
Abstract
Background
Hyptis suaveolens (L.) Poit., native to the American tropics, is a pantropical annual plant and a major invasive species throughout India. It was anticipated that the availability of sunlight, coupled with its superior reproductive potential, persistent propagule bank, and dispersal ability, could lead to an increase in the growth and spread of this invader, thus potentially impeding herbaceous growth and diversity in non-native areas. Clarifying its ecological fitness and competitive performance will be useful to manage the spread of H. suaveolens in natural ecosystems that are facing a wide range of anthropogenic pressures.
Methods
The present study is a three-tier experiment. In the first tier, a field study was conducted to assess the patterns of H. suaveolens abundance and herbaceous species diversity in response to light availability (sun, 842–1072 µmol m–2 s−1 and shade 253–341 µmol m–2 s−1) in the tropical dry deciduous ecosystems in the Vindhyan highlands, India. Furthermore, the impact of H. suaveolens abundance on the resident native and non-native species abundance and diversity was also studied. In the second tier, a randomized common garden experiment was conducted to understand the trait fitness of H. suaveolens in sun (940 µmol m–2 s−1) and shade (300 µmol m–2 s−1) conditions. In the third tier, a plant growth chamber experiment with high-light (940 µmol m–2 s−1) and low-light (300 µmol m–2 s−1) treatments was done to learn how H. suaveolens partitions its biomass between aboveground and belowground plant parts.
Results
The field study indicated that the sunlit areas had a higher abundance of H. suaveolens and a lower diversity of resident herbaceous species than the shaded areas. The common garden experiment showed that sun-dwelling H. suaveolens individuals performed better in germinative, vegetative, eco-physiological, and reproductive traits than the shade-dwelling individuals. The growth chamber experiment exhibited that plants grown in high-light environment had greater seed germination, seedling recruitment, and aboveground biomass than those grown in low-light environment, whereas plants grown in low-light environment exhibited a higher root mass ratio than the high-light individuals. These results suggest that H. suaveolens individuals mask the understory vegetation owing to higher seedling recruitment, relative growth rate, photosynthetic performance, resource acquisition-allocation, and reproductive output in response to high-light conditions.
Conclusions
The study concludes that light significantly controls the invasive population dynamics of H. suaveolens in dry deciduous forests. In high-light areas, H. suaveolens populations dominate the forest understory with suboptimal shade tolerance. In shade environment, H. suaveolens maintains a persistent soil seed bank along with ‘Oskar individuals’ that become active in response to high-light availability. The modus operandi is a ‘sit and wait’ strategy. The current study provides insights on prioritizing areas for H. suaveolens management that will potentially reduce the risk of biological invasions on the native species diversity of tropical regions.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Biological invasion is one of the most serious and biggest ecological threats to global biodiversity (Lenzner et al. 2020; Bellard et al. 2022). A significant number of invasive plant species have been reported to spread widely and pose detrimental effects on native vegetation through their ability to colonize and proliferate, viz.: Ageratum conyzoides L. (Singh et al. 2011; Sun et al. 2012), Alternanthera philoxeroides Mart. (Pan et al. 2007; Tao et al. 2009), Chromolaena odorata (L.) R.M. King & H. Rob. (Li et al. 2022), Imperata cylindrica (L.) Raeusch. (Hameed et al. 2009), Ipomoea cairica L. (Liu et al. 2012), Lantana camara L. (Goyal and Sharma 2019), Miconia calvescens DC. (Leary et al. 2018), Mikania micrantha Kunth (Banerjee et al. 2017), Parthenium hysterophorus L. (Adkins and Shabbir 2014), Prosopis juliflora (Sw.) DC. (Shirke et al. 2018), etc. Enhanced efficiency in capturing and utilizing available space, water, nutrients, and light is reported to be a strategy acquired by the invasive species for their success in the non-native ranges (van Kleunen et al. 2010; Heberling and Fridley 2013; Ziller et al. 2019). Light is an essential resource that affects the performance of invasive species, especially in disturbed and heterogeneous habitats (Chazdon et al. 1996). Morphological and physiological plant responses to different light conditions are manifested in their light capture and utilization efficiency to maximize carbon acquisition (Bazzaz and Carlson 1982; Poorter 1999). Under resource surplus environments, an invader’s ability to generate rapid biomass is often considered a key factor for facilitating invasion success and could be a function of resource acquisition strategy (Leishman et al. 2007). Thus, the resource acquisition strategy exhibited by invasive plants in response to heterogeneous light availability in new environments may enhance our understanding of the mechanism behind the competitive fitness of invasive species. This mechanism could be utilized as a basis to control the naturalization and colonization of invasive species in natural ecosystems (Holt 1995).
An invader’s performance ability can be deciphered through its varied plant functional traits and plastic responses to a wide range of environments (Kueffer et al. 2013; van Kleunen et al. 2015; Funk et al. 2016) that are de facto important determinants of its establishment, proliferation, and spread (Hui et al. 2023). Plastic responses are reported as environment-induced changes in morphology, physiology, and reproductive traits accounting for the interactions between a species’ development scheme and habitat characteristics (Leishman et al. 2000; Sharma and Esler 2008). In high-light, the invasive plants with pronounced photosynthetic performance (Fridley 2012) show rapid and high germination than do natives (van Kleunen and Johnson 2007; Sharma et al. 2010; Flores-Moreno and Moles 2013), thus providing invasive plants with higher chances of recruitment and survival during early life-stages in native environments (Honig et al. 1992; Klink 1996; Chrobock et al. 2011). However, it may not be true for all native plant species in most of the local environments; there is likely to be a pool of resident native species with similar plasticity and high germination, but their abundances are likely to be reduced by local enemies (Keane and Crawley 2002). Thus, understanding the plastic responses of an invader under varied light conditions could be of immense importance to study its recruitment, establishment, and proliferation in the regions that are receiving or are likely to receive comparatively high-light availability created by anthropogenic disturbances.
Vindhyan highlands, situated between the Indo-Gangetic plains and the Narmada valley, cover an area of ∼ 1086 km of west-central Indian states, namely Bihar, Gujarat, Madhya Pradesh, and Uttar Pradesh (Pant et al. 1992). The region is reported as one of the most threatened forested areas under high risk of anthropogenic disturbance and deforestation (Roy et al. 2002; Sagar and Singh 2003; Goparaju and Sinha 2015). Ecological field studies from the area reported that owing to rapid industrialization, urbanization, and biotic pressure, the forest cover is decreasing thus leading to the conversion of forested area into open marginal lands with light heterogeneity (Jha 1990; Singh et al. 1989, Sagar et al. 2003; Sharma 2007a). Studies have highlighted the need to conserve the remnant natural forest areas that are experiencing the loss of species-rich ecosystems of Vindhyan highlands (Sagar et al. 2003; Sagar and Singh 2003; Sharma 2007a). Anthropogenic disturbances in natural forests lead to increased light availability, which in turn facilitates environmental heterogeneity and may create micro-niches with altered resource availability for the invasive species (Kerns and Day 2017; Meyer et al. 2021). Dominant plant invasive species colonize fragmented dry deciduous forest areas of Vindhyan highlands, thus could lead to the loss of native plant diversity (Sharma et al. 2009; Raghubanshi and Tripathi 2009).
Agrawal (2002) reported that Hyptis suaveolens (L.) Poit., an aggressive plant invader, had spread extensively in the intact forests of the Vindhyan highlands. In interior forest openings of the Vindhyan highlands, heavy infestations of H. suaveolens monoculture thickets have also been reported (Sharma 2007b; Sharma et al. 2009; Sharma and Raghubanshi 2009; Sharma et al. 2017). Kumar et al. (2019) advocated that habitat size, land use, and management practices affect the H. suaveolens invasive population and reported a severe infestation of H. suaveolens in open areas of water stream networks of Indian Western Himalayan regions. Studies have indicated that H. suaveolens forms dense monospecific thickets, potentially reducing light penetration to the forest floor, thus masking the growth of other herbaceous species (Raizada 2006; Sharma et al. 2009, 2017; Afreen et al. 2018). This strategy may explain its negative effects on native species diversity in disturbed environments. According to Padalia et al. (2014), under the current changing environmental scenario, the potential distribution of H. suaveolens is vast and would be far more extensive than its current geographical extent. Therefore, identifying a mechanism that restricts the growth and abundance of H. suaveolens in response to the changing environment could be a need for the conservation and management of natural ecosystems. However, there is less systematic study to elucidate H. suaveolens abundance/dominance pattern in response to light heterogeneity that may provide a clue to understanding the distribution and expansion of H. suaveolens in natural habitats.
It needs to be factored in here that the tremendously increasing anthropogenic activities (encroachment and deforestation) create forest gaps and allow light penetration to the ground surface, which facilitates the rapid spread of H. suaveolens in natural forests. Assessing the impact of H. suaveolens dominance on native plant species composition and diversity and understanding the implications of H. suaveolens’ trait responses to imminent changes in resource availability—light heterogeneity—could be of utmost importance for the restoration and conservation of degraded ecosystems. Field observations provide insights that can be tested by controlled laboratory experiments. Conversely, the results of controlled experiments produce ingenious approaches to be tried and investigated in field studies (Farrugia et al. 2010). Moreover, common garden experiments with similar or comparable field environmental conditions assure reasonable internal validity while adhering to a degree of naturalism necessary for generalizability (de Villemereuil et al. 2016). Based on these principles, a three-tier study was conducted for a thorough understanding of the invasion potential of H. suaveolens.
