
Abstract
Background
Gastric cancer has a high incidence and mortality rate. Angiogenesis is necessary for tumor infiltration and metastasis and affects patient prognosis. YKL-39 has monocyte chemotactic activity and pro-angiogenic activity in some tumors. In this study, we investigated the relationship between YKL-39 and tumor-associated macrophages and microangiogenesis in gastric cancer to determine its potential as a prognostic biomarker.
Materials and methods
A total of 119 patients with gastric cancer who had undergone gastrectomy at the 940th Hospital of the Joint Security Force between 2014 and 2018 were included in this study. We assayed the protein expression of YKL-39, CD68, and CD34 by immunohistochemistry in tissues of 119 patients with gastric cancer, as well as the intracellular expression of YKL-39 and CD68 by immunofluorescence. Data were analyzed with SPSS Statistics 25.0 to explore the impact of expression of YKL-39, CD68, and CD34 in gastric cancer patients and the relationship among them.
Results
Our results show that YKL-39 was expressed in both the nucleus and cytoplasm of gastric cancer cells and tumor mesenchyme. YKL-39 protein expression was associated with the depth of tumor infiltration, lymph node metastasis, and TNM stage; CD68 protein expression was associated with lymph node metastasis and TNM stage; CD34 protein expression was not associated with clinicopathological characteristics. Expression of YKL-39 was positively correlated with CD68 and CD34 (p < 0.001), and high expression of YKL-39 was associated with poor prognosis (p < 0.05).
Conclusion
In gastric cancer, YKL-39 expression is positively correlated with the degree of tumor-associated macrophage infiltration and angiogenesis, and is a potential prognostic marker for gastric cancer.
Similar content being viewed by others
Introduction
Gastric cancer (GC) is one of the most common gastrointestinal malignancies, ranking fifth in incidence and fourth in lethality globally according to the latest global cancer statistics [1]. With advances in technology such as endoscopy, an increasing number of patients are being diagnosed with GC and treated in a timely manner, but the overall 5-year survival rate is still less than 40% [2]. Most GC patients are already in the middle or late stages when they are diagnosed, with a median overall survival (OS) of less than 12 months [3]. Angiogenesis supports the growth and metastasis of GC cells by providing nutrients and oxygen [4]; when the tumor has advanced metastasis or cannot be excised, the current treatment is based on palliative chemotherapy, and anti-angiogenic therapy can be used as an effective adjuvant treatment [5]. The commonly used anti-angiogenic drugs include bevacizumab, which targets vascular endothelial growth factor-A (VEGF-A), ramucirumab, which targets vascular endothelial growth factor receptor 2, and ziv-aflibercept, which targets VEGF-A isoforms, placental growth factor (PLGF), and vascular endothelial growth factor-B etc .[6]. However, tumors can develop resistance to anti-angiogenic drugs through a variety of mechanisms, including upregulation of alternative pro-angiogenic signaling pathways, resistance of tumor stromal cells to anti-angiogenic drugs, adaptation of tumor cells to hypoxic environments, and alternative mechanisms of tumor vascularization [7]. Therefore, it is of great significance to find new anti-angiogenic targets.
Tumor-associated macrophages (TAM) are the most common tumor-infiltrating immune cells in the tumor microenvironment (TME), accounting for more than 50% of immune cells in the TME and promoting tumorigenesis through various mechanisms such as stimulating angiogenesis, increasing tumor cell invasion and migration, and inhibiting anti-tumor immunity [8, 9]. Macrophages are stimulated by different chemokines released by tumors and stromal cells to differentiate into two phenotypes with dramatic differences: M1 macrophages with antitumor effects and M2 macrophages with pro-tumor effects [8]. M2 macrophages, which occupy the majority of TAM, can produce a variety of pro-angiogenic factors such as VEGF-A and tumor necrosis factor α (TNFα) in hypoxic areas to maintain tumor growth [10]. TAM infiltration in multiple tumors is positively correlated with angiogenesis [11, 12], and it has been shown that the emergence of anti-VEGF therapy resistance is associated with the aggregation of TAMs in TME [13, 14].
