–2–3], II (1–8–5–5–25) [1–1–1], III (3[or 4]–5–5–5–22) [1–1–0], IV (2–6–5–5–22) [1–1–0]; homologies of setae and solenidia indicated in Table 1.

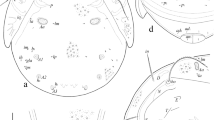
Nothrus anauniensis Canestrini and Fanzago, 1876, adult. 1 Dorsal view 2 Rostral seta 3 Lamellar seta 4 Bothridial seta 5 Notogastral seta c1 6 Notogastral seta c2 7 Notogastral seta h2 8 Notogastral seta p1 9 Anal seta an1 10 Adanal seta ad1 11 Adanal seta ad2
Material examined
Five specimens (all females): Taiwan, Miaoli County, Tai’an Township, SheiPa National Park, Xuejian Area, 24°25.464’N, 121°00.796 ‘E, 1855 m a.s.l., sample #32, soil and lichen, 13.XI.2017 (collected by J.-R. Liao and H.C. Lee).
Remarks
1. Nothrus anauniensis has a cosmopolitan distribution except the Antarctic region (Subías 2004, updated 2018). Adult of this species was redescribed and illustrated by several authors: e.g., Hammen (1950; as N. biciliatus), Sellnick and Forsslund (1955; as N. biciliatus), Hammer (1966; as N. biciliatus), Mahunka (1978; as N. pseudoborussicus), Seniczak and Norton (1993), Olszanowski (1996), Bayartogtokh (2010) and Ermilov and Hugo-Coetzee (2012). The specimens from Taiwan (data in this paper) are similar in general appearance to those from the redescriptions, however, partial morphological were observed:
-
(1)
Bothridial setae rod-like, dilated distally in specimens from Taiwan (it correspond to data from Hammen 1950; Hammer 1966; Olszanowski 1996) versus rod-like, not dilated distally in data from Sellnick and Forsslund (1955), Mahunka (1978), Seniczak and Norton (1993), Bayartogtokh (2010), Ermilov and Hugo-Coetzee (2012).
-
(2)
Notogastral setae h2 considerably longer than p1 in specimens from Taiwan (it correspond to data from Sellnick and Forsslund 1955; Seniczak and Norton 1993) versus h2 and p1 only slightly different in length in data from Hammen (1950), Hammer (1966), Mahunka (1978), Olszanowski (1996), Bayartogtokh (2010), Ermilov and Hugo-Coetzee (2012).
-
(3)
Some notogastral setae heavily barbed, other setae smooth or slightly barbed in specimens from Taiwan (it correspond to data from Mahunka 1978) versus smooth or slightly barbed, not heavily barbed in data from Hammen (1950), Sellnick and Forsslund (1955), Hammer (1966), Seniczak and Norton (1993), Olszanowski (1996), Bayartogtokh (2010), Ermilov and Hugo-Coetzee (2012).
-
(4)
Dorsal notogastral setae comparatively long (c1 d1, d2 almost reaching the insertions of d1, d2, e1) in specimens from Taiwan (it correspond to data from Sellnick and Forsslund 1955) versus comparatively short (c1 d1, d2 clearly not reaching the insertions of d1, d2, e1) in data from Hammen (1950), Hammer (1966), Mahunka (1978), Seniczak and Norton (1993), Olszanowski (1996), Bayartogtokh (2010), Ermilov and Hugo-Coetzee (2012).
-
(5)
Caudal notogastral ridges well-developed in specimens from Taiwan (it correspond to data from Sellnick and Forsslund 1955; Hammer 1966; Seniczak and Norton 1993; Ermilov and Hugo-Coetzee 2012) versus not developed or indistinctly developed in data from Hammen (1950), Mahunka (1978), Olszanowski (1996), Bayartogtokh (2010).
2. Based on the supplementary description of N. anauniensis from Taiwan and other redescriptions (e.g., Hammen 1950; Sellnick and Forsslund 1955; Hammer 1966; Mahunka 1978; Seniczak and Norton 1993; Olszanowski 1996; Bayartogtokh 2010; Ermilov and Hugo-Coetzee 2012), we propose the following diagnostic morphological traits for this species:
Large species, size 697–1050 × 348–500. Surface of prodorsum and notogaster alveolate. Rostral, lamellar and interlamellar setae covered with broad phylliform cerotegument, rarely ro simple. Bothridial setae long, rod-like, sometimes slightly dilated distally, sparsely barbed. Notogastral setae of medium size, covered with broad, phylliform cerotegument, h2 smooth, other setae barbed; h2 longer than p1. Distance between c1–c2 shorter than between c2–c3. Epimeral and anogenital setae covered with narrow phylliform cerotegument, smooth. Leg tarsi with 3 claws.