The field, common garden, and growth chamber studies provide a new approach for exploring the mechanisms that lead to the spread of H. suaveolens in natural dry tropical ecosystems. Interestingly, field-based studies of H. suaveolens under variable environmental conditions are scarce and field-based plant trait performance studies in tandem with common garden and growth chamber experiments of H. suaveolens under heterogeneous light conditions are totally missing. Thus, the present study is essential for the conservation and management of the depleting diversity of the Vindhyan dry tropical ecosystem. With this as the rationale, the study (i) assesses the changes in herbaceous species composition in response to variable light regimes, (ii) understands the pattern of native and non-native herbaceous species diversity with an increasing abundance of H. suaveolens, (iii) explores native, non-native, and H. suaveolens abundance in sun and shade conditions in fields, and (iv) looks into how high-light and low-light conditions modulate H. suaveolens trait responses through germinative, vegetative, eco-physiological, and reproductive life-history traits in controlled systems.
Materials and methods
The study is comprised of field, common garden, and growth chamber experiments. The field study was conducted to estimate abundance of H. suaveolens and richness and diversity of native and non-native species in the sun and shade conditions of Vindhyan highlands. The common garden experiment was carried out to estimate the life-history traits viz.: germinative, vegetative, eco-physiological, and reproductive traits of H. suaveolens individuals in sun and shade conditions. A growth chamber experiment was conducted to study aboveground and belowground plant biomass partitioning of H. suaveolens in response to high-light and low-light treatments and corroborate common garden experiment findings. Conclusively entire study was integrated to understand the performance of H. suaveolens life-history traits and subsequent effects of its dominance on the diversity of ground vegetation in natural ecosystems of Vindhyan highlands.
Study species
Hyptis suaveolens, a pantropical invader, is a native plant of tropical America. It is a rigid annual herb of aggressive nature and produces an ample amount of seeds (∼ 3000 seeds m2) (Sharma et al. 2009). It is known that prolific seed production, small seed size, persistent propagule bank, and superior reproduction could confer high colonization ability and, thereby, the abundance of this species (Raizada 2006). In India, it is a notably prominent invader in natural areas and wastelands (Verma and Mishra 1992; Mudgal et al. 1997; Raizada 2006; Sharma et al. 2009; Sharma and Raghubanshi 2009; Sharma et al. 2017; Afreen et al. 2020).
Field study
Study area
The study was conducted in the Vindhyan highlands of west-central India. Based on a reconnaissance survey of the entire Vindhyan highlands, ten sites (24° 33′ – 24° 56′ N; 83° 04′ – 83°13′ E) in Sonebhadra district (also known as golden valley) of Uttar Pradesh, India were randomly selected to represent the entire range of environmental conditions (viz.: topography, anthropogenic pressures, forest clearings, forest edges, light availability, and thickets of H. suaveolens). The area experiences a tropical monsoon climate with three distinct seasons: summer, winter, and rainy (Singh et al. 2010). The average annual temperature and rainfall are 25.2 °C and 997 mm, respectively. The soils show a rocky sandy-loam texture with nutrient-deficient Ultisols and are reddish to dark grey (Singh et al. 1989).
The region is characterized by intensive anthropogenic disturbances such as the widespread cutting of trees and understory vegetation for fuel and fodders (Sagar et al. 2003; Raghubanshi and Tripathi 2009). The region is experiencing the conversion of forest areas into agricultural fields, pastures, and sites for mining and quarrying activities (Goparaju and Sinha 2015). These activities are transforming the intact natural forest areas of Vindhyan highlands into non-contiguous vegetation patches of varying sizes of homogeneous (dominated by monospecific thickets) and heterogeneous (mixed species) plant communities (Jha and Singh 1990; Singh et al. 1989; Sagar and Singh 2006). Consequently, this creates spatial gradients of light availability to understorey vegetation. Occasionally during summer, anthropogenic fires augment ecosystems’ susceptibility to plant invasions (Hiremath and Sundaram 2005).
The overstory vegetation is tropical dry deciduous forest locally dominated by Buchanania lanzan Spreng., Boswellia serrata Roxb., Diospyros melanoxylon Roxb., Hardwickia binata Roxb., Lannea coromandelica (Houtt.) Merr., Lagerstroemia parviflora Roxb., Madhuca longifolia (L.) J.F. Macbr., Senegalia catechu (L.f.) P.J.H. Hurter & Mabb., Shorea robusta C.F. Gaertn., Tectona grandis L.f., Terminalia anogeissiana Gere & Boatwr., Terminalia elliptica Willd., etc. (Sagar et al. 2003; Sagar and Singh 2006; Sharma and Raghubanshi 2009). These tree species are native to India (Champion and Seth 1968). The understory vegetation is mostly dominated by annual and perennial herbaceous species [Alysicarpus vaginalis (L.) DC, Corchorus trilocularis L., Grona triflora (L.) H. Ohashi & K. Ohashi, Leucas biflora (Vahl) Sm., Senna tora (L.) Roxb., Sida spinosa L., etc.] with few subshrubs [Crotalaria albida B. Heyne ex Roth, Eragrostis tenella (L.) P. Beauv. ex Roem. & Schult., Xanthium strumarium L. etc.], shrubs (Lantana camara L., Euphorbia tithymaloides L. etc.), climbers (Basella alba L., Physalis angulata L. etc.), and tree seedlings. Among the listed herbaceous plants, C. trilocularis, G. triflora, S. spinosa, E. tenella, X. strumarium, L. camara, and P. angulata are non-native species (Tripathi 2003; Sharma et al. 2009; Afreen et al. 2018). Kushwaha et al. (2011) showed that the temporal pattern of leaf flush and leaf fall for deciduous tree species exhibits considerable overlapping timings and durations of deciduousness (leafless period) in the dry deciduous forest of Vindhyan highlands. Such varied temporal pattern of deciduousness does not allow the canopy foliar cover to be completely leafless, thus creating a light heterogeneous environment for understory vegetation.
Vegetation sampling and light estimation
On each site, ten random plots were marked, and within each plot, a 1 × 1 m quadrat was randomly laid to sample the ground vegetation and estimate light intensity. Thus, 100 quadrats (10 sites ×10 quadrats) were used in the analysis. For the estimation of light intensity of each 1 × 1 m quadrats, photosynthetic photon flux density (PPFD) was measured using a handheld quantum sensor (CI-340, CID Bio-Science, Inc., USA) in the months of August–October 2018–2019. During this period, the durations of vegetative pheno-phases (i.e. leaf flush and leaf fall) vary over several weeks to a few months amongst deciduous tree species, leading to considerable variation in seasonal light availability to understory vegetation (Kushwaha and Singh 2005; Kushwaha et al. 2010). To nullify the site-specific effects, the highest light-receiving quadrats from each of the ten sites were clubbed into a (total of ten 1 × 1 m quadrats) highest light-receiving plot and named P1. In the next step, slightly lower light-receiving quadrats from each of the ten sites were clubbed into a slightly lower light-receiving plot and named P2. This exercise was repeated for the remaining light-level plots (P3 to P10). Thus, the P1 plot had maximum light while P10 had minimum. Further, based on the PPFD values, these plots were divided into the sun (high-light, 842–1072 µmol m–2 s−1), and shade (low-light, 253–341 µmol m–2 s−1) groups. Consequently, the plots P1 to P5 were under the sun, and P6 to P10 were under shade groups (Supplementary Table 1).
During the peak growing season of 2018–2019, quadrat-wise total numbers of individuals of each herbaceous species were recorded to understand the composition and diversity of ground vegetation. The peak growing season for understory herbaceous species lies between August and November before the onset of the dry period (December–June) (Kushwaha et al. 2011). Plant individuals for each 1 × 1 m quadrat were enumerated to estimate alpha diversity. Plant individuals were defined as healthy recruited plants that had lost their first cotyledonary leaves (Martins et al. 2011). A runner (stoloniferous plant) can represent multiple individuals, but all shoots are considered a single individual at the point where the shoots and roots emerge (Nabe-Nielsen 2001). Raunkiær (1934) classification of life-form spectra was used to characterize the species assemblage in each quadrat. The life forms and growth forms were identified with the help of the Flora of Raipur, Durg, and Rajnandgaon (Verma et al. 1985). The enumerated species were categorized into native and non-native with the help of plant taxonomists, Flora of Raipur, Durg, and Rajnandangaon (Verma et al. 1985), Flora of Allahabad (Verma and Mishra 1992) and previously published research articles from the study area (Tripathi 2003; Sharma et al. 2009; Afreen et al. 2018).
Species richness, a component of alpha diversity (Sagar and Singh 1999) was determined by measuring the number of species m− 2 (Whittaker 1972). Fisher’s alpha diversity was calculated using BioDiversity Professional software (McAleece 1997). To determine the species dominance for each plot, Importance Value Indices (IVIs = relative frequency + relative density + relative dominance) for each species were calculated (Curtis 1959). For trait analysis, ten mature and healthy individuals of H. suaveolens of each quadrat were selected for the estimation of aboveground biomass (g m−2), number of seeds per plant, seed size (mm), and seed weight (mg). Each parameter was represented by taking the means of total quadrats of each plot (N = 10 quadrats for each plot) of each sun and shade category (10 quadrats × 5 plots × 2 light levels = 100 quadrats). Finally, the study reports the IVI of each species, diversity parameters, and functional traits for each plot of the sun (P1 to P5) and shade (P6 to P10) conditions.