Chitinase 3-like protein 2(CHI3L2), also known as YKL-39, belongs to the family of chitinase-like proteins (CLPs) that function as both cytokines and growth factors [15]. Human CLPs include chitinase 3-like protein 1 (YKL-40), YKL-39 and stabilin-1 interacting chitinase-like protein (SI-CLP). YKL-39 was originally found in human synoviocytes and chondrocytes and plays a role in regulating autoimmunity and participating in tissue remodeling [16]. It has been shown that YKL-39 expression is elevated in degenerative pathologies and diseases characterized by tissue remodeling, such as osteoarthritis, multiple sclerosis, Alzheimer’s disease, and amyotrophic lateral sclerosis [17,18,19]. Recent studies have reported that YKL-39 has monocyte chemotactic and pro-angiogenic activity and is expressed in M2 macrophages from breast, glioma, and kidney cancers, and affects tumor angiogenesis; overexpressed YKL-39 is associated with poor prognosis [20,21,22]. However, there are no reports on the relationship between YKL-39 expression and GC biological behavior and the prognosis of GC patients.
CD68 is a highly glycosylated transmembrane protein associated with lysosomal particles and is currently the most widely used marker for immunohistochemical recognition of macrophages [23]. CD68 is a major biomarker for detecting total TAMs in a variety of malignant cancers including breast, colorectal, lung, ovarian, and lymphoma [24]. The commonly used vascular markers are CD31, CD34, and factor VIII, among which CD31 mainly marks neovascularization and CD34 marks all vascular endothelium including neovascularization and mature vessels [25].
In this study, we used immunohistochemistry (IHC) and immunofluorescence (IF) to detect the expression of YKL-39, CD68, and CD34 in GC tissues to determine the relationship between YKL-39 expression and macrophage infiltration and angiogenesis.
Material and methods
Patients and tissue samples
This study included 119 patients diagnosed with GC through histopathologic evaluation on gastroscopic biopsy or surgical tissue specimens; 101 cases were male and 18 cases were female, ranging in age from 29 to 81 years, with a median age of 59.03 years. All patients underwent gastrectomy between 2014 and 2018 at the 940th Hospital of the Joint Security Force, Gansu, China. Patients who had preoperative treatment, such as radiotherapy, chemotherapy, or other medical interventions and those diagnosed with autoimmune diseases were excluded from the study. Formalin-fixed, paraffin-embedded (FFPE) GC tissue samples were obtained from these patients after surgery.
Data collection and follow-up
Demographic and clinicopathologic characteristics, including age, sex, TNM stage, histologic grade, and lymph node metastases were assessed. The TNM classification was based on the 8th edition of AJCC GC staging. The patients were followed up for 5 years by inpatient, outpatient and telephone contact. In this study, we used OS to evaluate prognosis. OS was defined as the period from the date of surgery to the date of death or last visit. The median follow-up time was 44.18 months (range 3–114 months). Informed consent was obtained from all patients.
Tissue microarray (TMA) construction
FFPE specimens were stained with H&E and examined by experienced pathologists. At least three representative regions were selected and marked on H&E slices each tumor. The corresponding part on the wax block was selected as the material selection site according to the mark on the H&E slice. TMAs containing the 119 FFPE specimens were constructed by using a Manual Tissue Arrayer (TM1, boyikang, Bejing, China). All three marked regions on each tissue with a diameter of 2.0 mm were removed using a TMA instrument and placed into the acceptor wax block from top to bottom in a right-to-left order.