Nothrus xuejianensis Ermilov sp. n. (Figs 21–34)
Diagnosis
Body size: 713–763 × 332–381. Surface of prodorsum and notogaster alveolate. Rostral, lamellar and interlamellar setae covered with broad, phylliform cerotegument, smooth. Bothridial setae long, rod-like, sparsely barbed. Notogastral setae covered with broad, phylliform cerotegument, smooth; h2, c1, d1, d2, e1 longest. Distance between c1–c2 shorter than between c2–c3. Subcapitular setae h and m2 covered with narrow phylliform cerotegument, smooth, m1 and a setiform, barbed. Epimeral setal formula 7–4–6–7. Epimeral and anogenital setae covered with narrow phylliform cerotegument, smooth. Leg tarsi with one claw.
Description
Measurements. Body length: 730 (holotype, female), 713–763 (3 paratypes, 3 females); notogaster width: 348 (holotype), 332–381 (3 paratypes).
Integument (Figs 21, 32, 33). Body color yellow-brownish, covered with thin layer of gel-like cerotegument. Surface densely microfoveolate (visible under high magnification, × 1000). Prodorsum, notogaster and genital plates with rounded or elongate oval alveoles (their diameter or length up to 16).
Prodorsum (Figs 21–24). One unpaired mediobasal (in interbothridial region) and one pair of laterobasal (anteriorly to bothridia) depressions poorly visible. Rostrum broadly rounded, with distinct medial indentation. Rostral (20–24), lamellar (41–49) and interlamellar (53–61) setae covered with broad, phylliform cerotegument, smooth, ro and le inserted on tubercles, le–le connected by thin ridge. Bothridial setae very long (278–287), rod-like, sparsely barbed. Bothridial ridges distinct. Exobothridial setae (10–12) covered with narrow, phylliform cerotegument, smooth.
Notogaster (Figs 21, 25–28, 32, 33). Anterior notogastral margin slightly convex. Posterior part evenly rounded (not dilated). Weakly convex in dorsocentral and lateral parts and with circummarginal furrows between them. Caudal ridges not developed. Sixteen pairs of notogastral setae covered with broad, phylliform cerotegument, smooth, inserted on small tubercles; h2, c1, d1, d2, e1 (77–90) longer than f1, h1 (61–69), c3, cp, e2 (49–57) and c2, f2, h3, p1–p3 (36–41). Setae c1 d1, d2 reaching the insertions of d1, d2, e1, respectively. Distance between setae c1–c2 shorter than between c2–c3. Lyrifissures ia, im, ip, ih, ips and opisthonotal gland openings well visible, located in typical positions.
Gnathosoma. Generally, the morphology of gnathosoma, palps and chelicerae is similar to N. anauniensis. Subcapitulum longer than wide (157–164 × 135–143). Subcapitular setae h (20–22) and m2 (8) covered with narrow phylliform cerotegument, smooth, m1 (20–22) and a (24–26) setiform, barbed. Two pairs of adoral setae (24–26) present; or1 setifrom, smooth, or2 dilated distally, truncate, smooth. Palps (86–90) with setation 0–1–1–3–9(+1ω). Postpalpal setae (12–14) simple, erect. Chelicerae (157–164) with two setiform, barbed setae; cha (53–57) longer than chb (36–41). Trägårdh’s organ elongate triangular.
Epimeral and lateral podosomal regions (Fig. 32). Anterior tectum of epimere I distinctly incised medially. Epimeral setal formula 7–4–6–7. Setae short (14–16), covered with narrow phylliform cerotegument, smooth.
Anogenital region (Figs 29–33). Nine pairs of genital (16–20), two pairs of anal (16–20) and 3 pairs of adanal (ad1, 41–45; ad2, ad3, 22–24) setae covered with narrow phylliform cerotegument, smooth. Lyrifissures ian and iad clearly visible.
Legs (Fig. 34). Tarsi with one strong smooth claw. Generally, the morphology of leg segments, setae and solenidia is similar to N. anauniensis. Formulas of leg setation and solenidia: I (1–9–5–6–27) [1–2–3], II (1–8–5–5–25) [1–1–1], III (3[or 4]–5–5–5–22) [1–1–0], IV (2–6–5–5–22) [1–1–0]; homologies of setae and solenidia indicated in Table 1.