Common garden experiment
Seeds of H. suaveolens were collected from a single population growing in the natural habitats of selected plots and stored at room temperature in paper bags. The common garden experiment was set up in a garden plot at the University of Delhi, Delhi, India. Equal proportions (v/v/v) of garden soil, solarite, and coco peat were used to prepare the soil composition of the sun and shade experimental setups. H. suaveolens seeds were sown in 250 pots (∼ 1700 cm3) filled with experimental soil. Equal numbers of pots were kept in full sunlight for sun treatment and under a system covered with ‘Tygen’ plastic mesh for shade treatment. The ‘Tygen’ plastic mesh covering system reduced the light intensity by ∼ 32% of full sunlight (sun, 940 µmol m–2 s−1 and shade, 300 µmol m–2 s−1). Sun and shade treatments were designed to match the conditions of field study plots. Moisture levels of the pots were measured using the Theta probe (Delta-T devices Ltd., England) on every alternate day and kept constant at field capacity. Seed germination and seedling recruitment were recorded. Seedlings were counted as recruited after they had lost their cotyledons (Martins et al. 2011). Pots were randomly rotated weekly within each treatment level to maintain a homogeneous effect.
The performance of H. suaveolens due to sun and shade treatments was analyzed by estimating the germinative, vegetative, eco-physiological, and reproductive life-history traits. For germinative traits i.e.: maximum germination percentage (MGP) and maximum recruitment percentage (MRP) were quantified. Plant height (cm) and number of leaves per plant were recorded to understand vegetative traits. Chlorophyll content index (CCI) (CCM-200 plus, Apogee Instruments, Inc., USA), photosynthetic rate (A) (CI-340, CID Bio-Science, Inc., USA), stomatal conductance (C), transpiration rate (E), specific leaf area (SLA), leaf construction cost (LCC), relative growth rate (RGR), net assimilation rate (NAR), and leaf mass ratio (LMR) were estimated to understand the responses of eco-physiological traits of H. suaveolens to the sun and shade treatments. Reproductive traits were assessed by estimating number of seeds per plant, seed size (mm), and seed mass (mg).
From the date of sowing (DOS), a set of 3 plants with three replicates from each treatment was randomly harvested fortnightly until the end of the experiment. Harvested plants were transported to the laboratory for further analysis. Plants were then separated into leaves, stems, and seeds. For each plant, leaf area (cm2) was estimated using a leaf area meter (Model 211, Systronics, India), and fresh biomass (g) of each plant part was weighed. All plant parts were oven-dried at 80 °C for 72 h to a constant weight, and then dry biomass (g) was computed. Harvested plant data was used to calculate the SLA (leaf area/dry weight). Water use efficiency (WUE) was calculated as ratio of photosynthetic rate (A) and transpiration rate (E). For each sampled plant, dried leaves were used to estimate the LCC (Barthod and Epron 2005; Singh et al. 2011; Goyal et al. 2018).
RGR for plant height (RGRh, cm cm−1 fortnight−1) and plant biomass (RGRb, g g−1 fortnight−1) were calculated using the expression: RGR = [(logef – logei)/(dt)] based on growth parameters at first and consecutive harvests (Kramer and Kozlowski 1979). Here, f and i are the final and initial measured values, and dt is the time period. Selected variables were plant height (cm) and plant biomass (g). NAR (g cm−3 fortnight−1) was calculated using the expression: RGRb = NAR×LMR×SLA (McKenna and Shipley 1999; Grotkopp and Rejmánek 2007). Here, leaf mass ratio (LMR) is the ratio of leaf dry weight (g) and plant dry weight (g).
Performance-related trait values of H. suaveolens from the common garden experiment were used to calculate the plasticity index (PI). The plasticity index, ranging from 0 (no plasticity) to 1 (maximal plasticity), was computed as the difference between maximum and minimum mean values divided by maximum mean values of all studied traits for sun- and shade-dwelling H. suaveolens individuals. Comparative assessment of plasticity was done by dividing these values into three range categories, viz.: low (PI, 0.00–0.33), medium (PI, 0.34–0.66), and high (PI, 0.67–1.00) plastic responses as suggested by Valladares et al. (2005) and Gratani et al. (2014).
Growth chamber experiment
The insight gained from the common garden experiment was used to set up the growth chamber experiment to study the selected life-history traits, i.e., germinative, vegetative, and eco-physiological traits, and understand the pattern of aboveground and belowground plant biomass partitioning of H. suaveolens under controlled environment. The biomass partitioning of H. suaveolens was assessed to understand the effects of light stress as it was hypothesized that plants allocate more biomass to aboveground than belowground under high-light conditions. Mixed experimental soil and seeds used for the common garden experiment were also used for the growth chamber experiment. H. suaveolens seeds were sown in small gardening pots (height 18 cm and diameter 20 cm) with three replicates of 100 seeds each for high-light and low-light treatments following a completely randomized design. High- and low-light conditions of growth chambers were maintained, similar to those of sun and shade treatments of common garden experiments (high-light, 940 µmol m–2 s−1 and low-light, 300 µmol m–2 s−1). The ambient temperature and relative humidity of the plant growth chamber (Model C 70, Saveer biotech limited, India) were kept constant at 28 °C and 50%, respectively. Soil moisture was maintained constant at the field capacity.
Seed germination and seedling recruitment were counted until the end of the experiment, and germinative traits, i.e., maximum germination percentage (MGP) and maximum recruitment percentage (MRP), were calculated. For each plant, plant height (cm) and the number of leaves were recorded to understand the response of vegetative traits to light variability. Forty-five and sixty days old seedlings were harvested for biomass estimation. Individuals were separated into stems, leaves, and roots, which were weighed to estimate dry biomass (g). Leaf area (cm2) was estimated using a leaf area meter (Model 211, Systronics, India). Eco-physiological traits were assessed by estimating SLA, CCI, RGRb, and NAR. Stem mass ratio (SMR, stem biomass/plant biomass), root mass ratio (RMR, root biomass/plant biomass), and aboveground-belowground biomass ratio (ABBR, aboveground biomass/belowground biomass) were estimated to understand the pattern of aboveground and belowground plant biomass partitioning of H. suaveolens (Engelkes et al. 2008; Peng and Yang 2016) to the varied light levels.
For a better understanding of H. suaveolens invasion success, its life-history traits were schematically depicted and divided into four subsequent stages: propagule phase, germination phase, recruitment phase, and fecundity phase (Harper 1977).
Statistical analyses
The species richness of native and non-native herbaceous species was correlated with the biomass of H. suaveolens. Herbaceous species diversity indices were linearly regressed with the IVI of H. suaveolens to understand the effect of H. suaveolens’s abundance on the diversity of ground vegetation of the study area. Linear regression was used to test the effect of PPFD (light intensity) on the abundance (IVI) of H. suaveolens, native and non-native species. The plots (P1–P10) were ordinated using Principal Component Analysis (PCA) ordination with the help of BioDiversity Professional software (McAleece 1997) to understand the differences in species composition due to variable light conditions. Furthermore, to have an overall measure of species richness of the study site, a plot-based rarefaction curve (classical species–area curves type IV, suggested by Scheiner 2003) was plotted between the cumulative number of species of each plot as dependent and the number of plots sampled as independent variables (Gotelli and Colwell 2001; Hsieh et al. 2016) using BioDiversity Professional software (McAleece 1997). The effect of light conditions (sun and shade) on H. suaveolens’s life-history traits was analyzed using a one-way analysis of variance (ANOVA). Pearson correlation was performed to evaluate relationships among studied plant traits of the common garden experiment. Paired T-test was also performed to analyze differences in trait values between light treatments (high- and low-light) for studied plant traits yielded by the growth chamber experiment. Statistical analyses were conducted using SPSS 16.0 (SPSS Inc. 2007, Chicago, USA).
Results
Field study
The floristic survey of the study area depicted 59 species (Supplementary Table 1). The sun and shade areas harboured 59 species belonging to 23 families and 42 species belonging to 19 families, respectively. Among these families, Poaceae, Malvaceae, Acanthaceae, and Leguminosae were the most species-rich families in both areas. The recorded herbaceous species were an assemblage of annuals, perennials, and a few annual species showing a tendency to be perennial. The sunlit area had 26 herbs, 13 subshrubs, two mat-forming herbs, and one climber species, while the shaded area had 39 herbs, 17 subshrubs, two mat-forming herbs, and one climber species (Supplementary Table 1). The mean IVI value of H. suaveolens in the sun area was 94 times greater than in the shaded area. The IVI values ranged from 0.13 (Triumfetta pentandra A. Rich.) to 82.50 (Hyptis suaveolens) and from 0.07 (Xanthium strumarium L.) to 9.6 (Ceropegia bulbosa Roxb.) for the sun and shade areas, respectively. Thus, H. suaveolens was heavily dominated in sunlit areas than shaded areas (hereinafter, sun and shade areas will be used as H. suaveolens invaded and non-invaded areas, respectively).