IHC
All TMA slides with a thickness of 4 μm were deparaffinized using xylene and rehydrated using graded ethanol. Then, the slides were immersed and boiled in ethylene diamine tetraacetic acid (pH 9.0) for 30 min in a pressure cooker for antigen retrieval. Endogenous peroxidase was inhibited by 3% H2O2 for 15 min. For YKL-39 staining, recombinant monoclonal rabbit anti-human YKL-39 (1:150; ThermoFisher, USA) was applied. For TAM evaluation, monoclonal mouse anti-human CD68 (ready-to-use; maixin, China) was applied. For micro vessel density (MVD) evaluation, monoclonal mouse anti-human CD34 (ready-to-use, maixin, China) was used. Specimens were incubated with all antibodies overnight at 4 °C. After washing with PBS, the slides were incubated with secondary antibody at 37 °C for 10 min. The following steps were performed: color development, counterstaining, differentiation, dehydration, and transparency. Finally, the slides fixed with neutral resin.
IHC slices were examined at a low magnification (× 100) and the most representative five high-magnification (× 400) fields were selected for staining assessment. A semi-quantitative IHC scoring criterion was used to determine the YKL-39 protein expression levels in tumor. The percent positivity of staining cells ranged from 0 to 4: 0, none; 1, 1–25%; 2, 26–50%; 3, 51–75%; 4, 76–100%. The intensity of staining was graded from 0 to 3 (0, no staining; 1, weak; 2, moderate; and 3, strong). We obtained the final IHC score by multiplying the proportion score by the intensity score. A median score of 6 (> 6 or ≤ 6) was selected as the cutoff to distinguish patients with YKL-39-positive or YKL-39-negative expression. CD68 expression was determined by the number of positive cells expressed, not the intensity of staining. The proportion of cells staining positively for CD68 to tumor interstitial cells was calculated as the CD68-positive rate which was used as a score, ranging from 0 to 10. By counting the staining results and calculating the median, it was determined that 6 was the cutoff point to distinguish between high and low expression of CD68, with a score ≥ 6 for high expression and < 6 for low expression [20]. The number of CD34-positive labeled vessels was determined with each vessel counted as one point. Five × 200 fields of view were observed and the average value was calculated. A median of 17.7 was calculated by counting the staining results to distinguish between dense and sparse angiogenesis [26].
IF staining and confocal microscopy
Formalin fixed, paraffin embedded sections were deparaffinized by xylene and rehydrated. The slices were placed in a pressure cooker with sodium citrate and boiled, then cooled gradually to room temperature to complete the antigen repair. Blocking solution consisted of 0.3% Triton + 10 mg/mL bovine serum albumin + PBS and used for 1 h. The primary antibodies used include YKL-39 (1:100, Bioss, China) and CD68 (1:50, BOSTER, China) and were incubated with the tissue overnight at 4 °C. The secondary antibodies used were goat anti-mouse-Alexa Fluor 594 (1:200) and goat anti-rabbit-Alexa Fluor 488 (1:200) and were used according to instructions. The secondary antibodies were dropped onto the tissues and incubated for 2 h at room temperature and protected from light. After cleaning the slides with PBS dilution, anti-fluorescence quenching sealing liquid (including DAPI) was added and the slides were stored at 4 °C and protected from light.
Confocal laser scanning microscopy was performed with a Leica TCS SP8 laser scanning spectral confocal microscope (OLYMPUS, FV10-ASW2.1 Viewer, Japan), equipped with a 63 £ 1.32 objective. Excitation was with laser emitting at 405 nm, 488 nm, and 559 nm. All two-or three-color images were acquired using a sequential scan mode.
Statistical analysis
SPSS 25.0 was used for statistical analysis. The relationship between YKL-39, CD68, CD34, and clinicopathological characteristics of patients was tested by a χ² test. Correlation analysis used Spearman's rank correlation coefficient. Survival analyses were conducted using the Kaplan-Meier method and differences in survival were examined using the log-rank test. Univariate and multivariate analyses were conducted using the Cox proportional hazards regression models. Statistical significance was defined as a value of p < 0.05.
