
Abstract
Background
GABAA receptor subunit composition has a profound effect on the receptor’s physiological and pharmacological properties. The receptor β subunit is widely recognised for its importance in receptor assembly, trafficking and post-translational modifications, but its influence on extrasynaptic GABAA receptor function is less well understood. Here, we examine the pharmacological properties of a potentially native extrasynaptic GABAA receptor that incorporates the β1 subunit, specifically composed of α4β1δ and α4β1 subunits.
Results
GABA activated concentration-dependent responses at α4β1δ and α4β1 receptors with EC50 values in the nanomolar to micromolar range, respectively. The divalent cations Zn2+ and Cu2+, and the β1-selective inhibitor salicylidine salicylhydrazide (SCS), inhibited GABA-activated currents at α4β1δ receptors. Surprisingly the α4β1 receptor demonstrated biphasic sensitivity to Zn2+ inhibition that may reflect variable subunit stoichiometries with differing sensitivity to Zn2+. The neurosteroid tetrahydro-deoxycorticosterone (THDOC) significantly increased GABA-initiated responses in concentrations above 30 nM for α4β1δ receptors.
Conclusions
With this study we report the first pharmacological characterisation of various GABAA receptor ligands acting at murine α4β1δ GABAA receptors, thereby improving our understanding of the molecular pharmacology of this receptor isoform. This study highlights some notable differences in the pharmacology of murine and human α4β1δ receptors. We consider the likelihood that the α4β1δ receptor may play a role as an extrasynaptic GABAA receptor in the nervous system.
Similar content being viewed by others


Background
GABAA receptors are pentameric ligand-gated ion channels that mediate phasic and tonic inhibition in the central nervous system [1]. When activated by GABA the integral ion channel opens, rendering the receptor permeable to chloride ions. The co-assembly of combinations of 19 different subunits (α1-6, β1-3, γ1-3, δ, ε, θ, π and ρ1-3) into homo- or hetero-pentameric receptors results in multiple different GABAA receptor isoforms with different function, pharmacology and neuronal location [2-4]. The stoichiometry of the majority of these receptors is considered to be 2α:2β:1γ/δ, predominantly arranged as β-α-β-α-γ/δ in an anticlockwise manner when viewed from the extracellular space [5-7]. However, the arrangement for δ subunit-containing receptors seems to be dependent on the experimental conditions [6,8-12].
Receptors incorporating α1, α2 or α3 subunits most commonly assemble with β and γ subunits, and are generally located at the synapse where they mediate phasic inhibition; whereas α4 and α6-containing receptors assemble with β and δ subunits, and are found extrasynaptically, mediating tonic inhibition [13]. The δ subunit is found almost exclusively in the extrasynaptic domain and forms α4βδ or α6βδ receptors in the cortex and cerebellum, respectively [2]. However, it has also been suggested that δ can co-assemble with α1 subunits in hippocampal interneurons [14,15].
The existence of α4β2/3δ receptors in brain regions such as the hippocampal dentate gyrus is well established, but there is a paucity of data which characterises the potential native expression of the α4β1δ receptor [2]. This is largely due to the limited number of β1-subtype-selective compounds available [16], and the fact that no β1 subunit knock-out mouse has been generated. As a consequence, most previous studies investigating α4βδ receptors have focused on those subtypes containing the β2 or β3 subunits as these are confirmed to be native receptor combinations [2,17,18]. However, co-regulation of the expression levels of α4, β1 and δ subunits in, for example, the periaqueductal grey matter throughout the female oestrous cycle has been reported, suggesting that α4β1δ receptors may perform a hitherto overlooked important physiological role [19]. Also, experimental evidence for the co-expression of α4, β1 and δ subunits at extrasynaptic sites exists for pyramidal hippocampal neurons [20]. Furthermore, recent studies have emphasized the importance of β1-containing receptors in relation to sleep control [21] and ataxia [22], and polymorphic mutants of the β1 subunit are thought to be important in determining the extent of alcohol consumption [23]. Therefore, since α4, β1 and δ subunits are co-expressed in several brain regions [24], the possibility exists that these subunits can co-assemble into native functional receptors, most likely at extrasynaptic sites, raising the prospect that such a receptor subtype may be important for neuronal physiology.