The PCA ordination based on IVI of component species for different plots is presented in Fig. 1. The ordination diagram showed two distinct clusters (each having five plots) indicating the changes in species composition due to light regimes (Fig. 1). The mean IVI values of resident herbaceous species significantly decreased with the increasing dominance of H. suaveolens under sun conditions (Fig. 2a). There was a reduction of 51.55% and 73.42% in species richness and alpha diversity, respectively, in the invaded area compared to the non-invaded area. For non-invaded areas, the proportions of native and non-native species were 61.19% and 38.81%, respectively. Interestingly, the proportion of non-native species (88.10%) was exceedingly higher than that of native species (11.90%) in the invaded area (Supplementary Table 1).

Principal component analysis (PCA) ordination of ten dry tropical plots (P1 to P10) based on the importance value indices (IVI) of component herbaceous species distributed in two light regimes (sun, 842–1072 µmol m–2 s−1 and shade, 253–341 µmol m–2 s−1)

(a) Relationships between importance value indices (IVI) of native, non-native and Hyptis suaveolens individuals and photosynthetic photon flux density (PPFD); (b) Rarefaction curves of plant species richness of ten dry deciduous plots (P1 to P10)
The results of plot-based rarefaction curves showed clubbing of curves into two distinct groups, i.e., plots 1, 2, 3, 4, and 5 into the invaded group and plots 6, 7, 8, 9, and 10 into the non-invaded group (Fig. 2b). In the rarefaction curves, the curves of the non-invaded group occupied a higher position than the curves of the invaded group, indicating higher species richness for the non-invaded area than the invaded area. Thus, light regimes caused distinct differences in herbaceous species composition and richness in the Vindhyan highland. Results showed that the diversity of herbaceous species decreased with the increasing dominance of H. suaveolens (Fig. 3). Sun-dwelling H. suaveolens individuals showed more than two times higher biomass than shade-dwelling individuals. Additionally, results indicated enhanced light-mediated dominance of H. suaveolens and a reduction of total herbaceous richness (Fig. 4a), as well as reduced richness and alpha diversity of native and non-native groups in sun than shade conditions (Fig. 4b). These patterns reflected the light-induced masking of H. suaveolens on the herbaceous diversity of Vindhyan highlands.

Response of herbaceous diversity to the dominance (IVI, importance value indices) of Hyptis suaveolens in the studied field plots of Vindhyan highlands, India suggesting a masking effect of H. suaveolens on other resident herbaceous species

(a) Plant biomass (g m−2) of Hyptis suaveolens as axis Y1 and herbaceous species richness as axis Y2, and (b) species richness and alpha-diversity of native and non-native plant species in response to two light levels (sun, 842–1072 µmol m–2 s−1 and shade, 253–341 µmol m–2 s−1) in Vindhyan highlands suggesting enhanced light-mediated dominance of H. suaveolens and reduction of native and non-native herbaceous diversity
The H. suaveolens individuals in sunlit areas had 3.2 times more seeds per plant than the shade-dwelling individuals. Based on cumulative results from all ten plots, plant biomass showed significantly negative relations with those of seed size (r2 = 0.23, p < 0.05) and seed mass (r2 = 0.30, p < 0.05). Sun-dwelling H. suaveolens individuals produced smaller and lighter seeds (in terms of seed size and seed mass). In contrast, shade-dwelling individuals produced more significant and heavyweight seeds, suggesting more remarkable spatial dispersal ability and fitness of H. suaveolens in sunlit areas than in shaded areas.
Common garden experiment
Sun-dwelling H. suaveolens individuals showed significantly higher seed germination and seedling recruitment than shade-dwelling H. suaveolens (Table 1). Shade-dwelling H. suaveolens were shorter, with a lower number of leaves per plant as compared to sun-dwelling plants. Similar to the field study, sun-dwelling individuals were more robust and sturdy in terms of biomass per plant as compared to shade-dwelling individuals (Fig. 5). Leaf area was more than two times higher for shade-dwelling individuals than the sun-dwelling individuals. In contrast, SLA was significantly lower for plants grown in sun treatment. Sun-dwelling individuals exhibited higher RGRh, RGRb, NAR, photosynthetic rate, stomatal conductance, and transpiration rate than shade-dwelling individuals. Seed size and seed weight per plant were lower for sun-dwelling individuals than for shade-dwelling individuals of H. suaveolens (Table 1). Similar to the field study, the number of seeds produced per plant was high (193) for sun-dwelling individuals, whereas shade-dwelling individuals produced a significantly low number (∼ 32) of seeds per plant (Table 1). These results favor the dominance of H. suaveolens in sunlit areas than in shaded areas.

Plant biomass of Hyptis suaveolens individuals grown in two different light treatments in the common garden experiment indicating rapid biomass generation in sun condition. Close and open circles signify dry biomass values for sun (high-light, 940 µmol m–2 s−1) and shade (low-light, 300 µmol m–2 s−1) inhabiting plant individuals
Plasticity index (PI)
Ten of the 19 parameters selected for comparative plasticity assessment reflected medium plastic responses (Table 1). Across the studied life-history traits, the plasticity index (PI) ranged from 0.02 (for LCC) to 0.83 (for the number of seeds per plant). For germinative traits, PI values were medium for the MGP and high for the MRP. H. suaveolens showed medium plasticity in its vegetative traits, except for leaf per plant which had a high range of plasticity. Likewise, eco-physiological traits of H. suaveolens also reflected medium plasticity except for CCI, A, and LCC which were in a low range of plastic responses. The NAR showed a high range of plasticity. For reproductive traits, the number of seeds per plant fell under a high range of plasticity, whereas seed size and seed mass expressed a low range of plasticity (Table 1). Thus, except for the CCI, A, and LCC eco-physiological traits, all the studied traits exhibited a medium to high range of plastic responses.
Plant growth chamber experiment
Paired T-test showed significant differences in maximum germination percentage (t = –4.84, p < 0.001), root mass ratio (t = 15.00, p < 0.001), and aboveground-belowground biomass ratio (t = –19.34, p < 0.001) due to differences in light conditions. Results showed higher plant biomass generation of H. suaveolens for individuals grown in the high-light than in the low-light conditions (Fig. 6). Under high-light, H. suaveolens exhibited almost twice MGP and MRP than in low-light. In the low-light, plant individuals were shorter with a lower number of leaves than plants grown in high-light. Similar to the field study and common garden experiment, the high-light plants showed higher plant biomass than plants grown in low-light. Specific leaf area was three times higher for low-light than high-light. The high-light plants exhibited significantly higher CCI (4.68), RGRb (66.80), and NAR (3.92) than the low-light plants (Table 1).

Key selected life-history traits’ performance of Hyptis suaveolens, viz., maximum germination percentage (MGP), root mass ratio (RMR), plant biomass, and aboveground-belowground biomass ratio (ABBR) under two different light treatments in growth chamber experiment (LL – low-light, 300 µmol m–2 s−1 and HL – high-light, 940 µmol m–2 s−1) indicating differential resource allocation to the aboveground-belowground plant systems. Histogram affixed with different letters were significantly different from each other in row at p < 0.05
Further, the analysis suggested 1.5 and 5 times higher SMR and ABBR for high-light plants than the low-light plants, respectively. Interestingly, plants grown in the low-light condition showed four times higher RMR than the plants grown in the high-light condition (Fig. 6). It suggests that plants grown in high-light conditions performed better in terms of aboveground biomass accumulation, whereas plants grown in low-light conditions exhibited better performance in terms of belowground biomass accumulation. Thus, high-light condition supports significant proliferation of H. suaveolens, and low-light individuals survive by allocating plant biomass to the roots. The results of the plant growth chamber experiment further strengthen our confidence in the results of the common garden experiment in terms of seed germination and subsequent seedling performance.
Relations between and among studied plant traits of field study, common garden experiment, and growth chamber experiment
The collation of the three-tier study (field study, common garden experiment, and plant growth chamber experiment) related to various life-history traits of H. suaveolens is illustrated in Fig. 7. Analyses of germinative life-history traits showed an inclination of H. suaveolens populations towards successful establishment and formation of monospecific thickets under high-light conditions due to rapid seed germination (r2 = 0.53, p < 0.001) and seedling recruitment (r2 = 0.64, p < 0.001).

Relationships between and among stand-level, seed-level, and plant-level traits of Hyptis suaveolens under sun (high-light) and shade (low-light) conditions. These relations are shown in black and blue lines for high-light and low-light conditions, respectively. Solid and dotted lines represent significant and non-significant correlations, respectively. Thick and thin lines signify positive and negative correlations, respectively. Values in parenthesis represent the plasticity index (PI) of corresponding traits in response to variable light conditions (l, m, and h denote low, medium, and high PI value ranges, respectively) (see methods section for details)
Based on vegetative, eco-physiological, and reproductive life-history traits, under high-light conditions, plant height (r2 = 0.96, p < 0.001) and leaf area (r2 = 0.62, p < 0.001) were negatively related to available light, whereas number of leaves (r2 = 0.92, p < 0.001) and plant biomass (r2 = 0.61, p < 0.001) showed significantly positive relations with available light intensity. Under the high-light condition, SLA (r2 = 0.57, p < 0.001) was negatively related to light availability. On the other hand, CCI (r2 = 0.58, p < 0.001), photosynthetic rate (r2 = 0.65, p < 0.001), transpiration rate (r2 = 0.51, p < 0.001), RGRb (r2 = 0.87, p < 0.001), and NAR (r2 = 0.89, p < 0.001) were positively and significantly related to the available light. It can be noted that under low-light conditions, the eco-physiological traits of H. suaveolens, except SLA (r2 = 0.42, p < 0.001), did not respond well to the light intensity, thus, exhibiting suboptimal performance during the shade condition. Further, in high-light, the number of seeds per plant (r2 = 0.82, p < 0.001) exhibited a significantly positive relation to available light, while seed size (r2 = 0.84, p < 0.001) and seed mass (r2 = 0.71, p < 0.001) were negatively related to available light. It indicates that high-light availability provides a substantial competitive advantage to H. suaveolens and facilitates its wide-range dispersion. Additionally, larger seeds accumulate more nutritional reserves for plant use during the initial development of the seedling.