Results
Relationship between YKL-39, CD68, and CD34 expression and clinicopathological characteristics of GC patients
Among the 119 IHC staining results, 69 (57.98%) cases showed elevated expression of YKL-39, 68 (57.14%) cases showed elevated expression of CD68, and 59 (49.58%) cases showed elevated expression of CD34.
We analyzed the correlation between YKL-39, CD68, and CD34 expression and patient gender, age, depth of tumor infiltration, lymph node metastasis, distant metastasis, TNM stage, and degree of differentiation. YKL-39 protein expression was associated with the depth of tumor infiltration (p = 0.018), lymph node metastasis (p = 0.029), and TNM stage (p = 0.003). Tumor infiltration was deeper, lymph node metastasis was more frequent, and TNM stage was higher, and YKL-39 expression was higher. CD68 protein expression was associated with lymph node metastasis (p = 0.048) and TNM stage (p = 0.029), with more lymph node metastasis and higher TNM stage associated with higher CD68 expression. There was no significant correlation between CD34 expression and clinicopathological characteristics (Table 1).
Correlation between YKL-39 expression and TAMs infiltration and angiogenesis in GC tissue
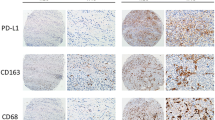
IHC staining showed that YKL-39 was expressed in both the nucleus and cytoplasm of GC tumor cells and stromal cells (Fig. 1A). IHC staining for CD68 showed cytoplasmic staining, diffusely distributed in the tumor stromal cells, and was used to label TAMs (Fig. 1A). MVD is an important index of tumor angiogenic activity and intensity. To evaluate tumor angiogenesis, MVD was evaluated by IHC staining with CD34. To determine the relationship between YKL-39 protein expression and TAMs infiltration and MVD, we performed a correlation analysis with Spearman’s correlation coefficient (Table 2). The results show that positive reactions for both CD68 and CD34 were increased in the samples with higher YKL-39 expression. YKL-39 expression significantly correlates with CD68 and CD34 expression in GC (p < 0.001).

YKL-39, CD68 and CD34 IHC staining result and IF staining results. A IHC staining images of high and low expression of YKL-39, CD68, and CD34 protein in gastric adenocarcinoma. (× 200, horizontal lines represent 50 μm). B IF staining showed co-expression of YKL-39 (green fluorescence) and CD68 (red fluorescence) in gastric adenocarcinoma tissue (× 20, horizontal lines represent 500 μm)
To determine the sites of expression of YKL-39 and CD68 in GC tissues, IF and confocal microscopy analysis was performed on samples from 10 patients with GC. The results show that YKL-39 was expressed in both tumor cells and tumor mesenchyme, and co-expressed with macrophages in the tumor mesenchyme (Fig. 1B).
Survival analysis and the prognostic value of YKL-39, CD68 and CD34 expression in patients with GC
Of the 119 cases included in this study, 75 were followed up and 44 were lost to follow-up, due to communication problems (change of phone number, refusal to answer). Among the 75 patients with follow-up, 28 patients had died during the last follow-up period. To explore the prognostic effects of YKL-39, CD68, and CD34 protein expression in GC, we plotted Kaplan-Meier survival curves and performed log-rank test (Fig. 2A–C). The results showed that patients with high YKL-39 expression had a significantly worse prognosis than those with low YKL-39 expression (p < 0.001, hazard ratio = 1.4). Patients expressing high CD68 and CD34 had a worse prognosis than those with low expression (p < 0.001).