Few studies have investigated the properties of either human [25-27] or murine [23,28] recombinant α4β1δ receptors, and none of these have fully addressed the characterisation of classic GABAA receptor ligands. In these studies, interesting pharmacological differences between murine and human α4β1δ receptors are manifest: human receptors expressed in Xenopus oocytes [25] are constitutively active and display a GABA EC50 in the mid-nanomolar range whereas the rat/mouse receptor expressed in oocytes or HEK293 cells display low-micromolar sensitivity towards activation by GABA [23,28]. To address this apparent species-dependent pharmacology and to address the effects of commonly used GABA receptor ligands, we report the pharmacological profiles for several GABAA receptor ligands at recombinant murine α4β1 and α4β1δ receptors expressed in Xenopus laevis oocytes.
Results
GABA concentration response relationships for α4β1 and α4β1δ receptors
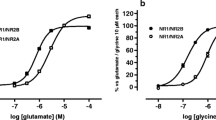
GABA concentration response curves were generated for α4β1 and α4β1δ receptors to evaluate GABA potency at these receptors. Oocytes were challenged with increasing concentrations of GABA (Figure 1A). Depending upon expression levels, maximal currents to GABA ranged from 100 to 3000 nA for cells expressing α4β1 receptors and from 300 to 3000 nA for cells expressing α4β1δ receptors. The holding currents were between 0 and -40 nA and the receptors were not constitutively active. The GABA EC50 values were 0.89 μM for α4β1δ receptors and 2.7 μM for α4β1 receptors (Figure 1B, Table 1). GABA concentration-response data were fitted as monophasic curves, with Hill coefficients of 1.1 and 0.87 for α4β1δ and α4β1 receptors, respectively (Table 1).

Examples of GABA-activated currents recorded from cDNA-injected Xenopus oocytes expressing α4β1δ and α4β1 receptors. A, Representative membrane currents for α4β1δ receptors (upper panel) and α4β1 receptors (lower panel) in response to increasing concentrations of GABA. The oocytes were voltage clamped at -60 mV. B, GABA concentration response curves for α4β1δ (n = 6) and α4β1(n = 5) receptors. All data points represent means ± SEMs.
Inhibition of GABA responses by divalent cations
Zn2+ has been shown to be a useful pharmacological tool to distinguish αβ receptors from αβδ/γ receptors in recombinant expression systems, as δ/γ subunit-containing GABAA receptors are less sensitive to inhibition by Zn2+ compared to their αβ counterparts [29,30]. The increased potency at αβ receptors is attributed to the incorporation of a third β subunit that contains an important histidine residue for coordinating Zn2+ ions at the top of channel lining M2 region [30,31].
We evaluated Zn2+-inhibition of the GABA response for both α4β1 and α4β1δ receptor isoforms to investigate whether Zn2+ can distinguish between these receptor subtypes. Oocytes were pre-incubated for 1 min with various concentrations of Zn2+ followed by co-application of Zn2+ and GABA at an EC75 (3 μM for α4β1δ and 8 μM for α4β1). As expected, Zn2+ exhibited a higher potency at α4β1 than α4β1δ receptors. For α4β1δ receptors, Zn2+ inhibited the GABA response with an IC50 of 3.3 μM (Figure 2A,B, Table 1). By contrast, for α4β1 receptors, biphasic inhibitory behaviour was observed with a high potency component (IC50A = 0.13 nM) accounting for ~25% of the total inhibition, and a more dominant lower potency component (IC50B = 56 nM) accounting for the remaining (~75%) inhibition (Figure 2A,B, Table 1).