Nothrus anauniensis Canestrini and Fanzago, 1876, adult. 12 Ventral view (gnathosoma and legs except trochanters I–III not shown) 13 Posterior part of body, lateral view 14 Subcapitulum, ventral view 15 Palp, right, antiaxial view 16 Chelicera, right, antiaxial view

Nothrus anauniensis Canestrini and Fanzago, 1876, adult. 17 Leg I, right, antiaxial view 18 Leg II, right, antiaxial view 19 Leg III, right, antiaxial view 20 Leg IV, right, antiaxial view

Nothrus xuejianensis Ermilov sp. n., adult. 21 Dorsal view 22 Rostral seta 23 Interlamellar seta 24 Bothridial seta 25 Notogastral seta c1 26 Notogastral seta c2 27 Notogastral seta h2 28 Notogastral seta p1 29 Anal seta an1 30 Adanal seta ad1 31 Adanal seta ad2

Nothrus xuejianensis Ermilov sp. n., adult. 32 Ventral view (gnathosoma and legs except trochanters I–III not shown) 33 Posterior part of body, lateral view 34 Distal part of leg tarsus I, right, antiaxial view
Material examined
Holotype (female) and two paratypes (two females): Taiwan, Miaoli County, Tai’an Township, SheiPa National Park, Xuejian Area, 24°25.464’N, 121°00.796 ‘E, 1855 m a.s.l., sample #31, moss, 13.XI.2017 (collected by J.-R. Liao and H.C. Lee). One paratype (one female): Taiwan, Yilan County, Nan’ao Township, Cueifong Lake Circular Trail, 24°30.771’ N, 121°36.616′ E, 1885 m a.s.l., sample #24, soil, 5.XI.2017 (collected by J.-R. Liao and H.C. Lee).
Type deposition
The holotype (in ethanol with drop of glycerol) is deposited in the collection of NTU. Three paratypes (all in ethanol with drop of glycerol) are deposited in the collection of TSUMZ.
Etymology
The specific name xuejianensis refers to the Xuejian Area, where the holotype and some paratypes were collected.
Remarks
In general morphological traits (body elongate oval, without dilatation posteriorly; bothridial setae very long, rod-like; notogastral setae with broad, phylliform cerotegument, dorsomedial setae comparatively long; monodactylous legs), Nothrus xuejianensis Ermilov sp. n. is morphologically most similar to Nothrus meakanensis Fujikawa, 1999 from Japan and Nepal, but differs from the latter by the notogastral setae h2 with broad, phylliform cerotegument, similar to dorsomedial setae c1, d1, d2, e1 in length (versus h2 with thin cerotegument, considerably longer than other notogastral setae) and presence of two pairs of subcapitular setae m (versus 3 pairs).
References
Aoki J (1991) Oribatid mites of high altitude forests of Taiwan I. Mt. Pei-ta-wu Shan. Acta Arach 40:75–84
Aoki J (1995) Oribatid mites of high altitude forests of Taiwan II. Mt. nan-hu-ta Shan. Spec Bul Jpn Soc Coleopterology, Tokyo 4:123–130
Balogh J, Balogh P (2002) Identification keys to the oribatid mites of the Extra-Holarctic regions. Vol. 1. Well-Press Publishing Limited, Miskolc, p 453
Bayartogtokh B (2010) Oribatid mites of Mongolia (Acari: Oribatida). KMK, Moscow
Bayartogtokh B, Chatterjee T, Chan BK, Ingole B (2009) New species of marine littoral mites (Acari: Oribatida) from Taiwan and India, with a key to the World’s species of Fortuynia and notes on their distributions. Zool Stud 48(2):243–261
Beck L (1962) Beiträge zur Kenntnis der neotropischen oribatidenfauna. 2. Nothridae, Camisiidae, Heterobelbidae (Arach., Acari). Senck. Biol 43(5):385–407
Canestrini G, Fanzago F (1876) Nuovi Acari italiani. Atti della Societa Veneto-trentina di Scienze naturali residente in Padova 5:99–111
Chu YI, Aoki J (1997) Fauna of oribatid mite at Fu-Shan forest litter and humus layer. Chin J Ent 17(3):172–178
Ermilov SG, Hugo-Coetzee EA (2012) The oribatid mite genus Nothrus Koch, 1836 (Acari: Oribatida: Nothridae) of South Africa, including a key to African species. Zootaxa 3243:29–51