Survival analysis and the prognostic value of YKL-39, CD68 and CD34 expression in patients with GC. Kaplan–Meier survival curves of A YKL-39, B CD68, C CD34 expression in GC patients. D Kaplan-Meier survival curves of OS among the four patient groups stratified by YKL-39 and CD68 expression. E Kaplan-Meier survival curves of OS among four patient groups stratified by YKL-39 and CD34 expression. F Results of univariate COX regression analysis of YKL-39, CD68, CD34 expression, and clinicopathological information. G Results of multivariate COX regression analysis of YKL-39, CD68, CD34 expression, and clinicopathological information
As shown in Fig. 2D, when combining CD68 and YKL-39 to analyze prognosis, patients in the high CD68 low YKL-39 group had a significantly better prognosis than those in the high CD68 high YKL-39 group. The same situation also occurred when combining CD34 and YKL-39 to analyze the prognosis of patients (Fig. 2E) (p < 0.001). From Fig. 2D, patients in the high CD68-low YKL-39 group had the best prognosis. In contrast, as shown in Fig. 2E, the prognosis of patients in the high CD34-low YKL-39 group and low CD34-low YKL-39 groups did not differ significantly, indicating that CD34 expression did not have a significant impact on the prognosis of patients.
To further discern the independent prognostic role of YKL-39 in GC, we performed Cox regression analysis. The results of the univariate independent prognostic analysis showed a statistically significant relationship between age, N-stage, YKL-39 expression, MVD and survival outcome (p < 0.05) (Fig. 2F). Multivariate COX regression results showed that only YKL-39 expression was an independent prognostic factor for GC (p < 0.05) (Fig. 2G). Our results demonstrate that YKL-39 may serve as an independent prognostic factor for GC.
Discussion
Despite advances in diagnostic and therapeutic techniques, GC retains a median survival of less than 12 months for patients with advanced GC [3]. Therefore, there is an urgent need to explore new biomarkers to predict the prognosis of GC patients as well as to guide individualized treatment. In this study we measured the expression of YKL-39, CD68, and CD34 in GC cells and mesenchyme by IHC, determined the distribution of YKL-39 and TAMs in GC tissue by IF, and analyzed the role of YKL-39 in GC. Our results indicate that YKL-39 an independent prognostic factor in GC.
Both chitinases and CLPs contain Glyco-18 structural domains, and in humans, chitinases include acidic mammalian chitinase and chitotriosidase, which cleave chitin polymers into oligosaccharides and release monosaccharides from the ends of chitin polymers [27]. CLPs do not possess glycosyl hydrolase activity due to the substitution of the catalytic glutamate in the active site DxxDxDxE motif terminus by leucine, isoleucine or tryptophan, but due to the Glyco-18 structural domain, they still have sugar-binding properties [28,29,30]. The human CLPs include YKL-40, YKL-39, and SI-CLP, of which YKL-40 (CHI3L1) is the best-studied and is upregulated in a variety of inflammatory and neurodegenerative diseases and cancers [16]. YKL-40 mediates activation of mitogen-activated protein kinases and phosphoinositide 3-kinase pathways through phosphorylation of extracellular signal-regulated kinase 1/2 and protein kinase B. Expression of YKL-40 is elevated in a variety of cancers, including breast cancer, colon cancer, lung cancer, prostate cancer, bladder cancer, and GC, and promotes tumor progression by promoting tumor cell proliferation, rapid invasion and migration, and angiogenesis [31]. Previous studies have confirmed that anti-YKL-40 treatment showed good anti-angiogenic effects in animal tumor models both in vivo and in vitro [32]. YKL-39 and YKL-40 show some similarity in size and amino acid sequence, which suggests that YKL-39 may have a similar biological function to YKL-40 [15]. Our results suggest that YKL-39 expression is associated with TAMs and angiogenesis in GC. As the most vital immune cells in the TME, the infiltration status of TAMs significantly affects the patient’s response to treatment and their prognosis [33, 34]. TAMs can stimulate solid tumor development and invasion by secreting a variety