Pharmacological modulation of GABA responses at α4β1δ receptors by various inhibitors and the neurosteroid, THDOC. A, Representative membrane currents showing inhibition of GABA (EC75) by 1 μM Zn2+ at α4β1 (upper) and α4β1δ (lower) receptors. B, Zn2+ concentration-inhibition relationships for α4β1 (n = 6) and α4β1δ (n = 6) receptors. C, Representative currents showing the degree of desensitization when activated by EC75 GABA in the absence (upper) and presence (lower) of 1 μM Cu2+. D, Cu2+ concentration-inhibition relationship for GABA EC75 desensitized responses at α4β1δ receptors by increasing concentrations of Cu2+ (n = 12). E, SCS concentration-inhibition relationship for GABA EC20 peak responses at α4β1δ receptors by increasing concentrations of SCS (n = 4). F, Response of α4β1δ receptors to increasing concentrations of THDOC co-applied with an EC7 GABA concentration. The responses were normalised to a preceding application of GABA EC7 in the absence of THDOC (n = 9). All data shown are means ± SEMs.
Cu2+ is another well-established inhibitor of GABA currents at γ subunit-containing GABAA receptors [32], but its inhibitory profile at δ subunit-containing receptors has only recently been explored [33]. This revealed that Cu2+ was far more potent as an inhibitor of steady-state GABA responses at α6β3δ receptors (IC50 = 65 nM) compared to α1β2γ2 receptors (IC50 = 85 μM) expressed in human embryonic kidney 293 (HEK293) cells [33]. To further investigate the inhibitory selectivity of Cu2+ at α4β1δ receptors, we co-applied increasing concentrations of Cu2+ with GABA (EC75: 3 μM) for 70 s, and measured the steady-state responses, reproducing the experimental conditions used by McGee et al. [33] to determine the IC50 values. This response was taken as the fraction of the peak response of a previously-applied GABA concentration, and normalized to the steady-state response of GABA EC75 in the absence Cu2+ (Figure 2C). An almost complete inhibition of the α4β1δ GABA-activated response was observed with increasing concentrations of Cu2+ (IC50 = 0.82 μM) (Figure 2D, Table 1).
Inhibition by the β1 subunit-selective blocker SCS
The salicylic acid derivative, SCS, is one of only a few compounds known to selectively target β1 subunit-containing GABAA receptors over β2- and β3-containing receptors [34]. SCS is reported to inhibit GABA-activated responses of α2β1γ2, α2β1γ1 and α2β1γ1θ receptors, but the pharmacological profile of SCS at α4 or δ-containing receptors, has not been addressed. To examine the interaction of SCS with α4β1δ receptors, we evaluated the inhibition of the GABA EC20 response following pre-application of the antagonist. In this instance, a low EC20 concentration was chosen to enable a direct comparison between our study and that obtained previously [34]. SCS inhibited the GABA response with an IC50 of 4.2 nM, but did not cause complete inhibition of the GABA current, attaining a steady-state level of inhibition at 67.2 ± 3.2% of control (Figure 2E, Table 1).
Neurosteroid potentiation at α4β1δ receptors
Neurosteroids are potent modulators at most, if not all, GABAA receptor isoforms [35]. Some studies report that neurosteroids are more potent at recombinant δ subunit-containing receptors compared to their γ subunit-containing counterparts [17,36,37]. Accordingly, we evaluated the sensitivity of α4β1δ receptors to neurosteroid-induced potentiation. Increasing concentrations of THDOC were co-applied with an approximately EC7 GABA concentration. A significant increase in the GABA peak-response was recorded with THDOC concentrations of 30 and 100 nM (P < 0.05, one-way analysis of variance (ANOVA) with Dunnett’s post test, Figure 2F).