Ermilov SG, Liao J-R (2017a) Contribution to the knowledge of the oribatid mite genus Brassiella (Acari: Oribatida: Caloppiidae). Biologia 72(9):1041–1048. https://doi.org/10.1515/biolog-2017-0115
Ermilov SG, Liao J-R (2017b) Neotype designations and supplementary descriptions of Taiwanoppia subtropica Tseng, 1982 and Muliercula chiayiensis Tseng, 1984 (Acari: Oribatida: Oppiidae, Scheloribatidae). Syst Appl Acarol 22(7):897–914. https://doi.org/10.11158/saa.22.7.1
Ermilov SG, Liao J-R (2017c) New faunistic and taxonomic data on oribatid mites (Acari, Oribatida) of Taiwan. Syst Appl Acarol 22(6):824–840. https://doi.org/10.11158/saa.22.6.8
Fujikawa T (1999) Eight new species of the genus Nothrus. Edaphologia 63:5–54
Hammen L van der (1950) On two interesting Oribatei (Acari) from the Sint Pietersberg (Southern Limburg, Netherlands). In: Mededelingen van de Commissie inzake wetenschappelijk onderzoek van de St Pietersberg No 11. Naturhist Maandbl 39(2):129–132
Hammer M (1966) Investigations on the oribatid fauna of New Zealand. Part I. Det Kong Dansk Vidensk Selsk. Biol Skr 15(2):1–108
Koch CL (1835) Deutschlands Crustaceen. Myriapoden und Arachniden. Regensburg. Heft:1–3
Koch CL (1839) Deutschlands Crustaceen. Myriapoden und Arachniden. Regensburg. Heft:28–30
Lochynska M (2008) Nothrid mites from Ecuador – recapitulation (Acari: Oribatida: Nothridae). Genus 19(1):143–149
Mahunka S (1978) Neue und interessante milben aus dem Genfer museum XXVII. A first survey of the oribatid (Acari) fauna of Mauritius, Reunion and the Seychelles I. Rev Suisse Zool 85(1):177–236
Norton RA (1977) A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal DL (ed) Biology of oribatid mites. SUNY College of environmental science and forestry, Syracuse, pp 33–61
Norton RA, Behan-Pelletier VM (2009) Suborder Oribatida. Chapter 15. In: Krantz GW, Walter DE (eds) A manual of acarology. Texas Tech University press, Lubbock, pp 430–564
Ohkubo N (1995) A new oppiid mite (Acari: Oribatida) from Taiwan. Acta Arach 44(1):61–64
Olszanowski Z (1996) A monograph of the Nothridae and Camisiidae of Poland (Acari, Oribatida, Crotonioidea). Genu (Suppl):1–201
Sellnick M, Forsslund KH (1955) Die Camisiidae Schwedens. Ark Zool 8(4):473–530
Seniczak S, Norton RA (1993) The morphology of juvenile stages of moss mites of the family Nothridae (Acari, Oribatida). III. Zool Anz 230(1–2):19–33
Sitnikova LG (1975) The family Nothridae Berlese, 1896. In: Ghilyarov MS (ed) Key to soil inhabiting mites, Sarcoptiformes. Nauka press, Moscow, pp 72–75
Subías LS (2004) Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Graellsia 60:3–305. Online version accessed in January 2018, 605 pp.; http://bba.bioucm.es/cont/docs/RO_1.pdf
Travé J, Vachon M (1975) François Grandjean. 1882–1975 (Notice biographique et bibliographique). Acarologia 17(1):1–19
Tseng Y (1982) Taxonomical study of oribatid mites from Taiwan (Acarina: Astigmata) (I). Chin J Ent 2(1):53–106
Tseng Y (1984) Taxonomical study of oribatid mites from Taiwan (Acarina: Astigmata) (II). Chin J Ent 4:27–74
Weigmann G (2006) Hornmilben (Oribatida). Die Tierwelt Deutschlands. Teil 76. Goecke and Evers, Keltern
Acknowledgments
We cordially thank two anonymous reviewers for the valuable comments; and SheiPa National Park for permission of collections.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
No potential conflict of interest was reported by the authors.
Rights and permissions
About this article

Cite this article
Ermilov, S.G., Liao, JR. To the knowledge of oribatid mites of the genus Nothrus (Acari, Oribatida, Nothridae) from Taiwan. Biologia 73, 513–521 (2018). https://doi.org/10.2478/s11756-018-0062-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2478/s11756-018-0062-1