Inputs and outputs (number of propagules and recruited plant individuals, respectively) for each phase of life history traits are quantified based on the field study and experimental data and presented in Fig. 8. Results indicated that in the propagule phase, bulk reproductive output potentially drives the invasion success (species establishment and proliferation) of H. suaveolens (Fig. 8). Plant individuals in germinative and reproductive phases give rise to established populations for growth and expansion. In the fecundity phase, the H. suaveolens individuals provide propagules for bulk reproductive output (perennial rootstocks + soil seed bank + seeds from mature plant individuals), which translates into the field abundance of H. suaveolens populations.

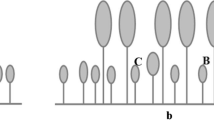
Illustration depicting different life-history stages and reproductive output of the Hyptis suaveolens individuals (non-overlapping generations) in response to sun (high-light) and shade (low-light) conditions. H. suaveolens’s life cycle is divided into four stages: Propagule phase (PP), including dormant seed banks (SB) and perennial rootstocks (PR); Germination phase (GP), which denotes the number of seed germination events in the field; Recruitment phase (RP), corresponds to established plant individuals; and Fecundity phase (FP), signifies the period of fruiting and total seed number (reproductive output). FA field abundance of H. suaveolens populations; d dormancy; f1, f2 and f3 fractions of bulk reproductive output corresponds to RP and FP; S sun and SH shade. Values in bold (sun) and non-bold (shade) denote the numbers of plant individuals corresponding to mentioned life cycle stages
Discussion
The PCA ordination yielded two distinct groups based on the importance value indices (IVI) of component herbaceous species, and the light conditions of the study area explained these groups. The most striking result from the present study is that the native plant species abundance, richness, and diversity of ground vegetation are low in open areas characterised by high-light availability and dominance of H. suaveolens. Thus, the light-modulated dominance of H. suaveolens might have changed the herbaceous species composition of the Vindhyan highlands. These observations are consistent with numerous other studies that have reported the negative impacts of invasive species abundance on native species richness in forest and grassland ecosystems (Pyšek and Pyšek 1995; Morris et al. 2002; Levine et al. 2003; Hulme and Bremner 2006; Stinson et al. 2007; Sharma et al. 2009; Sharma and Raghubanshi 2009; Sharma et al. 2017; Afreen et al. 2018).
Spatial heterogeneity, management practices, and environmental factors such as land disturbances (Minor et al. 2009; Vilà et al. 2011; Kumar et al. 2019), species composition (Stohlgren et al. 1998), soil properties (Fukano et al. 2013), and abiotic conditions (Popp and Kalwij 2021) play a significant role in the establishment and spread of invasive species. Studies have reported that light availability acts as a significant determinant of plant growth, reproduction (Godoy et al. 2011; Gratani et al. 2014; Goyal et al. 2018), plant abundance, and diversity (Stevens and Carson 1999; Onoda et al. 2014; Storch et al. 2018). The results of common garden experiment revealed enhanced germination and recruitment of H. suaveolens individuals under the sun than in shade conditions. The findings correspond to the common hypothesis of invasion biology that in the early life-history stage, invasive plants show enhanced seedling recruitment and survival that promote invasion success in introduced ranges (Rejmánek and Richardson 1996; Colautti et al. 2006; Pyšek and Richardson 2007). The enhanced performance of H. suaveolens individuals to light availability is suggested by taller plant individuals with an almost four times number of leaves, greater plant biomass, and higher SLA in high-light than in low-light conditions.
H. suaveolens exhibited opportunistic resource acquisition potential through a higher leaf number and a greater mass-based light-saturated photosynthetic rate, as suggested by higher values of CCI, A, RGRb, and NAR in high-light conditions. Since plant individuals depend on transpiration water loss to maintain homeostasis in high-light conditions (Ducrey 1992; James and Bell 2000), high values of stomatal conductance and transpiration rate in the sunlit area further characterized the fitness and adaptability of H. suaveolens to a sufficiently higher level of light intensity. Enhanced performance of vegetative and reproductive traits, which characterized species fitness in response to light availability, have been suggested to be a vital attribute of an invader (Daehler 2003; Funk 2008; Molina-Montenegro et al. 2012). H. suaveolens showed a greater plasticity index related to leaf production, biomass generation, and seed production under the sunlit condition than under the shade condition. These traits facilitate the survival and fitness of invasive plant species under high-light conditions (Reich et al. 2003; Rozendaal et al. 2006; Valladares and Niinemets 2008; van Kleunen et al. 2011; Feng and van Kleunen 2014; Liu et al. 2016). Robakowski et al. (2018) showed that the superior plastic responses of a species to available light facilitate the competitive advantage of aggressive colonizers over other co-occurring plant species in a wide range of environments.
The plant growth chamber experiment results revealed high MGP and MRP, increased plant height, and high plant biomass under high-light conditions, which were similar to the findings of common garden experiments. Plants grown in high-light conditions had greater RGRb than those in low-light conditions, possibly achieved by low SLA and higher NAR. As expected, an opposite trend was observed in plants grown in low-light conditions. Similar to the present study, other studies also reported that plants respond to light heterogeneity by changing growth patterns and/or modulation of biomass partitioning to aboveground and belowground plant parts (Poorter and Nagel 2000; Dasti et al. 2002; Freschet et al. 2018; Cleland et al. 2019; Umaña et al. 2021). Plants grown in low-light allocate substantial carbon to their root biomass, thus exhibiting higher RMR in response to low-light availability. This pattern reflected the survival strategy of H. suaveolens during stress conditions if shade acted as a stress to the H. suaveolens individuals. Correspondingly, H. suaveolens individuals showed high SMR and ABBR in high-light conditions than in low-light conditions. This biomass partitioning pattern reflects a trade-off between stem growth and allocation of biomass to roots.
Invasive plant species tend to exhibit enhanced trait response owing to differential energy allocation to their resource acquisition structures (Fenollosa et al. 2017). Opportunistic light energy acquisition and differential resource allocation to the aboveground-belowground plant systems probably facilitated the enhanced trait responses of H. suaveolens to high-light environments (Freschet et al. 2018; Cleland et al. 2019). Thus, rapid modulations of resource acquisition structures through alteration at the bioenergetics level showed the proclivity of H. suaveolens towards ‘Oskar Syndrome’. The ‘Oskar Syndrome’ reflects the phenomenon of resource (e.g. available light) acquisition and allocation by a plant species, facilitating a dynamic increase in biomass with exposure to high-light conditions (Silvertown 1984; Closset-Kopp et al. 2011). Probably, H. suaveolens individuals characterized by quick biomass accumulation along with the production of a greater number of small-sized and lightweight seeds, and higher interspecific competition from its large biomass may have caused low native plant diversity in high-light conditions (Raizada 2006; Schwarzkopf et al. 2009). This strategy enables H. suaveolens to quickly dominate the available space for the area, biomass, and reproductive fitness. Correspondingly, the findings of common garden and plant growth chamber experiments indicate that light availability is critical for seedling survival, plant growth, and reproduction of H. suaveolens in forest edges and clearing of Vindhyan highlands.
Based on the results of the present study, interspecific light competition may have acted as a driver for the invasion success of H. suaveolens and subsequent loss of species diversity, owing to its enhanced light capture and opportunistic resource utilization ability, and rapid biomass accumulation. Further, high fecundity per plant coupled with smaller and lightweight seeds produced by H. suaveolens individuals confer the superiority of its reproductive output in sunlit habitats and, thereby, its dominance. High reproductive output also promotes the chances for a species’ propagules to acquire a favourable environment for subsequent establishment and reproduction (Lockwood et al. 2005; Martínez-Ghersa and Ghersa 2006; Simberloff 2009; Wilson et al. 2009; Cassey et al. 2018; Alzate et al. 2020). Thus, opportunistic resource acquisition-allocation induced high bulk reproductive output of H. suaveolens in heavily infested areas facilitated its dominance and invasion pressure in locally disturbed habitats.