Discussion
In this study we investigated the pharmacological profiles of several GABAA receptor ligands at murine α4β1δ receptors. The GABA potency of 0.89 μM (EC50) at α4β1δ receptors determined in this study is similar to that previously reported for the rat receptor (2.02 μM) expressed in Xenopus oocytes [28], and the mouse/rat receptor (1.99 μM) expressed in HEK293 cells [23]. However, a much higher GABA potency (EC50 of 24 nM) has been reported for the human α4β1δ receptor expressed in Xenopus oocytes [25]. The GABA potency for human α4β1 (0.72 μM) is reported to be somewhat lower [25] than the murine combination studied here. Furthermore, constitutive activity in the absence of GABA was reported for the human recombinant α4β1δ receptor [25], but this was not evident in our studies with the murine equivalent. From these observations it is possible that differences in constitutive activity may explain the considerable differences in GABA potency reported between species observed by others and in this study. However, several other factors may also influence the observed potency shift, such as differences in relative subunit expression levels which may significantly impact on the final receptor stoichiometry. In particular, δ-containing receptors are prone to such variations in stoichiometry. This is evident from studies in which variations in the amount of RNA injected into Xenopus oocytes apparently yield different receptor populations [38]; likewise, concatemeric receptors also demonstrate that alternative assemblies of functional δ-containing receptors are possible [12,39]; and from reports that both high and low affinity conformations of the related α4β3δ receptor apparently exist [25,40].
In this study we report the first characterisation of specific GABA antagonists at α4β1δ receptors. The approximately 60-fold potency shift in the IC50 values for Zn2+ from low micromolar to mid nanomolar, when comparing α4β1δ and α4β1, is consistent with prior observations made for α4β3δ and α4β3 receptors [29]. Our determination of monophasic Zn2+ potency suggests it is highly unlikely that there is any contamination with binary α4β1 receptors in our α4β1δ-expressing oocytes. Also, the inability of Zn2+ to inhibit the response below the holding current baseline corroborates the absence of any constitutive activity caused by homomeric β1 receptors [41].
It has previously been suggested that the stoichiometry of binary GABAA receptors containing only α and β subunits is dependent on the type of α and β subunit present, with a 3α:2β stoichiometry predominating for α1β2 receptors, but a 2α:3β stoichiometry preferred for α4β2 receptors [11]. The biphasic inhibitory behaviour of Zn2+ in our study would suggest the presence of a mixed GABAA receptor population, perhaps with both 3α:2β and 2α:3β stoichiometries expressed in the oocyte. This conclusion is rationalized by the fact that an additional β subunit will contribute another H267 residue, known to be critical for Zn2+ inhibition [31], which may underpin the higher sensitivity component to Zn2+ in the biphasic inhibition curves.
This study also evaluated the pharmacology of additional compounds at α4β1δ known to interact with GABAA receptors. We have established that the pharmacological profile of the β1-selective inhibitor SCS, at α4β1δ receptors, is in agreement with the values reported for α2β1γ2 receptors when expressed in Xenopus oocytes (IC50 = 4.36 nM and maximum inhibition of ~33% [34], suggesting that α4 and δ subunits do not strongly influence the molecular interaction with SCS.
Our data evaluating the inhibition of desensitized GABA responses by Cu2+ suggest that the maximal inhibition of Cu2+ is greater at α4β1δ (98% inhibition at 10 μM: expressed in oocytes) than at α6β3δ receptors (expressed in HEK293) where the maximal inhibition of the steady-state response was reported to be ~ 68% [33]. Although this difference may conceivably be due to the different expression systems, it is equally likely that it reflects some subunit-selectivity (α4 > α6) for Cu2+.
As hypothesized, THDOC concentrations at 30 nM and 100 nM were able to potentiate low (EC7) GABA concentrations, suggesting that α4β1δ receptors are likely to be modulated by physiologically relevant concentrations of neurosteroids, as akin to many other GABAA receptor subtypes [35]. This is unsurprising given that the highly conserved neurosteroid binding site for potentiating GABA responses is contained within the receptor α subunit.