of pro-angiogenic factors, such as VEGF-A, PLGF and transforming growth factor-β [35]. The level of TAMs is positively correlated with the degree of MVD in a variety of tumors [11, 36]. In recent studies, YKL-39 has been identified in breast cancer as having monocyte chemotactic activity and pro-angiogenic activity, which is consistent with the functions of YKL-39 we found in GC. YKL-39 does not work directly on tumor cells but can influence tumor progression by altering the state of the TME, and elevated YKL-39 levels are associated with increased risk of metastasis and post-chemotherapy drug resistance in breast cancer [21]. Our results also show that higher YKL-39 protein expression in GC tissues was associated with deeper tumor infiltration, more lymph node metastases, and higher TNM stage. Similarly, high YKL-39 protein expression in renal cell carcinoma is significantly associated with tumor relapse, size, grade, and T-stage [22]. The Kaplan-Meier survival curve and log-rank test demonstrates that YKL-39 expression correlated with the prognosis of GC patients, and multivariate Cox regression analysis indicated that YKL-39 expression in tumors was an independent predictor of GC progression. These results argue that YKL-39 is a valuable prognostic biomarker for GC. In the study by Liu et al, YKL-39 was shown to be associated with tumor immune infiltration and affect the prognosis of patients with glioma, and high expression of YKL-39 is an independent adverse prognostic factor in glioma patients [20]. In renal cell carcinoma, high expression of YKL-39 is associated with poor patient prognosis and tumor recurrence [22]. In summary, combined with the findings on the function of the YKL-39 and its effect on prognosis in other tumors, we suggest that YKL-39 may act aggressively in malignant tumors. Our results demonstrate that YKL-39 is associated with TAMs infiltration and tumor angiogenesis in GC, and we suggest that YKL-39 has the potential to become a target for anti-TAMs aggregation and anti-tumor angiogenesis in GC. Our study indicates that the specific function of YKL-39 in GC needs to be further confirmed by cellular experiments.
Conclusion
In summary, our results demonstrate that YKL-39 expression in GC is positively correlated with the depth of tumor infiltration, lymph node metastasis, TNM stage, TAMs infiltration and tumor angiogenesis, and is significantly associated with the prognosis of GC patients. Our results indicate that YKL-39 is associated with angiogenesis and is involved in the progression of GC, and that YKL-39 has the potential to be a novel prognostic biomarker and therapeutic target for GC.
Availability of data and materials
Not applicable.
Abbreviations
- GC:
-
Gastric cancer
- OS:
-
Overall survival
- VEGF-A:
-
Vascular endothelial growth factor-A
- PLGF:
-
Placental growth factor
- TAM:
-
Tumor-associated macrophages
- TME:
-
Tumor microenvironment
- TNFα:
-
Tumor necrosis factor α
- CLPs:
-
Chitinase-like proteins
- SI-CLP:
-
Stabilin-1 interacting chitinase-like protein
- IHC:
-
Immunohistochemistry
- IF:
-
Immunofluorescence
- FFPE:
-
Formalin-fixed, paraffin-embedded
- TMA:
-
Tissue microarray
- MVD:
-
Micro vessel density
References
Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries [J]. CA Cancer J Clin. 2021;71(3):209–49.
Nienhuser H, Schmidt T. Angiogenesis and Anti-Angiogenic Therapy in Gastric Cancer [J]. Int J Mol Sci. 2017;19(1):43.
Digklia A, Wagner AD. Advanced gastric cancer: current treatment landscape and future perspectives [J]. World J Gastroenterol. 2016;22(8):2403–14.
Hsieh HL, Tsai MM. Tumor progression-dependent angiogenesis in gastric cancer and its potential application [J]. World J Gastrointest Oncol. 2019;11(9):686–704.
Sisic L, Strowitzki MJ, Blank S, et al. Postoperative follow-up programs improve survival in curatively resected gastric and junctional cancer patients: a propensity score matched analysis [J]. Gastric Cancer. 2018;21(3):552–68.
Jaszai J, Schmidt MHH. Trends and Challenges in Tumor Anti-Angiogenic Therapies [J]. Cells. 2019;8(9):1102.
Vasudev NS, Reynolds AR. Anti-angiogenic therapy for cancer: current progress, unresolved questions and future directions [J]. Angiogenesis. 2014;17(3):471–94.