Disturbances in fragmented forest edges and openings in forest interiors cause light heterogeneity, which leads to shifting of the species composition (Broadbent et al. 2008; Legner et al. 2014), owing to differential responses of species traits’ to light availability (Godoy et al. 2011; Carrión-Tacuri et al. 2013). The present study suggests that the H. suaveolens individuals rapidly proliferate, generate high biomass, produce a significantly high number of seeds, and dominate in high-light environments. In low-light environments, H. suaveolens maintains a self-sustaining population of plant individuals showing low performance and probably acting as ‘Oskar individuals’ (See Closset-Kopp et al. 2007). In the event of anthropogenic disturbances, increased light availability causes shade-dwelling ‘Oskar individuals’ to outperform the co-occurring native plant species through high resource acquisition in a short span of time. The modus operandi suggests that H. suaveolens individuals follow a ‘sit and wait’ strategy. The ‘sit and wait’ strategy may also explain the invasion success of other invasive plant species such as Ailanthus altissima, Acer platanoides, Quercus rubra, and Robinia pseudoacacia (Knapp and Canham 2000; Sanford et al. 2003; Lee et al. 2004). Likewise, Fridley et al. (2023) have designated the term ‘superinvaders’ to such forest invaders that rapidly grow than other co-occurring native plant species in high-light conditions due to enhanced resource acquisition and allocation while maintaining the stable population in the shade conditions. Owing to this peculiar strategy, H. suaveolens probably altered the available light and space in its vicinity, leading to the decline of native species.
Conclusion
The present field, common garden, and growth chamber studies provide compelling evidence that H. suaveolens abundance causes competitive exclusion of co-occurring native species in open forest areas with high-light conditions because it rapidly proliferates, generates high biomass, produces a significantly high number of seeds, and dominates in such environments. The current study provides insights on prioritizing areas/habitats infested by H. suaveolens to manage the declining native species. The study suggests that the forest habitats with open disturbed areas and subsequent light heterogeneity show a high invasion risk and should be given the highest priority for the management of invasive population. The insights gained from the study will aid in reducing the risk of biological invasions on the native species diversity in tropical regions, which is crucial for human wellbeing. Nevertheless, the study warrants multi-species common garden experiments to further reveal the mechanistic processes of competitive exclusion led by H. suaveolens.
Data Availability
The datasets generated during and/or analysed during the current study are available from the corresponding author upon reasonable request.
References
Adkins S, Shabbir A (2014) Biology, ecology and management of the invasive parthenium weed (Parthenium hysterophorus L.). Pest Manag Sci 70(7):1023–1029
Afreen T, Srivastava P, Singh H, Singh JS (2018) Effect of invasion by Hyptis suaveolens on plant diversity and selected soil properties of a constructed tropical grassland. J Plant Ecol 11(5):751–760
Afreen T, Singh V, Yadav VK, Singh RP, Singh H (2020) Impact of rainfall variability on the ecophysiology of Hyptis suaveolens: a study in the constructed tropical grassland. Environ Monit Assess 192:1–11
Agrawal M (2002) Atmospheric deposition and budgeting of the nutrients and trace elements around industrial sites at Singrauli region (Report, No. 19/93). Ministry of Environment and Forest, India
Alzate A, Onstein RE, Etienne RS, Bonte D (2020) The role of preadaptation, propagule pressure and competition in the colonization of new habitats. Oikos 129(6):820–829
Banerjee AK, Ghosh S, Dewanji A (2017) Do plants have a choice of traits to be modulated? Evidence from an invasive plant Mikania micrantha Kunth in different urban environments. Am J Plant Sci 8(4):835–855
Barthod S, Epron D (2005) Variations of construction cost associated to leaf area renewal in saplings of two co-occurring temperate tree species (Acer platanoides L. and Fraxinus excelsior L.) along a light gradient. Ann For Sci 62(6):545–551
Bazzaz FA, Carlson RW (1982) Photosynthetic acclimation to variability in the light environment of early and late successional plants. Oecologia 54(3):313–316
Bellard C, Marino C, Courchamp F (2022) Ranking threats to biodiversity and why it doesn’t matter. Nat Commun 13(1):2616
Broadbent EN, Asner GP, Keller M, Knapp DE, Oliveira PJ, Silva JN (2008) Forest fragmentation and edge effects from deforestation and selective logging in the Brazilian Amazon. Biol Conserv 141(7):1745–1757
Carrión-Tacuri J, Rubio-Casal AE, de Cires A, Figueroa ME, Castillo JM (2013) Effect of low and high temperatures on the photosynthetic performance of Lantana camara L. leaves in darkness. Russ J Plant Physiol 60(3):322–329
Cassey P, Delean S, Lockwood JL, Sadowski JS, Blackburn TM (2018) Dissecting the null model for biological invasions: a meta-analysis of the propagule pressure effect. PLoS Biol 16(4):e2005987
Champion HG, Seth SK (1968) A revised survey of the forest types of India. Manager of Publications
Chazdon RL, Pearcy RW, Lee DW, Fetcher N (1996) Photosynthetic responses of tropical forest plants to contrasting light environments. In: Mulkey SS, Chazdon RL, Smith AP (eds) Tropical forest plant ecophysiology. Chapman Hall, New York, pp 5–55
Chrobock T, Kempel A, Fischer M, van Kleunen M (2011) Introduction bias: cultivated alien plant species germinate faster and more abundantly than native species in Switzerland. Basic Appl Ecol 12(3):244–250
Cleland EE, Lind EM, DeCrappeo NM, DeLorenze E, Wilkins RA, Adler PB, Seabloom EW et al (2019) Belowground biomass response to nutrient enrichment depends on light limitation across globally distributed grasslands. Ecosystems 22:1466–1477
Closset-Kopp D, Chabrerie O, Valentin B, Delachapelle H, Decocq G (2007) When Oskar meets Alice: does a lack of trade-off in r/K-strategies make Prunus serotina a successful invader of European forests? For Ecol Manag 247(1–3):120–130
Closset-Kopp D, Saguez R, Decocq G (2011) Differential growth patterns and fitness may explain contrasted performances of the invasive Prunus serotina in its exotic range. Biol Invasions 13:1341–1355
Colautti RI, Grigorovich IA, MacIsaac HJ (2006) Propagule pressure: a null model for biological invasions. Biol Invasions 8:1023–1037
Curtis JT (1959) The vegetation of Wisconsin: an ordination of plant communities. University of Wisconsin
Daehler CC (2003) Performance comparisons of co-occurring native and alien invasive plants: implications for conservation and restoration. Annu Rev Ecol Syst 34(1):183–211
Dasti AA, Fatima K, Malik SA (2002) How photoperiod affects growth rate and biomass allocation pattern: a comparative study on three genotypes of Arabidopsis thaliana? Asian J Plant Sci 1:176–179
de Villemereuil P, Gaggiotti O, Mouterde M, Till-Bottraud I (2016) Common garden experiments in the genomic era: new perspectives and opportunities. Heredity 116(3):249–254
Ducrey M (1992) Variation in leaf morphology and branching pattern of some tropical rain forest species from Guadeloupe (French West Indies) under semi-controlled conditions. Ann Sci For 49(6):553–570
Engelkes T, Morriën E, Verhoeven K (2008) Successful range-expanding plants experience less above-ground and below-ground enemy impact. Nature 456(7224):946–948
Farrugia P, Petrisor BA, Farrokhyar F, Bhandari M (2010) Research questions, hypotheses and objectives. Can J Surg 53(4):278–281
Feng Y, van Kleunen M (2014) Responses to shading of naturalized and non-naturalized exotic woody species. Ann Bot 114(5):981–989
Fenollosa E, Munné-Bosch S, Pintó-Marijuan M (2017) Contrasting phenotypic plasticity in the photoprotective strategies of the invasive species Carpobrotus edulis and the coexisting native species Crithmum maritimum. Physiol Plant 160(2):185–200
Flores-Moreno H, Moles AT (2013) A comparison of the recruitment success of introduced and native species under natural conditions. PLoS ONE 8(8):e72509
Freschet GT, Violle C, Bourget MY, Scherer-Lorenzen M, Fort F (2018) Allocation, morphology, physiology, architecture: the multiple facets of plant above- and below-ground responses to resource stress. New Phytol 219(4):1338–1352
Fridley JD (2012) Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 485(7398):359–362
Fridley JD, Bellingham PJ, Closset-Kopp D, Daehler CC, Dechoum MS, Martin PH, Murphy HT, Rojas‐Sandoval J, Tng D (2023) A general hypothesis of forest invasions by woody plants based on whole‐plant carbon economics. J Ecol 111(1):4–22
Fukano Y, Tachiki Y, Yahara T, Iwasa Y (2013) Soil disturbances can suppress the invasion of alien plants under plant–soil feedback. Ecol Model 260:42–49
Funk JL (2008) Differences in plasticity between invasive and native plants from a low resource environment. J Ecol 96(6):1162–1173
Funk JL, Standish RJ, Stock WD, Valladares F (2016) Plant functional traits of dominant native and invasive species in Mediterranean climate ecosystems. Ecology 97(1):75–83