Conclusions
In summary, we have extended the pharmacological profile of the α4β1δ receptor, a subtype that has been studied surprisingly little compared to its β2- and β3-containing counterparts, despite its presence in many significant brain areas. We report a high GABA potency (EC50 of 0.89 μM), a weaker sensitivity to Zn2+ compared to α4β1 receptors, and a high sensitivity to SCS and THDOC.
The observed discrepancy in potency compared to the human ortholog highlights the need for caution when making comparisons of pharmacological parameters across species for this particular GABA receptor subtype. For example, in addition to the different GABA potencies, there are notable differences in constitutive activity for this receptor isoform. Whether these differences translate into the native behaviour of this receptor remains to be determined. Finally, the different expression pattern of the β1 subunit compared to β2 and β3 subunits [24] suggests that β1-containing subtypes may offer novel therapeutic targets and opportunities, and emphasizes a greater need for subtype-selective ligands and/or genetically modified mice to better understand the native function and pharmacology of this GABAA receptor isoform. Indeed recent findings suggest a clinically relevant role for β1-containing extrasynaptic GABAA receptors in the nucleus accumbens mediating addictive behaviours related to excessive alcohol intake [23].
Methods
Mouse α4 and β1 and rat δ cDNA were subcloned into a pRK5 expression vector using methods previously described [42]. γ-Aminobutyric acid (GABA), CuCl2 and tetrahydro-deoxycorticosterone (THDOC) were purchased from Sigma (Dorset, UK), ZnCl2 from BDP Biochemical (Poole, UK) and salicylidine salicylhydrazide (SCS) from Tocris (Bristol, UK).
Expression of GABA subunits
Oocytes from Xenopus laevis toads were prepared as described [43]. Briefly, ovaries were removed using procedures approved by the UK Animals (Scientific Procedures) Act 1986 and local (UCL) ethics committee. To obtain free oocytes, ovarian lobes were cut into pieces and incubated in OR2 (85 mM NaCl, 5 mM HEPES, 1 mM MgCl2, pH adjusted to 7.6 with KOH) supplemented with 2-2.5 mg/mL collagenase type 1 (Lorne Laboratories, Danehill, UK) for 3-4 hrs. Oocytes were washed several times in OR2 and subsequently in Barth’s solution (88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO3)2, 0.41 mM CaCl2, 0.82 mM MgSO4, 2.4 mM NaHCO3, 10 mM HEPES, pH adjusted to 7.6 with NaOH) to isolate cells and remove follicular membranes. After preparation, the oocytes were injected with 27 or 55 nL of 30 ng/μL cDNA in a ratio of 1:1 (α4β1) or 1:1:5 (α4β1δ). Oocytes were subsequently maintained at 18°C in Barth’s solution supplemented with gentamycin (Invitrogen, Paisley, UK).
Electrophysiological recordings
Recordings were performed at room temperature 2-4 days (α4β1) or 3-6 days (α4β1δ) post injection using two-electrode voltage-clamp (Axoclamp 2B amplifier, Digidata 1322A interface and pClamp 8 (Molecular Devices, Wokingham, UK)). Electrodes were filled with 3 M KCl and had resistances of 0.5 to 3 MΩ. The oocytes were clamped at a holding potential of -60 mV and were perfused with buffer (100 mM NaCl, 2 mM KCl, 2 mM CaCl2, 1 mM MgCl2, 5 mM HEPES, pH adjusted to 7.4 with NaOH) throughout the recordings. All currents were adjusted when necessary for voltage drop using equation 1:
Where Imeasured is the observed current, Vh is the holding potential (mV), Vrev is the Cl− reversal potential for oocytes (usually ~ -30 mV), and ΔV is the change in the holding potential (mV) due to loss of clamp during at the peak of large GABA-evoked currents. Drugs were applied using a fast application system coupled to a small bath volume (~1 ml), enabling drug onset times of ~ 10 ms.