Fu LQ, Du WL CMH, et al. The roles of tumor-associated macrophages in tumor angiogenesis and metastasis [J]. Cell Immunol. 2020;353:104119.
Cassetta L, Pollard JW. Targeting macrophages: therapeutic approaches in cancer [J]. Nat Rev Drug Discov. 2018;17(12):887–904.
Chanmee T, Ontong P, Konno K, et al. Tumor-associated macrophages as major players in the tumor microenvironment [J]. Cancers (Basel). 2014;6(3):1670–90.
Shieh YS, Hung YJ, Hsieh CB, et al. Tumor-associated macrophage correlated with angiogenesis and progression of mucoepidermoid carcinoma of salivary glands [J]. Ann Surg Oncol. 2009;16(3):751–60.
Osinsky S, Bubnovskaya L, Ganusevich I, et al. Hypoxia, tumour-associated macrophages, microvessel density, VEGF and matrix metalloproteinases in human gastric cancer: interaction and impact on survival [J]. Clin Transl Oncol. 2011;13(2):133–8.
Ramjiawan RR, Griffioen AW, Duda DG. Anti-angiogenesis for cancer revisited: Is there a role for combinations with immunotherapy? [J]. Angiogenesis. 2017;20(2):185–204.
Loges S, Schmidt T, Carmeliet P. Mechanisms of resistance to anti-angiogenic therapy and development of third-generation anti-angiogenic drug candidates [J]. Genes Cancer. 2010;1(1):12–25.
Kzhyshkowska J, Yin S, Liu T, et al. Role of chitinase-like proteins in cancer [J]. Biol Chem. 2016;397(3):231–47.
Yeo IJ, Lee CK, Han SB, et al. Roles of chitinase 3-like 1 in the development of cancer, neurodegenerative diseases, and inflammatory diseases [J]. Pharmacol Ther. 2019;203:107394.
Sanfilippo C, Longo A, Lazzara F, et al. CHI3L1 and CHI3L2 overexpression in motor cortex and spinal cord of sALS patients [J]. Mol Cell Neurosci. 2017;85:162–9.
Mollgaard M, Degn M, Sellebjerg F, et al. Cerebrospinal fluid chitinase-3-like 2 and chitotriosidase are potential prognostic biomarkers in early multiple sclerosis [J]. Eur J Neurol. 2016;23(5):898–905.
Sanfilippo C, Malaguarnera L, Di Rosa M. Chitinase expression in Alzheimer's disease and non-demented brains regions [J]. J Neurol Sci. 2016;369:242–9.
Liu L, Yang Y, Duan H, et al. CHI3L2 is a novel prognostic biomarker and correlated with immune infiltrates in gliomas [J]. Front Oncol. 2021;11:611038.
Liu T, Larionova I, Litviakov N, et al. Tumor-associated macrophages in human breast cancer produce new monocyte attracting and pro-angiogenic factor YKL-39 indicative for increased metastasis after neoadjuvant chemotherapy [J]. Oncoimmunology. 2018;7(6):e1436922.
Pusztai C, Yusenko MV, Banyai D, et al. M2 macrophage marker chitinase 3-like 2 (CHI3L2) associates with progression of conventional renal cell carcinoma [J]. Anticancer Res. 2019;39(12):6939–43.
Albini A, Bruno A, Noonan DM, et al. Contribution to tumor angiogenesis from innate immune cells within the tumor microenvironment: implications for immunotherapy [J]. Front Immunol. 2018;9:527.
Larionova I, Tuguzbaeva G, Ponomaryova A, et al. Tumor-associated macrophages in human breast, colorectal, lung, ovarian and prostate cancers [J]. Front Oncol. 2020;10:566511.
Yugawa K, Itoh S, Yoshizumi T, et al. Prognostic impact of tumor microvessels in intrahepatic cholangiocarcinoma: association with tumor-infiltrating lymphocytes [J]. Mod Pathol. 2021;34(4):798–807.