Godoy O, Saldana A, Fuentes N, Valladares F, Gianoli E (2011) Forests are not immune to plant invasions: phenotypic plasticity and local adaptation allow Prunella vulgaris to colonize a temperate evergreen rainforest. Biol Invasions 13:1615–1625
Goparaju L, Sinha D (2015) Forest cover change analysis of dry tropical forests of Vindhyan highlands in Mirzapur district, Uttar Pradesh using satellite remote sensing and GIS. Ecol Quest 22:23–37
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4(4):379–391
Goyal N, Sharma GP (2019) It takes two to tango: variable architectural strategies boost invasive success of Lantana camara L. (Sensu Lato) in contrasting light environments. Biol Invasions 21:163–174
Goyal N, Shah K, Sharma GP (2018) Does intrinsic light heterogeneity in Ricinus communis L. monospecific thickets drive species’ population dynamics? Environ Monit Assess 190:1–12
Gratani L, Crescente MF, D’amato V, Ricotta C, Frattaroli AR, Puglielli G (2014) Leaf traits variation in Sesleria nitida growing at different altitudes in the Central Apennines. Photosynthetica 52(3):386–396
Grotkopp E, Rejmánek M (2007) High seedling relative growth rate and specific leaf area are traits of invasive species: phylogenetically independent contrasts of woody angiosperms. Am J Bot 94(4):526–532
Hameed M, Ashraf M, Naz N (2009) Anatomical adaptations to salinity in cogon grass [Imperata cylindrica (L.) Raeuschel] from the Salt Range, Pakistan. Plant Soil 322:229–238
Harper JL (1977) Population Biology of plants. Academic Press, London
Heberling JM, Fridley JD (2013) Resource-use strategies of native and invasive plants in Eastern North American forests. New Phytol 200(2):523–533
Hiremath AJ, Sundaram B (2005) The fire-lantana cycle hypothesis in Indian forests. Conserv Soc 3(1):26–42
Holt JS (1995) Plant responses to light: a potential tool for weed management. Weed Sci 43(3):474–482
Honig MA, Cowling RM, Richardson DM (1992) The invasive potential of Australian banksias in South African fynbos: a comparison of the reproductive potential of Banksia ericifolia and Leucadendron laureolum. Aust J Ecol 17(3):305–314
Hsieh TC, Ma KH, Chao A (2016) iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol Evol 7(12):1451–1456
Hui C, Pyšek P, Richardson DM (2023) Disentangling the relationships among abundance, invasiveness and invasibility in trait space. npj Biodiversity 2:13
Hulme PE, Bremner ET (2006) Assessing the impact of Impatiens glandulifera on riparian habitats: partitioning diversity components following species removal. J Appl Ecol 43(1):43–50
James SA, Bell DT (2000) Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances. Tree Physiol 20(15):1007–1018
Jha CS, Singh JS (1990) Composition and dynamics of dry tropical forest in relation to soil texture. J Veg Sci 1(5):609–614
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17(4):164–170
Kerns BK, Day MA (2017) The importance of disturbance by fire and other abiotic and biotic factors in driving cheatgrass invasion varies based on invasion stage. Biol Invasions 19:1853–1862
Klink CA (1996) Germination and seedling establishment of two native and one invading African grass species in the Brazilian cerrado. J Trop Ecol 12(1):139–147
Knapp LB, Canham CD (2000) Invasion of an old-growth forest in New York by Ailanthus altissima sapling growth and recruitment in canopy gaps. J Torrey Bot Soc 127(4):307–315
Kramer PJ, Kozlowski TT (1979) Physiology of Woody Plants. Academic Press, New York, p 811
Kueffer C, Pyšek P, Richardson DM (2013) Integrative invasion science: model systems, multi-site studies, focused meta-analysis and invasion syndromes. New Phytol 200(3):615–633
Kumar M, Padalia H, Nandy S, Singh H, Khaiter P, Kalra N (2019) Does spatial heterogeneity of landscape explain the process of plant invasion? A case study of Hyptis suaveolens from Indian Western Himalaya. Environ Monit Assess 191:1–17
Kushwaha CP, Singh KP (2005) Diversity of leaf phenology in a tropical deciduous forest in India. J Trop Ecol 21(1):47–56
Kushwaha CP, Tripathi SK, Singh GS, Singh KP (2010) Diversity of deciduousness and phenological traits of key Indian dry tropical forest trees. Ann For Sci 67(3):310
Kushwaha CP, Tripathi SK, Singh KP (2011) Tree specific traits affect flowering time in Indian dry tropical forest. Plant Ecol 212:985–998
Leary J, Mahnken B, Wada C, Burnett K (2018) Interpreting life-history traits of miconia (Miconia calvescens) through management over space and time in the East Maui Watershed, Hawaii (USA). Invasive Plant Sci Manag 11(4):191–200
Lee C-S, Cho H-J, Yi H (2004) Stand dynamics of introduced black locust (Robinia pseudoacacia L.) plantation under different disturbance regimes in Korea. For Ecol Manage 189(1–3):281–293
Legner N, Fleck S, Leuschner C (2014) Within-canopy variation in photosynthetic capacity, SLA and foliar N in temperate broad-leaved trees with contrasting shade tolerance. Trees 28:263–280
Leishman MR, Wright IJ, Moles AT, Westoby M (2000) The evolutionary ecology of seed size. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities. CABI Publishing, Wallingford, UK, pp 31–57
Leishman MR, Haslehurst T, Ares A, Baruch Z (2007) Leaf trait relationships of native and invasive plants: community and global scale comparisons. New Phytol 176(3):635–643
Lenzner B, Latombe G, Capinha C, Bellard C, Courchamp F, Diagne C, Essl F (2020) What will the future bring for biological invasions on islands? An expert-based assessment. Front Ecol Evol 8:280
Levine JM, Vila M, Antonio CMD, Dukes JS, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Proc Royal Soc B 270(1517):775–781
Li YP, Feng YL, Li WT, Tomlinson K, Liao ZY, Zheng YL, Zhang JL (2022) Leaf trait association in relation to herbivore defense, drought resistance, and economics in a tropical invasive plant. Am J Bot 109(6):910–921
Liu G, Huang QQ, Lin ZG, Huang FF, Liao HX, Peng SL (2012) High tolerance to salinity and herbivory stresses may explain the expansion of Ipomoea cairica to salt marshes. PLoS ONE 7(11):e48829
Liu Y, Wayne D, Daniel P, Emily H, Yanhao F, van Kleunen M (2016) Does greater specific leaf area plasticity help plants to maintain a high performance when shaded? Ann Bot 118(7):1329–1336
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20(5):223–228
Martínez-Ghersa MA, Ghersa CM (2006) The relationship of propagule pressure to invasion potential in plants. Euphytica 148:87–96
Martins VF, Haddad CR, Semir J (2011) Responses of the invasive Ricinus communis seedlings to competition and light. N Z J Bot 49(2):263–279
McAleece N (1997) Biodiversity Pro. http://www.sams.ac.uk/research/software/research/software/bdpro.
McKenna MF, Shipley B (1999) Interacting determinants of interspecific relative growth: empirical patterns and a theoretical explanation. Ecoscience 6(2):286–296
Meyer SE, Callaham MA, Stewart JE, Warren SD (2021) Invasive species response to natural and anthropogenic disturbance. In: Poland TM, Patel-Weynand T, Finch DM, Miniat CF, Hayes DC, Lopez VM (eds) Invasive species in forests and rangelands of the United States: a comprehensive science synthesis for the United States forest sector. Springer Nature, pp 85–110
Minor ES, Tessel SM, Engelhardt KAM, Lookingbill TR (2009) The role of landscape connectivity in assembling exotic plant communities: a network analysis. Ecology 90(7):1802–1809
Molina-Montenegro MA, Penuelas J, Munné-Bosch S, Sardans J (2012) Higher plasticity in ecophysiological traits enhances the performance and invasion success of Taraxacum officinale (dandelion) in alpine environments. Biol Invasions 14:21–33
Morris LL, Walck JL, Hidayati SN (2002) Growth and reproduction of the invasive Ligustrum sinense and native Forestiera ligustrina (Oleaceae): implications for the invasion and persistence of a non-native shrub. Int J Plant Sci 163(6):1001–1010
Mudgal V, Khanna KK, Hazra PK (1997) Flora of Madhya Pradesh II. BSI 403–404
Nabe-Nielsen J (2001) Diversity and distribution of lianas in a neotropical rain forest, Yasuní National Park, Ecuador. J Trop Ecol 17(1):1–19
Onoda Y, Saluñga JB, Akutsu K, Aiba SI, Yahara T, Anten NP (2014) Trade-off between light interception efficiency and light use efficiency: implications for species coexistence in one‐sided light competition. J Ecol 102(1):167–175
Padalia H, Srivastava V, Kushwaha SPS (2014) Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: comparison of MaxEnt and GARP. Ecol Inform 22(1):36–43
Pan XY, Geng YP, Sosa A, Zhnag WJ, Li B, Chen JK (2007) Invasive Alternanthera philoxeroides: biology, ecology and management. Acta Phytotaxonomica Sinica 45(6):884–900
Pant DN, Das KK, Roy PS (1992) Mapping of tropical dry deciduous forest and land use in part of Vindhyan range using satellite remote sensing. J Indian Soc Remote Sens 20:9–20
Peng Y, Yang Y (2016) Allometric biomass partitioning under nitrogen enrichment: evidence from manipulative experiments around the world. Sci Rep 6:28918
Poorter L (1999) Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits. Funct Ecol 13:396–410
Poorter H, Nagel O (2000) The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Funct Plant Biol 27(12):1191–1191
Popp MR, Kalwij JM (2021) Abiotic conditions shape the relationship between indigenous and exotic species richness in a montane biodiversity hotspot. Plant Ecol 222(4):421–432