Generation of concentration response curves
Agonist concentration response curves were generated by measuring peak responses normalized to a standard concentration of GABA applied between each or every second application (typically an EC10-20 concentration). To pool data from different oocytes, the data were fitted and normalized to the calculated peak of the curve using GraphPad Prism (version 6.0) according to equation 2;
where I and Imax are the GABA activated current and maximal GABA current, respectively, and the EC50 is the GABA concentration inducing a half-maximal response and n is the Hill slope.
Zn2+ and SCS inhibition curves were generated by pre-application (for 60 s) of the inhibitor followed by co-applications of the inhibitor and GABA. For each co-application, the peak-response was measured. For Cu2+-inhibition curves, Cu2+ and GABA were co-applied and the steady-state response after 70 s was measured and normalized to a preceding GABA-application. All monophasic inhibition-curves were fitted to a modified version of equation 2. The biphasic Zn2+-inhibition curve was fitted to equation 3:
where IN / and IN represent the normalised GABA-induced current in the presence and absence of Zn2+ at a concentration (B) respectively. (IC50)A/B define the concentrations of Zn2+ producing 50% inhibition of the GABA-induced current and i and j represent weighting factors for the components A and B, where i + j = 1.
Potentiation by the neurosteroid THDOC was measured by co-application of GABA (EC7) and THDOC until a peak response was attained. Normalized responses were pooled and plotted as mean ± SEM from at least four different oocytes taken from at least two different Xenopus toads. For statistical evaluation, a repeated measures ANOVA with Dunnett’s post test was used. The significance level was set at *P < 0.05, **P < 0.01, ***P < 0.001. For inhibition curves, an F-test was performed to determine whether the data was either monophasic or biphasic.
Abbreviations
- GABA:
-
γ-aminobutyric acid
- HEK293:
-
Human embryonic kidney 293
References
Lüscher B, Keller CA. Regulation of GABAA receptor trafficking, channel activity, and functional plasticity of inhibitory synapses. Pharmacol Ther. 2004;102:195–221.
Olsen RW, Sieghart W. International Union of Pharmacology. LXX. Subtypes of γ-aminobutyric acidA receptors: classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol Rev. 2008;60:243–60.
D’Hulst C, Atack JR, Kooy RF. The complexity of the GABAA receptor shapes unique pharmacological profiles. Drug Discov Today. 2009;14:866–75.
Sigel E, Steinmann ME. Structure, function, and modulation of GABAA receptors. J Biol Chem. 2012;287:40224–31.
Smart TG, Paoletti P. Synaptic neurotransmitter-gated receptors. Cold Spring Harb Perspect Biol. 2012;4:a009662.
Barrera NP, Betts J, You H, Henderson RM, Martin IL, Dunn SMJ, et al. Atomic force microscopy reveals the stoichiometry and subunit arrangement of the α4β3δ GABAA receptor. Mol Pharmacol. 2008;73:960–7.
Barrera NP, Henderson RM, Edwardson JM. Determination of the architecture of ionotropic receptors using AFM imaging. Pflugers Arch. 2008;456:199–209.
Patel B, Mortensen M, Smart TG. Stoichiometry of δ subunit containing GABAA receptors. Br J Pharmacol. 2014;171:985–94.
Chang Y, Wang R, Barot S, Weiss DS. Stoichiometry of a recombinant GABAA receptor. J Neurosci. 1996;16:5415–24.
Neish CS, Martin IL, Davies M, Henderson RM, Edwardson JM. Atomic force microscopy of ionotropic receptors bearing subunit-specific tags provides a method for determining receptor architecture. Nanotechnology. 2003;14:864–72.
Wagoner KR, Czajkowski C. Stoichiometry of expressed α4β2δ γ-aminobutyric acid type A receptors depends on the ratio of subunit cDNA transfected. J Biol Chem. 2010;285:14187–94.
Baur R, Kaur KH, Sigel E. Structure of α6β3δ GABAA receptors and their lack of ethanol sensitivity. J Neurochem. 2009;111:1172–81.