Teixeira LR, Almeida LY, Silva RN, et al. Young and elderly oral squamous cell carcinoma patients present similar angiogenic profile and predominance of M2 macrophages: comparative immunohistochemical study [J]. Head Neck. 2019;41(12):4111–20.
Di Rosa M, Distefano G, Zorena K, et al. Chitinases and immunity: ancestral molecules with new functions [J]. Immunobiology. 2016;221(3):399–411.
Schimpl M, Rush CL, Betou M, et al. Human YKL-39 is a pseudo-chitinase with retained chitooligosaccharide-binding properties [J]. Biochem J. 2012;446(1):149–57.
Fusetti F, von Moeller H, Houston D, et al. Structure of human chitotriosidase. Implications for specific inhibitor design and function of mammalian chitinase-like lectins [J]. J Biol Chem. 2002;277(28):25537–44.
Kzhyshkowska J, Larionova I, Liu T. YKL-39 as a potential new target for anti-angiogenic therapy in cancer [J]. Front Immunol. 2019;10:2930.
Areshkov PO, Avdieiev SS, Balynska OV, et al. Two closely related human members of chitinase-like family, CHI3L1 and CHI3L2, activate ERK1/2 in 293 and U373 cells but have the different influence on cell proliferation [J]. Int J Biol Sci. 2012;8(1):39–48.
Shao R. YKL-40 acts as an angiogenic factor to promote tumor angiogenesis [J]. Front Physiol. 2013;4:122.
Heinhuis KM, Ros W, Kok M, et al. Enhancing antitumor response by combining immune checkpoint inhibitors with chemotherapy in solid tumors [J]. Ann Oncol. 2019;30(2):219–35.
Kim R, An M, Lee H, et al. Early Tumor-Immune Microenvironmental Remodeling and Response to First-Line Fluoropyrimidine and Platinum Chemotherapy in Advanced Gastric Cancer [J]. Cancer Discov. 2022;12(4):984–1001.
Chen Y, Song Y, Du W, et al. Tumor-associated macrophages: an accomplice in solid tumor progression [J]. J Biomed Sci. 2019;26(1):78.
Valkovic T, Dobrila F, Melato M, et al. Correlation between vascular endothelial growth factor, angiogenesis, and tumor-associated macrophages in invasive ductal breast carcinoma [J]. Virchows Arch. 2002;440(6):583–8.
Acknowledgements
We acknowledge the grants and support from: Colleges and Universities Innovation Ability Improvement Project of Gansu Province, China (No. 2021B-166), Double first-class scientific research key projects in Gansu Province (No. GSSYLXM-21), Foundation of Key Laboratory of Dunhuang Medicine and Transformation Constructed by Chinese Ministry of Education and Gansu Province (NO.DHYX20-07), Scientific Research and Innovation Fund of Gansu University of Traditional Chinese Medicine (2020KCYB-9), and the staffs from the Department of Pathology, 940 Hospital.
Funding
This work was supported by the Colleges and Universities Innovation Ability Improvement Project of Gansu Province, China (No. 2021B-166); Double first-class scientific research key projects in Gansu Province (No. GSSYLXM-21); Foundation of Key Laboratory of Dunhuang Medicine and Transformation Constructed by Chinese Ministry of Education and Gansu Province (No. DHYX20-07); and Scientific Research and Innovation Fund of Gansu University of Traditional Chinese Medicine (2020KCYB-9).
Author information
Authors and Affiliations
Contributions
L.X., P.W., and X.Z. wrote the manuscript text and L.X and W.C. prepared all figures and tables. All authors reviewed the manuscript. All authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional research committee (Medical Ethics Committee of the 940th Hospital of the Joint Security Force) and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xue, L., Chu, W., Wan, F. et al. YKL-39 is an independent prognostic factor in gastric adenocarcinoma and is associated with tumor-associated macrophage infiltration and angiogenesis. World J Surg Onc 20, 362 (2022). https://doi.org/10.1186/s12957-022-02830-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12957-022-02830-9