Pyšek P, Pyšek A (1995) Invasion by Heracleum mantegazzianum in different habitats in the Czech Republic. J Veg Sci 6(5):711–718
Pyšek P, Richardson DM (2007) Traits associated with invasiveness in alien plants: where do we stand? In: Nentwig W (ed) Biological invasions, Ecological Studies Vol 193. Springer, Berlin, pp 97–125
Raghubanshi AS, Tripathi A (2009) Effect of disturbance, habitat fragmentation and alien invasive plants on floral diversity in dry tropical forests of Vindhyan Highland: a review. Trop Ecol 50(1):57–69
Raizada P (2006) Ecological and vegetative characteristics of a potent invader, Hyptis suaveolens Poit. from India. Lyonia 11(2):115–120
Raunkiær C (1934) The life forms of plants and statistical plant geography. Clarendon Press, Oxford
Reich PB, Wright IJ, Cavender-Bares J, Craine JM, Oleksyn J, Westoby M, Walters MB (2003) The evolution of plant functional variation: traits, spectra, and strategies. Int J Plant Sci 164(S3):S143–S164
Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77(6):1655–1661
Robakowski P, Bielinis E, Sendall K (2018) Light energy partitioning, photosynthetic efficiency and biomass allocation in invasive Prunus serotina and native Quercus petraea in relation to light environment, competition and allelopathy. J Plant Res 131:505–523
Roy PS, Dutt CBS, Joshi PK (2002) Tropical forest assessment and management. Trop Ecol 43(1):21–38
Rozendaal DMA, Hurtado VH, Poorter L (2006) Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Funct Ecol 20(2):207–216
Sagar R, Singh JS (1999) Species diversity and its measurement. Botnica 49:9–16
Sagar R, Singh JS (2003) Predominant phenotypic traits of disturbed tropical dry deciduous forest vegetation in northern India. Community Ecol 4(1):63–71
Sagar R, Singh JS (2006) Tree density, basal area and species diversity in a disturbed dry tropical forest of northern India: implications for conservation. Environ Conserv 33(3):256–262
Sagar R, Raghubanshi AS, Singh JS (2003) Tree species composition, dispersion and diversity along a disturbance gradient in a dry tropical forest region of India. For Ecol Manag 186(1–3):61–71
Sanford NL, Harrington RA, Fownes JH (2003) Survival and growth of native and alien woody seedlings in open and understory environments. For Ecol Manage 183(1–3):377–385
Scheiner SM (2003) Six types of species-area curves. Glob Ecol Biogeogr 12(6):441–447
Schwarzkopf KT, Trevisan MC, Silva AJF (2009) A matrix model for the population dynamics of Hyptis suaveolens, an annual weed. Ecotropicos 22(1):23–36
Sharma GP (2007b) New reports of invasion in Vindhyan highlands: Hyptis suaveolens. Asia Pac Inv Sp 7:4–5
Sharma GP, Esler KJ (2008) Phenotypic plasticity among Echium plantagineum populations in different habitats of Western Cape, South Africa. S Afr J Bot 74(4):746–749
Sharma GP, Raghubanshi AS (2009) Plastic responses to different habitat type contribute to Hyptis suaveolens Poit. invasiveness in the dry deciduous forest of India. Ambio 38(6):342–344
Sharma GP (2007a) Habitat fragmentation induced plant species invasion in Vindhyan highlands. Ph.D. Thesis. Banaras Hindu University, Varanasi, India
Sharma GP, Raizada P, Raghubanshi AS (2009) Hyptis suaveolens: an emerging invader of Vindhyan plateau, India. Weed Biol Manag 9(3):185–191
Sharma GP, Muhl SA, Esler KJ, Milton SJ (2010) Competitive interactions between the alien invasive annual grass Avena fatua and indigenous herbaceous plants in South African renosterveld: the role of nitrogen enrichment. Biol Invasions 12:3371–3378
Sharma A, Batish DR, Singh HP, Jaryan V, Kohli RK (2017) The impact of invasive Hyptis suaveolens on the floristic composition of the periurban ecosystems of Chandigarh, North-western India. Flora 233:156–162
Shirke PA, Pathre UV, Sane PV (2018) Adaptation strategies of two leaf cohorts of Prosopis juliflora produced in spring and monsoon. Photosynthetica 56:468–477
Silvertown JW (1984) Introduction to Plant Population Ecology. Longman, London
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst 40:81–102
Singh JS, Raghubanshi AS, Singh RS, Srivastava SC (1989) Microbial biomass acts as a source of plant nutrients in dry tropical forests and savanna. Nature 338(6215):499–500
Singh JS, Roy PS, Murthy MSR, Jha CS (2010) Application of landscape ecology and remote sensing for assessment, monitoring and conservation of biodiversity. J Indian Soc Remote Sens 38:365–385
Singh V, Singh H, Sharma GP, Raghubanshi AS (2011) Eco-physiological performance of two invasive weed congeners (Ageratum conyzoides L. and Ageratum houstonianum Mill.) in the Indo-Gangetic plains of India. Environ Monit Assess 178:415–422
SPSS Inc. (2007) SPSS for Windows, Version 16.0 release. SPSS Inc., Chicago
Stevens MHH, Carson WP (1999) Plant density determines species richness along an experimental fertility gradient. Ecology 80(2):455–465
Stinson K, Kaufman S, Durbin L, Lowenstein F (2007) Impacts of garlic mustard invasion on a forest understory community. Northeast Nat 14(1):73–88
Stohlgren TJ, Bull KA, Otsuki Y, Villa CA, Lee M (1998) Riparian zones as havens for exotic plant species in the central grasslands. Plant Ecol 138(1):113–125
Storch D, Bohdalková E, Okie J (2018) The more-individuals hypothesis revisited: the role of community abundance in species richness regulation and the productivity-diversity relationship. Ecol Lett 21(6):920–937
Sun P, Mantri N, Moller M, Shen J, Shen Z, Jiang B, Chen C, Miao Q, Lu H (2012) Influence of light and salt on the growth of alien invasive tropical weed Ageratum conyzoides. Aust J Crop Sci 6(4):739–748
Tao Y, Chen F, Wan K, Li X, Li J (2009) The structural adaptation of aerial parts of invasive Alternanthera philoxeroides to water regime. J Plant Biol 52:403–410
Tripathi A (2003) Community Diversity Assessment in Vindhyans. Ph.D. Thesis. Banaras Hindu University, Varanasi, India
Umaña MN, Cao M, Lin L, Swenson NG, Zhang C (2021) Trade-offs in above- and below-ground biomass allocation influencing seedling growth in a tropical forest. J Ecol 109(3):1184–1193
Valladares F, Niinemets Ü (2008) Shade tolerance, a key plant feature of complex nature and consequences. Annu Rev Ecol Evol Syst 39:237–257
Valladares F, Arrieta S, Aranda I, Lorenzo D, Sánchez-Gómez D, Tena D, Suárez F, Pardos JA (2005) Shade tolerance, photoinhibition sensitivity and phenotypic plasticity of Ilex aquifolium in continental Mediterranean sites. Tree Physiol 25(8):1041–1052
van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13(2):235–245
van Kleunen M, Johnson SD (2007) South African Iridaceae with rapid and profuse seedling emergence are more likely to become naturalized in other regions. J Ecol 95:674–681.
van Kleunen M, Schlaepfer DR, Glaettli M, Fischer M (2011) Preadapted for invasiveness: do species traits or their plastic response to shading differ between invasive and non-invasive plant species in their native range? J Biogeogr 38(7):1294–1304
van Kleunen M, Dawson W, Essl F, Pergl J, Winter M, Weber E, Pyšek P et al (2015) Global exchange and accumulation of non-native plants. Nature 525(7567):100–103
Verma BK, Mishra BK (1992) Flora of Allahabad district. Uttar Pradesh, India
Verma DM, Pant PC, Hanf MI (1985) Flora of Raipur, Durg, and Rajnandgaon
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a metaanalysis of their effects on species, communities and ecosystems. Ecol Lett 14(7):702–708
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21(2–3):213–251
Wilson JR, Dormontt EE, Prentis PJ, Lowe AJ, Richardson DM (2009) Something in the way you move: dispersal pathways affect invasion success. Trends Ecol Evol 24(3):136–144
Ziller SR, de Sá Dechoum M, Dudeque Zenni R (2019) Predicting invasion risk of 16 species of eucalypts using a risk assessment protocol developed for Brazil. Austral Ecol 44(1):28–35
Acknowledgements
KS thanks University Grants Commission, India, for the Research Fellowship. Insights gained during the National Symposium on Biological Invasions, 2021 held at Centre for Biological Control (CBC), Eastern Cape, South Africa enriched ideation in the manuscript.
Funding
GPS acknowledges support from the Science and Engineering Research Board, Department of Science and Technology, India. RS acknowledges funding from the Science and Engineering Research Board, Department of Science and Technology, India (File No. SERB-EEQ /2021/000356) and IOE, Institute of Science, Banaras Hindu University.
Author information
Authors and Affiliations
Contributions
Conceptualisation: KS and GPS; data collection: KS; data analysis: KS, GPS and RS; writing-original draft preparation: KS, GPS and RS; writing-review and editing: KS, GPS and RS. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not Applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1:
Species abundance (IVI) in studied 10 field plots under the sun (high-light) and shade (low-light) environments in Vindhyan highlands, India (n = native and nn = non-native)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shah, K., Sharma, G.P. & Sagar, R. An arrow in the quiver: evaluating the performance of Hyptis suaveolens (L.) Poit. in different light levels. Ecol Process 13, 30 (2024). https://doi.org/10.1186/s13717-024-00504-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-024-00504-w