Farrant M, Nusser Z. Variations on an inhibitory theme: phasic and tonic activation of GABAA receptors. Nat Rev Neurosci. 2005;6:215–29.
Glykys J, Peng Z, Chandra D, Homanics GE, Houser CR, Mody I. A new naturally occurring GABAA receptor subunit partnership with high sensitivity to ethanol. Nat Neurosci. 2007;10:40–8.
Milenkovic I, Vasiljevic M, Maurer D, Höger H, Klausberger T, Sieghart W. The parvalbumin-positive interneurons in the mouse dentate gyrus express GABAA receptor subunits alpha1, beta2, and delta along their extrasynaptic cell membrane. Neuroscience. 2013;254:80–96.
Sergeeva OA, Kletke O, Kragler A, Poppek A, Fleischer W, Schubring SR, et al. Fragrant dioxane derivatives identify β1-subunit-containing GABAA receptors. J Biol Chem. 2010;285:23985–93.
Brown N, Kerby J, Bonnert TP, Whiting PJ, Wafford KA. Pharmacological characterization of a novel cell line expressing human α4β3δ GABAA receptors. Br J Pharmacol. 2002;136:965–74.
Herd MB, Haythornthwaite AR, Rosahl TW, Wafford KA, Homanics GE, Lambert JJ, et al. The expression of GABAA β subunit isoforms in synaptic and extrasynaptic receptor populations of mouse dentate gyrus granule cells. J Physiol. 2008;586:989–1004.
Griffiths JL, Lovick TA. GABAergic neurones in the rat periaqueductal grey matter express α4, β1 and δ GABAA receptor subunits: plasticity of expression during the estrous cycle. Neuroscience. 2005;136:457–66.
Mangan PS, Sun C, Carpenter M, Goodkin HP, Sieghart W, Kapur J. Cultured hippocampal pyramidal neurons express two kinds of GABAA receptors. Mol Pharmacol. 2005;67:775–88.
Yanovsky Y, Schubring S, Fleischer W, Gisselmann G, Zhu X-R, Lübbert H, et al. GABAA receptors involved in sleep and anaesthesia: β1- versus β3-containing assemblies. Pflugers Arch. 2012;463:187–99.
Gee KW, Tran MB, Hogenkamp DJ, Johnstone TB, Bagnera RE, Yoshimura RF, et al. Limiting activity at β1-subunit-containing GABAA receptor subtypes reduces ataxia. J Pharmacol Exp Ther. 2010;332:1040–53.
Anstee QM, Knapp S, Maguire EP, Hosie AM, Thomas P, Mortensen M, et al. Mutations in the Gabrb1 gene promote alcohol consumption through increased tonic inhibition. Nat Commun. 2013;4:2816.
Hörtnagl H, Tasan RO, Wieselthaler A, Kirchmair E, Sieghart W, Sperk G. Patterns of mRNA and protein expression for 12 GABAA receptor subunits in the mouse brain. Neuroscience. 2013;236:345–72.
Karim N, Wellendorph P, Absalom N, Bang LH, Jensen ML, Hansen MM, et al. Low nanomolar GABA effects at extrasynaptic α4β1/β3δ GABAA receptor subtypes indicate a different binding mode for GABA at these receptors. Biochem Pharmacol. 2012;84:549–57.
Absalom N, Eghorn LF, Villumsen IS, Karim N, Bay T, Olsen JV, et al. α4βδ GABAA receptors are high-affinity targets for γ-hydroxybutyric acid (GHB). Proc Natl Acad Sci U S A. 2012;109:13404–9.
Karim N, Gavande N, Wellendorph P, Johnston GAR, Hanrahan JR, Chebib M. 3-Hydroxy-2’-methoxy-6-methylflavone: a potent anxiolytic with a unique selectivity profile at GABAA receptor subtypes. Biochem Pharmacol. 2011;82:1971–83.
Lovick TA, Griffiths JL, Dunn SMJ, Martin IL. Changes in GABAA receptor subunit expression in the midbrain during the oestrous cycle in Wistar rats. Neuroscience. 2005;131:397–405.
Stórustovu SI, Ebert B. Pharmacological characterization of agonists at δ-containing GABAA receptors: Functional selectivity for extrasynaptic receptors is dependent on the absence of γ2. J Pharmacol Exp Ther. 2006;316:1351–9.
Hosie AM, Dunne EL, Harvey RJ, Smart TG. Zinc-mediated inhibition of GABAA receptors: discrete binding sites underlie subtype specificity. Nat Neurosci. 2003;6:362–9.
Wooltorton JR, McDonald BJ, Moss SJ, Smart TG. Identification of a Zn2+ binding site on the murine GABAA receptor complex: dependence on the second transmembrane domain of β subunits. J Physiol. 1997;505:633–40.
Fisher JL, Macdonald RL. The role of an α subtype M2-M3 His in regulating inhibition of GABAA receptor current by zinc and other divalent cations. J Neurosci. 1998;18:2944–53.
McGee TP, Houston CM, Brickley SG. Copper block of extrasynaptic GABAA receptors in the mature cerebellum and striatum. J Neurosci. 2013;33:13431–5.
Thompson SA, Wheat L, Brown NA, Wingrove PB, Pillai GV, Whiting PJ, et al. Salicylidene salicylhydrazide, a selective inhibitor of β1-containing GABAA receptors. Br J Pharmacol. 2004;142:97–106.
Hosie AM, Wilkins ME, da Silva HMA, Smart TG. Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature. 2006;444:486–9.
Belelli D, Casula A, Ling A, Lambert JJ. The influence of subunit composition on the interaction of neurosteroids with GABAA receptors. Neuropharmacology. 2002;43:651–61.
Wohlfarth KM, Bianchi MT, Macdonald RL. Enhanced neurosteroid potentiation of ternary GABAA receptors containing the δ subunit. J Neurosci. 2002;22:1541–9.
You H, Dunn SMJ. Identification of a domain in the δ subunit (S238-V264) of the α4β3δ GABAA receptor that confers high agonist sensitivity. J Neurochem. 2007;103:1092–101.
Kaur KH, Baur R, Sigel E. Unanticipated structural and functional properties of δ-subunit-containing GABAA receptors. J Biol Chem. 2009;284:7889–96.
Meera P, Wallner M, Otis TS. Molecular basis for the high THIP/gaboxadol sensitivity of extrasynaptic GABAA receptors. J Neurophysiol. 2011;106:2057–64.
Krishek BJ, Moss SJ, Smart TG. Homomeric β1 γ-aminobutyric acidA receptor-ion channels: evaluation of pharmacological and physiological properties. Mol Pharmacol. 1996;49:494–504.
Connolly CN, Wooltorton JR, Smart TG, Moss SJ. Subcellular localization of γ-aminobutyric acid type A receptors is determined by receptor β subunits. Proc Natl Acad Sci U S A. 1996;93:9899–904.
Gielen MC, Lumb MJ, Smart TG. Benzodiazepines modulate GABAA receptors by regulating the preactivation step after GABA binding. J Neurosci. 2012;32:5707–15.
Acknowledgements
This work was supported by the Lundbeck Foundation (grant number R83-A8000) and the Drug Research Academy. We thank Duncan Laverty for his technical and experimental guidance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ISV, PW and TGS designed the experiments. ISV conducted the experiments. ISV, PW and TGS analysed and interpreted the data. ISV, PW and TGS wrote or contributed to the writing of the manuscript. All authors read and approved the final manuscript.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Villumsen, I.S., Wellendorph, P. & Smart, T.G. Pharmacological characterisation of murine α4β1δ GABAA receptors expressed in Xenopus oocytes. BMC Neurosci 16, 8 (2015). https://doi.org/10.1186/s12868-015-0148-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12868-015-0148-4